 Germán A. Kopprio1,2,3*
Germán A. Kopprio1,2,3* Sucharit B. Neogi4
Sucharit B. Neogi4 Harunur Rashid5
Harunur Rashid5 Cecilia Alonso6
Cecilia Alonso6 Shinji Yamasaki4
Shinji Yamasaki4 Boris P. Koch7
Boris P. Koch7 Astrid Gärdes2
Astrid Gärdes2 Rubén J. Lara3
Rubén J. Lara3- 1Department of Chemical Analytics and Biogeochemistry, Leibniz-Institute of Freshwater Ecology and Inland Fisheries, Berlin, Germany
- 2Tropical Marine Microbiology, Leibniz Centre for Tropical Marine Research, Bremen, Germany
- 3Marine Biogeochemistry, Instituto Argentino de Oceanografía, Consejo Nacional de Investigaciones Científicas y Técnicas – Universidad Nacional del Sur, Bahía Blanca, Argentina
- 4Graduate School of Life and Environmental Sciences, Osaka Prefecture University, Izumisano, Japan
- 5Department of Fisheries Management, Bangladesh Agricultural University, Mymensingh, Bangladesh
- 6Microbial Ecology of Aquatic Systems, Centro Universitario Región Este, Universidad de la República, Rocha, Uruguay
- 7Ecological Chemistry, Alfred Wegener Institute, Helmholtz Centre for Polar and Marine Research, Bremerhaven, Germany
The highly populated coasts of the Bay of Bengal are particularly vulnerable to water-borne diseases, pollution and climatic extremes. The environmental factors behind bacterial community composition and Vibrio distribution were investigated in an estuarine system of a cholera-endemic region in the coastline of Bangladesh. Higher temperatures and sewage pollution were important drivers of the abundance of toxigenic Vibrio cholerae. A closer relation between non-culturable Vibrio and particulate organic matter (POM) was inferred during the post-monsoon. The distribution of operational taxonomic units (OTUs) of Vibrio genus was likely driven by salinity and temperature. The resuspension of sediments increased Vibrio abundance and organic nutrient concentrations. The δ13C dynamic in POM followed an increasing gradient from freshwater to marine stations; nevertheless, it was not a marker of sewage pollution. Bacteroidales and culturable coliforms were reliable indicators of untreated wastewater during pre and post-monsoon seasons. The presumptive incorporation of depleted-ammonium derived from ammonification processes under the hypoxic conditions, by some microorganisms such as Cloacibacterium and particularly by Arcobacter nearby the sewage discharge, contributed to the drastic 15N depletion in the POM. The likely capacity of extracellular polymeric substances production of these taxa may facilitate the colonization of POM from anthropogenic origin and may signify important properties for wastewater bioremediation. Genera of potential pathogens other than Vibrio associated with sewage pollution were Acinetobacter, Aeromonas, Arcobacter, and Bergeyella. The changing environmental conditions of the estuary favored the abundance of early colonizers and the island biogeography theory explained the distribution of some bacterial groups. This multidisciplinary study evidenced clearly the eutrophic conditions of the Karnaphuli estuary and assessed comprehensively its current bacterial baseline and potential risks. The prevailing conditions together with human overpopulation and frequent natural disasters, transform the region in one of the most vulnerable to climate change. Adaptive management strategies are urgently needed to enhance ecosystem health.
Introduction
The genus Vibrio comprises a diverse group of more than 140 species, within which V. cholerae, V. parahaemolyticus, V. vulnificus, V. fluvialis, and V. mimicus are etiological agents of human and animal disease causing gastroenteritis, sepsis and necrosis. Toxigenic and non-toxigenic populations of Vibrio species coexist generally in the aquatic environment. Vibrio cholerae is a natural inhabitant of estuaries and some strains are responsible of cholera disease. Cholera gravis cases are characterized by profuse watery diarrhea which leads to a life-threatening dehydration (Sack et al., 2004). Recent outbreaks in developing countries, particularly after natural disasters, classified cholera as a re-emerging disease (Morens et al., 2004; Vouga and Greub, 2016). Notably, the low-lying coastal areas of the Bay of Bengal are not only claimed as an endemic region for the pandemic serogroups of V. cholerae but also as the pivotal base of the global spread of cholera (Mutreja et al., 2011).
Cholera toxin and toxin co-regulated pilus are the main virulence factors of the pandemic serogroups of V. cholerae: O1 and O139. In endemic areas, the incidence of cholera in human populations is coupled with the distribution of V. cholerae in aquatic ecosystems. Temperature, salinity and plankton abundance are important factors explaining the distribution of V. cholerae (Huq et al., 2005; Constantin de Magny et al., 2008). Under adverse environmental conditions, V. cholerae is usually detected in a non-culturable state of reduced metabolism but with the potential to grow exponentially under favorable settings. In the last years, organic matter and suspended sediments were identified as key drivers for the distribution of Vibrio species (Lara et al., 2009; Neogi et al., 2018). Since only a few studies offer scattered information about these drivers, more investigations are needed for an integral assessment of the biogeochemical factors behind Vibrio ecology.
Vibrio species tend to form biofilms in seston and present a wide variety of enzymes to degrade and assimilate organic matter playing unique roles in biogeochemical cycles (Lara et al., 2011; Neogi et al., 2018; Liang et al., 2019). Moreover, Vibrio populations interact with other microorganisms during the process of nutrient remineralization, which is performed by a variety of microbial taxa with different metabolic and physiological capabilities. The precise way in which organic and inorganic nutrients influence dynamics of aquatic bacterial populations remains largely unresolved (Bunse and Pinhassi, 2017). Biological interactions are emerging as a key factor driving bacterial communities and many virulence features of V. cholerae evolved not only in response to environmental factors but also to biotic pressures (Schwartz et al., 2019). Studies combining the ecology of V. cholerae and other Vibrio populations in relation with bacterial community composition are urgently needed in cholera endemic areas.
The coastline of the Bengal delta is largely influenced by monsoon rainfalls, recurrent climatic disasters and pollutants. Among the diverse groups of bacteria existing in coastal waters, some can respond quickly to changing environmental conditions and are reliable indicators of hydrological changes and water pollution (Cao et al., 2017; Kegler et al., 2017). Sewage impacts on human and ecosystem health by eutrophication, spread of water-borne diseases, and as a source of heavy metals and persistent organic pollutants (reviewed by Schwarzenbach et al., 2010). Stable isotopes of nitrogen and carbon are other indicators of pollution and the origin of the organic matter. An elevated signature of δ15N in the suspended particulate matter is generally a marker of anthropogenic impacts; however, recent studies evidenced the marked depletion of 15N in heavily polluted systems (Ke et al., 2017; Kopprio et al., 2018; Hong et al., 2019). The combination of bacterial and isotopic indicators offers a tool of higher resolution in ecological studies of coastal systems.
Little work has been performed to understand the effect of the composition and distribution of organic matter influencing the population dynamics of Vibrio species along with the changes in coexisting estuarine bacterial communities in cholera endemic regions. Therefore, this study systematically explored a polluted estuary in Bangladesh with the following aims: (1) to elucidate the role of water quality and nutrient biogeochemistry on the dynamic of Vibrio species and culturable bacteria; (2) to study the seasonal distribution of bacterial communities across the estuarine and pollution gradient as well as their influence on the isotopic signature of the particulate organic matter; and (3) to identify potential pathogens and key bacteria for biogeochemical processes and bioremediation. We hypothesized a strong influence of sewage and sediment resuspension in the distribution of toxigenic V. cholerae, Vibrio species and genera of potential pathogens. Other global hypothesis was that the specific effects of sewage input and sediment resuspension on aquatic bacterial community and organic matter composition are clearly discernible. The integrated assessment of hydrological changes and pollution on the dynamic of biogeochemical and bacterial markers influencing ecosystem health enhances the multidisciplinary scope of this study.
Materials and Methods
Study Site
The Karnaphuli estuary is characterized by semidiurnal tides and flows across the southern banks of the Chittagong port city into the east coastline of the Bay of Bengal (Figure 1). In the last decades, Chittagong metropolitan area has experienced a considerable population growth and nowadays comprises of more than 4 million inhabitants. As it occurs in most of the coastal cities of Bangladesh, Chittagong has insufficient water treatment facilities and the effluents from diverse sources are discharged practically raw in the estuary endangering ecosystem and public health (Wang et al., 2016). The lack of safe drinking water and sanitation, and the prevalence of water-borne diseases are key challenges for Chittagong’s authorities and stakeholders (Lara et al., 2009). In the last decades the region experienced strong cyclones, which killed thousands of people and produced severe floods.
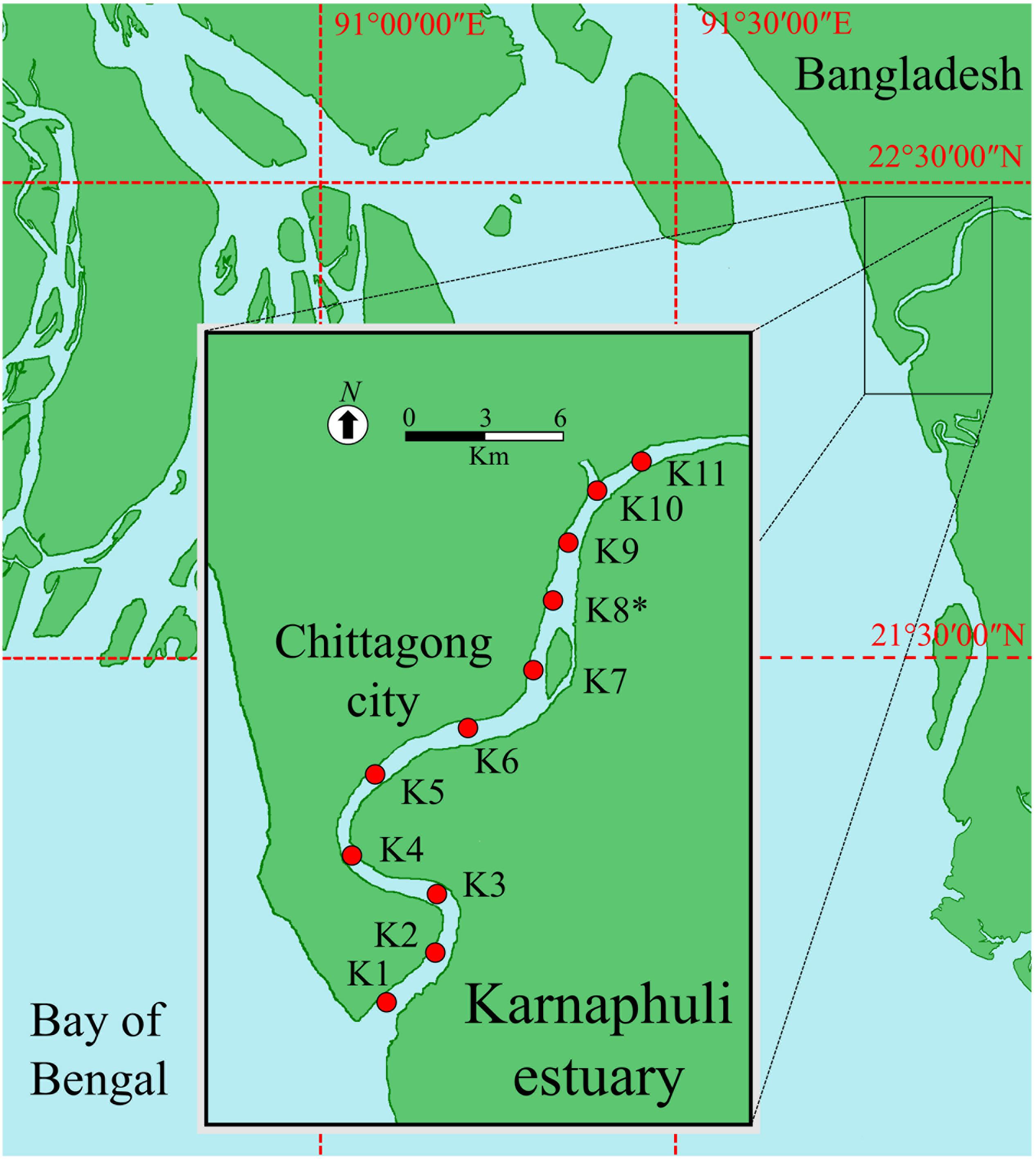
Figure 1. Location of the Karnaphuli estuary and sampling stations in the Bay of Bengal. (*) strongly influenced by sewage pollution.
