
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol., 24 September 2020
Sec. Microbial Symbioses
Volume 11 - 2020 | https://doi.org/10.3389/fmicb.2020.562140
 Kirk J. Grubbs1†
Kirk J. Grubbs1† Frank Surup2†
Frank Surup2† Peter H. W. Biedermann3†
Peter H. W. Biedermann3† Bradon R. McDonald4†
Bradon R. McDonald4† Jonathan L. Klassen4†Caitlin M. Carlson4Jon Clardy2*
Jonathan L. Klassen4†Caitlin M. Carlson4Jon Clardy2* Cameron R. Currie4*
Cameron R. Currie4*Symbiotic microbes help a myriad of insects acquire nutrients. Recent work suggests that insects also frequently associate with actinobacterial symbionts that produce molecules to help defend against parasites and predators. Here we explore a potential association between Actinobacteria and two species of fungus-farming ambrosia beetles, Xyleborinus saxesenii and Xyleborus affinis. We isolated and identified actinobacterial and fungal symbionts from laboratory reared nests, and characterized small molecules produced by the putative actinobacterial symbionts. One 16S rRNA phylotype of Streptomyces (XylebKG-1) was abundantly and consistently isolated from the galleries and adults of X. saxesenii and X. affinis nests. In addition to Raffaelea sulphurea, the symbiont that X. saxesenii cultivates, we also repeatedly isolated a strain of Nectria sp. that is an antagonist of this mutualism. Inhibition bioassays between Streptomyces griseus XylebKG-1 and the fungal symbionts from X. saxesenii revealed strong inhibitory activity of the actinobacterium toward the fungal antagonist Nectria sp. but not the fungal mutualist R. sulphurea. Bioassay guided HPLC fractionation of S. griseus XylebKG-1 culture extracts, followed by NMR and mass spectrometry, identified cycloheximide as the compound responsible for the observed growth inhibition. A biosynthetic gene cluster putatively encoding cycloheximide was also identified in S. griseus XylebKG-1. The consistent isolation of a single 16S phylotype of Streptomyces from two species of ambrosia beetles, and our finding that a representative isolate of this phylotype produces cycloheximide, which inhibits a parasite of the system but not the cultivated fungus, suggests that these actinobacteria may play defensive roles within these systems.
Ambrosia beetles are a diverse group of insects (∼3,400 species) that cultivate fungi for food (Kirkendall et al., 1997; Farrell et al., 2001). Adult beetles generally bore into dead or dying trees, establishing a nest in the xylem. They actively inoculate the tunnel walls of the nest with spores of their mutualistic fungus, which grows and forms a layer of nutrient rich aleurioconidia [“ambrosial growth” (Beaver, 1989)] on the woody tissue of the host plant and serves as the sole source of nutrition for adults and developing beetle larvae. Ambrosia beetles vector their fungal mutualist in specialized structures called mycangia or mycetangia (Francke-Grosmann, 1956, 1975). Nutritional symbioses with fungi evolved at least eleven times independently in bark- and ambrosia beetles (Scolytinae and Platypodinae: Coleoptera; Farrell et al., 2001; Kirkendall et al., 2015). Specific ambrosia beetle species associate with specific ambrosia fungi (Batra, 1985; Beaver, 1989; Biedermann et al., 2013; Mayers et al., 2015), although some beetles appear to rely on a community of cultivars. Fungal cultivars from the scolytine weevil genera Xyleborus and Xyleborinus are mostly in the Ascomycota genera Ambrosiella (Ceratocystidaceae: Microascales) and Raffaelea (Ophiostomataceae: Ophiostomatales), which convergently evolved as beetle cultivars 30–60 million years ago (Cassar and Blackwell, 1996). Whereas many phloem-boring bark beetles gain extra nutrition through associations with fungi (e.g., Dendroctonus sp.), the xylem-boring ambrosia beetles that we study here are true fungus-farmers and obligately rely on their cultivars for food (Six, 2012; Biedermann et al., 2013). Nectria, Penicillium, and Aspergillus species are common associates of these beetles, but are typically found at low abundances within nests. They are regarded as competitors, parasites, or pathogens of the ambrosia beetle mutualism (Beaver, 1989; Biedermann et al., 2013).
In addition to ambrosia beetles, active farming of fungi also occurs in attine ants, and macrotermitine termites (Mueller et al., 2001, 2005; Aanen et al., 2002), and nutritional symbioses with fungi are widespread in insects (Dejean et al., 2005; Lauth et al., 2011; Leroy et al., 2011; Li et al., 2012; Toki et al., 2012; Hajek et al., 2013). Reliance on fungi by these insects exposes them to potential parasite pressure in the form of pathogens or competitors of their symbionts. For example, the fungal mutualist of attine ants is impacted by a specialized and potentially virulent fungal parasite (Currie et al., 1999; Currie, 2001). To help defend the cultivar from this parasite the ants use actinobacterial symbionts that produce antibiotics (Currie et al., 1999, 2006; Cafaro et al., 2011). A similar type of defensive symbiosis has been shown in the fungus-associated bark beetle Dendroctonus frontalis (Scott et al., 2008), and has been further suggested in the Mediterranean Pine Engraver bark beetle, Orthotomicus erosus (Human et al., 2017), as well as fungus-growing termites (Visser et al., 2012). Beyond defending fungal mutualists in agricultural associations, Actinobacteria are well adapted for insect dispersal [e.g., by desiccation-resistant, hydrophobic spores that stick to the surface of insects (Ruddick and Williams, 1972)] and fulfill different defensive capacities in other insect systems. Within antennal glands, Beewolves (Philanthus spp.) cultivate Actinobacteria that they transfer into brood cells and onto developing cocoons in order to prevent infection by a wide range of pathogens (Kaltenpoth et al., 2005). Actinobacteria and the antibiotic secondary metabolites they produce have been identified in several species of mud daubers (Oh et al., 2011; Poulsen et al., 2011). Furthermore, Actinobacteria have been isolated from several additional ant species (Seipke et al., 2012; Hanshew et al., 2015) and the gypsy moth (Hanshew et al., 2013).
The majority of insect defensive symbioses characterized have involved Actinobacteria, which is not surprising as Actinobacteria, especially Streptomyces, are well known producers of bioactive secondary metabolites (Flórez et al., 2015). Over 10,000 biologically active compounds have been identified from Actinobacteria, accounting for ∼45% of known microbial metabolites (Berdy, 2005). The phylum Actinobacteria is composed of Gram-positive bacteria and is one of the largest in the domain Bacteria. They are common soil microbes, and studies have also identified them as dominant community members in both freshwater (Sekar et al., 2003), and marine (Stach and Bull, 2005) habitats. As such, Actinobacteria are common microbiome constituents in many environments.
The fruit-tree pinhole borer Xyleborinus saxesenii Ratzeburg and the sugarcane shot-hole borer Xyleborus affinis Eichhoff colonize a wide variety of dying or recently dead tree species and are two of the most widespread ambrosia beetles worldwide (Schedl, 1963; Wood, 1982). Both species are facultatively eusocial depending on the viability of the wood resource and may settle the same nest for multiple generations: Adult offspring of a single, sib-mated foundress typically delay dispersal from their mothers’ tunnel system and help her with nest-hygiene, brood-care, and fungus-farming (Biedermann et al., 2009, 2011, 2012). Unique for Holometabola, ambrosia-beetle larvae also help in these cooperative tasks (Biedermann and Taborsky, 2011). The beetles’ activity and presence is necessary to maintain the fruiting and monocultures of their fungal cultivars (Beaver, 1989; Kirkendall et al., 1997). Both species are obligately dependent on Raffaelea ambrosia fungi (Roeper and French, 1981; Biedermann et al., 2013). Experiments in X. saxesenii showed that these cultures are protected against pathogenic fungi, such as Paecilomyces variotii and Fusarium merismoides, by larvae and adults in unknown ways (Biedermann and Taborsky, 2011) and it is possible that this defense involves “microbial helpers.”
Here we describe actinobacterial symbionts of X. saxesenii and X. affinis ambrosia beetles and explore their potential function in helping defend nests against an antagonistic fungus that was isolated from X. saxesenii. Using specific media, we isolated both Actinobacteria and fungi from laboratory reared nests. Actinobacterial isolates were characterized using 16S rRNA gene sequencing and tested for their ability to inhibit the growth of both mutualistic and parasitic fungal isolates from the same nests. Active compounds were isolated using bioassay-guided HPLC fractionation, chemically characterized using NMR spectroscopy and mass spectrometry, and further tested using bioassays to confirm growth inhibition activity. We sequenced the genome of one actinobacterial isolate (Grubbs et al., 2011) to confirm this strains’ phylogenetic identification, and identified a putative biosynthetic gene cluster for one of the characterized active compounds. Based on these results, we propose a mutualism between two species of ambrosia beetle and Actinobacteria, in which the bacterial symbiont produces cycloheximide to inhibit the growth of fungal competitors of the mutualistic cultivar fungus.
Xyleborus affinis and X. saxesenii females (∼20 each) were collected at the Southern Research Station in Pineville, LA (31°20′N, 92°24′W; 123 ft asl) with four ethanol (95%) baited Lindgren funnel traps in October 2007. Live beetles were placed in sterile plastic tubes with wet filter paper, stored at 4°C for up to 3 days, surface sterilized by immersing in 70% ethanol and deionized water for a few seconds, and then reared on artificial medium in glass tubes following Biedermann et al. (2009; Peer and Taborsky, 2004). Briefly, beetles were reared in sterile glass tubes (Bellco culture tubes 18 × 150 mm) filled with the standard medium for rearing xyleborine ambrosia beetles. A single female per glass tube was put onto the medium and usually started boring tunnels as if in wood (N = 20 tubes/species). About one third of these beetle colonies successfully established brood and these were maintained in the lab at room temperature with indirect sunlight.
