 Eduardo L. Almeida
Eduardo L. Almeida Andrés Felipe Carrillo Rincón
Andrés Felipe Carrillo Rincón Stephen A. Jackson
Stephen A. Jackson Alan D. W. Dobson
Alan D. W. Dobson- 1School of Microbiology, University College Cork, Cork, Ireland
- 2Environmental Research Institute, University College Cork, Cork, Ireland
Plastics, such as the polyethylene terephthalate (PET), are widely used for various industrial applications, due to their physicochemical properties which are particularly useful in the packaging industry. However, due to improper plastic waste management and difficulties in recycling, post-consumer plastic waste has become a pressing issue for both the environment and for human health. Hence, novel technologies and methods of processing plastic waste are required to address these issues. Enzymatic-assisted hydrolysis of synthetic polymers has been proposed as a potentially more efficient and environment-friendly alternative to the currently employed methods. Recently, a number of PET hydrolases have been described, and in particular a PETase derived from Ideonella sakaiensis 201-F6 (IsPETase), which appears to be the most efficient and substrate-specific bacterial PET hydrolase enzyme discovered to date. In order to further investigate this class of PETase-like enzymes, we employed an in silico-based screening approach on the biotechnologically relevant genus Streptomyces, including terrestrial and marine isolates; in a search for potential PETase homologs. From a total of 52 genomes analyzed, we were able to identify three potential PETase-like enzymes, all of which were derived from marine-sponge associated Streptomyces isolates. A candidate PETase-like gene (SM14est) was identified in Streptomyces sp. SM14. Further in silico characterization of the SM14est protein sequence and its predicted three-dimensional structure were performed and compared to the well-characterized IsPETase. Both the serine hydrolase motif Gly-x1-Ser-x2-Gly and the catalytic triad Ser, Asp, His are conserved in both sequences. Molecular docking experiments indicated that the SM14est enzyme possessed the capacity to bind plastics as substrates. Finally, polyesterase activity was confirmed using a polycaprolactone (PCL) plate clearing assay which is a model substrate for the degradation of plastics; following heterologous expression of SM14est in Escherichia coli, with secretion being facilitated by the native Streptomyces signal peptide. These findings provide further insights into this important class of PETase-like enzymes.
Introduction
Plastics are materials that have been produced on a large scale from the 1950s onwards, and since then have been widely used for various applications, and have become almost indispensable in modern society (Jambeck et al., 2015; Geyer et al., 2017; Lebreton and Andrady, 2019). In the 1960s, plastics accounted for less than 1% of municipal solid waste in the United States, but steadily increased to around 10% by 2005 in countries with middle to high income (Jambeck et al., 2015; Geyer et al., 2017). This was largely due to their advantageous properties, such as their low production cost and bio-inertia, which are particularly useful for the packaging industry, when compared to other materials. This has resulted in the use of plastics in the packaging sector, which accounts for around 40% of the plastic converter demand in Europe (PlasticsEurope, 2018; Lebreton and Andrady, 2019). However, some of these aforementioned characteristics have resulted in plastics becoming a critical problem from an environmental perspective; as many synthetic plastics are highly recalcitrant to biodegradation and can persist for long periods of time in the environment (Wei and Zimmermann, 2017b).
In 2017, there was an estimated worldwide plastics production of 348 million tons – an increase from the 335 million tons estimated for the previous year, and this does not include polyethylene terephthalate (PET)-, polyamide (PA)-, and polyacryl-fibers (PlasticsEurope, 2018). At the current rate, this number is expected to double in the next 20 years (Lebreton and Andrady, 2019). In the past decade, plastic waste management policies have helped considerably in reducing post-consumer plastic waste being disposed into the environment. For example, in Europe more collected plastic waste (31.1% of 27.1 million tons of collected plastic waste in 2016) was submitted to recycling rather than to landfills, for the first time. However, landfills and incineration for energy recovery still account for 27.3 and 41.6% of the collected plastic post-consumer waste, respectively (PlasticsEurope, 2018).
Notwithstanding this, the aforementioned metrics do not take into account global mismanaged plastic waste which enters the natural environment, at locations others than landfills. A recent study has estimated that between 60 and 99 million metric tons of mismanaged plastic waste was produced globally in 2015, and that this number could triple by 2060 (Lebreton and Andrady, 2019). Mismanaged plastic waste is particularly concerning when it effects the marine environment. It has been calculated that in 2010 between 4.8 and 12.7 million metric tons of plastic waste entered our oceans, and this data only accounted for coastal countries (Jambeck et al., 2015). Sunlight and other weathering effects cause the fragmentation of plastic debris into milli- and micro-metric particles (<5 mm), which are defined as micro-plastics (Geyer et al., 2017; Lebreton and Andrady, 2019). These micro-plastics are now believed to be ubiquitous in soil and aquatic environments, and are commonly ingested by animals (Santillo et al., 2017; Lebreton and Andrady, 2019). This is especially concerning, since micro-plastics can adsorb and concentrate pollutants present in the ocean and transfer them along the food chain, particularly to seafood species that are consumed by humans (Santillo et al., 2017). Highlighting the issue of the ubiquitous presence of micro-plastics in the marine environment, a recent study detected the ingestion of micro-plastics by deep-sea amphipods, at depths ranging from 7,000 to 10,890 m (Jamieson et al., 2019). Additionally, the deepest ever sub diving recorded to date has registered the presence of plastics on the ocean floor, at a depth of 10,927 m (Street, 2019). It is alarming to find plastics, which are materials with a history of less than a century of large-scale production, already being so widespread in nature; with the potential for extensive negative impacts, many of which have yet to be fully realized. Hence, better plastic waste management and processing solutions are urgently required.
Currently, the majority of plastic waste recycling is based on mechanical recycling (collection, sorting, washing, and grinding) (Ragaert et al., 2017). However, the presence of organic and inorganic impurities in post-consumer plastic waste presents a huge challenge for mechanical recycling (Drzyzga and Prieto, 2019). On the other hand, chemical recycling has been applied as an alternative for improved plastic waste management processes, in which the plastic polymers can be converted into raw materials that can be used for the synthesis of chemicals, fuels, or virgin plastics (Drzyzga and Prieto, 2019). Strictly chemical methods, however, require the use of toxic chemicals and high temperatures, and can also be quite costly (Awaja and Pavel, 2005; Wei and Zimmermann, 2017a). Therefore, enzymatic hydrolysis of synthetic polyesters has been proposed as a potentially more efficient and environment friendly method for the recycling of plastic waste (Wei and Zimmermann, 2017a; Drzyzga and Prieto, 2019).
