 Natalie Pauly1*
Natalie Pauly1* Jens A. Hammerl1
Jens A. Hammerl1 Mirjam Grobbel1
Mirjam Grobbel1 Bernd-Alois Tenhagen1
Bernd-Alois Tenhagen1 Annemarie Käsbohrer1,2
Annemarie Käsbohrer1,2 Sandra Bisenius3Jannika Fuchs4Sabine Horlacher5Holger Lingstädt6
Sandra Bisenius3Jannika Fuchs4Sabine Horlacher5Holger Lingstädt6 Ute Mauermann7Silke Mitro8Margit Müller9Stefan Rohrmann10Arthur P. Schiffmann11Birgit Stührenberg12Pia Zimmermann13
Ute Mauermann7Silke Mitro8Margit Müller9Stefan Rohrmann10Arthur P. Schiffmann11Birgit Stührenberg12Pia Zimmermann13 Stefan Schwarz14
Stefan Schwarz14 Diana Meemken15
Diana Meemken15 Alexandra Irrgang1*
Alexandra Irrgang1*- 1German Federal Institute for Risk Assessment, Berlin, Germany
- 2Unit for Veterinary Public Health and Epidemiology, University of Veterinary Medicine, Vienna, Austria
- 3Institute for Fish and Fishery Products (LAVES), Cuxhaven, Germany
- 4Chemical and Veterinary Investigation Office, Karlsruhe, Germany
- 5Chemical and Veterinary Investigation Office, Fellbach, Germany
- 6State Office for Consumer Protection Saxony-Anhalt, Stendal, Germany
- 7Food and Veterinary Institute, Oldenburg, Germany
- 8State Investigation Institute for Health and Veterinary Services, Chemnitz, Germany
- 9Chemical and Veterinary Investigation Office Rhein-Ruhr-Wupper, Krefeld, Germany
- 10Chemical and Veterinary Investigation Office, Arnsberg, Germany
- 11Hessian State Laboratory, Gießen, Germany
- 12Chemical and Veterinary Investigation Office, Detmold, Germany
- 13Bavarian Health and Food Safety Authority, Oberschleißheim, Germany
- 14Institute of Microbiology and Epizootics, Freie Universität Berlin, Berlin, Germany
- 15Institute of Food Safety and Food Hygiene, Freie Universität Berlin, Berlin, Germany
After first detections of carbapenemase-producing Enterobacteriaceae (CPE) in animals, the European Union Reference Laboratory for Antimicrobial Resistance has provided a protocol for the isolation of carbapenemase-producing Escherichia (E.) coli from cecum content and meat. Up to now, only few isolates were recovered using this procedure. In our experience, the choice of the selective agar is important for the efficacy of the method. Currently, the use of the prevailing method fails to detect CPE that exhibit a low resistance against carbapenems. Thus, this study aims to evaluate the suitability of selective media with antibiotic supplements and commercial ChromID® CARBA agar for a reliable CPE detection. For comparative investigations, detection of freeze-dried carbapenemase-resistant bacteria was studied on different batches of the ChromID® CARBA agar as well as on MacConkey agar supplemented with 1 mg/L cefotaxime and 0.125 mg/L meropenem (McC+CTX+MEM). The suitability of the different media was assessed within a time of 25 weeks, starting at least six weeks before expiration of the media. Carbapenem-resistant isolates exhibiting a serine-based hydrolytic resistance mechanism (e.g., blaKPC genes) were consistently detected over 25 weeks on the different media. In contrast, carbapenemase producers with only slightly reduced susceptibility and exhibiting a zinc-catalyzed activity (e.g., blaVIM, blaNDM, and blaIMP) could only be cultivated on long-time expired ChromID® CARBA, but within the whole test period on McC+CTX+MEM. Thus, ChromID® CARBA agar appears to be not suitable for the detection of CPE with slightly increased minimum inhibitory concentrations (MIC) against carbapenems, which have been detected in German livestock and thus, are of main interest in the national monitoring programs. Our data are in concordance with the results of eleven state laboratories that had participated in this study with their ChromID® CARBA batches routinely used for the German CPE monitoring. Based on the determined CPE detection rate, we recommend the use of McC+CTX+MEM for monitoring purposes. This study indicates that the use of ChromID® CARBA agar might lead to an underestimation of the current CPE occurrence in food and livestock samples.
Introduction
Carbapenems are important antimicrobial substances with broad activity against almost all β-lactams. They are essential to treat severe human infections with multidrug-resistant Gram-negative bacteria. Due to their high impact in human medicine, dissemination of carbapenem-resistant bacteria should be avoided (Sharland et al., 2018; European Medicines Agency [EMA], 2019). Actually, more than 2,000 variants of carbapenemase resistance genes have been described that were allocated to the β-lactamases of Ambler classes A, B, and D (Codjoe and Donkor, 2017; Grundmann et al., 2017). Klebsiella pneumoniae carbapenemases (KPC) and defined OXA enzymes are common Ambler class A and D carbapenemases, respectively, with a serine residue in their active sites (Codjoe and Donkor, 2017). New Delhi metallo-β-lactamases (NDM) and Verona integron-encoded metallo-β-lactamases (VIM) are predominant Ambler class B enzymes. Their resistance mechanism depends on the interaction of the carbapenems with zinc ions in the active site of the enzymes (Codjoe and Donkor, 2017). Carbapenemase genes are often located on mobile genetic elements (i.e., transposons/integrons, plasmids, phages), which support horizontal transmission of the genes between different bacteria (Carattoli, 2009; Miriagou et al., 2010; Tato et al., 2010). The genes are often co-located with genes mediating resistance to other antimicrobial classes. This leads to an important reduction of therapeutic options in human medicine (Woodford et al., 2014). In addition, various chromosomal mechanisms are also involved in resistance development against carbapenems, i.e., changes in membrane permeability due to the loss of specific outer membrane porins (Miriagou et al., 2010).
For a long time, carbapenemase-producing Enterobacteriaceae (CPE) were almost exclusively associated with human medicine. However, the number of reported isolates from wildlife, companion animals, livestock, and food increases worldwide (Köck et al., 2018). To get deeper insights into the dynamics of occurring CPE, the European Union (EU) set up a carbapenem-resistance monitoring for Escherichia coli isolates from food products and livestock (Schrijver et al., 2018). Aside from clinical settings, the carbapenemase enzymes OXA, VIM, NDM, and KPC possess the highest impact in wildlife, pets, and the food chain (Grundmann et al., 2017; Köck et al., 2018). In Germany, France, Italy, Spain, and the Netherlands, reports have been published about carbapenem-resistant Enterobacteriaceae (CRE) in companion animals (i.e., dogs, cats, horses) (Stolle et al., 2013; Schmiedel et al., 2014; Gonzalez-Torralba et al., 2016; Melo et al., 2017; Vittecoq et al., 2017; Pulss et al., 2018), seafood (i.e., shrimps, blue mussels, cockles) (Roschanski et al., 2016; Ceccarelli et al., 2017), wild animals (i.e., yellow-legged gulls, black kite) (Fischer et al., 2013b; Vergara et al., 2017; Vittecoq et al., 2017), and food of livestock animals (i.e., beef, chicken meat, pork) (Schwaiger et al., 2008; Leverstein-van Hall et al., 2011; Poirel et al., 2012; Zurfluh et al., 2016; Ceccarelli et al., 2017; Fischer et al., 2017; Pulss et al., 2017; Randall et al., 2017; Roschanski et al., 2017, 2018). In 2011, VIM-1-producing Enterobacteriaceae (Salmonella Infantis and E. coli) were found in several chicken and pig farms in Germany (Borowiak et al., 2017; Falgenhauer et al., 2017; Fischer et al., 2017; Irrgang et al., 2017, 2019; Roschanski et al., 2018). The European Union Reference Laboratory for Antimicrobial Resistance (EURL-AR) together with national experts on antimicrobial resistances developed a culture-dependent procedure for the isolation of carbapenemase-producing E. coli from meat and cecum content samples1. As no validated method for isolates from food exists, this method is currently used in the annual monitoring on carbapenemase-producing E. coli from livestock and food in Germany and other European member states. In Germany, we recognized on several occasions within the monitoring and during additional studies that the method failed to detect CPE that were identified in the same sample using a different method. One example was an isolate in 2017, which was detected in the monitoring on ESBL/AmpC β-lactamases but could not be detected within the German CPE monitoring. It was a VIM-1-producing E. coli from the cecum content of a fattening pig at slaughter that exhibited minimum inhibitory concentrations (MIC) ranging around the current cutoff values for the tested carbapenems. These cutoff values are considerably lower than the MIC of CPE of human clinical origin (European Food Safety Authority [EFSA], and European Centre for Disease Prevention and Control [ECDC],, 2017, 2019). Thus, there was the necessity to improve the isolation method for CPE with low carbapenem MIC values from food and livestock samples.