We conducted targeted isolation of Actinobacteria from each of three X. saxesenii and X. affinis colonies in triplicate, aseptically sampling each tube three times in a biosafety cabinet. Briefly, the nest inside the solid rearing substrate was shaken out of the tube and tunnel-wall material, containing the layer of the mutualistic fungus (henceforth termed nest material), as well as individual beetles were collected with sterile metal probes/tweezers from the exposed tunnels. X. saxesenii nest material (0.05 g per sample), adults (2 pooled individuals per sample), and larvae (5 pooled individuals per sample) were sampled; only nest material (0.05 g per sample) and adults (2 pooled individuals per sample) were sampled from X. affinis. The amount of material was chosen such that all samples were of approximately equal volume. All samples were chosen at random and homogenized in 500 μL of autoclaved, 0.22 μm filtered, deionized water; 100 μL of each was evenly spread on dried chitin agar plates (15 g agar, 3 g chitin, 0.575 g K2HPO4, 0.375 g MgSO4 × 7H2O, 0.275 g KH2PO4, 0.0075 g FeSO4 × 7H2O, 0.00075 g MnCl2 × 4H2O, and 0.00075 g ZnSO4 × 7H2O dissolved in 750 mL deionized water) in duplicate and allowed to dry before wrapping with parafilm. Plates were incubated at 30°C for 3 weeks, after which colony forming units (CFUs) were counted and eight of each morphotype per plate were transferred to yeast malt extract agar (YMEA: 4 g yeast extract, 10 g malt extract, 4 g dextrose, and 15 g agar dissolved in 1 L). Colonies on YMEA plates were allowed to grow at 30°C for 2 weeks, visually inspected for morphological properties characteristic of Actinobacteria, and sub-cultured as necessary to obtain pure cultures. Three 0.05 g samples of artificial medium from tubes that were not inoculated with beetles were also plated in duplicate on YMEA without antibiotics to screen for contamination and possible presence of Actinobacteria in the beetle medium. All media used for actinobacterial isolation had filter-sterilized cycloheximide (0.05 g/L) and nystatin (10,000 units/mL) added after autoclaving and cooling to suppress fungal growth.
Fungal symbionts were isolated from three X. saxesenii nests, sampled three times each. X. affinis were not sampled for fungi. Nest material was scraped using a sterile metal probe and inoculated on potato dextrose agar plates (PDA; Difco, Sparks, MD, United States) with penicillin (0.05 g/L) and streptomycin (0.05 g/L) added after autoclaving and cooling to suppress bacterial growth, and incubated at 30°C for 1 week. During incubation, fast growing fungi were sub-cultured onto fresh PDA plates and the agar on which they grew was fully removed to prevent overgrowth of the entire original isolation plate. Two different fungi were obtained in pure culture by successive rounds of scraping a small amount of material from the edge of each colony and then plating on individual PDA plates.
The 16S rRNA gene was sequenced from eight Actinobacteria isolates obtained from both X. saxesenii and X. affinis for a total of 16. In an effort to maximize the possibility of capturing any phylogenetic diversity, and thereby discover if multiple species were present, the strains that were sequenced were chosen based on morphological differences rather than origin. Only two morphologies were observed with the only differences being that the spores of one morphology were slightly darker than the other. The 16S rRNA gene PCR primers used were the Actinobacteria-specific F243 (5′-GGATGAGCCCGCGGCCTA-3′) and R1378 (5′-CGGTGTGTACAAGGCCCGGGAACG-3′; Heuer et al., 1997), and in separate reactions the general bacterial primers pA (5′-AGAGTTTGATCCTGGCTCAG-3′) and pH (5′-AAGGAGGTGATCCAGCCCGCA-3′) to increase coverage length (Edwards et al., 1989). The cycle parameters used for each primer set was similar to those above except the annealing temperatures were 58°C and 54°C, respectively, and the elongation time was 95 s for primers pA and pH. Each PCR reaction was composed of 12.5 μL GoTaq master mix (Promega), 1 μL of template DNA, and 40 μM of each primer in a final volume of 25 μL. The EF-α and 18S rRNA genes were sequenced for two each of the isolated Raffaelea sulphurea and the putative antagonistic fungus Nectria spp. DNA was extracted as previously described (Cafaro and Currie, 2005). PCR primers NS1 (5′-GTAGTCATATGCTTGTCTC-3′) and NS4 (5′-CTTCCGTCAATTCCTTTAAG-3′) were used to amplify the 18S rRNA gene (White et al., 1990), using thermocycling parameters: 95°C for 2 min, 35 cycles of 95°C for 45 s, 42°C for 45 s, 72°C for 90 s, 72°C for 5 min and hold at 4°C. EF-α gene PCR primers 983F (5′-GCYCCYGGHCAYCGTGAYTTYAT) and 2218R (5′-ATGACACCRACRGCRACRGTYTG; Rehner and Buckley, 2005) were used with similar cycling parameters, except annealing temperature and elongation time were 55°C and 130 s.
PCR amplicons were purified by adding 0.8 μL ExoSap-IT (USB) to 2 μL of PCR product diluted in 5.25 μL of autoclaved deionized water and incubating this mixture at 37°C for 15 min and then at 80°C for 15 min. Sanger sequencing reactions contained: 1 μL BigDye Terminator v. 3.1 (Applied Biosystems), 1.5 μL Big Dye Buffer (Applied Biosystems) and 1 μL of 10 μM primer, and the entire cleaned amplicon solution. Sequencing PCR conditions were 95°C for 3 min, 35 cycles of 95°C for 20 s, 45°C for 30, 60°C for 4 min, 72°C for 7 min, and hold at 4°C. Excess dye terminators were removed using CleanSeq beads (Agencourt Biosciences) and samples were resuspended in 40 μL of sterile ddH2O and sequenced at the University of Wisconsin-Madison Biotechnology Center using an ABI 377 instrument (Applied Biosystems).
Growth inhibition assays were conducted between one S. griseus XylebKG-1 like strain (see section “Results”) isolated from each of the three X. saxesenii nests and both isolated fungal species by first inoculating the Actinobacterium in the center of a PDA plate and allowing it to grow for 2 weeks. A small amount of test fungus was then inoculated at the edge of this Petri plate and grown at 30°C for 2 weeks, after which a zone of inhibition (ZOI) was determined by measuring the shortest distance between the bacterium and the fungus.
All sequences were assembled using Bionumerics v6.5 (Applied Maths), searched against the GenBank Nucleotide Sequence Database (Benson et al., 2012) using BLAST (Altschul et al., 1990) to determine a preliminary identity, and then aligned in MEGA5 (Tamura et al., 2011) using MUSCLE (Edgar, 2004). 18S rRNA and EF-α sequences were aligned and trimmed individually and subsequently concatenated to increase phylogenetic resolving power. To ensure codons were not split by gaps, alignments were inspected in MEGA5 for consistent reading frames. Substitution models were chosen using the model selection module of MEGA5. Maximum likelihood phylogenies were inferred using 500 bootstrap replicates using MEGA5.
The genome of S. griseus XylebKG-1 has previously been sequenced (Grubbs et al., 2011) allowing us to generate a genome based phylogeny for this isolate. Proteins from all complete Streptomyces genomes were predicted using prodigal (Hyatt et al., 2010) for consistency and annotated using HMMer (Finn et al., 2011) models generated from KEGG (Ogata et al., 1999) gene families, of which 1,364 KEGG gene families were conserved in all genomes. For these gene families, the proteins with the highest HMMer bitscore from each genome were aligned using MAFFT (Katoh and Standley, 2013) and then converted to a nucleotide alignment. These alignments were concatenated and a phylogeny generated using RAxML (Stamatakis, 2006) with 100 rapid bootstraps.
The genomes of Streptomyces griseus subsp. griseus NBRC13350 [NC_010572.1] and S. griseus XylebKG-1 were aligned using progressive Mauve (Darling et al., 2010) with default parameters.
One- and two-dimensional NMR spectra were acquired using a Varian Inova spectrometer with a frequency of 600 MHz for 1H and 150 MHz for 13C nuclei. All compounds were dissolved in CD3OD. HPLC/MS analysis was performed on an Agilent 1200 Series HPLC/6130 Series mass spectrometer. High resolution spectra were obtained on a Waters Micromass Q-TOF Ultima ESI-TOF mass spectrometer.
Streptomyces griseus XylebKG-1 was cultivated on PDA plates for 5–10 days. Seed biomass for 1 L cultures was produced by adding 1 cm2 of a single mature PDA culture to three 500 mL Erlenmeyer flasks containing 85 mL modified yeast peptone maltose medium (YPM: 2 g/L yeast extract, 2 g/L bactopeptone, and 4 g/L D-mannitol). These were incubated at 28°C with shaking at 250 rpm for 48 h. Twenty-five ml of each culture was added to eight 1 L of YPM in 4 L Erlenmeyer flasks and incubated for 72 h at 28°C with shaking at 250 rpm. Supernatants and mycelia were processed separately after cultures were centrifuged at 7000 rpm for 30 min. Culture supernatants were adjusted to pH 6 and extracted twice with an equal volume of ethyl acetate. After evaporation in vacuo, residues were resuspended in 2 mL MeOH/H2O (8:2). Mycelia were lyophilized and each extracted with 50 mL acetone and 50 mL methanol. After evaporation in vacuo, crude extracts were resuspended in 2 mL methanol. Crude supernatant and mycelium extracts were tested for inhibition of Nectria sp.; only the extracts of the crude supernatant showed significant assay activity. Crude supernatant extracts were purified using a 2 g pre-packed C18 Sep-Pak resin and fractionated by eluting with a gradient of pure water to pure methanol. The pure water flow through and 10% methanol fractions exhibited the highest anti-Nectria activity. These fractions were therefore combined and fractionated by gel chromatography using Sephadex LH-20 with methanol as the mobile phase (column 60 × 2.5 cm). Active fractions were combined and subsequently purified by reversed-phase HPLC (Agilent 1100 Series HPLC system, Supelco Discovery HS C18 column, 250 × 10 mm, 2 mL/min). HPLC conditions used: 2 min 80% A, 20% B in 28 min to 100% B (A: water, B: methanol). The fraction most active against Nectria sp. was eluted from 17.5 and 18 min and contained 2.7 mg of cycloheximide (1).
Streptomyces griseus XylebKG-1 strain was cultivated on agar plates (300 mL) of YPM, PDA, oat media (20 g/L oat meal, 2.5 mL/L trace element solution, 3 g/L CaCl2⋅2 H2O, 1 g/L Fe(III)-citrate, 0.2 g/L MnSO4, 0.1 g/L ZnCl2, 25 mg/L CuSO4⋅5 H2O, 20 mg/L Na2B4O7⋅10 H2O, 4 mg/L CoCl2, and 10 mg/L Na2MoO4⋅2 H2O), soy mannitol media (20 g/L soy meal, 20 g/L mannitol), starch-glucose-glycerol media (10 g/L glucose, 10 g/L glycerol, 10 g/L starch, 2.5 mL/L cornsteep liquor, 5 g/L casein-peptone, 2 g/L yeast extract, 1 g/L NaCl, and 3 g/L CaCO3), ISP1 media (5 g/L pancreatic digest of casein, yeast extract 3 g/L), ISP2 media (4 g/L yeast extract, 10 g/L malt extract, and 4 g/L dextrose), and 1187 media [10 g/L starch, 2 g/L (NH4)2SO4, 1 g K2HPO4, 1 g/L MgSO4⋅7 H2O, 1 g/L NaCl, 2 g CaCO3, and 5 mL/L trace element solution] for 7 days at 30°C. All plates were extracted with ethyl acetate.
