
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 18 December 2020
Sec. Extreme Microbiology
Volume 11 - 2020 | https://doi.org/10.3389/fmicb.2020.540404
This article is part of the Research Topic Polar and Alpine Microbiological and Biogeochemical Processes in the Warming World View all 13 articles
 Ingeborg J. Klarenberg1,2*
Ingeborg J. Klarenberg1,2* Christoph Keuschnig3
Christoph Keuschnig3 Denis Warshan2
Denis Warshan2 Ingibjörg Svala Jónsdóttir2
Ingibjörg Svala Jónsdóttir2 Oddur Vilhelmsson1,4,5
Oddur Vilhelmsson1,4,5Lichens are traditionally defined as a symbiosis between a fungus and a green alga and or a cyanobacterium. This idea has been challenged by the discovery of bacterial communities inhabiting the lichen thalli. These bacteria are thought to contribute to the survival of lichens under extreme and changing environmental conditions. How these changing environmental conditions affect the lichen-associated bacterial community composition remains unclear. We describe the total (rDNA-based) and potentially metabolically active (rRNA-based) bacterial community of the lichen Cetaria islandica and its response to long-term warming using a 20-year warming experiment in an Icelandic sub-Arctic tundra. 16S rRNA and rDNA amplicon sequencing showed that the orders Acetobacterales (of the class Alphaproteobacteria) and Acidobacteriales (of the phylum Acidobacteria) dominated the bacterial community. Numerous amplicon sequence variants (ASVs) could only be detected in the potentially active community but not in the total community. Long-term warming led to increases in relative abundance of bacterial taxa on class, order and ASV level. Warming altered the relative abundance of ASVs of the most common bacterial genera, such as Granulicella and Endobacter. The potentially metabolically active bacterial community was also more responsive to warming than the total community. Our results suggest that the bacterial community of the lichen C. islandica is dominated by acidophilic taxa and harbors disproportionally active rare taxa. We also show for the first time that climate warming can lead to shifts in lichen-associated bacterial community composition.
The notion that lichens harbor diverse bacterial and fungal communities has challenged the traditional view of the lichen as a symbiosis between a fungus (mycobiont) and an alga and/or a cyanobacterium (photobiont) (González et al., 2005; Spribille et al., 2016). Nonetheless, the first lichen-associated bacteria were already discovered in the 1920s (Uphof, 1925). To date, bacterial communities of a wide-range of lichen species have been revealed by molecular approaches (Cardinale et al., 2006, 2008; Grube et al., 2009; Hodkinson and Lutzoni, 2009; Bjelland et al., 2011; Mushegian et al., 2011; Weiss et al., 2011; Sigurbjörnsdóttir et al., 2015; Park et al., 2016). Alphaproteobacteria usually dominate the lichen microbiome, but other taxa such as Actinobacteria, Firmicutes, Acidobacteria, Betaproteobacteria, Deltaproteobacteria, and Gammaproteobacteria are also found. These bacteria can form highly structured, biofilm-like assemblages on fungal surfaces and within the lichen thallus (Grube et al., 2009). The bacterial communities inhabiting the lichen thalli play important roles in the lichen holobiont (the lichen and its microbiome), by contributing to nutrient supply, resistance against biotic and abiotic stresses, production of vitamins and support of fungal and algal growth by the production of hormones, detoxification of metabolites and degradation of senescing parts of the lichen thallus (Grube et al., 2015; Sigurbjörnsdóttir et al., 2015). Thereby, the bacterial part of the lichen holobiont is suggested to contribute to the survival of lichens under extreme and changing environmental conditions.
The composition of associated bacterial communities of lichens may be shaped by intrinsic and extrinsic factors. Among intrinsic factors affecting the lichen microbiome are thallus age (Cardinale et al., 2012b), mycobiont species, and photobiont species (Grube et al., 2009; Weiss et al., 2011; Hodkinson et al., 2012; Wedin et al., 2016; Coleine et al., 2019). The composition of lichen bacterial communities can also be influenced by extrinsic factors such as sunlight exposure and substrate type (Cardinale et al., 2012b; Park et al., 2016), geography and local habitat (Cardinale et al., 2012a; Hodkinson et al., 2012; Printzen et al., 2012; West et al., 2018), altitude (Coleine et al., 2019), drought (Cernava et al., 2019), and arsenic contamination (Cernava et al., 2018). Some lichens can adapt to changing environmental factors by switching photobionts depending on the ecological niche of the photobiont (Domaschke et al., 2013; Rolshausen et al., 2018). Some lichens have also been shown to be able to acclimate to higher temperature by increasing their respiration (Lange and Green, 2005) or net photosynthesis (Colesie et al., 2018). However, not all lichens are able to adapt to changing environments in these ways. Lichens might also be able to acclimate through changes in their associated bacterial communities. This strategy has been demonstrated for several environmental factors, such as drought (Cernava et al., 2019) and arsenic contamination (Cernava et al., 2018). Substrate type is another extrinsic factor that can influence the composition of bacterial communities (Cardinale et al., 2012b). Therefore, changes in C (carbon) or N (nitrogen) availability in the environment, for instance as a result of changes in plant litter quality due to shrubification (McLaren et al., 2017), might be factors altering the structure of the lichen microbiome. However, little is known about the effect of long-term environmental changes on the bacterial communities associated with lichens.
High-latitudes are especially rich in lichen species and biomass (Cornelissen et al., 2007; Nash, 2008), where they make significant contributions to ecosystem functioning (Asplund and Wardle, 2017). Mat-forming lichens such as Cetraroid species contribute to primary production and nutrient cycling, control soil chemistry and water retention (Cornelissen et al., 2007). Currently, climate in high-latitudes warms twice as fast as elsewhere (IPCC, 2019) resulting in increased abundance of shrubs, particularly in the low and sub-Arctic (Elmendorf et al., 2012; Myers-Smith et al., 2019). Direct effects of warming on lichens include changes in C-based secondary compounds (Asplund et al., 2017) and increased biomass (Biasi et al., 2008). Warming also has indirect effects on lichens. In many low and sub-Arctic tundra ecosystems shrubification results in increased shading and greater amounts of litter, which can lead to decreased lichen photosynthesis rates causing a decline in lichen biomass (Nash and Olafsen, 1995; Cornelissen et al., 2001; Elmendorf et al., 2012; Fraser et al., 2014; Alatalo et al., 2017). Yet, the effect of long-term warming on the bacterial communities of lichens in high-latitudes needs to be investigated.
In this study we investigate the total and potentially metabolically active bacterial community of the lichen Cetraria islandica (L.) Ach. (English “Iceland moss”) and its response to two decades of warming in open top chambers (OTCs) in an Icelandic sub-Arctic alpine dwarf-shrub heath. C. islandica is a mat-forming chlorolichen with foliose thalli and forms a major component of the vegetation in Arctic, sub-Arctic and alpine environments throughout the northern hemisphere (Kärnefelt et al., 1993). 16S rRNA and rDNA sequencing was used to characterize the potentially active and total bacterial community in control plots and OTCs. We also quantified 16S rRNA gene abundance by quantitative PCR to compare the absolute abundance of bacteria in the controls and OTCs. Finally, it was recently demonstrated that N2-fixing bacteria could associate with chlorolichens (Almendras et al., 2018). Thus, we also quantified the number of nifH genes by quantitative PCR in order to test if the C. islandica microbiome could potentially perform N2-fixation and how warming influences the abundance of associated N2-fixers.
We predicted that long-term warming and the associated increase in tundra shrubs and litter will lead to an increase in heterotrophic, biopolymer-degrading bacterial taxa and a higher incidence of potentially lichenivorous or lichenopathogenic bacteria, such as shown for the plant phyllosphere (Aydogan et al., 2018). Thus, in terms of taxonomic composition, we expected an increase in detritivorous taxa, endosymbionts and pathogens of fungi such as chitinolytic bacteria (Kobayashi and Crouch, 2009), whereas the relative abundance of cold-adapted and facultatively lithotrophic bacteria may decrease.
We also hypothesized that the potentially metabolically active (16S rRNA based) community shows a larger change in richness, diversity and community structure to the warming treatment than the total bacterial community, as has been shown for the effect of drought (Bastida et al., 2017).
The study site is located in a Betula nana heath in the Icelandic central highlands at an altitude of 450 m. According to Köppen’s climate definitions, the sampling site, called Aukúluheii (65°16′N, 20°15′W, 480 m above sea level) is situated in the lower Arctic. The vegetation is characterized as a relatively species-rich dwarf shrub heath, with B. nana being the most dominant vascular species and the moss Racomitrium lanuginosum and the lichen C. islandica as the dominating cryptogam species (Jonsdottir et al., 2005).
Ten OTCs were set up to simulate a warmer summer climate in August 1996 in a fenced area to exclude sheep grazing (Hollister and Webber, 2000; Jonsdottir et al., 2005). The OTCs raise the mean daily temperature by 1–2°C during summer and minimize secondary experimental effects such as differences in atmospheric gas concentration and reduction in ambient precipitation. Control plots were established adjacent to the OTCs, but without any treatment, thus exposing the environment to ambient temperatures. Air temperatures measured in the growing season from 1999 to 2002 at the surface of the cryptogam layer indicated an average increase of 0.7–1.0°C in the OTCs (Jonsdottir et al., 2005). Air temperature measured 10 cm above the moss layer in summer 2016 was on average 1.4°C higher in the OTC than in the control plot of one of the plot pairs (t = −8.2, P < 0.001) (Supplementary Table 1). Relative humidity measured in the same plot pair and period was 3% lower in the OTC than in the control plot (t = 26.9, P < 0.001) (Supplementary Table 1). Temperatures on the moss surface measured in all OTCs and control plots from mid-August 2018 to mid-June 2019 were on average 0.22°C higher in the OTCs compared to the control plots (t = −16.4, P < 0.001) (Supplementary Table 1).
