 Yanru Chen1,2†
Yanru Chen1,2† Guiming Fu
Guiming Fu
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 14 January 2021
Sec. Food Microbiology
Volume 11 - 2020 | https://doi.org/10.3389/fmicb.2020.592421
Special-flavor Baijiu is a unique Baijiu in Jiangxi Province, China, whose uniqueness mainly depends on the unique production process of special-flavor Baijiu Daqu. However, the microbial structure and physicochemical indices of different parts of the special-flavor Baijiu Daqu are still unknown. This greatly reduces the actual value of Daqu in the production of special-flavor Baijiu. Therefore, culture-dependent and Illumina MiSeq sequencing methods were used to analyze the microbial structure of special-flavor Baijiu Daqu. The results indicated that there was a complicated microbial diversity in Chinese special-flavor Baijiu Daqu. The predominant bacterial communities were Bacillales, Lactobacillales, and Rhodospirillales, while Saccharomycetales and Eurotiales were the predominant fungal communities. Significant differences in microbial community and distribution were shown between the surface and central parts of Daqu. Acetobacter and Pichia genera were the predominant microorganisms in the surface part of Daqu, whereas Aspergillus, Kroppenstedtia, Oceanobacillus, and Bacillus genera were the predominant microorganisms in the central part of Daqu. Meantime, the different microbial distributions between the surface and central parts of Daqu caused the significant differences in the physicochemical indices. These results can provide an important theoretical basis for improving the brewing process and the quality of special-flavor Baijiu.
Chinese Baijiu is an old distilled alcoholic beverage, which plays a crucial role in Chinese tradition, culture, and people’s daily diet. Daqu is a crude fermentation starter, which serves as one of the carriers of microorganisms during the fermentation process of Chinese Baijiu (Zheng et al., 2011). The production of Chinese Daqu adopts a natural inoculation process, and its microbial community comes from raw materials and the air, wherein a complicated microbial community suitable for the specific environment in Daqu gradually forms (Quigley et al., 2012). The main microbial communities of Daqu can be divided into bacteria, mold, and yeast. “Saccharification sees mold, yeast to ferment, raw fragrance by bacteria” is an interesting statement that indicates the role of Daqu microorganisms during the fermentation of Chinese Baijiu. Studies have proved that the aroma profile of Chinese Baijiu is the result of the metabolic activity of its microbial community (Li et al., 2017). Therefore, Daqu is one of the key impact factors that determine the quality and flavor of Chinese Baijiu.
The traditional method is to use the whole Daqu for Chinese Baijiu fermentation, and its use is more dependent on human experience (Zheng et al., 2012). However, with the development of detection methods, the changes in physicochemical indices, enzyme systems, and microbial communities of different parts of Daqu gradually become the new standard for Daqu’s application in Chinese Baijiu fermentation (Yang Y. et al., 2019). It has been reported that there was a significant difference in microbial community and distribution between the surface and central parts of Jiang-flavor Baijiu Daqu, and there was a significant difference in volatile compounds between the surface and the center of Daqu (Jin et al., 2019). Yang Y. et al. (2019) found that different microbial communities could influence the activity of different fermentation-related enzymes between the surface and central parts of medium- and high-temperature Daqu, such as esterase, saccharifying enzyme, and acid protease. These enzymes have an important influence on the formation of liquor flavor. Therefore, it is important to study the physicochemical indices and microbial communities between the different parts of Daqu to promote the scientific development of the modern Chinese Baijiu fermentation industry.
Special-flavor Baijiu, which is one of 12 types of flavored Baijiu in China, has a distinctive aroma with a high concentration of odd-numbered fatty acid ethyl acetate as the primary flavor compound, which contributes to the harmonious and strong flavor (Zheng and Han, 2016). The geographical and climatic conditions in the middle reaches of Ganjiang River in the Jiangxi Province of China provide a suitable environment for the natural inoculation of microorganisms in the special-flavor Daqu. For medium- to high-temperature Daqu, its manufacturing technique adopts an open tempering process that involves natural inoculation. In general, Chinese traditional Daqu is made from a mixture of barley and wheat. However, the production process of special-flavor Daqu is unique because it uses vinasse from pits as one of the raw materials and selectively inoculates acid-tolerant microorganisms in the natural environment (Li et al., 2020). However, the microbial communities and physicochemical indices in different parts of special-flavor Daqu are still unknown.
Therefore, the objectives of this study were to analyze the microbial structure of special-flavor Baijiu Daqu and to evaluate the potential effects between the microbial structure and physicochemical indices of different parts of Daqu by using culture-dependent and Illumina MiSeq sequencing methods. It is of great significance to guide the production of Daqu and improve the quality of special-flavor Baijiu.
Special-flavor Daqu was obtained from the Jiangxi Zhangshugong Wine and Spirits Co. Ltd (longitude 115.53° and latitude 28.07°). Special-flavor Daqu was made from a mixture of 50% flour, 40% wheat bran, and 10% vinasse, which were mixed with warm water after grinding. The Daqu brick (27 cm × 17 cm × 8 cm) was constructed from the mixture and spontaneously incubated and fermented in a Qu room, which is a special open room near the Ganjiang River. Special-flavor Daqu belongs to medium- to high-temperature Daqu, whose fermentation temperature is between 40 and 50°C. The total fermentation period of special-flavor Daqu generally takes approximately 30 days through solid-state fermentation, and the dimension of mature Daqu blocks is 24 cm × 14.5 cm × 7.5 cm (Figure 1).

Figure 1. Schema of traditional method of making special-flavor Baijiu Daqu.
Mature special-flavor Daqu samples (after 30 days of fermentation) were collected according to the description of Shi et al. (2009). We randomly selected three mature Daqu bricks from the room where Daqu is stored. Each Daqu block was divided into two parts: the surface layer of 1.5 cm thick was named the surface part of Daqu, and the remaining central part was named the central part of Daqu. The central part of each Daqu is about 1.0 kg, and the surface part is 0.7 kg. All samples were made in triplicate. Daqu samples were crushed into powder by an antiseptic mortar and stored at −80°C for further experiments.
The content of total acidity was determined according to the method of potentiometric titration. The contents of moisture and reducing sugar were determined according to the atmospheric drying method and 3,5-dinitrosalicylic acid (DNS) method, respectively. The activity of α-amylase was determined according to the starch–iodine method. The activity of acid protease was determined according to the Folin phenol method; one unit of acid protease activity was defined as the amount of tyrosine produced by decomposing casein for 1 min under the conditions of the measurement. The ability of saccharification was determined by measuring released reducing sugars using the DNS method. One unit of saccharification ability was defined as the decomposition of soluble starch to produce 1 mg of glucose per hour under the assay conditions (Li Z.M. et al., 2015; Li et al., 2020). The ability of esterification was determined according to the method of Kumar and Satyanarayana (2009). One unit of esterifying ability was defined as the content of ethyl caproate produced after 100-h reactions.
The microbial count was determined according to the method of Du and Xu (2012) and Li Z.M. et al. (2015). The dilution was coated on lysogeny broth (LB) agar medium and cultured at 37°C for 24 h in a thermostatic incubator (FYL-YS-280L, Beijing, China) for the isolation and enumeration of total aerobic bacteria. Similarly, lactic acid bacteria (LAB) were isolated and enumerated on de Man, Rogosa, and Sharp (MRS) agar medium and were cultured at 37°C for 48 h. Yeast and mold were isolated and enumerated on potato dextrose and Rose Bengal Agar medium, respectively, and were cultured at 28°C for 48 h.
Bacterial genomic DNA was isolated using a TIANamp Bacteria DNA Kit (Tiangen, Beijing, China) following the manufacturer’s specifications. The domain of 16S rDNA sequence was amplified using forward primer 27F (5′-AGAGTTTGATCCTGGCTCAG-3′) and reverse primer 1492R (5′-GGTTACCTTGTTACGACTT-3′) (Han et al., 2013). Fungal genomic DNA was extracted by using a plant genomic DNA kit (Tiangen, Beijing, China), according to the manufacturer’s specifications. For the yeast strains, forward primer NL1 (5′-GCATATCAATAAGCGGAGGAAAAG-3′) and reverse primer NL4 (5′-GGTCCGTGTTTCAAGACGG-3′) were used to amplify the D1/D2 domain of the 26S rRNA gene (Wu et al., 2013). For mold, forward primer ITS1 (5′-TCCGTAGGTGAACCTGCGG-3′) and reverse primer ITS4 (5′-TCCTCCGCTTATTGATATGC-3′) were used to amplify the ITS regions (Ng et al., 2012). PCR amplification was conducted with PCR equipment (ABI GeneAmp 9700, United States) and reagents in a 25-μl reaction solution. The final PCR products were analyzed by 0.8% agarose gel electrophoresis and stored at −20°C for further sequencing analysis.
