
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 26 August 2020
Sec. Infectious Agents and Disease
Volume 11 - 2020 | https://doi.org/10.3389/fmicb.2020.02009
 Tatiana C. A. Pinto1
Tatiana C. A. Pinto1 Natália S. Costa1
Natália S. Costa1 Sandrine E. C. M. Pina1
Sandrine E. C. M. Pina1 Aline R. V. Souza1
Aline R. V. Souza1 Laura M. A. Oliveira1
Laura M. A. Oliveira1 Camille A. B. Moura1
Camille A. B. Moura1 Fabíola C. O. Kegele1,2
Fabíola C. O. Kegele1,2 Vânia L. C. Merquior3
Vânia L. C. Merquior3 Ana Caroline N. Botelho1
Ana Caroline N. Botelho1 José M. Peralta1
José M. Peralta1 Lúcia M. Teixeira1*
Lúcia M. Teixeira1*Streptococcus pneumoniae remains a major agent of invasive diseases, especially in children and the elderly. The presence of pneumococcal capsule, pneumococcal surface protein A (PspA), and pilus type 1 (PI-1) and the ability of colony phase variation are assumed to play important roles in the virulence potential of this microorganism. Differences in the capsular polysaccharide allow the characterization of more than 90 pneumococcal serotypes; among them, serotype 14 and serogroup 9 stand out due to their prevalence in the pre- pneumococcal conjugate vaccine era and frequent association with penicillin non-susceptibility. Here we investigated the distribution of PI-1 and pspA genes and colony phase variants among 315 S. pneumoniae isolates belonging to serotype 14 and serogroup 9, recovered over 20 years in Brazil, and correlated these characteristics with penicillin susceptibility and genotype as determined by multilocus sequence typing. All strains were shown to carry pspA genes, with those of family 2 (pspA2) being the most common, and nearly half of the strains harbored P1-1 genes. The pspA gene family and the presence of PI-1 genes were conserved features among strains belonging to a given clone. A trend for increasing the occurrence of pspA2 and PI-1 genes over the period of investigation was observed, and it coincided with the dissemination of CC156 (Spain9V-3) clone in Brazil, suggesting a role for these virulence attributes in the establishment and the persistence of this successful clone. Opaque variant was the colony phenotype most frequently observed, regardless of clonal type. On the other hand, the transparent variant was more commonly associated with penicillin-non-susceptible pneumococci and with strains presenting evidence of recombination events involving the genes coding for polysaccharide capsule and PspA, suggesting that pneumococcal transparent variants may present a higher ability to acquire exogenous DNA. The results bring to light new information about the virulence potentials of serotype 14 and serogroup 9 S. pneumoniae isolates representing the major clones that have been associated with the emergence and the dissemination of antimicrobial resistance in our setting since the late 1980s.
Streptococcus pneumoniae is a leading cause of invasive infections and can also asymptomatically colonize the upper respiratory tract of variable numbers of individuals (Weiser et al., 2018). The polysaccharide capsule is considered as the major virulence factor of this species, and its antigenic diversity allows the characterization of pneumococcal isolates into more than 90 serotypes (Geno et al., 2015). Additionally, several other virulence factors contribute to the pathogenic potential of this microorganism, including the pneumococcal surface protein A (PspA), which reduces opsonization and clearance of bacteria by the host immune system (Kadioglu et al., 2008), and the pilus type 1 (PI-1), which is believed to mediate the attachment of pneumococci to human epithelial cells and the extracellular matrix (Nelson et al., 2007; Hilleringmann et al., 2008; Iovino et al., 2020). PspA is known to occur in virtually all pneumococcal strains and exhibits polymorphic regions, being classified in three families and six clades (Hollingshead et al., 2000; Croney et al., 2012). The occurrence of PI-1, in turn, seems to be restricted to certain pneumococcal clones (Sjöström et al., 2007; Aguiar et al., 2008; Imai et al., 2011; Selva et al., 2012; Metcalf et al., 2016), but its distribution among pneumococcal isolates circulating in different geographic areas, including Brazil, has not been extensively investigated yet. The virulence of S. pneumoniae is also associated with phase variation, a phenomenon characterized by the expression of two colony phenotypes, opaque and transparent. These variants may differ from each other in multiple virulence-associated characteristics (Li et al., 2016; Li and Zhang, 2019). In addition, PI-1 was shown to have an on/off regulation, which has been associated with phase variation (Basset et al., 2011; Danne et al., 2014).
The efforts led by the Pneumococcal Molecular Epidemiology Network (PMEN) have helped in tracking successful pneumococcal clones disseminated worldwide, which are usually associated with antimicrobial resistance1. In Brazil, we have shown that four major clonal complexes (CC) were responsible for the emergence and the dissemination of antimicrobial resistance among S. pneumoniae strains of serotypes 14 and 9 circulating in the country since the late 1980s (Pinto et al., 2016). Among them, three are related to PMEN clones, including CC156 (related to Spain9V-3), CC66 (related to Tennessee14-18), and CC15 (related to England14-9), while the fourth, namely CC5401, is characterized as a regional clone. Despite the importance of such CCs in the dissemination of antimicrobial resistance, little is known about other characteristics that could potentially influence their pathogenic potential. Therefore, in the present study, we have evaluated the distribution of three virulence-associated characteristics (presence of pspA and PI-1 coding genes, as well as colony phase variation phenotypes), among S. pneumoniae isolates belonging to major clones of serotype 14 or serogroup 9 circulating in Brazil, which were recognized to be responsible for the emergence and the expansion of penicillin non-susceptibility in our setting.
A total of 315 S. pneumoniae isolates were investigated, encompassing 216 of serotype 14 and 99 of serogroup 9. All the isolates were previously characterized regarding capsular type, antimicrobial susceptibility profile, and multilocus sequence typing (Pinto et al., 2016). A number of them (89 isolates) were also previously characterized by multiple locus variable-number tandem repeat (VNTR) analysis (MLVA; Costa et al., 2016).