Due to the prevailing deficit in reliable detection of CPE exhibiting low MIC values, this study aims to determine the sensitivity of the selective cultivation after enrichment of the samples for the laboratory routine in detecting CPE. Thus, the suitability of different selective media for the recovery of CPE was assessed. Beside the frequently used ChromID® CARBA agar (bioMérieux, Nürtingen, Germany) (Nordmann et al., 2012; Girlich et al., 2013; Papadimitriou-Olivgeris et al., 2014), which is also predominantly used by the German federal state laboratories, we also determined the sensitivity of MacConkey agar supplemented with cefotaxime and meropenem over a time interval of 25 weeks.
Materials and Equipment
Bacterial Strains
Five carbapenem-resistant isolates were used for the evaluation of the different selective media (Table 1). We chose three CPE (E. coli: CP-3 and CP-9; Salmonella Corvallis: CP-5), which had previously been recovered from German food and livestock samples (Fischer et al., 2013b; Irrgang et al., 2017), one carbapenemase-producing Vibrio parahaemolyticus (CP-8) from seafood and one carbapenem-resistant K. pneumoniae isolate from a pig slaughterhouse that lacks carbapenemase production (CR-1). As references for quality assurance concerning the selectivity of the media, the ESBL-producing E. coli isolate AR-1 (Wilkinson et al., 2012) and E. coli ATCC 25922 were used.

Table 1. Strains and species used in this study with their resistance determinants (threefold determined MIC values, variations are indicated with a “/” between the values).
Further information on the bacterial isolates used in this study is summarized in Table 1. Detailed information on the MIC values of the isolates for 14 antimicrobials (ampicillin, cefotaxime, ceftazidime, chloramphenicol, ciprofloxacin, colistin, florfenicol, gentamicin, kanamycin, nalidixic acid, streptomycin, sulfamethoxazole, tetracycline, and trimethoprim) is provided in the Supplementary Table S1.
Media and Supplements
If not stated otherwise, all bacteria were cultivated in lysogeny broth (LB) at 37°C for 16–20 h under shaking conditions (120–160 rpm). Solid media agars containing 0.8% (w/v) Bacto agar (Oxoid, Dassel, Germany) were used. If necessary, antimicrobial substances were supplemented to the media as indicated. All media (listed in Table 2) of this study were stored at 4–6°C in a refrigerator and only exposed to light during processing of the experiments.

Table 2. List of media, reagents, materials, and equipment.
Methods
Antimicrobial Susceptibility Testing
The susceptibility to antimicrobials of the isolates was determined by broth microdilution using defined antimicrobial substances and concentrations from the harmonized EU panel [plates EUVSEC and EUVSEC2; TREK Diagnostic Systems (Thermo Fisher Scientific, Schwerte, Germany)]. Resistance testing was conducted according to the EN ISO20776-1:2006 (European Food Safety Authority [EFSA], and European Centre for Disease Prevention and Control [ECDC],, 2018), and MIC values were interpreted based on EUCAST definitions2 using epidemiological cutoff values. The testing was repeated three times (Supplementary Table S1). As quality control strain, E. coli ATCC 25922, was used.
Preparation of Freeze-Dried Bacteria
To ensure that equal amounts of bacterial cells were used throughout the whole study, a single batch of freeze-dried isolates was prepared using an Epsilon 2-10D LSC lyophilization instrument (Christ, Osterode am Harz, Germany) with the following procedure. Bacterial enrichments were prepared by inoculating 2 mL LB broth, supplemented with 1 mg/L cefotaxime (CTX), with a single colony of the carbapenem-resistant or ESBL-producing isolates, while cultivation of E. coli strain ATCC 25922 was conducted in LB without antimicrobials. The inoculated broths were cultured for 16–20 h at 37 ± 2°C on a rotational shaker (180 rpm). Following incubation, 150 μL of the respective cultures was transferred into 2 mL of fresh LB for further incubation at 37 ± 2°C until an optical density (OD600 nm) between 0.5 and 0.6 was reached. Thereafter, the culture was diluted to 1:10,000 (103 to 104 CFU/mL) and centrifuged at 4,000 g for 10 min. The bacterial cell pellet was resuspended in lyophilization medium [glutamic acid 10.0 g/L; NaOH (8%) 30.0 mL/L; Bacto tryptone 25.0 g/L; sucrose 50.0 g/L; thiourea (CH4N2S) 2.0 g/L] and was applied in aliquots of 1 mL to the borosilicate glasses. Lyophilization was conducted according to the standardized and validated procedure of the German Federal Institute for Risk Assessment (BfR) (freezing; condenser cooling to −50°C; vacuuming the drying chamber; main drying and post-drying). After lyophilization, the bacterial concentrations (CFU/mL) of the freeze-dried bacteria were determined under selective and non-selective conditions. The bacterial concentration after resuspension, plating on LB agar without any supplement and incubation overnight at 37°C accounted between 7.5 and 45 × 103 CFU/mL.
Enumeration of Bacteria on Different Media
To evaluate the performance of different agar types and the impact of increasing storage time of ChromID® CARBA plates, bacterial growth on different media was determined weekly over a period of 25 weeks as described below. Therefore, the respective lyophilisates of the bacteria were resuspended in 1 mL double-distilled water and incubated for 30–60 min at room temperature for adaption of the bacteria. To exclude potential effects of the commercial agar of the growth performance of the bacteria, five different ChromID® CARBA batches were tested. Aliquots (50 μL) of the lyophilisate suspensions were plated on ChromID® CARBA plates. For comparative analysis of the commercial agar to other media, the same lyophilisate suspensions were also applied to LB agar and to MacConkey agar supplemented with 1 mg/L CTX and 0.125 mg/L meropenem (MEM) (McC+CTX+MEM). Both LB agar and McC+CTX+MEM were produced in-house at the BfR. After an incubation period of 16–18 h at 37 ± 2°C, the bacterial concentration (CFU/mL) was determined. For comparability, the storage time the bacterial growth on the five ChromID® CARBA batches was calculated according to the expiry date.
Freeze-dried bacteria were also provided to 11 participating federal state laboratories for comparative investigation. The laboratories of Baden-Württemberg (Fellbach, Karlsruhe), Bavaria (Oberschleißheim), Hesse (Gießen), Lower Saxony (Cuxhaven, Oldenburg), North Rhine-Westphalia (Arnsberg, Detmold, Krefeld), Saxony (Chemnitz), and Saxony-Anhalt (Stendal) conducted growth investigation according to the specifications of the BfR. Overall, the selectivity of the ChromID® CARBA agar was tested on seven different batches. In contrast to the long-term procedure of the BfR, the federal state laboratories investigated the growth performance of the different isolates on different selective media at three defined time points: (i) at the expiry date, (ii) 2 weeks before, and (iii) 5 weeks after the expiry date. The systematic procedure of all steps is shown in Figure 1.