Naramycin B (0.7 mg, 2) was isolated from the crude extract of ISP1 cultivation by semipreparative HPLC (gradient 22% to 40% acetonitrile in 25 min, Supelco Discovery HS C18 column, 250 × 10 mm). Actiphenol (1.6 mg, 3) was isolated from the crude extract of 1187 cultivation by preparative HPLC (gradient 65% to 100% methanol in 25 min, column Phenomenex Luna C18 250 × 21 mm. Dihydromaltophilin (2.4 mg, 4) was isolated from extracts of PDA and 1187 cultivations by preparative HPLC (gradient 50 to 100% acetonitrile in 25 min, column Phenomenex Luna C18 250 × 21 mm), (gradient 80 to 100% acetonitrile in 25 min, column Phenomenex Luna C18 250 × 21 mm), and semi-preparative HPLC (gradient 35% to 50% acetonitrile in 25 min, column Supelco C18 250 × 8 mm).
Cycloheximide (1): white amorphous powder; 1H, 13C NMR were identical to a commercial sample obtained from Sigma-Aldrich (Jeffs and McWilliams, 1981), ESI-MS m/z [M + Na]+ 304.1, [M + H]+ 282.1, [M–H]– 280.1; HR-ESI-MS m/z 282.1718 [M + H]+ (calculated for C15H24NO4, 282.1700).
Naramycin B (2): white amorphous powder; 1H NMR data were consistent with those previously published for this metabolite (Berg et al., 1982); ESI-MS m/z [M + Na]+ 304.1, [M + H]+ 282.1, [M–H]– 280.1.
Actiphenol (3): white amorphous powder; 1H NMR data were consistent with those previously reported (Lakhvich et al., 1993); ESI-MS m/z [M + Na]+ 298.0, [M + H]+ 276.2, [M–H]– 274.1.
Dihydromaltophilin (4): white amorphous powder; 1H NMR and 13C NMR data were consistent with those previously reported (Graupner et al., 1997); ESI-MS m/z 513.3 [M + H]+, 511.3 [M-H]–; HR-ESI-MS 513.2964 [M + H]+ (calculated for C29H41N2O6 513.2965).
Minimum inhibitory concentrations were determined using Nectria sp. and R. sulphurea grown in liquid YPM for 3 days. Cultures were diluted 1:1000 with fresh YPM and 200 μL per well transferred into 96-well plates containing various amounts of commercial cycloheximide (100, 50, 20, 10, 5, 2, and 1 μg) and dihydromaltophilin (5, 2, 0.5, and 0.2 μg). These 96-well plates were incubated for 72 h at 30°C, after which the optical density was measured at 600 nm using a SpectraMax M5® Plate Reader. Naramycin B (2) and actiphenol (3) were inactive against both fungi up to concentrations of 10 μg/200 μL.
To determine the antifungal activity of cycloheximide (1) and dihydromaltophilin (4) in an agar plate dilution assay, Nectria sp. and R. sulphurea were grown in 20 mL liquid potato dextrose media for 7 and 21 days, respectively, at 30°C while shaking at 250 rpm. 1 mL of each culture was used to inoculate PDA plates. Paper disks (6 mm diameter) were soaked with solutions of 30, 2, and 0.2 μL of 1 and 20 μL, 2 μL of 4, both in methanol (concentration 1 mg/mL), dried, and applied to the surface of the agar plates. Plates were grown at 30°C for 5–7 days, when inhibition zones were recorded.
The biosynthetic cluster in the high quality draft S. griseus XylebKG-1 genome sequence [NZ_ADFC00000000.2] was predicted using antiSMASH v2.0 (Blin et al., 2013) and specific comparison to the previously published cycloheximide biosynthesis gene cluster from Streptomyces sp. YIM56141 (Yin et al., 2014). Gene cluster functional annotations were derived from the antiSMASH output, homologous annotations in the Streptomyces sp. cycloheximide biosynthesis gene cluster, and retrobiosynthetic logic.
At least one CFU having a morphology consistent with Actinobacteria was observed from 72% of adults, 33% of larvae, and 61% of nest samples from X. saxesenii (Table 1). On average, more Actinobacteria were cultured from nests than adults or larvae, with mean ± SEM CFUs/sample of 110 ± 56, 3.4 ± 1.4, and 1.1 ± 0.6, respectively (Figure 1). No growth of Actinobacteria was observed from media not inoculated with beetles (Table 1 and Figure 1). Thus, the medium serving as a possible source of bacterial isolates can be eliminated. All isolates had similar morphologies and growth patterns. Isolations from X. affinis nests and adults also resulted in CFUs of a single actinobacterial morphotype similar to that isolated from X. saxesenii.
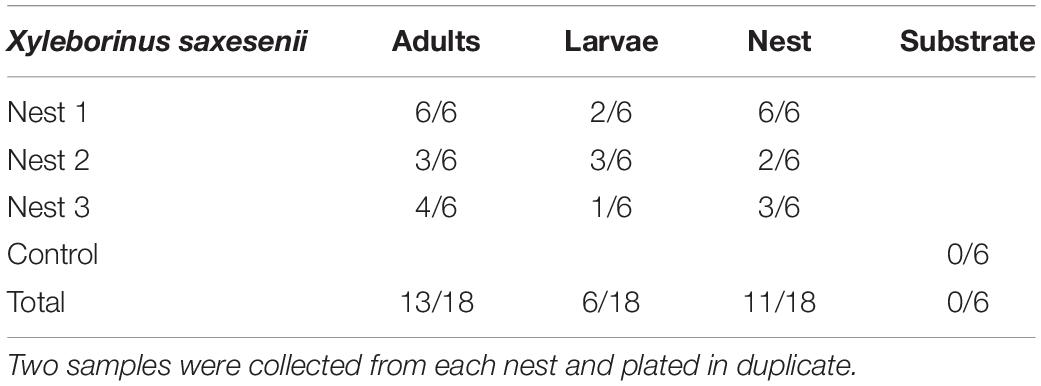
Table 1. Number of plates yielding actinobacterial growth.
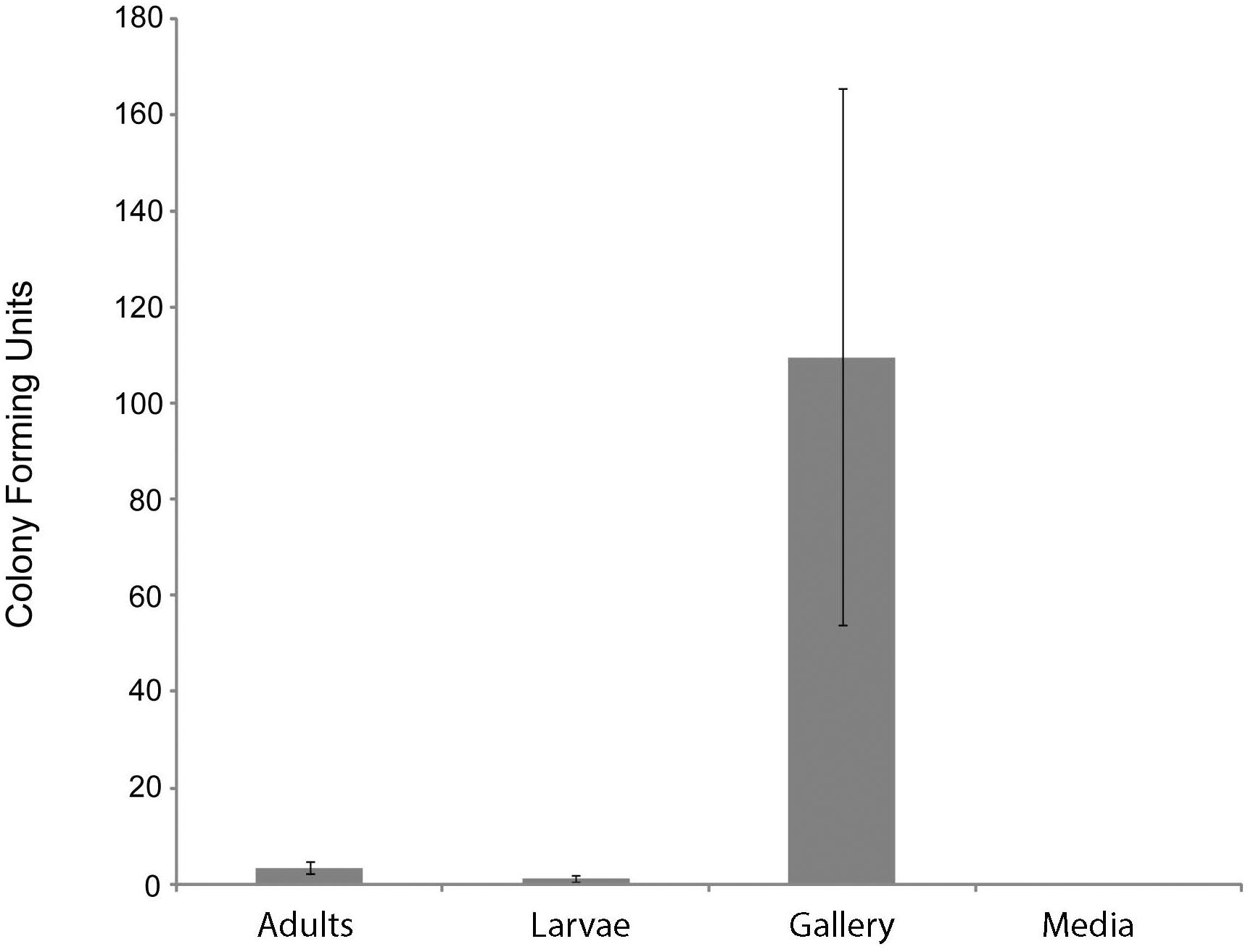
Figure 1. Number of Actinobacteria cultured from components of X. saxesenii nests. Means ± standard errors of the mean are displayed. See methods for culture conditions.