In addition, reference strains belonging to five worldwide disseminated pneumococcal lineages (Spain9v-3 ST156, England14-9 ST9, Tennessee14-18 ST67, Netherlands14-35 ST124, and Netherlands15B-37 ST199), characterized by the PMEN1 were included. Such lineages were previously shown to be associated with certain isolates included in the present study (Pinto et al., 2016).
The determination of pspA gene type was carried out by PCR as previously described (Pimenta et al., 2006), with a modification in the MgCl2 concentration (used at 3 mM in all PCR mixtures in the present study). Bacterial DNAs were obtained by using the Chelex® 100 resin (Bio-Rad Laboratories, United States) as previously described (Pinto et al., 2013).
pspA typing (pspA1, pspA2, and pspA3) was performed for all 315 isolates included in the study by using specific primers for family 1 (LSM12 and SKH63, Swiatlo et al., 1997; Vela-Coral et al., 2001), family 2 (LSM12 and SKH52, Swiatlo et al., 1997; Vela-Coral et al., 2001), and family 3 (SKH41 and SKH42; Hollingshead et al., 2006). The isolates that were not initially amplified using an annealing temperature of 62°C were retested under the same cycling conditions but using annealing temperatures of 58°C and, subsequently, 55°C. The pspA clade (1–6) was determined for a subset of 34 strains selected to represent the most important clones previously detected (Pinto et al., 2016) and required an additional sequencing step after the PCR amplification of the clade-defining region (CDR) using primers LSM12 and SKH2 (Swiatlo et al., 1997; Hollingshead et al., 2000). The amplification products were purified using ExoSAP-IT (Affymetrix-USB, United States) according to the manufacturer’s instructions and were run on an ABI 3130 Genetic Analyzer (Applied Biosystems, United States). The sequences were edited and aligned with BioEdit v7.0.9.0 (Hall, 1999).
The pspA gene families and clades were also determined for the reference strains of the five PMEN clones included in the study. S. pneumoniae reference strains were included as controls for pspA clade 1 (strain BG9739), clade 2 (strain EF10197), clade 3 (strain AC122), clade 4 (strain BG7561), clade 5 (ATCC 6303), and clade 6 (strain BG6380).
The recombination events among pspA genes of 15 S. pneumoniae strains, selected to represent different sequence types (ST) within the clonal complex CC156, were identified in silico using the recombination detection program (RDP) package, which incorporates the RDP, GENECONV, Maxchi, Chimera, 3Seq, Bootscan, and SiSscan programs (Martin et al., 2015), to predict the recombination signals from aligned DNA sequences. The recombination events were scored as significant only if at least three out of seven individual programs in the package identified the events with p < 0.05.
The presence of PI-1 coding genes was evaluated among all 315 pneumococcal strains included in the study and also among the reference strains of five PMEN clones. Bacterial DNAs were obtained by using the Chelex® 100 resin (Pinto et al., 2013), and the presence of genes associated with PI-1 production was evaluated using the primer set described by Moschioni et al. (2008). Controls for the presence (pneumococcal reference strain TIGR4) and absence (pneumococcal reference strain R6) of PI-1 genes were included in each set of reactions.
A microscopic examination of pneumococcal colony phase variation was carried out for the 235 strains as previously described (Weiser et al., 1994). Briefly, bacteria were grown on 5% sheep blood agar plates (Plast Labor, Brazil) for 18–24 h at 37°C under 5%-CO2 atmosphere. A single colony of each strain was inoculated onto a plate containing Todd-Hewitt agar (BBL, United States), supplemented with 0.5% yeast extract (Difco, United States), and incubated for 18–24 h at 37°C in 5% CO2. Subsequently, the colonies on each plate were submitted to microscopic examination and were assigned to either opaque or transparent colony morphology by using a stereoscopic microscope equipped with a × 40 magnifying glass (TecNival, Brazil).
The evaluation of the distribution of colony phase variants among penicillin-susceptible and penicillin-non-susceptible strains and also among ST156 and ST162 isolates was carried out by applying the one-way analysis of variance (ANOVA) test. The distribution of colony variants, pspA types, and PI-1 coding genes over the years was investigated by linear regression analysis. GraphPad Prism software v5.0 was used to perform all the statistical analysis and p < 0.05 were considered as significant.
The characteristics of the 315 S. pneumoniae isolates analyzed in the present study are included in Supplementary Table S1.
All 315 isolates harbored pspA genes, demonstrating the wide occurrence of these genes among pneumococcal isolates as previously suggested (McDaniel et al., 1998; Croney et al., 2012; Blumental et al., 2015; Kawaguchiya et al., 2018; Knupp-Pereira et al., 2019). A total of 30.8% (97 isolates) harbored pspA1 genes, while 69.2% (218 isolates) had pspA2 genes. The pspA3 genes were not detected. These observations reinforce the concept that about 98% of the pneumococcal strains circulating worldwide are estimated to belong to families 1 or 2, while the occurrence of family 3 is rare (Ochs et al., 2008; Blumental et al., 2015; Kawaguchiya et al., 2018; Knupp-Pereira et al., 2019). The isolates possessing the pspA1 gene that were submitted to CDR sequencing (10 in total) were included in clade 1, while the pspA2 isolates analyzed (24 in total) belonged to clade 3. Types of pspA genes were highly conserved among strains within each clone (Table 1). The association of CC156 with pspA2 and clade 3 observed in this study has already been detected among isolates from different countries (Melin et al., 2008; Rolo et al., 2009; Sadowy et al., 2010), as well as the association of CC15 with pspA1 and clade 1 (Ito et al., 2007; Melin et al., 2008; Rolo et al., 2009), showing that the PMEN-related clones identified in Brazil share characteristics with those circulating internationally.