In the past decade, a number of bacterial enzymes capable of degrading synthetic polyesters, including the widely used PET, have been identified (Wei and Zimmermann, 2017a; Kawai et al., 2019). These enzymes are commonly classified as members of the cutinase, lipase and esterase classes of enzymes, and to date have mainly been identified in thermophilic actinomycetes, particularly in the genus Thermobifida (Silva et al., 2011; Wei and Zimmermann, 2017a). More recently, Yoshida et al. (2016) isolated a bacterium from a PET plastic bottle recycling plant in Sakai, Japan, that was capable of degrading and assimilating PET as its major energy and carbon source – namely Ideonella sakaiensis 201-F6. The protein identified as being responsible for the hydrolysis of PET (ISF6_4831) was then defined as a PETase (or PET hydrolase) enzyme (EC 3.1.1.101) (Yoshida et al., 2016). The PETase from Ideonella sakaiensis 201-F6 has been shown to possess a relatively higher enzymatic activity and substrate specificity for PET than other previously described PET hydrolases, in addition to the ability to degrade PET at moderate temperatures (around 30°C) (Yoshida et al., 2016; Joo et al., 2018). Since then, a number of studies have been undertaken in a concerted effort to characterize this enzyme and the underlying metabolic and biochemical processes involved in the degradation of PET (Han et al., 2017; Chen et al., 2018; Joo et al., 2018; Liu et al., 2018).
The Streptomyces genus, member of the Actinomycetales order, is well-known to produce compounds and enzymes of industrial and clinical interest, particularly antibiotics, for which it is considered the largest producer in the microbial world (Watve et al., 2001; Hwang et al., 2014; Ser et al., 2017; Spasic et al., 2018). Recent efforts to exploit the biotechnological potential of Streptomyces species have largely focused on the identification of bioactive small molecules and secondary metabolites biosynthetic gene clusters (Chevrette et al., 2019; Manteca and Yagüe, 2019). In this respect, the focus has started to shift toward Streptomyces isolates derived from varied niche environments, such as those isolated from the marine environment, which are still not well-characterized and majorly unexplored organisms, when compared to the previously more commonly studied soil-derived isolates (Dharmaraj, 2010; Hassan et al., 2017; Jin et al., 2018; Xu et al., 2018; Almeida et al., 2019).
Streptomyces isolates from soil ecosystems have also been studied for their synthetic polyesters-degrading capabilities (Calabia and Tokiwa, 2004; Shivlata and Satyanarayana, 2015). These include Streptomyces sp. strain MG (Tokiwa and Calabia, 2004) and Streptomyces thermoviolaceus (Chua et al., 2013), which can degrade polycaprolactone (PCL); together with Streptomyces bangladeshensis 77T-4, which degrades poly(D-3-hydroxybutyrate) (PHB) (Hsu et al., 2012). Given that marine Streptomyces sp. SNG9 had previously been reported to degrade PHB (Mabrouk and Sabry, 2001), coupled with the fact that marine Streptomyces isolates are likely to have been exposed to plastics and/or microplastics in marine ecosystems – in particular those isolates which are associated with marine sponges (phylum Porifera), which filter large quantities of seawater (up to 24,000 L of water per day/Kg sponge) on a daily basis to obtain nutrients (Taylor et al., 2007; Food and Agriculture Organization of the United Nations, 2017; Godefroy et al., 2019) – we reasoned that marine sponge-derived Streptomyces species may possess enzymes with an ability to degrade synthetic polymers. In this study, we screened a number of Streptomyces species, including both terrestrial and marine-derived isolates, using an in silico-based analysis to interrogate their genomes for potential PETase homologs. A candidate PETase-like gene was identified in Streptomyces sp. SM14 and enzyme activity was confirmed following heterologous expression of this gene in Escherichia coli. This is the first report of a PETase-like enzyme being identified in a marine sponge-derived Streptomyces spp. isolate, and we believe that this study provides further insights into our current knowledge of this important class of enzymes.
Materials and Methods
Data Sets
The reference data set was comprised of 15 amino acid sequences of enzymes with previously demonstrated synthetic polyesters-degrading capabilities (Table 1) (Yoshida et al., 2016; Wei and Zimmermann, 2017a; Danso et al., 2018; Joo et al., 2018; Kawai et al., 2019). A lipase from Streptomyces exfoliatus (PDB ID: 1JFR), which is a cutinase-like enzyme (Wei et al., 1998; Kawai et al., 2019), was also included in the reference data set (Table 1), which although not possessing demonstrated polyester-degrading activity, served as an outgroup for the subsequent in silico analyses.
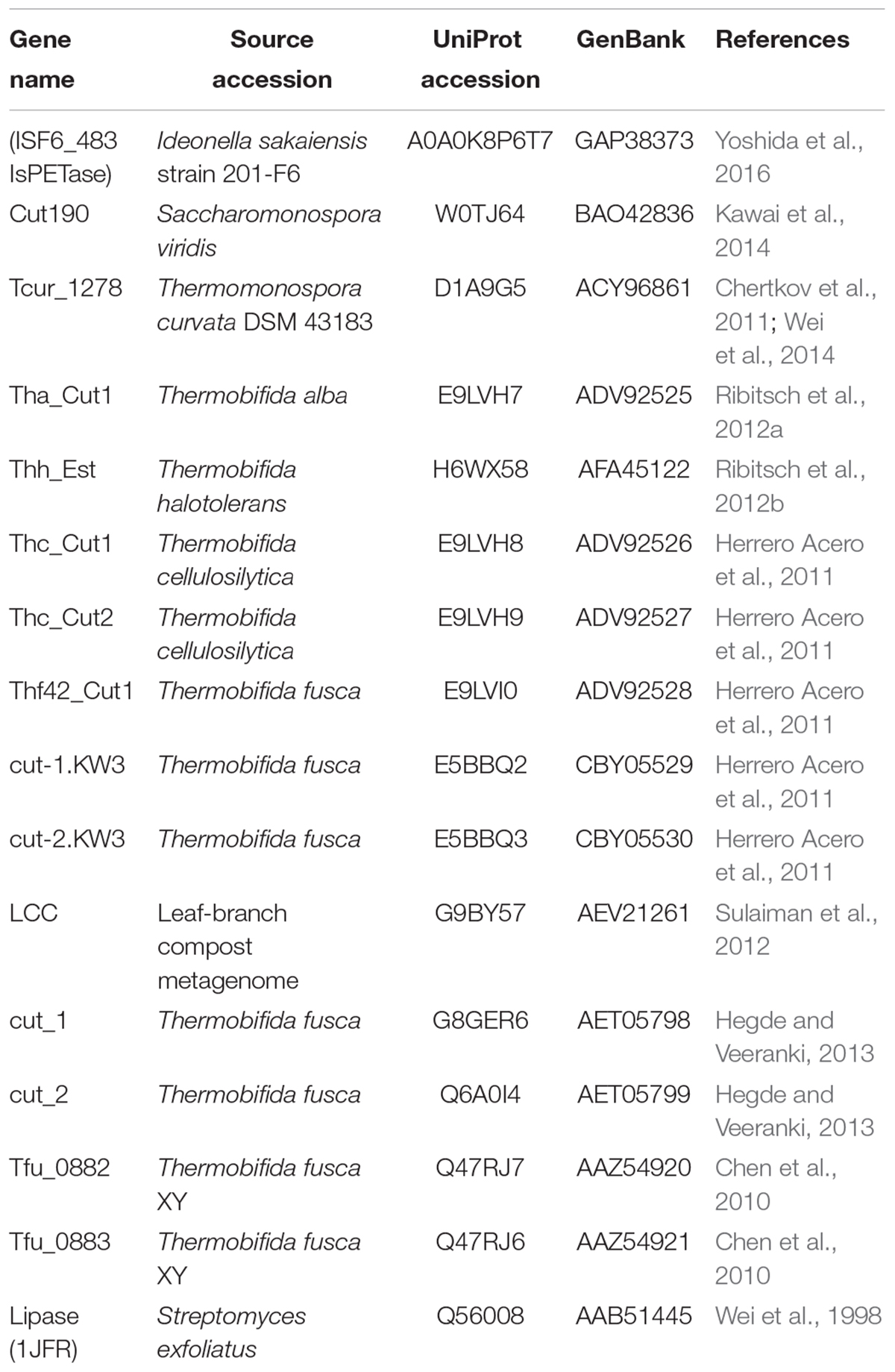
Table 1. Reference data set comprising of 15 PETase-like enzymes with demonstrated PET-degrading activity, including the ISF6_483 protein from Ideonella sakaiensis strain 201-F6 (IsPETase), and additionally the cutinase-like lipase from Streptomyces exfoliatus (PDB ID: 1JFR).