The response of the vegetation was monitored by a detailed vegetation analysis after peak biomass at a few year intervals using the point intercept method following standard protocols of the International Tundra Experiment (ITEX: 100 points per plot, all hits (intercepts) per species recorded in each point through the canopy; relates to biomass) (Jonsdottir et al., 2005). In 2014, the abundance of B. nana was on average 2.5 times larger in the OTCs than in the control plots and litter was 2.7 times more abundant in the OTCs than in the control plots (Jónsdóttir, unpublished data). In this study we use these data on abundance (total number of hits per plot) of B. nana and litter to test the effect of vegetation change on the richness, diversity, community structure of the lichen associated bacterial community as well 16S rRNA and nifH gene abundance.
Per warmed (OTC) and control plot, the upper parts (2 × 2 cm) of five lichen thalli were randomly selected and collected with sterile tweezers. The samples were immediately soaked in RNAlater (Ambion) to prevent RNA degradation and kept cool until storage at −80°C. The lichen samples were collected in June 2017.
Prior to RNA and DNA extraction, the samples were washed with RNase free water to get rid of soil particles and RNAlater and ground for six minutes using a Mini-Beadbeater and two sterile steel beads. RNA and DNA were extracted simultaneously using the RNeasy PowerSoil Total RNA Kit (Qiagen) and the RNeasy PowerSoil DNA Elution Kit (Qiagen), following the manufacturer’s instructions. DNA and RNA concentrations were measured with a Qubit Fluorometer (Life Technologies) and purity was assessed with a NanoDrop (NanoDrop Technologies) and integrity by Bioanalyzer (Agilent Technologies). cDNA was synthesized using the High-Capacity cDNA Reverse Transcription Kit (Thermofisher) following the manufacturer’s instructions and quantified on a Qubit Fluorometer (Life Technologies). From the 100 DNA and 100 cDNA samples, we selected 48 DNA samples (24 from each treatment) and 48 cDNA samples (24 from each treatment) for sequencing based on the RNA and DNA quantity and quality. Library preparation and sequencing of the V3–V4 region of the 16S rRNA gene on an Illumina MiSeq platform was performed by Macrogen (Seoul), using the standard Illumina protocol. The primer pair 337F/805R and the same PCR condition described in Klindworth et al. (2013) were used.
In order to obtain high-resolution data, we processed the raw sequences using the DADA2 pipeline (Callahan et al., 2016, 2017). Hereby, sequences are not clustered into operational taxonomic units (OTUs), but exact sequences or amplicon sequence variants (ASVs). Forward reads were truncated at 260 bp and reverse reads at 250 bp. Assembled ASVs were assigned taxonomy against the SILVA_132 database (Quast et al., 2013) using the Ribosomal Database Project (RDP) naïve Bayesian classifier (Wang et al., 2007) in DADA2. We discarded samples with less than 10,000 non-chimeric sequences and/or less than 50 ASVs. We removed ASVs assigned to chloroplasts and mitochondria, singletons, doubletons and ASVs occurring in only one sample. In total, for 82 samples, 1,954 ASVs remained. The data were normalized using cumulative-sum scaling (CSS) (Paulson et al., 2013) to account for uneven sequencing depths.
The 16S rDNA based community is hereafter sometimes referred to as the DNA based community and the 16S rRNA (cDNA) based community is hereafter referred to as the cDNA based community. We interpret the cDNA based community as the “potentially metabolically active bacterial community,” acknowledging that 16S rRNA is not a direct indicator of activity but rather protein synthesis potential (Blazewicz et al., 2013).
We used all 100 DNA extractions (50 replicates per treatment) for quantification of nifH and 16S rRNA genes, which was performed by quantitative PCR (Corbett Rotor-Gene) using the primer set PolF/PolR and 341F/534R, respectively (Poly et al., 2001). The specificity of the nifH primers for our samples was confirmed by SANGER sequencing of 10 clone fragments. Standards for nifH reactions were obtained by amplifying one cloned nifH sequence with flanking regions of the plasmid vector (TOPO TA cloning Kit, Invitrogen). Standard curves were obtained by serial dilutions (E = 0.9–1.1, R2 = > 0.99 for all reactions). Each reaction had a volume of 20 μL, containing 1× QuantiFast SYBR Green PCR Master Mix (Qiagen), 0.2 μL of each primer (10 μM), 0.8 μL BSA (5 μg/μL), 6.8 μL RNase free water, and 2 μL template. The cycling program was 5 min at 95°C, 30 cycles of 10 s at 95°C and 30 s at 60°C.
Statistical analyses were conducted in R version 3.6.1. Richness (number of ASVs) and Shannon diversity were calculated with the R packages “vegan” (Oksanen et al., 2013) and “phyloseq” (McMurdie and Holmes, 2013).
Differences in 16S rRNA and nifH gene abundance, ASV richness and Shannon diversity between the treatments and the DNA and cDNA were assessed with Bayesian (Markov chain Monte Carlo) generalized linear models using the R package “MCMCglmm” (Hadfield, 2010) with treatment as a fixed and plot as a random factor to take into account the variation caused by pseudoreplication. We considered differences significant if the modeled 95% confidence intervals did not overlap. In addition to the effect of treatment, we included B. nana and litter abundance in these generalized linear models. The effect of B. nana and/or litter abundance was considered significant if 95% High Posterior Density Credible Interval (95% CrI) were not overlapping zero.
Distances between the community composition of the control and OTC samples were based on Bray-Curtis distances. The effect of the treatment, B. nana abundance and litter abundance on the bacterial community composition was tested by permutational-MANOVA (PERMANOVA) (Anderson, 2001) analysis of the Bray-Curtis distance matrices using the adonis function in the R package “vegan” with plot as strata. Principal coordinate analysis was used to ordinate the Bray-Curtis distance matrices and to visualize the relationships between samples from OTC and control plots.
For the comparisons of relative abundances of taxa on phylum, class and order level between the warmed and the control samples, pseudoreplicates were averaged by the OTC or control plot they originated from. These average relative abundances were then compared using Wilcoxon rank-sum tests.
We used two methods to determine taxa sensitive to warming. For both methods, we used the average abundances of ASVs in each plot. First, differential abundance of bacterial ASVs between warmed and control samples was assessed using DESeq2 (Love et al., 2014) on the non-CSS normalized datasets with the R package “DESeq2” (Love et al., 2014). The adjusted P-value cut-off was 0.1 (Love et al., 2014). Differential abundance analysis only uses ASVs present in both the OTC and control samples. The second method we used to find taxa sensitive to warming, was indicator species analysis. To find bacterial taxa indicative for the warming or the control treatment, correlation-based indicator species analysis was done with all possible site combinations using the function multipatt of the R package “indicSpecies” (De Caceres and Legendre, 2009) based on 103 permutations. The indicator species analysis takes into account ASVs present in both OTC and control samples, but also ASVs present in only one of the treatments. We combined results of the DESeq2 and indicator species analysis for a final list of ASVs sensitive to warming.
For visualizations of the data, we showed all samples when we could account for pseudoreplication (Bayesian generalized linear models and Permanovas) and we showed plot averages when we compared between the control and warmed treatment (Wilcoxon rank-sum tests).
The ASV richness and Shannon diversity of the bacterial communities associated with C. islandica were not significantly affected by the warming treatment (Figure 1 and Supplementary Figures 1–4). However, we found that B. nana abundance tended to positively influence the cDNA-based bacterial richness and Shannon diversity (Supplementary Figures 3, 4). A significant difference was found for the richness, which was higher for the cDNA-based bacterial community than the DNA-based community in the warmed treatment (Supplementary Figure 7). The cDNA-based Shannon index tended to be higher than the DNA-based Shannon index (Figure 1 and Supplementary Figure 5).
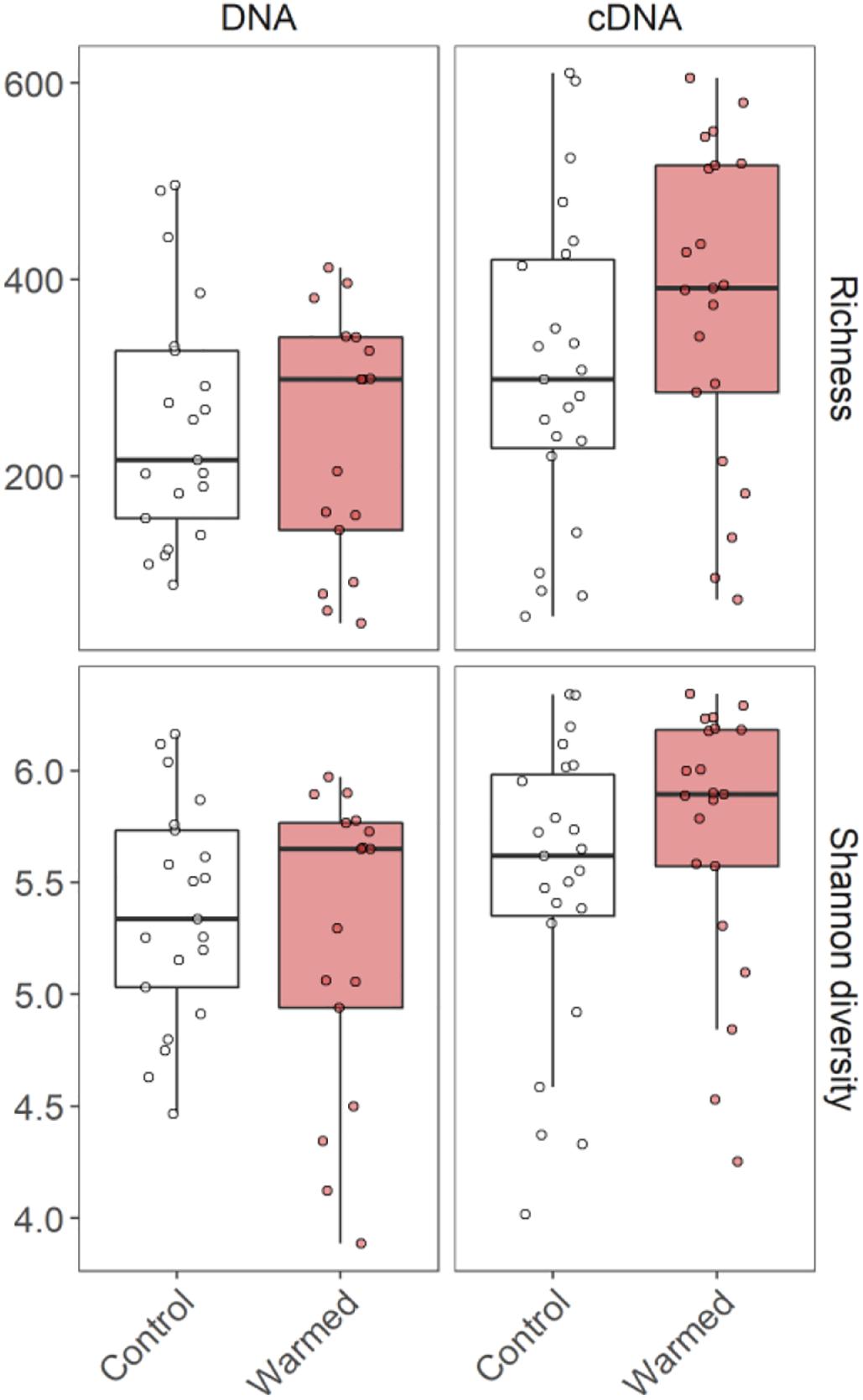
Figure 1. ASV richness and Shannon diversity of the DNA and cDNA-based bacterial communities associated with the lichen C. islandica in control and warmed samples. N = 82, n = 21 for DNA control, n = 17 for DNA warmed, n = 23 for cDNA control and n = 21 for cDNA warmed.