Eight representative Actinobacteria from various samples of the X. saxesenii and X. affinis systems were identified using 16S rRNA gene sequencing. The 16S rRNA gene sequences (1,123 bp) from all 16 were 100% identical and were most similar to that of S. griseus subsp. griseus NBRC13350 [NC_010572.1] when queried against the NCBI nr database using BLASTN, a result confirmed by phylogenetic analysis (Figure 2 and Supplementary Figure 1). A phylogeny constructed using the genome sequence for one of these strains, S. griseus XylebKG-1 (XylebKG-1) isolated from X. saxesenii [NZ_ADFC00000000.2] confirmed the close relationship between this strain and S. griseus subsp. griseus NBRC13350 [NC_010572.1], generating a tight clade in all bootstrap replicates produced (Figure 3). This is consistent with their high genomic similarity suggested previously using average nucleotide identity (Grubbs et al., 2011). Note that although the progressive Mauve algorithm aligns both genomes as one homology block (except for the extreme 5′and 3′ends), this block contains some regions of negligible sequence homology. These regions typically represent secondary metabolite biosynthetic gene clusters of unknown function that are not conserved between these two genomes (data not shown).
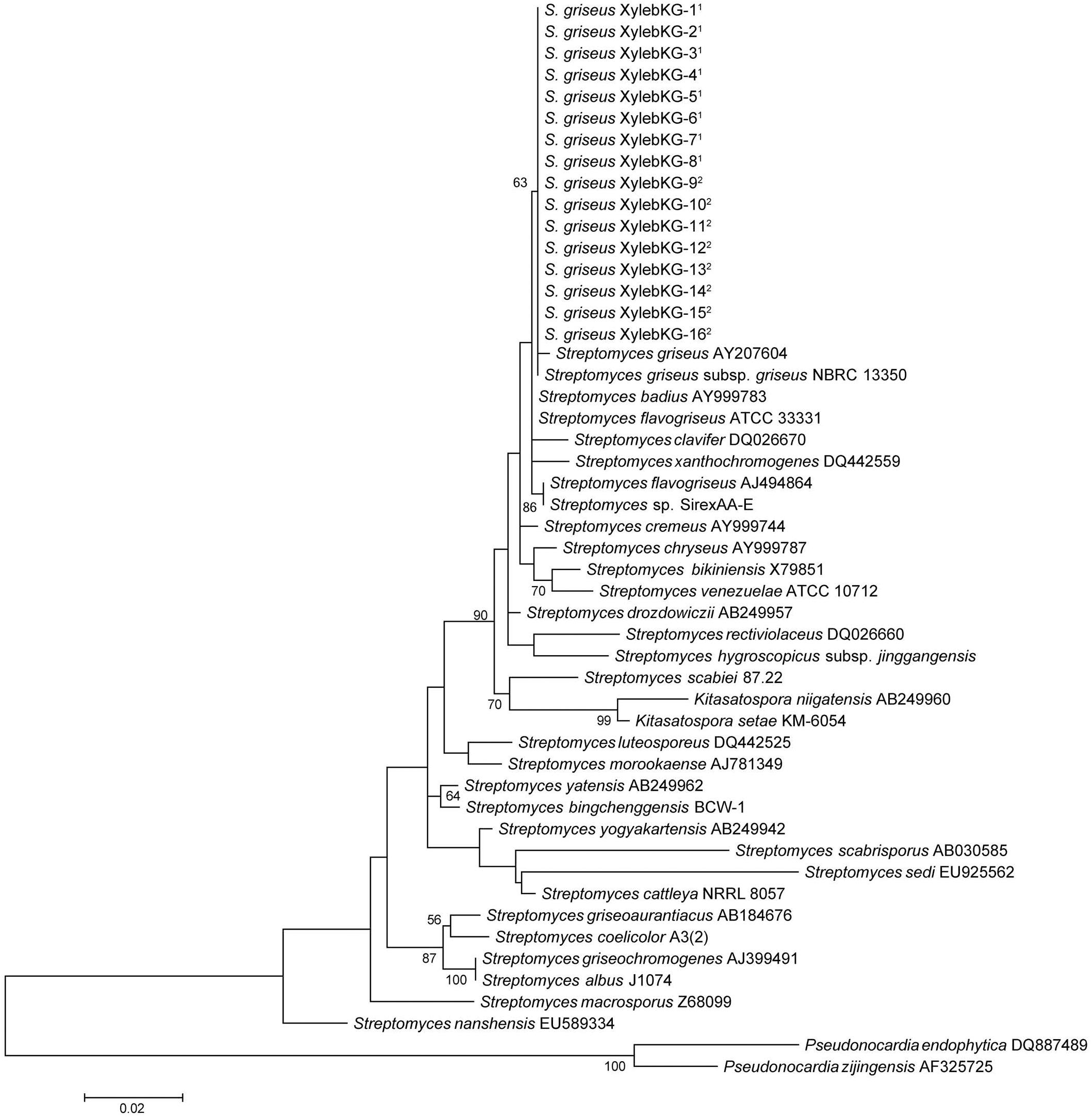
Figure 2. Maximum likelihood 16S phylogeny of the XylebKG-1 clade and its relatives, constructed using MEGA5. Molecular phylogeny based on 1,123 bp of 16S rRNA gene sequence. The Tamura 3-parameter substitution model was used with discrete gamma-distributed rate variation having 5 categories and a proportion of invariable sites, selected by MEGA5 as best fitting the data. The percent node conservation >50% in 500 bootstrap replicates is indicated, and the scale bar indicates the number of substitutions per site. MEGA5’s initial heuristic tree search was applied using an initial neighbor-joining tree of pairwise distances estimated using the Maximum Composite Likelihood method. 1- Indicates X. affinis origin. 2- Indicates X. saxesenii origin. Accession numbers in order from top to bottom of phylogeny: AY207604, AB184347, AY999783, NC_016114, DQ026670, DQ442559, AJ494864, NC_015953, AY999744, AY999787, X79851, NR_102857, AB249957, DQ026660, MF590728, NC_013929, AB249960, NR_112082, DQ442525, AJ781349, AB249962, NC_016582, AB249942, AB030585, EU925562, NC_017586, AB184676, NC_003888, AJ399491, CP004370, Z68099, EU589334, DQ887489, and AF325725.

Figure 3. Multilocus phylogeny constructed from 1,364 gene families conserved in all Streptomyces genomes analyzed. Alignments were done using MAFFT and the phylogeny generated using RAxML. Numbers above the branches based on 100 rapid bootstraps and the scale bar indicates the number of substitutions per site.
Our work supports a symbiosis between the S. griseus XylebKG -1clade and X. saxesenii ambrosia beetles. First, strains were consistently isolated having the same culture morphology from nests, larvae, and adults, and a random subset of these had 100% identical 16S rRNA sequences. Second, Actinobacteria were found to be very abundant within the nest material samples of the investigated X. saxesenii strains (approximately 110 Streptomyces CFUs per sample). Their recovery rate of 3.4 Streptomyces CFUs per adult individual is comparable with other established symbioses, like the Dendroctonus bark beetle system [average of 7.7 Streptomyces CFUs per individual (Scott et al., 2008)] or mud daubers [maximum average of 3.1 Streptomyces CFUs per individual (Poulsen et al., 2011)]. Third, Streptomyces are vectored by the beetles, likely within their bodies, as artificial medium was sterile and beetles were surface sterilized before being allowed to initiate nests. Fourth, the isolation of the XylebKG-1 Actinobacteria 16S phylotype from X. affinis further supports an association with ambrosia beetles, and suggests its potentially wider phylogenetic distribution within these insects.
Two fungi were consistently isolated from X. saxesenii nests. One type was identified as R. sulphurea using a dichotomous key (Batra, 1967) and confirmed by 18S rRNA and EF-α gene sequencing. This fungus has been repeatedly isolated from X. saxesenii and is known as the main cultivar of this beetle (Batra, 1967; Francke-Grosmann, 1975; Biedermann et al., 2013). The second fungus we isolated was identified as a close relative of the ascomycetous genus Nectria based on 18S rRNA and EF-α gene sequences and both BLAST and phylogenetic analyses (Figure 4). The consistent isolation of this Nectria sp. suggests that it is vectored by the ambrosia beetles. Nectria species are frequently isolated in low numbers from Scolytine beetles (Persson et al., 2009), and unpublished 18S rRNA 454-pyrosequencing data from Biedermann et al. suggest that they are commonly present in the nests of ambrosia beetles. Given that only Raffaelea and Ambrosiella species are producing nutritional fruiting structures for feeding ambrosia beetles and Nectria spp. are known pathogens of both insects (Mauchline et al., 2011) and trees (Ehrlich, 1934), it is likely a parasite of the system.