The Streptomyces genomes data set comprised of 52 Streptomyces genome sequences obtained from GenBank (Benson et al., 2018), including 23 genomes from terrestrial isolates, and 29 from marine isolates (Supplementary Table S1). Open reading frames (ORFs) and their respective translated amino acid sequences were obtained using Prokka (Seemann, 2014).
Bacterial Strains
Streptomyces strain SM14 was isolated from the sponge Haliclona simulans (class Demospongiae, order Haplosclerida, family Chalinidae) which was sampled by SCUBA diving at a depth of 15 m in Kilkieran Bay, Galway, Ireland (N 53°18′56.6′′, W 09°40′08.4′′) as previously described (Kennedy et al., 2009). The NEB® 5-alpha and the BL21(DE3) competent E. coli cells were obtained from New England Biolabs, Inc., United States.
Protein Homology Search and Phylogeny Analysis
Potential PETase-like proteins were identified in the Streptomyces genomes data set by performing an homology search using BLASTP (e-value threshold of 1e-30, maximum subject sequence length of 400 aa) (Altschul et al., 1990; Camacho et al., 2009). Protein alignments were performed using Muscle (Edgar, 2004), and phylogeny analysis was performed using MEGA X (maximum likelihood statistical method; 500 bootstrap replications; 50% bootstrap cut-off value; LG + G + F model) (Kumar et al., 2018).
PCL Plate Clearing Assay
Polycaprolactone (PCL) plate clearing assays were performed based on previously described studies (Nishida and Tokiwa, 1993; Murphy et al., 1996; Nishida et al., 1998; Nawaz et al., 2015). PCL is a synthetic polyester that has previously been used as a model substrate to assess both PETase and cutinase enzymatic activities (Nyyssölä et al., 2013; Danso et al., 2018). PCL with an average molecular weight of 80,000 was used (Sigma-Aldrich®). PCL emulsion was prepared with 1% m/v of PCL in acetone, at 50°C with magnetic stirring. Water, agar (1.5% m/v) and LB medium (2% m/v) were added to the emulsion, at 50°C with magnetic stirring until the acetone evaporated. The medium was then autoclaved and poured into plates. Streptomyces sp. SM14 and transformed E. coli BL21(DE3) containing the pET20b:SM14est vector construct were inoculated onto the plates and incubated at 28°C for up to 12 days. For enzyme activity assessment using E. coli as the heterologous host, isopropyl-β-D-thiogalactopyranoside (IPTG) was added to the medium at a concentration of 0.5 mM and plates were incubated at 28°C for up to 4 days. As a negative control, E. coli BL21(DE3) containing the pET-20b(+) plasmid without the insert did not show any PCL-degrading activity (results not shown). It has recently been reported that E. coli BL21(DE3) can be employed as a host system in screens for polyesterase activity, as it does not possess PCL-degrading capabilities (Molitor et al., 2019). Additionally, 12 other marine sponge-derived isolates were also assayed for polyesterase activity using the PCL plate clearing assay. SM1, SM3, SM4, SM7, SM8, SM9, SM11, SM13, SM17, and FMC008 which had previously been isolated from the marine sponge Haliclona simulans Kennedy et al., 2009), together with B188M101 and B226SN101 isolated from the deep sea sponges Lissodendoryx diversichela and Inflatella pellicula respectively (Jackson et al., 2018); were grown in LB medium + 1% PCL emulsion at 28°C for 12 days (data not shown).
Protein Structure Analysis, Modeling, and Molecular Docking
Amino acid sequence analysis was performed and graphically represented using ESPript 3.0 (Robert and Gouet, 2014). In silico protein structure prediction was performed using the SWISS-DOCK webserver (Guex et al., 2009; Benkert et al., 2011; Bertoni et al., 2017; Bienert et al., 2017; Waterhouse et al., 2018), and the UCSF Chimera software was used for structure analysis and three-dimensional model rendering (Pettersen et al., 2004). Molecular docking experiments were performed using AutoDock Vina, MGLtools1, AutoDockTools (ADT) and UCSF Chimera (Sanner, 1999; Pettersen et al., 2004; Trott and Olson, 2009), with the model substrate BHET [Bis(2-hydroxyethyl) terephthalate, Zinc database ID: ZINC02040111] molecule as the ligand (Irwin and Shoichet, 2005; Irwin et al., 2012). BHET has previously been used as a model substrate for PET degradation both in vitro and in silico through molecular docking studies (Hantani et al., 2018; Joo et al., 2018; Liu et al., 2018).
