 Jennifer Scott1
Jennifer Scott1 Monica Sueiro-Olivares1
Monica Sueiro-Olivares1 Waqar Ahmed2,3Christoph Heddergott4
Waqar Ahmed2,3Christoph Heddergott4 Can Zhao1
Can Zhao1 Riba Thomas1Michael Bromley1
Riba Thomas1Michael Bromley1 Jean-Paul Latgé4†
Jean-Paul Latgé4† Sven Krappmann5
Sven Krappmann5 Stephen Fowler2,6
Stephen Fowler2,6 Elaine Bignell1
Elaine Bignell1 Jorge Amich1*
Jorge Amich1*- 1Manchester Fungal Infection Group, Division of Infection, Immunity and Respiratory Medicine, Faculty of Biology, Medicine and Health, School of Biological Sciences, University of Manchester, Manchester Academic Health Science Centre, Manchester, United Kingdom
- 2Respiratory and Allergy Research Group, Division of Infection, Immunity and Respiratory Medicine, Faculty of Biology, Medicine and Health, School of Biological Sciences, University of Manchester, Manchester, United Kingdom
- 3Manchester Institute of Biotechnology, University of Manchester, Manchester, United Kingdom
- 4Unité des Aspergillus, Institut Pasteur, Paris, France
- 5Mikrobiologisches Institut – Klinische Mikrobiologie, Immunologie und Hygiene, Universitätsklinikum Erlangen, Friedrich-Alexander-Universität Erlangen-Nürnberg, Erlangen, Germany
- 6NIHR Manchester Biomedical Research Centre – Manchester Academic Health Science Centre, Manchester University Hospitals NHS Foundation Trust, Manchester, United Kingdom
Pathogen-pathogen interactions in polymicrobial infections are known to directly impact, often to worsen, disease outcomes. For example, co-infection with Pseudomonas aeruginosa and Aspergillus fumigatus, respectively the most common bacterial and fungal pathogens isolated from cystic fibrosis (CF) airways, leads to a worsened prognosis. Recent studies of in vitro microbial cross-talk demonstrated that P. aeruginosa-derived volatile sulfur compounds (VSCs) can promote A. fumigatus growth in vitro. However, the mechanistic basis of such cross-talk and its physiological relevance during co-infection remains unknown. In this study we combine genetic approaches and GC-MS-mediated volatile analysis to show that A. fumigatus assimilates VSCs via cysteine (CysB)- or homocysteine (CysD)-synthase. This process is essential for utilization of VSCs as sulfur sources, since P. aeruginosa-derived VSCs trigger growth of A. fumigatus wild-type, but not of a ΔcysBΔcysD mutant, on sulfur-limiting media. P. aeruginosa produces VSCs when infecting Galleria mellonella and co-infection with A. fumigatus in this model results in a synergistic increase in mortality and of fungal and bacterial burdens. Interestingly, the increment in mortality is much greater with the A. fumigatus wild-type than with the ΔcysBΔcysD mutant. Therefore, A. fumigatus’ ability to assimilate P. aeruginosa derived VSCs significantly triggers a synergistic association that increases the pathobiology of infection. Finally, we show that P. aeruginosa can promote fungal growth when growing on substrates that resemble the lung environment, which suggests that this volatile based synergism is likely to occur during co-infection of the human respiratory airways.
Introduction
The filamentous fungus Aspergillus fumigatus and the bacterium Pseudomonas aeruginosa are common respiratory pathogens. Both are able to infect the lungs of immunosuppressed individuals and also of patients with certain underlying diseases, such as bronchiectasis, chronic obstructive pulmonary disease, hospital acquired pneumonia and, particularly, cystic fibrosis (CF). CF is the result of mutations in the CF transmembrane conductance regulator (CFTR) gene, which encodes a chloride channel. Dysfunction of this channel leads to a failure to secrete chloride ions and to increased water absorption from the airway epithelium, resulting in the formation of viscous mucus that is resistant to mucociliary clearance. As a result, a perfect environment for polymicrobial colonization is developed (Ratjen and Doring, 2003).
Aspergillus fumigatus and Pseudomonas aeruginosa are, respectively, the most prevalent fungal and bacterial pathogens isolated from CF airways (Shoseyov et al., 2006; Amin et al., 2010; Brandt et al., 2018). Recent studies have reported prevalences in CF patients ranging from 31 to 46% for P. aeruginosa (Registry CFFP, 2016, Registry UCF, 2016; Reece et al., 2017), up to 57% for A. fumigatus (Bakare et al., 2003; Baxter et al., 2013a; Burgel et al., 2016), and a prevalence of 15.8% for co-infection (Zhao et al., 2018). Co-infection with both pathogens is associated with a more rapid decline in CF pulmonary function and more respiratory exacerbations, all leading to a worse prognosis (Amin et al., 2010; Reece et al., 2017). Therefore, both pathogens should be considered and targeted in order to improve CF patient outcomes. In support of this, it was shown that an intravenous antibiotic treatment targeting Pseudomonas could also reduce the presence of Aspergillus in CF patient sputum. Treatment translated into a greater improvement in the forced expiratory volume in the first second (FEV1) for patients initially co-infected than in those only infected with Pseudomonas (Baxter et al., 2013b). To understand such cross effects and exploit them to the patients’ benefit a profound insight of the pathogen-pathogen interaction taking place during infection is required.
In the environment microbes constantly interact through molecular signals, which can trigger synergistic and antagonistic associations. Such interactions also occur during polymicrobial infections, and are being increasingly recognized as important pathogenicity factors (Singer, 2010; Peters et al., 2012). The relevance of volatile organic compounds (VOCs), molecular messengers that can exert long distance interactions (Schulz-Bohm et al., 2017), has thus far been largely overlooked. However, the fact that exhaled VOCs are promising diagnostic biomarkers for a number of infections (Ahmed et al., 2017) demonstrates that pathogenic microbes produce these molecules in human tissues, which suggests that they may be at sufficient levels to mediate microbial cross-talk during polymicrobial infections.
Recently, it has been reported that P. aeruginosa-derived dimethyl sulfide (DMS) enhances A. fumigatus growth in vitro, which was proposed to be due to its exploitation as a sulfur source (Briard et al., 2016). We had previously shown that A. fumigatus can utilize volatile sulfur compounds (VSCs) as the sole sulfur source (Amich et al., 2013). Therefore, in this study we aimed to unravel the mechanistic basis of A. fumigatus assimilation of VSCs, and the relevance of P. aeruginosa-derived VSCs for pathogen-pathogen interactions in vivo.
Results
A. fumigatus Assimilates VSCs via Cysteine Synthase-(CysB) and Homocysteine Synthase (CysD) Incorporation of Sulfide
We previously demonstrated that VSCs are produced from the catabolism of methionine (but not from other sulfur sources, Supplementary Figure S1A) and can be assimilated and exploited as the sole sulfur source (Amich et al., 2013). However, it remains unknown whether sulfur acquisition via VSCs is reliant upon gaseous or soluble sources. Aiming to discern between VSC uptake from media or air A. fumigatus was cultured on methionine-containing media, in the inner of two concentrically positioned agar plates, for 3 days to promote VSC absorption into sterile S-depleted distal media. At day 3, the inner plate was removed and the outer S-depleted medium ventilated to ensure elimination of any volatiles, after which A. fumigatus was inoculated. In this experimental set-up, fungal growth should only occur if the VSCs had been absorbed in the agar media. However, no fungal growth was observed (Supplementary Figure S1B), which suggests that A. fumigatus likely assimilates VSCs directly from the air.
To precisely identify the VSCs that serve as S-sources, we grew the fungus on minimal medium containing sulfate or methionine as the sole sulfur source and sampled in the headspace of petri dishes using solid-phase microextraction (SPME) and gas chromatography mass spectrometry (GC-MS) (Labows et al., 1980). No VSCs were detected when measuring headspace samples from sterile media. Five major VSCs were present in headspaces sampled from methionine- but not sulfate-cultured A. fumigatus (Figure 1) and their production occurred independently of nitrogen metabolism (Figure 1). VSC2 (dimethyl sulfide, DMS) and VSC3 (dimethyl disulfide, DMDS) were previously described to be exploited as S-sources when produced by P. aeruginosa (Briard et al., 2016). VSC4 (dithiapentane, DTP) is likely degraded to DMDS (Bellesia et al., 1996), and thus its assimilation likely follows the same route. VSC5 (2-methylthiolan-3-one, MTO) is possibly derived from homocysteine catabolism (Nawrath et al., 2010), but since growth on homocysteine does not trigger distal growth on S-free medium (Supplementary Figure S1A), it is unlikely to be utilized as a major volatile S-source. DMS and DMDS can both be degraded to methanethiol (VSC1), either by a DMDS reductase (Smith and Kelly, 1988), or a DMS monooxygenase (De Bont et al., 1981). Sulfur from methanethiol (MT) could then be assimilated into the metabolism through two different routes. It may be degraded to formaldehyde, hydrogen peroxide, and sulfide by the action of MT oxidase (Suylen et al., 1987; Gould and Kanagawa, 1992), and therefore sulfide (S2–) would be incorporated into cysteine by the action of CysB or to homocysteine by the action of CysD (Figure 2A). As an alternative route, MT has also been described to be entirely added to O-acetylhomoserine by the action of CysD to directly yield methionine, bypassing the methionine synthase dependent methylation of homocysteine (Bolten et al., 2010; Figure 2A).
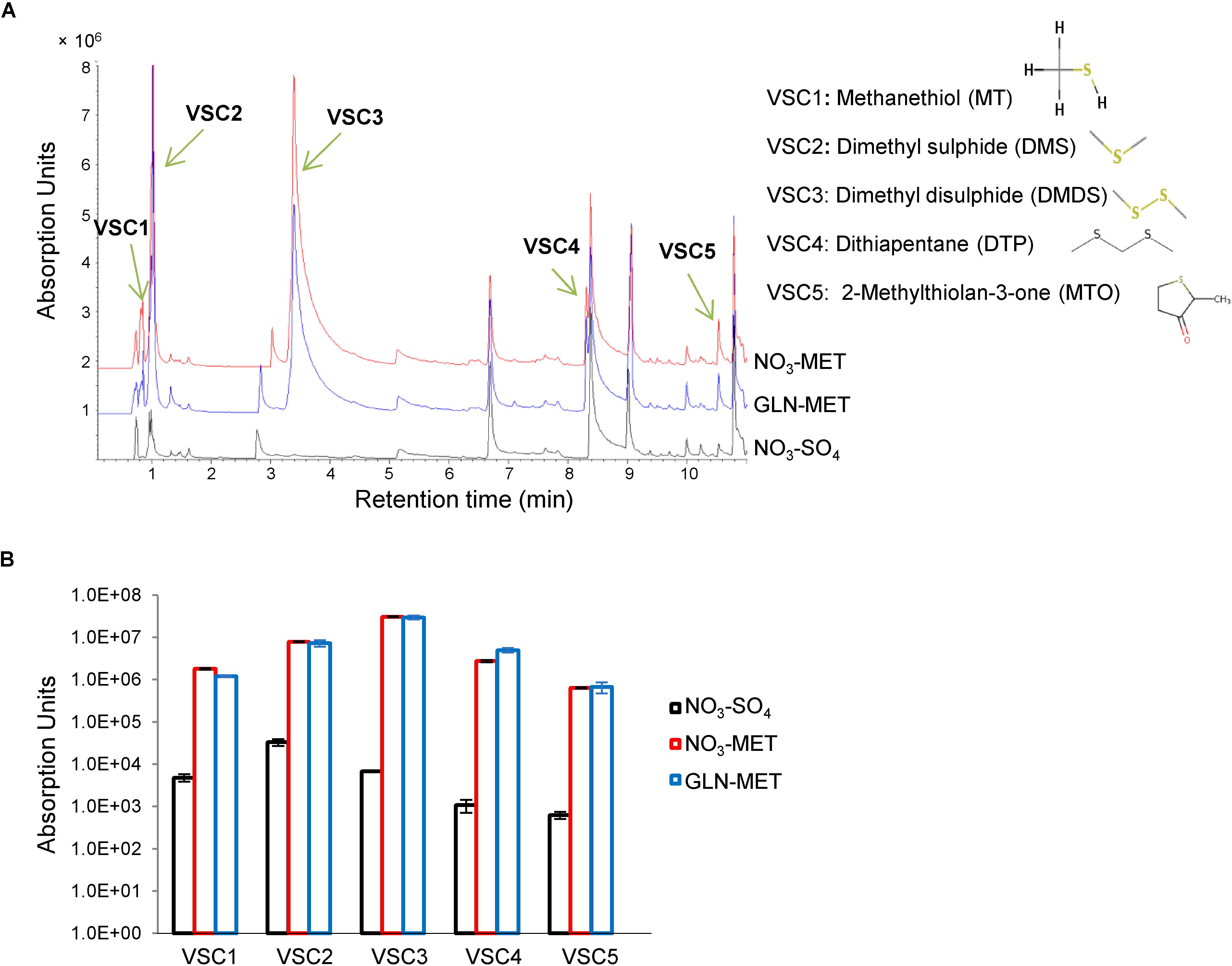
Figure 1. Gas chromatography mass spectrometry (GC-MS) identifies 5 major VSCs produced from Aspergillus fumigatus catabolism of methionine. A. fumigatus was cultured for 3 days on various solid media before the headspace was analyzed using solid-phase microextraction (SPME) and gas chromatography mass spectrometry (GC-MS) to detect VSCs. (A) Chromatogram showing the peaks for the 5 VSCs. These volatiles are produced only on methionine (not on sulfate) and independently of the nitrogen source (ammonium or glutamine). The exact VSCs could be identified and their names and formula are shown. (B) Quantification of Absorption Units for each of the volatiles in three biological replicates.