Sampling Strategy
The Karnaphuli estuary was sampled at 11 stations (Figure 1) during pre and post-monsoon seasons (May 15th and December 8th of 2016, respectively). The pre-monsoon campaign was carried out 3 days before the cyclone Roanu, which forced half a million people to leave their homes and caused some fatalities. Water and Suspended Particulate Matter (SPM) were sampled with a motor boat across a ∼20 Km transect, starting at station 1 (K1) with marine water and finishing at K11 with freshwater. The station with a strong influence of the sewage discharge of Chittagong city corresponded to K8. To separate larger aggregates and organisms from nanoplankton, water was fractionated with a net (20 μm pore size) at K1, K3, K5, K7, and K11. To simulate the likely effect of water runoff, coastal sediments were resuspended in a known volume of estuarine water (∼2 g L–1) at K3, K5, K8 and K11.
In situ Measurements and Preparations
Temperature, salinity, conductivity, pH and dissolved oxygen were measured in situ with a multiparametric meter WTW 3430 (Xylem Analytics). Water and SPM were collected at 30 cm below the surface and stored in clean and sterile 2 L plastic bottles. The nanoplankton fraction (SPM < 20 μm) and SPM enriched with coastal sediments (SPM-Sed) were sampled at the same depth and stored in bottles with the mentioned characteristics. Water for dissolved nutrients was filtered with a Syringe filter Minisart PES High Flow (Sartorius) and stored in chemically clean HDPE 50 mL bottles. For Catalyzed Reporter Deposition – Fluorescence In Situ Hybridization (CARD-FISH) studies, water samples were fixed with buffered particle-free paraformaldehyde (1% final concentration) and kept in PE 50 mL bottles. All samples were transported in insulated plastic boxes and processed within 6 h.
Biogeochemical Analyses
Bottles for dissolved nutrients were frozen at −20°C until further analyses. All samples were filtered through precombusted GF/F glass-fiber filters (Whatman). Filters for chlorophyll determinations were preserved frozen at −20°C and those for particulate organic matter (POM) and stable isotope measurements were dried overnight at 50°C and stored in a vacuum desiccator. Dissolved inorganic nutrients were determined according to standard methods with an auto-analyzer (Evolution III, Alliance Instrument). Dissolved organic carbon (DOC) was measured by high temperature catalytic oxidation with a Shimadzu TOC-VCPN analyzer and total dissolved nitrogen (TDN) was measured simultaneously by chemiluminescence detection with a Shimadzu TNM-1. Dissolved organic nitrogen (DON) was calculated by difference as DON = TDN – (ammonium + nitrate + nitrite). Pigments were extracted in ethanol ≥99.5% (Roth) and an overnight incubation at 4°C in the darkness. Chlorophyll a content was estimated photometrically after Marker et al. (1980).
All dried filters were acidified (1N HCl) to remove inorganic carbon and placed in tin or silver capsules for particulate organic nitrogen and carbon (PON and POC) determination, respectively. The capsules were oxidized at 1000°C under pure oxygen in an elemental analyzer (EURO EA, HEKAtech). Carbon and nitrogen stable isotopes (13C and 15N) were measured with a mass spectrometer (Thermo Finnigan Delta Plus, Thermo Fisher Scientific) coupled with an elemental analyzer (Flash EA 1112, Thermo Fisher Scientific). Acetanilide (HEKAtech) and peptone were used as internal standards. Stable isotopes were reported as delta (δ) values in parts per thousand (denoted as ‰), carbon relative to Pee Dee Belemnite and nitrogen relative to nitrogen in air. The isotope ratios were measured in accordance with the following reference standards: IAEA-N-1, IAEA-N-2, NBS 22, and USGS-24.
Culturable Bacteria
Water was directly spread or placed after filtration through sterile mixed cellulose membranes of 0.45 μm pore size (ME 25, Whatman) on Thiosulfate Citrate Bile salts Sucrose (TCBS, Roth) agar plates. Colony Forming Units (CFUs) of presumptive Vibrio were counted after overnight incubation at 37°C. Water (25 mL) was enriched in Alkaline Peptone Water (APW, Roth) and incubated overnight at the same temperature. To estimate the colonies of V. cholerae growing in TCBS, 30 presumptive colonies were isolated and enriched in APW. Water was also spread in duplicate directly or after serial dilutions in 1× phosphate buffered saline (PBS) on Mac Conkey agar (Roth) for the determination of coliforms and on Plate Counting Agar (PCA, Roth) for terrestrial heterotrophic bacteria. CFUs were counted after overnight incubation at 37°C and 30°C, respectively.
Card-Fish
Samples for CARD-FISH were kept overnight at 4°C and known volumes (10–50 mL) were filtered through GTTP Isopore membranes (Merck Millipore) of 0.2 μm pore size, and stored in sterile Petri dishes at −20°C. The protocol for CARD-FISH was performed basically after Pernthaler et al. (2002). Briefly, cells were permeabilized with lysozyme (Sigma Aldrich) solution (10 mg mL–1) and endogenous peroxidases were inactivated with HCl (0.01 N). The Horseradish Peroxidase (HRP) – labeled probes (Biomers.net) used for hybridization were: Vchomim1276 for Vibrio cholerae and V. mimicus and Vibrio-GV for V. parahaemolyticus, V. vulnificus and several other Vibrio species excluding V. cholerae (Table 1). The HRP-probes (50 ng μL–1) were mixed in a relation 1:100 with the hybridization buffer containing 50% formamide for Vchomim1276 and 30% for Vibrio-GV. After an incubation of 2 h at 35°C, samples were rinsed in the corresponding washing buffer and subsequently in 1× PBS. The CARD step was performed in the darkness for 15 min at 37°C, with a tyramide-containing amplification buffer (1 μg μL–1 tyramide conjugated with Alexa 488, Thermo Fisher Scientific). Filters were counterstained with DAPI (1 μg mL–1) and cells were counted across 50 random grids (120 × 120 μm). The abundance (cells mL–1) was calculated considering the total surface and volume filtered.

Table 1. Primers and probes used for catalysed reporter deposition – fluorescence in situ hybridization (CARD-FISH), quantitative polymerase chain reaction (qPCR) and 16S rRNA studies.
DNA Extraction
Water (200–100 mL) was filtered through the mentioned GTTP membranes (Merck Millipore), stored at −20°C and the DNA was extracted after Boström et al. (2004). Briefly, cells were lysed firstly with lysozyme (1 mg mL–1) and then treated with SDS (1%) and proteinase K (Thermo Fisher Scientific, 100 μg mL–1). DNA was recovered after incubation with 0.6 volumes of isopropanol (ROTIPURAN, Roth) and two centrifugation steps for 20 min at 20,000 g and 4°C. In case of inhibition, the extracts were diluted or cleaned with a genomic DNA clean-up kit (NucleoSpin, Macherey-Nagel). For culturable bacteria, DNA was extracted using a simple boiling method. DNA concentrations were measured in a NanoQuant plate with an infinite M200 PRO Multimode reader (TECAN).
Quantitative Polymerase Chain Reaction (qPCR)
The detection of V. cholerae was based on the qPCR protocol of Bliem et al. (2015). The protocol was modified in two duplex strategies in our Bio-Rad CFX Connect real time system: (1) the ompW probe (Table 1) was labeled with 6-carboxyfluorescein (FAM, Biomers.net) and the internal control (IC) with 6-hexachloro-fluorescein phosphoramidite (HEX, Biomers.net); and (2) ctxA was labeled with HEX and IC with FAM. For ompW detection, all qPCR were performed in a final reaction volume of 20 μL containing 1× SensiFAST Probe Lo-ROX master mix (Bioline), ompW-F1 (150 nM), ompW-F2 (50 nM), ompW-R (200 nM), ompW-FAM (200 nM), IC-F (100 nM), IC-R (100 nM), IC-HEX (25 nM), and sample and IC templates. For ctxA quantification, the primers were: ctxA-F (200 nM), ctxA-R (200 nM), ctxA-HEX (200 nM), IC-F (100 nM), IC-R (100 nM), IC-FAM (50 nM). The qPCR conditions were 1 min of denaturation at 95°C followed by 45 cycles at 95°C for 10 s and at 60°C for 30 s. Each run contained a dilution series of DNA from V. cholerae O1 N 16961 and negative controls. The qPCR reactions were managed and analyzed using the software Bio-Rad CFX Manager 3.0. The genomic units (GU) of V. cholerae were calculated based on the genome length and the molar mass of V. cholerae O1 N 16961, DNA concentrations and total filtered volume.
16S rRNA Diversity
The V3-V4 hypervariable region of the 16S rRNA gene was amplified by the primers Bact-341F and Bact-785R (Table 1) and sequenced by the company LGC Genomics on an Illumina MiSeq platform. After primer removal and demultiplexing, sequences were trimmed using trimmomatic v0.36 (Bolger et al., 2014) and merged with PEAR v0.9.8 (Zhang et al., 2014). Quality trimmed sequences were 54,917 ± 25,626 (mean ± SD) per sample. Clustering of Operational Taxonomic Units (OTUs) was performed with Minimum Entropy Decomposition MED v2.1 (Eren et al., 2015) and the OTU number per sample was 20,676 ± 12,550. For taxonomic classification, OTU representatives were submitted to SilvaNGS (v1321) using a threshold of sequence similarity of one for clustering (Quast et al., 2013). Singletons, doubletons, archaea, chloroplasts, mitochondria were removed from the analysis. Demultiplexed and primer-clipped sequences were deposited at the European Nucleotide Archive using the data brokerage service of the German Federation for Biological Data (Diepenbroek et al., 2014) with the accession number PRJEB35775.
Statistical Analysis
Removal of OTUs with poor alignment quality, pooling of taxa and calculations of abundance and diversity were performed in R v3.5.1 (R Core Team, 2018) and additional packages. Statistics and graphics were carried out with R, XLSTAT (Addinsoft, 2018), PRIMER v6 and Xact 7.21d. Permutational multivariate analysis of variance (PERMANOVA) was used to evaluate differences between seasons and fractions in culturable bacteria and Vibrio counts and in main orders, genera and OTUs. In case of significant differences among fractions, pair-wise tests were performed and the variables were compared with Kruskal-Wallis one-way analysis of variance (ANOVA) and Dunn pair-wise comparisons. Two Canonical Correspondence Analyses (CCA 1 and 2) were carried out using the environmental parameters as explanatory variables of the distribution of: CCA 1) Culturable bacteria and Vibrio abundance (counts or genomic units L–1); and CCA 2) OTU numbers for the main bacterial orders (total n at each order). For the CCAs, the environmental and the biological variables were logarithmical [log (x + 1)] and square-root transformed, respectively. Linearity was checked with the Monte Carlo test after 1000 permutations. Relationships between and within biological and environmental variables were also studied using Pearson correlations (r) based on the log-transformed data. The 70 most abundant OTUs of those genera with a sequence relative abundance higher than 1% at any of the K8 samples were displayed in a heat map, K3 and K11 were included as “control” sites.
Results
Distribution and Seasonality of Vibrio Across the Estuarine Continuum
The ompW gene for V. cholerae was positive in all water samples directly enriched with APW and in the 83% of the non-cultured samples. The ctxA gene followed the opposite trend, with 25% of the positive cases in the culturable fraction and 38% in the non-enriched cases. The samples derived from the presumptive colonies in TCBS were positive for ompW in the 23% of the cases but negative for ctxA. The CFUs in TCBS or culturable Vibrio counts (CVCs) were significantly positively correlated with Vibrio-GV counts (r = 0.70, p < 0.001) and Vibrio OTUs (r = 0.55, p < 0.001). A total of 74 different OTUs for Vibrio were detected.