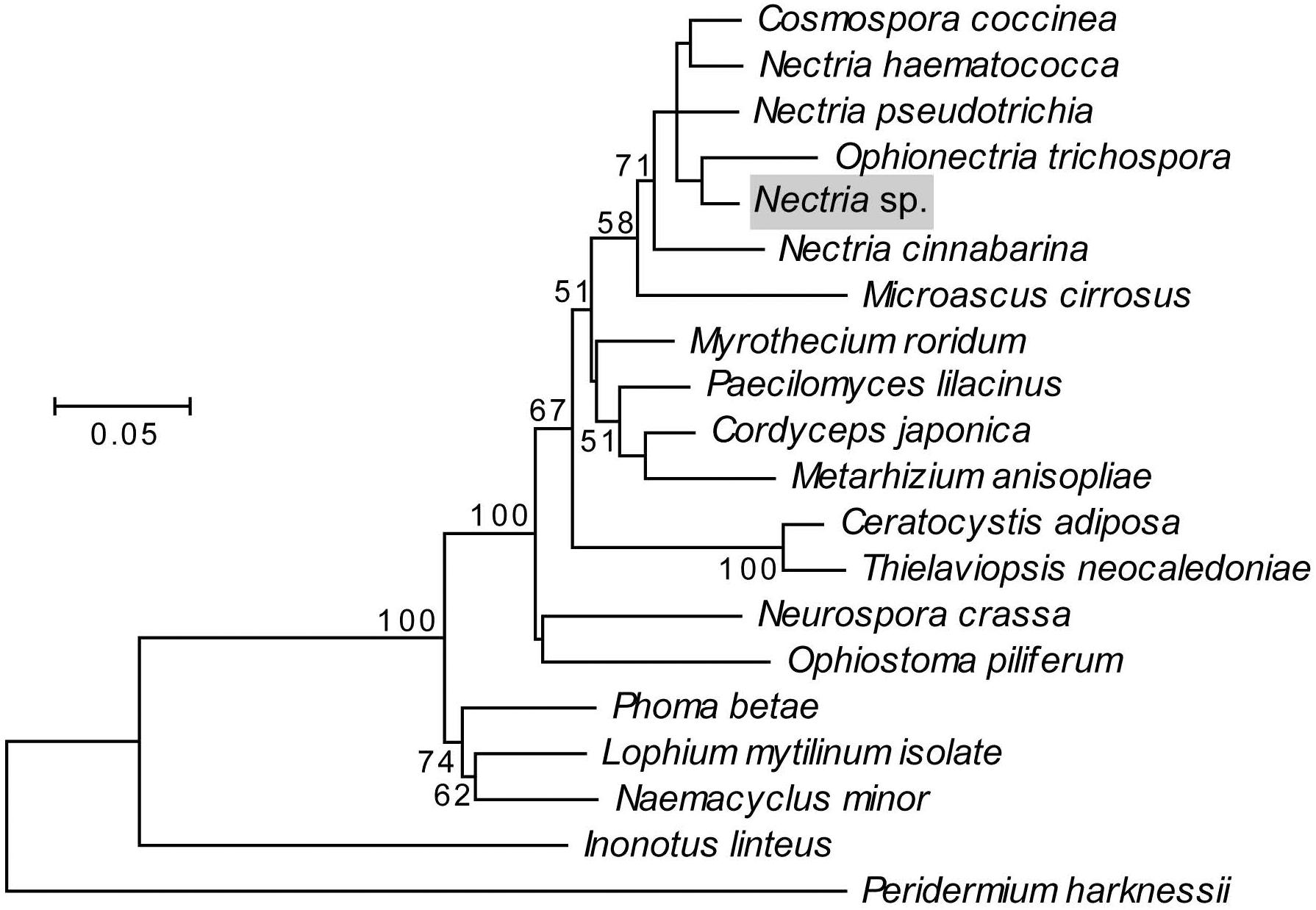
Figure 4. Maximum likelihood phylogeny of the fungal antagonist, highlighted in gray, constructed using concatenated 18S rRNA and EF-α genes in MEGA5. The Tamura-Nei substitution model was used with discrete gamma-distributed rate variation having 5 categories and a proportion of invariable sites, selected by MEGA5 as best fitting the data. The percent node conservation >50% in 500 bootstrap replicates is indicated, and the scale bar indicates the number of substitutions per site. MEGA5’s initial heuristic tree search was applied using an initial neighbor-joining tree of pairwise distances estimated using the Maximum Composite Likelihood method. Accession numbers for 18S sequences in order of phylogeny top to bottom: AY489702.1, AY489697.1, AY342011.1, NG_013128.1, Nectria sp., AB003949.1, M89994.1, AY489676.1, AY526475.2, AY245669.1, AB027337.1, EU984263.1, FJ176288.1, AY046271.1, DQ471003.1, EU754079.1, DQ678030.1, FJ176812.1, GU461973.1, and AY665785.1. Accession numbers for EF-α sequences in order of phylogeny top to bottom: HM484515.1, AY489624.1, HM534878.1, AF543779.1, Nectria sp., AF543785.1, HM569658.1, AY489603.1, GU979990.1, DQ522330.1, AY445082.1, HM569644.1, HM569649.1, XM_959775.2, DQ677926.1, DQ471074.1, GU349075.1, FJ238400.1, GU461936, and DQ234567.
To explore the potential that S. griseus XylebKG-1 function as defensive symbionts of X. saxesenii, S. griseus XylebKG-1’s ability to inhibit the growth of R. sulphurea and Nectria sp. isolated from this host was examined. Whereas S. griseus XylebKG-1 only marginally inhibited the growth of R. sulphurea (average zone of inhibition = 0.52 mm; Figure 5), it significantly inhibited the growth of Nectria sp. (average zone of inhibition = 26.2 mm; Figure 5). The strength of inhibition significantly differed between these fungi (t-test, P = 1.03E-27, n = 29, and n = 30 for Nectria sp. and R. sulphurea bioassays, respectively).
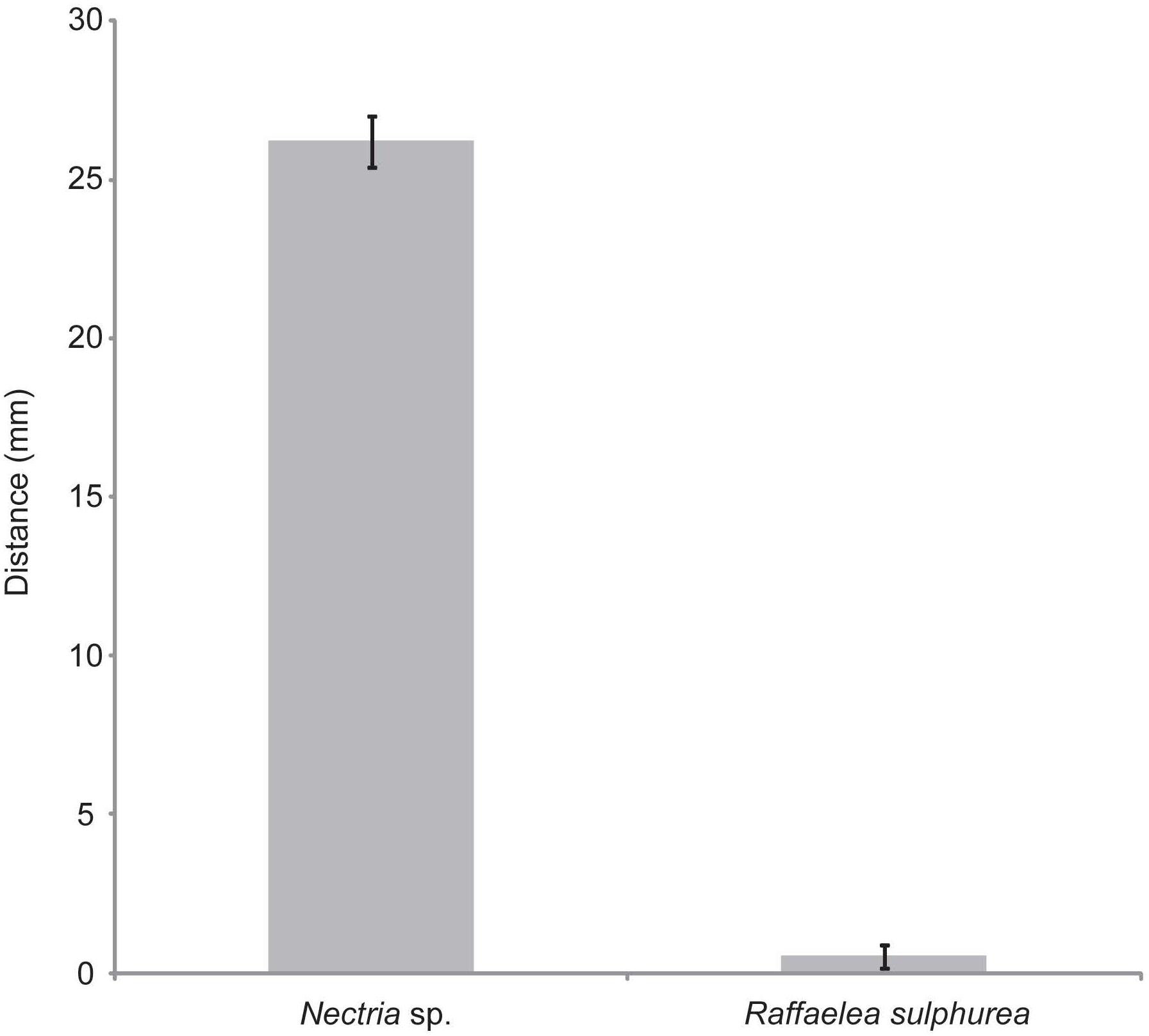
Figure 5. Plate bioassays of S. griseus XylebKG-1 versus Nectria sp. and R. sulphurea. See methods for assay conditions and media used. Average values are shown from 29 and 30 trials (Nectria sp. and R. sulphurea, respectively), with error bars representing standard error.
Cultivation in liquid yeast peptone media led to bioactivity guided isolation of cycloheximide (1) using Nectria sp. as the indicator organism. HR-ESI-MS provided the molecular formula C15H23NO4; 1H and 13C NMR data solely matched to the known compound cycloheximide (1) (Figure 6). In addition, the isolated metabolite showed the same retention time, UV spectrum and ESI-MS pattern as a commercially acquired cycloheximide standard. Furthermore, 1 was produced by S. griseus XylebKG-1 cultivated on agar plates of 8 different media (yeast peptone maltose, potato dextrose, oat, soy mannitol, starch glycerol glucose, ISP1, ISP2, and 1187 media), consistent with a robust synthesis of compound 1 under diverse growth conditions. In addition to 1, two byproducts were isolated by preparative HPLC from a culture of S. griseus XylebKG-1 in ISP1 media and identified as naramycin B (2) and actiphenol (3) (Jeffs and McWilliams, 1981; Berg et al., 1982; Lakhvich et al., 1993).
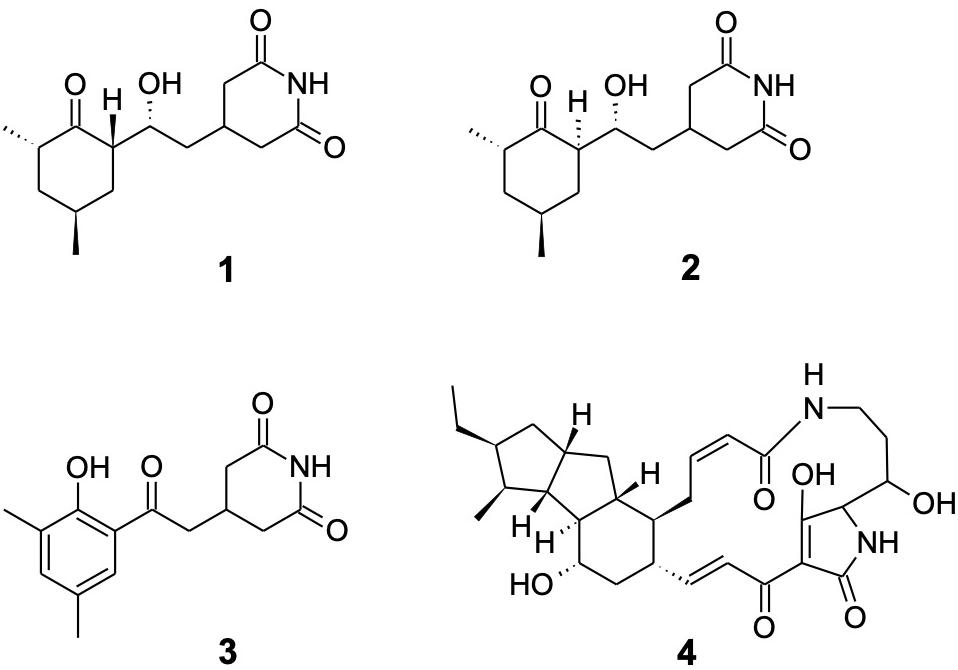
Figure 6. Metabolites isolated from S. griseus XylebKG-1. Cycloheximide (1); naramycin B (2); actiphenol (3); and dihydromaltophilin (4).