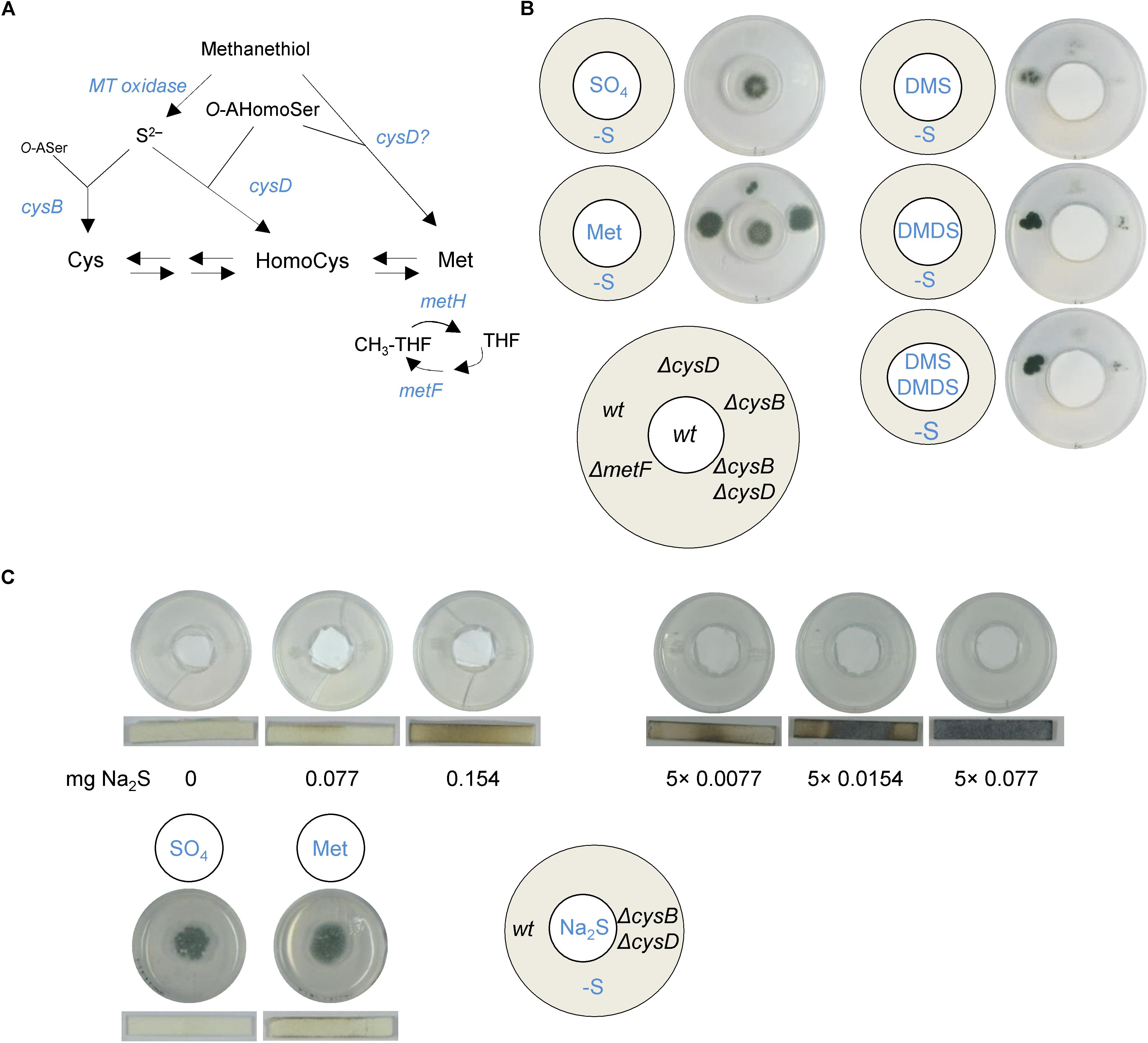
Figure 2. VSCs are incorporated into the trans-sulfuration pathway by the action of CysB and CysD. (A) Outline of the possible routes for VSCs assimilation. Sulfide could be liberated from methanethiol (MT) by an MT oxidase action and incorporated into the metabolism through the action of CysB or CysD. Alternatively, MT could be directly incorporated into homocysteine by CysD to yield methionine, bypassing the requirement of MetH (cysB, cysteine synthase; cysD, homocysteine synthase; metH, methionine synthase; metF, methyl-tetrahydrofolate reductase). (B) Aspergillus derived VSCs and pure DMDS and DMS were not able to trigger growth of a ΔmetF strain. The volatiles triggered growth of the single ΔcysB and ΔcysD mutant strains, although to a much lesser extent than the wild-type, but did not trigger growth of the double ΔcysBΔcysD mutant. ∼8.5 μg of DMS and ∼10.5 μg of DMDS were added every 12 h (5×) to the filter disk in the inner plate. (C) The use of Hydrogen Sulfide Test Strips, which turn dark upon contact with H2S, detected that A. fumigatus produces low amounts of sulfide (comparable to that liberated from ∼0.077 mg of Na2S) from methionine catabolism. Sulfide released from the Na2S donor could not trigger growth of the distal A. fumigatus colony, even if added in higher amounts or in repeated occasions (5 times, once every 12 h). All plates in (B,C) were incubated at 37°C for 3 days.
To elucidate the assimilation route, we constructed mutant strains deleted in genes that are required for each pathway and investigated their ability to grow on S-free media when exposed to VSCs derived from methionine catabolism or to pure DMDS and DMS (Figure 2B). If CysD was the only gene product responsible for MT assimilation, a ΔcysD mutant should not be able to grow on S-free media; however, this strain showed some distal growth (Figure 2B), proving that CysB can also incorporate volatiles and therefore that the MT oxidase route is active. Given that we could not identify any obvious orthologs of MT oxidases described in other microorganisms in A. fumigatus’ genome, we evaluated the MT oxidase activity potential of A. fumigatus protein extracts by measuring the accumulation of formaldehyde upon addition of MT and observed that indeed the fungus seems to have MT oxidase activity (Supplementary Figure S2). To determine if the alternative route is also active we constructed a ΔmetF strain (Figure 2A) to induce a genetic blockade upon the recycling of methylated tetrahydrofolate (CH3-THF), which is required for methionine synthase-dependent methylation of homocysteine. We noticed that ΔmetF grew very poorly on MM supplemented with methionine, but did grow normally if amino acids (aac) were supplemented (Supplementary Figure S3A); this is not surprising as tetrahydrofolate (THF) mediates the interconversion of serine and glycine and folates play a role in histidine and aromatic amino acid metabolism (Lucock, 2000; Tzin and Galili, 2010). We observed that the ΔmetF strain was not able to grow on volatiles, implying that the CysD enzyme cannot bypass the MetF dependent requirement of CH3-THF to synthesize methionine and therefore that the alternative route is not active (Figure 2B). Finally, a ΔcysBΔcysD double mutant could not grow on A. fumigatus derived or externally added volatiles, whereas the single mutant strains ΔcysB and ΔcysD did grow, although to a much lower degree (Figure 2B). This indicates that assimilation of VSCs must go through the conversion of MT into sulfide and its subsequent incorporation by CysD or CysB, and that both enzymatic actions are required for efficient assimilation of S2–. Reintroduction of the CysB encoding gene in the ΔcysBΔcysD strain reconstituted its ability to grow on sulfate and to cross-feed from VSCs, demonstrating that the growth defect of the double mutant is specific to the mutations (Supplementary Figure S3B).
Therefore, A. fumigatus can use DMS and DMDS as the sole S-sources by converting them to sulfide, which is assimilated into the metabolism. This raises the question of whether H2S may be solely responsible for the cross-feeding effect. Since our instruments cannot detect H2S we used hydrogen sulfide test strips to determine if this gas is produced by A. fumigatus and a chemical donor (Na2S) to investigate its potential to cross-feed a distal colony. We found that A. fumigatus does produce low amounts of H2S when grown on methionine, but not on sulfate (Figure 2C). However, a distal A. fumigatus colony could not cross-feed from pure sulfide, even if we attempted higher amounts of the compound and also adding it on repeated occasions (Figure 2C). We could differentiate between the absence of cross-feeding effects and the toxicity of H2S as the addition of the highest concentrations used (5 × 0.0154 mg and 5 × 0.077 mg) caused the disappearance of the background residual growth (Figure 2C).
P. aeruginosa Derived VSCs Are Exploited as S-Sources by A. fumigatus, Promoting Its Growth on Low Concentrations of Organic Sulfur
It has recently been reported that P. aeruginosa promotes A. fumigatus distal growth in vitro through the production of DMS (Briard et al., 2016). To determine if the exploitation of VSCs as a sulfur source drives this stimulatory effect, we tested the effect of P. aeruginosa VSCs on distal growth of A. fumigatus wild-type and ΔcysBΔcysD strains (Figure 3). P. aeruginosa derived VSCs triggered growth of wild-type Aspergillus, but not of the ΔcysBΔcysD mutant, on S-depleted media (Figure 3A). Actually, the growth profile of the A. fumigatus mutants was identical to that observed in Figure 2C, indicating that they are assimilated through the same route. In contrast to A. fumigatus, P. aeruginosa produced VSCs (i.e., triggered distal fungal growth) when the medium contained an excess of sulfate, but not when it contained an excess of methionine, even if the production of H2S was higher from methionine (Figure 3A). This suggests that the bacterium may be producing VSCs through a different mechanism and also supports our previous observation that sulfide cannot be directly exploited as an S-source.