Some level of clustering between the control and warmed samples could be observed in the principal coordinate analysis (Figure 2). Based on the results of a PERMANOVA, the warmed lichen associated bacterial communities were significantly different from the communities in the control samples (DNA: R2 = 0.07 and P = 0.001; RNA: R2 = 0.07 and P = 0.005) (Supplementary Tables 2, 3). In addition, litter abundance was associated with variation in the cDNA-based bacterial community (Permanova: R2 = 0.04 and P = 0.05).
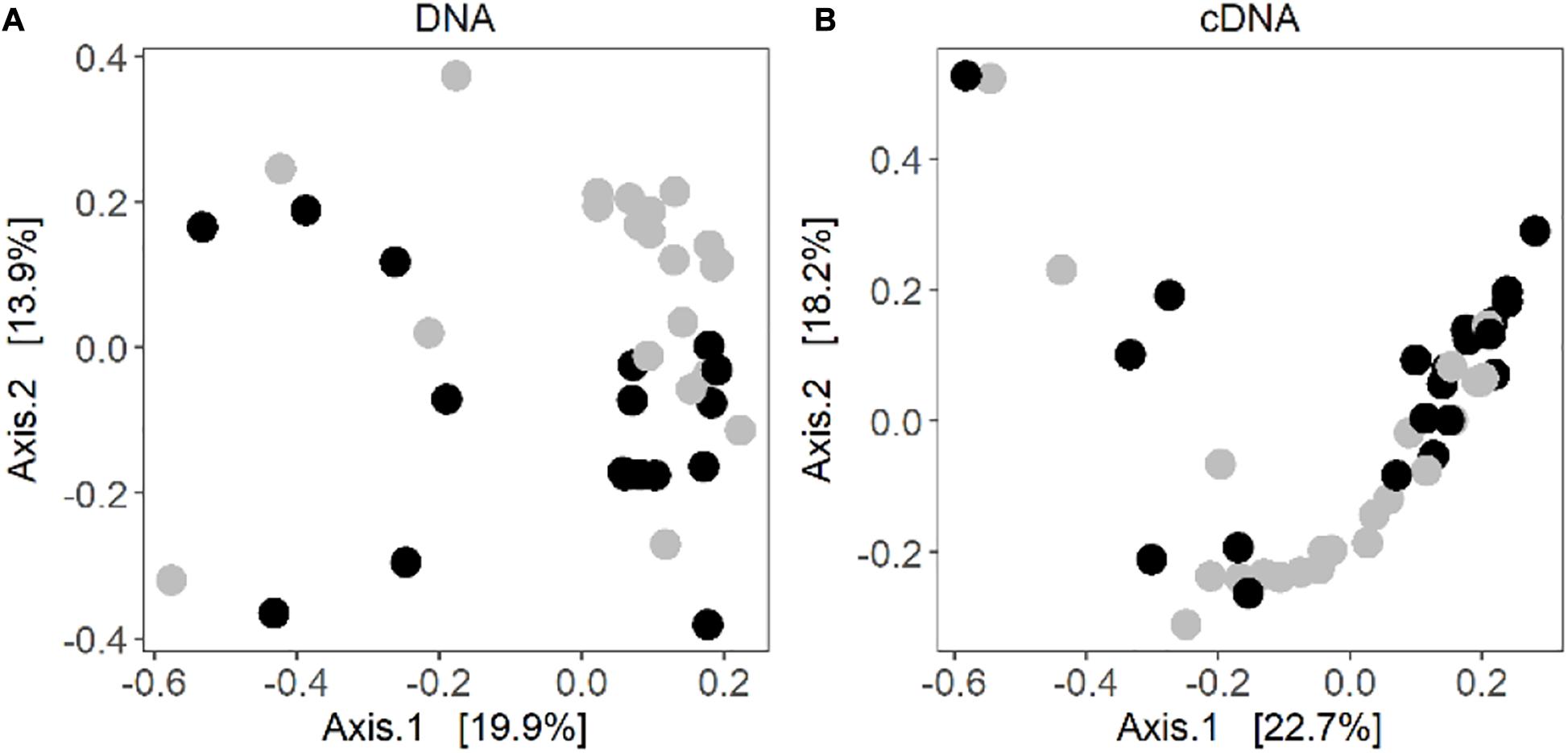
Figure 2. Principal coordinate analysis of Bray-Curtis distances of the (A) DNA-based community composition and (B) cDNA-based community composition of the lichen C. islandica. Warmed samples are represented as black circles and control samples as gray circles. N = 28, n = 21 for DNA control, n = 17 for DNA warmed, n = 23 for cDNA control and n = 21 for cDNA warmed.
The bacterial community found associated with the lichen C. islandica is described at the phylum level (Figure 3A) and at the class and order level (Figure 4). No clear differences were found for the relative abundance at the phylum level between the control and warmed treatment for the total bacteria community (Figure 3A). Similarly, we did not detect differences between the control and warmed treatment in the cDNA-based bacterial community at the phylum level (Figure 3A). The total bacterial community was dominated by Proteobacteria and Acidobacteria (DNA: 58 and 34% average relative abundance across all control and warmed samples, for Proteobacteria and Acidobacteria, respectively). Proteobacteria and Acidobacteria were also the main phyla in the cDNA-based bacterial community (cDNA: 63 and 29%, respectively) (Figure 3A). At lower taxonomic level, the orders Acetobacterales and Acidobacteriales were the dominant taxa (DNA: 44%; cDNA: 51%, DNA: 34%; cDNA 29%, Figure 4A). Within the acidobacterial family Acetobacteraceae, about 14% could not be assigned to a genus (Supplementary Figure 7). The most abundant genera in the cDNA and DNA-based bacterial communities were the proteobacterial genera Acidiphilium (DNA: 8%, cDNA: 11%) and Endobacter (DNA: 19%, cDNA: 20%) and the acidobacterial genera Bryocella (DNA: 10%, cDNA: 9%) and Granulicella (DNA: 15%, cDNA: 11%) (Supplementary Figure 7).
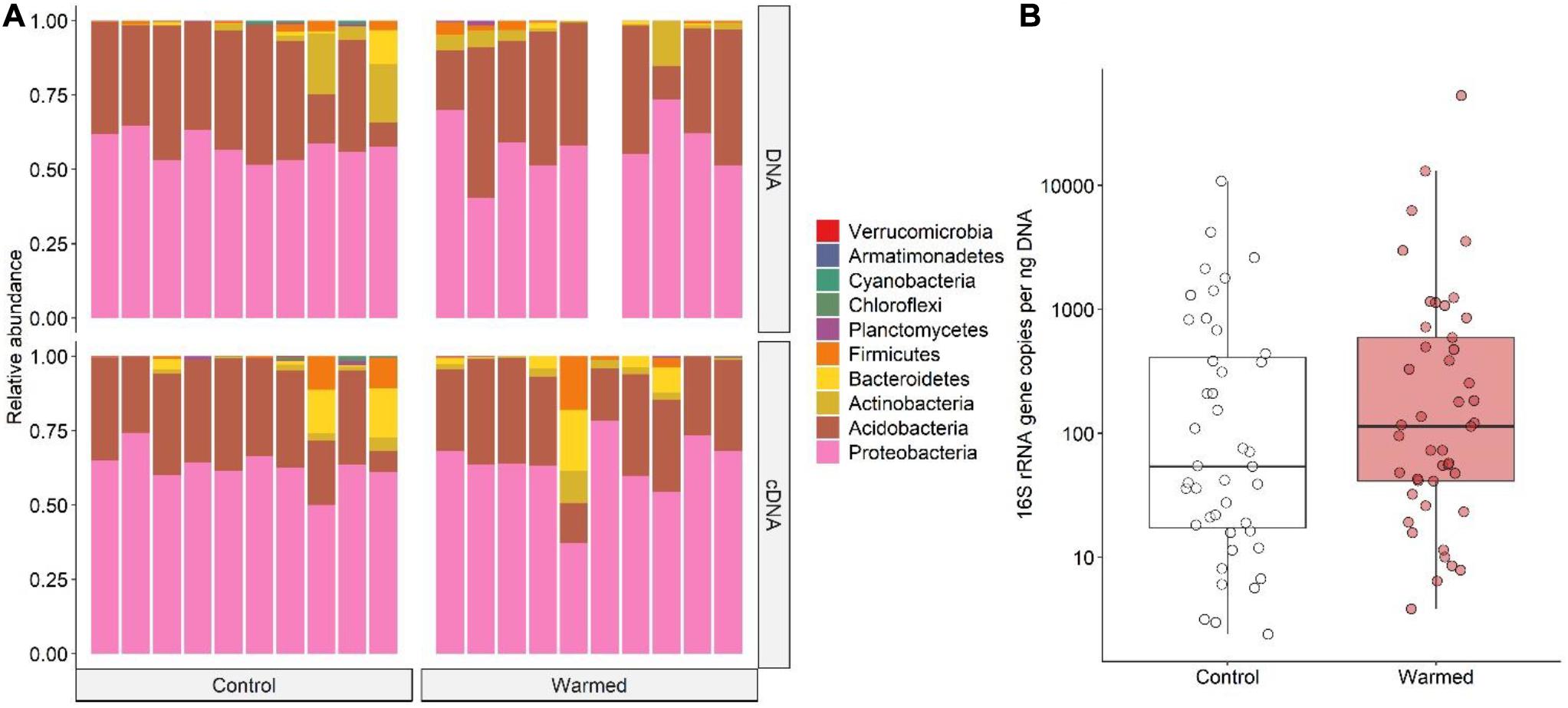
Figure 3. (A) Barplots of the bacterial community composition associated with the lichen C. islandica in the control and warmed plots. Data are presented at the phylum level for DNA and cDNA. Bars represent pooled samples from each separate plot. (B) 16S rRNA gene abundance per ng extracted DNA in control (white) and warmed (red) samples. N = 88, nwarmed = 45, ncontrol = 43.