Because the alteration of growth media frequently results in a substantially changed metabolite pattern, a switch in growth media can be utilized to explore the metabolic potential of bacterial strains. In the case of S. griseus XylebKG-1, cultivation in PD and ISP4 resulted in the biosynthesis of an additional antifungal metabolite. After isolation by preparative HPLC, the molecular formula C29H40N2O6 (determined by HR-ESI-MS) and NMR data identified the compound as dihydromaltophilin (4; Graupner et al., 1997).
In plate bioassays, cycloheximide inhibited the isolated Nectria sp. (zones of inhibition: 30 μl, 44 mm; 2 μl, 18 mm; and 0.2 μg, 9 mm) but not R. sulphurea (no inhibition observed). Liquid culture assays confirmed this result, indicating a minimum inhibitory concentration of cycloheximide toward Nectria sp. of 0.02 mM. R. sulphurea grew in all test conditions (cycloheximide concentration up to 2.7 mM), although it did exhibit slower growth at higher concentrations of cycloheximide (data not shown). Dihydromaltophilin similarly inhibited both Nectria sp. and R. sulphurea (zones of inhibition: 20 μl, 20/22 mm; 2 μl, 12/13 mm). Naramycin-B and actiphenol were non-inhibitory under all conditions tested.
A putative cycloheximide biosynthetic gene cluster was identified in the S. griseus XylebKG-1 genome using antiSMASH v2.0 (Blin et al., 2013) based on its homology to the cycloheximide biosynthetic cluster from Streptomyces sp. YIM56141 (GenBank accession [JX014302.1]; Yin et al., 2014), and its being consistent with retrobiosynthetic logic (data not shown). All proteins predicted in the gene cluster from Streptomyces sp. YIM56141 were present in the S. griseus XylebKG-1 genome, except that the polyketide synthase cheE homolog was annotated as two separate genes in S. griseus XylebKG-1 (labeled chxE1 and chxE2 in Figure 7). This entire genomic region is conserved in the genome of S. griseus subsp. griseus NBRC13350 [NC_010572.1]. BLAST searches using dihydromaltophilin biosynthetic cluster genes previously identified by Yu et al. (2007) [EF028635], revealed highly similar protein sequences with the top blast hits of the first six genes (expect values: 0.0, 0.0, 0.0, 0.0, 0.0, and 2e-105) in the cluster being in the same relative positions. Similar to the pine beetle symbiont Streptomyces sp. SPB78 in which no homologs were found for ferredoxin-like and arginase proteins (Blodgett et al., 2010), BLAST indicated top hits for these genes in separate areas of the genome with much higher expect values (expect values: 8e-009 and 2e-010 respectively). Evolutionary conservation of these clusters from before the adaptation of S. griseus XylebKG-1 as a symbiont of ambrosia beetles suggests the potential for similar and/or complementary regulation and activity of the metabolites that they produce.
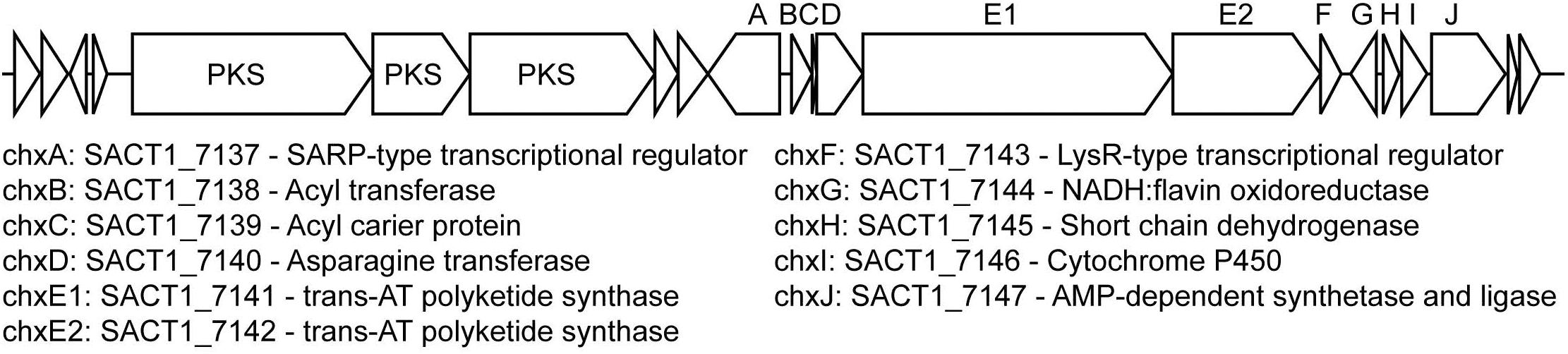
Figure 7. Putative cycloheximide biosynthetic gene cluster in S. griseus XylebKG-1, predicted using antiSMASH v2.0, homology to a cycloheximide biosynthetic gene cluster from Streptomyces sp. YIM56141, and retrobiosynthetic logic. Letters above the cluster indicate gene names, with corresponding locus names and annotations indicated below. An upstream biosynthetic gene cluster predicted to be unrelated to cycloheximide biosynthesis is also shown, with its polyketide synthase (PKS) genes indicated.
Our findings are consistent with S. griseus XylebKG-1 being a potential defensive symbiont of ambrosia beetles. Cycloheximide’s specific inhibition of the antagonist Nectria sp., but not the mutualist R. sulphurea, supports a defensive role for its production by XylebKG. This parallels similar results obtained for fungus-growing ants and D. frontalis, where their associated Actinobacteria inhibited the growth of a fungal parasite and not the fungal mutualist (Currie et al., 2006; Scott et al., 2008). Cycloheximide inhibits protein synthesis in eukaryotic cells and as such is toxic to most eukaryotes (Blodgett et al., 2010), including fungi. Interestingly, species in the fungal order Ophiostomatales (including Raffaelea spp.), are known to largely be resistant to cycloheximide (Harrington, 1981; Human et al., 2017). Although not determined here, X. affinis also cultivates a cycloheximide-insensitive Raffaelea sp. as food (Roeper and French, 1981). The association of two cycloheximide-insensitive Raffaelea spp. mutualists with isolates closely related to S. griseus XylebKG-1 in two ambrosia beetle species further supports the role of XylebKG-1 isolates as defensive symbionts, as evolutionarily stable relationships are expected to promote such complementarity. In contrast, dihydromaltophilin production inhibits the growth of both Nectria sp. and R. sulphurea. The production of dihydromaltophilin under only a few growth conditions could suggest that it does not have an active role in the ambrosia beetle system, but rather is a remnant from before S. griseus XylebKG-1 became associated with these beetles. In this regard it is worth noting that dihydromaltophilin analogs were found at low production levels in the D. frontalis system (Blodgett et al., 2010). Alternatively, dihydromaltophilin production may be regulated to avoid inhibition of R. sulphurea, or selected for activity versus other organisms not considered in this study.
In this study we consistently isolated a single Streptomyces morphotype and phylotype from both X. saxesenii and X. affinis that inhibited the growth of the parasitic fungus Nectria sp., but not the mutualistic R. sulphurea, likely via the production of cycloheximide. Its ubiquity suggests that XylebKG-1 may be a defensive mutualist of these ambrosia beetles that inhibits the growth of all but a few fungi, including its mutualistic fungal food source. Future studies should include natural nests collected from a wider range of species and geographies to establish the breadth and prevalence of XylebKG-1 in bark and ambrosia beetle nests (Human et al., 2017). These studies should also include greater phylogenetic power as 16S rRNA gene analyses is not sufficient to resolve species within the S. griseus clade (Rong and Huang, 2010). Cycloheximide should be assayed in vivo to confirm the relevance of its in vitro activity, and any other compounds also produced determined in vivo (e.g., those produced by the biosynthetic gene cluster adjacent to the cycloheximide cluster), if they exist. Furthermore, the presence and activity of XylebKG-1 may also vary during beetle development, e.g., cycloheximide may be used to clear new nests of contaminating fungus in preparation for the agricultural symbiont. The antibiosis of cycloheximide includes a large non-specific range of fungi and as such other fungal symbionts, aside from Nectria sp., may also be inhibited. We have identified a putative defensive Actinobaterium and an antagonistic fungal symbiont in two ambrosia beetles, potentially expanding the interactions from bipartite to quadripartite.
The original contributions presented in the study are publicly available. This data can be found at NCBI under accession numbers MT853219, MT853258–MT853261.
Most biological experiments were carried out by KJG, most chemical experiments and bioassays by FS. KJG, CRC, and PHWB wrote the manuscript with support from JC, JLK, BRM, and FS. KJG, PHWB, JLK, FS, and CMC analyzed datasets and helped with figure design. CRC and JC helped supervise the project. All authors contributed to the article and approved the submitted version.
This work was supported by a grant from the National Institute for Health GM09634 and U19 AI10967 (to JC and CRC) and a National Institute of Food and Agriculture, U.S. Department of Agriculture grant, under identification number WISO1321 (KJG and CRC). Funding to PHWB was provided by a Marie Curie Intra-European Fellowship (IEF; Project Number 626279) and by the German Research Foundation (DFG; Emmy Noether grant number BI 1956/1-1). FS was funded by a German Academic Exchange Service (DAAD) postdoctoral grant. BRM was supported by the Molecular Biosciences Training Grant, NIH National Research Service Award T32 GM07215. JLK was funded by a postdoctoral fellowship from the National Science and Engineering Research Council (Canada).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2020.562140/full#supplementary-material
Supplementary Methods | Additional details concerning analytical chemistry methods.
Supplementary Figure 1 | Updated Maximum likelihood 16S phylogeny of the XylebKG-1 clade and its relatives.
Supplementary Figure 2 | 1H spectrum of cycloheximide (CD3OD).
Supplementary Figure 3 | 1H spectrum of naramycin B (CD3OD).
Supplementary Figure 4 | 1H spectrum of dihydromaltophilin (CD3OD).