Heterologous Expression
An E. coli codon-optimized version of the PETase-like gene was designed, and was synthesized by Eurofins Genomics (Ebersberg, Germany). A 5′ NdeI restriction site, a C-terminal His6 tag, and a stop codon followed by a 3′ XhoI restriction site were added to the gene sequence. Alignment of the nucleotide sequences of the original SM14est gene and the codon-optimized version is shown in Supplementary Figure S1. The synthetic gene was PCR amplified using Phusion Green High-Fidelity DNA Polymerase (Thermo ScientificTM) (primers and conditions detailed in Supplementary Table S2), and was subcloned into the pET-20b(+) plasmid (Novagen®), resulting in the pET20b:SM14est vector construct (Figure 1), using the NEB® 5-alpha competent E. coli (New England Biolabs, Inc., United States) for vector construction and maintenance. The signal peptide of the native protein was predicted using SignalP 5.0 (Almagro Armenteros et al., 2019), and it was maintained in the final construct. The expression vector was then transformed into BL21(DE3) competent E. coli (New England Biolabs, Inc., United States) for heterologous protein expression. Confirmation of the insert was performed via (1) restriction digestion of the plasmid DNA with the NdeI and XhoI restriction enzymes followed by gel electrophoresis analysis and (2) via Sanger sequencing of the insert region of the plasmid, amplified using the T7 standard vector primers (Supplementary Table S2).
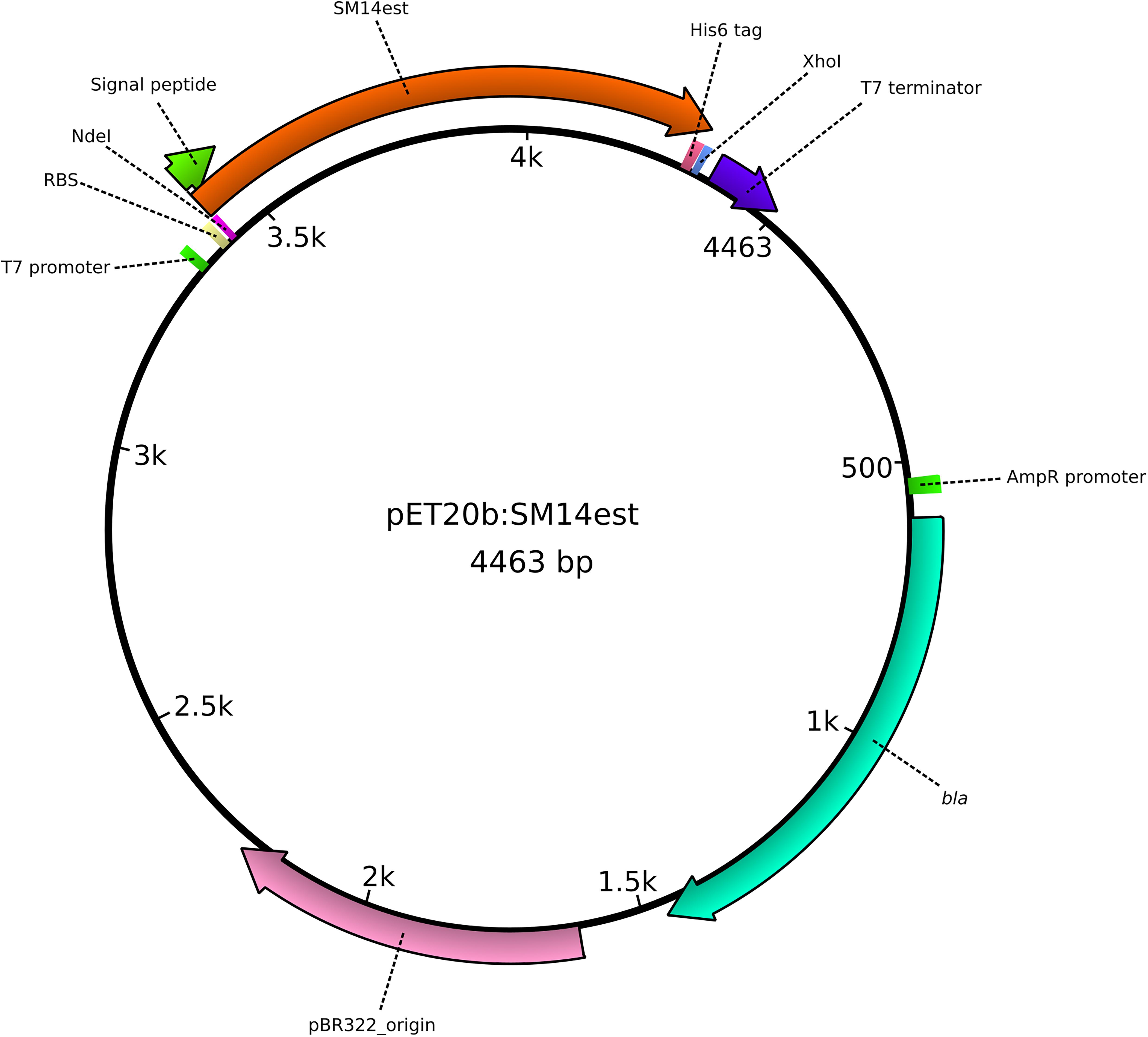
Figure 1. Graphical representation of the pET20b:SM14est plasmid, constructed for the heterologous expression of the SM14est protein in Escherichia coli. The insert (SM14est) and other important features of the plasmid are represented and labeled accordingly.
Results and Discussion
In silico Screening of PETase-Like Proteins in Streptomyces Genomes and Enzyme Activity Assessment
Previous studies have identified enzymes with plastic-degrading capabilities which have been isolated from different organisms, amongst these were the Ideonella sakaiensis strain 201-F6, and isolates from the genera Thermobifida, Thermomonospora, and Saccharomonospora (Wei and Zimmermann, 2017a; Danso et al., 2018; Joo et al., 2018; Kawai et al., 2019). This class of enzymes is commonly referred to as PETase or PETase-like, due to their ability to hydrolyse PET, although at different levels of efficiency. From the bacterial PETase-like class of enzymes discovered to date, the PETase from Ideonella sakaiensis strain 201-F6 (referred to from now on as IsPETase) is the one that has received most attention from the scientific community; as it is considered the enzyme which displays the best PET hydrolase activity and substrate specificity which has been discovered thus far (Yoshida et al., 2016; Joo et al., 2018; Kawai et al., 2019). A total of 15 of the most prominent PETase-like enzymes were selected to build the reference data set (Table 1), which was then used to search for potential homologous proteins in the Streptomyces genomes data set. As described previously, the Streptomyces genomes data set comprised of 52 genome sequences, including 23 terrestrial Streptomyces isolates, and 29 isolates derived from the marine environment – 20 of which had previously been isolated by our group and for which genome sequences were available (Supplementary Table S2) (Kennedy et al., 2009; Almeida et al., 2018; Jackson et al., 2018).