The PCR products of 16S rRNA gene of bacteria, 26S rRNA gene of yeast, and ITS gene of molds were sequenced by Shenggong Bio, Shanghai, China. And the manual alignment of the sequences was conducted with DNASTAR software (version 4.0). Sequence homologies were inspected by comparing the acquired DNA sequence with those in the NCBI library1.
We took out one third of each of the three parallel samples to make a mixed sample for Illumina MiSeq Sequencing research. Metagenomic DNA was extracted using a Soil Genomic DNA Purification Kit (Tiangen, Beijing, China) following the manufacturer’s instructions. For bacteria, the 16S rRNA gene was amplified according to the method of Li et al. (2020). The 18S rRNA gene fragments in the fungi were amplified by using the bar-coded primer pairs of SSU0817F (5′-TTAGCATGGAATAATRRAATAGGA-3′) and SSU1196R (5′-TCTGGACCTGGTGAGTTTCC-3′) (Rousk et al., 2010). The PCR amplification program was operated as described by Li L. et al. (2015). The PCR products were sent to Majorbio Bio-Pharm Technology Co., Ltd. (Shanghai, China) and run on an Illumina MiSeq PE300 platform. The raw 16S rRNA and 18S rRNA gene sequencing reads were demultiplexed, quality-filtered by fastp version 0.20.0, and merged by FLASH version 1.2.7 with the following criteria: (i) The 300-bp reads were truncated at any site receiving an average quality score of <20 over a 50-bp sliding window, truncated reads shorter than 50 bp were discarded, and reads containing ambiguous characters were also discarded. (ii) Only overlapping sequences longer than 10 bp were assembled according to their overlapped sequence. The maximum mismatch ratio of the overlap region was 0.2. Reads that could not be assembled were discarded. (iii) Samples were distinguished according to the barcode and primers, and the sequence direction was adjusted, with exact barcode matching and two nucleotide mismatches in primer matching. Operational taxonomic units (OTUs) with 97% similarity cutoff were clustered using UPARSE version 7.1, and chimeric sequences were identified and removed. The taxonomy of each OTU representative sequence was analyzed by RDP Classifier version 2.2 against the 16S rRNA database (Silva v138) using a confidence threshold of 0.7.
The experiment was performed three times, except for Illumina MiSeq sequencing. And statistical analysis was plotted with Excel 2013 and Origin 8.0 software. And the significant differences (P < 0.05) between the means of samples were tested by one-way analysis of variance (ANOVA) using the SPSS 17.0 software (SPSS Inc., Chicago, IL, United States). MEGA 5.0 software was used to construct the phylogenetic tree.
As shown in Table 1, the moisture and acidity contents in the central part of Daqu were significantly higher (P < 0.05) than that of the surface part of Daqu, and the content of acidity in the central part was 110% higher than the surface part of Daqu. However, the content of reducing sugars, the activity of acid protease and α-amylase, the ability of esterification and saccharification in the central part of Daqu were significantly lower (P < 0.05) than those of the surface part of Daqu. Among them, the content of reducing sugars in the surface part was 146% higher than that of the central part of Daqu. The activity of acid protease and α-amylase in the surface part was 171% and 120% higher than that of the central part of Daqu, respectively. And the ability of esterification and saccharification in the surface part was 122% and 114% higher than that of the central part of Daqu, respectively.

Table 1. Physicochemical indices of Daqu of special-flavor liquor relative.
Table 2 showed the microbial counts of bacteria and fungi in the different parts of Daqu samples, including total aerobic bacteria, LAB, yeast, and mold. There was no significant difference (P > 0.05) in the total quantities of bacteria between the surface and central parts of Daqu. The LAB amounts in the central part of Daqu were significantly higher (P < 0.05) than those of the surface part of Daqu. Compared with yeast and mold, bacteria were the dominant microorganism in Daqu. The counts of mold in the central part of Daqu were fewer than that of the surface part of Daqu. In addition, yeast had low levels in both the surface and central parts of Daqu, and yeast was primarily focused on the surface part of Daqu.

Table 2. Microbial count in surface and central parts of Daqu.
A total of 100 bacterial strains were randomly selected from the agar plates. Based on the gene sequences of partial 16S rRNAs, the phylogenetic tree showed the remarkable classification among the 14 bacterial species (Table 3 and Figure 2). The bacteria were divided into five categories at the phylotype level, namely, Lactobacillales, Bacillales, Rhodospirillales, Enterobacteriales, and Micrococcales, which accounted for 44%, 40%, 2%, 10%, and 4% of the bacterial community in Daqu, respectively. And 94% of bacteria were under the three categories of Lactobacillales, Bacillales, and Rhodospirillales. Bacillales mainly appeared in the central part of Daqu, and Bacillus licheniformis was the most abundant bacterial community. However, there was no significant difference in the distribution of Lactobacillales in the different parts of Daqu. Enterococcus faecalis had the largest number of 17 strains in Lactobacillales, while Lactobacillus plantarum and Pediococcus pentosaceus had six strains. Acetobacter pasteurianus with 10 strains belonged to Rhodospirillales and was mainly detected on the surface part of Daqu. Enterobacteriales, including Staphylococcus capitis, Klebsiella pneumoniae, and Enterobacteriales sp., accounted for 4% of the bacteria in Daqu and mainly appeared on the surface part of Daqu.
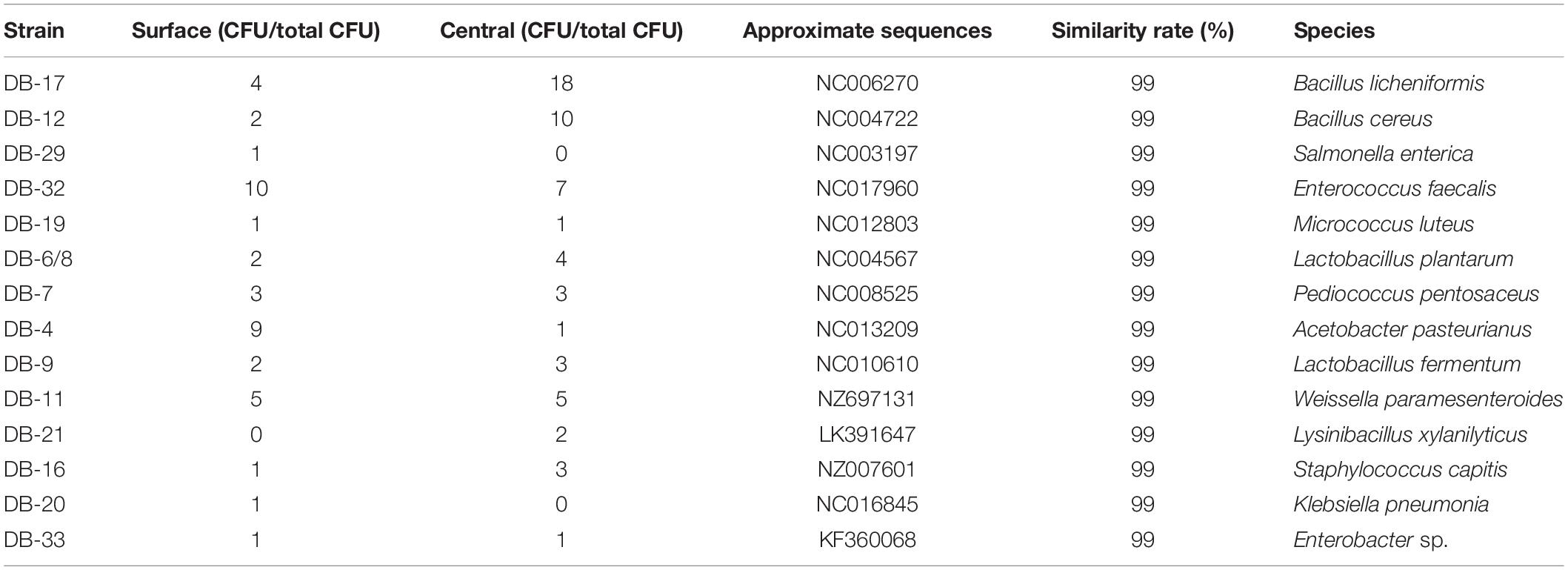
Table 3. Blast and distribution information of bacterial strains of special-flavor Daqu.