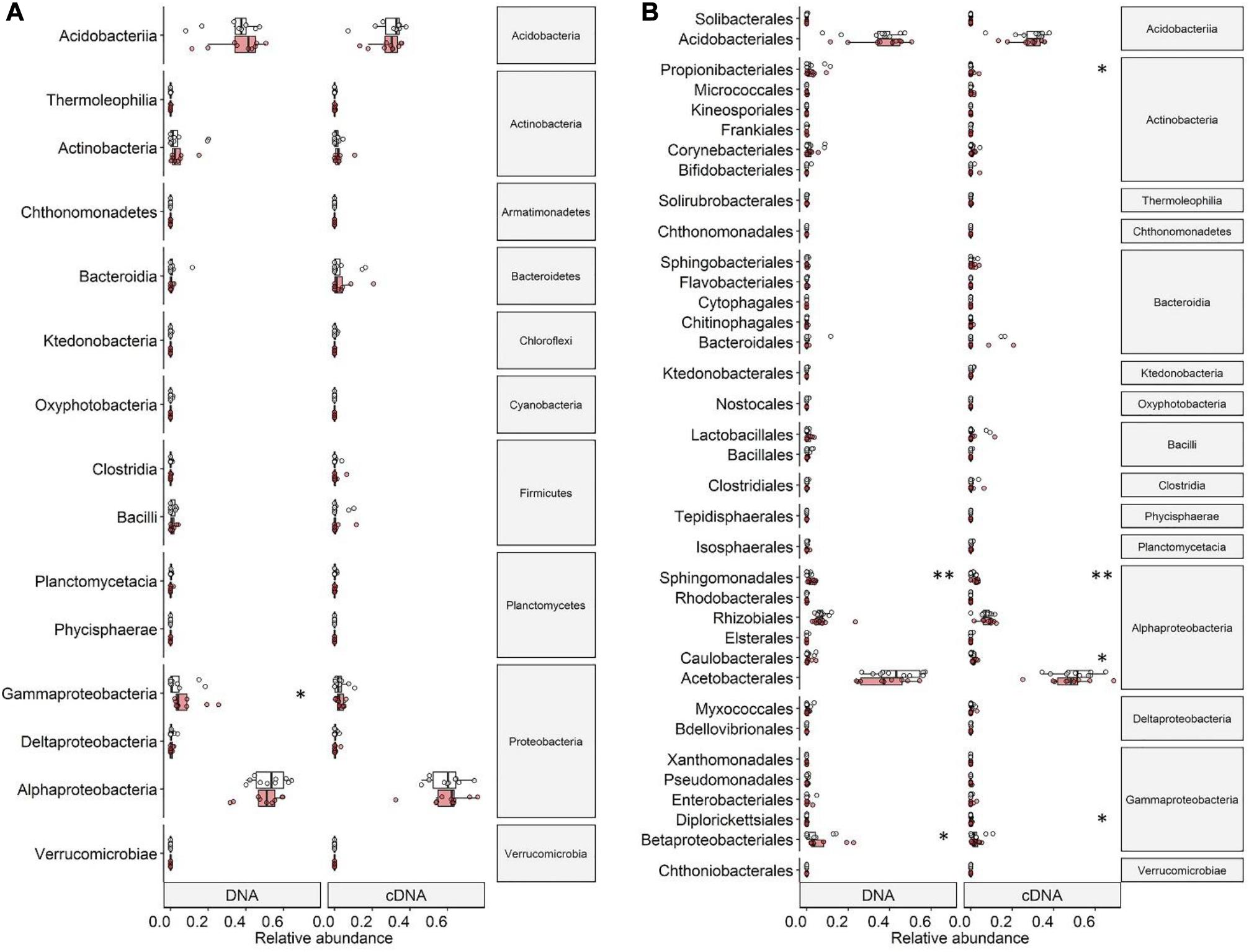
Figure 4. Relative abundances of (A) classes and (B) orders of DNA- and cDNA-based bacterial communities associated with the lichen C. islandica in control (white) and warmed (red) samples. Points indicate average relative abundance values per control or warmed plot. Boxplots represent minimum values, first quartiles, medians, third quartiles, and maximum values. Significance levels (∗p < 0.05 and ∗∗p < 0.01) are based on Wilcoxon rank-sum tests.
A total of 295 ASVs were only detected in the cDNA-based samples and not in the DNA-based samples. These taxa belonged to abundant genera such as Endobacter and Acidiphilium (Supplementary Tables 4, 5). Genera that were exclusively found in the cDNA-based community were Lactococcus, Lachnospiraceae NK4A136 group, Ktedonobacter, Methylovirgula, Frigoribacterium, Amnibacterium, Rhizobacter, Telmatobacter, Kineosporia, and Acidiphilium (Supplementary Tables 4, 5).
The bacterial load was on average 671 and 1944 16S rRNA copies per ng DNA for control and warmed plots, respectively (Figure 3B). However, no differences could be found between the overall 16S rRNA gene copy numbers, or nifH gene copy numbers in the control and warmed plots (Figure 3B and Supplementary Figures 8, 9). Nevertheless, B. nana abundance positively affected both nifH and 16S rRNA gene copy numbers, whereas litter abundance negatively affected nifH gene copy numbers (Supplementary Figures 8, 9).
In the DNA-based samples, the only proteobacterial class found significantly affected by warming were the Gammaproteobacteria with an increase of 50% in the warmed samples (Wilcoxon rank-sum test, P = 0.033; Figure 4A). Most of this increase was due to an increase in relative abundance of the order Betaproteobacteriales (P = 0.046) (Figure 4B). The order Sphingomonadales (Alphaproteobacteria) increased by a factor of 2.5 in relative abundance (P = 0.009) (Figure 4B). In the cDNA-based bacterial community, we did not detect differences in relative abundance on Proteobacterial class level between the control and warmed treatment (Figure 4A). Changes were found on order level with the alphaproteobacterial order Caulobacterales being twice as abundant in the cDNA-based bacterial community in the warmed plots (P = 0.038) than in the control plots. Similarly, the order Sphingomonadales was 2.7 times more active in the warmed treatment (P = 0.007) (Figure 4B). The order Diplorickettsiales (Gammaproteobacteria) was four times as abundant in the cDNA-based bacterial community in the warmed plots (P = 0.040) (Figure 4B).
While the phylum Actinobacteria was not among the most common phyla (DNA: 4%, cDNA: 2%), its order Propionibacteriales was 20 times as abundant in the cDNA-based bacterial community in the warmed plots compared to the control plots (P = 0.040) (Figure 4B).
For the DNA-based communities, we detected 61 ASVs with a higher relative abundance in the warmed samples with a total relative abundance of 1% (Supplementary Table 4). We detected 96 ASVs with a lower relative abundance in the warmed samples compared to the control samples making up 1.7% of the total abundance (Supplementary Table 4). For the cDNA-based bacterial communities, we detected 190 ASVs with a higher relative abundance in the warmed samples (2.12%) and 77 ASVs with a lower relative abundance in the warmed samples compared to the control samples (0.9%) (Supplementary Table 5).
Of the ASVs only detected in cDNA-based bacterial community, 14 ASVs had a higher relative abundance in the warmed plots. All these rare ASVs belonged to the Proteobacteria, except one ASV that was classified as Bacteroidetes.
Amplicon sequence variants within the Proteobacteria showed mainly increased relative abundance in the warmed samples (Figure 5 and Supplementary Tables 4, 5). Only ASVs classified under the genus Acidiphilium had more often a lower relative abundance in the warmed samples as well as a few ASVs of the genera Acidisphaera and Endobacter. In the cDNA-based samples, more proteobacterial ASVs with increased relative abundances under warming were detected than in the DNA-based samples.