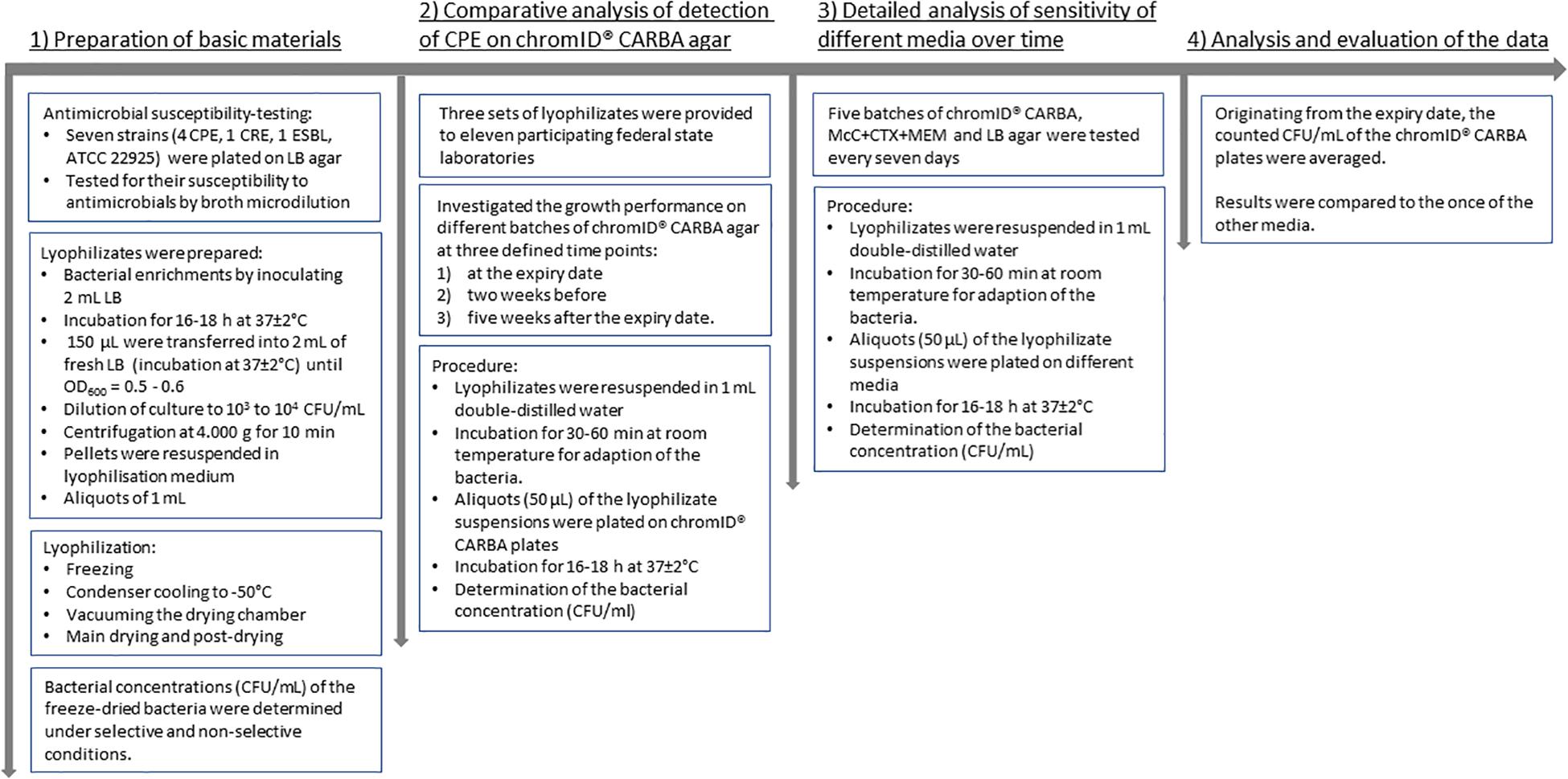
Figure 1. Flow diagram of the systematic procedure.
Results
Commercial selective medium is not suitable for routine monitoring purposes on isolates from livestock and food that exhibit low carbapenem MIC values.
Therefore, the commercial ChromID® CARBA agar was tested out of date because of previous observations and experiences. Freeze-dried bacteria, which were produced within this study, showed growth between 5 × 102 CFU/mL and 4.5 × 104 CFU/mL (Figure 2). Over the 25-week period, the bacterial titer of the isolates did not change significantly under non-selective conditions (LB). However, we observed a slight decrease in the bacterial titer for the isolates AR-1, CR-1, and CP-8 over the first time points of enumeration (Figure 2). After 25 weeks, all isolates were still reliably detectable under non-selective conditions (Figure 2).
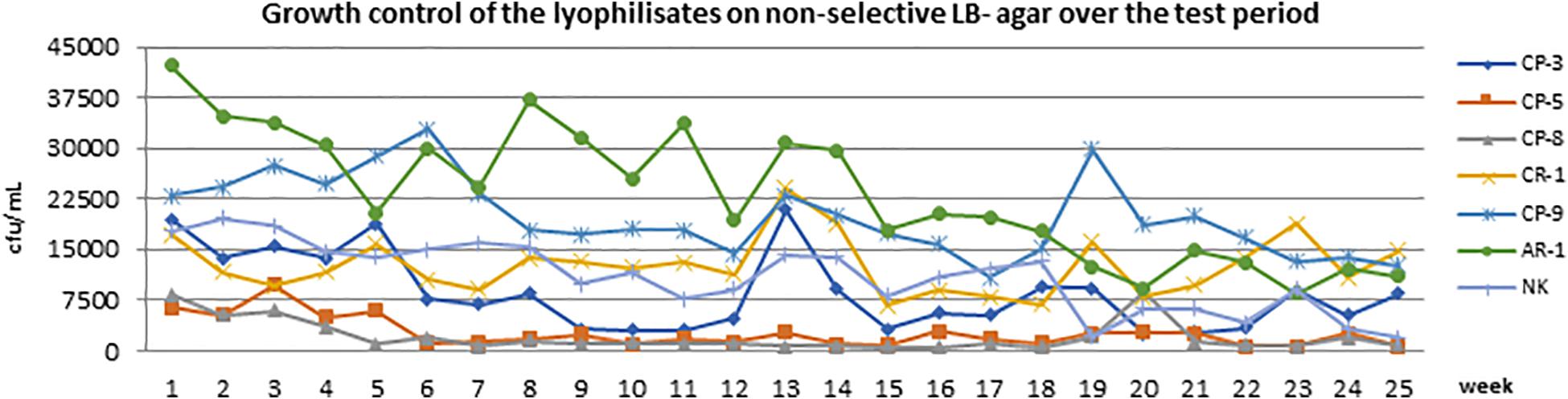
Figure 2. The growth of all lyophilisates on non-selective LB agar over the entire test period of 25 weeks. The x-axis (abscissa) shows the time in weeks, and the y-axis (ordinate) shows CFU/mL.
The results of the comparative analysis are summarized in Supplementary Table S2. Results of comparative testing of the different federal state laboratories were very similar. They indicated that only one isolate (CP-9) could be detected on the used agar batches at all three time points. This respective isolate, a human E. coli which carried the gene blaKPC–2, exhibited a high MIC values against the tested carbapenems (Table 1). None of the other isolates showed bacterial growth at the selected time points.