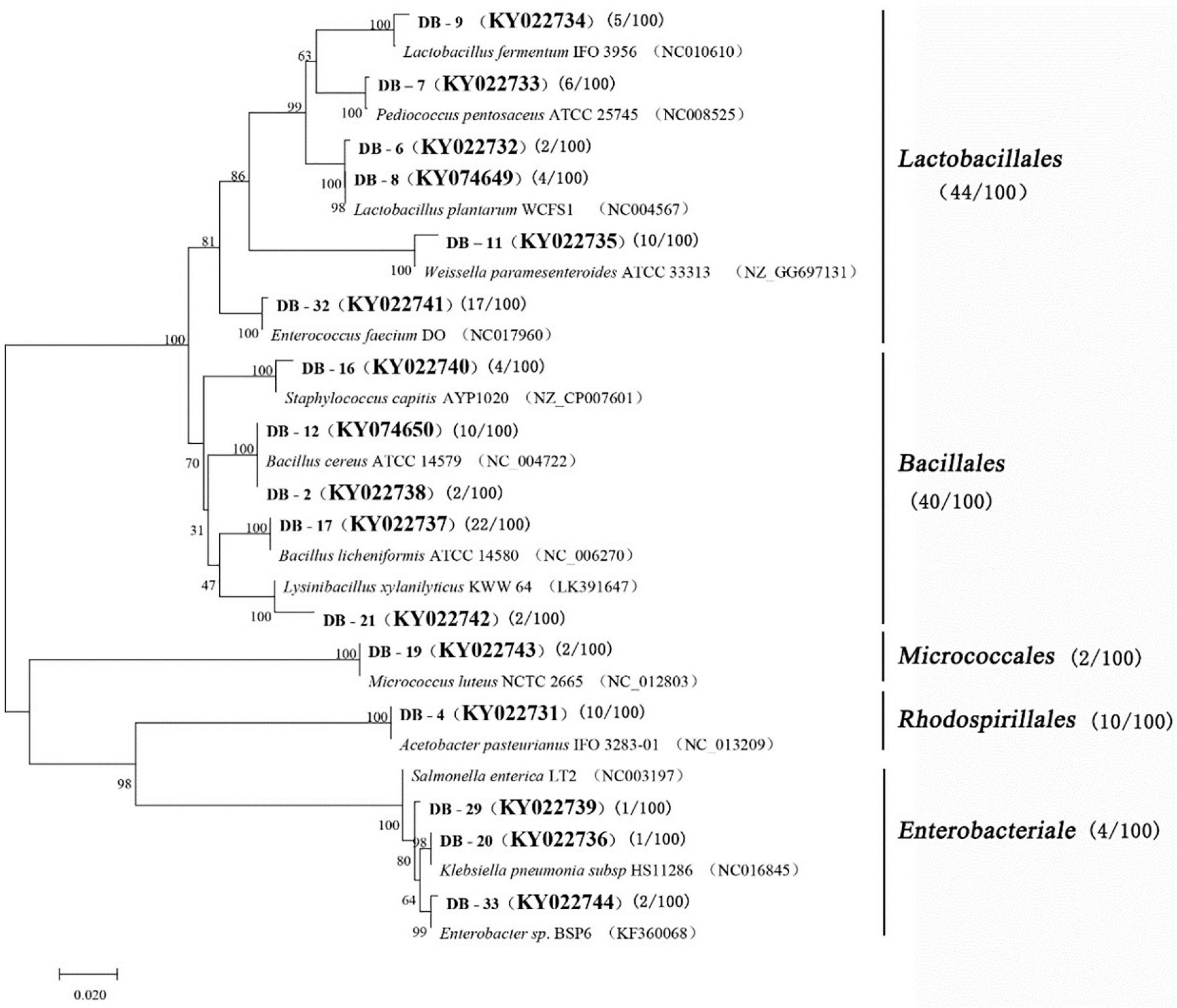
Figure 2. Phylogenetic tree based on l6S rRNA gene sequences.
A total of 120 fungal strains were randomly selected, and a phylogenetic tree was constructed for the sequence fragments from the fungus (Table 4 and Figure 3). The fungus was divided into three categories at phylotype level, namely, Saccharomycetales, Eurotiales, and Capnodiales, which had 60, 58, and 2 strains in the Daqu, respectively. Saccharomycetales mainly appeared on the surface part of Daqu, including Wickerhamomyces anomalus (47 strains), Pichia anomalus (17 strains), and Saccharomycetales cerevisiae (one strain). Aspergillus and Penicillium belonged to Eurotiales, and Aspergillus genus included Aspergillus varians (23 strains), Aspergillus niger (12 strains), Aspergillus carbonarius (11 strains), and Aspergillus flavus (one strain). Penicillium genus included Penicillium citrinum (three strains), Penicillium chrysogenum (two strains), Penicillium crustosum (three strains), and Penicillium corylophilum (three strains).
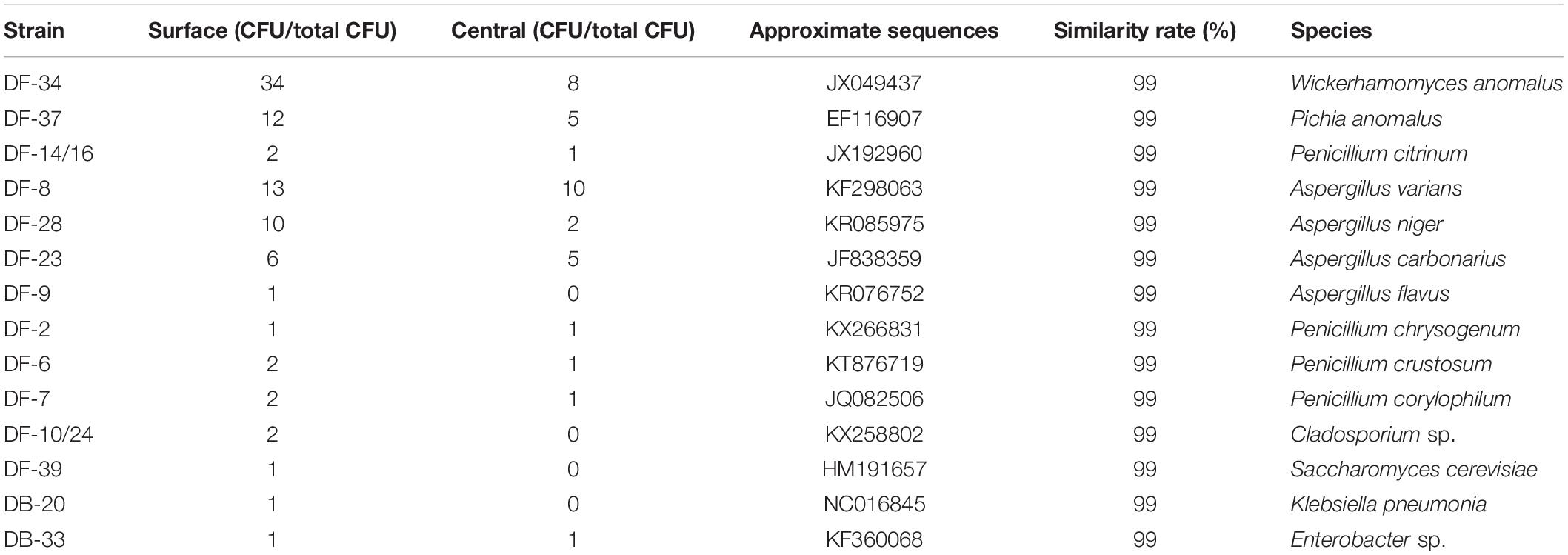
Table 4. Blast and distribution information of fungal strains of special-flavor Daqu.
