
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 17 December 2020
Sec. Antimicrobials, Resistance and Chemotherapy
Volume 11 - 2020 | https://doi.org/10.3389/fmicb.2020.517462
This article is part of the Research Topic Discovery of novel plant-derived compounds with antibacterial actions against antibiotic-resistant bacteria View all 24 articles
 Weifeng Yang1†
Weifeng Yang1† Qing Wei2,5†
Qing Wei2,5† Qian Tong3
Qian Tong3 Kaiyu Cui1
Kaiyu Cui1 Gaiying He1Longfei Lin4Lvyan Z. Ma5
Gaiying He1Longfei Lin4Lvyan Z. Ma5 Pierre Cornelis6,7Yi Wang1*
Pierre Cornelis6,7Yi Wang1*Pseudomonas aeruginosa is an opportunistic pathogen that can infect a wide variety of hosts including humans, plants, and animals. The production of virulence factors is the determinant of the infection paradigm and is under orchestrated regulation via cell-to-cell communication process called quorum sensing (QS). To disable QS circuits and prevent bacterial infections, a large battery of anti-QS agents, particularly from traditional Chinese medicine have been developed. Here, we used P. aeruginosa as a model microorganism to investigate the effect of traditional Chinese medicine Tanreqing (TRQ) formula on bacterial pathogenicity. Phenotypic analysis showed that TRQ treatment could completely inhibit the production of phenazine pyocyanin and moderately inhibit the production of virulence factors such as rhamnolipids, elastase, and alkaline protease. Further transcriptomic analyses revealed that TRQ treatment could significantly attenuate the expression of QS-regulated genes in P. aeruginosa and TRQ-treated P. aeruginosa regulon shared a large overlap with QS regulon. Component contribution to QS inhibition shed light on the indispensable role of all five components in TRQ formula. Further genetic analysis indicated that upstream regulators of QS systems, including two-component systems GacS/GacA and PprA/PprB, were both inhibited by TRQ treatment. Finally, our TRQ formula could efficiently protect Caenorhabditis elegans from killing by P. aeruginosa. Altogether, we have proved TRQ formula as an effective and specific agent to attenuate bacterial virulence and combat bacterial infections.
Pseudomonas aeruginosa is an important human opportunistic pathogen that causes a panel of acute infections such as urinary tract infections, acute ulcerative keratitis and burn wound infections as well as pulmonary infections, as often seen in cystic fibrosis patients (Lyczak et al., 2000, 2002; Haussler, 2004). This bacterium is noted for its production of a large number of extracellular virulence factors including the phenazine pigment pyocyanin, lectins, siderophores, proteases and rhamnolipids that are required to initiate a successful acute infection (Jimenez et al., 2012). To efficiently coordinate these virulence gene expression, P. aeruginosa employs a ubiquitous bacterial cell-to-cell communication process called quorum sensing (QS) to produce, secrete, and detect the chemical signaling molecules termed autoinducers in response to fluctuations in cell population density (Camilli and Bassler, 2006; Papenfort and Bassler, 2016).
Two canonical LuxI/LuxR QS circuits (LasI/LasR and RhlI/RhlR) exist in P. aeruginosa (Schuster and Greenberg, 2006; Venturi, 2006). Both LasI and RhlI are autoinducer synthases that catalyze the formation of signal molecule N-3-(oxododecanoyl)-homoserine lactone (3-oxo-C12-HSL) and N-butyryl homoserine lactone (C4-HSL), respectively. LasR and RhlR function both to bind their respective autoinducers and to activate the transcription of downstream genes (Miller and Bassler, 2001). The two regulatory circuits act in tandem to control the expression of a large number of P. aeruginosa virulence factors (Parsek and Greenberg, 2000; Papenfort and Bassler, 2016).
In addition to classical autoinducers, P. aeruginosa produces quinolone signals including 2-heptyl-4-quinolone (HHQ) and 2-heptyl-3-hydroxy-4-quinolone (PQS) to regulate virulence gene expression (Diggle et al., 2006, 2007; Heeb et al., 2011). Both HHQ and PQS regulate gene expression via acting as the ligand of the MvfR regulator, also known as PqsR (Deziel et al., 2005; Xiao et al., 2006; Diggle et al., 2007). Although a fourth QS signal molecule, termed IQS, has been suggested (Lee et al., 2013), it later appeared that IQS is in fact aeruginaldehyde, a by-product of the siderophore pyochelin biosynthesis pathway and not of the AmbABCDE as proposed previously (Ye et al., 2014; Rojas Murcia et al., 2015; Cornelis, 2019). A yet uncharacterized autoinducer produced by PqsE that interacts with RhlR has been recently reported (Mukherjee et al., 2018).
In P. aeruginosa, QS systems not only act as central regulatory circuits for the expression of virulence factors, but also are involved in the development of biofilms, sessile communities of bacterial cells and the causative agent of chronic infections (Davies et al., 1998). Therefore, QS systems represent ideal and attractive targets for the design of novel therapeutics (Rasmussen and Givskov, 2006). Extensive studies have been undertaken to search for novel anti-QS drugs that could override bacterial communication signals and investigate their potential roles in the attenuation of virulence in P. aeruginosa (Hentzer et al., 2003; Muh et al., 2006; Borlee et al., 2010; Tan et al., 2013; Starkey et al., 2014).
Up to now, almost all P. aeruginosa QS systems have corresponding anti-QS drugs that could reduce the virulence in in intro and in vivo infection models (Fan et al., 2013; O’Loughlin et al., 2013; Starkey et al., 2014; Li et al., 2018). For example, azithromycin, a macrolide antibiotic, has been shown to inhibit the QS system in P. aeruginosa through several mechanisms (Nalca et al., 2006; Skindersoe et al., 2008; Perez-Martinez and Haas, 2011). However, the development of resistance to these anti-QS compounds and concerns over the safety of synthetic chemicals have been viewed as serious side effects of these types of compounds (D’Costa et al., 2011; Wright, 2011). To resolve these global health threats and determine effective anti-QS compounds, the use of natural products, particularly traditional Chinese medicine (TCM), has become attractive due to their broad spectrum of secondary metabolites and low potential to develop resistance (Koh et al., 2013; Asfour, 2018; Chong et al., 2018).
The Tanreqing (TRQ) injection is a Chinese herbal preparation made from Scutellariae radix (Huang Qin in Chinese, HQ), Lonicerae flos (Jin Yin Hua in Chinese, JYH), Forsythiae fructus (Lian Qiao in Chinese, LQ), Ursi fel (Xiong Dan in Chinese, XD) and Naemorhedi cornu (Shan Yang Jiao in Chinese, SYJ). According to TCM theory, TRQ formula could clear heat, detoxify and remove phlegm and has been used in China as a treatment for respiratory tract infections, pneumonia and chronic obstructive pulmonary disease (Liu et al., 2016). However, the mode of action of TRQ is still not well understood in respect to its antibacterial activity.
In this study, we used P. aeruginosa as a model microorganism to investigate the inhibitory mechanism of TRQ against bacterial infections. We found that TRQ formula could completely inhibit the expression of the three QS systems in P. aeruginosa at sub-minimum inhibitory concentrations (sub-MIC) in vitro and in vivo, suggesting that TRQ is a highly specific and effective agent to attenuate bacterial virulence and combat bacterial infections.
Bacterial strains and plasmids used in this study are shown in Table 1. Both P. aeruginosa and E. coli strains were grown in Lysogeny broth (LB) with aeration at 37°C. When required, LB agar plates were used to streak bacterial colonies. In addition, antibiotics were added into the growth medium at the following concentrations: 300 μg ml–1 carbenicillin (Cb) for P. aeruginosa, 100 μg ml–1 ampicillin (Amp) for E. coli.

Table 1. Strains and plasmids used in this study.
Overnight cultures of bacterial strains in LB were diluted (1:100) in 3 ml LB medium (30 μl culture into 3 ml LB broth) and precultures incubated aerobically at 37°C in a shaker at 200 rpm to an OD600nm of 0.5. The precultures were further diluted (1:100) in 1 ml LB medium (10 μl precultures in 990 μl LB broth). Growth was then analyzed in 10 × 10-wells microtiter plates containing 294 μl LB medium to which 6 μl of diluted precultures containing 105 cells was added to obtain a final 1:5000 dilution. Control wells contained only the medium without preculture. TRQ treatment was prepared in a 1:4, 1:8, 1:16 dilution using LB medium. The microtiter plates were incubated for 72 h at 37°C in a Bioscreen incubator (Life Technologies, Finland) using the following settings: shaking for 20 s every 3 min and absorbance measured every 30 min at 600 nm. Each culture was prepared in triplicate.
Pyocyanin production was visualized by plating the bacteria on Pseudomonas-agar (P-agar or King’s A medium), followed by 48 h incubation. Pyocyanin production resulted in a deep blue coloration of the medium. Quantification of pyocyanin was modified from previous studies (Vinckx et al., 2010). Briefly, the agar was collected in a falcon tube and 10 ml of chloroform was added per 12.5 g of agar medium. The phenazine pigment was extracted during 2 h incubation at 37°C after which 2 ml 0.5 M HCl was added and shaken vigorously. The pink top layer was recovered, and its absorbance was measured at 520 nm.
Other QS-regulated virulence factors including rhamnolipid, elastase, and alkaline protease were detected using methods described previously (Aybey and Demirkan, 2016). TRQ was diluted with an 1/4 ratio in all assay medium and the inhibitory effect was analyzed against untreated controls. All experiments were repeated three times with three technical replicates.
The GFP reporter plasmid pProbe-AT′ was used to construct transcriptional fusions with the corresponding tested promoters (Miller et al., 2000). The promoter region was amplified by PCR using I-5 Hi-Fi DNA Hotstart polymerase (MCLAB). The DNA fragment was digested and cloned into digested pProbe-AT′. The resulting plasmids were mobilized into P. aeruginosa by conjugation using E. coli S17-1 as the donor strain. Bacteria with reporter fusion were growth at 37°C and data were recorded at 2 h intervals after treatment with TRQ. The promoter activity was evaluated by measuring GFP intensity values captured with Ex/Em 488/520 via calibrating with the corresponding OD600 (Synergy H4 Hybrid Reader, BioTek). The gene expression level was expressed as relative GFP intensity.
Animal virulence test was done according to the previously described method (Moy et al., 2006; Fito-Boncompte et al., 2011). Briefly, C. elegans wild-type Bristol strain N2 worms were grown at 20°C on nematode growth medium (NGM) agar plates using E. coli OP50 as the nutrient. Synchronous nematode cultures were achieved by exposure to a sodium hypochlorite-sodium hydroxide solution. The resulting eggs were then incubated at 20°C for 24 h to obtain L1 stage worms and transferred to fresh NGM plates to grow until the worms reached L4 life stage. P. aeruginosa cells were grown as described above in LB broth overnight and harvested. 109 bacteria cells were spread onto NGM plates and incubated at 37°C overnight. P. aeruginosa plates were cooled down to room temperature and 30 L4-synchronized worms infected with P. aeruginosa were placed on NGM plates with or without TRQ and incubated at 20°C for bacterial survival assays. C. elegans survival data were recorded at a regular interval of each day using an Motic K-400 LED Stereo microscope. Worms were considered dead if they were unresponsive to gentle touch with a platinum wire pick. An infection assay was performed with three independent replicates and repeated three times.
Overnight cultures of P. aeruginosa in LB were used to inoculate fresh LB medium in an 1:1000 dilution in the absence and presence of TRQ (1:4 dilution ratio). After 6 h (EXP) and 12 h (STA) of incubation, one ml of culture was immediately fixed with 2 ml of RNA Protect Reagent (Qiagen), following the manufacturer’s instructions, and the fixed cell pellets were frozen at −80°C until further use. All experiments were performed with three technical replicates. Total RNA was extracted using TRIzol® Reagent according to the manufacturer’s instructions (Invitrogen) and genomic DNA was removed using RNase-free DNase I (TaKaRa). Then RNA quality was determined using 2100 Bioanalyzer (Agilent) and quantified using the ND-2000 (NanoDrop Technologies). High-quality RNA sample (OD260/280 = 1.8∼2.2, OD260/230 ≥ 2.0, RIN ≥ 6.5, 28S:18S ≥ 1.0, >10 μg) was used to construct the sequencing library.