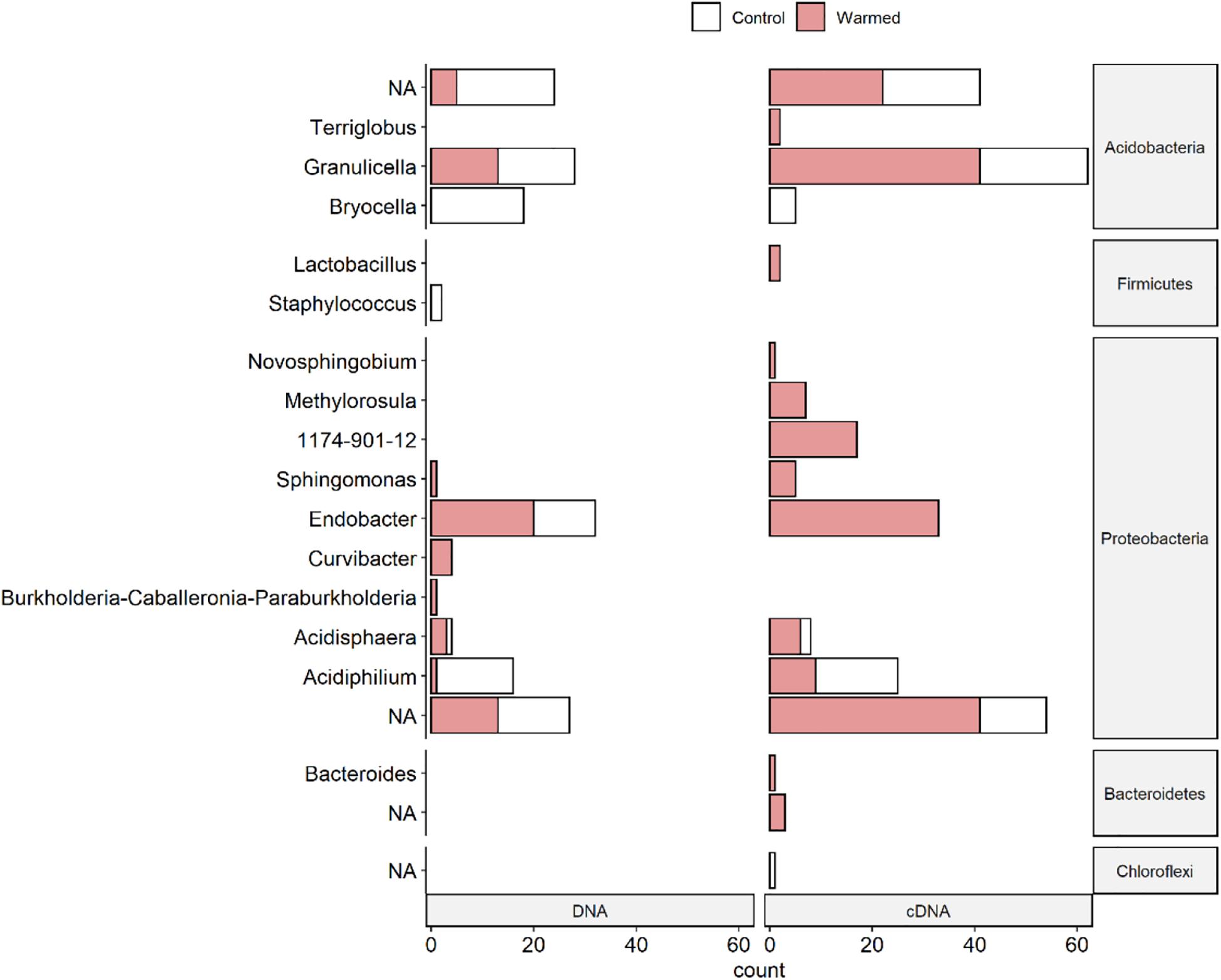
Figure 5. Number of ASVs (amplicon sequence variants) per genus sensitive to warming for DNA and cDNA-based bacterial communities associated with the lichen C. islandica. Sensitivity to warming was determined by differential abundance analysis (DESeq2) and indicator species analysis. ASVs not assigned to genus level are called “NA.” The number of ASVs indicative for the OTC (warmed) treatment is indicated in red and the number of ASVs indicative for the control treatment is indicated in white.
Acidobacterial ASVs showed mixed differences between the control and warmed samples (Figure 5 and Supplementary Tables 4, 5). ASVs of the genus Bryocella were less abundant under warming in both the DNA- and cDNA-based samples. ASVs of the genus Granulicella were equally more and less abundant in the warmed DNA-based samples, but had more often higher relative abundances in the warmed cDNA-based samples (Figure 5 and Supplementary Tables 4, 5).
We assessed the effect of long-term (20 years) warming by OTCs on the bacterial community composition associated with the lichen C. islandica in an Icelandic sub-Arctic alpine dwarf-shrub heath. The community was dominated by Acidobacteria and Proteobacteria in both total and potentially active bacterial communities in both control and warmed plots. Warming did not induce compositional or structural changes at higher taxonomical levels. Nevertheless, we found indications of multiple warming-induced shifts in the community composition at the class, order and ASV levels. The most prominent increases in relative abundance were found in several genera belonging to the Proteobacteria. Our results illustrate that the long-term warming treatment affects the bacterial community composition of the lichen symbiosis at fine taxonomical levels.
While the dominance of the class Alphaproteobacteria has been described as a general characteristic of lichen bacterial communities (Cardinale et al., 2008; Weiss et al., 2011; Printzen et al., 2012), a striking feature of the C. islandica microbiome is the strong dominance of the family Acetobacteriaceae (Alphaproteobacteria). While the presence of Acetobacteriaceae in lichens has been observed before, notably in the reindeer lichen Cladonia arbuscula (Cardinale et al., 2008), this strong dominance does seem unusual. Indeed, the Rhizobiaceae, often held to be the dominant Alphaproteobacteria in lichens (Weiss et al., 2011; Hodkinson et al., 2012), only make up a minor part of the C. islandica bacteriome. The second dominant group in the C. islandica bacteriome are members of the family Acidobacteriaceae (Acidobacteria). Even at the genus level, the C. islandica bacteriome is surprisingly homogeneous, with approximately half of the reads being assigned to only four genera, the acetobacterial genera Endobacter and Acidiphilium, and the acidobacterial genera Granulicella and Bryocella. This pronounced dominance of presumptively acidophilic taxa is noteworthy. Acidobacteria were also reported earlier in living parts of bog and tundra (Pankratov, 2012) and lichens in Alpine soil crusts (Muggia et al., 2013). The presence of acidophilic taxa may be explained by organic acid secondary metabolites produced by C. islandica, such as protolichesterinic and fumaroprotocetraric acids (Xu et al., 2018).
Another feature of the C. islandica microbiota is the difference between the potentially metabolically active and total community. The richness of the potentially active bacterial communities was higher compared the to total bacterial communities. One possible explanation for this is that the detection of taxa in 16S rRNA sequences, but not in 16S rDNA sequences, can occur when rare taxa have a high metabolic potential. The occurrence of these “phantom taxa” could be a result of cDNA synthesis errors that do not occur in the rDNA samples, but are introduced in the rRNA sequences. Another explanation could be variation in metabolic activity among taxa (Campbell et al., 2011; Baldrian et al., 2012; Klein et al., 2016; Jia et al., 2019). Rare taxa have been observed to be disproportionally active compared to abundant members (Jones and Lennon, 2010) and thereby might contribute more to ecosystem functioning than one would expect based on their abundance (Jousset et al., 2017). The rare bacterial taxa of C. islandica were mostly composed of not assigned genera and members of the genera Endobacter, Acidiphilium, Lactococcus, Mucilaginibacter, and Bacteroides. The fermenting bacteria from the genus Lactococcus have been described before in a bioreactor as being rare while having high potential activity levels (Lawson et al., 2015).
N2-fixation is an important process in N-limited tundra ecosystems and previous work has shown that biocrust chlorolichens can show significant nitrogenase activity (Torres-Cruz et al., 2018). As C. islandica is a chlorolichen and does not have N2-fixing Cyanobacteria as a photobiont, this raises the question if other taxa could be N2-fixers. The N2-fixing capability of C. islandica is unknown, but the presence of nifH genes indicates that potential N2-fixers are present in the C. islandica bacterial community. Indeed, we detected putative N2-fixers such as Curvibacter (Ding, 2004) and members of the Burkholderiaceae. On the other hand, lichens might also obtain nutrients dissolved in precipitation or through runoff from taller vegetation, or via a moisture gradient resulting in upward movement of soil moisture and dissolved nutrients (Longton, 1992). Nevertheless, our data indicate that the indirect effect of warming through changes in litter and B. nana abundance can influence the abundance of N2-fixing bacteria. More studies on the N2-fixation capabilities of chlorolichens in tundra ecosystems are necessary to elucidate their role as N2-fixers.
The C. islandica-associated microbiota was found to be markedly different to that of the moss Racomitrium lanuginosum which was studied in the same warming experiment (Klarenberg et al., 2019), further supporting the host-specific selection of bacteria from the environment and symbiotic nature of both bryophyte and lichen holobionts proposed in the recent literature (Aschenbrenner et al., 2016; Holland-Moritz et al., 2018). Specifically, we found that C. islandica harbored a less rich and diverse bacterial community than R. lanuginosum, and the microbiota composition was profoundly different. Whereas the moss was dominated by the genera Haliangium, Acidiphilium, Nostoc, Conexibacter, Granulicella, Solibacter, and Bryobacter, the lichen was dominated by a few genera (Bryocella, Granulicella, Acidiphilium, and Endobacter) as reported herein. The same difference between the bacterial diversity of a lichen and a moss was shown by Aschenbrenner et al. (2017).
While we did not see any significant changes in richness or diversity of the bacterial community with warming, the warmed bacterial community structure significantly differed from the control community, both for the total as well as the potentially active communities. Overall, the potentially active community tended to be more affected by warming than the total bacterial community. For instance, more indicator taxa were found in the potentially active community and many more of these indicators were found in the warmed treatment. In addition, indirect effects of warming via shrubification and litter modification were found to affect the bacterial community. A positive effect of B. nana abundance was found on the richness and diversity of the potentially active community as well as on 16S rRNA and nifH gene abundance. Litter abundance was positively associated with nifH gene abundance and with the structure of the potentially active bacterial community.
At a coarse taxonomic level, the bacterial community structure was quite similar between the control and warmed treatment. One possible explanation for the similarity between the richness, diversity and composition of the warmed and control lichen bacterial communities could be that over long periods of warming bacterial communities acclimatize (Bradford et al., 2008; Crowther and Bradford, 2013; Romero-Olivares et al., 2017). Nonetheless, at lower taxonomic levels (class, order, and ASV) we detected differences in relative abundances. Shifts in individual taxa can affect microbe–microbe and microbe–host interactions and potentially change functionality or stability of the lichen-associated bacterial communities and thereby influence host health and ecosystem functioning, as proposed for plant–microbiomes (Agler et al., 2016; van der Heijden and Hartmann, 2016; Aydogan et al., 2018).