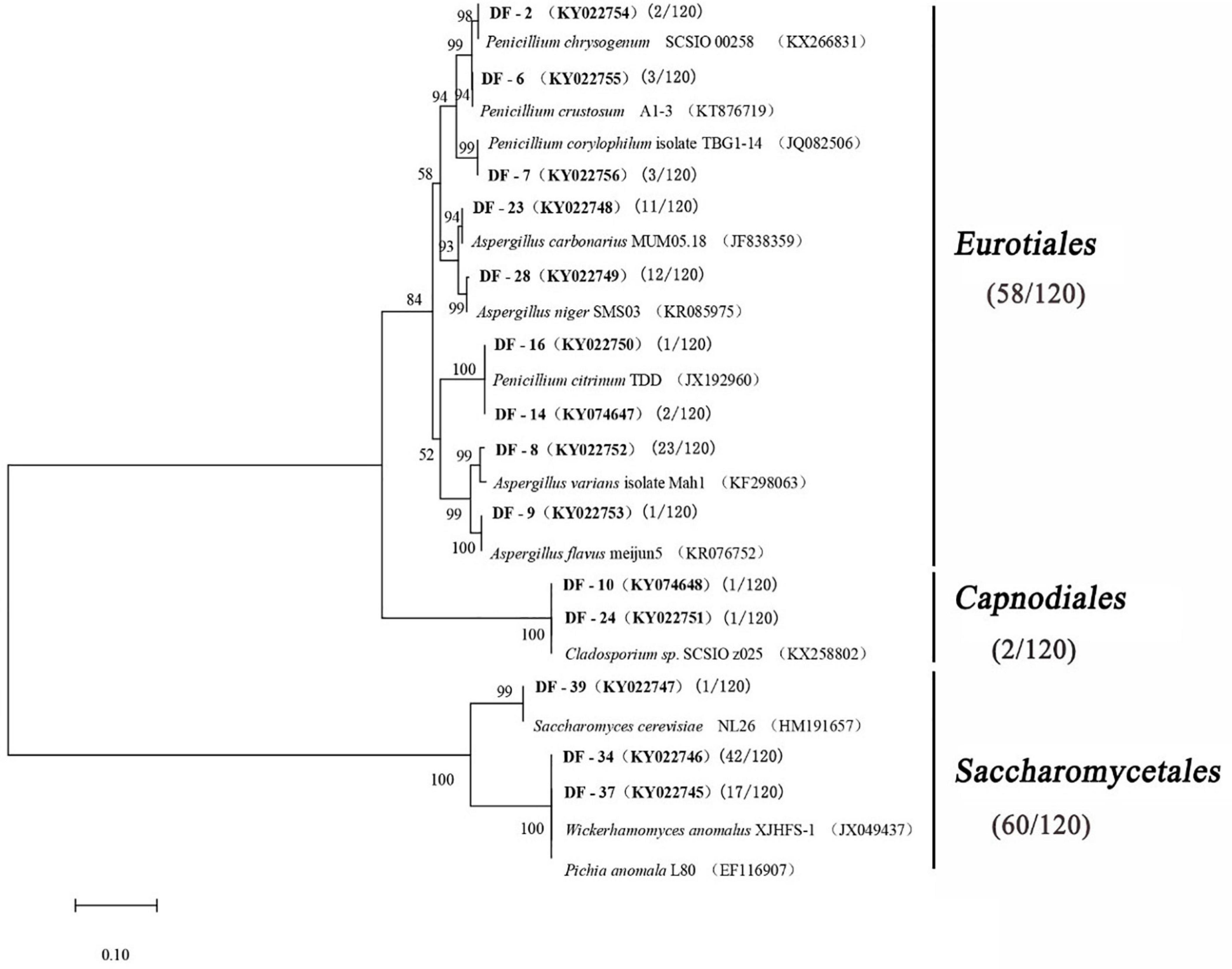
Figure 3. Phylogenetic tree based on the 26S rRNA and ITS gene sequences.
Table 5 showed the diversity, richness, and coverage estimations of each data set. The Shannon diversity index, a measurement of overall diversity, indicated the diversity of the microorganisms. Good’s coverage result, an estimator of sampling completeness, highlighted good overall sampling with levels of 99–100%. A total of 93,466 Illumina MiSeq sequencing reads were obtained. Approximately 16,886–24,092 effective reads with different phylogenetic OTUs ranging from 13 to 89 were obtained from the Daqu samples for microbial communities. Based on the results of Illumina MiSeq sequencing, the number of bacterial OTUs was more than that of fungal OTUs. Bacterial diversity on the surface part of Daqu was lower than that on the central part of Daqu, whereas fungal diversity was similar between the surface and central parts of Daqu.
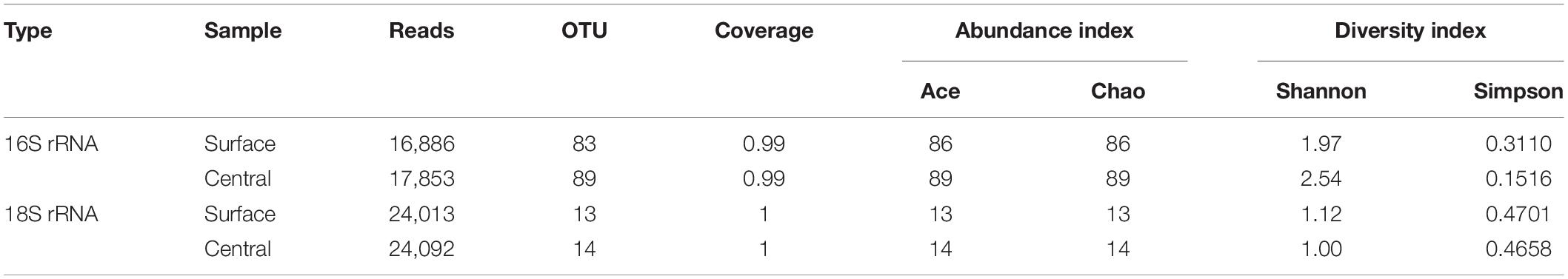
Table 5. Alpha-diversity of special-flavor Daqu.
The sequencing results also showed that different parts of Daqu had various microbial community structures. For bacterial communities (Figures 4A,B), three phyla were identified on the surface and central parts of Daqu. Proteobacteria and Firmicutes were the dominant bacterial communities on the surface part of Daqu, which averaged 55.32% and 42.86% of the bacterial communities, respectively. However, bacterial communities on the central part of Daqu were dominated by Firmicutes, which averaged 95.03% of the communities. At the genus level (Figures 5A,B), over half of the classified bacteria was Acetobacter genus (53.4%), followed by Lactobacillus (25.52%) and Pediococcus (7.42%) genera in the surface part of Daqu. However, Lactobacillus, Kroppenstedtia, Oceanobacillus, and Bacillus genera were the abundant strains in the central part of Daqu, which accounted for 15.7%, 29.9%, 21.9%, and 10.7%, respectively. Moreover, Lactococcus, Weissella, Enterococcus, and Enterobacter genera occurred throughout the surface and central parts of Daqu.
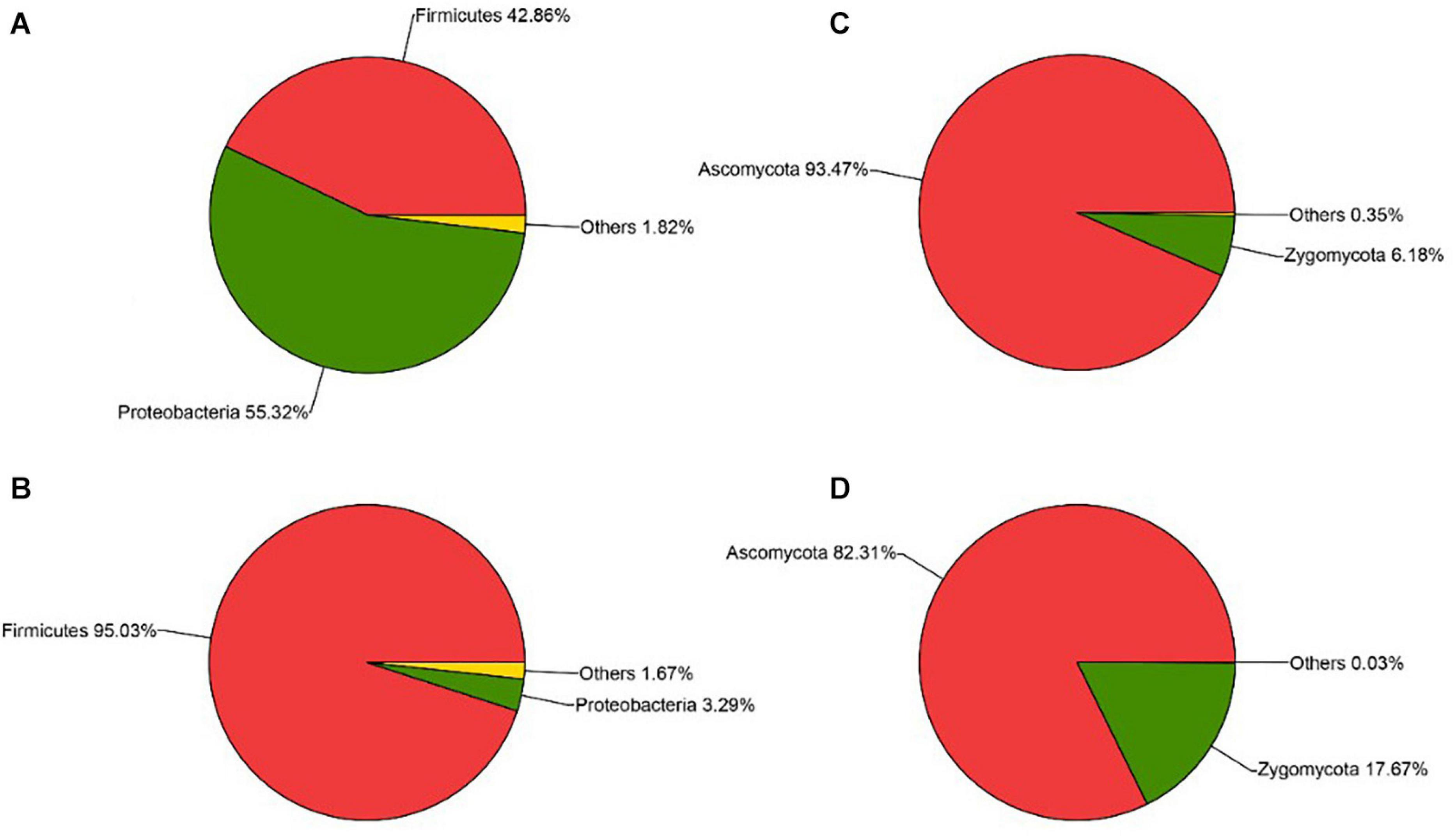
Figure 4. The relative abundances of surface Daqu (A) and central Daqu (B) in bacterial communities at the phylum level, and the relative abundances of surface Daqu (C) and central Daqu (D) in fungal communities at the phylum level.