RNA-seq strand-specific libraries were prepared following TruSeq RNA sample preparation Kit from Illumina (San Diego, CA, United States), using 5 μg of total RNA. Briefly, rRNA was removed by RiboZero rRNA removal kit (Epicenter), fragmented using fragmentation buffer. cDNA synthesis, end repair, A-base addition and ligation of the Illumina-indexed adaptors were performed according to Illumina’s protocol. Libraries were then size selected for cDNA target fragments of 200∼300 bp on 2% Low Range Ultra Agarose followed by PCR amplified using Phusion DNA polymerase (NEB) for 15 PCR cycles. After quantification by TBS380 Mini-Fluorometer, paired-end libraries were sequenced by Shanghai Biozeron Biotechnology Co., Ltd. (Shanghai, China) with the Illumina HiSeq PE 2 × 151 bp read length.
The raw paired end reads were trimmed and quality controlled by Trimmomatic with default parameters (Bolger et al., 2014). Then clean reads were separately aligned to the reference genome (Pseudomonas aeruginosa PAO1, Accession number NC_002516) with orientation mode using Rockhopper software (McClure et al., 2013; Tjaden, 2015), which was a comprehensive and user-friendly system for computational analysis of bacterial RNA-seq data. As input, Rockhopper takes RNA sequencing reads generated by high-throughput sequencing technology to calculate gene expression levels with default parameters.
To identify DEGs (differential expression genes) between two different samples, the expression level for each transcript was calculated using the fragments per kilobase of read per million mapped reads (RPKM) method. The method edgeR was used for differential expression analysis (Robinson et al., 2010). The DEGs between two samples were selected using the following criteria: the logarithmic of fold change was greater than 2 and the false discovery rate (FDR) should be less than 0.05. To understand the functions of these differential expressed genes, GO functional enrichment and KEGG pathway analysis were carried out by Goatools (Klopfenstein et al., 2018) and KOBAS (Xie et al., 2011), respectively. DEGs were significantly enriched in GO terms and metabolic pathways when their Bonferroni-corrected P-value was less than 0.05. The RNA-seq datasets have been deposited in National Center for Biotechnology Information (NCBI) with an accession number GSE141753.
The data of virulence factor production, transcriptional analysis, and virulence test were analyzed by one-way ANOVA. Student’s t-test was used when one-way ANOVA revealed significant differences (P < 0.05). Survival data were analyzed via the Kaplan–Meier method and the log-rank test was used to compare the significant differences between subgroups (P < 0.01). All statistical analyses were performed with GraphPad Prism statistical software (GraphPad Software, La Jolla, CA, United States) with the assistance of Excel (Microsoft).
It has been established that TRQ could affect the viability of bacterial cells (Yang et al., 2018). Therefore, we measured the growth of P. aeruginosa PAO1 in the presence or absence of different concentrations of TRQ (1/4, 1/8, 1/16 dilution ratio). As can be seen from Figure 1A, we found that P. aeruginosa growth kinetics were unaffected at all tested concentrations, indicating that TRQ formula has no inhibitory effect on planktonic P. aeruginosa growth. It was also interesting to notice that when different concentrations of TRQ were added to P. aeruginosa cultures, we observed that the production of phenazine pyocyanin was greatly reduced. We further used Pseudomonas isolation agar as growth medium to determine the production of pyocyanin and detected that pyocyanin production was significantly decreased in both dose- and time-dependent manner by TRQ treatment (Figures 1B,C). Furthermore, we determined that virulence factors including rhamnolipid (Figure 1D), elastase (Figure 1E) and alkaline protease (Figure 1F) were all significantly repressed by TRQ treatment, indicating TRQ might function as an anti-virulence agent against P. aeruginosa.
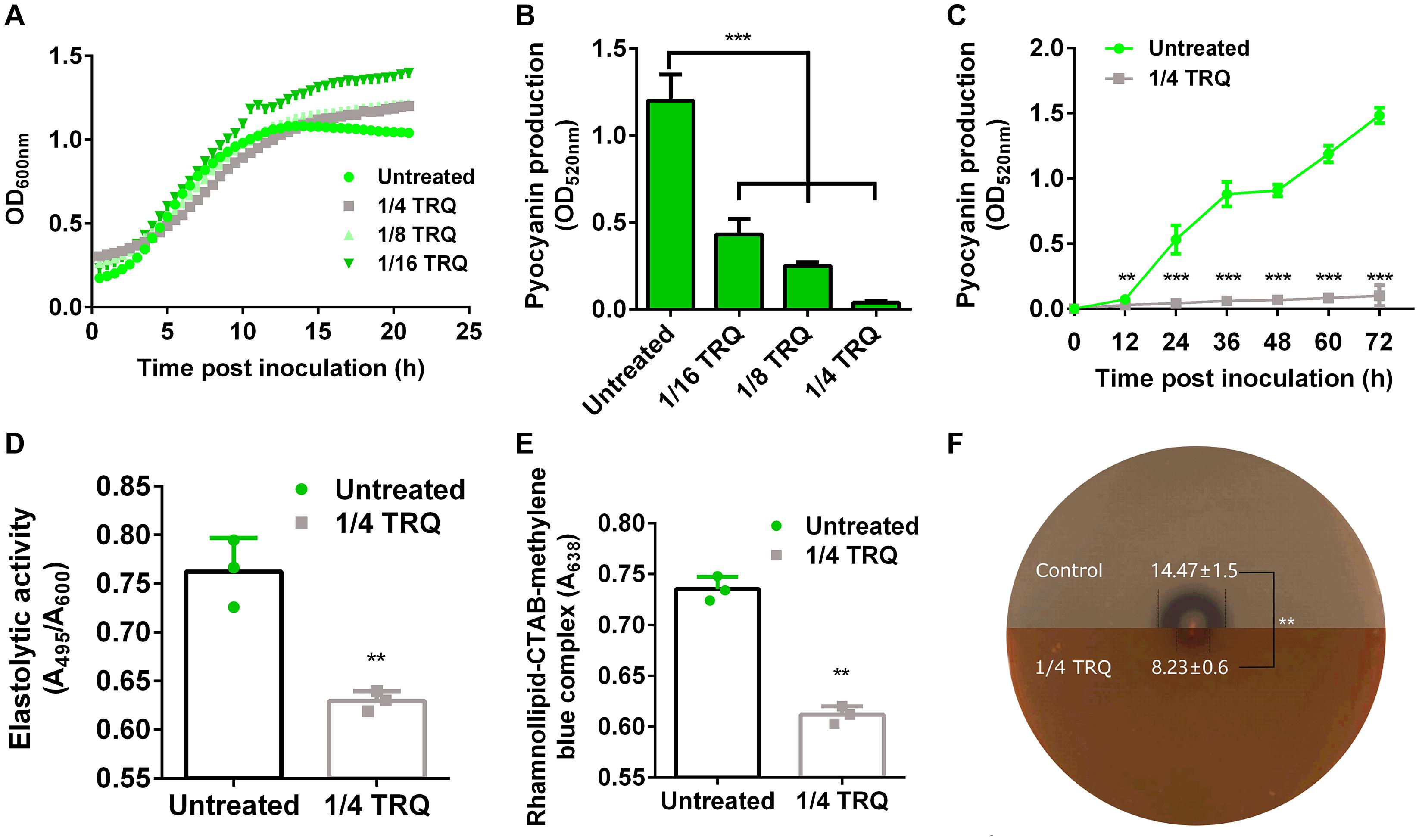
Figure 1. TRQ inhibits the production of virulence factors in P. aeruginosa. (A) TRQ treatment does not affect P. aeruginosa growth. (B) TRQ inhibits pyocyanin production in a dose-dependent manner. (C) TRQ inhibits pyocyanin production in a time-dependent manner. (D) Effect of TRQ on elastase activity. PAO1 was cultivated in LB medium at 37°C for 24 h in the presence of TRQ and then added to elastin-Congo red. Elastolytic activity was performed in elastin-Congo red buffer. (E) Effect of TRQ on rhamnolipid production. PAO1 was cultivated in fresh LB broth with and without TRQ at 37°C for 24 h and added to agar-free CTAB medium and incubated for 48 h. The amount of rhamnolipid-CTAB-methylene blue complex biomass in the presence of TRQ was determined. (F) Effect of TRQ on alkaline protease activity. The production of alkaline protease was measured using skimmed milk agar plate. A colony from overnight cultures of PAO1 was inoculated using a toothpick at the center of the plates and then incubated at 37°C for 12 h. The production of alkaline protease was confirmed by the formation of a clear zone around bacterial colony. The halo diameters were determined and shown in corresponding region (mm in unit). The upper half plate is untreated control and the bottom half plate is TRQ-containing plate. The dashed lines indicate the halo. Statistical analysis was based on pairwise comparisons (Student’s t-test). **P < 0.01; ***P < 0.001. Error bars represent the SD of three replicates.
Since the production of virulence factors such as pyocyanin are under the control of QS systems in P. aeruginosa, we therefore determined whether the expression of these QS systems was affected by TRQ treatment. Three QS signal receptors were chosen and their transcriptional fusions with GFP reporters were constructed to probe the relative expression level of these QS systems. As can be seen from Figure 2, the transcriptional expression of lasR, rhlR, and mvfR was all greatly repressed by TRQ treatment in a dose-dependent manner. Our results confirmed that TRQ could indeed inhibit the expression of QS systems in P. aeruginosa and thus inhibit the production of virulence factors such as pyocyanin.
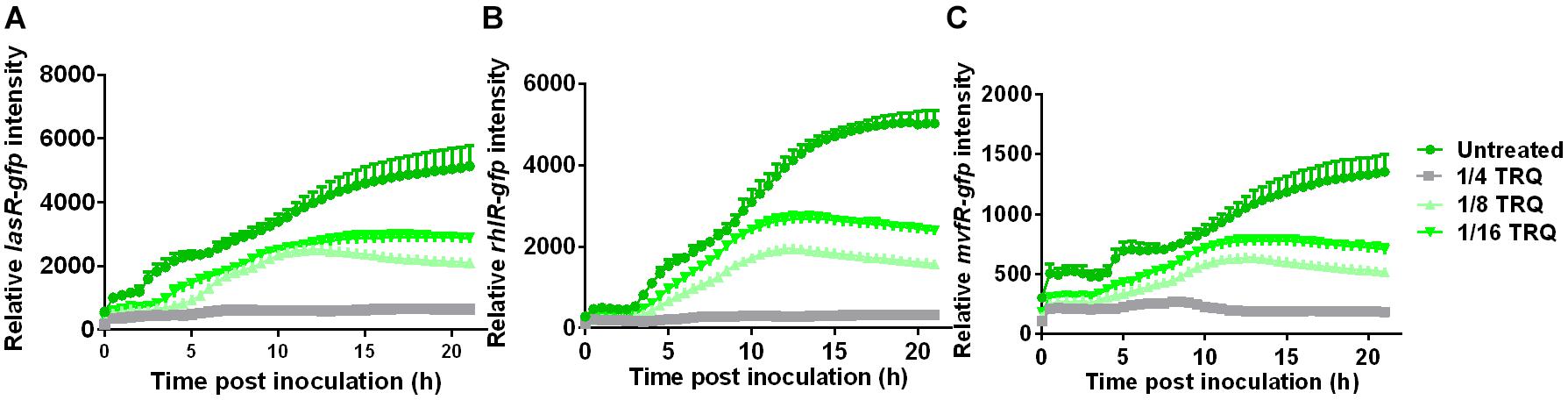
Figure 2. TRQ treatment inhibits the expression of QS regulator genes in P. aeruginosa. las-gfp expression analysis (A), rhlR-gfp expression analysis (B), and mvfR-gfp analysis (C) by TRQ treatment at different concentrations (untreated control, 1/4 TRQ, 1/8 TRQ, and 1/16 TRQ).