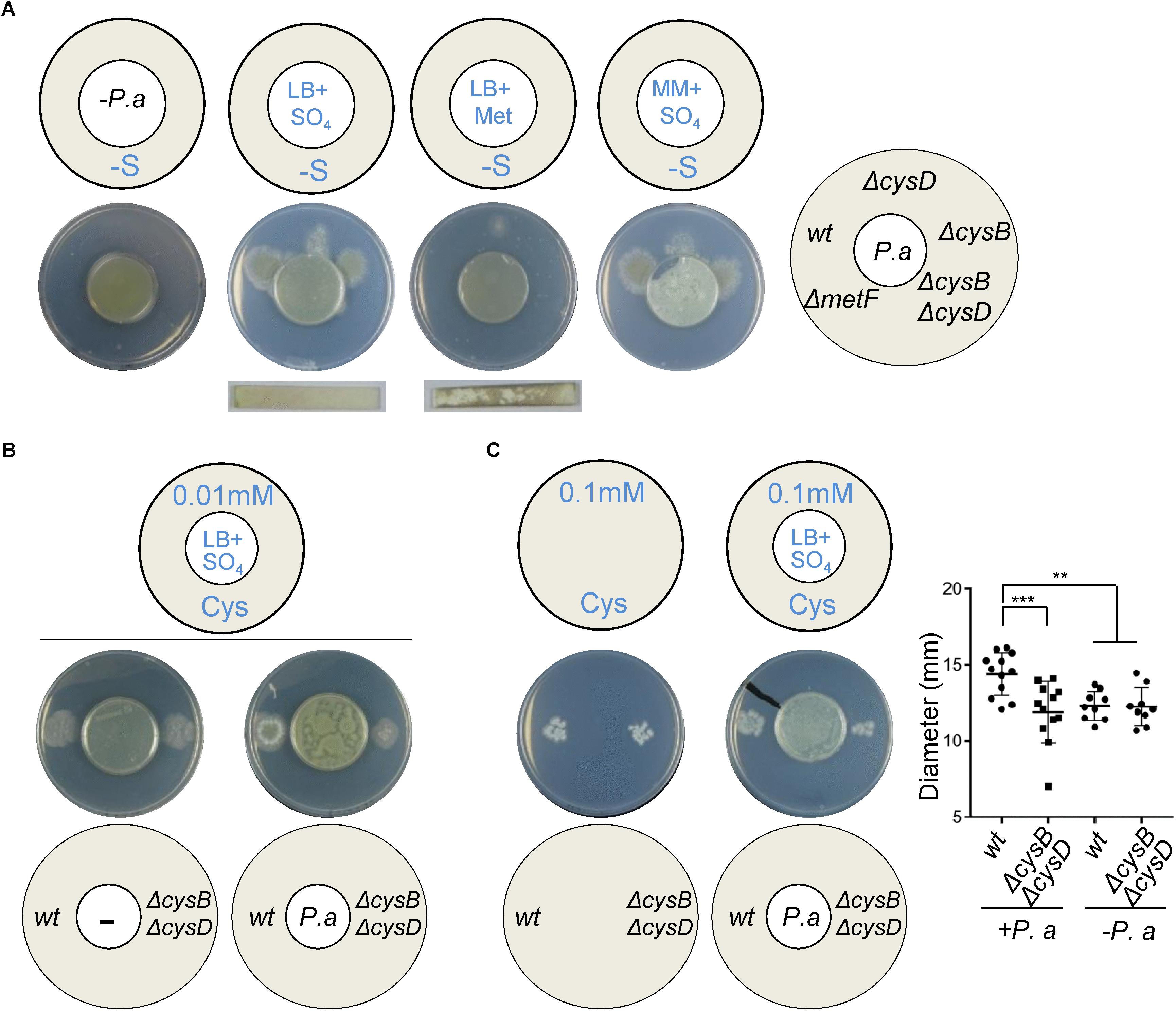
Figure 3. Pseudomonas aeruginosa derived VSCs promote A. fumigatus growth in sulfur limiting conditions. (A) When P. aeruginosa (P.a) was inoculated on a sulfate containing inner plate it was able to trigger distal growth of A. fumigatus wild-type, ΔcysB, and ΔcysD strains on the outer S-free plates. The ΔmetF and ΔcysBΔcysD strains could not utilize P. aeruginosa derived VSCs as a sulfur source. The use of H2S strips showed that P.a produced a small amount of H2S on sulfate and more on methionine, but this could not cross-feed A. fumigatus. Plates were incubated at 37°C for 3 days. (B) On media containing very low cysteine (0.01 mM) as the sole S-source, both wild-type ΔcysBΔcysD strains displayed limited growth. P. aeruginosa derived VSCs significantly enhanced A. fumigatus wild-type, not mutant, growth. Plates were incubated at 37°C for 3 days. (C) Measurement of colony diameters at 36 h post-inoculation on media containing low cysteine (0.1 mM) showed that P. aeruginosa derived VSCs significantly stimulated early A. fumigatus wild-type, not mutant, growth. Plates were incubated at 37°C for 36 h. ∗∗P < 0.01 and ∗∗∗P < 0.001.
Aspergillus fumigatus most likely encounters low concentrations of organic S-compounds to exploit as S-sources during its growth in living tissues (personal observations, reference (Amich et al., 2016) and explained in discussion). Therefore, we tested the capacity of P. aeruginosa derived VSCs to enhance fungal growth on low concentrations of cysteine. On very low cysteine (0.01 mM), both A. fumigatus wild-type and ΔcysBΔcysD strains displayed limited growth; interestingly P. aeruginosa derived VSCs significantly enhanced wild-type, but not mutant, growth (Figure 3B). On mid-low cysteine (0.1 mM) both A. fumigatus strains developed fully grown colonies at day 3 (Supplementary Figure S3C), but Pseudomonas derived volatiles exclusively stimulated early growth (day 1.5) of the wild-type strain (Figure 3C). These results show that A. fumigatus can exploit VSCs and cysteine as S-sources simultaneously; therefore the presence of VSCs can enhance fungal growth of the wild-type strain in otherwise limiting conditions.
Contrary to the positive effect of P. aeruginosa-derived VSCs on fungal growth, A. fumigatus derived VSCs from methionine catabolism did not trigger P. aeruginosa growth on an S-depleted plate (Supplementary Figure S4), indicating that P. aeruginosa cannot assimilate these volatiles.
P. aeruginosa Produces VSCs Inside Infected Galleria mellonella
To determine if P. aeruginosa produces VSCs when growing inside Galleria, we actively sampled the headspace within sealed flasks containing groups of ten infected or uninfected larvae 24 h after infection and measured it using GC-MS (Ahmed et al., 2018). We could detect a clear enrichment of several VSCs specifically in the flasks containing infected larvae, namely MT, DMS, DMDS, and dimethyl trisulfide (DMTriS) (Figures 4A,B). This demonstrates that P. aeruginosa does produce VSCs inside living Galleria during infection that can potentially be exploited by A. fumigatus during co-infection. We next tested if P. aeruginosa can trigger distal fungal growth from a homogenized Galleria (solidified with agar). Indeed P. aeruginosa inoculated on the “Galleria medium,” but not the medium alone, triggered distal A. fumigatus growth on S-depleted medium (Figure 4C). This proves that P. aeruginosa produces VSCs from the molecules contained in the Galleria organism than can then trigger distal fungal growth.
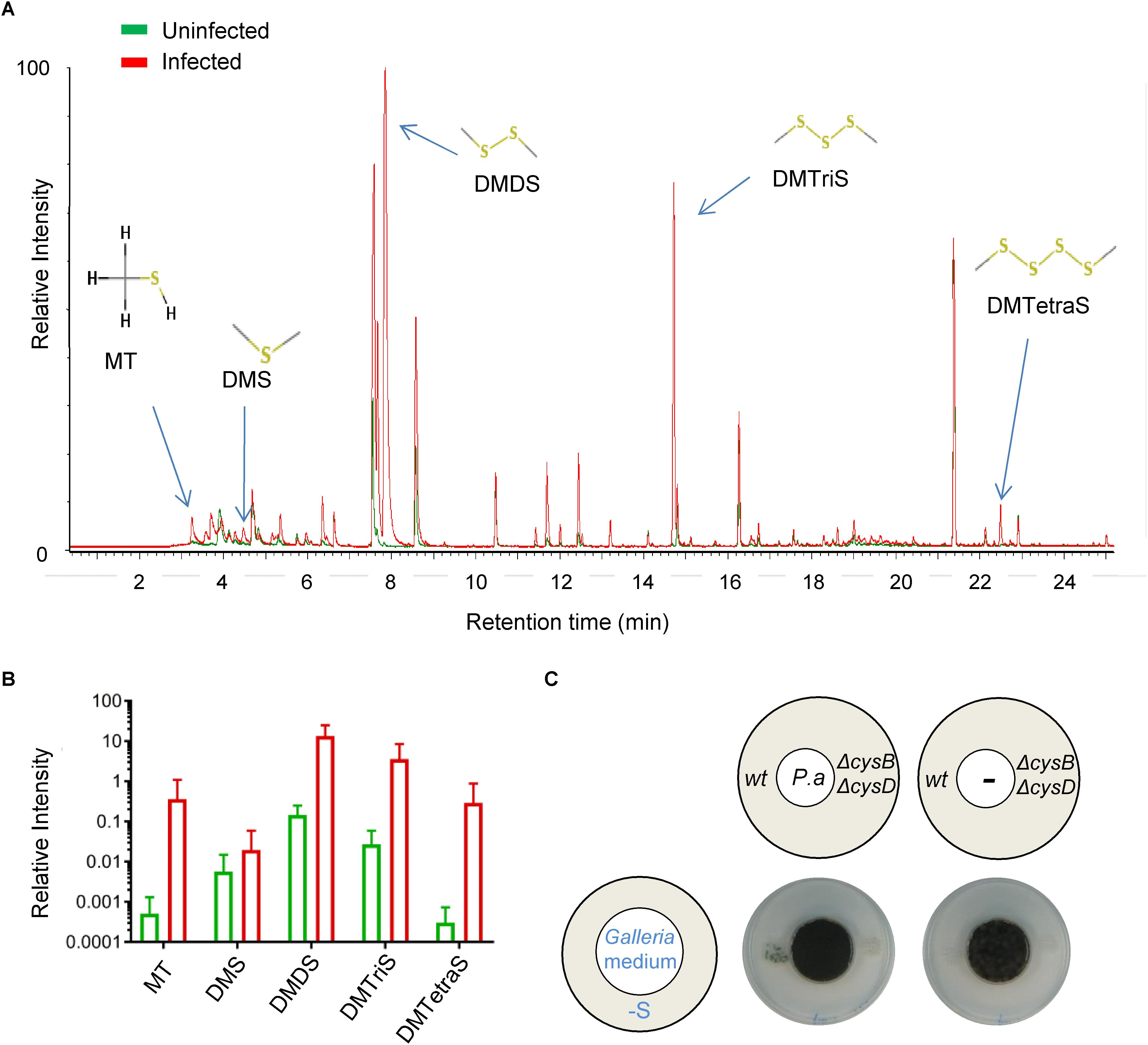
Figure 4. Pseudomonas aeruginosa produces VSCs inside infected Galleria, which are exploited as S-sources by A. fumigatus. Gas emitted from infected or uninfected Galleria accumulated in the headspace of flasks and was sampled 24 h after infection and analyzed by GC-MS. (A) Chromatogram showing an overlay of the peaks for an infected and an uninfected sample. (B) Relative normalized intensity (shown in logarithmic scale) of each of the VSCs peaks detected in two independent experiments with five biological replicates. VSCs accumulated in the flask containing infected Galleria. (C) P. aeruginosa (P. a) growing on a solid medium prepared from homogenized Galleria triggered distal A. fumigatus growth on sulfur depleted medium. Plates were incubated at 37°C for 3 days.