The highest number of ctxA GU in the SPM was found at the station receiving the sewage discharge (K8) during the warmer season (Figure 2). Other peaks of ctxA GU were associated to downstream stations of intermediate salinities. Ammonium, closely related to coliforms, and POC were positively correlated with ctxA GU (Table 2). Vchomim1276 counts in SPM presented maximum values at K8 during both seasons (Figure 2) and generally higher values during the post-monsoon. POC showed significant correlations with Vchomim1276 counts and ompW GU (Table 2). Salinity as an indicator of the estuarine gradient was positively correlated principally with Vibrio OTUs, CVCs and Vibrio-GV. Water temperature as an indicator of seasonality was significantly positively correlated with heterotrophic culturable bacteria and CVCs.
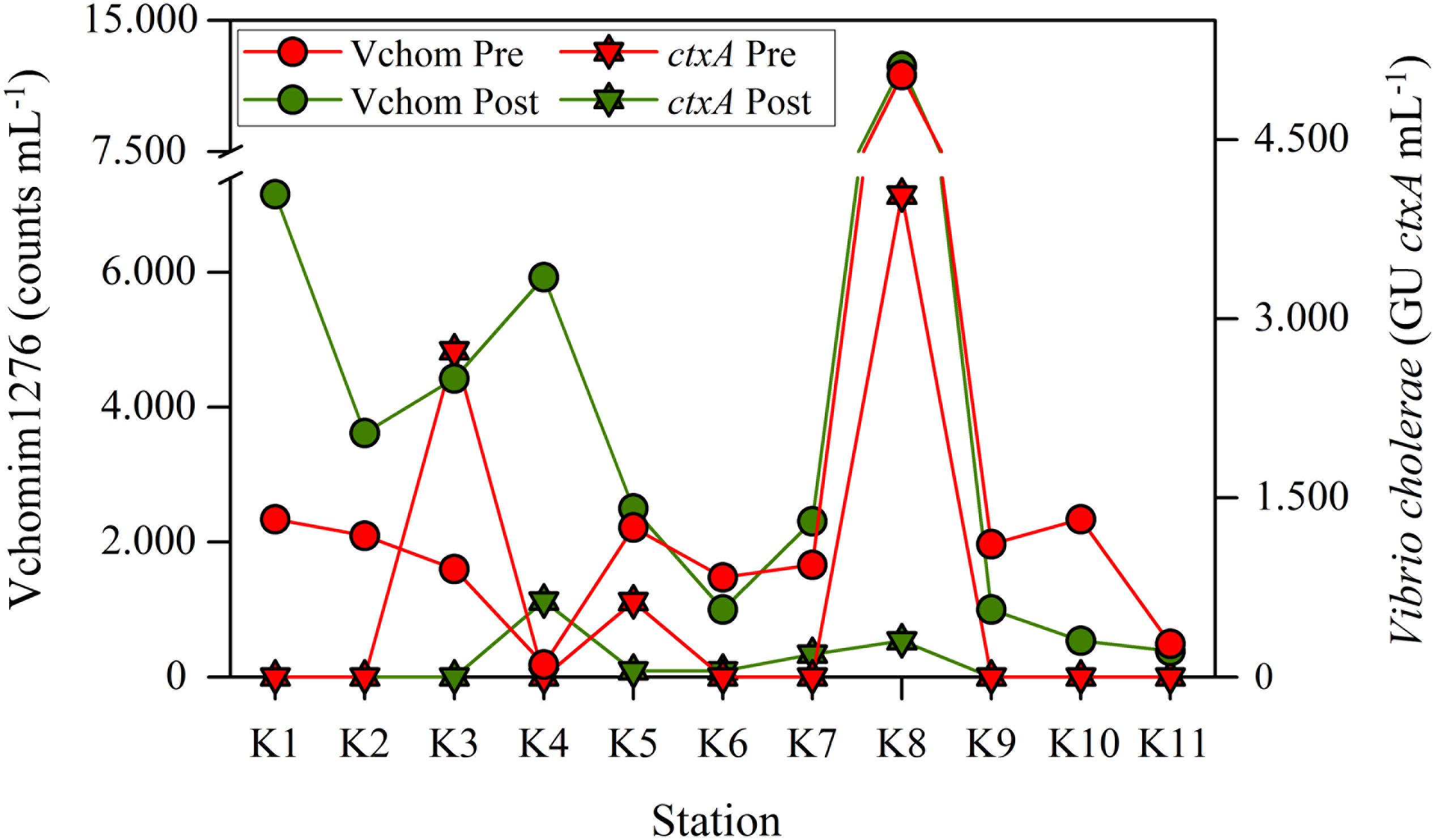
Figure 2. Seasonal abundances of Vibrio cholerae and V. mimicus based on CARD-FISH counts with the probe Vchomim1276 and genomic units (GU) of cholera toxin gene A (ctxA) across the estuarine gradient. Station K8 was strongly influenced by sewage pollution.
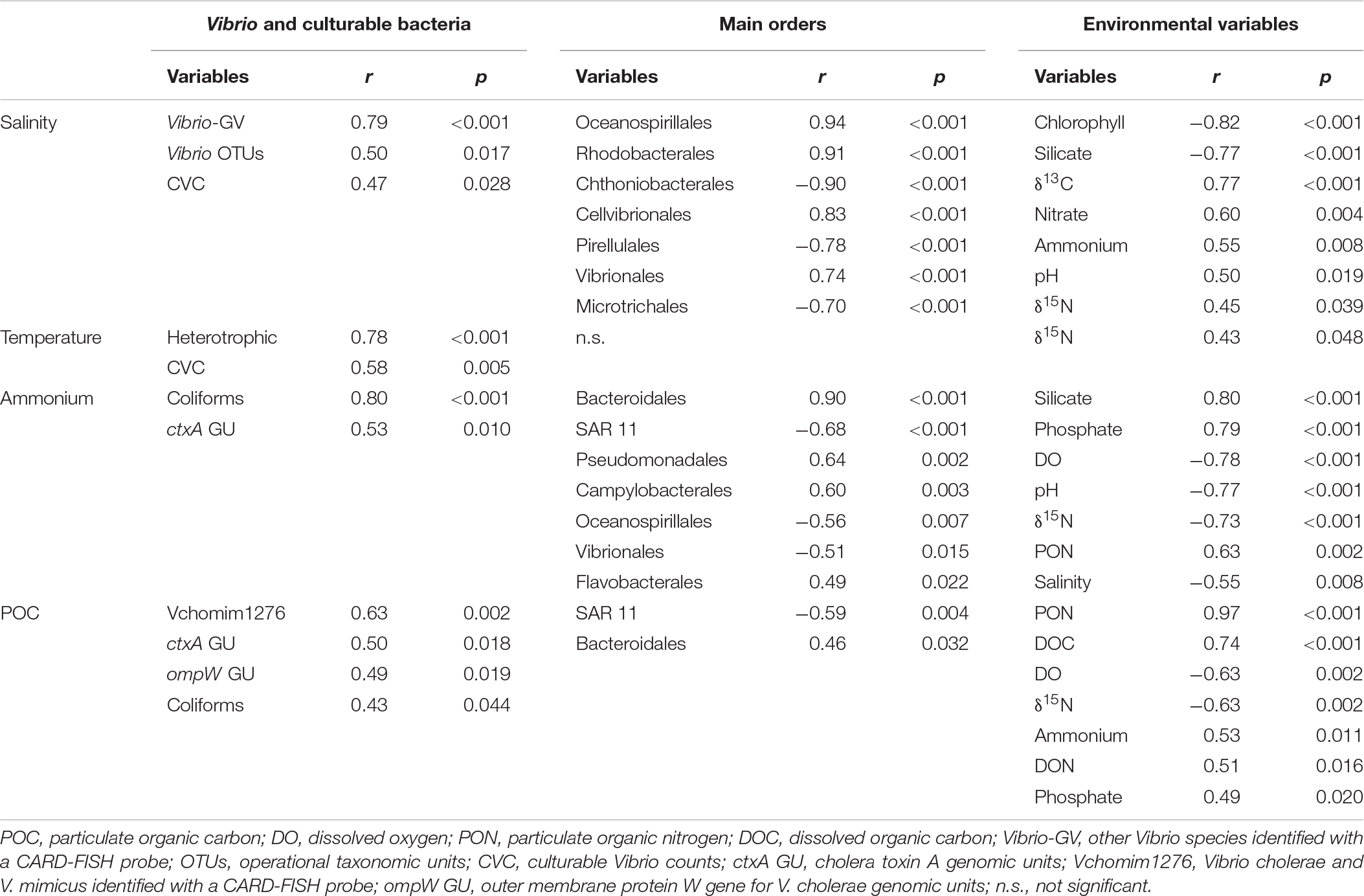
Table 2. Main correlations (Pearson coefficient = r) of salinity, temperature, ammonium and particulate organic carbon (POC) with biological and other environmental variables in the suspended particulate matter (n = 22).
PERMANOVA revealed significant differences between seasons (pseudo-F = 7.28, p < 0.001) and fractions (pseudo-F = 3.52, p = 0.013). Fractions refer to SPM, SPM < 20 μm and SPM-Sed. No interaction was detected between seasons and fractions (pseudo-F = 0.95, p = 0.954). According to pair-wise tests and considering the differences between fractions, SPM-Sed differed significantly from SPM (t = 2.20, p = 0.013) and SPM < 20 μm (t = 2.41, p = 0.006). No significant differences were found between SPM and SPM < 20 μm (t = 0.91, p = 0.451). SPM-Sed (Table 3) shows significantly higher values of Vchomim1276 and Vibrio-GV counts, ompW GU, DON, POC and PON. No significance differences in any parameter were found between SPM and SPM < 20 μm.
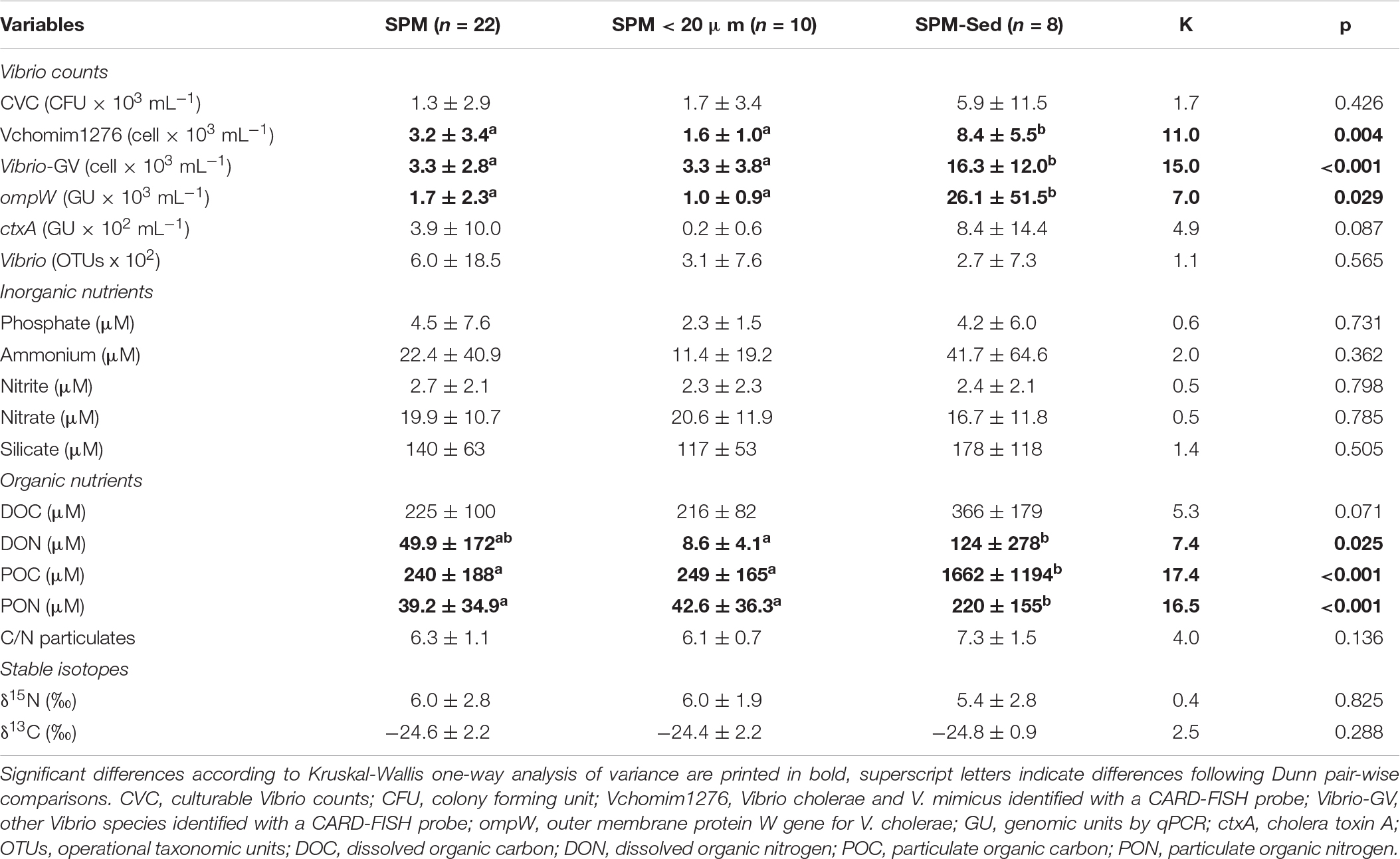
Table 3. Distribution of Vibrio counts, nutrients and stable isotopes (mean value ± standard deviation) in the fractions: suspended particulate matter (SPM), nanoplankton (SPM < 20 μm), and SPM enriched with sediments (SPM-Sed).