To gain insight into the regulatory breadth of TRQ treatment on P. aeruginosa, we carried out a global RNA-seq analysis of the transcriptional responses to determine the number of genes under both exponential (EXP) and stationary (STA) growth phases. P. aeruginosa cells were harvested, and RNA was extracted and processed according to the recommendations of the Illumina system for RNA-seq analyses. After data qualification control and processing, we finally obtained a comprehensive data set summarized in Supplementary Tables S1, S2. Clearly, PAO1 and TRQ-treated PAO1 demonstrated distinct gene expression profiles according to the heat map diagram (Figures 3A,B). A large number of genes were differentially expressed under both EXP and STA growth phases and showed overlap between these two conditions. In total, 371 genes were downregulated and 720 genes were upregulated in EXP and STA growth phases by TRQ treatment (Figure 3C). Functional classification analysis showed that genes encoding secreted factors and membrane proteins were largely represented as downregulated under both EXP and STA growth phases by TRQ treatment. In contrast, genes encoding carbon compound metabolism, transport of small molecules, and translation and post-translation modification were mostly up-regulated under EXP and STA growth phases (Figure 3D).
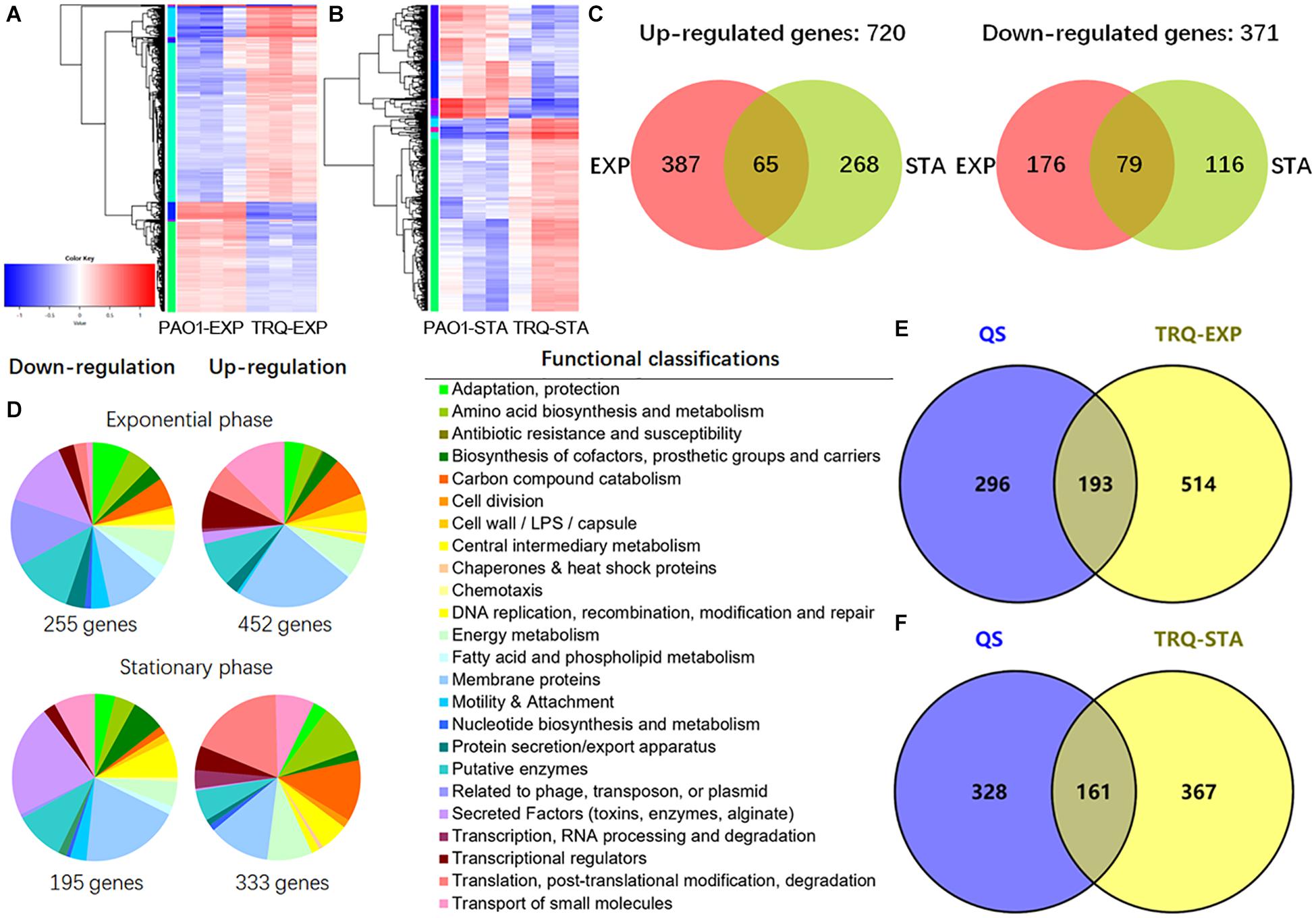
Figure 3. Transcriptome analyses of TRQ-treated P. aeruginosa cells. (A) Hierarchical clustering and heatmap analysis of the differentially expressed genes of TRQ-treated P. aeruginosa cells under exponential growth phase analysis (PAO1-EXP vs. TRQ-EXP) and stationary growth phase (PAO1-STA vs. TRQ-STA) (B). (C) Comparison of up-regulated and down-regulated genes in TRQ-treated P. aeruginosa cells in the exponential growth phase (EXP) and in stationary growth phase (STA). (D) Pie charts showing the relative abundance of functional classifications of differentially expressed genes of TRQ-treated P. aeruginosa under two growth conditions. Hypothetical genes were removed from the pies to ensure ease of reading. (E) TRQ-treated P. aeruginosa regulon shared overlap with custom QS regulon. Venn diagrams showing the overlap between QS regulon and TRQ-treated P. aeruginosa cells in the exponential growth phase and in the stationary growth phase (F).
As can be seen from Table 2, the expression of a large number of virulence genes was downregulated upon TRQ treatment, including these QS-regulated genes. Specifically, PQS signal synthase operon (pqsBCDE), hydrogen cyanide synthase operon (hcnABC), two phenazine biosynthesis operons (phzA1B1C1D1F1G1 and phzA2B2C2D2F2G2), lasA, lasB, lecB, lecA, rhlAB, chiC, alkaline protease operon (aprXDEFAI) were highly repressed upon TRQ treatment compared to non-treatment control. This finding was confirmed by phenotypic analysis (Figure 1B) and transcriptional analyses of QS regulator genes (Figure 2B) and further showed that all three QS systems in P. aeruginosa were inhibited by TRQ treatment. In addition, genes involved in type VI secretion (T6SS) were also downregulated by TRQ treatment.
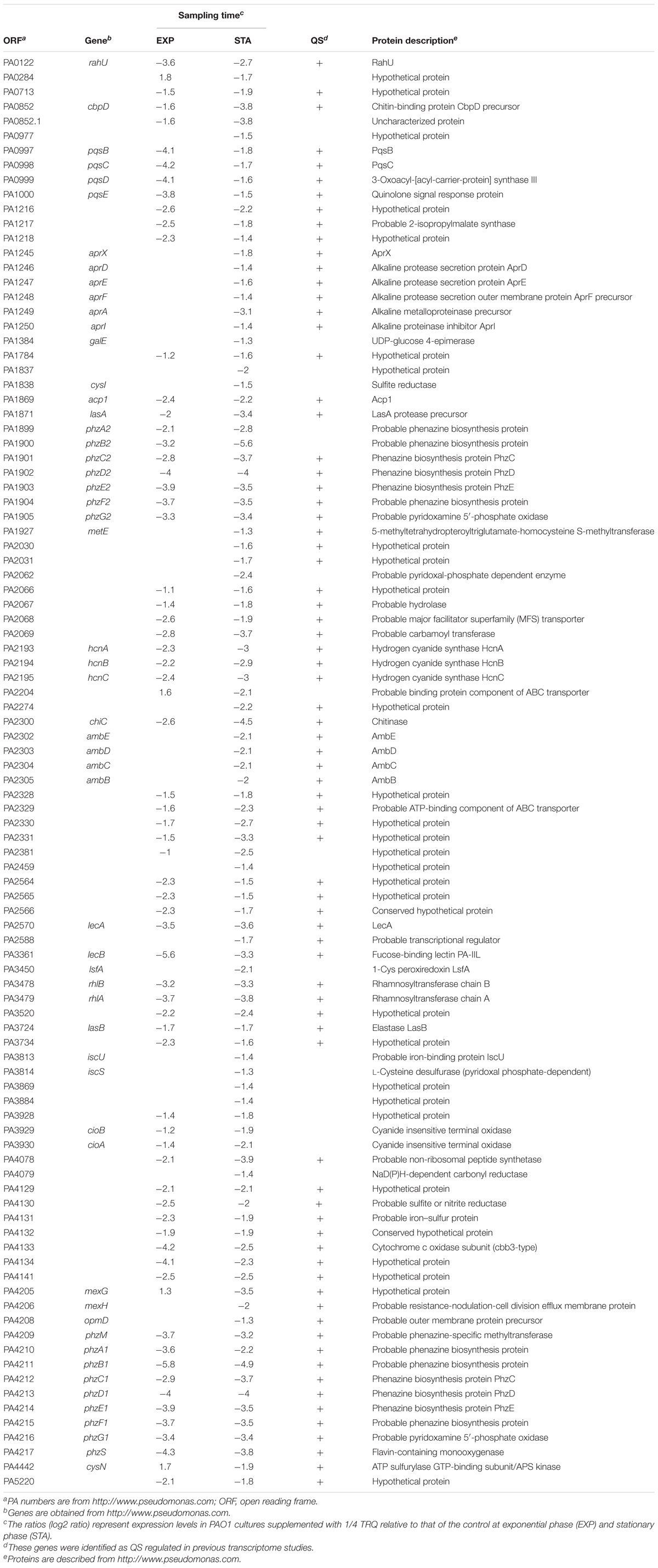
Table 2. Selected downregulated genes upon TRQ treatment.
Given the inhibition of QS systems, we aimed to find out the overlap between TRQ-treated P. aeruginosa regulon and QS regulon under typical growth conditions (LB medium in 500 mL Erlenmeyer flasks, grow at 37°C and shake at 200 rpm). We then integrated the previously identified QS regulon into a custom made QS regulon (Hentzer et al., 2003; Schuster et al., 2003) and compared it with the TRQ-treated P. aeruginosa regulon. We found that our determined regulons (TRQ-EXP and TRQ-STA) showed a close relationship with custom QS regulon (Figures 3E,F). More than 193 (TRQ-EXP) and 161 (TRQ-STA) genes were identified to overlap with the custom QS regulon (489 genes) in P. aeruginosa, suggesting that TRQ treatment has a considerable impact on the regulation of QS and clearly this dis-regulation of QS systems will contribute to the reduced virulent phenotypes.
Altogether, through our extensive RNA-seq analyses, we can conclude that the mode of action of TRQ is closely linked to the inhibition of QS systems in P. aeruginosa.
TRQ formula consists of five different components [Huang Qin (HQ), Jin Yin Hua (JYH), Lian Qiao (LQ), Xiong Dan (XD), and Shan Yang Jiao (SYJ)] (Yang et al., 2018). To further understand the roles played by these components, we used individual component to determine their effects on the expression of QS regulatory genes. As can be seen from Figures 4A–D, we showed that HQ, JYH, LQ, and XD had an inhibitory effect on the expression of all three QS regulator genes. To our surprise, SYJ demonstrated only little inhibitory effect on the expression of QS regulator genes (Figure 4E).