To investigate the suitability of the ChromID® CARBA agar for the detection of CPE exhibiting low carbapenem MIC values, the growth ability of the aforementioned isolates was investigated at the NRL-AR over a time period of 25 weeks using five different batches of the agar. The diagrams 2A–2E show that at the beginning of the test, bacterial growth could only be detected for isolate CP-9, exhibiting high MIC values for the carbapenems ertapenem, imipenem, and meropenem (Table 1 and Supplementary Table S1). Slight bacterial growth could be detected for the isolates CP-3, CP-5, and CP-8 2 weeks before expiry date (week 7) (Figures 3A–C). This detection increased over time and also after the expiry date. The carbapenemase-producing isolate CP-9 showed a highly reliable average growth rate between 5.0 × 103 and 2.5 × 104 CFU/mL at all points of investigations over the 25 weeks (Figure 3D). At the end of the trial, four carbapenem-resistant isolates (CP-3; CP-5; CP-8; and CR-1) showed increased growth rates (average: CP-3 with blaVIM–1: 4 to 32 CFU/mL; CP-5 with blaNDM–1: 4 to 368 CFU/mL; and CP-8 with blaVIM–1: 4 to 24 CFU/mL). The CR-1 isolate that exhibits no carbapenemase-production showed growth rates between 20 and 3920 CFU/mL and was first detected 1 week before the expiry date of the media (Figure 3E). The quality control isolate AR-1 (Table 1 and Supplementary Table S1) and the control strain ATCC 25922 did not grow on ChromID® CARBA plates during the investigated period.
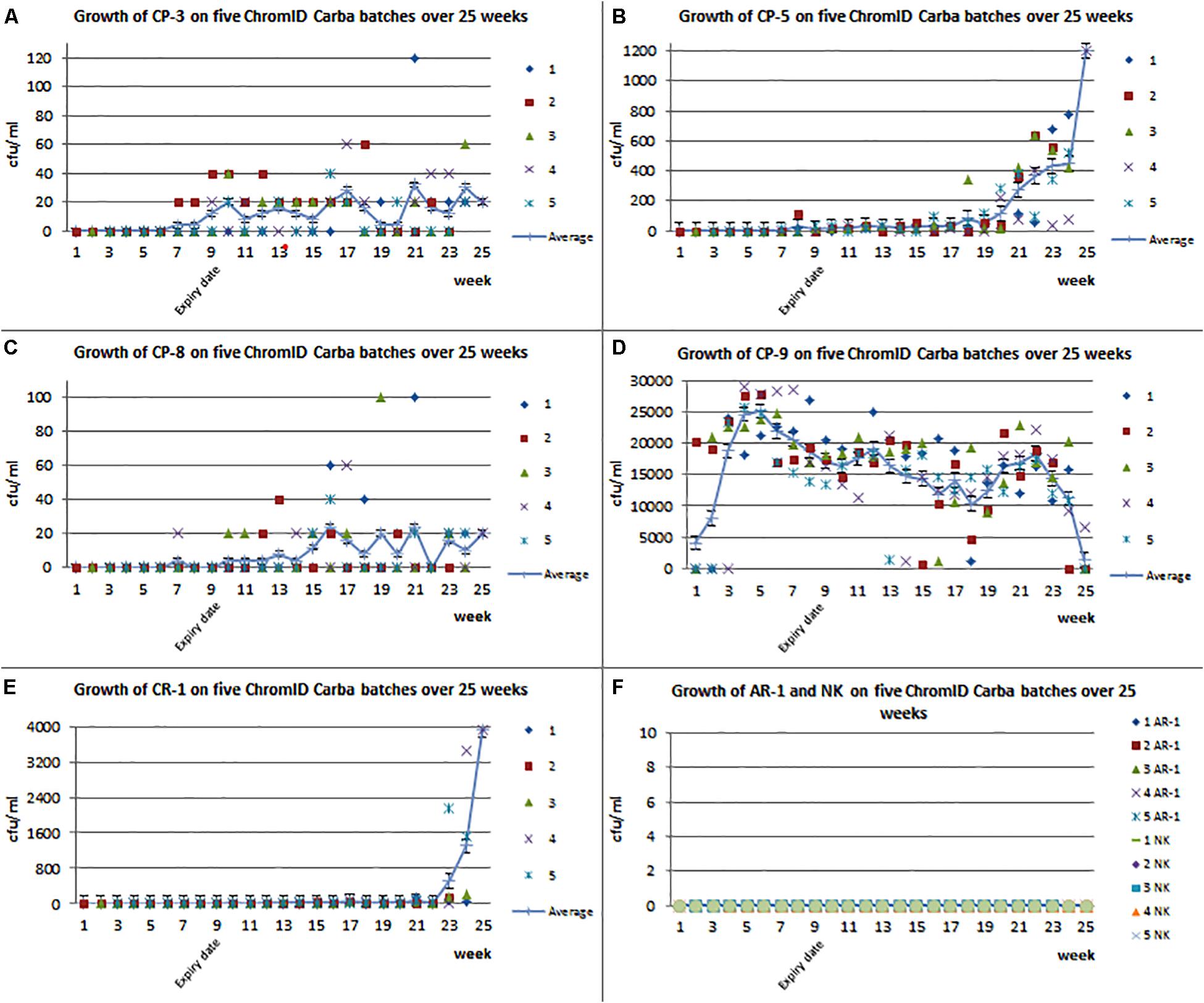
Figure 3. The x-axis shows the time in weeks where “9” the expiry date is. The y-axis shows CFU/mL. (A) Growth of CP-3 on five tested batches of ChromID® CARBA agar. Moreover, the average of all batches is shown. (B) Growth of CP-5 on five tested batches of ChromID® CARBA agar. Moreover, the average of all batches is shown. (C) Growth of CP-8 on five tested batches of ChromID® CARBA agar. Moreover, the average of all batches is shown. (D) Growth of CP-9 on five tested batches of ChromID® CARBA agar. Moreover, the average of all batches is shown. (E) Growth of CR-1 on five tested batches of ChromID® CARBA agar. Moreover, the average of all batches is shown. (F) Growth of the both control strains NK and AR-1 on five tested batches of ChromID® CARBA agar.
Supplementing 1 mg/L CTX and 0.125 mg/L MEM to the agar basis of the MacConkey medium can overcome growth inhibition during cultivation of low-carbapenem MIC-exhibiting isolates.
The growth performance of the abovementioned isolates was also determined on McC+CTX+MEM agar. In contrast to ChromID® CARBA agar, all carbapenem-resistant isolates were reliably detected at all time points within the 25-week period of investigation (Table 3). The growth of the CP-9 isolate was similar on McC+CTX+MEM, ChromID® CARBA agar as well as under non-selective conditions (LB agar) ranging between 2.4 × 103 and 3.3 × 104 CFU/mL. With the focus on the isolates (AR-1 and NK) used as quality controls, McC+CTX+MEM has a reliable specificity of up to 22 weeks, as within this period no bacterial growth was observed.
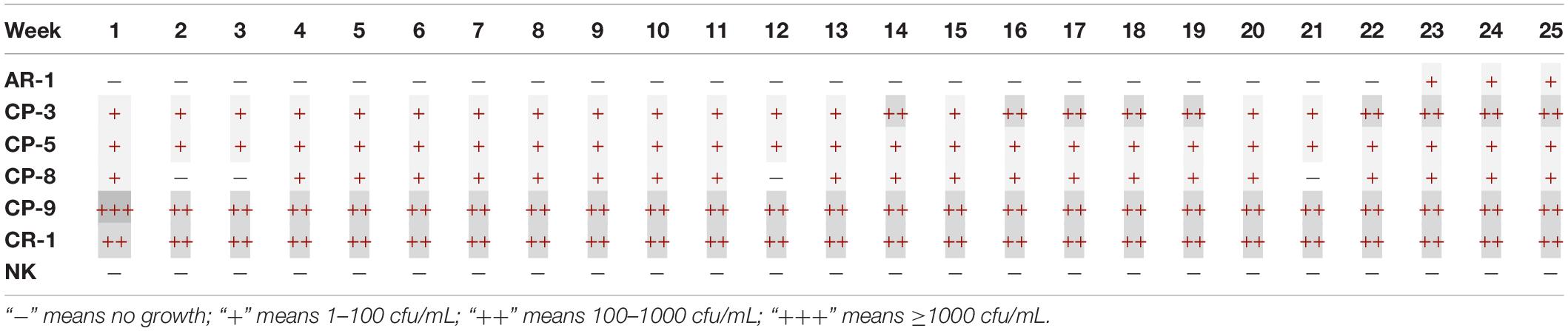
Table 3. Growth of the seven lyophilisates tested once a week over 25 weeks on MacConkey agar supplemented with 0.125 mg/L meropenem and 1 mg/L cefotaxime.