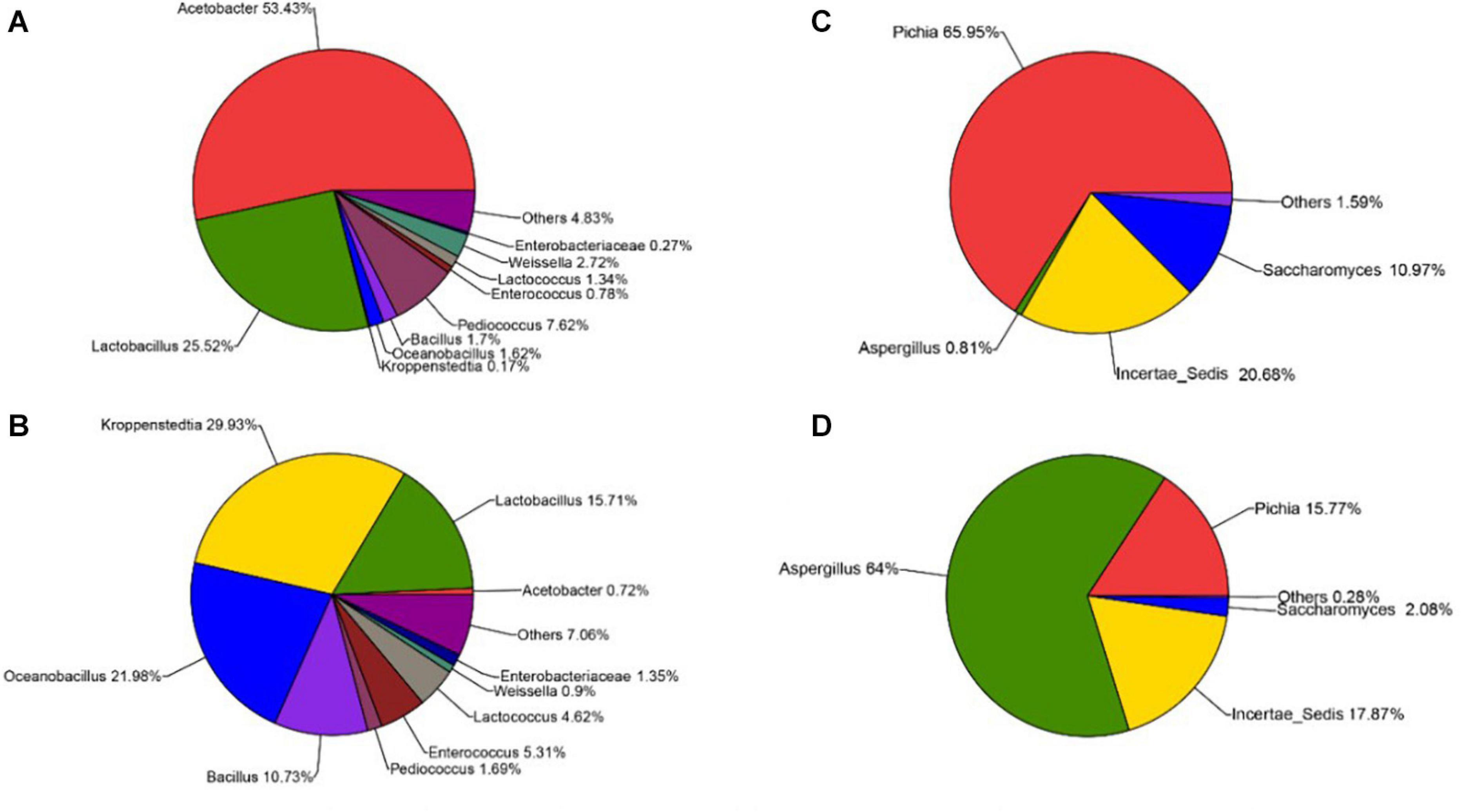
Figure 5. The relative abundances of surface Daqu (A) and central Daqu (B) in bacterial communities at the genus level, and the relative abundances of surface Daqu (C) and central Daqu (D) in fungal communities at the genus level.
Compared with that of bacteria, the fungal community structure was relatively low. At the phylum level, Ascomycota and Zygomycota were detected through sequencing analysis. Ascomycota was the predominant type in both the surface and central parts of Daqu, which accounted for 93.47% and 82.31% of the total sequences, respectively (Figures 4C,D). Figures 5C,D showed that Pichia, Aspergillus, and Saccharomyces were the predominant genera. The abundance of Pichia and Saccharomyces on the surface part of Daqu was higher than that on the central part of Daqu. However, the abundance of Aspergillus genus accounted for 64% of the central part of Daqu, but few on the surface part of Daqu.
Most of the microorganisms in the Chinese Baijiu brewing process come from pit mud, fermented grains, and Daqu. Among them, the main source is from Daqu (Zou et al., 2018). The traditional process of making Daqu is to naturally enrich a variety of microorganisms in Daqu. These microorganisms compete to grow in Daqu and finally adapt to the changing environment of Daqu to survive and reproduce in large numbers. Microbial communities in Daqu will also produce rich enzymes and metabolites in their growth and metabolism process and ultimately give the Chinese Baijiu a unique flavor (Gan et al., 2019). Some studies have found that Bacillus genus could break down starch by producing amylase in Daqu and used decomposition products to generate nitrogenous flavor compounds, such as diverse pyrazines (Mukherjee et al., 2009; Zheng et al., 2012). Meantime, the different microbial communities in the different parts of the Daqu block led to different physicochemical indices, such as the moisture content, acidity content, and temperature. Similarly, moisture and acidity contents in different parts of Daqu were also key environmental factors that could cause different microbial community distributions (Yan et al., 2013). Our study found that moisture and acidity contents in the central part of Daqu were significantly higher than those in the surface part of Daqu, and the counts of microorganisms were significantly different between the surface and central parts of Daqu. Li et al. (2019) also found that the moisture content was significantly different between the surface and central parts of Yanghe Daqu, and the different microbial communities were closely related to moisture content in Daqu. Therefore, it is of great significance to explore the relationship between the specific species of microbial community and physicochemical indices in Daqu for improving the quality and yield of Chinese special-flavor Baijiu.
We used culture-dependent and Illumina MiSeq sequencing methods to determine the specific species of the microbial communities in the different parts of special-flavor Baijiu Daqu. Our results showed that compared with yeast and mold, bacteria were the dominant microbial community in special-flavor Baijiu Daqu. Firmicutes and Proteobacteria were predominant in Daqu at the phylum level. Sun et al. (2016) and Gan et al. (2019) also found that Firmicutes and Proteobacteria were the predominant microbial communities in Jiang-flavor Daqu and Nong-flavor Daqu. This result indicated that Firmicutes and Proteobacteria may be the main microbial communities in the fermentation process of Chinese Daqu, and this also explained why the special-flavor Baijiu has the partial flavor of Jiang-flavor Baijiu and Nong-flavor Baijiu (Zheng and Han, 2016).