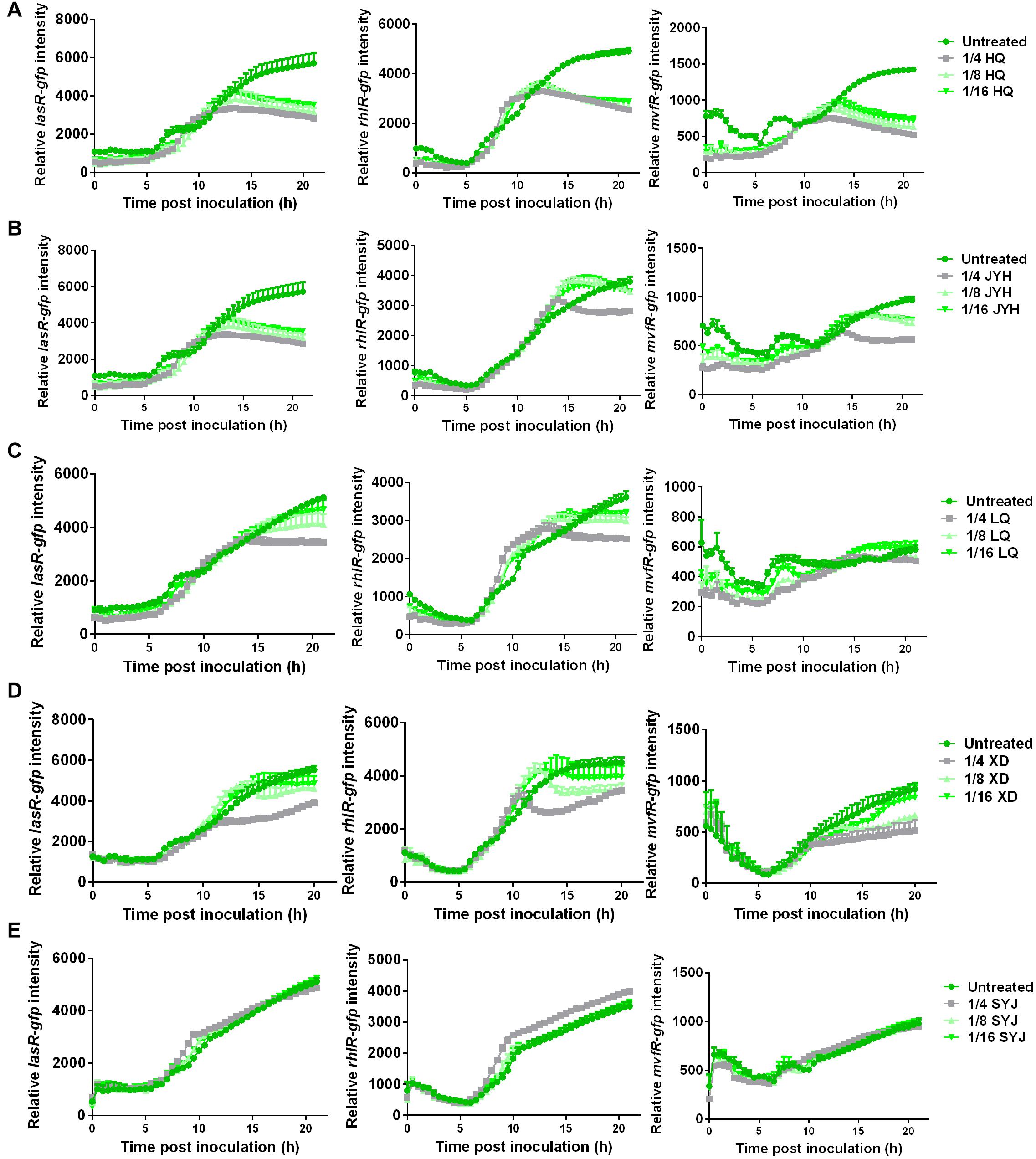
Figure 4. Component analyses of TRQ reveal sub-inhibitory effects on QS regulator genes. The same conditions were used as Figure 2. The left panel showed the expression analysis of lasR-gfp; the middle panel showed the expression of rhlR-gfp; the right panel showed the expression of mvfR-gfp. Five components in TRQ formula were tested individually for the gene expression analyses: (A) HQ; (B) JYH; (C) LQ; (D) XD; (E) SYJ.
To further understand the exact roles of these five components within TRQ formula, we calculated the individual contribution to the inhibition of QS regulatory genes. As shown in Figure 5, the inhibition of lasR expression depended on all five components with the contribution of unknown mixing effects (indicated as others in Figure 5A). The inhibition of rhlR expression depended mainly on the unknown mixing effects, and the five components showed minor effects on rhlR expression (Figure 5B). As for mvfR expression, HQ demonstrated the most significant effect and the mixing only showed a marginal effect on mvfR expression, indicating that these five components were sufficient for mvfR inhibition (Figure 5C).
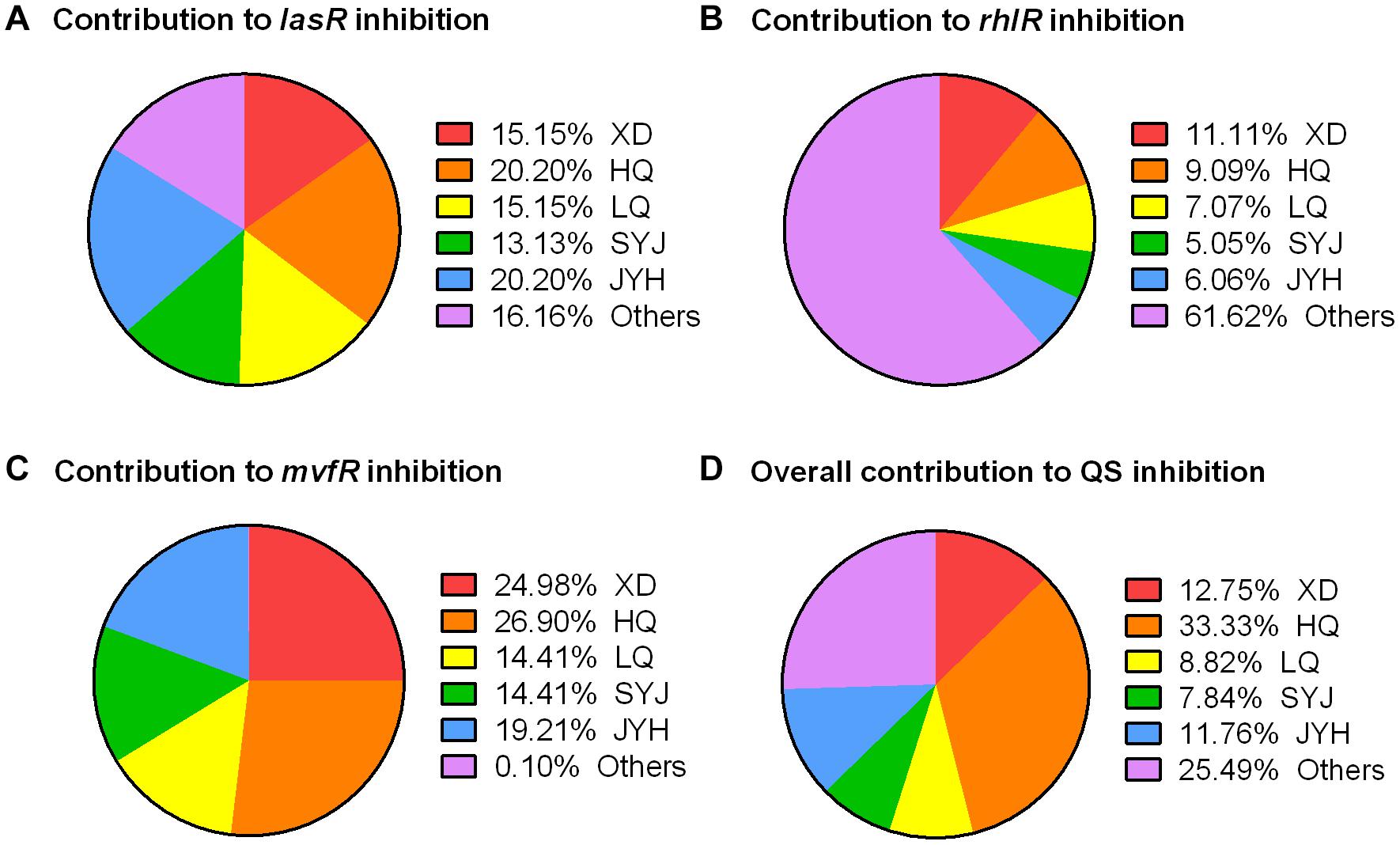
Figure 5. Contribution of five components in TRQ formula to QS inhibition. Contribution to lasR inhibition (A), rhlR inhibition (B), mvfR inhibition (C), and overall inhibition (D). Others indicate the mixing effect of five components in TRQ. Fixed data points were extracted from each treatment and calculated as the percentage calibrated by total inhibition (100%).
Altogether, we can conclude that all five components had inhibitory effects on QS systems in P. aeruginosa and, in particular HQ showed the most efficient role in QS inhibition with the assistance of the other four ingredients (Figure 5D).
Given that TRQ formula significantly attenuates the expression of QS systems in P. aeruginosa, we hypothesized whether there were upstream genetic determinants targeted by TRQ treatment. Previously, it was reported that two-component system (TCS) GacS/GacA positively regulated the expression of lasR expression in P. aeruginosa (Venturi, 2006). In addition, Vfr was also demonstrated to positively regulate the expression of global virulence factors and Vfr-dependent transcription was linked with its cofactor, cyclic AMP (cAMP), which is synthesized by two adenylate cyclases, CyaA (PA5272) and CyaB (PA3217) (Almblad et al., 2015). Based on these information, we generated transcriptional fusion reporter strains for gacS, gacA and cyaB and detected their transcriptional responses after TRQ treatment. As shown in Figures 6A–C, we could clearly observe that TRQ could reduce the expression of the TCS GacS/GacA and the cyaB adenylate cyclase gene, implying that TRQ could regulate these upstream genetic determinants of QS systems.