Discussion
Previous experiments in our laboratory showed that the ChromID® CARBA agar becomes more sensitive with increasing storage time. Based on the observations and experiences within the laboratory routine (International Organization for Standardization, 2006; Fischer et al., 2013a, b, 2017; Irrgang et al., 2017, 2019), detection of carbapenemase-producing bacteria is challenging using the EURL-AR method in combination with ChromID® CARBA agar. We assume that the concentration of different ingredients and thus the selectivity will decrease with time or after the expiry date. This justifies the choice of times to determine a possible decrease in antimicrobial concentration after the shelf life of ChromID® CARBA agar.
Within this study, we observed strong differences in the growth performance of the used carbapenem-resistant isolates on the selective media. These different growth performances exceed the observed slight decrease of AR-1, CR-1, and CP-8 during the first time points of enumeration, which seems to represent a natural adaption of the specific isolates to the prevailing cultivation conditions (Figure 2).
As previously reported (Fischer et al., 2012; Köck et al., 2018), carbapenem-resistant isolates from food products or livestock exhibit significantly lower MIC values for the respective carbapenems than clinical isolates. There may be different reasons for this observation. It could be due to molecular conditions, such as a variable number of gene copies, a different expression of the genes, or a different genetic localization (plasmid versus chromosomal DNA). It could also be due to the different antimicrobial selection pressures to which the strains are exposed. As carbapenems are not licensed for use in food animals, previous exposure of the strains with a livestock origin is unlikely. The chosen isolates for the evaluation of the selective cultivation step of the detection procedure represent different bacterial genera and carbapenem resistance genes as well as resistance mechanisms (Viau et al., 2012; Köck et al., 2018). The E. coli isolate CP-9 carries a blaKPC–2 gene and exhibited high MIC values against all tested carbapenems. CP-9 represents typical MIC values for carbapenems of human isolates (Supplementary Table S1). Other target isolates, except CP-5 (the NDM-producing Salmonella) exhibited carbapenem resistance but showed only MICs ranging around the current cutoff values (Supplementary Table S1). As the used isolates exhibited very low MIC values, we suppose that the remaining concentration of the supplemented antimicrobials is still high enough to prevent a growth of these isolates.
The KPC enzymes belong to Amber class A carbapenemases and use a serine-based hydrolytic mechanism to inactivate carbapenems. In contrast, the VIM-1 and NDM-1 enzymes belong to Ambler class B carbapenemases and convey the resistances by a zinc-catalyzing active site (Codjoe and Donkor, 2017). Viau et al. (2016) reported that most of their ten tested media performed reasonably well with different screening methods for the detection of class A enzymes. Performance with class B and D enzymes was more variable. However, Viau and co-workers used up to 180 different CRE originating from human clinical sources. They determined a good performance of the ChromID® CARBA medium if tested with rectal/perirectal swabs (Jeon et al., 2015). The differences between CPE from human samples and CPE from non-clinical animal samples have not been investigated systematically so far as the number of available non-clinical animal isolates is still limited. As the ingredients of the ChromID® CARBA agar are unknown and have not been provided by the manufacturers on request, it is difficult to assess the selective additives of the medium and to expect their influence on the cultivation of CPE with low MIC. It might be that an additive supports the serine-based hydrolytic resistance mechanism in a better way than the zinc-based mechanism.
Within our study, CP-9 was the only isolate that was consistently detected in all participating laboratories and with all selective media. The determined amount of the adjusted, freeze-dried bacteria ranged between 5 × 102 and 4.5 × 104 CFU/mL between the laboratories. The other carbapenem-resistant isolates (CP-3; CP-5; CP-8; and CR-1) showed no detectable growth on the ChromID® CARBA plates within the different federal state laboratories up to 5 weeks after expiration date of the plates (Supplementary Table S2). At this time point, the agar will not be used by accredited laboratories as the agar will be handled as expired according to the recommendations of the manufacturers. However, each of these isolates was reliably detected on McC+CTX+MEM (Table 3). The quality control isolate AR-1, a CTX-M-1-producing E. coli with a low phenotypical resistance against ertapenem (Table 1 and Supplementary Table S1), and the quality control strain ATCC 25922 did not grow on ChromID® CARBA plates at any time (Figure 3F). As they grew at the last three time points on McC+CTX+MEM, it can be assumed that the specificity of the plates did not decrease considerably within a period of 20 weeks after production (Table 3). In contrast, the number of CFU of the isolates CP-3; CP-5; CP-8; and CR-1 increased at the end of the investigated time interval. This indicates that the sensitivity of the commercial agar increased over time.
In contrast to our study, other groups found that ChromID® CARBA agar was one of the most sensitive and specific chromogenic medium for the detection of CPE (Vrioni et al., 2012; Papadimitriou-Olivgeris et al., 2014; Simner et al., 2015; Viau et al., 2016). However, in these studies, the MIC values of the CPE are in general much higher than the MIC values of the isolates tested here. In comparison, Vrioni et al. (2012) used clinical isolates with very high MIC values of up to over 32 mg/L to MEM, but they also used some strains with MIC values below the resistance breakpoint. The detection of the isolates with these high MIC values might correlate much more with the detection of CP-9 and is not comparable with the detection of isolates exhibiting low MICs (CP-3, CP-8, and CR-1).
Limited detection of CPE might be also attributable to a loss of the resistance-harboring plasmids during lyophilization (Wagman and Weneck, 1963; Berman et al., 1968; Wein et al., 2019). We can exclude this as an important factor in our study because we observed similar bacterial titers on selective and non-selective in-house media (data not shown). We had also confirmed the presence of the plasmid in some representative colonies from selective and non-selective media.
Overall, we have detected more CFU/mL by using the conventional MacConkey agar supplemented with antibiotics over the entire period for almost all isolates of this study. Due to the lacking resistance against carbapenems, the quality control strain ATCC 25922 could not be detected on McC+CTX+MEM, indicating the appropriate specificity of the medium. Some spontaneous growth of the E. coli isolate AR_1 was observed 22, 23, and 24 weeks after agar production, indicating that the prepared agar worked with a high specificity for up to 22 weeks. In contrast to this observation, the AR_1 isolate could not be detected on the ChromID® CARBA agar. The concentration of meropenem (0.125 mg/L) in the supplemented agar is lower than the current ECOFFs (0.25 mg/L) (Giske et al., 2013). Based on this concentration, CPE with low MIC values are also able to grow successfully and will further be detected within the monitoring programs. Viau et al. (2012) reported about “silent” dissemination of K. pneumoniae, because some isolates were not identified as carbapenem-resistant based on their MIC, even though they carried a carbapenemase gene and showed slightly increased MIC values against ertapenem and meropenem (Fischer et al., 2012).