By applying a sequence similarity search approach using BLASTP (Altschul et al., 1990; Camacho et al., 2009), 34 potential homologous proteins from 32 Streptomyces strains were identified, of which the majority were from marine isolates (22 in total). These were then selected for further phylogeny analysis using MEGA X (Kumar et al., 2018). The amino acid sequences of the reference data set, and the Streptomyces potential PETase-like homologs were aligned using the Muscle program (Edgar, 2004), and a maximum likelihood phylogenetic tree was generated, with 500 bootstrap replicates. The resulting consensus phylogenetic tree, with a 50% bootstrap value cut-off, comprised of four main clades (Figure 2). Clade number 1 appeared to include Streptomyces isolates that were previously reported to share genetic similarity to the type strain Streptomyces albus J1074, all of which shared > 99.50% 16S rRNA gene sequence similarity amongst each other (Ian et al., 2014; Zaburannyi et al., 2014). Clade number 2 showed less obvious similarities between all the members of the clade. While it included the strains SM1, SM3 and SM4, that were isolated from the marine sponge Haliclona simulans and shared high similarity in their 16S rRNA sequences (>99%) (Kennedy et al., 2009; Jackson et al., 2018), it also included the sponge isolate Streptomyces sp. 13-12-16 that shared less 16S rRNA similarity with the aforementioned strains (∼98%), and the soil isolate Streptomyces glaucescens strain GLA.O, which in addition to being isolated from a completely different environment, also shares < 99% 16S rRNA similarity with the others in the clade. Clade number 3 appeared to be the most diverse, with Streptomyces isolated from varied sources, including: soil, marine sediment, and those isolated from marine sponges, lichens and insects (Ohnishi et al., 2008; Kennedy et al., 2009; Bianchetti et al., 2013; Shin et al., 2013; Xu et al., 2018). Clade 3 also included the lipase from Streptomyces exfoliatus (indicated with an asterisk in Figure 2), suggesting that these enzymes are likely to be cutinase-like lipases (Wei et al., 1998; Kawai et al., 2019). Most interesting, however, was clade number 4, which clearly included all of the PETase-like enzymes used in the reference data set, indicating that they may share similar evolutionary processes and history that differentiate them from the other proteins considered in this analysis, which may possibly have led to their ability to degrade synthetic polyesters. It is noteworthy that 3 proteins from marine Streptomyces isolates were also included in clade 4, specifically protein sequences from the Streptomyces sp. SM12, Streptomyces sp. SM14, and Streptomyces xinghaiensis S187 isolates, which is a strong indicator that these enzymes may possess plastic-degrading capabilities.
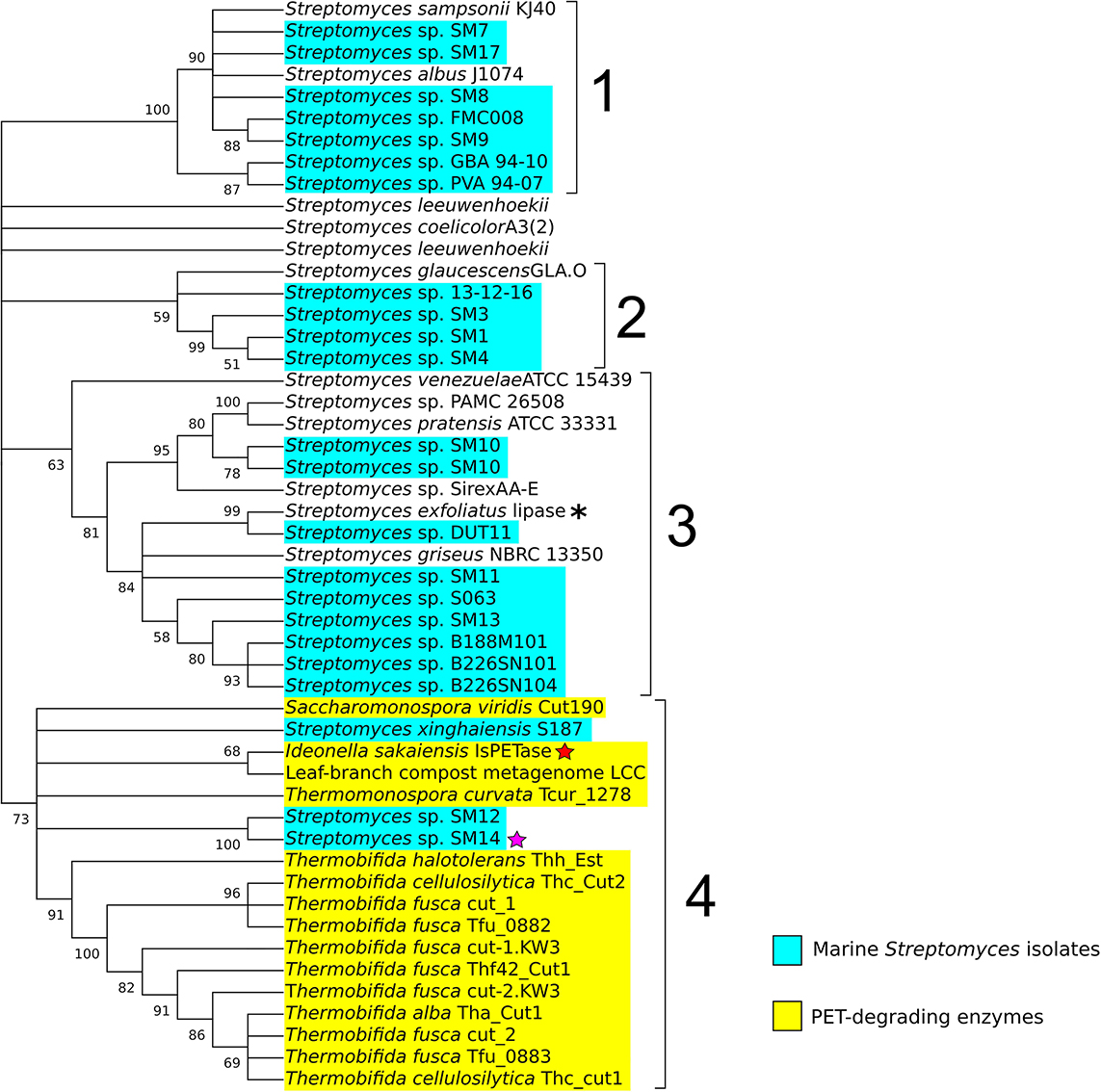
Figure 2. Phylogenetic tree of potential PETase homologs identified in the Streptomyces genomes, including terrestrial and marine (highlighted in cyan) Streptomyces isolates. The enzymes with known PET-degrading activity are highlighted in yellow. The red star indicates the Ideonella sakaiensis IsPETase, the purple star a PETase-like protein from the Streptomyces sp. SM14, and the asterisk the lipase from Streptomyces exfoliatus. The numbers in the branches indicate the percentage of bootstrap replicates (with a 50% cut-off from 500 replicates) in which the associated taxa clustered together. Repeated strain names indicate the presence of multiple proteins in their respective genomes that shared similarity with the reference PETases proteins.