P. aeruginosa Derived VSCs Promote A. fumigatus Growth in vivo, Which Accounts for an Increase in Mortality
To elucidate the relevance of VSC for the in vivo A. fumigatus – P. aeruginosa interaction, we developed a low-dose co-infection model in Galleria mellonella. Titration of infectious inocula of the A. fumigatus wild-type and ΔcysBΔcysD strains determined that 50 conidia caused moderate mortality of 15% (Supplementary Figure S5A). It is notable that ΔcysBΔcysD mutant is as virulent as the wild-type (Supplementary Figure S5A), proving that A. fumigatus exploits organic S-sources inside the larvae. Titration of the infection inocula for the P. aeruginosa clinical isolate ATCC 9027 showed that ∼10–20 CFU caused ∼26.7% mortality and ∼1–5 CFU resulted in ∼100% survival (Supplementary Figure S5B). Co-infection with 50 conidia of A. fumigatus wild-type and both ∼1–5 CFU and ∼10–20 CFU of P. aeruginosa triggered a ∼45% increase in mortality compared with the single infections (Figure 5A). Thus, relative to monomicrobial infections, co-infection leads to a threefold increase in mortality. Interestingly, co-infection with 50 conidia of ΔcysBΔcysD and Pseudomonas resulted in no increase in mortality for ∼1–5 CFU and of only 19% for ∼10–20 CFU (Figure 5A), representing a maximum of only a 1.4 fold increase in mortality. Actually, co-infection with A. fumigatus wild-type was significantly more virulent than with ΔcysBΔcysD: 92.5% vs. 64.1% mortality for ∼10–20 P. aeruginosa CFU (P = 0.0046) and 73% vs. 27.1% for ∼1–5 P. aeruginosa CFU (P = 0.001). To corroborate the observed synergism, we also assayed co-infection using the P. aeruginosa PAO1 reference strain. PAO1 was similarly virulent as the clinical isolate ATCC 9027 (Supplementary Figure S5C). In agreement with our previous result, co-infection of ∼1–5 CFU Pseudomonas PAO1 with Aspergillus wild-type synergistically increased mortality (2.8 fold with respect to monomicrobial infections) whilst co-infection with A. fumigatusΔcysBΔcysD enhanced mortality to a much lesser extent (1.4 fold) (Figure 5B). Again, co-infection with Aspergillus wild-type was significantly more virulent than co-infection with the mutant (93.3% vs. 46.7%, P = 0.005). Reintroduction of the CysB encoding gene in the double mutant background (cysB + ΔcysD) reconstituted the capacity of the strain to cause highest mortality in co-infection with P. aeruginosa PAO1 (Figure 5B). These results confirm that co-infection worsens the outcome of infection (i.e., enhances mortality) and suggests that there is a synergistic effect between P. aeruginosa and A. fumigatus which is significantly dependent on the fungal exploitation of VSCs. To check if this synergy produces enhanced fungal growth, as we had observed in vitro (Figure 3B), we measured fungal burden at day 4 post-infection in single and co-infected larvae by qPCR using a published quantification method (Gago et al., 2014; Figure 5C). We observed a general tendency toward increased fungal burden (∼0.25 genome equivalents) in larvae co-infected with A. fumigatus wild-type. Taking into account that several co-infected Galleria had already succumbed, and so were not included but likely would have had high burdens, we believe this increase is relevant. We also observed a significantly increased bacterial burden in Galleria co-infected with A. fumigatus wild-type, suggesting a reciprocal synergistic effect of the fungus on P. aeruginosa (Figure 5D).
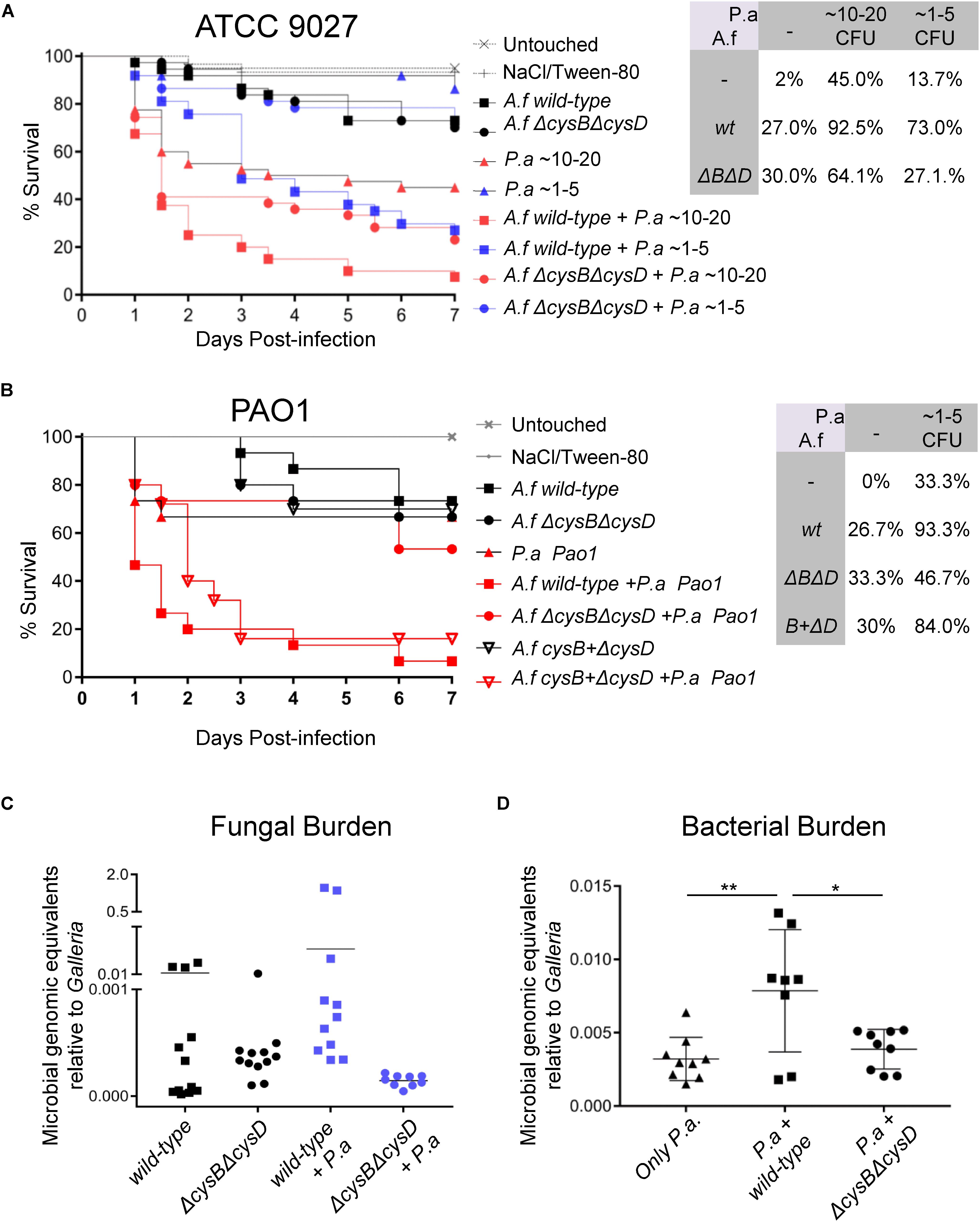
Figure 5. Pseudomonas aeruginosa derived VSCs synergize with A. fumigatus in co-infection, increasing G. mellonella mortality. (A) A G. mellonella model of infection with a low dose (50 conidia) of A. fumigatus (A. f) wild-type or ΔcysBΔcysD caused low mortality (27–30%). Infection with a low dose of P. aeruginosa ATCC 9027 caused only 13.7% mortality for 1–5 CFU or 45% for 10–20 CFU. P. aeruginosa co-infection with Aspergillus wild-type dramatically increased mortality to 73% for 1–5 CFUs and to 92.5% for 10–20 CFUs. In contrast, co-infection with the ΔcysBΔcysD mutant caused only a moderated increased in mortality to 27.1% for 1–5 CFUs and to 64.1% for 10–20 CFUs (Survival graph shows pool data of 3 independent experiments). (B) Co-infection of ∼1–5 CFU P. aeruginosa PAO1 with A. fumigatus (A. f) wild-type caused a strong increase in mortality (from 26–33% in monomicrobial infections to 93.3%, 2.8 fold). In contrast, co-infection with the A. fumigatus ΔcysBΔcysD mutant caused a much more moderate increase in mortality (to 46.7%, 1.4 fold). Reintroduction of the cysB gene in the ΔcysBΔcysD mutant (cySB + ΔcysD) reconstituted the synergism of mortality of con-infection (84%) (Survival graph shows pool data of 3 independent experiments). (C) Four days after infection, Galleria co-infected with A. fumigatus wild-type and P. aeruginosa ATCC 9027 showed a tendency toward having an increased fungal burden and (D) had significantly higher bacterial burdens. ∗∗P < 0.05 and ∗∗∗P < 0.01.
P. aeruginosa Produces VSCs From Artificial Sputum Medium (ASM) and From Mucins
To better mimic the conditions that the pathogens encounter in the airways of CF patients, we used a partially defined medium with a composition that closely resembles CF sputum (Kirchner et al., 2012). Both P. aeruginosa wild-type strains ATCC 9027 and PAO1 were able to grow on ASM and produced VSCs that triggered A. fumigatus wild-type, but not ΔcysBΔcysD, distal growth on S-depleted media (Figure 6A). A. fumigatus was able to grow on ASM, but did not produce VSCs unless methionine was supplemented to the medium (Figure 6A). Mucins are cysteine rich proteins and, accordingly, both A. fumigatus wild-type and ΔcysBΔcysD strains could exploit them as the sole S-source (Figure 6B), but volatiles were not derived (Figure 6C). In contrast, P. aeruginosa could produce VSCs from mucins that triggered growth of the distally inoculated A. fumigatus wild-type strain. Taken together, these results show that P. aeruginosa produces VSCs from mucins contained in ASM that can stimulate A. fumigatus growth, suggesting that bacterial utilization of mucins may release an additional exploitable sulfur source for the fungus.