Long-term warming decreased the relative abundance of ASVs belonging to the Acidobacterial genera Granulicella and Bryocella and the alphaproteobacterial genus Acidiphilium in the total bacterial communities. Acidiphilium and Granulicella have been observed in other lichen microbiomes (Bates et al., 2011; Pankratov, 2012; Park et al., 2016; Aschenbrenner et al., 2017). These genera are chemoorganotrophic or chemolithotrophic and might thus survive on C sources present in the lichen thallus. Granulicella encompasses several acidophilic, cold-adapted species described from tundra soil isolates (Männistö et al., 2012). It has hydrolytic properties such as the ability to degrade chitin (Pankratov and Dedysh, 2010; Pankratov, 2012; Park et al., 2016; Belova et al., 2018), which suggest a role for these bacteria in the degradation of senescing lichen thalli. While some ASVs of the genus Granulicella decreased in relative abundance, the increased potential activity might result in increased degradation of dead lichen material. In contrast, the decreased relative abundance and potential activity of Bryocella and Acidiphilium might result in slower degradation of dead lichen material. The genus Acidiphilium showed an increase in relative abundance in a moss microbiome in the same warming experiment (Klarenberg et al., 2019). This suggests that the responses of microbiome components to environmental change are at least in part dependent upon host vegetation identity rather than constituting a direct response of the bacteria themselves to extrinsic environmental factors. Thus leading to different outcomes for the various microbiomes within the same environment.
The alphaproteobacterial genera Acidisphaera, Sphingomonas, and Endobacter showed an increased relative abundance and potential activity with warming. Sphingomonas and Acidisphaera have been identified in other lichen bacterial communities (Cardinale et al., 2008). Endobacter is a poorly characterized genus of which only one species has been described (Ramirez-Bahena et al., 2013). Acidisphaera is chemoorganotrophic and contains bacteriochlorophyll (Hiraishi et al., 2000). Sphingomonas is known for its ability to degrade plant biomass, the utilization of recalcitrant matter in oligotrophic environments, and the use of sulfonated compounds as sources of C and sulfur (Aylward et al., 2013), which may be linked to the increase in litter abundance. The increase in relative abundance and potential activity of these genera in the warmed conditions might enhance C and nutrient availability in the lichen thalli.
Overall, all genera that dominated the bacterial community of C. islandica contained ASVs that were affected by the warming treatment in their relative abundances and potential metabolic activity. The genera that were affected in relative abundance are likely to play roles in nutrient recycling and supply in the lichen symbiosis. As most of these ASVs increased in relative abundance with warming, nutrient turnover in the lichen might be accelerated.
OTCs have been deployed to study warming effects in a wide range of ecosystems and plant responses correspond well to responses to natural climate warming (Hollister and Webber, 2000). We have shown that the OTC treatment leads to changes in the composition of the bacterial community associated with the lichen C. islandica, and that part of this change could be attributed to the increase in B. nana and litter abundance in the OTCs. This secondary effect of the OTC treatment may shield radiation from reaching the lichen layer or soil and thereby reduce the warming effect of the OTCs (Bokhorst et al., 2013), reducing the amount of PAR reaching the lichen and potentially affecting the bacterial communities. The increase in B. nana leaf litter may also increase C turnover as it is easily decomposable (McLaren et al., 2017) and thereby influence the lichen bacterial community. It should be noted that multiple caveats can be associated with the use of OTCs. Snow trapped in OTCs can increase temperatures at the soil surface, but at the same time decrease photosynthetically active radiation (Bokhorst et al., 2013). The walls of the OTCs may act as a barrier for new species to arrive (Richardson et al., 2000), even though it is unknown how important this side effect of the OTCs is on microbial communities. In addition, temperature-induced changes in lichen traits such as thallus nutrient content, as well as soil organic matter content, and soil moisture are environmental factors that could potentially influence the lichen bacterial community composition. Warming could also affect the secondary metabolites of the lichen (Asplund et al., 2017) and thereby alter the composition of the lichen microbiota.
In conclusion, we found that the bacterial community of C. islandica was dominated by acidophilic taxa and harbored rare, but potentially active taxa. Our results also showed that twenty years of warming and an increase dwarf-shrub and litter abundance can lead to changes in lichen bacterial communities at a fine taxonomic level as well as richness and diversity. The lichen microbiome plays an important role in the growth of lichens and climate-driven changes in the lichen microbiota, irrespective of whether they are due to direct or indirect effects of climate change, might affect decomposition of lichens and thereby nutrient cycling in sub-Arctic ecosystems.
The original contributions presented in the study are publicly available. This data can be found in NCBI, under accession PRJEB37116.
IJ, IK, and OV designed the study. IK conducted the sampling, the laboratory work, the bioinformatics processing, and the statistical analysis. CK performed the qPCR measurements. IK wrote the manuscript with contributions from OV, IJ, CK, and DW. All authors contributed to the article and approved the submitted version.
This work was funded by the MicroArctic Innovative Training Network grant supported by the European Commissions’s Horizon 2020 Marie Sklodowska-Curie Actions program under project number 675546.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We kindly thank Margrét Auður Sigurbjörnsdóttir for providing insightful comments on the manuscript.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2020.540404/full#supplementary-material
Agler, M. T., Ruhe, J., Kroll, S., Morhenn, C., Kim, S.-T., and Weigel, D., et al. (2016). Microbial hub taxa link host and abiotic factors to plant microbiome variation. PLoS Biol. 14:e1002352. doi: 10.1371/journal.pbio.1002352
Alatalo, J. M., Jägerbrand, A. K., Chen, S., and Molau, U. (2017). Responses of lichen communities to 18 years of natural and experimental warming. Ann. Bot. 120, 159–170. doi: 10.1093/aob/mcx053
Almendras, K., García, J., Carú, M., and Orlando, J. (2018). Nitrogen-fixing bacteria associated with peltigera cyanolichens and cladonia chlorolichens. Molecules 23:3077. doi: 10.3390/molecules23123077
Anderson, M. J. (2001). A new method for non-parametric multivariate analysis of variance. Austral. Ecol. 26, 32–46. doi: 10.1111/j.1442-9993.2001.01070.pp.x
Aschenbrenner, I. A., Cernava, T., Berg, G., and Grube, M. (2016). Understanding microbial multi-species symbioses. Front. Microbiol. 7:180. doi: 10.3389/fmicb.2016.00180
Aschenbrenner, I. A., Cernava, T., Erlacher, A., Berg, G., and Grube, M. (2017). Differential sharing and distinct co-occurrence networks among spatially close bacterial microbiota of bark, mosses and lichens. Mol. Ecol. 26, 2826–2838. doi: 10.1111/mec.14070
Asplund, J., Siegenthaler, A., and Gauslaa, Y. (2017). Simulated global warming increases usnic acid but reduces perlatolic acid in the mat-forming terricolous lichen Cladonia stellaris. Lichenologist 49, 269–274. doi: 10.1017/S0024282917000159
Asplund, J., and Wardle, D. A. (2017). How lichens impact on terrestrial community and ecosystem properties. Biol. Rev. 92, 1720–1738. doi: 10.1111/brv.12305
Aydogan, E. L., Moser, G., Müller, C., Kämpfer, P., and Glaeser, S. P. (2018). Long-term warming shifts the composition of bacterial communities in the phyllosphere of galium album in a permanent grassland field-experiment. Front. Microbiol. 9:144. doi: 10.3389/fmicb.2018.00144
Aylward, F. O., McDonald, B. R., Adams, S. M., Valenzuela, A., Schmidt, R. A., Goodwin, L. A., et al. (2013). Comparison of 26 sphingomonad genomes reveals diverse environmental adaptations and biodegradative capabilities. Appl. Environ. Microbiol. 79, 3724–3733. doi: 10.1128/AEM.00518-13
Baldrian, P., Kolařík, M., Štursová, M., Kopecký, J., Valášková, V., Větrovský, T., et al. (2012). Active and total microbial communities in forest soil are largely different and highly stratified during decomposition. ISME J. 6, 248–258. doi: 10.1038/ismej.2011.95
Bastida, F., Torres, I. F., Andrés-Abellán, M., Baldrian, P., López-Mondéjar, R., Větrovský, T., et al. (2017). Differential sensitivity of total and active soil microbial communities to drought and forest management. Glob. Change Biol. 23, 4185–4203. doi: 10.1111/gcb.13790