Berthoin et al. (2010) reported that the stability of meropenem in solutions depends on time, temperature, and concentration (Berthoin et al., 2010). These observations were supported by Keel et al. (2011). A potential effect of light on the stability of meropenem can be excluded, as the agar plates were only exposed to light during plating and visual inspection. Therefore, it was assumed that the stability of meropenem in the media might be limited. In our study, the McC+CTX+MEM agar was stably usable for up to 22 weeks without a loss of specificity when stored at 4 ± 2°C. The supplementation with cefotaxime supported the selective effect, but for OXA-48 producers this antibiotic should be omitted. Changes in sensitivity and specificity of MacConkey agar supplemented with meropenem only over time were not tested in our study. Adler et al. (2011) used MacConkey agar supplemented with imipenem at 1 mg/mL (McC+IMI) and compared this medium with a commercial selective medium (CHROMagar KPC, CHROMagar Company, Paris, France). They described a similar performance of both media during clinical evaluation. Moreover, they argued with the lower costs of McC+IMI to prefer it for detection of CRE in their study (Adler et al., 2011). We decided to use meropenem as supplement because it is more stable than imipenem (Zhanel et al., 2007). Based on our data, we recommend using McC+CTX+MEM for selective cultivation of carbapenem-resistant isolates from food products and livestock samples.
Another important difference between the different agars is that the ingredients of the in-house prepared agar are known. In McC+CTX+MEM agar, no further substances were supplemented for inhibition of accompanying bacteria. It is possible that the commercial selective medium comprises unspecified substances (e.g., zinc that is used to inhibit non-carbapenemase-producing bacteria), which may also suppress the growth of some CPE, e.g., with a low MIC. Although less selective substances are used, no increased difficulties were found with the accompanying flora in other experiments so far.
A limitation of this study is that only one batch of McC+CTX+MEM plates was used throughout the experiment. This was due to its function as a control medium, and it was not the primary aim to compare both media.
Conclusion
The results of our study indicate that the ChromID® CARBA agar is not suitable for the cultural detection of CPE with low MIC to carbapenems, while clinical isolates with higher MIC values can be detected reliably. Overall, the use of the ChromID® CARBA agar in monitoring programs may lead to an underestimation of the CPE occurrence in livestock and food as the method lacks sensitivity.
We further provide data, that MacConkey agar with 1 mg/L cefotaxime and 0.125 mg/L meropenem provides a better sensitivity for the detection of such isolates and may be more suitable for the carbapenemase monitoring in livestock and food.
Data Availability Statement
All datasets presented in this study are included in the article/Supplementary Material.
Ethics Statement
This study uses strains obtained from humans (E. coli CP-9), livestock (E. coli AR-1 and CP-3, Klebsiella pneumoniae CR-1), wildlife (Salmonella Corvallis CP-5) and seafood (Vibrio parahaemolyticus CP-8). All isolates were part of the strain collection of the German NRL AR and had been submitted to the NRL in the framework of its routine tasks in monitoring and surveillance programs or during previous scientific collaborations. None of the isolates was collected specifically for the current study. Therefore we did not consider it necessary to seek for ethical approval for using the isolates.
Author Contributions
JH, NP, and AI designed the study and developed the draft of the manuscript. NP performed the preparation of adjusted, freeze-dried bacteria, and the determination of the growth performance of the isolates at the NRL-AR. MG, AI, and JH initially characterized the carbapenem-resistant isolates. SB, JF, SH, HL, UM, MM, SR, AS, SS, BS, and PZ performed the individual growth investigations on the commercial agar at the different federal state laboratories. NP, AI, JH, MG, AK, B-AT, SS, and DM performed the data analysis. All authors supported the finalization of the manuscript and supported editing of the final version.
Funding
This work was financially supported by a grant of the German Federal Institute for Risk Assessment (43-002). The work of NP was supported by the European Joint Project (EJP) IMPART funded by the European Union’s Horizon 2020 Research and Innovation Program under Grant Agreement No. 773830.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We gratefully acknowledge the support of the federal state laboratories and authorities in collecting the samples and providing the recovered isolates in the framework of the different German monitoring programs. We also thank the laboratory team (Britta Lesniewsky, Anja Seeger, Tanja Skladnikiewicz-Ziemer, Silke Jahn, Romy Fuhrmeister, Yvonne Klaar, and Silvia Schmoger) of the NRL-AR for excellent technical assistance. We further acknowledge the support of Peter Bahn (German Federal Institute for Risk Assessment) during preparation of the freeze-dried bacteria.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2020.01678/full#supplementary-material
Footnotes
References
Adler, A., Navon-Venezia, S., Moran-Gilad, J., Marcos, E., Schwartz, D., and Carmeli, Y. (2011). Laboratory and clinical evaluation of screening agar plates for detection of carbapenem-resistant Enterobacteriaceae from surveillance rectal swabs. J. Clin. Microbiol. 49, 2239–2242. doi: 10.1128/jcm.02566-10
Berman, S., Altieri, P. L., Groffinger, A., Lowenthal, J. P., and Formal, S. B. (1968). Freeze-drying various strains of Shigella. Appl. Microbiol. 16, 1779–1781. doi: 10.1128/aem.16.11.1779-1781.1968
Berthoin, K., Le Duff, C. S., Marchand-Brynaert, J., Carryn, S., and Tulkens, P. M. (2010). Stability of meropenem and doripenem solutions for administration by continuous infusion. J. Antimicrob. Chemother. 65, 1073–1075. doi: 10.1093/jac/dkq044
Borowiak, M., Szabo, I., Baumann, B., Junker, E., Hammerl, J. A., Kaesbohrer, A., et al. (2017). VIM-1-producing Salmonella Infantis isolated from swine and minced pork meat in Germany. J. Antimicrob. Chemother. 72, 2131–2133. doi: 10.1093/jac/dkx101
Carattoli, A. (2009). Resistance plasmid families in Enterobacteriaceae. Antimicrob. Agents Chemother. 53, 2227–2238. doi: 10.1128/aac.01707-08
Ceccarelli, D., van Essen-Zandbergen, A., Veldman, K. T., Tafro, N., Haenen, O., and Mevius, D. J. (2017). Chromosome-Based blaOXA-48-Like Variants in Shewanella species isolates from food-producing animals, fish, and the aquatic environment. Antimicrob. Agents Chemother. 61:e01013-16.
Codjoe, F. S., and Donkor, E. S. (2017). Carbapenem resistance: a review. Med. Sci. 6:1. doi: 10.3390/medsci6010001
European Food Safety Authority [EFSA], and European Centre for Disease Prevention and Control [ECDC], (2017). The European Union Summary Report on Antimicrobial Resistance in Zoonotic and Indicator Bacteria from Humans, Animals and Food in 2015. Parma: EFSA. 176–192.
European Food Safety Authority [EFSA], and European Centre for Disease Prevention and Control [ECDC], (2018). The European Union summary report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2016. EFSA J. 16, 223–250.
European Food Safety Authority [EFSA], and European Centre for Disease Prevention and Control [ECDC], (2019). The European Union Summary Report on Antimicrobial Resistance in Zoonotic and Indicator Bacteria from Humans, Animals and Food in 2017. Parma: EFSA. 223–250.
European Medicines Agency [EMA] (2019). List of Medicinal Products Under Additional Monitoring. Amsterdam: EMA.