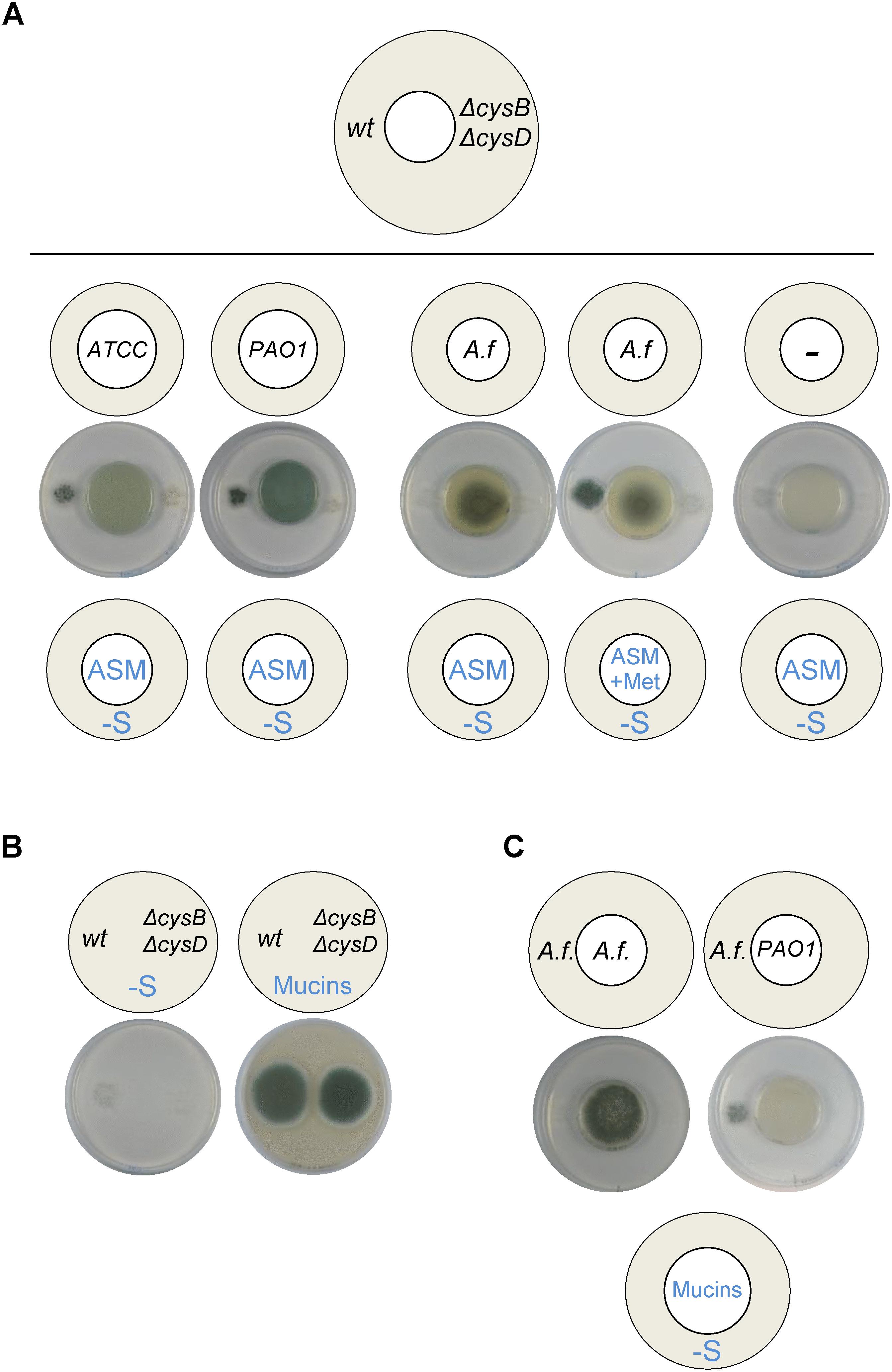
Figure 6. Pseudomonas aeruginosa produces VSCs from artificial sputum medium (ASM) and from mucins. (A) Growth of P. aeruginosa ATCC 9027 and PAO1 strains on ASM triggered distal A. fumigatus (A. f) growth. A. fumigatus needed supplementation of methionine to the ASM medium to trigger distal growth (i.e., to produce VSCs). (B) A. fumigatus wild-type and ΔcysBΔcysD strains could exploit mucins as the sole sulfur source. (C) When grown on mucins as the sole S-source wild-type P. aeruginosa, but not A. fumgiatus, produced VSCs, which triggered distal A. fumigatus growth. Plates were incubated at 37°C for 3 days.
P. aeruginosa Can Cross-Feed A. fumigatus When Growing on Murine Lung Tissue and on Human Bronchial Epithelial Cells
Aiming to determine if the observed synergistic effect may be relevant for co-infection in humans, we tested the capacity of P. aeruginosa to trigger distal A. fumigatus growth when inoculated on substrates that replicate the lung. We first inoculated P. aeruginosa directly on explanted diced murine lungs and observed that indeed bacterial growth on the lung tissues could trigger growth of distal A. fumigatus wild type, but not ΔcysBΔcysD, on S-depleted media (Figure 7). A. fumigatus was able to grow on diced lungs, but its growth did not cross-feed a distally inoculated colony. We then inoculated P. aeruginosa on human bronchial epithelial cells 16HBE and observed that the bacteria is also able to trigger distal A. fumigatus wild-type, not ΔcysBΔcysD, growth when feeding exclusively from human cells (Figure 7). A. fumigatus growth directly on cells was quite limited and did not cross-feed the distal colony. In conclusion, the cross-feeding effect took place when P. aeruginosa grew on substrates that mimic the airways, indicating that the observed synergism is likely to take place in human co-infection.
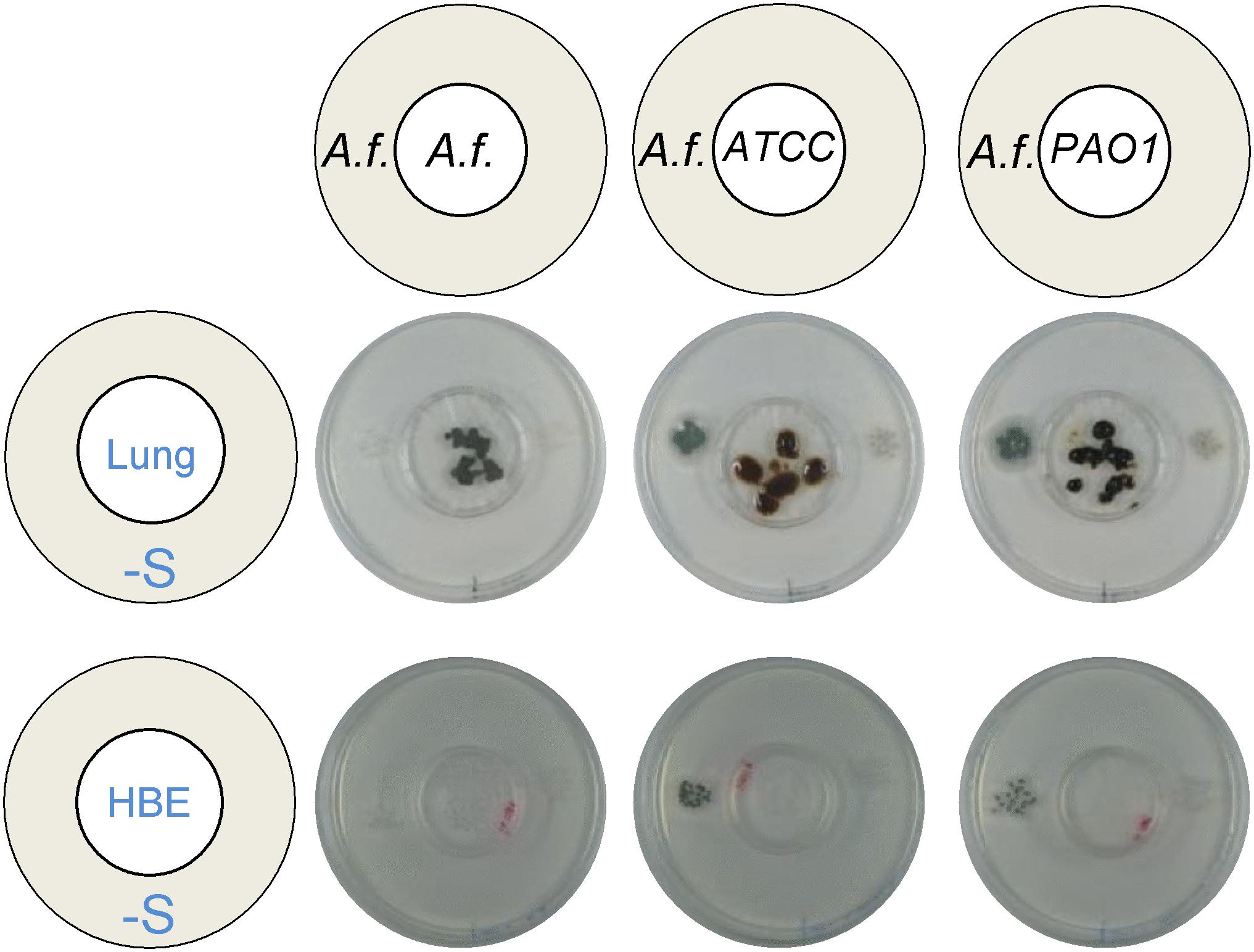
Figure 7. Pseudomonas aeruginosa produces VSCs from substrates that replicate the lung. Both A. fumigatus (A. f) and P. aeruginosa ATCC 9027 and PAO1 strains could grow directly on explanted diced murine lungs and on human bronchial epithelial cells (16-HBE), but only P. aeruginosa produced volatiles that triggered distal growth of A. fumigatus on a sulfur-free plate. Plates were incubated at 37°C for 3 days.
Discussion
The relevance of pathogen-pathogen interactions in polymicrobial infections is being more and more recognized (Singer, 2010; Peters et al., 2012). However, A. fumigatus – P. aeruginosa interactions in co-infection are largely unknown. Several in vitro studies have reported negative effects of P. aeruginosa on A. fumigatus (Kerr et al., 1999; Mowat et al., 2010; Penner et al., 2016; Shirazi et al., 2016; Briard et al., 2017; Sass et al., 2018). However, clinical evidence suggests the opposite, indicating a synergistic association in vivo. For instance, a longitudinal study in adolescents with CF found that A. fumigatus was the only species associated with an increased risk for infection with P. aeruginosa (Hector et al., 2016) and another recent study showed that P. aeruginosa infection in CF patients was associated with higher incidence of subsequent Aspergillus infection (Granchelli et al., 2018). In addition, antibiotic treatment targeting Pseudomonas has been observed to reduce Aspergillus’ presence in CF patient’s sputum (Baxter et al., 2013b). Most of the negative effects described in vitro were observed when setting the pathogens in direct contact. However, given the size of the human lungs, we speculate that the organisms likely occupy physically separated niches, and therefore the action of long distance acting molecules should be very relevant. Surprisingly, despite the increasing number of studies investigating pathogen-pathogen interactions, the role of volatiles in intermicrobial cross-talk in vivo has been largely overlooked. Recently it has been described that the Pseudomonas derived volatile sulfur compound DMS promotes A. fumigatus growth in vitro (Briard et al., 2016). In this study we have scrutinized the pathway for VSCs assimilation in A. fumigatus and explored the hypothesis that Pseudomonas derived VSCs could be utilized by A. fumigatus in vivo, producing a synergistic effect in co-infection.