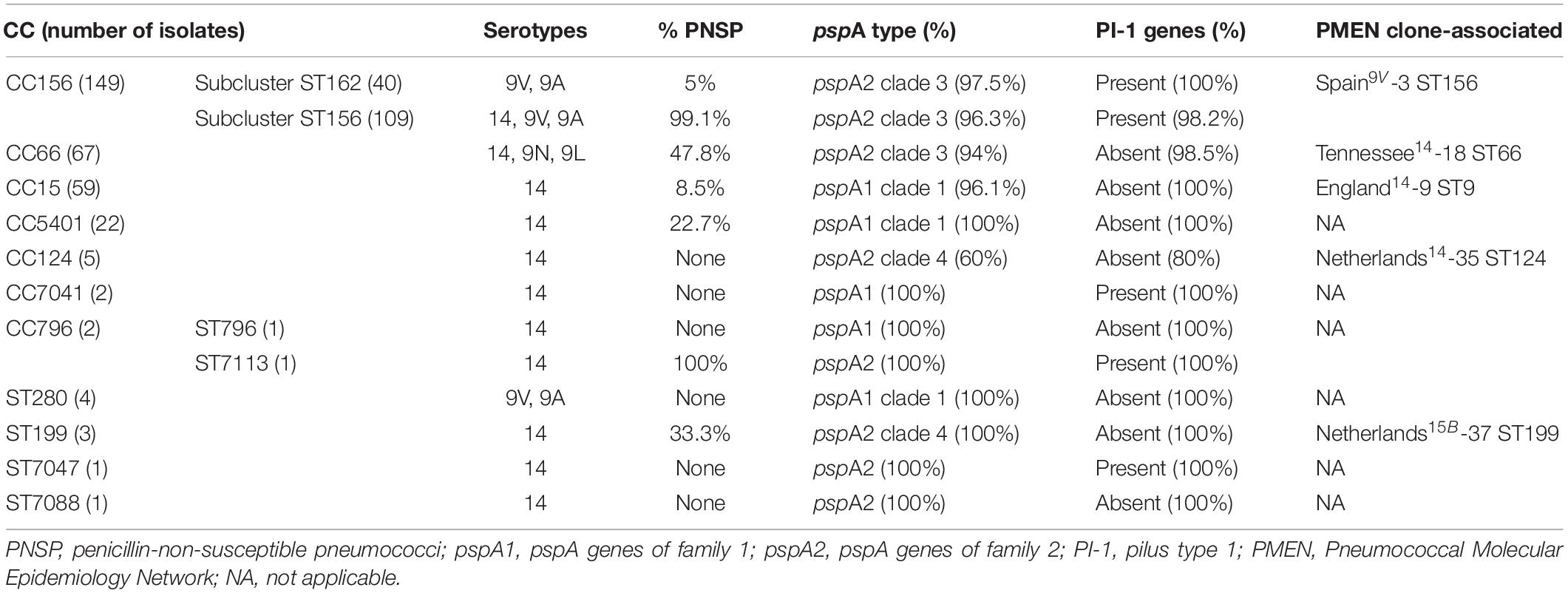
Table 1. Characteristics of the clonal complexes (CC) and singleton sequence types (ST) of the 315 Streptococcus pneumoniae isolates included in the present study.
Around 50% of the isolates (total of 154) had PI-1 genes, and the presence of these determinants varied according to the clone (Table 1). CC156, CC7041, ST7113, and ST7047 were the only genotypes associated with PI-1. Accordingly, previous studies have revealed that PI-1 is harbored by certain pneumococcal clones, especially among the internationally disseminated ST156 (Sjöström et al., 2007; Aguiar et al., 2008; Imai et al., 2011; Selva et al., 2012; Horácio et al., 2016). In turn, CC7041, ST7113, and ST7047 are genotypes described only in Brazil up to date (accessed on April 4th 2020)2, and, thus, their association with PI-1 is being described here for the first time.
Among the five PMEN clones investigated in the present study, all had pspA genes and two harbored PI-1 coding genes, including Spain9V-3 ST156 and Netherlands15B-37 ST199 (Table 2). In general, the reference strains of PMEN clones presented profiles of pspA type and PI-1 genes identical to those of clinical isolates associated with these clones (Table 1). The only exception regarding the presence of PI-1 genes within Netherlands15B-37 ST199 isolates is that, while the respective reference strain harbored PI-1 genes (Table 2), three clinical isolates belonging to ST199 evaluated in the present study did not (Table 1). This finding is in contrast with a previous report indicating the association of PI-1 with clinical isolates belonging to CC199 from Italy (Del Grosso et al., 2013). However, ST199 isolates usually belong to serogroups 15 or 192, and the ones included in the present study were of serotype 14, representing the first serotype 14 variants of this clone to be reported (Pinto et al., 2016). We have also previously shown that these serotype 14 ST199 variants might have been generated by capsular switching events since they were genetically more closely related to strains belonging to serotype 15B than to other isolates of serotype 14 (Costa et al., 2016). Previous studies have indicated that capsular switching events in pneumococci can lead to other simultaneous recombination events spanning a genome region of approximately 750 kb, which includes the capsular locus and the pilus type 1 operon (Sjöström et al., 2007; Croucher et al., 2014; Metcalf et al., 2016); thus, in these serotype 14 ST199 variants, PI-1 coding genes could have been lost during recombination events in the capsular locus that led to serotype switching.
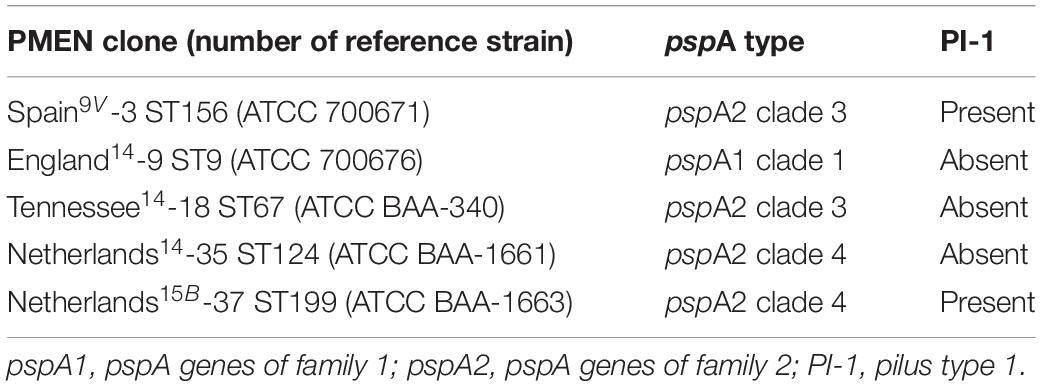
Table 2. Profiles of pspA gene types and pilus type 1 coding genes of five reference strains representative of Pneumococcal Molecular Epidemiology Network (PMEN) Streptococcus pneumoniae clones included in the present study.