The CCA (Figure 3) ordinated samples and culturable bacteria and Vibrio based on the environmental parameters and suggested also seasonal differences. The constrained inertia was 71.6% of the total inertia and the relation among the variables were considered linear (pseudo-F = 1.61, p < 0.001). All pre-monsoon samples were located at the negative side of the CC1 (which explains the 45% of the variation) together with culturable Vibrio, heterotrophic bacteria and Vibrio OTUs. These variables were linked mainly to temperature, δ15N and nitrate. At the positive side of CC1 were ordinated all post-monsoon samples together with coliforms and Vibrio counts (qPCR and CARD-FISH). Organic matter, ammonium and chlorophyll were also located at the positive side of CC1. At the positive extreme of CC2 (which explains the 32% of the variation) were ordinated the samples of K8 with higher ammonium and DON values and some freshwater samples with elevated chlorophyll values. The samples more influenced by the sea were at the positive extreme of CC2.
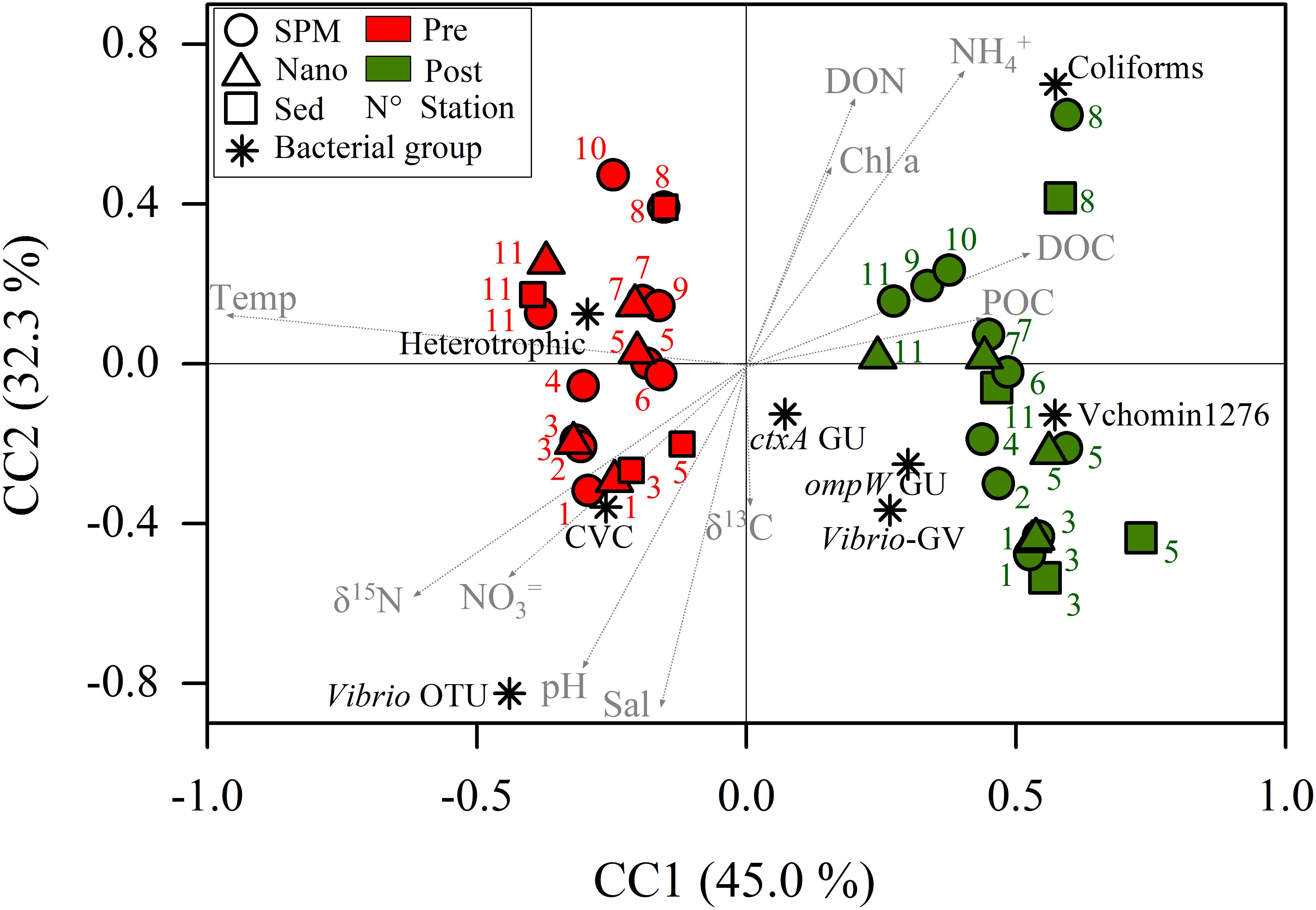
Figure 3. Canonical correspondence analysis (CCA) using environmental parameters as explanatory variables of culturable bacteria and Vibrio counts. SPM, suspended particulate matter; Nano, SPM after 20 μm net filtration; Sed, SPM enriched with sediments; Pre, Pre-monsoon; Post, Post-monsoon; Temp, temperature; Sal, salinity; DON, dissolved organic nitrogen; DOC, dissolved organic carbon; Chl a, chlorophyll a; POC, particulate organic carbon; CVC, culturable Vibrio counts; OTU, operational taxonomic units; GU, genomic units by qPCR; ctxA, cholera toxin A; ompW, outer membrane protein W gene for Vibrio cholerae; Vchomim1276, CARD-FISH counts for V. cholerae and V. mimicus; Vibrio-GV; CARD-FISH counts for other Vibrio species.
Bacterial Communities: Estuarine Gradient and Pollution Impact
The orders Rhodobacterales and Oceanospirillales presented higher relative sequence abundances at the marine stations (Supplementary Figure S1). The main genera in the Rhodobacterales were Marivivens (63% of the total OTUs for the order Rhodobacterales) and Roseobacter clade CHAB-I-5 lineage (11%), while in the Oceanospirillales were Bermanella (22%), Oleiphilus (17%), Halomonas (9%), and Marinomonas (7%). The relative sequence abundances of Vibrionales (96% Vibrio) were also higher at the marine stations and particularly during the Pre-monsoon season (up to 20%, Supplementary Figure S1). Salinity as an indicator of the estuarine gradient was positively correlated with Oceanospirillales, Rhodobacterales, Cellvibrionales and Vibrionales, while negatively mainly with Chthoniobacterales, Pirellulales and Microtrichales (Table 2). The Microtrichales (96% CL500-29 marine group) and Synechococcales (100% Cyanobium PCC-6307) were the dominant orders at the freshwater stations.
Other abundant orders at intermediate stations (Supplementary Figure S1) were the Betaproteobacteriales (45% Burkholderiaceae unclassified and 5% Dechloromonas) and Micrococcales (70% Candidatus Aquiluna and 5% Candidatus Planktoluna). The Bacillales (85% Bacillus) dominated the samples enriched with sediments at some stations during the post-monsoon. Ammonium was significantly correlated with Bacteroidales, Pseudomonadales, Campylobacterales and Flavobacteriales (Table 2). Moreover, POC was positively correlated with Bacteroidales and coliforms. The principal genera in the order Bacteroidales were Prevotella 9 (50%) and Macellibacteroides (16%), in Pseudomonadales were Acinetobacter (76%) and Pseudomonas (23%), in Campylobacterales were Arcobacter (93%) and Sulfurospirillum (6%), and in Flavobacteriales were Cloacibacterium (35%) and Flavobacterium (28%).
PERMANOVA indicated no significant differences between seasons (pseudo-F = 1.16, p = 0.289) and fractions (pseudo-F = 0.994, p = 0.407) at order level. At genera level, PERMANOVA did not evidence significant differences between seasons (pseudo-F = 1.95, p = 0.103) and fractions (pseudo-F = 0.84, p = 0.530). Following the same trend, no significant differences were found between seasons (pseudo-F = 2.1, p = 0.054) and fractions (pseudo-F = 0.68, p = 0.800) at OTU level. The non-seasonality in the distribution of the main orders was also indicated by the CCA ordination (Figure 4). The constrained inertia was the 72.8% of the total inertia and the relations between the data sets were considered linear (pseudo-F = 4.39, p < 0.001). The CC1 (which explained the 41.8% of the variation) ordinated at its negative side Vibrionales, Oceanospirillales, Cellvibrionales and Rhodobacterales. These orders were grouped principally together with salinity, δ13C and nitrate. The marine stations (K1, K2 and K3) were located at the negative extreme of CC1. At the positive side were ordinated most of the freshwater stations together with the Chthoniobacterales, Pirellulales, Synechococcales, Microtrichales and Frankiales. These variables were principally associated with chlorophyll a. The positive side of CC2 grouped the samples from K8 and K7, principally together with Bacteroidales, Campylobacterales, Flavobacteriales, ammonium and organic matter.
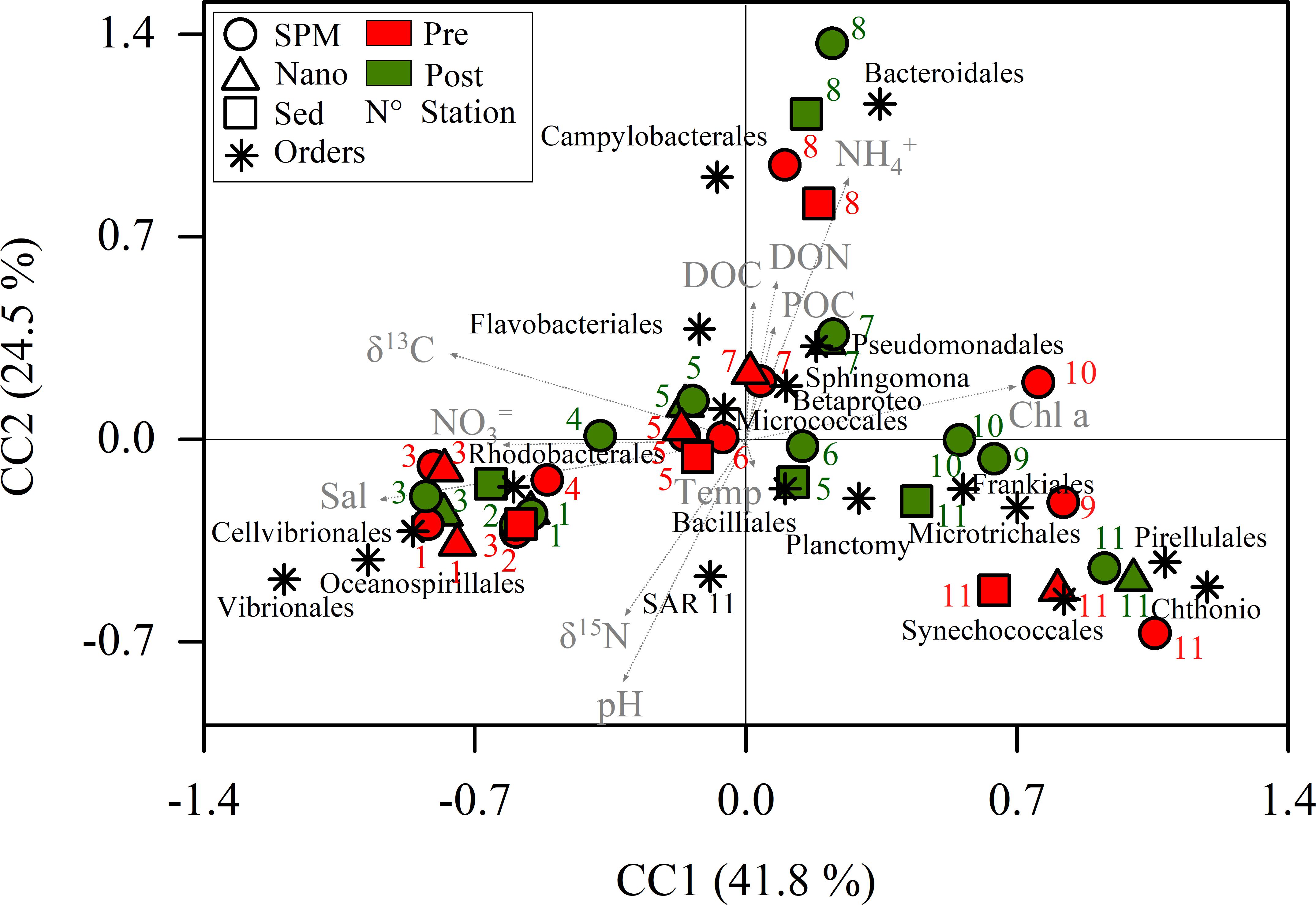
Figure 4. Canonical correspondence analysis (CCA) using environmental parameters as explanatory variables of the main orders. SPM, suspended particulate matter; Nano, SPM after 20 μm net filtration; Sed, SPM enriched with sediments; Pre, Pre-monsoon; Post, Post-monsoon; Temp, temperature; Sal, salinity; DON, dissolved organic nitrogen; DOC, dissolved organic carbon; Chl a: chlorophyll a; POC, particulate organic carbon; Betaproteo, Betaproteobacteriales; Sphingomonas, Sphingomonadales; Planctomy, Planctomycetales; Chthonio, Chthoniobacterales.