In our study, two bacterial groups, Bacillales and Lactobacillales, were the dominant bacterial communities in special-flavor Daqu. However, we found an interesting phenomenon that Bacillales, including the genera of Bacillus, Kroppenstedtia, and Oceanobacillus, were only dominant in the central part of Daqu. This was consistent with the results of Illumina MiSeq sequencing. Li et al. (2013) also found that the members of the Bacillales family were dominant in the central part of Fen-Daqu. However, the reason for the high number of Bacillus genus in the central part of special-flavor Daqu needs further study in the future. Jin et al. (2019) found that Bacillus genus was the α-amylase producer, which could consecutively transform the raw material of Daqu into the key metabolite, such as pyruvate, and lead to further production of organic acid and ethanol in Jiang-flavor Baijiu Daqu. This may be one of the reasons that the acidity in the central of Daqu was significantly higher than that in the surface part of special-flavor Baijiu Daqu. We also found that B. licheniformis was the most abundant species in Bacillales. This observation was consistent with result of light-flavor Baijiu Daqu (Zhang et al., 2014). And Wang P. et al. (2017) found that B. licheniformis could not only increase the content of some flavor compounds, such as aromatic substances and pyrazines, but also influence the related fermentation–enzyme activity of Daqu, such as increasing the activity of α-amylase. However, we found that the activity of α-amylase in the surface part of Daqu was significantly higher than that in the central part of Daqu. The reason for this phenomenon may be that the number of fungal communities in the surface part of Daqu was higher than that in the central part of Daqu, and the ability of some fungal communities to produce the activity of α-amylase was significantly higher than that of B. licheniformis, resulting in the different α-amylase activities between the surface and central parts of Daqu. α-Amylase is a key enzyme that breaks down the starch of Daqu to produce reducing sugar and can provide energy and substrate for the growth of microorganisms and the formation of flavor compounds (Wu et al., 2009). The result of reducing sugar content also indirectly proved this conclusion. We found that the content of reducing sugar in the surface part of Daqu was significantly higher than that in the central part.
In our study, Lactobacillales mainly included Lactobacillus, Pediococcus, Enterococcus, Lactococcus, and Weissella genera, wherein Lactobacillus genus was the most abundant bacterial community. Moreover, a considerable number of LAB, including Enterococcus faecium, Pediococcus pentosaceus, Lactococcus plantarum, Lactococcus fermentum, and Weissella paramesenteroides were identified by the culture-dependent method. These bacterial communities also generally appeared in other Chinese traditional Daqus (Sun et al., 2016; Wang X.D. et al., 2017). Zou et al. (2018) found that the members of LAB were the main producers of lactic acid, which is subsequently found in the synthesis of ethyl lactate by esterification in Nong-flavor Daqu. These microorganisms could survive in high-acid concentration and control spoilage bacteria by secreting bacteriocin. Lactobacillales had multiple metabolic products, wherein lactic acid was its main metabolic product and the main basic substance for the formation of ethyl lactate and other flavor compounds. Moreover, lactic acid could reduce the harsh taste of Baijiu, enhance the mellow taste of the liquor body, prolong liquor aftertaste, and strengthen the sweetness of the Chinese Baijiu (Wang X.S. et al., 2017). Therefore, these bacterial communities were the crucial microbial communities in the Daqu. In addition, our study showed a higher abundance of Acetobacter genus in Daqu; Acetobacter genus occurred in the natural environment of several plants, such as fruits and grains, which could disintegrate the different types of sugars and alcohols to organic acids in oxygen as final substances by a special aerobic metabolism method (Sengun and Karabiyikli, 2011). Sun et al. (2016) found that Acetobacter genus play an important role during the brewing process of Nong-flavor Baijiu and could contribute to the accumulation of ethyl acetate, which has a significant positive effect on the flavor of Nong-flavor Baijiu. Therefore, Acetobacter genus in the Daqu may be an important bacterial community that provides the flavor compound during the special-flavor Baijiu fermentation process. In the current study, the relative abundance of Acetobacter genus was more than 50% in the surface part of Daqu but few in the central part of Daqu. This result may be due to Acetobacter genus being a type of aerobic bacteria. The lack of oxygen led to the reduction of Acetobacter genus in the central part of Daqu. Lactobacillus and Acetobacter genera are the main acid-producing bacteria in Daqu, and in our study, the abundance in the surface part of special-flavor Baijiu Daqu was significantly higher than that in the central part. On the contrary, the content of acidity in the surface part of Daqu was significantly lower than that of the central part. In addition to the Bacillus genus we mentioned earlier, which was also the main acid-producing bacteria, another reason may be due to the high esterification ability in the surface part of Daqu, which resulted in a large amount of organic acids being used as precursors to generate ester compounds (Liu et al., 2017). The result of esterification ability also indirectly proved this conclusion. However, the difference of acid content changes and the accumulation of flavor compounds in different parts of special-flavor Baijiu Daqu need further study in the future.
Our study also indicated that the diversity of fungal community was relatively low compared with the bacteria community, which was consistent with the previous studies in other traditional Daqu (Fan et al., 2018). The reason for this phenomenon may be that the bacteria were able to survive in harsher environments and exhibited better tolerance compared with fungus under conditions of high moisture content, acidity content, and temperature (Huang et al., 2017). In addition, the counts of yeast had the lowest level in both the surface and central parts of Daqu. Among them, S. cerevisiae was not detected by Illumina MiSeq sequencing, and only one strain was obtained and isolated by the culture-dependent method. This result was also observed by using the microbial composition analysis of Fen-Daqu (Zheng et al., 2012). The reason for this phenomenon may be that S. cerevisiae and other yeasts were vulnerable to environmental factors, such as low moisture (below 15%) and high temperature (over 50°C), resulting in the most yeast being killed in the process of medium–high temperature Daqu (Li et al., 2013; Gou et al., 2015). Therefore, only few yeasts could survive in the maturation of Daqu. In our study, the low moisture content (<13.14%) also proved this conclusion and may be the reason for this result. Wang and Xu (2015) found that the number of molds in Daqu was higher than that of yeast, because they had relatively high tolerance to low moisture content and could form spores to resist higher temperatures. However, the acidity content in the central part of Daqu was significantly higher than that in the surface part of Daqu, which was not suitable for the growth of molds, resulting in the number of molds in the central part of Daqu being lower than that in the surface part. Saccharomycetales and Eurotiales dominated the fungal communities in our study. The culture-dependent method was used to determine fungal communities which were composed of the Wickerhamomyces, Pichia, and Aspergillus genera. Likewise, the Pichia and Aspergillus genera were detected with a relative abundance of more than 50% in the surface and central parts of Daqu by Illumina MiSeq sequencing, respectively. Some studies have indicated that the Wickerhamomyces and Pichia genera were ester-producing yeasts with the high ability of saccharification and esterification. Pichia could generate the etherification of various alcohols, such as ethanol, geraniol, isoamyl alcohol, and 2-phenylethanol, resulting in increasing concentrations of esters with a fruity aroma, because alcohols and acids are key substrates for ester synthesis (You et al., 2016). Our study also found that the abundance of Wickerhamomyces and Pichia genera on the surface part of special-flavor Baijiu Daqu was significantly higher than that on the central part. At the same time, the results of saccharification and esterification ability in the surface part of special-flavor Baijiu Daqu were significantly higher than those of the central part also prove that they were undoubtedly important functional fungal communities in special-flavor Daqu.
Our study also found that Aspergillus was predominant in special-flavor Daqu, including A. varians, A. niger, and A. carbonarius, which were also consistent with a previous study on the fungal community of Maotai Daqu (Wang et al., 2008). Esterification ability is an important indicator of the ability in Daqu to produce flavor compounds. It refers to the ability of acid to combine with alcohol to remove one water molecule to form an ester (Kumar and Satyanarayana, 2009). Meantime, acid protease can decompose the protein in Daqu raw materials into small-molecule substances (polypeptides, free amino acids, and so on), which has a positive effect on the formation and quality of Chinese Baijiu flavor (Huang et al., 2017). Hu et al. (2017) found that a considerable number of Aspergillus genus could produce high esterification ability and acid protease activity, which is of great significance for improving the quality of Jiang-flavor Daqu. However, there was a significant difference in the distribution of Aspergillus genus between the surface and central parts of Daqu. The Aspergillus genus was mainly located in the central part of Daqu. However, our research found that the acid protease activity and esterification ability in the central part of Daqu were significantly lower than that in the surface part of Daqu. This may be because the partial metabolic mechanism of Aspergillus genus was inhibited in a highly acidic environment, resulting in the decreasing acid protease activity and esterification ability. However, the specific mechanism behind the decrease of acid protease activity and esterification ability needs further study. In other words, these results showed that the microbial community had a unique distribution in special-flavor Baijiu Daqu. Meantime, the distribution of the unique microbial community in special-flavor Baijiu Daqu also had a significant impact on the enzyme activity in different parts of Daqu.