Not all 315 pneumococcal isolates were tested for colony phenotype. Although this represents a limitation of the study, it is expected that the results obtained from the subset of 235 (almost 75% of the total) isolates are highly representative of the total sampling as they were selected to encompass the different variants included in the study. The majority of the 235 pneumococcal isolates submitted to the determination of colony phenotype (128 isolates, 54.5%) consisted of exclusively opaque variants, while 19.6% (46 isolates) were exclusively transparent and 25.9% (61 isolates) presented mixed phenotypes comprising both opaque and transparent variants. The opaque phenotype was the most common overall, corroborating previous observations (Arai et al., 2011). The distribution of opaque and transparent variants did not correlate with pspA types or presence of PI-1 coding genes. The four major clonal complexes included in the study (CC156, CC66, CC15, and CC5401) likewise presented very similar distributions of both opaque and transparent variants, with a slightly higher frequency of transparent variants within CC5401 (Table 3).

Table 3. Distribution of colony phase variants among the four major clonal complexes (CCs) of serotypes 9 and 14 Streptococcus pneumoniae isolates included in this study.
On the other hand, the distribution of opaque and transparent variants between penicillin-susceptible (PSP) and penicillin-non-susceptible (PNSP) isolates was markedly different, with a significantly higher frequency of transparent variants and a lower occurrence of opaque variants within PNSP isolates (Table 4; p = 0.0126). Similarly, PNSP occurrence among isolates showing the transparent phenotype (56.5%) was much more frequent than among those presenting the opaque phenotype (37.5%). PNSP in this study comprised all isolates that were shown to present penicillin minimal inhibitory concentrations (MIC) ≥0.12 μg/ml (Pinto et al., 2016).

Table 4. Distribution of colony phase variants among serotypes 9 and 14 Streptococcus pneumoniae isolates according to the penicillin susceptibility profile.
We have shown earlier that the emergence of PNSP and the increment on penicillin MICs among pneumococcal isolates circulating in our setting were mainly due to the introduction of ST156 in the mid-1990s (Pinto et al., 2016). Additionally, it has been suggested that ST156 was derived from a penicillin-susceptible ancestor, the genotype ST162, which is a single-locus variant (SLV) of ST156 (Sjöström et al., 2007). Therefore, although ST156 and ST162, as well as their SLVs, are components of CC156, they can be analyzed as two different subclusters, especially with regard to penicillin susceptibility (Table 1). Interestingly, a significant difference (p = 0.0151) in the distribution of colony phenotypes was also seen between the two subclusters included in CC156 (Table 3). The isolates belonging to subcluster ST156, associated with penicillin non-susceptibility, presented higher and lower frequencies of transparent and opaque variants, respectively, when compared to subcluster ST162. Such observations may indicate a correlation between the transparent phenotype and penicillin non-susceptibility among the pneumococcal isolates investigated, but further studies are required to better elucidate this question.
The opaque phenotype was the most common overall and also within each one of all the clones investigated in this study, except for ST199 isolates, which we have shown to be a probable result of capsular switching events (Costa et al., 2016). Among these three serotype 14 ST199 variants, two were exclusively composed of transparent variants, while the third was composed of a mixed phenotype, indicating that the genotype in which capsular recombination events were assumed to occur was also the only genotype where the transparent phenotype was more frequent.
The association of the transparent phenotype with higher rates of recombination in pspA genes was also detected in this study. Among the 15 pneumococcal isolates submitted to in silico evaluation of recombination events in pspA genes, five had significant signals of recombination; all of them belonged to subcluster ST156 and three were shown to be exclusively composed of transparent variants, while the other two comprised mixed phenotypes. On the other hand, none of the isolates presenting exclusively opaque variants had significant signals of recombination in pspA genes (Table 5).
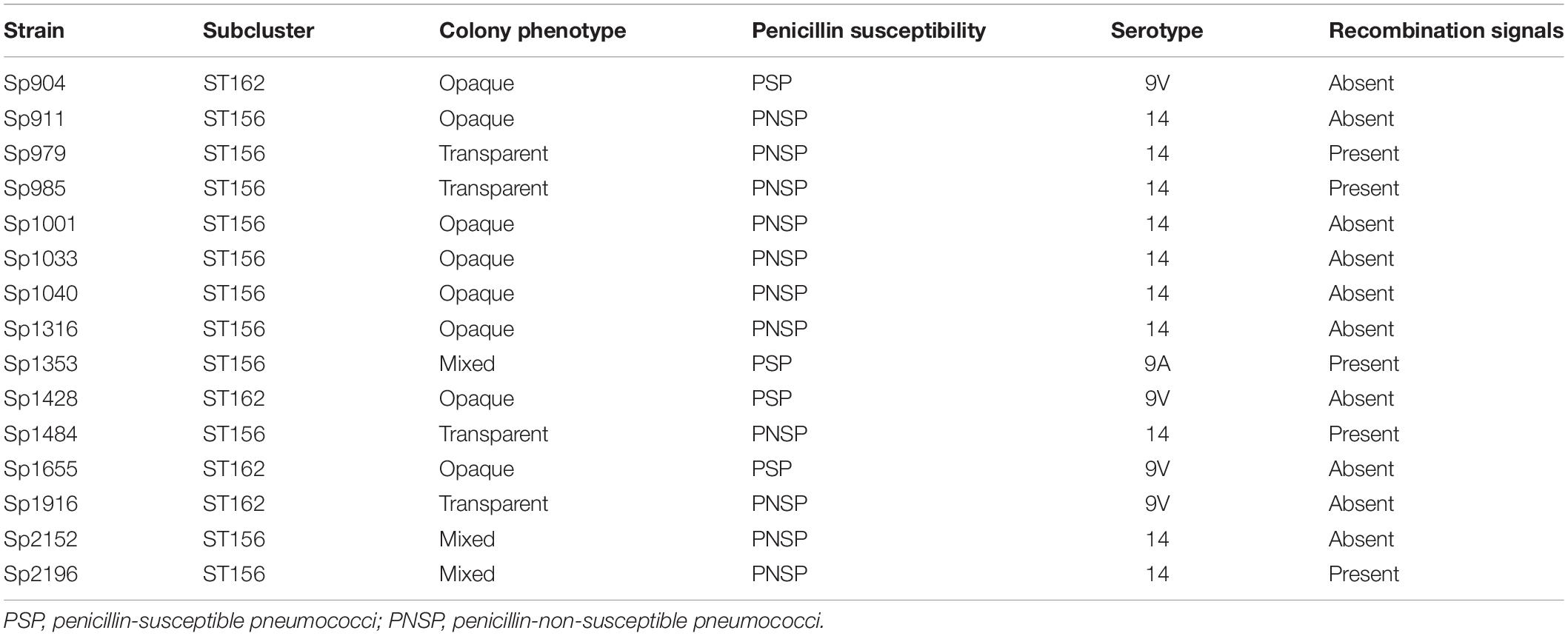
Table 5. Characteristics of the 15 Streptococcus pneumoniae isolates belonging to different subclusters of clonal complex 156 (CC156) submitted to in silico analysis of recombination events in pspA genes by using the recombination detection program software.