Subsequent amino acid sequence analysis showed that the SM12 and SM14 proteins are in fact identical, so additional analysis proceeded with the SM14 strain. The enzyme activity was confirmed with a PCL plate clearing assay – a model substrate to assess PETase and cutinase enzymatic activities (Molitor et al., 2019), in which the SM14 strain was grown in LB medium + 1% PCL emulsion at 28°C for 12 days (Figure 3). The zone of clearing demonstrates the synthetic polyester-degrading capability of the Streptomyces sp. SM14 isolate (Figure 3), which is presumably due to the protein identified from the in silico screening (Figure 2). Therefore, for the purposes of this study, the SM14 protein will from now on be referred to as SM14est, as it is likely to be a potential polyesterase enzyme. The SM14est gene sequence was deposited in the GenBank database under the accession number BK010828.
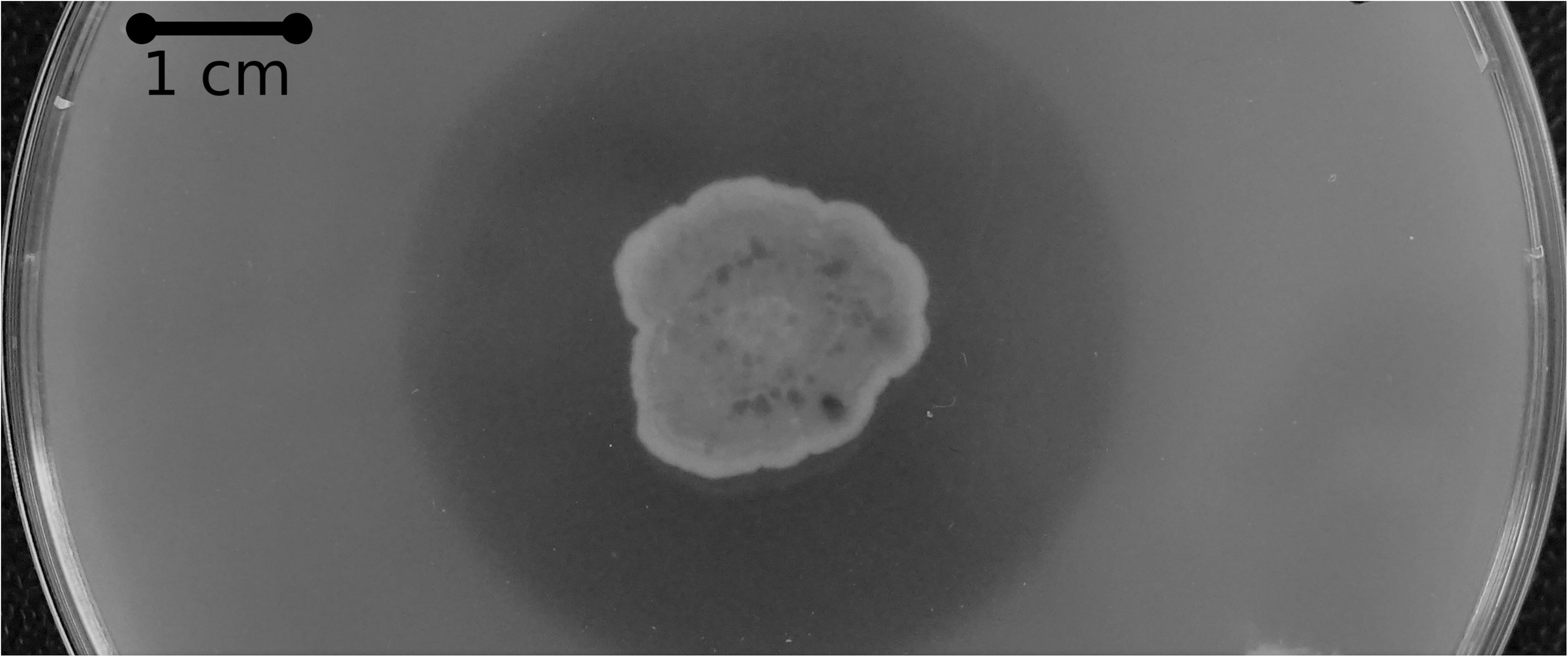
Figure 3. PCL plate clearing assay with the Streptomyces sp. SM14 strain incubated at 28°C after 12 days.
In addition, we screened 12 other marine Streptomyces isolates from the different clades (clade 1: SM7, SM8, SM9, SM17, and FMC008) (Clade 2: SM1, SM3, SM4) and (Clade 3: SM11, SM13, B188M101, and B226SN104) of the phylogenetic tree identified from the in silico screening (Figure 2) using the PCL plate clearing assay. No zones of clearing were however observed following growth of each strain in LB medium + 1% PCL emulsion at 28°C for 12 days (data not shown).
Protein Structure Analysis
Amino Acid Sequence, Conserved Residues, and Domains
An amino acid sequence comparison between the SM14est and the IsPETase was performed, using the previously described PETase enzyme sites as reference (Joo et al., 2018). The amino acid sequences were aligned using the Muscle algorithm in MEGA X (Edgar, 2004; Kumar et al., 2018), and the alignment and amino acid residues were analyzed in ESPript 3.0 (Figure 4) (Robert and Gouet, 2014). The amino acid alignment showed that 41% of the amino acids in SM14est were identical to the IsPETase, and that an additional 19% of the SM14est amino acids shared similar biochemical properties to the IsPETase. The serine hydrolase motif Gly-x1-Ser-x2-Gly is conserved in both sequences (residues in IsPETase: Gly158-Trp159-Ser160-Met161-Gly162; residues in SM14est: Gly154-His155-Ser156-Met157-Gly158). The catalytic triad is also conserved in both sequences (residues in IsPETase: Ser160, Asp206, His237; and in SM14est: Ser156, Asp202, His234) (Figure 4 and Supplementary Table S3), which is to be expected given that this catalytic triad has been shown to be crucial for enzymatic activity in this class of enzymes. In previous site-direct mutagenesis experiments performed with the IsPETase protein, substitution of any of the residues within the catalytic triad resulted in a complete disruption of the catalysis process (Joo et al., 2018; Liu et al., 2018). One major difference between the IsPETase and our SM14est is that the former possesses two disulphide bonds (the first between Cys273 and Cys289, and the second between Cys203 and Cys239), while the latter has none. Although disulphide bonds are generally related to higher protein thermostability, it has been proposed that the second disulphide bond in IsPETase is connected to its enzymatic activity, since it is positioned in close proximity to the enzyme’s active sites, and substitution of this disulphide bond via site-directed mutagenesis experiments resulted in a drastic decrease in PET hydrolysis (Joo et al., 2018; Liu et al., 2018). However, the requirement of this extra disulphide bond may be exclusive to the IsPETase, since other PETase-like cutinases display PET hydrolase activity and high thermostability without possessing this disulphide bond (Kawai et al., 2019).