We have shown that A. fumigatus assimilates VSCs through the action of CysB (cysteine synthase) or CysD (homocysteine synthase). The double mutant ΔcysBΔcysD cannot feed from inorganic sources (nor incorporate VSCs), but is completely virulent in both the G. mellonella and leukopenic murine models of infection (Supplementary Figures S5A, S6A). This strongly suggests that A. fumigatus predominately feeds from organic S-sources within the tissues of both the larvae and the mammalian lung. We have shown that in the Galleria model, co-infection of P. aeruginosa with wild-type A. fumigatus enhances virulence to a greater extent than co-infection with the ΔcysBΔcysD strain. It has been recently described that co-infection triggers a more pronounced pro-inflammatory response (Bergeron et al., 2017); indeed Bergeron et al. (2017) found a stronger immune response in Candida albicans – P. aeruginosa co-infection, which they proposed could be the reason of the observed increment in virulence and mortality. Therefore, it could be argued that in our model a stronger inflammatory response accounts for the increased mortality of co-infection, and that the double mutant is more sensitive to such hyperinflammation. However, a conidiocidal assay using human neutrophilic granulocytes showed that neutrophils kill conidia of the wild-type and double mutant strains similarly (Supplementary Figure S6B), indicating that the ΔcysBΔcysD strain is not more susceptible to attack by immune effector cells. Therefore, although a stronger pro-inflammatory response is likely to play a role in the pathology of co-infection, it does not explain the differences found between the A. fumigatus wild-type and mutant strains. We have shown that P. aeruginosa produces VSCs in vivo, which translates into an increased fungal burden in G. mellonella larvae co-infected with A. fumigatus wild-type, but not ΔcysBΔcysD. Interestingly, we have also observed an enhanced bacterial burden in larvae co-infected with wild-type, but not the mutant strain, Aspergillus. It has been reported that A. fumigatus enhances P. aeruginosa production of the virulence factor elastase (Smith et al., 2015). Similarly, it has been shown that C. albicans triggers production of P. aeruginosa virulence factors, which was suggested, is the leading cause of mortality in co-infection (Trejo-Hernandez et al., 2014). Consequently, we hypothesize that Pseudomonas derived VSCs enhance fungal growth, which then somehow stimulates bacterial growth and pathogenesis, contributing to an increase in the mortality of co-infection. Very recently Reece et al. (2018) have reported that P. aeruginosa causes increased mortality in G. mellonella when infected 24 h after a challenge with a non-lethal A. fumigatus dose. The authors proposed that this effect is likely due to one of two options: either the fungus is producing immunoregulatory toxins that suppress the immune system of G. mellonella, making them more susceptible to P. aeruginosa infection or the two species interaction causes a more virulent infection. Even if we believe they are not mutually exclusive and toxins may also play a role, our results support the second hypothesis: inoculated P. aeruginosa produces VSCs, which increases A. fumigatus growth, and the presence of the fungus enhances bacterial pathogenicity, all resulting in higher mortality. Interestingly, the authors assayed various A. fumigatus and P. aeruginosa isolates (reference strains and patient isolates) and observed different effects depending on the strain combination of co-infection. Although we have not made any attempt to address this, we speculate it might be partially explained by subtle differences in the volatile production capacities of the P. aeruginosa isolates, and/or their responses to interactions that occur in vivo.
We have observed in vitro that P. aeruginosa produces VSCs in the presence of sulfate. It has been shown that P. aeruginosa can feed from sulfate present in human respiratory tracts, and that it can also use sulfate contained within mucins as an S-source (Robinson et al., 2012). We have shown that it produces VSCs when grown on Artificial Sputum Medium and on mucins. G. mellonella hemolymph contains mucin-like proteins (Li et al., 2002), which we speculate the bacteria is exploiting as an S-source, which triggers VSCs production inside the infected larvae. It had previously been described that A. fumigatus is able to degrade mucins (Leger and Screen, 2000), and we have observed that the fungus can use them as a sulfur source in vitro. This is most likely due to the utilization of the enriched amino acid cysteine as the S-source, as suggested by the capacity of the ΔcysBΔcysD mutant to grow on mucins and the inability of the wild-type to produce VSCs. However, the gene(s) required for utilization of the sulfate contained within the mucins (which could be one or several of the three arylsulphatases encoded by its genome: AFUA_3G02290, AFUA_5G12940, and AFUA_8G02520) may not be expressed in the presence of cysteine (Amich et al., 2013), impeding its exploitation as an additional S-source. Therefore, we propose that during co-infection P. aeruginosa liberates an otherwise non-accessible sulfur source in the form of VSCs, which in turns makes the tissues less sulfur limiting for the fungus and promotes its growth. An alternative explanation could be that P. aeruginosa liberates and/or releases sulfate or other inorganic sulfur sources, which could be assimilated by A. fumigatus wild-type but not ΔcysBΔcysD. Although we cannot reject this possibility, we believe it is unlikely because the growth promoting effect would be restricted to the proximity of bacterial cells and also because the genes of the sulfate reduction assimilation pathway (required to assimilate inorganic S-sources) are downregulated in the presence of organic S-compounds (Amich et al., 2013).
Interestingly, we show that when P. aeruginosa grows on substrates that resemble the lung, namely explanted murine lungs and human bronchial epithelial cells, it can trigger distal A. fumigatus growth. A recent analysis of bronchoalveolar lavage fluid of CF patients detected sulfur-containing volatiles specifically in P. aeruginosa infected patients (Nasir et al., 2018). Therefore, it seems plausible that P. aeruginosa derived VSCs are able to trigger the synergistic effect described in this manuscript and be relevant for co-infection in the human airways.
In conclusion, we report a volatile dependent synergistic effect of Pseudomonas aeruginosa on A. fumigatus during co-infection of G. mellonella that causes higher fungal and bacterial burdens, resulting in increased mortality. This effect is likely relevant for the pathogen-pathogen interaction in patients’ lungs and thus has an impact on the outcome of co-infection.
Materials and Methods
Strains, Media and Culture Conditions
The Escherichia coli strain DH5α (Woodcock et al., 1989) was used for cloning procedures. Plasmid-carrying E. coli strains were grown at 37°C in LB liquid medium under selective conditions (100 μg⋅mL–1 ampicillin, 50 μg⋅mL–1 kanamycin or 10 μg⋅mL–1 gentamycin); Media was solidified with 1.5% agar. All plasmid constructs used in the course of this study are listed in Supplementary Table S1 and were generated using the Seamless Cloning (Invitrogen) technology as previously described (Amich et al., 2013, 2016).
The wild-type clinical isolate A. fumigatus strain ATCC 46645 served as reference recipient; gene knock-out derivative strains of this isolate were generated using a standard protoplasting protocol (Szewczyk et al., 2006). Once verified by Southern-blot, the mutants were grown on xylose to activate the encoded beta-recombinase, which self-excise the selection marker, a strategy previously described (Amich et al., 2013, 2016) (Supplementary Figure S7). Reversion of the cysB gene was directed to its natural locus using homology flanking regions. Mutants were selected by reconstitution of ability to grow on sulfate as sole sulfur source and checked by PCR. A. fumigatus strains were generally cultured on minimal medium (MM) (Amich et al., 2016) at 37°C. For selection in the presence of resistance markers 50 μg⋅mL–1 of hygromycin B or 100 μg⋅mL–1 of pyrithiamine (InvivoGen) was applied. In sulfur-free medium, MgCl2 substituted for MgSO4, and a modified mixture of trace elements lacking any sulfate salt was used. Artificial sputum medium (ASM) was prepared as previously described (Kirchner et al., 2012) and solidified with 1.5% agar.
Pseudomonas aeruginosa wild-type clinical isolate ATCC 9027 and reference strain PAO1 were used for cross-feeding experiments and G. mellonella infections. Strains were routinely grown on LB, which was supplemented with 5 mM methionine or 2 mM Na2SO4 for some experiments. “Galleria medium” was prepared by homogenizing a larva with sterile ultrapure water on a 70 μm cell-strainer (Falcon) with the use of a syringe plunger. This homogenate was mixed with agar (to 1.5% final concentration), solidified, and illuminated with UV light for 10 min to reduce bacterial contamination. Murine lungs were explanted from uninfected controls at the end of a different experiment. 16HBE cells (Cozens et al., 1994) were seeded in coated plates (NunclonTM Delta, ThermoFisher Scientific) with Dulbecco’s Modified Eagle Medium (DMEM, Gibco®) at a very high density (>100% confluency), left overnight to adhere, and then the medium was removed to inoculate P. aeruginosa directly on the cells. For G. mellonella infection, one single colony of an overnight grown plate was suspended in 1 mL NaCl 0.9%/Tween-20 0.002%. This solution was diluted serially to yield the desired concentrations, as confirmed each time by plating the dilutions on LB.
For cross-feeding experiments one colony of P. aeruginosa or 103 conidia of A. fumigatus were inoculated in the internal plate, and 102 conidia of each A. fumigatus strain in the external petri dish. Unless otherwise stated, plates were incubated at 37°C for 3 days inside zip-closed plastic bags. Na2S, DMS, and DMDS were purchased from Sigma and the amounts described in each experiment were added directly to Whatman filter paper placed inside the inner plate.
Extraction and Manipulation of Nucleic Acids
Standard protocols of recombinant DNA technology were carried out (Sambrook et al., 1989). Phusion® high-fidelity DNA polymerase (Thermo Fisher Scientific) was generally used in polymerase chain reactions and essential cloning steps were verified by sequencing. Fungal genomic DNA was prepared following the protocol of Kolar et al. (1988) and Southern analyses were carried out as described (Southern, 1975, 2006), using the Amersham ECL Direct Labeling and Detection System® (GE Healthcare).
Detection of Volatile Sulfur Compounds Derived From Aspergillus fumigatus in vitro
Volatile sulfur compounds measurement was performed as described (Briard et al., 2016). Briefly, 106 conidia per ml A. fumigatus spores were inoculated in 5 mL of sulfur free MM with 2 mM Na2SO4 or 5 mM methionine as sulfur source and incubated in 40 mL vials with screw caps with a rubber/PTFE septum for 3 days at 37°C. Direct sampling was performed in the headspace of culture vials using SPME fibers [2 cm composite coating carboxen/divinylbenzene/polydimethylsiloxane (Car/DVB/PDMS)]. Vial septa were pierced with the needle of the filament holder followed by fiber exposure for 30 min in the incubator at 37°C. Volatiles were injected into Agilent HP-5MS GC-MS column (length 30 m, internal diameter 0.25 mm, and film thickness 0.25 μm) using a Front SS Inlet He in Pulsed Splitless mode.
MT Oxidase Activity
Aspergillus fumigatus protein extracts were isolated from ground mycelia in lysis buffer (50 mM HEPES; 1% NP-40; 0.5 mM EDTA; 0.1% SDS; 1mM DTT) with 1× complete protease inhibitor cocktail (Roche). 500 μM methanethiol was added or not to 0.5 mg protein extracts in 200 μl of lysis buffer and incubated for 6 h at 37°C in sealed tubes. Formaldehyde was measured using Purplad® reagent (Sigma) as previously described (Boden et al., 2011). Briefly, 100 μl of a 34 mM dilution of Purpald® in 2 M NaOH was added to the tubes, incubated for 15 min and OD measured at 550 nm. The increment in absorbance was normalized with lysis buffer and converted to μM of formaldehyde using a standard curve.
Galleria mellonella Infection
Sixth-stage instar larval G. mellonella moths (15 to 25 mm in length) were ordered from the Live Foods Company (Sheffield, United Kingdom). Infections were performed according to Kavanagh and Fallon (2010). Randomly selected groups of 15 larvae were injected with 20 μl of NaCl 0.9%/Tween-20 0.002% containing varying doses of A. fumigatus conidia or P. aeruginosa CFUs, as detailed in each experiment. A. fumigatus was always injected into the last right proleg, whilst P. aeruginosa was always injected in the left proleg. Braun Omnican 50-U 100 0.5-mL insulin syringes with integrated needles were used. In each experiment an untouched and a saline injected control were included, to verify that mortality was not due to the health status of the larvae or the injection method.
Detection of Volatile Sulfur Compounds Derived From Pseudomonas aeruginosa in Infected Galleria
The headspace of flasks containing 10 G. mellonella uninfected or infected with P. aeruginosa (5 flasks/group) were sampled after a 24 h incubation at 37°C, using a method described previously (Ahmed et al., 2018). Briefly, a total volume of 200 mL was sampled from a 100 mL glass flask containing Galleria, at a flow of 100 mL/min, onto conditioned silco-treated stainless steel multi-sorbent tubes packed TenaxTA/Carbograph5TD. This sorbent bed was chosen to maximize the range of Volatile Organic Compounds (VOCs) captured, whilst minimizing water retention. Sorbent tubes were stored at 4°C before analysis, for up to 7 days.