Although phase variation in pneumococci has been investigated over the past decades, the molecular mechanisms involved in this phenomenon are still unclear (Weiser et al., 1994; Manso et al., 2014; Li et al., 2016; Li and Zhang, 2019). Earlier studies have shown that it appears to be independent of in vitro growth conditions including pH, temperature, and osmolarity (Kim et al., 1999). In addition, genetic analyses have demonstrated that BOX elements (repeat sequences in the pneumococcal genome) can have a role in the frequency of phase variation (Saluja and Weiser, 1995; Li et al., 2016). Many of the 315 pneumococcal isolates included in this study were previously analyzed by MLVA (Costa et al., 2016), a methodology that targets BOX elements. This previous study showed interesting differences in the number of repeat sequences in certain BOX elements between ST156 and ST162 isolates. In the BOX element represented by locus Spneu40, for example, two or three repeats were found in ST162 isolates, while a number of eight repeats were observed in ST156 isolates (Costa et al., 2016).
In the present investigation, an increasing trend for the occurrence of isolates harboring pspA2 and PI-1 genes was observed over the years, in parallel with a decreasing tendency of isolates showing pspA1 genes and the opaque phenotype (Figure 1). These trends might actually reflect the natural fluctuation of clonal complexes over time as it was previously shown that ST156, which is associated with PI-1 and pspA2 genes and a higher frequency of transparent variants, has been increasing steadily in occurrence since the mid-1990s (Pinto et al., 2016). However, changes in the occurrence of PI-1 and PspA families have been observed in the United States after the implementation of the seven-valent pneumococcal conjugate vaccine (Regev-Yochay et al., 2010; Croney et al., 2012). Since this study only included isolates recovered before the introduction of the first pneumococcal conjugate vaccine (10-valent PCV) offered to the general population by the Brazilian public health service, the continuous tracking of these characteristics among isolates obtained in more recent years is required and may reveal the impact of PCV10 in the distribution of virulence-associated characteristics among pneumococci.
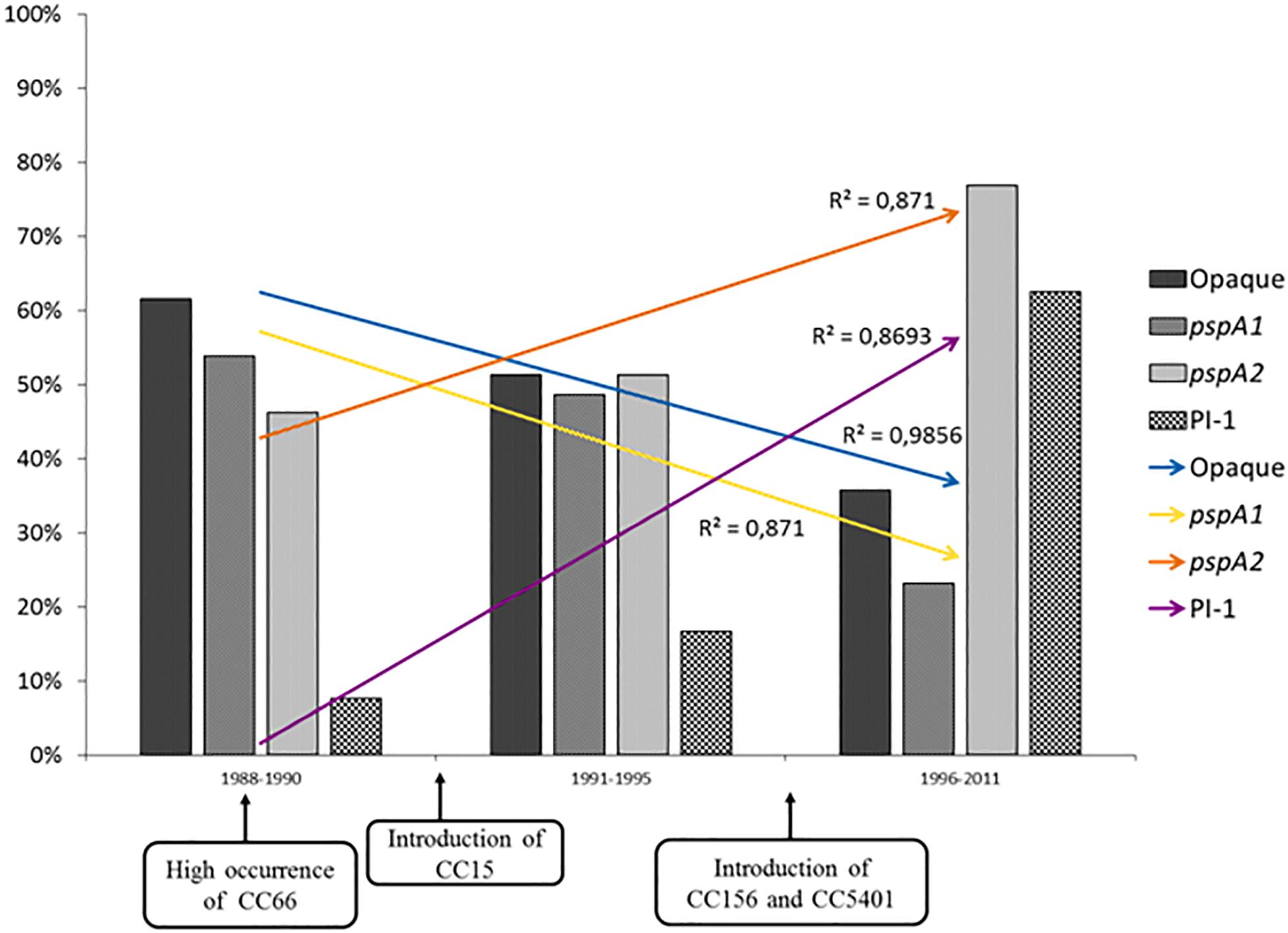
Figure 1. Distribution of opaque variants, pspA gene types, and pilus type 1 (PI-1) coding genes among serotypes 9 and 14 Streptococcus pneumoniae isolates investigated in this study over the years. Opaque, isolates with opaque phenotype; pspA1, pspA genes of family 1; pspA2, pspA genes of family 2.