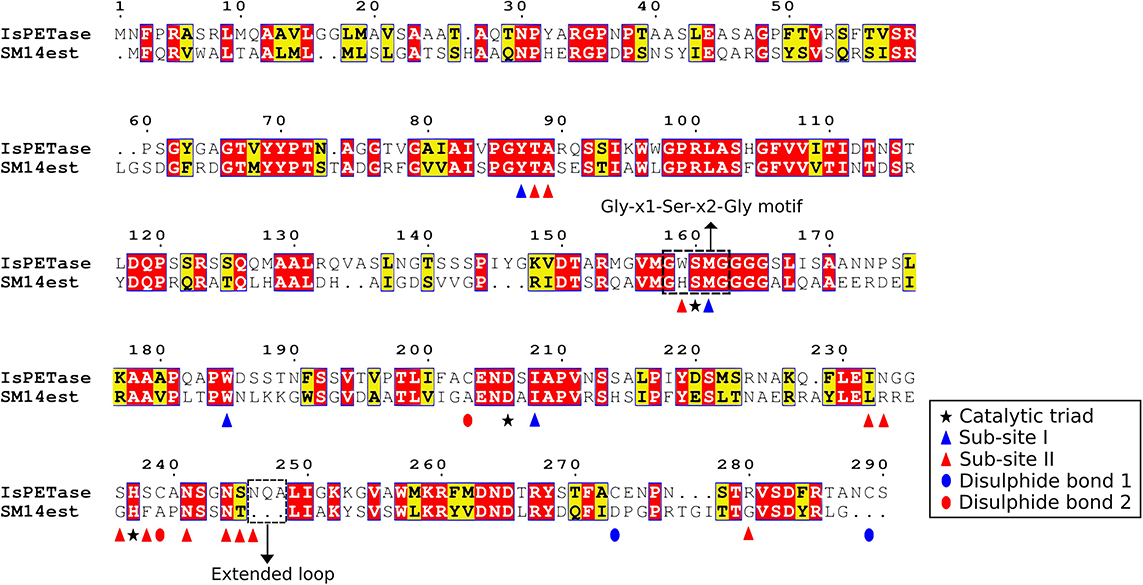
Figure 4. Amino acid sequence alignment of the IsPETase and the SM14est proteins, generated using MEGA X, Muscle, and ESPript 3.0. Identical residues are highlighted in red boxes, and the ones with similar biochemical properties are highlighted in yellow boxes. The serine hydrolase motif Gly-x1-Ser-x2-Gly and the IsPETase extended loop are highlighted in black boxes; the catalytic triad is indicated by a star; the sub-site I and sub-site II are indicated by a blue and a red triangle, respectively. The disulphide bond 1 and disulphide bond 2 are indicated by blue and red ellipses, respectively.
Previous molecular docking analysis of the IsPETase, using a four-monohydroxyethyl terephthalate (MHET) molecule that would mimic PET [2-hydroxyethyl-(monohydroxyethyl terephthalate)4 or 2-HE(MHET)4] as ligand, together with site-directed mutagenesis analysis, suggests that the enzyme possess two sub-sites (Joo et al., 2018). Sub-site I has been proposed to be responsible for the binding of the first MHET moiety, and thus for stabilization of the ligand. Meanwhile, sub-site II has been proposed to be responsible for accommodating the other three MHET moieties, partially leading to the superior PET degradation by the IsPETase in comparison to other PETase-like enzymes. The sub-site I, which consists of 4 residues, is conserved in both IsPETase and SM14est (residues in IsPETase: Tyr87, Met161, Trp185, Ile208; residues in SM14est: Tyr88, Met157, Trp181, Ile204) (Figure 4 and Supplementary Table S3). This implies that both enzymes have a similar mode of binding to the substrate. However, major differences exist between the two sequences in the 12-residue sub-site II region (residues in IsPETase: Thr88, Ala89, Trp159, Ile232, Asn233, Ser236, Ser238, Asn241, Asn244, Ser245, Asn246, Arg280; residues in SM14est: Thr89, Ala90, His155, Leu229, Arg230, Gly233, Phe235, Asn238, Asn241, Thr242, GAP, Gly277) (Figure 4 and Supplementary Table S3). These differences in the sub-site II region in both proteins could lead to different binding affinities to the moieties of the PET polymer. Another important difference between the two protein sequences is in the loop connecting β8 and α6, in which IsPETase appears to possess 3 extra amino acids in comparison to SM14est (Asn246, Gln247, Ala248) (Figure 4). This extended loop has been proposed to be an important structural feature of the IsPETase. When compared to a cutinase from Thermobifida fusca KW3, the extended loop seems to provide a conformation that allowed the formation of a continuous cleft on sub-site II – and hence accommodation of the third and fourth MHET moieties. This may represent another structural characteristic that could potentially explain the superior enzymatic activity of IsPETase (Joo et al., 2018).
It has been suggested that the aforementioned protein structural differences between the IsPETase and the cutinase from Thermobifida fusca KW3 – namely the absence of two disulphide bonds; differences in the sub-site II residues and the lack of an extended loop – could result in a reduced efficiency in the degradation of PET when compared to the IsPETase. Additionally, it has been proposed that proteins with these characteristics could be classified into the type I category of PETase-like enzymes, which also seems to be the case for the SM14est protein (Joo et al., 2018).
Protein Three-Dimensional Structure Prediction and Molecular Docking
The function and stability of proteins is closely linked to its conformation, or folded/native state (Lumry and Eyring, 1954; England and Haran, 2011; Balcão and Vila, 2015). To provide further insights into the potential functionality and conformation of the SM14est protein, a three-dimensional structure of the protein was in silico predicted using SWISS-MODEL (Benkert et al., 2011; Bertoni et al., 2017; Bienert et al., 2017; Waterhouse et al., 2018). The cutinase 1 (Thc_Cut1) from Thermobifida cellulosilytica (PDB ID: 5LUI) was used (Ribitsch et al., 2017) as a template for the model prediction, which generated a model with a GMQE score of 0.76 and a QMEAN Z-score of −1.76, indicating a reliable predicted model (Figure 5B). When compared to the structure of the IsPETase (Figure 5A), the predicted structure of the SM14est shows many similarities, with both belonging to the α/β hydrolase superfamily (Ollis et al., 1992; Nardini and Dijkstra, 1999; Hotelier et al., 2004), displaying a similar arrangement of 9 β-sheets and 7 α-helixes (Figure 5). The arrangement of the catalytic triad residues is also quite similar, as highlighted in Figures 5A,B, which may partially explain the synthetic polyester-degrading activity of these enzymes. With respect to observed potential differences between the two protein structures, the most striking differences – as previously shown in the amino acid sequence comparison, were the lack of disulphide bonds in the SM14est, and the absence of an extended loop between β8 and α6. The latter, as previously mentioned, has been proposed to be linked to proper accommodation of the MHET moieties constituting the PET polymer, and therefore the superior enzymatic activity of the IsPETase (Joo et al., 2018).