Supplementary Figure 5 | HMBC spectrum (600 MHz, methanol-d4) of cycloheximide (1).
Aanen, D. K., Eggleton, P., Rouland-Lefevre, C., Guldberg-Frøslev, T., Rosendahl, S., and Boomsma, J. J. (2002). The evolution of fungus-growing termites and their mutualistic fungal symbionts. Proc. Natl. Acad. Sci. U.S.A. 99, 14887–14892. doi: 10.1073/pnas.222313099
Altschul, S. F., Gish, W., Miller, W., Myers, E. W., and Lipman, D. J. (1990). Basic local alignment search tool. J. Mol. Biol. 215, 403–410.
Batra, L. R. (1967). Ambrosia fungi: a taxonomic revision, and nutritional studies of some species. Mycologia 59, 976–1017. doi: 10.2307/3757271
Batra, L. R. (1985). Ambrosia beetles and their associated fungi: research trends and techniques. Proc. Plant Sci. 94, 137–148.
Beaver, R. A. (1989). “Insect-fungus relationships in the bark and ambrosia beetles,” in Insect-Fungus Interactions, eds N. Wilding, N. M. Collins, P. M. Hammond, and J. F. Webber (London: Academic), 121–143. doi: 10.1016/b978-0-12-751800-8.50011-2
Benson, D. A., Cavanaugh, M., Clark, K., Karsch-Mizrachi, I., Lipman, D. J., Ostell, J., et al. (2012). GenBank. Nucleic Acids Res. 41, D36–D42.
Berdy, J. (2005). Bioactive microbial metabolites. J. Antibiot. 58, 1–26. doi: 10.1007/978-1-4757-9522-6_1
Berg, D., Schedel, M., Schmidt, R. R., Ditgens, K., and Weyland, H. (1982). Naramycin B, an antibiotic from Streptomyces griseus strain 587 with herbicidal properties-fermentation, isolation, and identification. Z. Naturforsch. C 37, 1100–1106. doi: 10.1515/znc-1982-11-1210
Biedermann, P. H., Klepzig, K. D., and Taborsky, M. (2009). Fungus cultivation by ambrosia beetles: behavior and laboratory breeding success in three xyleborine species. Environ. Entomol. 38, 1096–1105. doi: 10.1603/022.038.0417
Biedermann, P. H., Klepzig, K. D., and Taborsky, M. (2011). Costs of delayed dispersal and alloparental care in the fungus-cultivating ambrosia beetle Xyleborus affinis Eichhoff (Scolytinae: Curculionidae). Behav. Ecol. Sociobiol. 65, 1753–1761. doi: 10.1007/s00265-011-1183-5
Biedermann, P. H., Klepzig, K. D., Taborsky, M., and Six, D. L. (2013). Abundance and dynamics of filamentous fungi in the complex ambrosia gardens of the primitively eusocial beetle Xyleborinus saxesenii Ratzeburg (Coleoptera: Curculionidae. Scolytinae). FEMS Microbiol. Ecol. 83, 711–723. doi: 10.1111/1574-6941.12026
Biedermann, P. H., Peer, K., and Taborsky, M. (2012). Female dispersal and reproduction in the ambrosia beetle Xyleborinus saxesenii Ratzeburg (Coleoptera; Scolytinae). Mitt. Dtsch. Ges Allg. Angew. Entomol. 18, 231–235.
Biedermann, P. H., and Taborsky, M. (2011). Larval helpers and age polyethism in ambrosia beetles. Proc. Natl. Acad. Sci. U.S.A. 108, 17064–17069. doi: 10.1073/pnas.1107758108
Blin, K., Medema, M. H., Kazempour, D., Fischbach, M. A., Breitling, R., Takano, E., et al. (2013). antiSMASH 2.0—a versatile platform for genome mining of secondary metabolite producers. Nucleic Acids Res. 41, W204–W212.
Blodgett, J. A., Oh, D. C., Cao, S., Currie, C. R., Kolter, R., and Clardy, J. (2010). Common biosynthetic origins for polycyclic tetramate macrolactams from phylogenetically diverse bacteria. Proc. Natl. Acad. Sci. U.S.A. 107, 11692–11697. doi: 10.1073/pnas.1001513107
Cafaro, M. J., and Currie, C. R. (2005). Phylogenetic analysis of mutualistic filamentous bacteria associated with fungus-growing ants. Can. J. Microbiol. 51, 441–446. doi: 10.1139/w05-023
Cafaro, M. J., Poulsen, M., Little, A. E., Price, S. L., Gerardo, N. M., Wong, B., et al. (2011). Specificity in the symbiotic association between fungus-growing ants and protective Pseudonocardia bacteria. Proc. R. Soc. : Biol. Sci. 278, 1814–1822. doi: 10.1098/rspb.2010.2118
Cassar, S., and Blackwell, M. (1996). Convergent origins of ambrosia fungi. Mycologia 88, 596–601. doi: 10.2307/3761153
Currie, C. R. (2001). Prevalence and impact of a virulent parasite on a tripartite mutualism. Oecologia 128, 99–106. doi: 10.1007/s004420100630
Currie, C. R., Mueller, U. G., and Malloch, D. (1999). The agricultural pathology of ant fungus gardens. Proc. Natl. Acad. Sci. U.S.A. 96, 7998–8002. doi: 10.1073/pnas.96.14.7998
Currie, C. R., Poulsen, M., Mendenhall, J., Boomsma, J. J., and Billen, J. (2006). Coevolved crypts and exocrine glands support mutualistic bacteria in fungus-growing ants. Science 311, 81–83. doi: 10.1126/science.1119744
Darling, A. E., Mau, B., and Perna, N. T. (2010). progressiveMauve: multiple genome alignment with gene gain, loss and rearrangement. PLoS One 5:e11147. doi: 10.1371/journal.pone.0011147
Dejean, A., Solano, P. J., Ayroles, J., Corbara, B., and Orivel, J. (2005). Arboreal ants build traps to capture prey. Nature 434, 973–973. doi: 10.1038/434973a
Edgar, R. C. (2004). MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 32, 1792–1797. doi: 10.1093/nar/gkh340
Edgar, R. C. (2004). MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 32, 1792–1797. doi: 10.1093/nar/gkh340
Edwards, U., Rogall, T., Bl cker, H., Emde, M., and B ttger, E. C. (1989). Isolation and direct complete nucleotide determination of entire genes. Characterization of a gene coding for 16S ribosomal RNA. Nucleic Acids Res. 17, 7843–7853.
Ehrlich, J. (1934). The beech bark disease: a nectria disease of fagus, following Cryptococcus fagi (Baer.). Can. J. Res. 10, 593–692. doi: 10.1139/cjr34-070
Farrell, B. D., Sequeira, A. S., O’Meara, B. C., Normark, B. B., Chung, J. H., and Jordal, B. H. (2001). The evolution of agriculture in beetles (Curculionidae: Scolytinae and Platypodinae). Evolution 55, 2011–2027. doi: 10.1111/j.0014-3820.2001.tb01318.x
Finn, R. D., Clements, J., and Eddy, S. R. (2011). HMMER web server: interactive sequence similarity searching. Nucleic Acids Res. 39(Suppl._2), W29–W37.
Flórez, L. V., Biedermann, P. H., Engl, T., and Kaltenpoth, M. (2015). Defensive symbioses of animals with prokaryotic and eukaryotic microorganisms. Natural Prod. Rep. 32, 904–936. doi: 10.1039/c5np00010f
Francke-Grosmann, H. (1956). Hautdrüsen als träger der pilzsymbiose bei ambrosiakäfern. Z. Morpholo. Oekol. Tiere 45, 275–308. doi: 10.1007/bf00430256
Francke-Grosmann, H. (1975). Zur epizoischen und endozoischen ubertragung der symbiotischen pilze des ambrosiakafers Xyleborus saxeseni (Coleoptera: Scolytidae). Entomol. German. 23–24, 279–292.
Graupner, P. R., Thornburgh, S., Mathieson, J. T., Chapin, E. L., Kemmitt, G. M., Brown, J. M., et al. (1997). Dihydromaltophilin; a novel fungicidal tetramic acid containing metabolite from Streptomyces sp. J. Antibiot. 50, 1014–1019. doi: 10.7164/antibiotics.50.1014
Grubbs, K. J., Biedermann, P. H., Suen, G., Adams, S. M., Moeller, J. A., Klassen, J. L., et al. (2011). Genome sequence of Streptomyces griseus strain XylebKG-1, an ambrosia beetle-associated actinomycete. Comparat. Study 193, 2890–2891. doi: 10.1128/jb.00330-11
Hajek, A. E., Nielsen, C., Kepler, R. M., Long, S. J., and Castrillo, L. (2013). Fidelity among Sirex woodwasps and their fungal symbionts. Microb. Ecol. 65, 753–762. doi: 10.1007/s00248-013-0218-z
Hanshew, A. S., Mason, C. J., Raffa, K. F., and Currie, C. R. (2013). Minimization of chloroplast contamination in 16S rRNA gene pyrosequencing of insect herbivore bacterial communities. J. Microbiol. Methods 95, 149–155. doi: 10.1016/j.mimet.2013.08.007
Hanshew, A. S., McDonald, B. R., Díaz Díaz, C., Djiéto-Lordon, C., Blatrix, R., and Currie, C. R. (2015). Characterization of actinobacteria associated with three ant-plant mutualisms. Microb. Ecol. 69, 192–203. doi: 10.1007/s00248-014-0469-3
Harrington, T. C. (1981). Cycloheximide sensitivity as a taxonomic character in Ceratocystis. Mycologia 73, 1123–1129. doi: 10.2307/3759682
Heuer, H., Krsek, M., Baker, P., Smalla, K., and Wellington, E. M. (1997). Analysis of actinomycete communities by specific amplification of genes encoding 16S rRNA and gel-electrophoretic separation in denaturing gradients. Appl. Environ. Microbiol. 63, 3233–3241. doi: 10.1128/aem.63.8.3233-3241.1997
Human, Z. R., Slippers, B., Wilhelm de Beer, Z., Wingfield, M. J., and Venter, S. N. (2017). Antifungal actinomycetes associated with the pine bark beetle. Orthotomicus erosus, in South Africa. South Afr. J. Sci. 113, 1–7.