Falgenhauer, L., Ghosh, H., Guerra, B., Yao, Y., Fritzenwanker, M., Fischer, J., et al. (2017). Comparative genome analysis of IncHI2 VIM-1 carbapenemase-encoding plasmids of Escherichia coli and Salmonella enterica isolated from a livestock farm in Germany. Vet. Microbiol. 200, 114–117. doi: 10.1016/j.vetmic.2015.09.001
Fischer, J., Rodriguez, I., Schmoger, S., Friese, A., Roesler, U., Helmuth, R., et al. (2012). Escherichia coli producing VIM-1 carbapenemase isolated on a pig farm. J. Antimicrob. Chemother. 67, 1793–1795. doi: 10.1093/jac/dks108
Fischer, J., Rodriguez, I., Schmoger, S., Friese, A., Roesler, U., Helmuth, R., et al. (2013a). Salmonella enterica subsp. enterica producing VIM-1 carbapenemase isolated from livestock farms. J. Antimicrob. Chemother. 68, 478–480. doi: 10.1093/jac/dks393
Fischer, J., San Jose, M., Roschanski, N., Schmoger, S., Baumann, B., Irrgang, A., et al. (2017). Spread and persistence of VIM-1 Carbapenemase-producing Enterobacteriaceae in three German swine farms in 2011 and 2012. Vet. Microbiol. 200, 118–123. doi: 10.1016/j.vetmic.2016.04.026
Fischer, J., Schmoger, S., Jahn, S., Helmuth, R., and Guerra, B. (2013b). NDM-1 carbapenemase-producing Salmonella enterica subsp. enterica serovar Corvallis isolated from a wild bird in Germany. J. Antimicrob. Chemother. 68, 2954–2956. doi: 10.1093/jac/dkt260
Girlich, D., Poirel, L., and Nordmann, P. (2013). Comparison of the SUPERCARBA, CHROMagar KPC, and Brilliance CRE screening media for detection of Enterobacteriaceae with reduced susceptibility to carbapenems. Diagn. Microbiol. Infect. Dis. 75, 214–217. doi: 10.1016/j.diagmicrobio.2012.10.006
Giske, C. G., Canton, R., Stefani, S., Skov, R., Glupczynski, Y., Nordmann, P. N., et al. (2013). EUCAST Guidelines for Detection of Resistance Mechanisms and Specific Resistances of Clinical and/or Epidemiological Importance. Sweden: EUCAST.
Gonzalez-Torralba, A., Oteo, J., Asenjo, A., Bautista, V., Fuentes, E., and Alos, J. I. (2016). Survey of carbapenemase-producing Enterobacteriaceae in companion dogs in Madrid, Spain. Antimicrob. Agents Chemother. 60, 2499–2501. doi: 10.1128/aac.02383-15
Grundmann, H., Glasner, C., Albiger, B., Aanensen, D. M., Tomlinson, C. T., Andrasevic, A. T., et al. (2017). Occurrence of carbapenemase-producing Klebsiella pneumoniae and Escherichia coli in the European survey of carbapenemase-producing Enterobacteriaceae (EuSCAPE): a prospective, multinational study. Lancet Infect. Dis. 17, 153–163.
International Organization for Standardization (2006). Clinical Laboratory Testing and In Vitro Diagnostic Test System - Susceptibility Testing of Infectious Agents and Evaluation of Performance of Antimicrobial Susceptibility Test Devices. Geneva: International Organization for Standardization.
Irrgang, A., Fischer, J., Grobbel, M., Schmoger, S., Skladnikiewicz-Ziemer, T., Thomas, K., et al. (2017). Recurrent detection of VIM-1-producing Escherichia coli clone in German pig production. J. Antimicrob. Chemother. 72, 944–946.
Irrgang, A., Tenhagen, B.-A., Pauly, N., Schmoger, S., Kaesbohrer, A., and Hammerl, J. A. (2019). Characterization of VIM-1-producing E. coli isolated from a german fattening pig farm by an improved isolation procedure. Front. Microbiol. 10:2256. doi: 10.3389/fmicb.2019.02256
Jeon, J. H., Lee, J. H., Lee, J. J., Park, K. S., Karim, A. M., Lee, C.-R., et al. (2015). Structural basis for carbapenem-hydrolyzing mechanisms of carbapenemases conferring antibiotic resistance. Int. J. Mol. Sci. 16, 9654–9692. doi: 10.3390/ijms16059654
Keel, R. A., Sutherland, C. A., Crandon, J. L., and Nicolau, D. P. (2011). Stability of doripenem, imipenem and meropenem at elevated room temperatures. Int. J. Antimicrob. Agents 37, 184–185. doi: 10.1016/j.ijantimicag.2010.06.043
Köck, R., Daniels-Haardt, I., Becker, K., Mellmann, A., Friedrich, A. W., Mevius, D., et al. (2018). Carbapenem-resistant Enterobacteriaceae in wildlife, food-producing, and companion animals: a systematic review. Clin. Microbiol. Infect. 24, 1241–1250. doi: 10.1016/j.cmi.2018.04.004
Leverstein-van Hall, M. A., Dierikx, C. M., Stuart, J. C., Voets, G. M., van den Munckhof, M. P., van Essen-Zandbergen, A., et al. (2011). Dutch patients, retail chicken meat and poultry share the same ESBL genes, plasmids and strains. Clin. Microbiol. Infect. 17, 873–880. doi: 10.1111/j.1469-0691.2011.03497.x
Melo, L. C., Boisson, M. N., Saras, E., Medaille, C., Boulouis, H. J., Madec, J. Y., et al. (2017). OXA-48-producing ST372 Escherichia coli in a French dog. J. Antimicrob. Chemother. 72, 1256–1258.
Miriagou, V., Cornaglia, G., Edelstein, M., Galani, I., Giske, C. G., Gniadkowski, M., et al. (2010). Acquired carbapenemases in Gram-negative bacterial pathogens: detection and surveillance issues. Clin. Microbiol. Infect. 16, 112–122. doi: 10.1111/j.1469-0691.2009.03116.x
Nordmann, P., Girlich, D., and Poirel, L. (2012). Detection of carbapenemase producers in Enterobacteriaceae by use of a novel screening medium. J. Clin. Microbiol. 50, 2761–2766. doi: 10.1128/jcm.06477-11
Papadimitriou-Olivgeris, M., Bartzavali, C., Christofidou, M., Bereksi, N., Hey, J., Zambardi, G., et al. (2014). Performance of chromID® CARBA medium for carbapenemases-producing Enterobacteriaceae detection during rectal screening. Eur. J. Clin. Microbiol. Infect. Dis. 33, 35–40. doi: 10.1007/s10096-013-1925-6
Poirel, L., Berçot, B., Millemann, Y., Bonnin, R. A., Pannaux, G., and Nordmann, P. (2012). Carbapenemase-producing Acinetobacter spp. in Cattle, France. Emerg. Infect. Dis. 18, 523–525.
Pulss, S., Semmler, T., Prenger-Berninghoff, E., Bauerfeind, R., and Ewers, C. (2017). First report of an Escherichia coli strain from swine carrying an OXA-181 carbapenemase and the colistin resistance determinant MCR-1. Int. J. Antimicrob. Agents 50, 232–236. doi: 10.1016/j.ijantimicag.2017.03.014
Pulss, S., Stolle, I., Stamm, I., Leidner, U., Heydel, C., Semmler, T., et al. (2018). Multispecies and clonal dissemination of OXA-48 carbapenemase in Enterobacteriaceae From Companion Animals in Germany, 2009-2016. Front. Microbiol. 9:1265. doi: 10.3389/fmicb.2018.01265
Randall, L. P., Lodge, M. P., Elviss, N. C., Lemma, F. L., Hopkins, K. L., Teale, C. J., et al. (2017). Evaluation of meat, fruit and vegetables from retail stores in five United Kingdom regions as sources of extended-spectrum beta-lactamase (ESBL)-producing and carbapenem-resistant Escherichia coli. Int. J. Food Microbiol. 241, 283–290. doi: 10.1016/j.ijfoodmicro.2016.10.036
Roschanski, N., Friese, A., von Salviati-Claudius, C., Hering, J., Kaesbohrer, A., Kreienbrock, L., et al. (2017). Prevalence of carbapenemase producing Enterobacteriaceae isolated from German pig-fattening farms during the years 2011-2013. Vet. Microbiol. 200, 124–129. doi: 10.1016/j.vetmic.2015.11.030
Roschanski, N., Friese, A., von Salviati-Claudius, C., Hering, J., Kaesbohrer, A., Kreienbrock, L., et al. (2018). Retrospective analysis of bacterial cultures sampled in german chicken-fattening farms during the years 2011-2012 revealed additional VIM-1 carbapenemase-producing Escherichia coli and a serologically rough Salmonella enterica Serovar Infantis. Front. Microbiol. 9:538. doi: 10.3389/fmicb.2018.00538
Roschanski, N., Guenther, S., Vu, T. T. T., Fischer, J., Semmler, T., Huehn, S., et al. (2016). VIM-1 carbapenemase-producing Escherichia coli isolated from retail seafood, Germany 2016. Euro Surveill. 22:17-00032.