Volatile organic compounds trapped onto the sorbent bed were analyzed using thermal desorption-gas chromatography-mass spectrometry (TD-GC-MS). In summary, sorbent tubes underwent a two-stage thermal desorption process where the initial desorption was carried out at 220°C for 5 min. Prior to this, an internal standard (1 ppmV 4-bromoflorobenzene in N2) was injected onto each tube. VOCs were purged off the tube using N2 and onto a cold trap (kept at 0°C), which was subsequently desorbed at 230°C for 3 min. VOCs were then transferred to the GC column at a constant flow of 2 mL/min using He as the carrier gas. The Agilent DB-1ms GC column (length 60 m, internal diameter 0.25 mm, and film thickness 0.5 μm) was housed within an Agilent 6890 GC oven (Agilent Technologies, West Lothian, United Kingdom) which underwent a ramped programs with an initial 40°C temperature (no hold), ramped to 250°C (18 min hold) at 5°C/min. The total GC cycle time was 60 min. After separation by GC, VOCs were transferred to a Waters GCT mass spectrometer (Waters Corporation, Manchester, United Kingdom), equipped with an EI + source (70 eV), and a time-of-flight mass analyzer. Data were acquired between m/z 29–400 in centroid mode, at a rate of 10 scans/s.
Data were analyzed by using Target Lynx (Masslynx software version 4.1, Waters Corporation, Manchester, United Kingdom). The exact masses of base peaks m/z 47, 62, 94, 126, and 158 were used to extract methanethiol, dimethyl sulfide, dimethyl disulfide, DMTriS, and dimethyl tetrasulfide, respectively, along with retention indices calculated using alkane chemical standards (C5–C15). Data were pre-processed using the Apex Trak algorithm (Waters Corporation, Manchester, United Kingdom) which included peak integration, smoothing and baseline correction. Data were then normalized to the internal standard (with an extracted base peak of m/z 174). DMS and DMDS were identified to MSI level 1 standard (Fiehn et al., 2007), a NIST library search was performed on the sample peak, together with retention time matching of their counterpart external standards. Other VSCs were identified to MSI level 2 i.e., NIST library search only.
Neutrophil Killing Assay
Human neutrophils were isolated from healthy donors’ blood using the EasySepTM Direct Human Neutrophil Isolation Kit (StemCell) following the manufacturer’s instructions for EasystepTM Magnets EasyEightsTM protocol. Purified neutrophils were set in culture in RPMI medium (Sigma) supplemented with 10% human serum (isolated from the same donor’s blood by centrifugation at 1200 × g 4 min), and conidia were added on top at a multiplicity of infection (MOI) ratio of 1:10. After 6 h, medium was collected and centrifuged at 4,000 rpm for 5 min; the pellet was resuspended in distilled water and finally resuspended in 1 ml 0.9% NaCl plus 0.01% Tween 20. Different dilutions were plated onto MM + cysteine in triplicates, and colony forming units were counted. For input controls, the same amount of conidia were incubated in RPMI + serum for 6 h without neutrophils and treated and plated as the experimental conditions.
Leukopenic Murine Model of Pulmonary Aspergillosis
The experiment was performed under United Kingdom Home Office project license PPL70/7324 and approved by the University of Manchester Ethics Committee. Outbred CD1 male mice (22–26 g) were purchased from Charles Rivers and left to rest for at least 1 week before the experiment. Mice were allowed access ad libitum to water and food throughout the experiment. Mice were immunosuppressed with 150 mg/kg of cyclophosphamide on days −3 and −1 and with 112 mg/kg cortisone acetate on day −1. On day 0 mice were anesthetized with isofluorane and intranasally infected with a dose of 105 conidia (40 μl of a freshly harvested spore solution of 2.5 × 106 conidia/mL). Immunosuppression was maintained by injecting 150 mg/kg cyclophosphamide every 3 days (days + 2 and + 5) and survival was followed for 7 days.
Statistical Analyses
All graphs were prepared using GraphPad Prism v7.04 software. For statistical analysis of survival curves the Log-Rank test was applied. In column graphs one-way ANOVA or Kruskal-Wallis (depending on the distribution) were utilized to determine differences among groups. Significance was considered when ∗P < 0.05, ∗∗P < 0.01, and ∗∗∗P < 0.001.
Data Availability Statement
Datasets for GC/MS experiments are available upon reasonable request to the corresponding author. All other datasets are included in the manuscript/Supplementary Files.
Ethics Statement
The animal study was reviewed and approved by the UK Home Office project license PPL70/7324 and approved by the University of Manchester Ethics Committee.
Author Contributions
JS performed the majority of the experiments and wrote the manuscript. MS-O helped JS with most of the experiments. WA performed and analyzed the in vivo measurements of VSCs. CH performed and analyzed the in vitro measurements of VSCs. CZ helped with the acquisition and analysis of volatile measurements. RT provided the support with Galleria experiments. MB supported the design of in vivo volatile experiments and revised the manuscript. J-PL supported the design in vitro volatile experiments and revised the manuscript. SK participated in the design of cross-feeding experiments and revised the manuscript. SF devised the volatile measurements and revised the manuscript. EB participated in the conception of the study and contributed to manuscript writing. JA designed the project and wrote the manuscript.
Funding
JA is recipient of a MRC Career Development Award (MR/N008707/1).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are indebted to Dr. Zorana Carter for inspiring discussions. Thanks are due to Dr. Pavlos Geranios for valuable advice with the sampling of volatiles from infected Galleria and Dr. Iain R. White for assisting with the TD-GC-MS analysis. We thank Dr. Matt Fletcher and Dr. Stephan Heeb for providing pME6032Gm plasmid. Help and encouragement from all members of the MFIG is highly appreciated.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2019.02311/full#supplementary-material
References
Ahmed, W. M., Geranios, P., White, I. R., Lawal, O., Nijsen, T. M., Bromley, M. J., et al. (2018). Development of an adaptable headspace sampling method for metabolic profiling of the fungal volatome. Analyst 143, 4155–4162. doi: 10.1039/c8an00841h
Ahmed, W. M., Lawal, O., Nijsen, T. M., Goodacre, R., and Fowler, S. J. (2017). Exhaled volatile organic compounds of infection: a systematic review. ACS Infect. Dis. 3, 695–710. doi: 10.1021/acsinfecdis.7b00088
Amich, J., Dumig, M., O’Keeffe, G., Binder, J., Doyle, S., Beilhack, A., et al. (2016). Exploration of sulfur assimilation of Aspergillus fumigatus reveals biosynthesis of sulfur-containing amino acids as a virulence determinant. Infect. Immun. 84, 917–929. doi: 10.1128/IAI.01124-15
Amich, J., Schafferer, L., Haas, H., and Krappmann, S. (2013). Regulation of sulphur assimilation is essential for virulence and affects iron homeostasis of the human-pathogenic mould Aspergillus fumigatus. PLoS Pathog. 9:e1003573. doi: 10.1371/journal.ppat.1003573
Amin, R., Dupuis, A., Aaron, S. D., and Ratjen, F. (2010). The effect of chronic infection with Aspergillus fumigatus on lung function and hospitalization in patients with cystic fibrosis. Chest 137, 171–176. doi: 10.1378/chest.09-1103
Bakare, N., Rickerts, V., Bargon, J., and Just-Nubling, G. (2003). Prevalence of Aspergillus fumigatus and other fungal species in the sputum of adult patients with cystic fibrosis. Mycoses 46, 19–23. doi: 10.1046/j.1439-0507.2003.00830.x
Baxter, C. G., Dunn, G., Jones, A. M., Webb, K., Gore, R., Richardson, M. D., et al. (2013a). Novel immunologic classification of aspergillosis in adult cystic fibrosis. J. Allergy Clin. Immunol. 132, 560–566.e10. doi: 10.1016/j.jaci.2013.04.007
Baxter, C. G., Rautemaa, R., Jones, A. M., Webb, A. K., Bull, M., Mahenthiralingam, E., et al. (2013b). Intravenous antibiotics reduce the presence of Aspergillus in adult cystic fibrosis sputum. Thorax 68, 652–657. doi: 10.1136/thoraxjnl-2012-202412
Bellesia, F., Pinetti, A., Bianchi, A., and Tirillini, B. (1996). Volatile compounds of the white truffle (Tuber magnatum Pico) from middle Italy. Flavour Fragr. J. 11, 239–243. doi: 10.1002/(sici)1099-1026(199607)11:4<239::aid-ffj573>3.0.co;2-a
Bergeron, A. C., Seman, B. G., Hammond, J. H., Archambault, L. S., Hogan, D. A., and Wheeler, R. T. (2017). Candida albicans and Pseudomonas aeruginosa interact to enhance virulence of mucosal infection in transparent zebrafish. Infect. Immun. 85:e00475-17. doi: 10.1128/IAI.00475-17
Boden, R., Borodina, E., Wood, A. P., Kelly, D. P., Murrell, J. C., and Schafer, H. (2011). Purification and characterization of dimethylsulfide monooxygenase from Hyphomicrobium sulfonivorans. J. Bacteriol. 193, 1250–1258. doi: 10.1128/JB.00977-10
Bolten, C. J., Schroder, H., Dickschat, J., and Wittmann, C. (2010). Towards methionine overproduction in Corynebacterium glutamicum–methanethiol and dimethyldisulfide as reduced sulfur sources. J. Microbiol. Biotechnol. 20, 1196–1203. doi: 10.4014/jmb.1002.02018
Brandt, C., Roehmel, J., Rickerts, V., Melichar, V., Niemann, N., and Schwarz, C. (2018). Aspergillus bronchitis in patients with cystic fibrosis. Mycopathologia 183, 61–69. doi: 10.1007/s11046-017-0190-0
Briard, B., Heddergott, C., and Latge, J. P. (2016). Volatile compounds emitted by Pseudomonas aeruginosa stimulate growth of the fungal pathogen Aspergillus fumigatus. mBio 7:e00219. doi: 10.1128/mBio.00219-16
Briard, B., Rasoldier, V., Bomme, P., ElAouad, N., Guerreiro, C., Chassagne, P., et al. (2017). Dirhamnolipids secreted from Pseudomonas aeruginosa modify anjpegungal susceptibility of Aspergillus fumigatus by inhibiting beta1,3 glucan synthase activity. ISME J. 11, 1578–1591. doi: 10.1038/ismej.2017.32
Burgel, P. R., Paugam, A., Hubert, D., and Martin, C. (2016). Aspergillus fumigatus in the cystic fibrosis lung: pros and cons of azole therapy. Infect. Drug Resist. 9, 229–238. doi: 10.2147/idr.s63621
Cozens, A. L., Yezzi, M. J., Kunzelmann, K., Ohrui, T., Chin, L., Eng, K., et al. (1994). CFTR expression and chloride secretion in polarized immortal human bronchial epithelial cells. Am. J. Respir. Cell Mol. Biol. 10, 38–47. doi: 10.1165/ajrcmb.10.1.7507342
De Bont, J. A. M., van Dijken, J. P., and Harder, W. (1981). Dimethyl sulphoxide and dimethyl sulphide as a carbon, sulphur and energy source for growth of Hyphomicrobium S. J. Gen. Microbiol. 127, 315–323. doi: 10.1099/00221287-127-2-315
Fiehn, O., Robertson, D., Griffin, J., van der Werf, M., Nikolau, B., Morrison, N., et al. (2007). The metabolomics standards initiative (MSI). Metabolomics 3, 175–178. doi: 10.1007/s11306-007-0070-6
Gago, S., Buitrago, M. J., Clemons, K. V., Cuenca-Estrella, M., Mirels, L. F., and Stevens, D. A. (2014). Development and validation of a quantitative real-time PCR assay for the early diagnosis of coccidioidomycosis. Diagn. Microbiol. Infect. Dis. 79, 214–221. doi: 10.1016/j.diagmicrobio.2014.01.029
Gould, W. D., and Kanagawa, T. (1992). Purification and properties of methyl mercaptan oxidase from Thiobacillus-Thioparus Tk-M. J. Gen. Microbiol. 138, 217–221. doi: 10.1099/00221287-138-1-217
Granchelli, A. M., Adler, F. R., Keogh, R. H., Kartsonaki, C., Cox, D. R., and Liou, T. G. (2018). Microbial interactions in the cystic fibrosis airway. J. Clin. Microbiol. 56:e00354-18. doi: 10.1128/JCM.00354-18
Hector, A., Kirn, T., Ralhan, A., Graepler-Mainka, U., Berenbrinker, S., Riethmueller, J., et al. (2016). Microbial colonization and lung function in adolescents with cystic fibrosis. J. Cyst. Fibros. 15, 340–349. doi: 10.1016/j.jcf.2016.01.004
Kavanagh, K., and Fallon, J. P. (2010). Galleria mellonella larvae as models for studying fungal virulence. Fungal Biol. Rev. 24, 79–83. doi: 10.1016/j.fbr.2010.04.001
Kerr, J. R., Taylor, G. W., Rutman, A., Hoiby, N., Cole, P. J., and Wilson, R. (1999). Pseudomonas aeruginosa pyocyanin and 1-hydroxyphenazine inhibit fungal growth. J. Clin. Pathol. 52, 385–387. doi: 10.1136/jcp.52.5.385
Kirchner, S., Fothergill, J. L., Wright, E. A., James, C. E., Mowat, E., and Winstanley, C. (2012). Use of artificial sputum medium to test antibiotic efficacy against Pseudomonas aeruginosa in conditions more relevant to the cystic fibrosis lung. J. Vis. Exp. 5:e3857. doi: 10.3791/3857
Kolar, M., Punt, P. J., van den Hondel, C. A., and Schwab, H. (1988). Transformation of Penicillium chrysogenum using dominant selection markers and expression of an Escherichia coli lacZ fusion gene. Gene 62, 127–134. doi: 10.1016/0378-1119(88)90586-0
Labows, J. N., McGinley, K. J., Webster, G. F., and Leyden, J. J. (1980). Headspace analysis of volatile metabolites of Pseudomonas aeruginosa and related species by gas chromatography-mass spectrometry. J. Clin. Microbiol. 12, 521–526.