Depleted values of δ15N in PON were evidenced at K8 in both seasons and even negative values during the post-monsoon (Figure 5). The maxima of ammonium, DON and POC, the second lowest concentration of dissolved oxygen, poorer values of the inverse Simpson Index (similar to K1), the lowest concentration of nitrate, and one of the minimum concentrations of nitrite were observed at the sewage during the post-monsoon. The trend in the mentioned parameters was similar during the pre-monsoon. The δ13C was not influenced by the sewage and its trend decreases from marine to freshwater stations. The main relations between salinity, temperature, ammonium and POC with other environmental parameters are summarized on Table 2. For example, salinity was positively significantly correlated with δ13C and nitrate, ammonium with phosphate and PON, and POC with PON and DOC. It is worth to mention the strong negative correlation between ammonium and δ15N. The influence of the sewage pollution was stronger in the water quality parameters during the post-monsoon and coincided with highest relative abundance of Arcobacter (Figure 5). Cloacibacterium presented the maximum of relative sequence abundance at K8 during the pre-monsoon. The Bacteroidales presented abundance peaks during both seasons. The 70 OTUs of the heatmap (Figure 6) represented more than the 50% of the total sequence abundance at K8. The OTUs found at both seasons were those of the genera: Paludibacter, Dechloromonas, C39 (Rhodocyclaceae), Prevotellaceae unclassified, Prevotella 9, Macellibacteroides, Acinetobacter, Tolumonas, and Streptococcus.
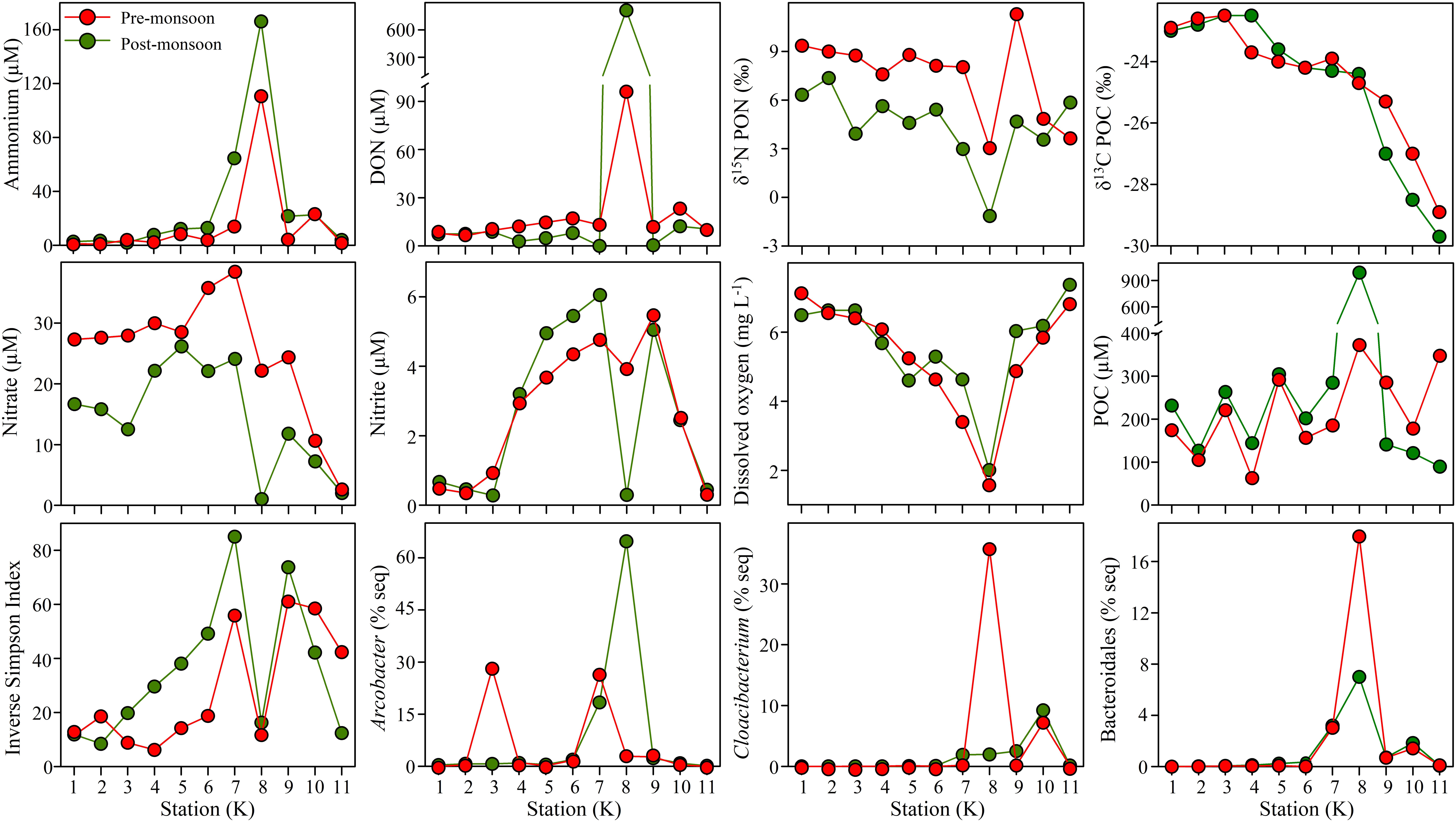
Figure 5. Dynamic of dissolved inorganic and organic nitrogen, particulate organic carbon, stable isotopes in the particulates, dissolved oxygen concentration, diversity, main genera at the sewage and order Bacteroidales. Station K8 was strongly influenced by sewage pollution.

Figure 6. Heatmap of the core OTUs at the sewage (K8) in comparison with K3 and K11. SPM, suspended particulate matter; SED, SPM enriched with sediments; Pre, pre-monsoon; Post, post-monsoon. (*) unclassified.
Discussion
Vibrio Dynamic: Culturability and Influence of the Environment
The observed endemic nature of V. cholerae populations, predominantly occurring in non-culturable state, is an important aspect of Vibrio survival withstanding physicochemical stresses in estuarine water, also reported in previous studies in the Bengal coastline (Alam et al., 2007; Neogi et al., 2012). A non-culturable state is a dormant phase to subsist unfavorable conditions can be easily reverted to an active phase in an appropriate environment or within the host. This study observed that culturable Vibrio counts were a good estimator for general Vibrio species. Additionally, results of this study showing a lower number of ctxA-positive strains after APW enrichment indicated a biasness of the culture method, preferentially inducing the growth of co-existent non-toxigenic populations of V. cholerae in coastal waters. Although culturable organisms represent one or two orders of magnitude lower than the total population and that culture methods decrease the number of toxigenic V. cholerae, bacterial cultures offer a rough and low-cost estimation of general Vibrio with an active metabolism.
The combination of copious amounts of organic matter from sewage and warmer temperatures favored likely the abundance of toxigenic V. cholerae during the pre-monsoon. Temperature is one of the most important drivers of the distribution of Vibrio and a strong predictor of Vibrio outbreaks (Huq et al., 2005; Siboni et al., 2016). Although the impact of sewage on main nutrients (e.g., ammonium and PON) was comparatively higher during the post-monsoon, its effect on ctxA abundance was not conspicuous. The detection of ctxA gene in estuarine waters, especially nearby the sewage discharge point, highlights the direct association of toxigenic V. cholerae with the potential to cause an outbreak. Vibrio cholerae has the ability to proliferate rapidly under nutrient pulses and untreated wastewater favors the occurrence of V. cholerae in aquatic systems (Gil et al., 2004). Moreover, the expression of pathogenic factors in V. cholerae is enhanced by anoxia (Xu et al., 2003) as probably occurred at the station influenced by the sewage outlet.
The ecology of V. cholerae strains without the ctxA gene is also relevant for human and ecosystem health. These strains may contain other toxins and pathogenic factors such as heat stable enterotoxin, type three-secretion system and cholix toxin (Awasthi et al., 2019). Moreover, they may be linked with the emergence of new pathogenic strains. Outbreaks of diarrhea and skin infections have been reported worldwide caused by V. cholerae without the genes to produce cholera toxin (Awasthi et al., 2019; Schwartz et al., 2019). On one hand, sewage pollution signifies toxigenic V. cholerae of fecal origin reaching the estuarine environment with the potential to impact on aquatic biota and to survive attached to SPM. On the other hand, this means also that environmental V. cholerae strains have the potential to exchange genetic information with toxigenic strains of clinical origin, both metabolically active at elevated temperatures. An environment rich in organic matter favors the horizontal transfer of genes and these settings probably facilitate the evolution of new epidemic serogroups and variant strains of toxigenic V. cholerae and V. mimicus (Neogi et al., 2019).
The load of organic matter in coastal water can be considered as a key driver behind the seasonal dynamic of non-culturable Vibrio across the estuarine and pollution gradient. The relation of non-culturable forms of Vibrio with the organic matter was closer during the post-monsoon as indicated by the CCA. Vibrio species tend to form biofilms in the SPM and display several enzymes for organic matter remineralization (Neogi et al., 2018; Liang et al., 2019). The particulates offer also protection from predation and unfavorable environmental conditions and sometimes are described as islands sheltering pathogenic organisms in the form of biofilms. The conditions of lower temperatures might have contributed to the non-culturable state of Vibrio during the post-monsoon.
A strong effect of salinity and higher temperatures on Vibrio OTUs was suggested by the CCA ordination; salinity was also correlated with Vibrio-GV, Vibrio OTUs and CVCs. Vibrio vulnificus and V. parahaemolyticus are generally detected at higher salinities than V. cholerae (e.g., Kopprio et al., 2017). Several Vibrio species, other than those estimated by the qPCR and CARD-FISH probes, influenced likely the OTU dynamic of this genus. Higher temperatures and salinity may have also a strong effect on Vibrio diversity. Most Vibrio species need at least some salinity for growth and this environmental factor was described as an important driver of Vibrio abundance and distribution (Lara et al., 2009, 2011). Furthermore, the observed abundance peaks of toxigenic V. cholerae at stations of intermediate salinities during the pre-monsoon might be related to their better survival potential in this salinity range. Among the halophilic Vibrio species, V. cholerae has extraordinary potential to tolerate lower salinities in the presence of organic nutrients in aquatic environment (Singleton et al., 1982).