This study highlights data on the virulence potential of serotype 14 and serogroup 9 S. pneumoniae isolates representing the major CCs that have been responsible for the emergence and the dissemination of antimicrobial resistance in our setting since the late 1980s. Particularly, the results indicate that important recombination events involving the genes coding for penicillin-binding proteins (leading to penicillin non-susceptibility), capsule polysaccharide (leading to serotype switching), and PspA are more associated with the transparent variants of pneumococci, which may present an improved ability to acquire exogenous DNA. The evaluation of such virulence-associated characteristics can help to better understand the evolution and the adaptation of pneumococcal clones over the years and, consequently, may be helpful in designing new strategies to prevent pneumococcal diseases.
All datasets generated for this study are included in the article/Supplementary Material.
TP, VM, JP, and LT designed the study. CM and FK performed the microscopic analysis of colony variants. NC, AS, and AB performed the experiments for pilus gene detection. SP, LO, and AB performed the experiments for pspA typing. TP performed in silico and statistical analysis. TP, VM, JP, and LT wrote the manuscript. All authors contributed to the article and approved the submitted version.
This work was supported in part by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES) – Finance Code 001, the Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), the Instituto Nacional de Pesquisa em Resistência Antimicrobiana (INPRA), and the Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We wish to thank Dr. Maria Cristina Brandileone of the Instituto Adolfo Lutz, São Paulo, Brazil, for the kind donation of pneumococcal reference strains for the PspA clades.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2020.02009/full#supplementary-material
TABLE S1 | Characteristics of the 315 Streptococcus pneumoniae isolates included in the present study. aND, not determined, bPenicillin minimum inhibitory concentration, cSequence type.
Aguiar, S. I., Serrano, I., Pinto, F. R., Melo-Cristino, J., and Ramirez, M. (2008). The presence of the pilus locus is a clonal property among pneumococcal invasive isolates. BMC Microbiol. 8:41. doi: 10.1186/1471-2180-8-41
Arai, J., Hotomi, M., Hollingshead, S. K., Ueno, Y., Briles, D. E., and Yamanaka, N. (2011). Streptococcus pneumoniae isolates from middle ear fluid and nasopharynx of children with acute otitis media exhibit phase variation. J. Clin. Microbiol. 49, 1646–1649. doi: 10.1128/jcm.01990-10
Basset, A., Turner, K. H., Boush, E., Sayeed, S., Dove, S. L., and Malley, R. (2011). Expression of the type 1 pneumococcal pilus is bistable and negatively regulated by the structural component RrgA. Infect. Immun. 79, 2974–2983. doi: 10.1128/iai.05117-11
Blumental, S., Granger-Farbos, A., Moïsi, J. C., Soullié, B., Leroy, P., Njanpop-Lafourcade, B. M., et al. (2015). Virulence factors of Streptococcus pneumoniae. Comparison between African and French invasive isolates and implication for future vaccines. PLoS One 10:e0133885. doi: 10.1371/journal.pone.0133885
Costa, N. S., Pinto, T. C., Merquior, V. L., Castro, L. F., da Rocha, F. S., Morais, J. M., et al. (2016). MLVA Typing of Streptococcus pneumoniae isolates with emphasis on serotypes 14, 9N and 9V: comparison of previously described panels and proposal of a novel 7 VNTR loci-based simplified scheme. PLoS One 11:e0158651. doi: 10.1371/journal.pone.00158651
Croney, C. M., Coats, M. T., Nahm, M. H., Briles, D. E., and Crain, M. J. (2012). PspA family distribution, unlike capsular serotype, remains unaltered following introduction of the heptavalent pneumococcal conjugate vaccine. Clin. Vaccine Immunol. 19, 891–896. doi: 10.1128/cvi.05671-11
Croucher, N. J., Chewapreecha, C., Hanage, W. P., Harris, S. R., McGee, L., van der Linden, M., et al. (2014). Evidence for soft selective sweeps in the evolution of pneumococcal multidrug resistance and vaccine escape. Genome Biol. Evol. 6, 1589–1602. doi: 10.1093/gbe/evu120
Danne, C., Dubrac, S., Trieu-Cuot, P., and Dramsi, S. (2014). Single cell stochastic regulation of pilus phase variation by an attenuation-like mechanism. PLoS Pathog. 10:e1003860. doi: 10.1371/journal.pone.1003860
Del Grosso, M., Camilli, R., D’Ambrosio, F., Petrucci, G., Melchiorre, S., Moschioni, M., et al. (2013). Increase of pneumococcal serotype 19A in Italy is due to expansion of the piliated clone ST416/CC199. J. Med. Microbiol. 62, 1220–1225. doi: 10.1099/jmm.0.061242-0
Geno, K. A., Gilbert, G. L., Song, J. Y., Skovsted, I. C., Klugman, K. P., Jones, C., et al. (2015). Pneumococcal capsules and their types: past, present, and future. Clin. Microbiol. Rev. 28, 871–899. doi: 10.1128/cmr.00024-15
Hall, T. (1999). BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nuclear Acids Symp. Series 41, 95–98.