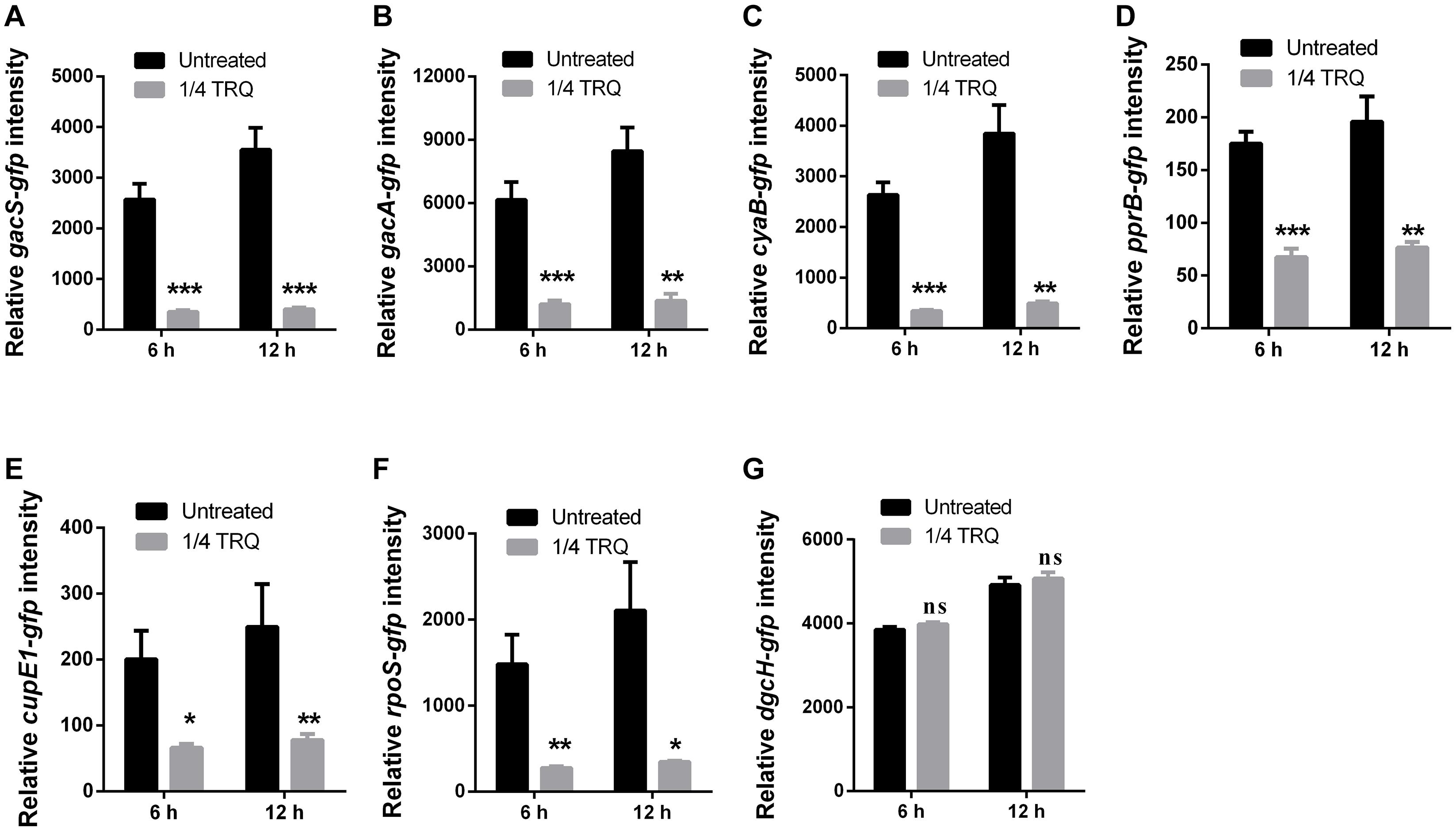
Figure 6. Upstream genetic determinants of QS systems impacted by TRQ treatment. (A) gacS-gfp; (B) gacA-gfp; (C) cyaB-gfp; (D) pprB-gfp; (E) cupE1-gfp; (F) rpoS-gfp; (G) dgcH-gfp. Relative fluorescence intensity was defined by GFP intensity calibrated with OD600nm. Statistical analysis was based on pairwise comparisons (Student’s t-test). ***P < 0.001; **P < 0.01; *P < 0.05; ns, non-significant. Error bars represent the SD.
Additionally, the two-component response regulator PprB was demonstrated to modulate the QS-regulated genes in P. aeruginosa, especially the PQS-mediated QS system (de Bentzmann et al., 2012). Furthermore, the TCS PprAB could trigger the hyper-biofilm phenotype with a unique adhesive signature made of BapA adhesin and CupE fimbriae (Giraud et al., 2011; de Bentzmann et al., 2012). More recently, the environmental signal for the TCS PprAB was found to be carbon starvation stress in P. aeruginosa and this response was dependent on the sigma factor RpoS (Wang et al., 2019). Therefore, we checked whether the TCS PprAB could be involved in TRQ-mediated QS inhibition. As can be seen from Table 3, RNA-seq analysis indicated that a large array of genes involved in carbon source metabolism were upregulated by TRQ treatment (Table 3), while the transcriptional expression of the TCS sensor kinase gene pprA was down-regulated by TRQ treatment (−1.6-fold reduction, log2 ratio, Supplementary Table S1). In addition, TRQ treatment was found to downregulate the expression of several PprAB-regulated genes including the cupE operon and tad operons (Supplementary Table S1). Therefore, it would be interesting to determine whether excess supply of carbon sources from the TRQ formula would repress the expression of pprAB. To this end, we used the transcriptional fusion of pprB, cupE1 and rpoS to verify our hypothesis. By adding TRQ to cultures of PAO1 derivatives, we could observe that the expression of these genes was significantly downregulated (Figures 6D–F). As a negative control, we used the previously characterized dgcH promoter fusion, which showed invariable under various environmental conditions and genetic backgrounds (Wei et al., 2019b). No significant change was observed in gene expression level (Figure 6G), consistent with its original RNA-seq data (see GEO dataset GSE141753).
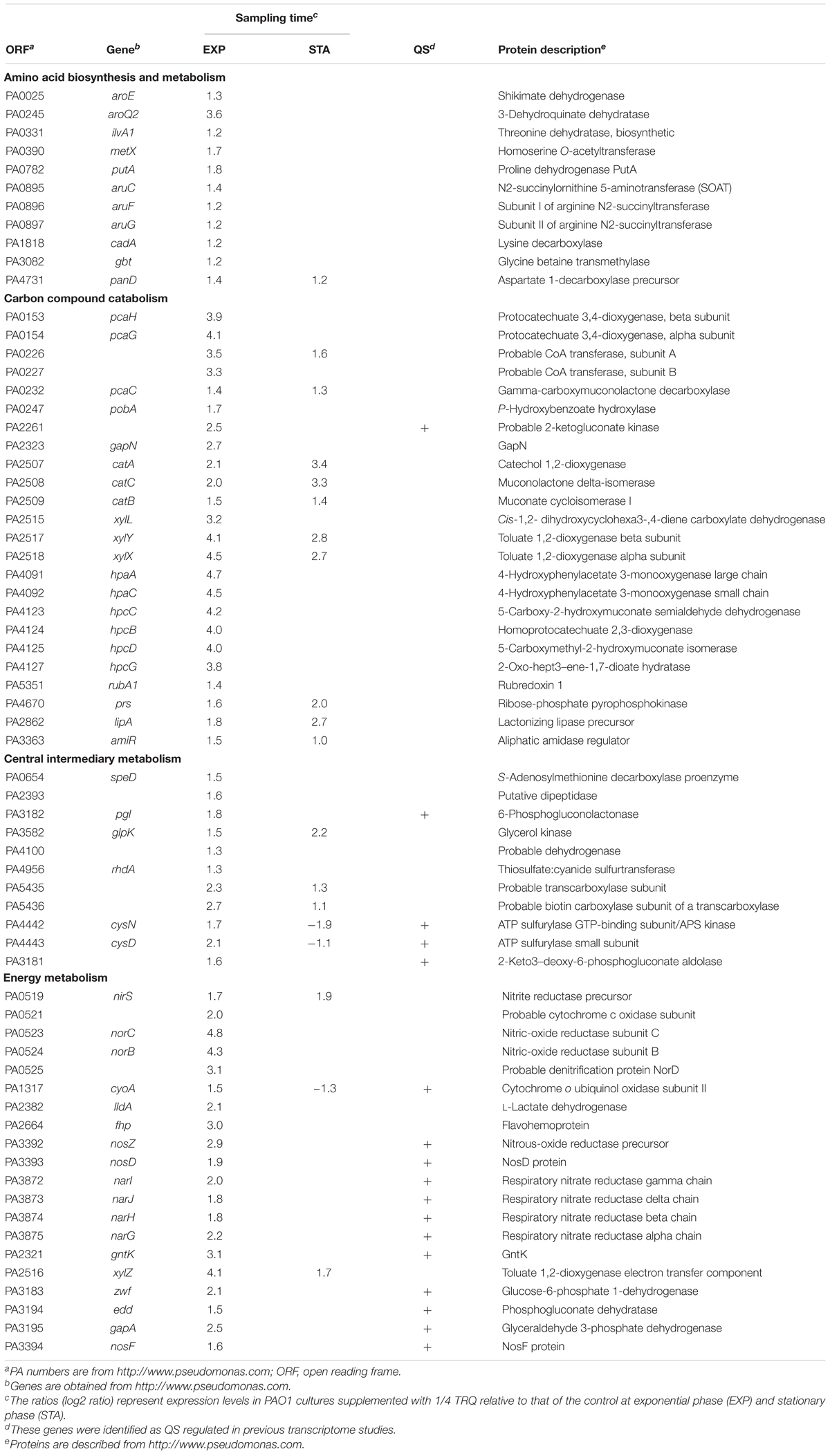
Table 3. Selected upregulated genes upon TRQ treatment.
Altogether, we could draw the conclusion that TRQ formula functions through at least two TCSs to downregulate QS systems in P. aeruginosa.
As we already observed that TRQ mainly targeted the QS regulatory systems in P. aeruginosa which mediates the pathogenesis in plant and animal models via the production of several virulence factors (Mahajan-Miklos et al., 1999; Rahme et al., 2000). We therefore checked the in vivo protection role of TRQ against P. aeruginosa infection. We fed C. elegans with TRQ-treated P. aeruginosa cells and then checked the survival rate of worms. As shown in Figure 7, we found that P. aeruginosa wild type strain was toxic to the worms since more than 50% of the population died after 4 days and 80% after 7 days. In contrast, TRQ-treated P. aeruginosa cells caused a significant increase in worm survival and this effect was TRQ concentration dependent. After 7 days, there were still more than 50% worms alive. Altogether, we demonstrated that TRQ could protect C. elegans from killing by P. aeruginosa.
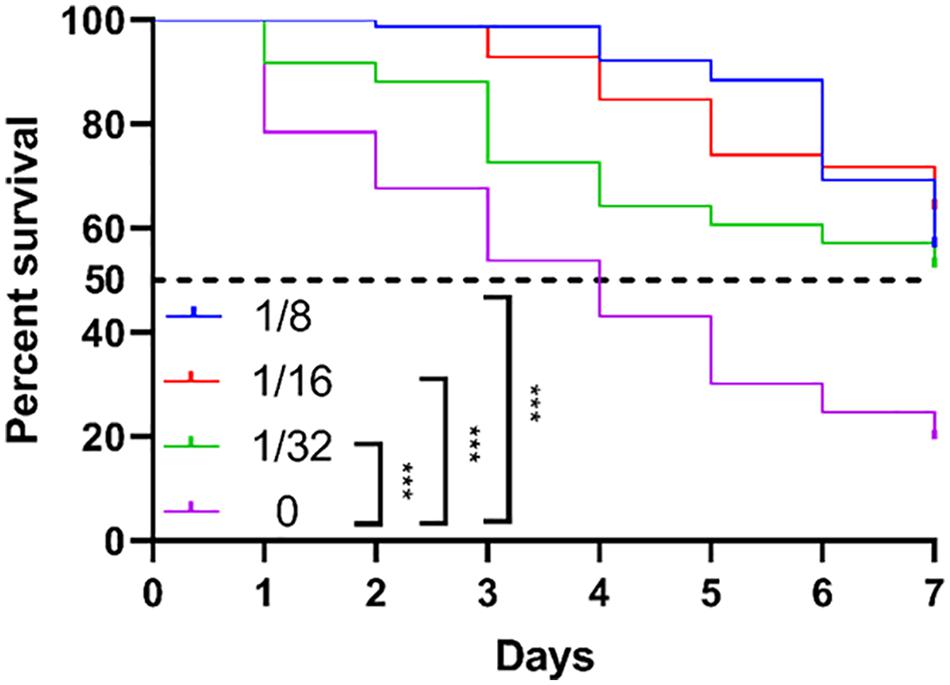
Figure 7. TRQ attenuates the virulence of P. aeruginosa toward C. elegans. The survival curve was determined by counting the percent of live worms within 7 days. The log-rank test was used to compare the significant differences between subgroups. ***P < 0.001.