The stations more influenced by the sea were characterized by surface waters with elevated values of δ13C, pH and nitrate. An elevated 13C signature in the POC is characteristic of the mouth of estuaries (e.g., Thornton and McManus, 1994; Kopprio et al., 2018). The POC enriched in 13C at the marine stations may be related to organic matter derived from phytoplankton or microzooplankton. Furthermore, the higher pH at the estuarine mouth may favor also the abundance of bicarbonate ions, which are enriched by about 8 ‰ compared to CO2 (Ostrom et al., 1997). Some microalgae such as diatoms and cyanobacteria are able to incorporate bicarbonate ions (Moschen et al., 2009) and may increase the δ13C in the POC at the marine stations. A higher pH potentially benefited the abundance of Vibrio species, which are generally cultured in alkaline media (e.g., APW). Some Vibrio species are able to reduce nitrate using it as an electron acceptor for respiration (Lara et al., 2011), but this process is unlikely under the well-oxygenated waters of the marine stations.
The coastal sediments of the Karnaphuli estuary were detected as important reservoirs for non-culturable V. cholerae, Vibrio species and organic matter. This is in congruence with previous studies claiming that benthic sediment of coastal zones provides shelter and nutrients for Vibrio populations (Vezzulli et al., 2009; Neogi et al., 2018). Coastal storms, particularly cyclones and heavy rains, increase water runoff and the resuspension of sediment together with several microorganisms (e.g., Lara et al., 2009). Usually after cyclones, the rural populations of Bangladesh suffer outbreaks of cholera and we inferred that the cyclone Roanu transported upstream V. cholerae, organic nutrients and salt water. There were no differences between the nanoplankton and SPM, most of Vibrio were detected as free-living or attached to particles lower than 20 μm. Nanoplankton is the fraction of seston, where the higher abundances of V. cholerae are usually observed (Lara et al., 2011; Neogi et al., 2018). However, the role of larger fractions as vector of V. cholerae should not be underestimated, Vibrio species comprise an important proportion of natural microbiome of zooplankton (Constantin de Magny et al., 2008; Grim et al., 2009).
Bacterial Community Composition and Potential Roles Across the Estuarine Continuum
The lack of seasonal differences at order, genera or OTU level may be related to the strong estuarine gradient. The Rhodobacterales (class Alphaproteobacteria) and the Oceanospirillales, Vibrionales and Cellvibrionales (Gammaproteobacteria) characterized the marine regions of the Karnaphuli estuary. Certain Alphaproteobacteria and Gammaproteobacteria in general are copiotrophic organisms equipped with a number of tools to degrade polymers and particles (Fuhrman et al., 2015). Large amounts of particulate and dissolved organic matter are characteristic of the watercourses flowing into the Bay of Bengal. Gammaproteobacteria and Alphaproteobacteria dominated the bacterial community composition of the waters of the Bay of Bengal (Ghosh and Bhadury, 2018; Rajpathak et al., 2018; Fernandes et al., 2019; Dhal et al., 2020) and these classes together with Deltaproteobacteria were detected abundantly in sediments of the Sundarbans (Basak et al., 2015). In this study, Deltaproteobacteria were mainly represented by Geobacter at K8 with relevance for the arsenic cycle (Gnanaprakasam et al., 2017).
The observed dynamic of Rhodobacterales and Oceanospirillales may reflect that these taxa were early colonizers of the allochthonous organic matter flowing into the Bay of Bengal. The Rhodobacterales are dominant and ubiquitous primary surface colonizer, while the Oceanospirillales are a fast-surface-colonizing group (Dang et al., 2008). According to the last authors, the production of extracellular polymeric substances (EPS) by Rhodobacterales facilitates the settlement of other bacterial communities. Marivivens may be responsible of this key role at the Karnaphuli estuary. Roseobacter clade CHAB-I-5 lineage may colonize particles derived from freshwater organisms, which suffer a considerable osmotic shock at the mouth of the estuary. Pelagic members of Roseobacter have been related to chlorophyll peaks in estuaries (Alonso et al., 2010) and play a major role in processing organic matter derived from phytoplankton (Sarmento and Gasol, 2012; Bakenhus et al., 2017). The POC presented typically an enriched 13C signature at the mouth of estuaries and several causes explaining this phenomenon were discussed in the previous section.
Cyanobium PCC-6307 was the main cyanobacteria at the freshwater stations. Synechococcales bacteria contribute to more than the 25% of the global photosynthesis (Fuhrman et al., 2015). Cyanobium PCC-6307 is characterized by strong bioactive compounds against the growth of other microorganisms (Costa et al., 2015) and may influence the freshwater communities. The Microtrichales, Micrococcales and Frankiales of the phylum Actinobacteria may produce also bioactive compounds and help to remineralize recalcitrant organic matter. Actinobacteria are widely distributed in aquatic and terrestrial systems and play a key role in the production of antibiotics, recycling of nutrients and degradation of complex polymers (Alvarez et al., 2017). The Chthoniobacterales (phylum Verrucomicrobia) and Pirellulales (Planctomycetes) were likely associated with the distribution of Cyanobium PCC-6307. According to Bunse and Pinhassi (2017), Verrucomicrobia are related with cyanobacterial blooms due to their diverse enzymes to degrade polysaccharides, while Planctomycetes are scavengers of semilabile dissolved organic matter derived from the bloom.
Sewage Impact: Bacterial Diversity, Presumptive Functions and Potential Pathogens
Cloacibacterium and Arcobacter as putative EPS producers, Bacteroidales and depletion of 15N were clear indicators of the impact of sewage. The δ13C dynamic did not indicate sewage pollution. Cloacibacterium, detected abundantly during the pre-monsoon, was isolated originally from municipal wastewater and together with other members of the order Flavobacteriales (Flavobacterium and Bergeyella detected in this study, phylum Bacteroidetes) are associated with the breakdown of complex organic matter (Bernardet et al., 2002). Cloacibacterium normanense may play a key role in phosphate removal (Allen et al., 2006) and produces EPS with flocculant and metal removal properties (Nouha et al., 2016; Wang et al., 2018), processes key for bioremediation or water treatment. Additionally, the production of EPS by Cloacibacterium may support its role as early colonizer of anthropogenic organic matter.
Arcobacter of the order Campylobacterales (phylum Epsilonbacteraeota) was a marker of sewage pollution and even negative δ15N values were detected during the post-monsoon. Similar to Cloacibacterium, Arcobacter species are known to secrete EPS and to form biofilms (Ross et al., 2001), and these properties give to members of this genus, a central role in processes of bioremediation and colonization of organic matter. Furthermore, several Arcobacter species are indicators of high levels of fecal pollution and emerging water-borne pathogens causing gastroenteritis, abortions and bacteremia (Collado et al., 2008; Khan et al., 2017). Certain Arcobacter species are able to fix atmospheric nitrogen (Collado et al., 2008) and this process may have been linked with the lower values of δ15N at the sewage. However, the fixation of atmospheric nitrogen is very unlikely at the dissolved nitrogen concentrations detected, and 15N depletion occurred also during the pre-monsoon. As mentioned in the introduction, there is recent evidence of depleted 15N values in POM from heavy-polluted aquatic systems. Under condition of elevated ammonium values and oxygen limitation, there is an accumulation of bacterial biomass with 15N-depleted values in POM (Lehmann et al., 2002; Kopprio et al., 2018). Values below −20 ‰ have been reported in ammonia emitted by waste (David Felix et al., 2013). Ammonia from ammonification processes in aqueous solution is transformed to ammonium and its incorporation to bacterial biomass depletes likely the signature of POM.
Nitrate and nitrite may be utilized by some bacteria as electron acceptors under hypoxic conditions at the sewage impacted station. Some bacteria, and particularly the Epsilonbacteraeota (mainly Arcobacter and Sulfurospirillum in this study), utilize a broad range of electron donors, both organic and inorganic, in absence of molecular oxygen. Low dissolved oxygen concentrations were measured at K8 (∼2 mg L–1), anoxic conditions are suspected in the waste-water (previous to be mixed with estuarine waters) or near the sediments at K8. The peak of Arcobacter coincided with negligible concentrations of nitrate and nitrite. Arcobacter species have been shown to degrade organic compounds with nitrate as electron acceptor (Roalkvam et al., 2015). Furthermore, the families Burkholderiaceae and Rhodocyclaceae (C39 and Dechloromonas, order Betaproteobacteriales) were described with denitrifying activity (Cao et al., 2017) and may reduce nitrate and nitrite at K8. The remaining nitrate after this fractionation may be characterized by an elevated 15N signature and may have contributed to an increment of δ15N of POM at the marine stations.
Prevotella 9, Prevotellaceae unclassified, Macellibacteroides and Paludibacter of the order Bacteroidales (phylum Bacteroidetes) were clear indicators of untreated wastewater and hypoxia during both seasons. Bacteroidales bacteria are used to assess sources of fecal pollution (Cao et al., 2017). For example, Prevotella, the main genus of the Bacteroidales in the Karnaphuli estuary, is a fiber fermenter associated with non-industrialized populations whose diet contains more dietary fiber (Chen et al., 2017). Other microbial indicators of wastewater input and hypoxia were Macellibacteroides, Anaerospora, Veillonellaceae unclassified, Streptococcus, Hypnocyclicus and Synergistaceae unclassified. For instance, Streptococcus has been linked to the gut microbiota recovery after V. cholerae infection (Hsiao et al., 2014); though the genus comprises important pathogens such as S. pneumonia and S. pyogenes. Gammaproteobacteria were supported likely by the copious amounts of organic matter at the sewage and intermediate stations and played a key function as early colonizers. Genera of potential pathogens other than Streptococcus, Arcobacter and Vibrio linked with sewage pollution were Acinetobacter and Aeromonas.
The proportions of Vibrio, Escherichia-Shigella and Enterococcus in the estuary might have been influenced by the selected primers for 16S rRNA analyses. Despite the higher number of GU and CARD-FISH counts for Vibrio at the sewage of the Karnaphuli estuary, the relative sequence abundances of this genus were below the threshold of 1%. A deeper study of Vibrio communities requires the use of specific primers like Vib-169F and Vib2-r (e.g., Siboni et al., 2016) but with the consequent loss of information about other organisms. Although culturable coliforms were associated to the sewage, other potential indicators such as Escherichia-Shigella and Enterococcus were not also parts of the core communities at K8. The V4 primer pair has been described as unsuitable for the detection of enteric pathogens in wastewater (Greay et al., 2019). Overall, the results of bacterial diversity obtained in this study may reflect the real dominance of other communities over Vibrio, Escherichia-Shigella and Enterococcus.
Conclusion and Outlook
Fecal pollution and warmer temperatures were the deduced drivers of the abundance of toxigenic V. cholerae. Vibrio outbreaks are expected to increase under climate change scenarios and waste-water management strategies are urgently needed to reduce the risks of Vibrio and other potential pathogens linked to sewage. The particulates influenced mostly non-culturable Vibrio, while salinity in combination with temperature was mostly related with Vibrio diversity. Coastal sediments were an important reservoir for Vibrio and organic matter. The protection of wetlands would help to reduce sediment resuspension during coastal storms or cyclones, and contribute with nutrient sequestration.
The island biogeography theory explained the dynamic of some bacterial groups in the Karnaphuli estuary. A few organisms act as early colonizers of allochthonous organic matter (freshwater or anthropogenic) and facilitate the later settlement of other bacteria. The presumptive production of EPS by Rhodobacterales, Arcobacter or Cloacibacterium may facilitate the mentioned role. The changing conditions of estuaries favor the abundance of early colonizers and the copiotrophic setting of the Karnaphuli estuary promoted the abundance of Gammaproteobacteria. Relevant bacteria for waste water treatment, bioremediation and further biotechnological approaches are already in the estuarine system.