Bates, S. T., Cropsey, G. W. G., Caporaso, J. G., Knight, R., and Fierer, N. (2011). Bacterial communities associated with the lichen symbiosis. Appl. Environ. Microbiol. 77, 1309–1314. doi: 10.1128/AEM.02257-10
Belova, S. E., Ravin, N. V., Pankratov, T. A., Rakitin, A. L., Ivanova, A. A., Beletsky, A. V., et al. (2018). Hydrolytic capabilities as a key to environmental success: chitinolytic and cellulolytic acidobacteria from acidic sub-arctic soils and boreal peatlands. Front. Microbiol. 9:2775. doi: 10.3389/fmicb.2018.02775
Biasi, C., Meyer, H., Rusalimova, O., Hämmerle, R., Kaiser, C., Baranyi, C., et al. (2008). Initial effects of experimental warming on carbon exchange rates, plant growth and microbial dynamics of a lichen-rich dwarf shrub tundra in Siberia. Plant Soil 307, 191–205. doi: 10.1007/s11104-008-9596-2
Bjelland, T., Grube, M., Hoem, S., Jorgensen, S. L., Daae, F. L., Thorseth, I. H., et al. (2011). Microbial metacommunities in the lichen-rock habitat: microbial metacommunities in the lichen-rock habitat. Environ. Microbiol. Rep. 3, 434–442. doi: 10.1111/j.1758-2229.2010.00206.x
Blazewicz, S. J., Barnard, R. L., Daly, R. A., and Firestone, M. K. (2013). Evaluating rRNA as an indicator of microbial activity in environmental communities: limitations and uses. ISME J. 7, 2061–2068. doi: 10.1038/ismej.2013.102
Bokhorst, S., Huiskes, A., Aerts, R., Convey, P., Cooper, E. J., Dalen, L., et al. (2013). Variable temperature effects of open top chambers at polar and alpine sites explained by irradiance and snow depth. Glob. Change Biol. 19, 64–74. doi: 10.1111/gcb.12028
Bradford, M. A., Davies, C. A., Frey, S. D., Maddox, T. R., Melillo, J. M., Mohan, J. E., et al. (2008). Thermal adaptation of soil microbial respiration to elevated temperature. Ecol. Lett. 11, 1316–1327. doi: 10.1111/j.1461-0248.2008.01251.x
Callahan, B. J., McMurdie, P. J., and Holmes, S. P. (2017). Exact sequence variants should replace operational taxonomic units in marker-gene data analysis. ISME J. 11, 2639–2643. doi: 10.1038/ismej.2017.119
Callahan, B. J., McMurdie, P. J., Rosen, M. J., Han, A. W., Johnson, A. J. A., and Holmes, S. P. (2016). DADA2: high-resolution sample inference from Illumina amplicon data. Nat. Methods 13, 581–583. doi: 10.1038/nmeth.3869
Campbell, B. J., Yu, L., Heidelberg, J. F., and Kirchman, D. L. (2011). Activity of abundant and rare bacteria in a coastal ocean. PNAS 108, 12776–12781. doi: 10.1073/pnas.1101405108
Cardinale, M., Grube, M., Castro, J. V., Müller, H., and Berg, G. (2012a). Bacterial taxa associated with the lung lichen Lobaria pulmonaria are differentially shaped by geography and habitat. FEMS Microbiol. Lett. 329, 111–115. doi: 10.1111/j.1574-6968.2012.02508.x
Cardinale, M., Puglia, A. M., and Grube, M. (2006). Molecular analysis of lichen-associated bacterial communities: lichen-associated bacterial communities. FEMS Microbiol. Ecol. 57, 484–495. doi: 10.1111/j.1574-6941.2006.00133.x
Cardinale, M., Steinová, J., Rabensteiner, J., Berg, G., and Grube, M. (2012b). Age, sun and substrate: triggers of bacterial communities in lichens: ecology of lichen-associated bacterial communities. Environ. Microbiol. Rep. 4, 23–28. doi: 10.1111/j.1758-2229.2011.00272.x
Cardinale, M., Vieira de Castro, J., Müller, H., Berg, G., and Grube, M. (2008). In situ analysis of the bacterial community associated with the reindeer lichen Cladonia arbuscula reveals predominance of Alphaproteobacteria: lichen-associated bacterial community. FEMS Microbiol. Ecol. 66, 63–71. doi: 10.1111/j.1574-6941.2008.00546.x
Cernava, T., Aschenbrenner, I. A., Soh, J., Sensen, C. W., Grube, M., and Berg, G. (2019). Plasticity of a holobiont: desiccation induces fasting-like metabolism within the lichen microbiota. ISME J. 13:547. doi: 10.1038/s41396-018-0286-7
Cernava, T., Vasfiu, Q., Erlacher, A., Aschenbrenner, I. A., Francesconi, K., Grube, M., et al. (2018). Adaptions of lichen microbiota functioning under persistent exposure to arsenic contamination. Front. Microbiol. 9:2959. doi: 10.3389/fmicb.2018.02959
Coleine, C., Stajich, J. E., Pombubpa, N., Zucconi, L., Onofri, S., Canini, F., et al. (2019). Altitude and fungal diversity influence the structure of Antarctic cryptoendolithic Bacteria communities. Environ. Microbiol. Rep. 11, 718–726. doi: 10.1111/1758-2229.12788
Colesie, C., Büdel, B., Hurry, V., and Green, T. G. A. (2018). Can Antarctic lichens acclimatize to changes in temperature? Glob. Change Biol. 24, 1123–1135. doi: 10.1111/gcb.13984
Cornelissen, J. H. C., Callaghan, T. V., Alatalo, J. M., Michelsen, A., Graglia, E., Hartley, A. E., et al. (2001). Global change and arctic ecosystems: is lichen decline a function of increases in vascular plant biomass? J. Ecol. 89, 984–994. doi: 10.1111/j.1365-2745.2001.00625.x
Cornelissen, J. H. C., Lang, S. I., Soudzilovskaia, N. A., and During, H. J. (2007). Comparative cryptogam ecology: a review of bryophyte and lichen traits that drive biogeochemistry. Ann. Bot. 99, 987–1001. doi: 10.1093/aob/mcm030
Crowther, T. W., and Bradford, M. A. (2013). Thermal acclimation in widespread heterotrophic soil microbes. Ecol. Lett. 16, 469–477. doi: 10.1111/ele.12069
De Caceres, M., and Legendre, P. (2009). Associations between species and groups of sites: indices and statistical inference. Ecology 90, 3566–3574. doi: 10.1890/08-1823.1
Ding, L. (2004). Proposals of Curvibacter gracilis gen. nov., sp. nov. and Herbaspirillum putei sp. nov. for bacterial strains isolated from well water and reclassification of [Pseudomonas] huttiensis, [Pseudomonas] lanceolata, [Aquaspirillum] delicatum and [Aquaspirillum] autotrophicum as Herbaspirillum huttiense comb. nov., Curvibacter lanceolatus comb. nov., Curvibacter delicatus comb. nov. and Herbaspirillum autotrophicum comb. nov. Int. J. Syst. Evol. Microbiol. 54, 2223–2230. doi: 10.1099/ijs.0.02975-0
Domaschke, S., Vivas, M., Sancho, L. G., and Printzen, C. (2013). Ecophysiology and genetic structure of polar versus temperate populations of the lichen Cetraria aculeata. Oecologia 173, 699–709. doi: 10.1007/s00442-013-2670-3
Elmendorf, S. C., Henry, G. H. R., Hollister, R. D., Björk, R. G., Bjorkman, A. D., Callaghan, T. V., et al. (2012). Global assessment of experimental climate warming on tundra vegetation: heterogeneity over space and time: warming effects on tundra vegetation. Ecol. Lett. 15, 164–175. doi: 10.1111/j.1461-0248.2011.01716.x
Fraser, R. H., Lantz, T. C., Olthof, I., Kokelj, S. V., and Sims, R. A. (2014). Warming-induced shrub expansion and lichen decline in the Western Canadian Arctic. Ecosystems 17, 1151–1168. doi: 10.1007/s10021-014-9783-3
González, I., Ayuso-Sacido, A., Anderson, A., and Genilloud, O. (2005). Actinomycetes isolated from lichens: evaluation of their diversity and detection of biosynthetic gene sequences. FEMS Microbiol. Ecol. 54, 401–415. doi: 10.1016/j.femsec.2005.05.004
Grube, M., Cardinale, M., de Castro, J. V., Müller, H., and Berg, G. (2009). Species-specific structural and functional diversity of bacterial communities in lichen symbioses. ISME J. 3, 1105–1115. doi: 10.1038/ismej.2009.63
Grube, M., Cernava, T., Soh, J., Fuchs, S., Aschenbrenner, I. A., Lassek, C., et al. (2015). Exploring functional contexts of symbiotic sustain within lichen-associated bacteria by comparative omics. ISME J. 9, 412–424. doi: 10.1038/ismej.2014.138
Hadfield, J. D. (2010). MCMC methods for multi-response generalized linear mixed models: the MCMCglmm R package. J. Stat. Softw. 33, 1–22. doi: 10.18637/jss.v033.i02
Hiraishi, A., Matsuzawa, Y., Kanbe, T., and Wakao, N. (2000). Acidisphaera rubrifaciens gen. nov., sp. nov., an aerobic bacteriochlorophyll-containing bacterium isolated from acidic environments. Int. J. Syst. Evol. Microbiol. 50, 1539–1546. doi: 10.1099/00207713-50-4-1539
Hodkinson, B. P., Gottel, N. R., Schadt, C. W., and Lutzoni, F. (2012). Photoautotrophic symbiont and geography are major factors affecting highly structured and diverse bacterial communities in the lichen microbiome: prokaryotic communities of the lichen microbiome. Environ. Microbiol. 14, 147–161. doi: 10.1111/j.1462-2920.2011.02560.x
Hodkinson, B. P., and Lutzoni, F. (2009). A microbiotic survey of lichen-associated bacteria reveals a new lineage from the Rhizobiales. Symbiosis 49, 163–180. doi: 10.1007/s13199-009-0049-3
Holland-Moritz, H., Stuart, J., Lewis, L. R., Miller, S., Mack, M. C., McDaniel, S. F., et al. (2018). Novel bacterial lineages associated with boreal moss species. Environ. Microbiol. 20, 2625–2638. doi: 10.1111/1462-2920.14288
Hollister, R. D., and Webber, P. J. (2000). Biotic validation of small open-top chambers in a tundra ecosystem. Glob. Change Biol. 6, 835–842. doi: 10.1046/j.1365-2486.2000.00363.x
IPCC (2019). Special Report on the Ocean and Cryosphere in a Changing Climate (SROCC). Geneva: IPCC.