Pseudomonas aeruginosa is one of the notorious human opportunistic pathogens that cause significant mortality and morbidity in immunocompromised patients. The pathogenicity of P. aeruginosa is largely dependent on the production of virulence factors such as pyocyanin, siderophores, elastases, and proteases, which are elaborately controlled by a hierarchal QS regulatory network (Papenfort and Bassler, 2016). This elegant regulatory strategy has attracted extensive attention to develop therapeutics against P. aeruginosa-related infections, particularly acute infections. A wide array of natural and synthetic compounds targeting QS systems have been developed and tested in vitro and in vivo (Starkey et al., 2014; Asfour, 2018; Dogan et al., 2019). For example, furanone C-30 (Hentzer et al., 2002), gingerol (Kim et al., 2015) and garlic extract (Rasmussen et al., 2005) have been reported to possess anti-QS activities. Recently, it has been demonstrated that ginkgolic acids from Pistacia lentiscus fruits inhibit P. aeruginosa virulence by targeting the pqs system (Tahrioui et al., 2020). Particularly, many traditional Chinese medicines have been shown to exert anti-QS and anti-virulence activities. However, the exact mechanism of QS inhibition seems to be still uncertain.
Previously, we have shown that TRQ could be used in synergy with traditional antibiotics to enhance its antibacterial effects against methicillin-resistant Staphylococcus aureus (Yang et al., 2018). Furthermore, we have demonstrated that TRQ formula could efficiently eradicate S. aureus biofilms in synergy with penicillin (Wang et al., 2011). However, the underlying mechanisms concerning the antibacterial and anti-biofilm activities of TRQ are still poorly understood. In this study, we focused on the Gram-negative bacterium P. aeruginosa as a model to investigate the mode of action of TRQ against bacterial infections. Surprisingly, we found that TRQ could target all three QS systems in P. aeruginosa, including las, rhl, and pqs (Lee et al., 2013). Through extensive bioinformatic analyses, we found that there was a significant overlap between differentially expressed genes in the exponential and stationary growth phase, suggesting that TRQ has prolonged effects on P. aeruginosa transcriptional response. Most of the QS-regulated genes have been found to be repressed by TRQ treatment, including two phenazine biosynthesis operons, lasA, lasB, lecA, lecB, rhlAB, chiC, and the alkaline protease operon (aprXDEFAI).
Interestingly, we also observed that the expression of pqsE was repressed in both exponential and stationary growth phases (Table 2). Recently, PqsE was reported to function as an alternative ligand synthase pairing with the RhlR receptor to control virulence and biofilm formation in P. aeruginosa (Mukherjee et al., 2018). The reduced pqsE gene expression would therefore disrupt the RhlR-mediated virulence gene expression. Additionally, the down-regulation of pqsBCDE will lead to quenching of the pqs system in P. aeruginosa and further contribute to reduced virulence (Deziel et al., 2005; Tahrioui et al., 2020). Collectively, all these evidences indicated that TRQ treatment could effectively target the pqs system and subsequently contribute to the inhibition of QS systems in P. aeruginosa.
The inhibition of rhl by TRQ might involve a complex mechanism since much less contribution (less than 40%) of the five components in TRQ has been observed, while unknown mixing effects play significant roles in rhlR suppression. In addition, the lack of RhlR structural information has proved the difficulty to analyze the protein-ligand interaction and to develop structure-function based therapeutics for this regulatory protein. However, several progresses have been made recently. For example, it was reported that meta-bromo-thiolactone (mBTL) could target both LasR and RhlR in vitro, while RhlR is the relevant in vivo target (O’Loughlin et al., 2013). Furthermore, RhlR responds to a suggested PqsE-encoded alternative ligand in addition to its canonical C4-HSL to promote a transcriptional program in the absence of RhlI (Mukherjee et al., 2017, 2018). Most recently, mBTL-complexed RhlR has allowed the first purification of RhlR protein (McCready et al., 2019). All these new findings will facilitate the understanding of rhl QS system and the future study of its inhibition mechanism.
The component contributions to QS inhibition analyses have shed light on the underlying mechanism of each component of TRQ and showed that all five components are necessary for the efficient and complete inhibition of QS systems, since we have noticed that none of each separate component could completely inhibit the expression of QS regulator genes. In addition, the unknown mixing effects of these five components are still not well understood. Therefore, in the future, we should focus on deciphering the individual contribution of TRQ components to determine the exact chemical mechanism within these five components, although we have already obtained the results that at least the HQ played the most important role in the inhibition of QS systems in our test conditions. It is interesting to note that multiple ingredients have been characterized within HQ and their potential involvement in TCS or QS inhibition has been reported (Supplementary Table S4). For instance, one of the HQ ingredients, scutellarein, might be involved in the inhibition of protein kinase activity (You et al., 2017). It is tempting to investigate the underlying mechanisms of action in future studies.
Our RNA-seq analyses also enabled us to further understand the upstream genetic determinants that mediate the inhibition of QS systems in P. aeruginosa. We are particularly interested in the finding that the TCSs GacS/GacA and PprA/PprB were downregulated by TRQ treatment, possibly through carbon source supply due to the existence of two nutrients within TRQ (i.e., SYJ and XD), which contain excess amounts of amino acids and other nutritional ingredients. The TCS PprA/PprB was previously shown to control some QS-regulated genes, CupE fimbriae expression and thus biofilm formation (Giraud et al., 2011), although we have not systematically examined the effect of TRQ on the formation of biofilms in P. aeruginosa in this study. Most recently, this TCS was shown to be triggered by carbon starvation stress and this process was dependent on the sigma factor RpoS (Wang et al., 2019). As for the TCS GacS/GacA, it was previously known that certain QS inhibitors could function through this pathway to repress the expression of QS-regulated genes in P. aeruginosa (Tan et al., 2014). Altogether, our results shed light on the multilevel inhibitory mechanisms of TRQ on the regulatory cascades in P. aeruginosa.
One of the technical issues concerning our study is that we used transcriptional fusion rather than qRT-PCR to validate our RNA-seq data. In principle, the transcriptional fusion analysis takes advantage of the accumulation of GFP fluorescence and serves as a reporter for the rate of transcription initiation from the promoter region. In contrast, RNA-seq, DNA microarray, and qRT-PCR report mRNA concentration, which is a balance between mRNA production and degradation (Zaslaver et al., 2006). We also realized that the transcriptional fusions are indeed plasmid-based constructs, therefore with increased copy number. Since the genes in fusion with GFP represent regulators which are not highly expressed (Supplementary Table S3), the advantage of using plasmid-borne fusions is evident. Specifically, the reason that these genes were not included in Table 2 and Supplementary Tables S1, S2 was due to their low fold change (between −2 and 2). Therefore, it appears that we did not detect them in the RNA-seq with stringent filtering. On the other hand, a factor two increase in transcripts of a regulator can have an effect more pronounced on the downstream genes, which is the case of the upstream targets by TRQ treatment. This is also the advantage of combining different methods and the reason we could identify the upstream genetic regulators that affect QS systems in our study.
In addition to inhibition of regulatory gene expression, TRQ might function through other mechanisms that would involve the possible interference with ligand-receptor interactions. For example, we have recently shown that one of the Chinese medicinal herb extracts (MHE) could interrupt the binding of MvfR to the pqsA promoter region, thus inhibiting the virulence factors production. Chemical analysis and molecular docking analysis showed that butyrolactone (BTL) and furfuryl alcohol (FA) within MHE could function as interference molecule(s) with the PQS signal to compete with MvfR (Wei et al., 2019a). Therefore, it would be interesting to identify the chemical components within TRQ to test for their performance and involvement in ligand-receptor binding interference.
Based on these facts, the complete inhibition of the three hierarchical QS systems in P. aeruginosa raises the possibility that TRQ is likely functioning as a multitarget inhibitor of bacterial virulence. Since we have seen that the expression of all three regulator genes (lasR, rhlR and mvfR) were completely repressed by TRQ treatment. One of the QS inhibition mechanisms might be the shutdown of las by TRQ and in turn the downregulation of pqs and rhl systems. The second hypothesis is the inhibition of the pqs system by TRQ with the consecutive shutdown of the rhl system. In addition to QS inhibition, we have unraveled that two TCSs, GacS/GacA and PprA/PprB, could be inhibited by TRQ treatment, further explaining the reason of QS inhibition and the final attenuation of P. aeruginosa virulence. Altogether, we summarized the conclusions from our study in Figure 8.
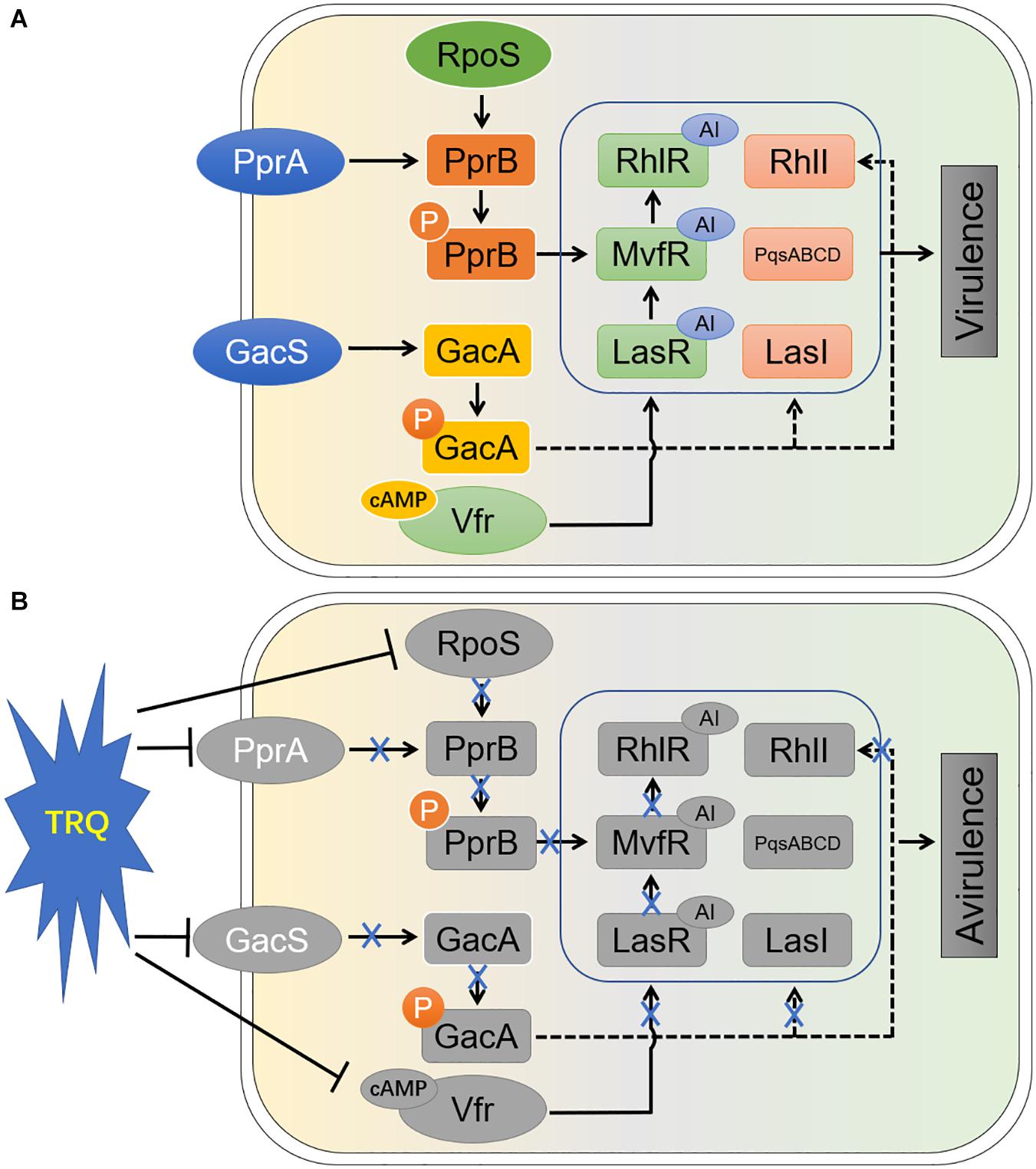
Figure 8. Schematic diagram of the mode of action of TRQ-mediated virulence inhibition in P. aeruginosa. Without introduction, the QS systems and their upstream regulators work smoothly (A). In our study (B), we have found that TRQ could efficiently inhibit the transcription of upstream regulators of QS systems in P. aeruginosa. Particularly, we found that two TCSs, GacS/GacA and PprA/PprB, were involved in these process. One of the sigma factors, RpoS, was also identified to positively affect the expression of PprA/PprB system (Wang et al., 2019). The expression of c-AMP synthesis was also inhibited by TRQ treatment, suggesting that the downstream Vfr-mediated QS activation via lasR was shutdown. In addition, TCS GacS/GacA was found to be involved in the regulation of QS signal synthesis by LasI and RhlI (Reimmann et al., 1997). With the introduction of TRQ, all those processes might be affected and thus the virulent phenotypes diminished (avirulence). AI, autoinducer; P, phosphorylated.