Leger, R. J. S., and Screen, S. E. (2000). In vitro utilization of mucin, lung polymers, plant cell walls and insect cuticle by Aspergillus fumigatus, metarhizium anisopliae and haematonectria haematococca. Mycol. Res. 104, 463–471. doi: 10.1017/s0953756299001525
Li, D., Scherfer, C., Korayem, A. M., Zhao, Z., Schmidt, O., and Theopold, U. (2002). Insect hemolymph clotting: evidence for interaction between the coagulation system and the prophenoloxidase activating cascade. Insect. Biochem. Mol. Biol. 32, 919–928. doi: 10.1016/s0965-1748(02)00030-9
Lucock, M. (2000). Folic acid: nutritional biochemistry, molecular biology, and role in disease processes. Mol. Genet. Metab. 71, 121–138. doi: 10.1006/mgme.2000.3027
Mowat, E., Rajendran, R., Williams, C., McCulloch, E., Jones, B., Lang, S., et al. (2010). Pseudomonas aeruginosa and their small diffusible extracellular molecules inhibit Aspergillus fumigatus biofilm formation. FEMS Microbiol. Lett. 313, 96–102. doi: 10.1111/j.1574-6968.2010.02130.x
Nasir, M., Bean, H. D., Smolinska, A., Rees, C. A., Zemanick, E. T., and Hill, J. E. (2018). Volatile molecules from bronchoalveolar lavage fluid can ‘rule-in’ Pseudomonas aeruginosa and ‘rule-out’ Staphylococcus aureus infections in cystic fibrosis patients. Sci. Rep. 8:826. doi: 10.1038/s41598-017-18491-8
Nawrath, T., Gerth, K., Muller, R., and Schulz, S. (2010). The biosynthesis of the aroma volatile 2-methyltetrahydrothiophen-3-one in the bacterium chitinophaga Fx7914. Chembiochemistry 11, 1914–1919. doi: 10.1002/cbic.201000296
Penner, J. C., Ferreira, J. A., Secor, P. R., Sweere, J. M., Birukova, M. K., Joubert, L. M., et al. (2016). Pf4 bacteriophage produced by Pseudomonas aeruginosa inhibits Aspergillus fumigatus metabolism via iron sequestration. Microbiology 162, 1583–1594. doi: 10.1099/mic.0.000344
Peters, B. M., Jabra-Rizk, M. A., O’May, G. A., Costerton, J. W., and Shirtliff, M. E. (2012). Polymicrobial interactions: impact on pathogenesis and human disease. Clin. Microbiol. Rev. 25, 193–213. doi: 10.1128/cmr.00013-11
Reece, E., Doyle, S., Greally, P., Renwick, J., and McClean, S. (2018). Aspergillus fumigatus Inhibits Pseudomonas aeruginosa in co-culture: implications of a mutually antagonistic relationship on virulence and inflammation in the CF airway. Front. Microbiol. 9:1205. doi: 10.3389/fmicb.2018.01205
Reece, E., Segurado, R., Jackson, A., McClean, S., Renwick, J., and Greally, P. (2017). Co-colonisation with Aspergillus fumigatus and Pseudomonas aeruginosa is associated with poorer health in cystic fibrosis patients: an Irish registry analysis. BMC Pulm. Med. 17:70. doi: 10.1186/s12890-017-0416-4
Registry CFFP (2016). Annual Data Report © 2017 Cystic Fibrosis Foundation. Bethesda: Cystic Fibrosis Trust.
Robinson, C. V., Elkins, M. R., Bialkowski, K. M., Thornton, D. J., and Kertesz, M. A. (2012). Desulfurization of mucin by Pseudomonas aeruginosa: influence of sulfate in the lungs of cystic fibrosis patients. J. Med. Microbiol. 61, 1644–1653. doi: 10.1099/jmm.0.047167-0
Sambrook, J., Fritsch, E. F., and Maniatis, T. (1989). Molecular Cloning: A Laboratory Manual. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Sass, G., Nazik, H., Penner, J., Shah, H., Ansari, S. R., Clemons, K. V., et al. (2018). Studies of Pseudomonas aeruginosa mutants indicate pyoverdine as the central factor in inhibition of Aspergillus fumigatus biofilm. J. Bacteriol. 200:e00345-17. doi: 10.1128/JB.00345-17
Schulz-Bohm, K., Martin-Sanchez, L., and Garbeva, P. (2017). Microbial volatiles: small molecules with an important role in intra- and inter-kingdom interactions. Front. Microbiol. 8:2484. doi: 10.3389/fmicb.2017.02484
Shirazi, F., Ferreira, J. A., Stevens, D. A., Clemons, K. V., and Kontoyiannis, D. P. (2016). Biofilm filtrates of Pseudomonas aeruginosa strains isolated from cystic fibrosis patients inhibit preformed Aspergillus fumigatus biofilms via apoptosis. PLoS One 11:e0150155. doi: 10.1371/journal.pone.0150155
Shoseyov, D., Brownlee, K. G., Conway, S. P., and Kerem, E. (2006). Aspergillus bronchitis in cystic fibrosis. Chest 130, 222–226. doi: 10.1378/chest.130.1.222
Singer, M. (2010). Pathogen-pathogen interaction: a syndemic model of complex biosocial processes in disease. Virulence 1, 10–18. doi: 10.4161/viru.1.1.9933
Smith, K., Rajendran, R., Kerr, S., Lappin, D. F., Mackay, W. G., Williams, C., et al. (2015). Aspergillus fumigatus enhances elastase production in Pseudomonas aeruginosa co-cultures. Med. Mycol. 53, 645–655. doi: 10.1093/mmy/myv048
Smith, N. A., and Kelly, D. P. (1988). Isolation and physiological characterization of autotrophic sulfur bacteria oxidizing dimethyl disulfide as sole source of energy. J. Gen. Microbiol. 134, 1407–1417. doi: 10.1099/00221287-134-6-1407
Southern, E. M. (1975). Detection of specific sequences among DNA fragments separated by gel electrophoresis. J. Mol. Biol. 98, 503–517. doi: 10.1016/s0022-2836(75)80083-0
Suylen, G. M. H., Large, P. J., Vandijken, J. P., and Kuenen, J. G. (1987). Methyl mercaptan oxidase, a key enzyme in the metabolism of methylated sulfur-compounds by Hyphomicrobium Eg. J. Gen. Microbiol. 133, 2989–2997. doi: 10.1099/00221287-133-11-2989
Szewczyk, E., Nayak, T., Oakley, C. E., Edgerton, H., Xiong, Y., Taheri-Talesh, N., et al. (2006). Fusion PCR and gene targeting in Aspergillus nidulans. Nat. Protoc. 1, 3111–3120. doi: 10.1038/nprot.2006.405
Trejo-Hernandez, A., Andrade-Dominguez, A., Hernandez, M., and Encarnacion, S. (2014). Interspecies competition triggers virulence and mutability in Candida albicans-Pseudomonas aeruginosa mixed biofilms. ISME J. 8, 1974–1988. doi: 10.1038/ismej.2014.53
Tzin, V., and Galili, G. (2010). The biosynthetic pathways for shikimate and aromatic amino acids in Arabidopsis thaliana. Arabidopsis Book 8:e0132. doi: 10.1199/tab.0132
Woodcock, D. M., Crowther, P. J., Doherty, J., Jefferson, S., DeCruz, E., Noyer-Weidner, M., et al. (1989). Quantitative evaluation of Escherichia coli host strains for tolerance to cytosine methylation in plasmid and phage recombinants. Nucleic Acids Res. 17, 3469–3478. doi: 10.1093/nar/17.9.3469
Keywords: P. aeruginosa – A. fumigatus interaction, co-infection, volatile sulfur compounds, volatile interaction, interkingdom interaction, polymicrobial infection
Citation: Scott J, Sueiro-Olivares M, Ahmed W, Heddergott C, Zhao C, Thomas R, Bromley M, Latgé J-P, Krappmann S, Fowler S, Bignell E and Amich J (2019) Pseudomonas aeruginosa-Derived Volatile Sulfur Compounds Promote Distal Aspergillus fumigatus Growth and a Synergistic Pathogen-Pathogen Interaction That Increases Pathogenicity in Co-infection. Front. Microbiol. 10:2311. doi: 10.3389/fmicb.2019.02311
Received: 16 July 2019; Accepted: 23 September 2019;
Published: 09 October 2019.
Edited by:
Agostinho Carvalho, University of Minho, PortugalReviewed by:
Falk Hillmann, Leibniz Institute for Natural Product Research and Infection Biology, GermanyFrancoise Botterel, Université Paris-Est Créteil, France
Copyright © 2019 Scott, Sueiro-Olivares, Ahmed, Heddergott, Zhao, Thomas, Bromley, Latgé, Krappmann, Fowler, Bignell and Amich. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jorge Amich, am9yZ2UuYW1pY2hlbGlhc0BtYW5jaGVzdGVyLmFjLnVr
†Present address: Jean-Paul Latgé, School of Medicine, University of Crete, Heraklion, Greece