Hilleringmann, M., Giusti, F., Baudner, B. C., Masignani, V., Covacci, A., Rappuoli, R., et al. (2008). Pneumococcal pili are composed of protofilaments exposing adhesive clusters of RrgA. PLoS Pathog. 4:e1000026. doi: 10.1371/journal.pone.1000026
Hollingshead, S. K., Baril, L., Ferro, S., King, J., Coan, P., Briles, D. E., et al. (2006). Pneumococcal surface protein A (PspA) family distribution among clinical isolates from adults over 50 years of age collected in seven countries. J. Med. Microbiol. 55, 215–221. doi: 10.1099/jmm.0.46268-0
Hollingshead, S. K., Becker, R., and Briles, D. E. (2000). Diversity of PspA: mosaic genes and evidence for past recombination in Streptococcus pneumoniae. Infect. Immun. 68, 5889–5900. doi: 10.1128/iai.68.10.5889-5900.2000
Horácio, A. N., Silva-Costa, C., Diamantino-Miranda, J., Lopes, J. P., Ramirez, M., Melo-Cristino, J., et al. (2016). Population structure of Streptococcus pneumoniae causing invasive disease in adults in Portugal before PCV13 availability for adults: 2008-2011. PLoS One 11:e0153602. doi: 10.1371/journal.pone.0153602
Imai, S., Ito, Y., Ishida, T., Hirai, T., Ito, I., Yoshimura, K., et al. (2011). Distribution and clonal relationship of cell surface virulence genes among Streptococcus pneumoniae isolates in Japan. Clin. Microbiol. Infect. 17, 1409–1414. doi: 10.1111/j.1469-0691.2010.03446.x
Iovino, F., Nannapaneni, P., Henriques-Normark, B., and Normark, S. (2020). The impact of the ancillary pilus-1 protein RrgA of Streptococcus pneumoniae on colonization and disease. Mol. Microbiol. 113, 650–658. doi: 10.1111/mmi.14451
Ito, Y., Osawa, M., Isozumi, R., Imai, S., Ito, I., Hirai, T., et al. (2007). Pneumococcal surface protein A family types of Streptococcus pneumoniae from community-acquired pneumonia patients in Japan. Eur. J. Clin. Microbiol. Infect. Dis. 26, 739–742. doi: 10.1007/s10096-007-0364-7
Kadioglu, A., Weiser, J. N., Paton, J. C., and Andrew, P. W. (2008). The role of Streptococcus pneumoniae virulence factors in host respiratory colonization and disease. Nat. Rev. Microbiol. 6, 288–301. doi: 10.1038/nrmicro1871
Kawaguchiya, M., Urushibara, N., Aung, M. S., Morimoto, S., Ito, M., Kudo, K., et al. (2018). Genetic diversity of pneumococcal surface protein A (PspA) in paediatric isolates of non-conjugate vaccine serotypes in Japan. J. Med. Microbiol. 67, 1130–1138. doi: 10.1099/jmm.0.000775
Kim, J. O., Romero-Steiner, S., Sørensen, U. B., Blom, J., Carvalho, M., Barnard, S., et al. (1999). Relationship between cell surface carbohydrates and intrastrain variation on opsonophagocytosis of Streptococcus pneumoniae. Infect. Immun. 67, 2327–2333. doi: 10.1128/iai.67.5.2327-2333.1999
Knupp-Pereira, P. A., Marques, N. T. C., Teixeira, L. M., Póvoa, H. C. C., and Neves, F. P. G. (2019). Prevalence of PspA families and pilus islets among Streptococcus pneumoniae colonizing children before and after universal use of pneumococcal conjugate vaccines in Brazil. Braz. J. Microbiol. 51, 419–425. doi: 10.1007/s42770-019-00179-y
Li, J., Li, J. W., Feng, Z., Wang, J., An, H., Liu, Y., et al. (2016). Epigenetic switch driven by DNA inversions dictates phase variation in Streptococcus pneumoniae. PLoS Pathog. 12:e1005762. doi: 10.1371/journal.pone.1005762
Li, J., and Zhang, J. R. (2019). Phase variation of Streptococcus pneumoniae. Microbiol. Spectr. 7:e005-18. doi: 10.1128/microbiolspec.GPP3-0005-2018
Manso, A. S., Chai, M. H., Atack, J. M., Furi, L., De Ste Croix, M., Haigh, R., et al. (2014). A random six-phase switch regulates pneumococcal virulence via global epigenetic changes. Nat. Commun. 5, 1–9.
Martin, D. P., Murrell, B., Golden, M., Khoosal, A., and Muhire, B. (2015). RDP4: detection and analysis of recombination patterns in virus genomes. Virus Evol. 1, 1–5.
McDaniel, L. S., McDaniel, D. O., Hollingshead, S. K., and Briles, D. E. (1998). Comparison of the PspA sequence from Streptococcus pneumoniae EF5668 to the previously identified PspA sequence from strain Rx1 and ability of PspA from EF5668 to elicit protection against pneumococci of different capsular types. Infect. Immun. 66, 4748–4754. doi: 10.1128/iai.66.10.4748-4754.1998
Melin, M. M., Hollingshead, S. K., Briles, D. E., Lahdenkari, M. I., Kilpi, T. M., and Käyhty, H. M. (2008). Development of antibodies to PspA families 1 and 2 in children after exposure to Streptococcus pneumoniae. Clin. Vaccine Immunol. 15, 1529–1535. doi: 10.1128/cvi.00181-08
Metcalf, B. J., Gertz, R. E. Jr., Gladstone, R. A., Walker, H., Sherwood, L. K., Jackson, D., et al. (2016). Strain features and distributions in pneumococci from children with invasive disease before and after 13-valent conjugate vaccine implementation in the USA. Clin. Microbiol. Infect. 22, e9–e29.
Moschioni, M., Donati, C., Muzzi, A., Masignani, V., Censini, S., Hanage, W. P., et al. (2008). Streptococcus pneumoniae contains 3 rlrA pilus variants that are clonally related. J. Infect. Dis. 197, 888–896.