The datasets generated for this study can be found in the accession number GSE141753.
YW conceived the project and supervised the study. QW, WY, QT, and KC performed the experiments and analyzed the data. GH, LL, and LM performed experiments, provided the reagents, technical assistance and interpretation of data for the project. QW, WY, PC, and YW wrote the draft. All authors added comments and corrections and approved the final version to be published.
This work was supported by Kaibao Pharmaceutical Company, Shanghai, and Basic Scientific Research Fund from Ministry of Finance of China (No. ZZ2017001), Office of International Cooperation, China Academy of Chinese Medical Sciences (GH2018001), Academician Expert Workstation, National Key Research and Development Program of China (No. 2019YFC1709305), and National Mega-project for Innovative Drugs (2019ZX09721001-005-002). QW is a CSC scholarship receiver and thanks for invitation from Prof. Per Saris in University of Helsinki, Finland.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We would like to thank the two reviewers for their constructive suggestions and comments.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2020.517462/full#supplementary-material
Supplementary Table 1 | TRQ-treated P. aeruginosa regulon in exponential growth phase.
Supplementary Table 2 | TRQ-treated P. aeruginosa regulon in stationary growth phase.
Supplementary Table 3 | Raw expression data of selected genes from TRQ-treated P. aeruginosa regulon.
Supplementary Table 4 | Main components of three herbal medicines of TRQ (Ru et al., 2014).
Almblad, H., Harrison, J. J., Rybtke, M., Groizeleau, J., Givskov, M., Parsek, M. R., et al. (2015). The cyclic AMP-Vfr signaling pathway in Pseudomonas aeruginosa is inhibited by cyclic di-GMP. J. Bacteriol. 197, 2190–2200. doi: 10.1128/jb.00193-15
Asfour, H. Z. (2018). Anti-quorum sensing natural compounds. J. Microsc. Ultrastruct. 6, 1–10. doi: 10.4103/jmau.jmau_10_18
Aybey, A., and Demirkan, E. (2016). Inhibition of quorum sensing-controlled virulence factors in Pseudomonas aeruginosa by human serum paraoxonase. J. Med. Microbiol. 65, 105–113. doi: 10.1099/jmm.0.000206
Bolger, A. M., Lohse, M., and Usadel, B. (2014). Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114–2120. doi: 10.1093/bioinformatics/btu170
Borlee, B. R., Geske, G. D., Blackwell, H. E., and Handelsman, J. (2010). Identification of synthetic inducers and inhibitors of the quorum-sensing regulator LasR in Pseudomonas aeruginosa by high-throughput screening. Appl. Environ. Microbiol. 76, 8255–8258. doi: 10.1128/aem.00499-10
Camilli, A., and Bassler, B. L. (2006). Bacterial small-molecule signaling pathways. Science 311, 1113–1116. doi: 10.1126/science.1121357
Chong, Y. M., How, K. Y., Yin, W. F., and Chan, K. G. (2018). The effects of Chinese herbal medicines on the quorum sensing-regulated virulence in Pseudomonas aeruginosa PAO1. Molecules 23:972. doi: 10.3390/molecules23040972
Cornelis, P. (2019). Putting an end to the Pseudomonas aeruginosa IQS controversy. Microbiologyopen 9:e962.
Davies, D. G., Parsek, M. R., Pearson, J. P., Iglewski, B. H., Costerton, J. W., and Greenberg, E. P. (1998). The involvement of cell-to-cell signals in the development of a bacterial biofilm. Science 280, 295–298. doi: 10.1126/science.280.5361.295
D’Costa, V. M., King, C. E., Kalan, L., Morar, M., Sung, W. W., Schwarz, C., et al. (2011). Antibiotic resistance is ancient. Nature 477, 457–461.
de Bentzmann, S., Giraud, C., Bernard, C. S., Calderon, V., Ewald, F., Plesiat, P., et al. (2012). Unique biofilm signature, drug susceptibility and decreased virulence in Drosophila through the Pseudomonas aeruginosa two-component system PprAB. PLoS Pathog. 8:e1003052. doi: 10.1371/journal.ppat.1003052
Deziel, E., Gopalan, S., Tampakaki, A. P., Lepine, F., Padfield, K. E., Saucier, M., et al. (2005). The contribution of MvfR to Pseudomonas aeruginosa pathogenesis and quorum sensing circuitry regulation: multiple quorum sensing-regulated genes are modulated without affecting lasRI, rhlRI or the production of N-acyl-L-homoserine lactones. Mol. Microbiol. 55, 998–1014. doi: 10.1111/j.1365-2958.2004.04448.x
Diggle, S. P., Cornelis, P., Williams, P., and Camara, M. (2006). 4-quinolone signalling in Pseudomonas aeruginosa: old molecules, new perspectives. Int. J. Med. Microbiol. 296, 83–91. doi: 10.1016/j.ijmm.2006.01.038
Diggle, S. P., Matthijs, S., Wright, V. J., Fletcher, M. P., Chhabra, S. R., Lamont, I. L., et al. (2007). The Pseudomonas aeruginosa 4-quinolone signal molecules HHQ and PQS play multifunctional roles in quorum sensing and iron entrapment. Chem. Biol. 14, 87–96. doi: 10.1016/j.chembiol.2006.11.014
Dogan, S., Gokalsin, B., Senkardes, I., Dogan, A., and Sesal, N. C. (2019). Anti-quorum sensing and anti-biofilm activities of Hypericum perforatum extracts against Pseudomonas aeruginosa. J. Ethnopharmacol. 235, 293–300. doi: 10.1016/j.jep.2019.02.020
Fan, H., Dong, Y., Wu, D., Bowler, M. W., Zhang, L., and Song, H. (2013). QsIA disrupts LasR dimerization in antiactivation of bacterial quorum sensing. Proc. Natl. Acad. Sci. U.S.A. 110, 20765–20770. doi: 10.1073/pnas.1314415110
Fito-Boncompte, L., Chapalain, A., Bouffartigues, E., Chaker, H., Lesouhaitier, O., Gicquel, G., et al. (2011). Full virulence of Pseudomonas aeruginosa requires OprF. Infect. Immun. 79, 1176–1186.
Giraud, C., Bernard, C. S., Calderon, V., Yang, L., Filloux, A., Molin, S., et al. (2011). The PprA-PprB two-component system activates CupE, the first non-archetypal Pseudomonas aeruginosa chaperone-usher pathway system assembling fimbriae. Environ. Microbiol. 13, 666–683. doi: 10.1111/j.1462-2920.2010.02372.x
Haussler, S. (2004). Biofilm formation by the small colony variant phenotype of Pseudomonas aeruginosa. Environ. Microbiol. 6, 546–551. doi: 10.1111/j.1462-2920.2004.00618.x
Heeb, S., Fletcher, M. P., Chhabra, S. R., Diggle, S. P., Williams, P., and Camara, M. (2011). Quinolones: from antibiotics to autoinducers. FEMS Microbiol. Rev. 35, 247–274. doi: 10.1111/j.1574-6976.2010.00247.x
Hentzer, M., Riedel, K., Rasmussen, T. B., Heydorn, A., Andersen, J. B., Parsek, M. R., et al. (2002). Inhibition of quorum sensing in Pseudomonas aeruginosa biofilm bacteria by a halogenated furanone compound. Microbiology 148, 87–102. doi: 10.1099/00221287-148-1-87
Hentzer, M., Wu, H., Andersen, J. B., Riedel, K., Rasmussen, T. B., Bagge, N., et al. (2003). Attenuation of Pseudomonas aeruginosa virulence by quorum sensing inhibitors. EMBO J. 22, 3803–3815. doi: 10.1093/emboj/cdg366
Jimenez, P. N., Koch, G., Thompson, J. A., Xavier, K. B., Cool, R. H., and Quax, W. J. (2012). The multiple signaling systems regulating virulence in Pseudomonas aeruginosa. Microbiol. Mol. Biol. Rev. 76, 46–65. doi: 10.1128/mmbr.05007-11
Kim, H. S., Lee, S. H., Byun, Y., and Park, H. D. (2015). 6-Gingerol reduces Pseudomonas aeruginosa biofilm formation and virulence via quorum sensing inhibition. Sci. Rep. 5:8656.
Klopfenstein, D. V., Zhang, L., Pedersen, B. S., Ramirez, F., Warwick Vesztrocy, A., Naldi, A., et al. (2018). GOATOOLS: a python library for gene ontology analyses. Sci. Rep. 8:10872.
Koh, C. L., Sam, C. K., Yin, W. F., Tan, L. Y., Krishnan, T., Chong, Y. M., et al. (2013). Plant-derived natural products as sources of anti-quorum sensing compounds. Sensors 13, 6217–6228. doi: 10.3390/s130506217
Lee, J., Wu, J., Deng, Y., Wang, J., Wang, C., Chang, C., et al. (2013). A cell-cell communication signal integrates quorum sensing and stress response. Nat. Chem. Biol. 9, 339–343. doi: 10.1038/nchembio.1225
Li, S., Chen, S., Fan, J., Cao, Z., Ouyang, W., Tong, N., et al. (2018). Anti-biofilm effect of novel thiazole acid analogs against Pseudomonas aeruginosa through IQS pathways. Eur. J. Med. Chem. 145, 64–73. doi: 10.1016/j.ejmech.2017.12.076
Liu, W., Jiang, H. L., Cai, L. L., Yan, M., Dong, S. J., and Mao, B. (2016). Tanreqing injection attenuates lipopolysaccharide-induced airway inflammation through MAPK/NF-kappaB signaling pathways in rats model. Evid. Based Complement. Alternat. Med. 2016:5292346.
Lyczak, J. B., Cannon, C. L., and Pier, G. B. (2000). Establishment of Pseudomonas aeruginosa infection: lessons from a versatile opportunist. Microbes Infect. 2, 1051–1060. doi: 10.1016/s1286-4579(00)01259-4
Lyczak, J. B., Cannon, C. L., and Pier, G. B. (2002). Lung infections associated with cystic fibrosis. Clin. Microbiol. Rev. 15, 194–222.
Mahajan-Miklos, S., Tan, M. W., Rahme, L. G., and Ausubel, F. M. (1999). Molecular mechanisms of bacterial virulence elucidated using a Pseudomonas aeruginosa-Caenorhabditis elegans pathogenesis model. Cell 96, 47–56. doi: 10.1016/s0092-8674(00)80958-7
McClure, R., Balasubramanian, D., Sun, Y., Bobrovskyy, M., Sumby, P., Genco, C. A., et al. (2013). Computational analysis of bacterial RNA-Seq data. Nucleic Acids Res. 41:e140.
McCready, A. R., Paczkowski, J. E., Cong, J. P., and Bassler, B. L. (2019). An autoinducer-independent RhlR quorum-sensing receptor enables analysis of RhlR regulation. PLoS Pathog. 15:e1007820. doi: 10.1371/journal.ppat.1007820
Miller, M. B., and Bassler, B. L. (2001). Quorum sensing in bacteria. Annu. Rev. Microbiol. 55, 165–199.