Other bacteria, including K. pneumoniae, Salmonella enterica, and Micrococcus luteus, had low number in both the surface and central parts of Daqu. Tang et al. (2019) found that the rich LAB in the Xiaoqu could inhibit the growth of these bacteria by producing large amounts of lactic acid. Similarly, some fungus also was found in our study, such as Penicillium genus. They were common in traditional Chinese Daqus, and the Penicillium genus would reduce Baijiu yield and exhibit adverse effects on Baijiu quality (bitter taste of base Baijiu) (Zheng et al., 2011; Yan et al., 2013). Of course, these bacteria also were discovered in Daqu samples maybe because of external pollution. Li et al. (2013) found that these microorganisms could be killed during the distillation process and did not survive in the Baijiu with high alcohol concentration. Therefore, more information on the microbial community needs to be studied during the special-flavor Baijiu fermentation process.
All in all, the dominant microbial communities in different parts of Daqu had been detected by culture-dependent and Illumina MiSeq sequencing methods, such as Lactobacillus, Bacillus, Weissella, Pediococcus, and Acetobacter genera. However, the results obtained by different methods were slightly different, and culture-dependent and Illumina MiSeq sequencing methods still have their own advantages and disadvantages (Wang X.D. et al., 2017; Yang F. et al., 2019). The culture-dependent method cultivates the microorganisms in Daqu through different selective media to obtain pure strains and then amplifies and sequences specific fragments. The sequence information obtained is complete, and a large amount of information is carried in the fragments, so that each strain information can be observed more intuitively and quickly. However, the disadvantage is that the workload is large, only cultivable microorganisms can be identified, and the obtained Daqu microbial diversity is incomplete (Zheng et al., 2012). In our study, Kroppenstedtia and Oceanobacillus genera were not detected by the culture-dependent method. In contrast, we had obtained the existence of these microbial communities through the Illumina MiSeq sequencing method in Daqu, and they have a higher abundance compared to other microbial communities. Our study indicated that the results obtained on microbial community through the Illumina MiSeq sequencing method are more comprehensive; we have identified 54 genera and can determine the distribution and relative abundance of various genera in Daqu, thereby further determining the dominant microbial community in different parts of Daqu. The Illumina MiSeq sequencing method also has the advantages of being a simple and fast operation (Sun et al., 2016). However, its disadvantage is that it is easily restricted by various factors, such as genome extraction, sequence depth, and algorithms. Meantime, compared with the culture-dependent method, the results of Illumina MiSeq sequencing did not detect Penicillium genus, possibly due to its very low content or insufficient sequence information, which could not be classified. These results showed that we could effectively explain the microbial community changes in Chinese Baijiu fermentation industry based on the advantages of culture-dependent and Illumina MiSeq sequencing methods.
In this study, we used culture-dependent and Illumina MiSeq sequencing methods to analyze the microbial structure of special-flavor Daqu. We found that bacterial diversity was higher than fungal diversity in Daqu and that there were significant differences in microbial composition, distribution, and physicochemical indices between the surface and central parts of Daqu. These results could provide a certain reference for the production and utilization of special-flavor Daqu in the future. However, more information on the function microbial community in special-flavor Baijiu Daqu need to be studied for a better understanding of the relation between microbial community and physicochemical indices.
The original contributions presented in the study are publicly available. This data can be found here: NCBI, accession numbers and links in Supplementary Table 1.
YC and KL participated in all the experimental processes and wrote the initial manuscript. TL and RL made the figures and tables. YW proofread the revised manuscripts. GF and FZ provided the design and financial support of this study. All authors contributed to the article and approved the submitted version.
This project was supported by the Open Project Program of Beijing Advanced Innovation Center for Food Nutrition and Human Health (No. 20161016).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank Jiangxi Zhangshugong Wine and Spirits Co., Ltd. for Daqu sampling assistance and support.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2020.592421/full#supplementary-material
Du, H., and Xu, Y. (2012). Determination of the microbial origin of geosmin in Chinese liquor. J. Agric. Food Chem. 60, 2288–2292. doi: 10.1021/jf204648e
Fan, G. S., Sun, B. G., Fu, Z. L., Xia, Y. Q., Huang, M. Q., Xu, C. Y., et al. (2018). Analysis of physicochemical indices, volatile flavor components, and microbial community of a light-flavor Daqu. J. Am. Soc. Brew. Chem. 76, 209–218. doi: 10.1080/03610470.2018.1424402
Gan, S. H., Yang, F., Sahu, S. K., Luo, R. Y., Liao, S. L., Wang, H. Y., et al. (2019). Deciphering the composition and function profile of the microbial communities in Chinese Moutai liquor. Front. Microbiol. 10:1540. doi: 10.3389/fmicb.2019.01540
Gou, M., Wang, H. Z., Yuan, H. W., Zhang, W. X., Tang, X. Q., and Kid, J. (2015). Characterization of the microbial community in three types of fermentation starters used for Chinese liquor production. J. I. Brew. 121, 620–627. doi: 10.1002/jib.272
Han, K., Li, Z. F., Peng, R., Zhu, L. P., Zhou, T., Wang, L. G., et al. (2013). Extraordinary expansion of a Sorangium cellulosum genome from an alkaline milieu. Sci. Rep. 3, 2101–2109. doi: 10.1038/srep02101
Hu, D. B., Qiu, S. Y., Zhou, H. X., and Wang, X. D. (2017). Relationships among physiochemical indices and hydrolyzing enzyme systems and enzymes-produced-ability in Jiangxiang Daqu. Modern Food Sci. Technol. 33, 99–106. doi: 10.13982/j.mfst.1673-9078.2017.2.016
Huang, Y. H., Yi, Z. L., Jin, Y. L., Zhao, Y. G., He, G. Z., Liu, D. Y., et al. (2017). New microbial resource: microbial diversity, function and dynamics in Chinese liquor starter. Sci. Rep. 7, 14577–14586. doi: 10.1038/s41598-017-14968-8
Jin, Y., Li, D. Y., Ai, M., Tang, Q. X., Huang, J., Ding, X. F., et al. (2019). Correlation between volatile profiles and microbial communities: a metabonomic approach to study Jiang-flavor liquor Daqu. Food Res. Int. 121, 422–432. doi: 10.1016/j.foodres.2019.03.021
Kumar, P., and Satyanarayana, T. (2009). Overproduction of glucoamylase by a deregulated mutant of a thermophilic mould Thermomucor indicae-seudaticae. Appl. Biochem. Biotech. 158, 113–125. doi: 10.1007/s12010-008-8342-9
Li, K. M., Chen, Y. R., Liu, T., Deng, M. F., Xu, Z. W., Fu, G. M., et al. (2020). Analysis of spatial distribution of bacterial community associated with accumulation of volatile compounds in Jiupei during the brewing of special-flavor liquor. LWT Food Sci. Technol. 130, 1–10. doi: 10.1016/j.lwt.2020.109620
Li, L., Sun, Y., Yuan, Z. D., Kong, X. Y., Wao, Y., Yang, L. L., et al. (2015). Effect of microalgae supplementation on the silage quality and anaerobic digestion performance of Manyflower silvergrass. Bioresour. Technol. 189, 334–340. doi: 10.1016/j.biortech.2015.04.029