Jia, Y., Leung, M. H. Y., Tong, X., Wilkins, D., and Lee, P. K. H. (2019). Rare taxa exhibit disproportionate cell-level metabolic activity in enriched anaerobic digestion microbial communities. mSystems 4, e208–e218. doi: 10.1128/mSystems.00208-18
Jones, S. E., and Lennon, J. T. (2010). Dormancy contributes to the maintenance of microbial diversity. PNAS 107, 5881–5886. doi: 10.1073/pnas.0912765107
Jonsdottir, I. S., Magnusson, B., Gudmundsson, J., Elmarsdottir, A., and Hjartarson, H. (2005). Variable sensitivity of plant communities in Iceland to experimental warming. Glob. Change Biol. 11, 553–563. doi: 10.1111/j.1365-2486.2005.00928.x
Jousset, A., Bienhold, C., Chatzinotas, A., Gallien, L., Gobet, A., Kurm, V., et al. (2017). Where less may be more: how the rare biosphere pulls ecosystems strings. ISME J. 11, 853–862. doi: 10.1038/ismej.2016.174
Kärnefelt, I., Mattsson, J.-E., and Thell, A. (1993). The lichen genera arctocetraria, cetraria, and cetrariella (parmeliaceae) and their presumed evolutionary affinities. Bryologist 96, 394–404. doi: 10.2307/3243869
Klarenberg, I. J., Keuschnig, C., Colmenares, A. J. R., Jungblut, A. D., Jónsdóttir, I. S., and Vilhelmsson, O. (2019). Long-term warming effects on the microbiome and nitrogen fixation associated with the moss Racomitrium lanuginosum in a subarctic alpine heathland. bioRxiv [Preprint]. doi: 10.1101/838581
Klein, A. M., Bohannan, B. J. M., Jaffe, D. A., Levin, D. A., and Green, J. L. (2016). Molecular evidence for metabolically active bacteria in the atmosphere. Front. Microbiol. 7:772. doi: 10.3389/fmicb.2016.00772
Klindworth, A., Pruesse, E., Schweer, T., Peplies, J., Quast, C., Horn, M., et al. (2013). Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 41:e1. doi: 10.1093/nar/gks808
Kobayashi, D. Y., and Crouch, J. A. (2009). Bacterial/fungal interactions: from pathogens to mutualistic endosymbionts. Annu. Rev. Phytopathol. 47, 63–82. doi: 10.1146/annurev-phyto-080508-081729
Lange, O. L., and Green, T. G. A. (2005). Lichens show that fungi can acclimate their respiration to seasonal changes in temperature. Oecologia 142, 11–19. doi: 10.1007/s00442-004-1697-x
Lawson, C. E., Strachan, B. J., Hanson, N. W., Hahn, A. S., Hall, E. R., Rabinowitz, B., et al. (2015). Rare taxa have potential to make metabolic contributions in enhanced biological phosphorus removal ecosystems. Environ. Microbiol. 17, 4979–4993. doi: 10.1111/1462-2920.12875
Longton, R. (1992). “The role of bryophytes and lichens in terrestrial ecosystems,” in Bryophytes and Lichens in a Changing Environment, eds J. Bates and A. M. Farmer (Oxford: Oxford University Press), 32–76. doi: 10.1017/cbo9780511565212.003
Love, M. I., Huber, W., and Anders, S. (2014). Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 15:550. doi: 10.1186/s13059-014-0550-8
Männistö, M. K., Rawat, S., Starovoytov, V., and Häggblom, M. M. (2012). Granulicella arctica sp. nov., Granulicella mallensis sp. nov., Granulicella tundricola sp. nov. and Granulicella sapmiensis sp. nov., novel acidobacteria from tundra soil. Int. J. Syst. Evol. Microbiol. 62, 2097–2106. doi: 10.1099/ijs.0.031864-0
McLaren, J. R., Buckeridge, K. M., van de Weg, M. J., Shaver, G. R., Schimel, J. P., and Gough, L. (2017). Shrub encroachment in Arctic tundra: Betula nana effects on above- and belowground litter decomposition. Ecology 98, 1361–1376. doi: 10.1002/ecy.1790
McMurdie, P. J., and Holmes, S. (2013). phyloseq: an R package for reproducible interactive analysis and graphics of microbiome census data. PLoS One 8:e61217. doi: 10.1371/journal.pone.0061217
Muggia, L., Klug, B., Berg, G., and Grube, M. (2013). Localization of bacteria in lichens from Alpine soil crusts by fluorescence in situ hybridization. Appl. Soil Ecol. 68, 20–25. doi: 10.1016/j.apsoil.2013.03.008
Mushegian, A. A., Peterson, C. N., Baker, C. C. M., and Pringle, A. (2011). Bacterial diversity across individual lichens. Appl. Environ. Microbiol. 77, 4249–4252. doi: 10.1128/AEM.02850-10
Myers-Smith, I. H., Grabowski, M. M., Thomas, H. J. D., Angers-Blondin, S., Daskalova, G. N., Bjorkman, A. D., et al. (2019). Eighteen years of ecological monitoring reveals multiple lines of evidence for tundra vegetation change. Ecol. Monogr. 89:e01351. doi: 10.1002/ecm.1351
Nash, T. H., and Olafsen, A. G. (1995). Climate change and the ecophysiological response of Arctic lichens. Lichenologist 27, 559–565. doi: 10.1016/s0024-2829(05)80014-3
Oksanen, J., Blanchet, F. G., Kindt, R., Legendre, P., Minchin, P. R., O’hara, R. B., et al. (2013). Package ‘vegan.’ Community ecology package, version 2, 296.
Pankratov, T. A. (2012). Acidobacteria in microbial communities of the bog and tundra lichens. Microbiology 81, 51–58. doi: 10.1134/S0026261711060166
Pankratov, T. A., and Dedysh, S. N. (2010). Granulicella paludicola gen. nov., sp. nov., Granulicella pectinivorans sp. nov., Granulicella aggregans sp. nov. and Granulicella rosea sp. nov., acidophilic, polymer-degrading acidobacteria from Sphagnum peat bogs. Int. J. Syst. Evol. Microbiol. 60, 2951–2959. doi: 10.1099/ijs.0.021824-0
Park, C. H., Kim, K. M., Kim, O.-S., Jeong, G., and Hong, S. G. (2016). Bacterial communities in Antarctic lichens. Antarctic Sci. 28, 455–461. doi: 10.1017/S0954102016000286
Paulson, J. N., Stine, O. C., Bravo, H. C., and Pop, M. (2013). Differential abundance analysis for microbial marker-gene surveys. Nat. Methods 10, 1200–1202. doi: 10.1038/nmeth.2658
Poly, F., Monrozier, L. J., and Bally, R. (2001). Improvement in the RFLP procedure for studying the diversity of nifH genes in communities of nitrogen fixers in soil. Res. Microbiol. 152, 95–103. doi: 10.1016/S0923-2508(00)01172-4
Printzen, C., Fernández-Mendoza, F., Muggia, L., Berg, G., and Grube, M. (2012). Alphaproteobacterial communities in geographically distant populations of the lichen Cetraria aculeata. FEMS Microbiol. Ecol. 82, 316–325. doi: 10.1111/j.1574-6941.2012.01358.x
Quast, C., Pruesse, E., Yilmaz, P., Gerken, J., Schweer, T., Yarza, P., et al. (2013). The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res. 41, D590–D596. doi: 10.1093/nar/gks1219
Ramirez-Bahena, M. H., Tejedor, C., Martin, I., Velazquez, E., and Peix, A. (2013). Endobacter medicaginis gen. nov., sp. nov., isolated from alfalfa nodules in an acidic soil. Int. J. Syst. Evol. Microbiol. 63, 1760–1765. doi: 10.1099/ijs.0.041368-0
Richardson, S. J., Hartley, S. E., and Press, M. C. (2000). Climate warming experiments: are tents a potential barrier to interpretation? Ecol. Entomol. 25, 367–370. doi: 10.1046/j.1365-2311.2000.00263.x
Rolshausen, G., Grande, F. D., Sadowska-Deś, A. D., Otte, J., and Schmitt, I. (2018). Quantifying the climatic niche of symbiont partners in a lichen symbiosis indicates mutualist-mediated niche expansions. Ecography 41, 1380–1392. doi: 10.1111/ecog.03457
Romero-Olivares, A. L., Allison, S. D., and Treseder, K. K. (2017). Soil microbes and their response to experimental warming over time: a meta-analysis of field studies. Soil Biol. Biochem. 107, 32–40. doi: 10.1016/j.soilbio.2016.12.026
Sigurbjörnsdóttir, M. A., Andrésson, ÓS., and Vilhelmsson, O. (2015). Analysis of the Peltigera membranacea metagenome indicates that lichen-associated bacteria are involved in phosphate solubilization. Microbiology 161, 989–996. doi: 10.1099/mic.0.000069
Spribille, T., Tuovinen, V., Resl, P., Vanderpool, D., Wolinski, H., Aime, M. C., et al. (2016). Basidiomycete yeasts in the cortex of ascomycete macrolichens. Science 353, 488–492. doi: 10.1126/science.aaf8287
Torres-Cruz, T. J., Howell, A. J., Reibold, R. H., McHugh, T. A., Eickhoff, M. A., and Reed, S. C. (2018). Species-specific nitrogenase activity in lichen-dominated biological soil crusts from the Colorado Plateau, USA. Plant Soil 429, 113–125. doi: 10.1007/s11104-018-3580-2
Uphof, J. C. T. (1925). Purple bacteria as symbionts of a lichen. Science 61:67. doi: 10.1126/science.61.1568.67
van der Heijden, M. G. A., and Hartmann, M. (2016). Networking in the plant microbiome. PLoS Biol. 14:e1002378. doi: 10.1371/journal.pbio.1002378
Wang, Q., Garrity, G. M., Tiedje, J. M., and Cole, J. R. (2007). Naive Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl. Environ. Microbiol. 73, 5261–5267. doi: 10.1128/AEM.00062-07
Wedin, M., Maier, S., Fernandez-Brime, S., Cronholm, B., Westberg, M., and Grube, M. (2016). Microbiome change by symbiotic invasion in lichens: microbiome change by symbiotic invasion in lichens. Environ. Microbiol. 18, 1428–1439. doi: 10.1111/1462-2920.13032
Weiss, S. T., Cropsey, G. W. G., Caporaso, J. G., Knight, R., and Fierer, N. (2011). Bacterial communities associated with the lichen symbiosis. Appl. Environ. Microbiol. 77, 1309–1314. doi: 10.1128/AEM.02257-10
West, N. J., Parrot, D., Fayet, C., Grube, M., Tomasi, S., and Suzuki, M. T. (2018). Marine cyanolichens from different littoral zones are associated with distinct bacterial communities. PeerJ 6:e5208. doi: 10.7717/peerj.5208
Keywords: lichen, lichen microbiome, tundra, climate change, host–microbiome, lichen-associated bacteria, long-term warming
Citation: Klarenberg IJ, Keuschnig C, Warshan D, Jónsdóttir IS and Vilhelmsson O (2020) The Total and Active Bacterial Community of the Chlorolichen Cetraria islandica and Its Response to Long-Term Warming in Sub-Arctic Tundra. Front. Microbiol. 11:540404. doi: 10.3389/fmicb.2020.540404
Received: 04 March 2020; Accepted: 30 November 2020;
Published: 18 December 2020.
Edited by:
David Anthony Pearce, Northumbria University, United KingdomReviewed by:
Soon Gyu Hong, Korea Polar Research Institute, South KoreaCopyright © 2020 Klarenberg, Keuschnig, Warshan, Jónsdóttir and Vilhelmsson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ingeborg J. Klarenberg, aW5nZWJvcmcua2xhcmVuYmVyZ0BnbWFpbC5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.