Miller, W. G., Leveau, J. H., and Lindow, S. E. (2000). Improved gfp and inaZ broad-host-range promoter-probe vectors. Mol. Plant Microbe Interact. 13, 1243–1250. doi: 10.1094/mpmi.2000.13.11.1243
Moy, T. I., Ball, A. R., Anklesaria, Z., Casadei, G., Lewis, K., and Ausubel, F. M. (2006). Identification of novel antimicrobials using a live-animal infection model. Proc. Natl. Acad. Sci. U.S.A. 103, 10414–10419. doi: 10.1073/pnas.0604055103
Muh, U., Schuster, M., Heim, R., Singh, A., Olson, E. R., and Greenberg, E. P. (2006). Novel Pseudomonas aeruginosa quorum-sensing inhibitors identified in an ultra-high-throughput screen. Antimicrob. Agents Chemother. 50, 3674–3679. doi: 10.1128/aac.00665-06
Mukherjee, S., Moustafa, D., Smith, C. D., Goldberg, J. B., and Bassler, B. L. (2017). The RhlR quorum-sensing receptor controls Pseudomonas aeruginosa pathogenesis and biofilm development independently of its canonical homoserine lactone autoinducer. PLoS Pathog. 13:e1006504. doi: 10.1371/journal.ppat.1006504
Mukherjee, S., Moustafa, D. A., Stergioula, V., Smith, C. D., Goldberg, J. B., and Bassler, B. L. (2018). The PqsE and RhlR proteins are an autoinducer synthase-receptor pair that control virulence and biofilm development in Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. U.S.A. 115, E9411–E9418.
Nalca, Y., Jansch, L., Bredenbruch, F., Geffers, R., Buer, J., and Haussler, S. (2006). Quorum-sensing antagonistic activities of azithromycin in Pseudomonas aeruginosa PAO1: a global approach. Antimicrob. Agents Chemother. 50, 1680–1688. doi: 10.1128/aac.50.5.1680-1688.2006
O’Loughlin, C. T., Miller, L. C., Siryaporn, A., Drescher, K., Semmelhack, M. F., and Bassler, B. L. (2013). A quorum-sensing inhibitor blocks Pseudomonas aeruginosa virulence and biofilm formation. Proc. Natl. Acad. Sci. U.S.A. 110, 17981–17986. doi: 10.1073/pnas.1316981110
Papenfort, K., and Bassler, B. L. (2016). Quorum sensing signal-response systems in Gram-negative bacteria. Nat. Rev. Microbiol. 14, 576–588. doi: 10.1038/nrmicro.2016.89
Parsek, M. R., and Greenberg, E. P. (2000). Acyl-homoserine lactone quorum sensing in gram-negative bacteria: a signaling mechanism involved in associations with higher organisms. Proc. Natl. Acad. Sci. U.S.A. 97, 8789–8793. doi: 10.1073/pnas.97.16.8789
Perez-Martinez, I., and Haas, D. (2011). Azithromycin inhibits expression of the GacA-dependent small RNAs RsmY and RsmZ in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 55, 3399–3405. doi: 10.1128/aac.01801-10
Rahme, L. G., Ausubel, F. M., Cao, H., Drenkard, E., Goumnerov, B. C., Lau, G. W., et al. (2000). Plants and animals share functionally common bacterial virulence factors. Proc. Natl. Acad. Sci. U.S.A. 97, 8815–8821. doi: 10.1073/pnas.97.16.8815
Rasmussen, T. B., Bjarnsholt, T., Skindersoe, M. E., Hentzer, M., Kristoffersen, P., Kote, M., et al. (2005). Screening for quorum-sensing inhibitors (QSI) by use of a novel genetic system, the QSI selector. J. Bacteriol. 187, 1799–1814. doi: 10.1128/jb.187.5.1799-1814.2005
Rasmussen, T. B., and Givskov, M. (2006). Quorum-sensing inhibitors as anti-pathogenic drugs. Int. J. Med. Microbiol. 296, 149–161. doi: 10.1016/j.ijmm.2006.02.005
Reimmann, C., Beyeler, M., Latifi, A., Winteler, H., Foglino, M., Lazdunski, A., et al. (1997). The global activator GacA of Pseudomonas aeruginosa PAO positively controls the production of the autoinducer N-butyryl-homoserine lactone and the formation of the virulence factors pyocyanin, cyanide, and lipase. Mol. Microbiol. 24, 309–319. doi: 10.1046/j.1365-2958.1997.3291701.x
Robinson, M. D., Mccarthy, D. J., and Smyth, G. K. (2010). edgeR: a Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 26, 139–140. doi: 10.1093/bioinformatics/btp616
Rojas Murcia, N., Lee, X., Waridel, P., Maspoli, A., Imker, H. J., Chai, T., et al. (2015). The Pseudomonas aeruginosa antimetabolite L-2-amino-4-methoxy-trans-3-butenoic acid (AMB) is made from glutamate and two alanine residues via a thiotemplate-linked tripeptide precursor. Front. Microbiol. 6:170. doi: 10.3389/fmicb.2015.00170
Ru, J., Li, P., Wang, J., Zhou, W., Li, B., Huang, C., et al. (2014). TCMSP: a database of systems pharmacology for drug discovery from herbal medicines. J. Cheminform. 6:13. doi: 10.1186/1758-2946-6-13
Schuster, M., and Greenberg, E. P. (2006). A network of networks: quorum-sensing gene regulation in Pseudomonas aeruginosa. Int. J. Med. Microbiol. 296, 73–81. doi: 10.1016/j.ijmm.2006.01.036
Schuster, M., Lostroh, C. P., Ogi, T., and Greenberg, E. P. (2003). Identification, timing, and signal specificity of Pseudomonas aeruginosa quorum-controlled genes: a transcriptome analysis. J. Bacteriol. 185, 2066–2079. doi: 10.1128/jb.185.7.2066-2079.2003
Skindersoe, M. E., Alhede, M., Phipps, R., Yang, L., Jensen, P. O., Rasmussen, T. B., et al. (2008). Effects of antibiotics on quorum sensing in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 52, 3648–3663.
Starkey, M., Lepine, F., Maura, D., Bandyopadhaya, A., Lesic, B., He, J., et al. (2014). Identification of anti-virulence compounds that disrupt quorum-sensing regulated acute and persistent pathogenicity. PLoS Pathog. 10:e1004321. doi: 10.1371/journal.ppat.1004321
Tahrioui, A., Ortiz, S., Azuama, O. C., Bouffartigues, E., Benalia, N., Tortuel, D., et al. (2020). Membrane-interactive compounds from Pistacia lentiscus L. thwart Pseudomonas aeruginosa virulence. Front. Microbiol. 11:1068. doi: 10.3389/fmicb.2020.01068
Tan, S. Y., Chua, S. L., Chen, Y., Rice, S. A., Kjelleberg, S., Nielsen, T. E., et al. (2013). Identification of five structurally unrelated quorum-sensing inhibitors of Pseudomonas aeruginosa from a natural-derivative database. Antimicrob. Agents Chemother. 57, 5629–5641. doi: 10.1128/aac.00955-13
Tan, S. Y., Liu, Y., Chua, S. L., Vejborg, R. M., Jakobsen, T. H., Chew, S. C., et al. (2014). Comparative systems biology analysis to study the mode of action of the isothiocyanate compound Iberin on Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 58, 6648–6659. doi: 10.1128/aac.02620-13
Tjaden, B. (2015). De novo assembly of bacterial transcriptomes from RNA-seq data. Genome Biol. 16:1.
Vinckx, T., Wei, Q., Matthijs, S., and Cornelis, P. (2010). The Pseudomonas aeruginosa oxidative stress regulator OxyR influences production of pyocyanin and rhamnolipids: protective role of pyocyanin. Microbiology 156, 678–686. doi: 10.1099/mic.0.031971-0
Wang, C., Chen, W., Xia, A., Zhang, R., Huang, Y., Yang, S., et al. (2019). Carbon starvation induces the expression of PprB-regulated genes in Pseudomonas aeruginosa. Appl. Environ. Microbiol. 85:e01705-19.
Wang, Y., Wang, T., Hu, J., Ren, C., Lei, H., Hou, Y., et al. (2011). Anti-biofilm activity of TanReQing, a traditional Chinese medicine used for the treatment of acute pneumonia. J. Ethnopharmacol. 134, 165–170. doi: 10.1016/j.jep.2010.11.066
Wei, Q., Bhasme, P., Wang, Z., Wang, L., Wang, S., Zeng, Y., et al. (2019a). Chinese medicinal herb extract inhibits PQS-mediated quorum sensing system in Pseudomonas aeruginosa. J. Ethnopharmacol. 248:112272. doi: 10.1016/j.jep.2019.112272
Wei, Q., Leclercq, S., Bhasme, P., Xu, A., Zhu, B., Zhang, Y., et al. (2019b). Diguanylate cyclases and phosphodiesterases required for basal-Level c-di-GMP in Pseudomonas aeruginosa as revealed by systematic phylogenetic and transcriptomic analyses. Appl. Environ. Microbiol. 85:e01194-19.
Wei, Q., Tarighi, S., Dotsch, A., Haussler, S., Musken, M., Wright, V. J., et al. (2011). Phenotypic and genome-wide analysis of an antibiotic-resistant small colony variant (SCV) of Pseudomonas aeruginosa. PLoS One 6:e29276. doi: 10.1371/journal.pone.0029276
Xiao, G., Deziel, E., He, J., Lepine, F., Lesic, B., Castonguay, M. H., et al. (2006). MvfR, a key Pseudomonas aeruginosa pathogenicity LTTR-class regulatory protein, has dual ligands. Mol. Microbiol. 62, 1689–1699. doi: 10.1111/j.1365-2958.2006.05462.x
Xie, C., Mao, X., Huang, J., Ding, Y., Wu, J., Dong, S., et al. (2011). KOBAS 2.0: a web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 39, W316–W322.
Yang, W., Liu, J., Blazekovic, B., Sun, Y., Ma, S., Ren, C., et al. (2018). In vitro antibacterial effects of Tanreqing injection combined with vancomycin or linezolid against methicillin-resistant Staphylococcus aureus. BMC Complement. Altern. Med. 18:169. doi: 10.1186/s12906-018-2231-8
Ye, L., Cornelis, P., Guillemyn, K., Ballet, S., and Hammerich, O. (2014). Structure revision of N-mercapto-4-formylcarbostyril produced by Pseudomonas fluorescens G308 to 2-(2-hydroxyphenyl)thiazole-4-carbaldehyde [aeruginaldehyde]. Nat. Prod. Commun. 9, 789–794.
You, L., Zhu, H., Wang, C., Wang, F., Li, Y., Li, Y., et al. (2017). Scutellarin inhibits Hela cell growth and glycolysis by inhibiting the activity of pyruvate kinase M2. Bioorg. Med. Chem. Lett. 27, 5404–5408. doi: 10.1016/j.bmcl.2017.11.011
Keywords: Pseudomonas aeruginosa, antimicrobial agent, quorum sensing, two-component system, Tanreqing
Citation: Yang W, Wei Q, Tong Q, Cui K, He G, Lin L, Ma LZ, Cornelis P and Wang Y (2020) Traditional Chinese Medicine Tanreqing Inhibits Quorum Sensing Systems in Pseudomonas aeruginosa. Front. Microbiol. 11:517462. doi: 10.3389/fmicb.2020.517462
Received: 04 December 2019; Accepted: 09 November 2020;
Published: 17 December 2020.
Edited by:
Ren-You Gan, Chinese Academy of Agricultural Sciences, ChinaReviewed by:
Gloria Soberón-Chávez, National Autonomous University of Mexico, MexicoCopyright © 2020 Yang, Wei, Tong, Cui, He, Lin, Ma, Cornelis and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yi Wang, cHJvZi53YW5neWlAZm94bWFpbC5jb20=
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.