Schmiedel, J., Falgenhauer, L., Domann, E., Bauerfeind, R., Prenger-Berninghoff, E., Imirzalioglu, C., et al. (2014). Multiresistant extended-spectrum beta-lactamase-producing Enterobacteriaceae from humans, companion animals and horses in central Hesse, Germany. BMC Microbiol. 14:187. doi: 10.1186/1471-2180-14-187
Schrijver, R., Stijntjes, M., Rodriguez-Bano, J., Tacconelli, E., Babu Rajendran, N., and Voss, A. (2018). Review of antimicrobial resistance surveillance programmes in livestock and meat in EU with focus on humans. Clin. Microbiol. Infect. 24, 577–590. doi: 10.1016/j.cmi.2017.09.013
Schwaiger, K., Schmied, E. M., and Bauer, J. (2008). Comparative analysis of antibiotic resistance characteristics of Gram-negative bacteria isolated from laying hens and eggs in conventional and organic keeping systems in Bavaria, Germany. Zoonoses Public Health 55, 331–341. doi: 10.1111/j.1863-2378.2008.01151.x
Sharland, M., Pulcini, C., Harbarth, S., Zeng, M., Gandra, S., Mathur, S., et al. (2018). Classifying antibiotics in the WHO Essential Medicines List for optimal use. Lancet Infect. Dis. 18, 18–20. doi: 10.1016/s1473-3099(17)30724-7
Simner, P. J., Gilmour, M. W., DeGagne, P., Nichol, K., and Karlowsky, J. A. (2015). Evaluation of five chromogenic agar media and the Rosco Rapid Carb screen kit for detection and confirmation of carbapenemase production in Gram-negative bacilli. J. Clin. Microbiol. 53, 105–112. doi: 10.1128/jcm.02068-14
Stolle, I., Prenger-Berninghoff, E., Stamm, I., Scheufen, S., Hassdenteufel, E., Guenther, S., et al. (2013). Emergence of OXA-48 carbapenemase-producing Escherichia coli and Klebsiella pneumoniae in dogs. J. Antimicrob. Chemother. 68, 2802–2808. doi: 10.1093/jac/dkt259
Tato, M., Coque, T. M., Baquero, F., and Canton, R. (2010). Dispersal of carbapenemase blaVIM-1 gene associated with different Tn402 variants, mercury transposons, and conjugative plasmids in Enterobacteriaceae and Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 54, 320–327. doi: 10.1128/aac.00783-09
Vergara, A., Pitart, C., Montalvo, T., Roca, I., Sabate, S., Hurtado, J. C., et al. (2017). Prevalence of extended-spectrum-beta-lactamase- and/or carbapenemase-producing Escherichia coli isolated from yellow-legged gulls from barcelona, Spain. Antimicrob. Agents Chemother. 61:e02071-16.
Viau, R., Frank, K. M., Jacobs, M. R., Wilson, B., Kaye, K., Donskey, C. J., et al. (2016). Intestinal carriage of carbapenemase-producing organisms: current status of surveillance methods. Clin. Microbiol. Rev. 29, 1–27. doi: 10.1128/cmr.00108-14
Viau, R. A., Hujer, A. M., Marshall, S. H., Perez, F., Hujer, K. M., Briceño, D. F., et al. (2012). “Silent” dissemination of Klebsiella pneumoniae isolates bearing K. pneumoniae carbapenemase in a long-term care facility for children and young adults in Northeast Ohio. Clin. Infect. Dis. 54, 1314–1321. doi: 10.1093/cid/cis036
Vittecoq, M., Laurens, C., Brazier, L., Durand, P., Elguero, E., Arnal, A., et al. (2017). VIM-1 carbapenemase-producing Escherichia coli in gulls from southern France. Ecol. Evol. 7, 1224–1232. doi: 10.1002/ece3.2707
Vrioni, G., Daniil, I., Voulgari, E., Ranellou, K., Koumaki, V., Ghirardi, S., et al. (2012). Comparative evaluation of a prototype chromogenic medium (ChromID CARBA) for detecting carbapenemase-producing Enterobacteriaceae in surveillance rectal swabs. J. Clin. Microbiol. 50, 1841–1846. doi: 10.1128/jcm.06848-11
Wagman, J., and Weneck, E. J. (1963). Preservation of bacteria by circulating-gas freeze drying. Appl. Microbiol. 11, 244–248. doi: 10.1128/aem.11.3.244-248.1963
Wein, T., Hülter, N. F., Mizrahi, I., and Dagan, T. (2019). Emergence of plasmid stability under non-selective conditions maintains antibiotic resistance. Nat. Commun. 10, 2595–2595.
Wilkinson, K. M., Winstanley, T. G., Lanyon, C., Cummings, S. P., Raza, M. W., and Perry, J. D. (2012). Comparison of four chromogenic culture media for carbapenemase-producing Enterobacteriaceae. J. Clin. Microbiol. 50, 3102–3104. doi: 10.1128/jcm.01613-12
Woodford, N., Wareham, D. W., Guerra, B., and Teale, C. (2014). Carbapenemase-producing Enterobacteriaceae and non-Enterobacteriaceae from animals and the environment: an emerging public health risk of our own making? J. Antimicrob. Chemother. 69, 287–291. doi: 10.1093/jac/dkt392
Zhanel, G., Wiebe, R., Dilay, L., Thomson, K., Rubinstein, E., Hoban, D. J., et al. (2007). Comparative review of the carbapenems. Drugs 67, 1027–1052. doi: 10.2165/00003495-200767070-00006
Zurfluh, K., Hindermann, D., Nuesch-Inderbinen, M., Poirel, L., Nordmann, P., and Stephan, R. (2016). Occurrence and features of chromosomally encoded carbapenemases in Gram-negative bacteria in farm animals sampled at slaughterhouse level. Schweiz. Arch. Tierheilkd. 158, 457–460. doi: 10.17236/sat00072
Keywords: carbapenemase, isolation, media, specificity, sensitivity
Citation: Pauly N, Hammerl JA, Grobbel M, Tenhagen B-A, Käsbohrer A, Bisenius S, Fuchs J, Horlacher S, Lingstädt H, Mauermann U, Mitro S, Müller M, Rohrmann S, Schiffmann AP, Stührenberg B, Zimmermann P, Schwarz S, Meemken D and Irrgang A (2020) ChromID® CARBA Agar Fails to Detect Carbapenem-Resistant Enterobacteriaceae With Slightly Reduced Susceptibility to Carbapenems. Front. Microbiol. 11:1678. doi: 10.3389/fmicb.2020.01678
Received: 30 March 2020; Accepted: 26 June 2020;
Published: 11 August 2020.
Edited by:
Shaolin Wang, China Agricultural University, ChinaCopyright © 2020 Pauly, Hammerl, Grobbel, Tenhagen, Käsbohrer, Bisenius, Fuchs, Horlacher, Lingstädt, Mauermann, Mitro, Müller, Rohrmann, Schiffmann, Stührenberg, Zimmermann, Schwarz, Meemken and Irrgang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Natalie Pauly, bmF0YWxpZS5wYXVseUBiZnIuYnVuZC5kZQ==; Alexandra Irrgang, YWxleGFuZHJhLmlycmdhbmdAYmZyLmJ1bmQuZGU=; YWxleC5pcnJnYW5nQGdtYWlsLmNvbQ==