Li, P., Lin, W. F., Liu, X., Wang, X. W., Gan, X., Luo, L. X., et al. (2017). Effect of bioaugmented inoculation on microbiota dynamics during solid-state fermentation of Daqu starter using autochthonous of Bacillus, Pediococcus, Wickerhamomyces and Saccharomycopsis. Food Microbiol. 61, 83–92. doi: 10.1016/j.fm.2016.09.004
Li, R. Y., Yang, Y., Zhang, L. Y., Tan, H. D., Jiang, L., Guo, F. X., et al. (2019). Microbial growth/decline rules in the fermentation process of medium/high-temperature Daqu. Liquor Mak. Sci. Technol. 5, 89–93. doi: 10.13746/j.njkj.2018322
Li, X. R., Ma, E. B., Yan, L. Z., Meng, H., Du, X. W., and Quan, Z. X. (2013). Bacterial and fungal diversity in the starter production process of Fen liquor, a traditional Chinese liquor. J. Microbiol. 51, 430–438. doi: 10.1007/s12275-013-2640-9
Li, Z. M., Chen, L., Bai, Z. H., Wang, D. L., Gao, L. P., and Hui, B. D. (2015). Cultivable bacterial diversity and amylase production in two typical light-flavor Daqus of Chinese spirits. Front. Life Sci. 8, 264–270. doi: 10.1080/21553769.2015.1041188
Liu, P. L., Xiong, X. M., Wang, S., and Miao, L. H. (2017). Population dynamics and metabolite analysis of yeasts involved in a Chinese miscellaneous-flavor liquor fermentation. Ann. Microbiol. 67, 1–13. doi: 10.1007/s13213-017-1286-y
Mukherjee, A. K., Borah, M., and Rai, S. K. (2009). To study the influence of different components of fermentable substrates on induction of extracellular α-amylase synthesis by Bacillus subtilis DM-03 in solid-state fermentation and exploration of feasibility for inclusion of α-amylase in laundry detergent formulations. Biochem. Eng. J. 43, 149–156. doi: 10.1016/j.bej.2008.09.011
Ng, K. P., Ngeow, Y. F., Yew, S. M., Hassan, H., Soo-Hoo, T. S., Na, S. L., et al. (2012). Draft genome sequence of Daldinia eschscholzii isolated from blood culture. Eukaryot Cell. 11, 703–704. doi: 10.1128/EC.00074-12
Quigley, L., O’Sullivan, O., Beresford, T. P., Ross, R. P., Fitzgerald, G. F., and Cotter, P. D. (2012). High-throughput sequencing for detection of subpopulations of bacteria not previously associated with artisanal cheeses. Appl. Environ. Microb. 78, 5717–5723. doi: 10.1128/AEM.00918-12
Rousk, J., Baath, E., Brookes, P. C., Lauber, C. L., Lozupone, C., Caporaso, J. G., et al. (2010). Soil bacterial and fungal communities across a pH gradient in an arable soil. ISME J. 4, 1340–1351. doi: 10.1038/ismej.2010.58
Sengun, I. Y., and Karabiyikli, S. (2011). Importance of acetic acid bacteria in food industry. Food Control 22, 647–656. doi: 10.1016/j.foodcont.2010.11.008
Shi, J. H., Xiao, Y. P., Li, X. R., Ma, E. B., Du, X. W., and Quan, Z. X. (2009). Analyses of microbial consortia in the starter of Fen liquor. Lett. Appl. Microbiol. 48, 478–485. doi: 10.1111/j.1472-765X.2009.02554.x
Sun, W. N., Xiao, H. Z., Peng, Q., Zhang, Q. G., Li, X. X., and Han, Y. (2016). Analysis of bacterial diversity of Chinese Luzhou-flavor liquor brewed in different seasons by Illumina Miseq sequencing. Ann. Microbiol. 66, 1293–1301. doi: 10.1007/s13213-016-1223-5
Tang, Q. X., He, G. Q., Huang, J., Wu, C. D., Jin, Y., and Zhou, R. Q. (2019). Characterizing relationship of microbial diversity and metabolite in Sichuan Xiaoqu. Front. Microbiol. 10:696. doi: 10.3389/fmicb.2019.00696
Wang, C. L., Shi, D. J., and Gong, G. L. (2008). Microorganisms in Daqu: a starter culture of Chinese Maotai-flavor liquor. World J. Microb. Biot. 24, 2183–2190. doi: 10.1007/s11274-008-9728-0
Wang, H. Y., and Xu, Y. (2015). Effect of temperature on microbial composition of starter culture for Chinese light aroma style liquor fermentation. Lett. Appl. Microbiol. 60, 85–91. doi: 10.1111/lam.12344
Wang, P., Wu, Q., Jiang, X. J., Wang, Z. Q., Tang, J. L., and Xu, Y. (2017). Bacillus licheniformis affects the microbial community and metabolic profile in the spontaneous fermentation of Daqu starter for Chinese liquor making. Int. J. Food Microbiol. 250, 59–67. doi: 10.1016/j.ijfoodmicro.2017.03.010
Wang, X. D., Ban, S. D., Hu, B. D., Qiu, S. Y., and Zhou, H. X. (2017). Bacterial diversity of Moutai-flavour Daqu based on high-throughput sequencing method. J. I. Brew. 123, 138–143. doi: 10.1002/jib.391
Wang, X. S., Du, H., and Xu, Y. (2017). Source tracking of prokaryotic communities in fermented grain of Chinese strong-flavor liquor. Int. J. Food Microbiol. 244, 27–35. doi: 10.1016/j.ijfoodmicro.2016.12.018
Wu, Q., Chen, L. Q., and Xu, Y. (2013). Yeast community associated with the solid state fermentation of traditional Chinese Maotai-flavor liquor. Int. J. Food Microbiol. 166, 323–330. doi: 10.1016/j.ijfoodmicro.2013.07.003
Wu, X. H., Zheng, X. W., Han, B. Z., Vervoort, J., and Robert Nout, M. J. (2009). Characterization of Chinese liquor starter, “Daqu”, by flavor type with H-1 NMR-Based nontargeted analysis. J. Agr. Food Chem. 57, 11354–11359. doi: 10.1021/jf902881p
Yan, Z., Zheng, X. W., Chen, J. Y., Han, J. S., and Han, B. Z. (2013). Effect of different Bacillus strains on the profile of organic acids in a liquid culture of Daqu. J. I. Brew. 119, 78–83. doi: 10.1002/jib.58
Yang, F., Chen, L. Q., Liu, Y. F., Li, J. H., Wang, L., and Chen, J. (2019). Identification of microorganisms producing lactic acid during solid-state fermentation of Maotai flavour liquor. J. I. Brew. 125, 171–177. doi: 10.1002/jib.537
Yang, Y., Li, J. R., Jiang, L., Jia, Y. W., and Wang, Q. B. (2019). Study on the difference and change regulation of biochemicalindices between parts of medium and high temperature Daqu. Food Ferment. Industr. 9, 1–10. doi: 10.13995/j.cnki.11-1802/ts.020446
You, L., Wang, S., Zhou, R. P., Hu, X. Q., Chu, Y. J., and Wang, T. (2016). Characteristics of yeast flora in Chinese strong-flavoured liquor fermentation in the Yibin region of China. J. I. Brew. 122, 517–523. doi: 10.1002/jib.352
Zhang, X., Zhao, J., and Du, X. (2014). Barcoded pyrosequencing analysis of the bacterial community of Daqu for light-flavour Chinese liquor. Lett. Appl. Microbiol. 58, 549–555. doi: 10.1111/lam.12225
Zheng, X. W., and Han, B. Z. (2016). Baijiu, Chinese liquor: history, classification and manufacture. J. Ethn. Foods 3, 19–25. doi: 10.1016/j.jef.2016.03.001
Zheng, X. W., Tabrizi, M. R., Robert Nout, M. J., and Han, B. Z. (2011). Daqu-a traditional Chinese liquor fermentation starter. J. I. Brew. 117, 82–90. doi: 10.1002/j.2050-0416.2011.tb00447.x
Zheng, X. W., Yan, Z., Han, B. Z., Zwietering, M. H., Samson, R. A., Boekhout, T., et al. (2012). Complex microbiota of a Chinese “Fen” liquor fermentation starter (Fen-Daqu), revealed by culture-dependent and culture-independent methods. Food Microbiol. 31, 293–300. doi: 10.1016/j.fm.2012.03.008
Keywords: special-flavor Baijiu, Daqu, microbial community, physicochemical indices, Illumina MiSeq sequencing
Citation: Chen Y, Li K, Liu T, Li R, Fu G, Wan Y and Zheng F (2021) Analysis of Difference in Microbial Community and Physicochemical Indices Between Surface and Central Parts of Chinese Special-Flavor Baijiu Daqu. Front. Microbiol. 11:592421. doi: 10.3389/fmicb.2020.592421
Received: 10 August 2020; Accepted: 03 December 2020;
Published: 14 January 2021.
Edited by:
Abd El-Latif Hesham, Assiut University, EgyptReviewed by:
Yuanyuan Qu, Dalian University of Technology, ChinaCopyright © 2021 Chen, Li, Liu, Li, Fu, Wan and Zheng. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Fuping Zheng, emhlbmdmcEBidGJ1LmVkdS5jbg==; emhlbmdmcEB0aC5idGJ1LmVkdS5jbg==; Guiming Fu, ZnVndWltaW5nQG5jdS5lZHUuY24=
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.