The δ13C dynamic was hypothetically driven by planktonic organic matter and pH, while the δ15N by microorganisms with a depleted 15N signature at the sewage. Nitrate and nitrite were likely used as a substrate for bacterial metabolism, particularly by Epsilonbacteraeota, under the hypoxic conditions of the sewage. Several members of Bacteroidales, as well as culturable coliforms and ammonium, were stronger markers of sewage and hypoxia during both seasons. The use of 16S rRNA diversity allows us only to infer some functions on bacterial communities, further studies from a metagenomic and metatranscriptomic approach will confirm key genes involved on bioremediation and biogeochemical processes and their expression. Moreover, to define a pathogen only with the genus is vague (e.g., Streptococcus) and the detection of pathogenic factors is desirable; nevertheless, this study give us an idea about the potential risks.
The coastline of Bangladesh is extremely prone to natural disasters and one of the most vulnerable areas to climate change. Higher temperatures, stronger and more frequent coastal storms or cyclones, salinization of upstream courses, eutrophication-like processes, floods and droughts; in combination with an increasing population will signify considerable challenges for water and food security. Low-cost and adaptive management strategies to sustain a good water quality in the estuary are urgently needed in order to reduce the impact of water-borne bacterial diseases, pollutants and eutrophication. The development of educational programs, strategies to cope natural disasters, sanitation and vaccination campaigns and early warning systems are also crucial to enhance social-ecological resilience.
Data Availability Statement
16S sequencing data are available at ENA (https://www.ebi.ac.uk/ena/data/view/PRJEB35775).
Author Contributions
GK, SN, RL, BK, SY, and AG performed the study design. GK, SN, HR, and CA carried out the field, laboratory work and data analyses. The first draft of the manuscript was written by GK and the final version was improved by contributions of all authors.
Funding
GK was benefited with a Leibniz-DAAD fellowship (91536278). The publication of this article was funded by the Open Access Fund of the Leibniz Association.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are grateful with H. Westphal for institutional support, with C. Bureau and K-U. Ludwichowski for nutrient determinations, with D. Dasbach for stable isotope analysis and with S. Peters for qPCR assessment. We also thank A. Oishee and S. Sarkar for field and laboratory assistance and C. Hassenrück for bioinformatic support. We also thank the reviewers for their constructive comments on earlier versions of this manuscript. To the memory of our colleague D. Peterke.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2020.00594/full#supplementary-material
FIGURE S1 | Relative sequence abundance of the main orders in the fractions comparing seasons and stations (K).
Footnotes
References
Alam, M., Sultana, M., Nair, G. B., Siddique, A. K., Hasan, N. A., Sack, R. B., et al. (2007). Viable but non-culturable Vibrio cholerae O1 in biofilms in the aquatic environment and their role in cholera transmission. Proc. Nat. Acad. Sci. U.S.A. 104:17801. doi: 10.1073/pnas.0705599104
Allen, T. D., Lawson, P. A., Collins, M. D., Falsen, E., and Tanner, R. S. (2006). Cloacibacterium normanense gen. nov., sp. nov., a novel bacterium in the family Flavobacteriaceae isolated from municipal wastewater. Int. J. Syst. Evol. Microbiol. 56, 1311–1316. doi: 10.1099/ijs.0.64218-0
Alonso, C., Gómez-Pereira, P., Ramette, A., Ortega, L., Fuchs, B. M., and Amann, R. (2010). Multilevel analysis of the bacterial diversity along the environmental gradient Río de La Plata-South Atlantic Ocean. Aquat. Microb. Ecol. 61, 57–72. doi: 10.3354/ame01439
Alvarez, A., Saez, J. M., Davila Costa, J. S., Colin, V. L., Fuentes, M. S., Cuozzo, S. A., et al. (2017). Actinobacteria: current research and perspectives for bioremediation of pesticides and heavy metals. Chemosphere 166, 41–62. doi: 10.1016/j.chemosphere.2016.09.070
Awasthi, S. P., Chowdhury, N., Neogi, S. B., Hinenoya, A., Hatanaka, N., Chowdhury, G., et al. (2019). Development of a multiplex PCR assay for the detection of major virulence genes in Vibrio cholerae including non-O1 and non-O139 serogroups. J. Microbiol. Meth. 157, 54–58. doi: 10.1016/j.mimet.2018.12.012
Bakenhus, I., Dlugosch, L., Billerbeck, S., Giebel, H.-A., Milke, F., and Simon, M. (2017). Composition of total and cell-proliferating bacterioplankton community in early summer in the North Sea – roseobacters are the most active component. Front. Microbiol. 8:1771. doi: 10.3389/fmicb.2017.01771
Basak, P., Majumder, N. S., Nag, S., Bhattacharyya, A., Roy, D., Chakraborty, A., et al. (2015). Spatiotemporal analysis of bacterial diversity in sediments of Sundarbans using parallel 16S rRNA gene tag sequencing. Microb. Ecol. 69, 500–511. doi: 10.1007/s00248-014-0498-y
Bernardet, J. F., Nakagawa, Y., and Holmes, B. (2002). Proposed minimal standards for describing new taxa of the family flavobacteriaceae and emended description of the family. Int. J. Syst. Evol. Microbiol. 52, 1049–1070. doi: 10.1099/00207713-52-3-1049
Bliem, R., Schauer, S., Plicka, H., Obwaller, A., Sommer, R., Steinrigl, A., et al. (2015). A novel triplex quantitative PCR strategy for quantification of toxigenic and non-toxigenic Vibrio cholerae in aquatic environments. Appl. Environ. Microbiol. 81, 3077–3085. doi: 10.1128/AEM.03516-14
Bolger, A. M., Usadel, B., and Lohse, M. (2014). Trimmomatic: a flexible trimmer for Illumina sequence data. Bioinformatics 30, 2114–2120. doi: 10.1093/bioinformatics/btu170
Boström, K. H., Simu, K., Hagström, A., and Riemann, L. (2004). Optimization of DNA extraction for quantitative marine bacterioplankton community analysis. Limnol. Oceanogr. Methods 2, 365–373. doi: 10.4319/lom.2004.2.365
Bunse, C., and Pinhassi, J. (2017). Marine bacterioplankton seasonal succession dynamics. Trends Microbiol. 25, 494–505. doi: 10.1016/j.tim.2016.12.013
Cao, Q., Wang, H., Chen, X., Wang, R., and Liu, J. (2017). Composition and distribution of microbial communities in natural river wetlands and corresponding constructed wetlands. Ecol. Eng. 98, 40–48. doi: 10.1016/j.ecoleng.2016.10.063
Chen, T., Long, W., Zhang, C., Liu, S., Zhao, L., and Hamaker, B. R. (2017). Fiber-utilizing capacity varies in Prevotella- versus Bacteroides-dominated gut microbiota. Sci. Rep. 7:2594. doi: 10.1038/s41598-017-02995-4
Collado, L., Inza, I., Guarro, J., and Figueras, M. J. (2008). Presence of Arcobacter spp. in environmental waters correlates with high levels of fecal pollution. Environ. Microbiol. 10, 1635–1640. doi: 10.1111/j.1462-2920.2007.01555.x
Constantin de Magny, G., Murtugudde, R., Sapiano, M. R. P., Nizam, A., Brown, C. W., Busalacchi, A. J., et al. (2008). Environmental signatures associated with cholera epidemics. Proc. Nat. Acad. Sci. U.S.A. 105, 17676–17681. doi: 10.1073/pnas.0809654105
Costa, M. S., Costa, M., Ramos, V., Leão, P. N., Barreiro, A., Vasconcelos, V., et al. (2015). Picocyanobacteria from a clade of marine Cyanobium revealed bioactive potential against microalgae, bacteria, and marine invertebrates. J. Toxicol. Environ. Health A 78, 432–442. doi: 10.1080/15287394.2014.991466
Dang, H., Li, T., Chen, M., and Huang, G. (2008). Cross-ocean distribution of Rhodobacterales bacteria as primary surface colonizers in temperate coastal marine waters. Appl. Environ. Microbiol. 74, 52–60. doi: 10.1128/aem.01400-07
David Felix, J., Elliott, E. M., Gish, T. J., McConnell, L. L., and Shaw, S. L. (2013). Characterizing the isotopic composition of atmospheric ammonia emission sources using passive samplers and a combined oxidation-bacterial denitrifier approach. Rapid Commun. Mass Sci. 27, 2239–2246. doi: 10.1002/rcm.6679
Dhal, P. K., Kopprio, G. A., and Gärdes, A. (2020). Insights on aquatic microbiome of the Indian Sundarbans mangrove areas. PLoS One 15:0221543. doi: 10.1371/journal.pone.0221543
Diepenbroek, M., Glöckner, F., Grobe, P., Güntsch, A., Huber, R., König-Ries, B., et al. (2014). “Towards an integrated biodiversity and ecological research data management and archiving platform: the german federation for the curation of biological data (GFBio),” in Informatik 2014 – Big Data Komplexität meistern. GI-Edition: Lecture Notes in Informatics (LNI) – Proceedings, eds E. Plödereder, L. Grunske, E. Schneider, and D. Ull (Bonn: Köllen Verlag), 1711–1724.