Hyatt, D., Chen, G. L., LoCascio, P. F., Land, M. L., Larimer, F. W., and Hauser, L. J. (2010). Prodigal: prokaryotic gene recognition and translation initiation site identification. BMC Bioinformatics 11:119. doi: 10.1186/1471-2105-11-119
Jeffs, P. W., and McWilliams, D. (1981). Carbon-13 nuclear magnetic resonance study of the biosynthesis of cycloheximide. Stereospecific incorporation of malonate into the glutarimide ring. J. Am. Chem. Soc. 103, 6185–6192. doi: 10.1021/ja00410a034
Kaltenpoth, M., Göttler, W., Herzner, G., and Strohm, E. (2005). Symbiotic bacteria protect wasp larvae from fungal infestation. Curr. Biol. 15, 475–479. doi: 10.1016/j.cub.2004.12.084
Katoh, K., and Standley, D. M. (2013). MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol. Biol. Evol. 30, 772–780. doi: 10.1093/molbev/mst010
Kirkendall, L. R., Biedermann, P. H. W., and Jordal, B. H. (2015). “Evolution and diversity of bark and ambrosia beetles,” in Bark Beetles: Biology and Ecology of Native and Invasive Species, eds F. E. Vega and R. W. Hofstetter (Cambridge, MA: Academic Press), 85–156. doi: 10.1016/b978-0-12-417156-5.00003-4
Kirkendall, L. R., Kent, D. S., and Raffa, K. F. (1997). “Interactions among males, females and offspring in bark and ambrosia beetles: the significance of living in tunnels for the evolution of social behavior,” in The Evolution of Social Behavior in Insects and Arachnids, eds J. C. Choe and B. J. Crespi (Cambridge: Cambridge University Press), 181–215.
Lakhvich, F. A., Buravskaya, T. N., and Akhrem, A. A. (1993). Synthesis of actiphenol and its normethyl analogues and 6-hydroxy derivatives. Chem. Nat. Comp. 29, 526–529. doi: 10.1007/bf00630584
Lauth, J., Ruiz-González, M. X., and Orivel, J. (2011). New findings in insect fungiculture: have ants developed non-food, agricultural products? Commun. Integ. Biol. 4, 728–730. doi: 10.4161/cib.17590
Leroy, C., Séjalon-Delmas, N., Jauneau, A., Ruiz-González, M. X., Gryta, H., Jargeat, P., et al. (2011). Trophic mediation by a fungus in an ant–plant mutualism. J. Ecol. 99, 583–590.
Li, X., Guo, W., and Ding, J. (2012). Mycangial fungus benefits the development of a leaf-rolling weevil. Euops chinesis. J. Insect Physiol. 58, 867–873. doi: 10.1016/j.jinsphys.2012.03.011
Mauchline, N., Hallett, I., Hill, G., and Casonato, S. (2011). Process of infection of armored scale insects (Diaspididae) by an entomopathogenic Cosmospora sp. J. Invertebr. Pathol. 108, 46–51. doi: 10.1016/j.jip.2011.07.002
Mayers, C. G., McNew, D. L., Harrington, T. C., Roeper, R. A., Fraedrich, S. W., Biedermann, P. H., et al. (2015). Three genera in the Ceratocystidaceae are the respective symbionts of three independent lineages of ambrosia beetles with large, complex mycangia. Fungal Biol. 119, 1075–1092. doi: 10.1016/j.funbio.2015.08.002
Mueller, U. G., Gerardo, N. M., Aanen, D. K., Six, D. L., and Schultz, T. R. (2005). The evolution of agriculture in insects. Annu. Rev. Ecol. Evol. Syst. 36, 563–595.
Mueller, U. G., Schultz, T. R., Currie, C. R., Adams, R. M., and Malloch, D. (2001). The origin of the attine ant-fungus mutualism. Q. Rev. Biol. 76, 169–197. doi: 10.1086/393867
Ogata, H., Goto, S., Sato, K., Fujibuchi, W., Bono, H., and Kanehisa, M. (1999). KEGG: kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 27, 29–34.
Oh, D. C., Poulsen, M., Currie, C. R., and Clardy, J. (2011). Sceliphrolactam, a polyene macrocyclic lactam from a wasp-associated Streptomyces sp. Organic Lett. 13, 752–755. doi: 10.1021/ol102991d
Peer, K., and Taborsky, M. (2004). Female ambrosia beetles adjust their offspring sex ratio according to outbreeding opportunities for their sons. J. Evol. Biol. 17, 257–264. doi: 10.1111/j.1420-9101.2003.00687.x
Persson, Y., Vasaitis, R., Långström, B., Öhrn, P., Ihrmark, K., and Stenlid, J. (2009). Fungi vectored by the bark beetle Ips typographus following hibernation under the bark of standing trees and in the forest litter. Microb.Ecol. 58, 651–659. doi: 10.1007/s00248-009-9520-1
Poulsen, M., Oh, D. C., Clardy, J., and Currie, C. R. (2011). Chemical analyses of wasp-associated Streptomyces bacteria reveal a prolific potential for natural products discovery. PLoS One 6:e16763. doi: 10.1371/journal.pone.0016763
Rehner, S. A., and Buckley, E. (2005). A Beauveria phylogeny inferred from nuclear ITS and EF1-α sequences: evidence for cryptic diversification and links to Cordyceps teleomorphs. Mycologia 97, 84–98. doi: 10.3852/mycologia.97.1.84
Roeper, R. A., and French, J. R. (1981). Ambrosia fungi of the western United States and Canada: beetle associations (Coleoptera: Scolytidae), tree hosts, and distributions [Ambrosiella, Raffaelea]. Northw. Sci. 55, 305–309.
Rong, X., and Huang, Y. (2010). Taxonomic evaluation of the Streptomyces griseus clade using multilocus sequence analysis and DNA–DNA hybridization, with proposal to combine 29 species and three subspecies as 11 genomic species. Int. J. Syst. Evol. Microbiol. 60, 696–703. doi: 10.1099/ijs.0.012419-0
Ruddick, S. M., and Williams, S. T. (1972). Studies on the ecology of actinomycetes in soil V. Some factors influencing the dispersal and adsorption of spores in soil. Soil Biol. Biochem. 4, 93–103. doi: 10.1016/0038-0717(72)90046-6
Scott, J. J., Oh, D. C., Yuceer, M. C., Klepzig, K. D., Clardy, J., and Currie, C. R. (2008). Bacterial protection of beetle-fungus mutualism. Science 322:63. doi: 10.1126/science.1160423
Seipke, R. F., Barke, J., Ruiz-Gonzalez, M. X., Orivel, J., Douglas, W. Y., and Hutchings, M. I. (2012). Fungus-growing Allomerus ants are associated with antibiotic-producing actinobacteria. Antonie van Leeuwenhoek 101, 443–447. doi: 10.1007/s10482-011-9621-y
Sekar, R., Pernthaler, A., Pernthaler, J., Warnecke, F., Posch, T., and Amann, R. (2003). An improved protocol for quantification of freshwater Actinobacteria by fluorescence in situ hybridization. Appl. Environ. Microbiol. 69, 2928–2935. doi: 10.1128/aem.69.5.2928-2935.2003
Six, D. L. (2012). Ecological and evolutionary determinants of bark beetle -fungus symbioses. Insects 3, 339–366. doi: 10.3390/insects3010339
Stach, E. M., and Bull, A. T. (2005). Estimating and comparing the diversity of marine actinobacteria. Antonie van Leeuwenhoek 87, 3–9. doi: 10.1007/s10482-004-6524-1
Stamatakis, A. (2006). RAxML-VI-HPC: maximum likelihood-based phylogenetic analyses with thousands of taxa and mixed models. Bioinformatics 22, 2688–2690. doi: 10.1093/bioinformatics/btl446
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M., and Kumar, S. (2011). MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 28, 2731–2739. doi: 10.1093/molbev/msr121
Toki, W., Tanahashi, M., Togashi, K., and Fukatsu, T. (2012). Fungal farming in a non-social beetle. PLoS One 7:e41893. doi: 10.1371/journal.pone.0041893
Visser, A. A., Nobre, T., Currie, C. R., Aanen, D. K., and Poulsen, M. (2012). Exploring the potential for actinobacteria as defensive symbionts in fungus-growing termites. Microb. Ecol. 63, 975–985. doi: 10.1007/s00248-011-9987-4
White, T. J., Bruns, T. D., Lee, S. B., and Taylor, J. W. (1990). Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. PCR Protoc. 18, 315–322. doi: 10.1016/b978-0-12-372180-8.50042-1
Wood, S. L. (1982). The Bark and Ambrosia Beetles of North and Central America (Coleoptera: Scolytidae), a Taxonomic Monograph. 6). Provo, UT: Brigham Young University.
Yin, M., Yan, Y., Lohman, J. R., Huang, S. X., Ma, M., Zhao, G. R., et al. (2014). Cycloheximide and actiphenol production in Streptomyces sp. YIM56141 governed by single biosynthetic machinery featuring an acyltransferase-less type I polyketide synthase. Organic Lett. 16, 3072–3075. doi: 10.1021/ol501179w
Keywords: symbiosis, mutualism, insect fungal interactions, antimicrobial, Insect symbiois
Citation: Grubbs KJ, Surup F, Biedermann PHW, McDonald BR, Klassen JL, Carlson CM, Clardy J and Currie CR (2020) Cycloheximide-Producing Streptomyces Associated With Xyleborinus saxesenii and Xyleborus affinis Fungus-Farming Ambrosia Beetles. Front. Microbiol. 11:562140. doi: 10.3389/fmicb.2020.562140
Received: 14 May 2020; Accepted: 19 August 2020;
Published: 24 September 2020.
Edited by:
Steve Perlman, University of Victoria, CanadaReviewed by:
Ryan Seipke, University of Leeds, United KingdomCopyright © 2020 Grubbs, Surup, Biedermann, McDonald, Klassen, Carlson, Clardy and Currie. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jon Clardy, am9uX2NsYXJkeUBobXMuaGFydmFyZC5lZHU=; Cameron R. Currie, Y3VycmllQHdpc2MuZWR1; Y3VycmllQGJhY3Qud2lzYy5lZHU=
†Present address: Kirk J. Grubbs, Global Digitalization – Bioinformatics, BASF Corporation, Morrisville, NC, United States; Frank Surup, Helmholtz Centre for Infection Research GmbH, Department Microbial Drugs, Braunschweig, Germany; Peter H. W. Biedermann, Albert-Ludwigs-University Freiburg, Chair for Forest Entomology and Protection, Stegen-Wittental, Germany; Bradon R. McDonald, Translational Genomics Research Institute, Phoenix, AZ, United States; Jonathan L. Klassen, Department of Molecular and Cell Biology, University of Connecticut, Storrs, CT, United States
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.