Nelson, A. L., Ries, J., Bagnoli, F., Dahlberg, S., Fälker, S., Rounioja, S., et al. (2007). RrgA is a pilus-associated adhesin in Streptococcus pneumoniae. Mol. Microbiol. 66, 329–340. doi: 10.1111/j.1365-2958.2007.05908.x
Ochs, M. M., Bartlett, W., Briles, D. E., Hicks, B., Jurkuvenas, A., Lau, P., et al. (2008). Vaccine-induced human antibodies to PspA augment complement C3 deposition on Streptococcus pneumoniae. Microb. Pathog. 44, 204–214. doi: 10.1016/j.micpath.2007.09.007
Pimenta, F. C., Ribeiro-Dias, F., Brandileone, M. C., Miyaji, E. N., Leite, L. C., and Sgambatti de Andrade, A. L. (2006). Genetic diversity of PspA types among nasopharyngeal isolates collected during an ongoing surveillance study of children in Brazil. J. Clin. Microbiol. 44, 2838–2843. doi: 10.1128/jcm.00156-06
Pinto, T. C., Kegele, F. C., Dias, C. A., Barros, R. R., Peralta, J. M., Merquior, V. L., et al. (2016). Streptococcus pneumoniae serotypes 9 and 14 circulating in Brazil over a 23-year period prior to introduction of the 10-valent pneumococcal conjugate vaccine: role of international clones in the evolution of antimicrobial resistance and description of a novel genotype. Antimicrob. Agents Chemother. 60, 6664–6672. doi: 10.1128/aac.00673-16
Pinto, T. C., Souza, A. R., de Pina, S. E., Costa, N. S., Borges Neto, A. A., Neves, F. P., et al. (2013). Phenotypic and molecular characterization of optochin-resistant Streptococcus pneumoniae isolates from Brazil, with description of five novel mutations in the ATPC gene. J. Clin. Microbiol. 51, 3242–3249. doi: 10.1128/jcm.01168-13
Regev-Yochay, G., Hanage, W. P., Trzcinski, K., Rifas-Shiman, S. L., Lee, G., Bessolo, A., et al. (2010). Re-emergence of the type 1 pilus among Streptococcus pneumoniae isolates in Massachusetts, USA. Vaccine 28, 4842–4846. doi: 10.1016/j.vaccine.2010.04.042
Rolo, D., Ardanuy, C., Fleites, A., Martín, R., and Liñares, J. (2009). Diversity of pneumococcal surface protein A (PspA) among prevalent clones in Spain. BMC Microbiol. 9:80. doi: 10.1186/1471-2180-9-80
Sadowy, E., Kuch, A., Gniadkowski, M., and Hryniewicz, W. (2010). Expansion and evolution of the Streptococcus pneumoniae Spain9V-ST156 clonal complex in Poland. Antimicrob. Agents Chemother. 54, 1720–1727. doi: 10.1128/aac.01340-09
Saluja, S. K., and Weiser, J. N. (1995). The genetic basis of colony opacity in Streptococcus pneumoniae: evidence for the effect of box elements on the frequency of phenotypic variation. Mol. Microbiol. 16, 215–227. doi: 10.1111/j.1365-2958.1995.tb02294.x
Selva, L., Ciruela, P., Blanchette, K., del Amo, E., Pallares, R., Orihuela, C. J., et al. (2012). Prevalence and clonal distribution of pcpA, psrP and Pilus-1 among pediatric isolates of Streptococcus pneumoniae. PLoS One 7:e41587. doi: 10.1371/journal.pone.0041587
Sjöström, K., Blomberg, C., Fernebro, J., Dagerhamn, J., Morfeldt, E., Barocchi, M. A., et al. (2007). Clonal success of piliated penicillin nonsusceptible pneumococci. Proc. Natl. Acad. Sci. U.S.A. 104, 12907–12912. doi: 10.1073/pnas.0705589104
Swiatlo, E., Brooks-Walter, A., Briles, D. E., and McDaniel, L. S. (1997). Oligonucleotides identify conserved and variable regions of pspA and pspA-like sequences of Streptococcus pneumoniae. Gene 188, 279–284. doi: 10.1016/s0378-1119(96)00823-2
Vela-Coral, M. C., Fonseca, N., Castañeda, E., Di Fabio, J. L., Hollingshead, S. K., and Briles, D. E. (2001). Pneumococcal surface protein A of invasive Streptococcus pneumoniae isolates from Colombian children. Emerg. Infect. Dis. 7, 832–836. doi: 10.3201/eid0705.017510
Weiser, J. N., Austrian, R., Sreenivasan, P. K., and Masure, H. R. (1994). Phase variation in pneumococcal opacity: relationship between colonial morphology and nasopharyngeal colonization. Infect. Immun. 62, 2582–2589. doi: 10.1128/iai.62.6.2582-2589.1994
Keywords: Streptococcus pneumoniae, pneumococcal surface protein A, pilus type 1, colony phase variation, penicillin-non-susceptible pneumococci
Citation: Pinto TCA, Costa NS, Pina SECM, Souza ARV, Oliveira LMA, Moura CAB, Kegele FCO, Merquior VLC, Botelho ACN, Peralta JM and Teixeira LM (2020) Virulence-Associated Characteristics of Serotype 14 and Serogroup 9 Streptococcus pneumoniae Clones Circulating in Brazil: Association of Penicillin Non-susceptibility With Transparent Colony Phenotype Variants. Front. Microbiol. 11:2009. doi: 10.3389/fmicb.2020.02009
Received: 23 April 2020; Accepted: 29 July 2020;
Published: 26 August 2020.
Edited by:
Sven Hammerschmidt, University of Greifswald, GermanyReviewed by:
Dodi Safari, Eijkman Institute for Molecular Biology, IndonesiaCopyright © 2020 Pinto, Costa, Pina, Souza, Oliveira, Moura, Kegele, Merquior, Botelho, Peralta and Teixeira. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lúcia M. Teixeira, bG10MkBtaWNyby51ZnJqLmJy
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.