Eren, A. M., Morrison, H. G., Lescault, P. J., Reveillaud, J., Vineis, J. H., and Sogin, M. L. (2015). Minimum entropy decomposition: unsupervised oligotyping for sensitive partitioning of high-throughput marker gene sequences. ISME J. 9, 968–979. doi: 10.1038/ismej.2014.195
Fernandes, G. L., Shenoy, B. D., Menezes, L. D., Meena, R. M., and Damare, S. R. (2019). Prokaryotic diversity in oxygen depleted waters of the Bay of Bengal inferred using culture-dependent and -independent methods. Indian J. Microbiol. 59, 193–199. doi: 10.1007/s12088-019-00786-1
Fuhrman, J. A., Cram, J. A., and Needham, D. M. (2015). Marine microbial community dynamics and their ecological interpretation. Nat. Rev. Microbiol. 13, 133–146. doi: 10.1038/nrmicro3417
Ghosh, A., and Bhadury, P. (2018). Investigating monsoon and post-monsoon variabilities of bacterioplankton communities in a mangrove ecosystem. Environ. Sci. Pollut. Res. 25, 5722–5739. doi: 10.1007/s11356-017-0852-y
Gil, A. I., Louis, V. R., Rivera, I. N. G., Lipp, E., Huq, A., Lanata, C. F., et al. (2004). Occurrence and distribution of Vibrio cholerae in the coastal environment of Peru. Environ. Microbiol. 6, 699–706. doi: 10.1111/j.1462-2920.2004.00601.x
Giuliano, L., De Domenico, M., De Domenico, E., Höfle, M. G., and Yakimov, M. M. (1999). Identification of culturable oligotrophic bacteria within naturally occurring bacterioplankton communities of the Ligurian sea by 16S rRNA sequencing and probing. Microb. Ecol. 37, 77–85. doi: 10.1007/s002489900132
Gnanaprakasam, E. T., Lloyd, J. R., Boothman, C., Ahmed, K. M., Choudhury, I., Bostick, B. C., et al. (2017). Microbial community structure and arsenic biogeochemistry in two arsenic-impacted aquifers in Bangladesh. mBio 8:1326. doi: 10.1128/mBio.01326-17
Greay, T. L., Gofton, A. W., Zahedi, A., Paparini, A., Linge, K. L., Joll, C. A., et al. (2019). Evaluation of 16S next-generation sequencing of hypervariable region 4 in wastewater samples: an unsuitable approach for bacterial enteric pathogen identification. Sci. Tot. Environ. 670, 1111–1124. doi: 10.1016/j.scitotenv.2019.03.278
Grim, C. J., Zo, Y.-G., Hasan, N. A., Ali, A., Chowdhury, W. B., Islam, A., et al. (2009). RNA colony blot hybridization method for enumeration of culturable Vibrio cholerae and Vibrio mimicus bacteria. Appl. Environ. Microbiol. 75, 5439–5444. doi: 10.1128/aem.02007-08
Hong, S., Lee, Y., Yoon, S. J., Lee, J., Kang, S., Won, E.-J., et al. (2019). Carbon and nitrogen stable isotope signatures linked to anthropogenic toxic substances pollution in a highly industrialized area of South Korea. Mar. Pollut. Bull. 144, 152–159. doi: 10.1016/j.marpolbul.2019.05.006
Hsiao, A., Ahmed, A. M. S., Subramanian, S., Griffin, N. W., Drewry, L. L., Petri, W. A., et al. (2014). Members of the human gut microbiota involved in recovery from Vibrio cholerae infection. Nature 515, 423–426. doi: 10.1038/nature13738
Huq, A., Sack, R. B., Nizam, A., Longini, I. M., Nair, G. B., Ali, A., et al. (2005). Critical Factors influencing the occurrence of Vibrio cholerae in the environment of Bangladesh. Appl. Environ. Microbiol. 71, 4645–4654. doi: 10.1128/aem.71.8.4645-4654.2005
Ke, Z., Tan, Y., Huang, L., Zhao, C., and Jiang, X. (2017). Spatial distributions of δ13C, δ15N and C/N ratios in suspended particulate organic matter of a bay under serious anthropogenic influences: daya Bay, China. Mar. Pollut. Bull. 114, 183–191. doi: 10.1016/j.marpolbul.2016.08.078
Kegler, H. F., Lukman, M., Teichberg, M., Plass-Johnson, J., Hassenrück, C., Wild, C., et al. (2017). Bacterial community composition and potential driving factors in different reef habitats of the Spermonde Archipelago, Indonesia. Front. Microbiol. 8:662. doi: 10.3389/fmicb.2017.00662
Khan, I. U. H., Cloutier, M., Libby, M., Lapen, D. R., Wilkes, G., and Topp, E. (2017). Enhanced single-tube multiplex PCR assay for detection and identification of six Arcobacter species. J. Appl. Microbiol. 123, 1522–1532. doi: 10.1111/jam.13597
Klindworth, A., Pruesse, E., Schweer, T., Peplies, J., Quast, C., Horn, M., et al. (2012). Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucl. Acids Res. 41:1. doi: 10.1093/nar/gks808
Kopprio, G. A., Dutto, M. S., Garzón Cardona, J. E., Gärdes, A., Lara, R. J., and Graeve, M. (2018). Biogeochemical markers across a pollution gradient in a patagonian estuary: a multidimensional approach of fatty acids and stable isotopes. Mar. Pollut. Bull. 137, 617–626. doi: 10.1016/j.marpolbul.2018.10.059
Kopprio, G. A., Streitenberger, M. E., Okuno, K., Baldini, M., Biancalana, F., Fricke, A., et al. (2017). Biogeochemical and hydrological drivers of the dynamics of Vibrio species in two Patagonian estuaries. Sci. Tot. Environ. 579, 646–656. doi: 10.1016/j.scitotenv.2016.11.045
Lara, R. J., Neogi, S. B., Islam, M. S., Mahmud, Z. H., Islam, S., Paul, D., et al. (2011). Vibrio cholerae in waters of the Sunderban mangrove: relationship with biogeochemical parameters and chitin in seston size fractions. Wetl. Ecol. Manag. 19, 109–119. doi: 10.1007/s11273-010-9204-0
Lara, R., Neogi, S., Islam, M., Mahmud, Z., Yamasaki, S., and Nair, G. (2009). Influence of catastrophic climatic events and human waste on Vibrio distribution in the Karnaphuli estuary, Bangladesh. Ecohealth 6, 279–286. doi: 10.1007/s10393-009-0257-6
Lehmann, M. F., Bernasconi, S. M., Barbieri, A., and McKenzie, J. A. (2002). Preservation of organic matter and alteration of its carbon and nitrogen isotope composition during simulated and in situ early sedimentary diagenesis. Geochim. Cosmochim. Acta 66, 3573–3584. doi: 10.1016/S0016-7037(02)00968-7
Liang, J., Liu, J., Wang, X., Lin, H., Liu, J., Zhou, S., et al. (2019). Spatiotemporal dynamics of free-living and particle-associated Vibrio communities in the Northern Chinese Marginal Seas. Appl. Environ. Microbiol. 85, 217–219. doi: 10.1128/AEM.00217-19
Marker, A. F. H., Nusch, E. A., Rai, H., and Riemann, B. (1980). The measurement of photosynthetic pigments in freshwaters and standardization of methods: conclusions and recommendations. Arch. Hydrobiol. Bull. 14, 91–106.
Morens, D. M., Folkers, G. K., and Fauci, A. S. (2004). The challenge of emerging and re-emerging infectious diseases. Nature 430, 242–249. doi: 10.1038/nature02759
Moschen, R., Lücke, A., Parplies, J., and Schleser, G. H. (2009). Controls on the seasonal and interannual dynamics of organic matter stable carbon isotopes in mesotrophic Lake Holzmaar, Germany. Limnol. Oceanogr. 54, 194–209.
Mutreja, A., Kim, D. W., Thomson, N. R., Connor, T. R., Lee, J. H., Kariuki, S., et al. (2011). Evidence for several waves of global transmission in the seventh cholera pandemic. Nature 477, 462–465. doi: 10.1038/nature10392
Neogi, S. B., Chowdhury, N., Awasthi, S. P., Asakura, M., Okuno, K., Mahmud, Z. H., et al. (2019). Novel cholera toxin variant and ToxT regulon in environmental Vibrio mimicus isolates: potential resources for the evolution of Vibrio cholerae hybrid strains. Appl. Environ. Microbiol. 85:1977. doi: 10.1128/AEM.01977-18
Neogi, S. B., Islam, M. S., Nair, G. B., Yamasaki, S., and Lara, R. J. (2012). Occurrence and distribution of plankton-associated and free-living toxigenic Vibrio cholerae in a tropical estuary of a cholera endemic zone. Wetl. Ecol. Manag. 20, 271–285. doi: 10.1007/s11273-012-9247-5
Neogi, S. B., Lara, R., Alam, M., Harder, J., Yamasaki, S., and Colwell, R. R. (2018). Environmental and hydroclimatic factors influencing Vibrio populations in the estuarine zone of the Bengal delta. Environ. Monit. Assess. 190:565. doi: 10.1007/s10661-018-6925-7
Nouha, K., Kumar, R. S., and Tyagi, R. D. (2016). Heavy metals removal from wastewater using extracellular polymeric substances produced by Cloacibacterium normanense in wastewater sludge supplemented with crude glycerol and study of extracellular polymeric substances extraction by different methods. Bioresour. Technol. 212, 120–129. doi: 10.1016/j.biortech.2016.04.021
Ostrom, N. E., Macko, S. A., Deibel, D., and Thompson, R. J. (1997). Seasonal variation in the stable carbon and nitrogen isotope biogeochemistry of a coastal cold ocean environment. Geochim. Cosmochim. Acta 61, 2929–2942. doi: 10.1016/S0016-7037(97)00131-2
Pernthaler, A., Pernthaler, J., and Amann, R. (2002). Fluorescence In situ hybridization and catalyzed reporter deposition for the identification of marine bacteria. Appl. Environ. Microbiol. 68, 3094–3101. doi: 10.1128/aem.68.6.3094-3101.2002
Quast, C., Pruesse, E., Yilmaz, P., Gerken, J., Schweer, T., Yarza, P., et al. (2013). The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucl. Acids Res. 41, 590–596. doi: 10.1093/nar/gks1219
R Core Team (2018). A Language And Environment For Statistical Computing. Vienna: R Foundation for Statistical Computing.
Rajpathak, S. N., Banerjee, R., Mishra, P. G., Khedkar, A. M., Patil, Y. M., Joshi, S. R., et al. (2018). An exploration of microbial and associated functional diversity in the OMZ and non-OMZ areas in the Bay of Bengal. J. Biosci. 43, 635–648. doi: 10.1007/s12038-018-9781-2
Roalkvam, I., Drønen, K., Stokke, R., Daae, F. L., Dahle, H., and Steen, I. H. (2015). Physiological and genomic characterization of Arcobacter anaerophilus IR-1 reveals new metabolic features in epsilonproteobacteria. Front. Microbiol. 6:987. doi: 10.3389/fmicb.2015.00987
Ross, N., Villemur, R., Deschenes, L., and Samson, R. (2001). Clogging of a limestone fracture by stimulating groundwater microbes. Water Res. 35, 2029–2037. doi: 10.1016/S0043-1354(00)00476-0
Sack, D. A., Sack, R. B., Nair, G. B., and Siddique, A. K. (2004). Cholera. Lancet 363, 223–233. doi: 10.1016/S0140-6736(03)15328-7
Sarmento, H., and Gasol, J. M. (2012). Use of phytoplankton-derived dissolved organic carbon by different types of bacterioplankton. Environ. Microbiol. 14, 2348–2360. doi: 10.1111/j.1462-2920.2012.02787.x
Schwartz, K., Hammerl, J. A., Göllner, C., and Strauch, E. (2019). Environmental and clinical strains of Vibrio cholerae Non-O1, Non-O139 from Germany possess similar virulence gene profiles. Front. Microbiol. 10:733. doi: 10.3389/fmicb.2019.00733
Schwarzenbach, R. P., Egli, T., Hofstetter, T. B., Gunten, U. V., and Wehrli, B. (2010). Global water pollution and human health. Annu. Rev. Env. Resour. 35, 109–136. doi: 10.1146/annurev-environ-100809-125342
Siboni, N., Balaraju, V., Carney, R., Labbate, M., and Seymour, J. R. (2016). Spatiotemporal dynamics of Vibrio spp. within the sydney harbour estuary. Front. Microbiol. 7:460. doi: 10.3389/fmicb.2016.00460
Singleton, F. L., Attwell, R. W., Jangi, M. S., and Colwell, R. R. (1982). Influence of salinity and organic nutrient concentration on survival and growth of Vibrio cholerae in aquatic microcosms. Appl. Environ. Microbiol. 43, 1080–1085.
Thornton, S. F., and McManus, J. (1994). Application of organic carbon and nitrogen stable isotope and C/N ratios as source indicators of organic matter provenance in estuarine systems: evidence from the Tay Estuary, Scotland. Estuar. Coast. Shelf Sci. 38, 219–233. doi: 10.1006/ecss.1994.1015
Vezzulli, L., Pezzati, E., Moreno, M., Fabiano, M., Pane, L., and Pruzzo, C. (2009). Benthic ecology of Vibrio spp. and pathogenic Vibrio species in a coastal Mediterranean environment (La Spezia Gulf, Italy). Microb. Ecol. 58, 808–818. doi: 10.1007/s00248-009-9542-8
Vouga, M., and Greub, G. (2016). Emerging bacterial pathogens: the past and beyond. Clin. Microbiol. Infect. 22, 12–21. doi: 10.1016/j.cmi.2015.10.010
Wang, A.-J., Kawser, A., Xu, Y.-H., Ye, X., Rani, S., and Chen, K. (2016). Heavy metal accumulation during the last 30 years in the Karnaphuli River estuary, Chittagong, Bangladesh. Springerplus 5:2079. doi: 10.1186/s40064-016-3749-1
Wang, X., Hua, Z., and Mao, H. (2018). Influential factors for metal ions removal using extracellular polymeric substances produced by Cloacibacterium normanense. Water Environ. J. 32, 650–656. doi: 10.1111/wej.12363
Xu, Q., Dziejman, M., and Mekalanos, J. J. (2003). Determination of the transcriptome of Vibrio cholerae during intra-intestinal growth and mid-exponential phase in vitro. Proc. Nat. Acad. Sci. U.S.A. 100, 1286–1291. doi: 10.1073/pnas.0337479100
Keywords: 15N depletion, Arcobacter, Cloacibacterium, organic matter, sewage, isotopes, Vibrio cholerae, 16S rRNA diversity
Citation: Kopprio GA, Neogi SB, Rashid H, Alonso C, Yamasaki S, Koch BP, Gärdes A and Lara RJ (2020) Vibrio and Bacterial Communities Across a Pollution Gradient in the Bay of Bengal: Unraveling Their Biogeochemical Drivers. Front. Microbiol. 11:594. doi: 10.3389/fmicb.2020.00594
Received: 03 December 2019; Accepted: 18 March 2020;
Published: 15 April 2020.
Edited by:
Stanley Chun Kwan Lau, The Hong Kong University of Science and Technology, Hong KongReviewed by:
Zhanfei Liu, The University of Texas at Austin, United StatesCharmaine Cheuk Man Yung, The Hong Kong University of Science and Technology, Hong Kong
Copyright © 2020 Kopprio, Neogi, Rashid, Alonso, Yamasaki, Koch, Gärdes and Lara. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Germán A. Kopprio, a29wcHJpb0BpZ2ItYmVybGluLmRl