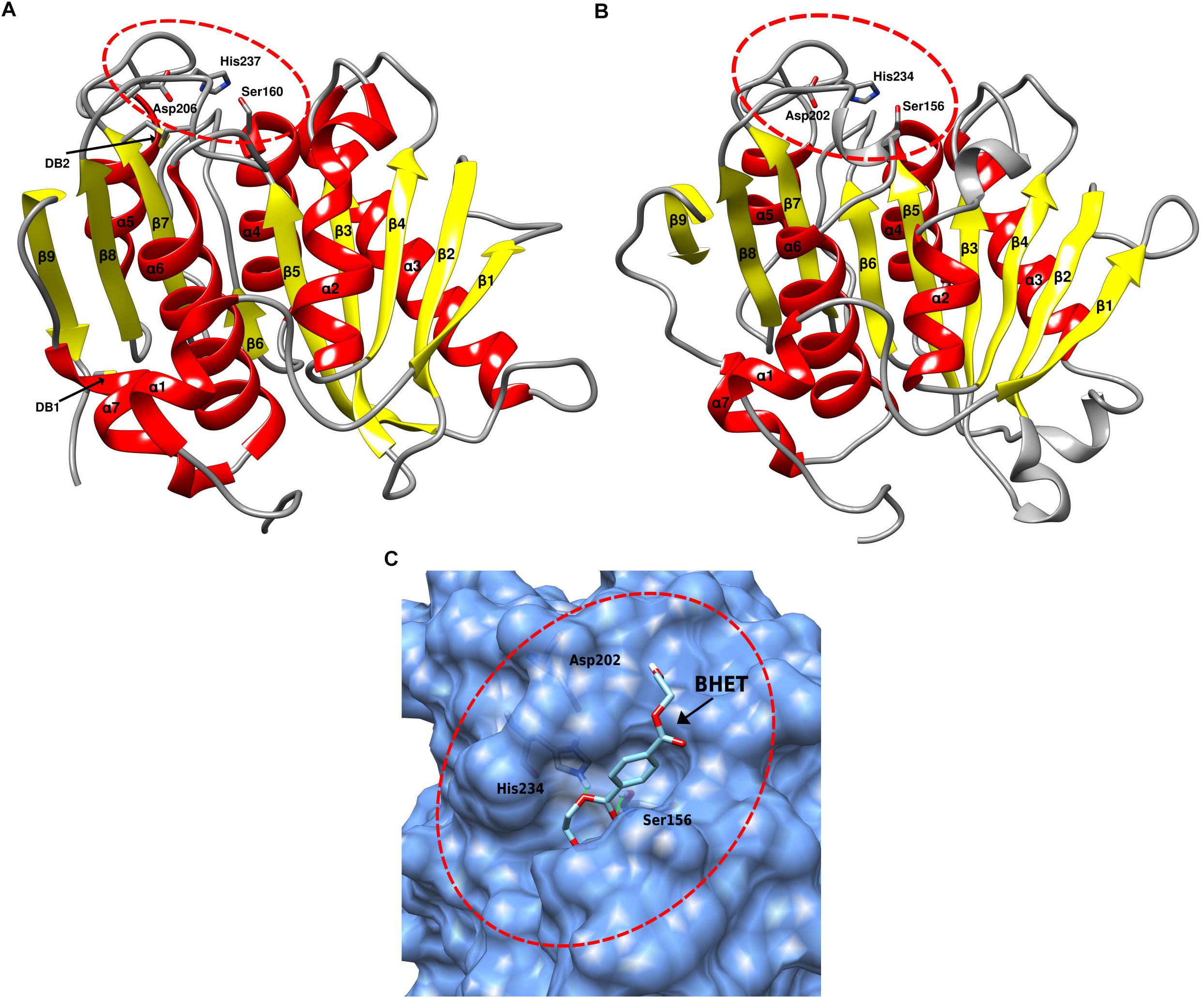
Figure 5. Three-dimensional protein structure comparison and molecular docking. (A) IsPETase three-dimensional structure (PDB ID: 5XJH), the catalytic triad (Ser160, Asp206, His237) is circled in red; the two disulphide bonds (DB1 and DB2) are indicated with arrows. (B) Predicted three-dimensional structure of the SM14est protein, generated using SWISS-MODEL, with the crystal structure of cutinase 1 from Thermobifida cellulosilytica as template (PDB ID: 5LUI). The catalytic triad (Ser156, Asp202, His234) is circled in red. (C) Molecular docking simulation performed using AutoDock Vina, with BHET as substrate, indicated with an arrow, detailing the binding pocket, which is circled in red. The catalytic triad residues are shown as sticks. Structures were analyzed and rendered using the UCSF Chimera software.
Molecular docking experiments were then performed, to analyze the likelihood of the SM14est enzyme possessing the capacity to bind plastics as substrates using an in silico-based approach; as well as determining the most probable binding mode of the protein to the ligand. To this end, the AutoDock Vina program was used for the protein-ligand molecular docking simulations (Trott and Olson, 2009), using the previously predicted SM14est structure and the BHET molecule as the ligand. The molecular docking experiment generated six binding modes with energy ≤ -5.0 kcal/mol, similar to the binding modes energy values that had previously been described in similar molecular docking experiment performed with IsPETase and BHET (Liu et al., 2018). The binding modes were analyzed in more detail using the UCSF Chimera software (Pettersen et al., 2004), and the best mode with the lowest binding energy is represented in Figure 5C, which highlights the binding pocket of the enzyme, the catalytic triad, and the proposed binding mode of the BHET molecule. The protein structure and molecular docking analyses results further emphasize the potential of the SM14est to degrade plastics, and highlight the structural features that may facilitate this enzymatic activity.
Heterologous Expression of SM14est
To determine whether the SM14est gene does possess polyesterase activity, the gene was firstly codon-optimized for E. coli to facilitate heterologous expression in this host. The gene was then cloned into the expression vector pET-20b(+), generating the construct pET20b:SM14est, in which the native signal peptide sequence was maintained (Figure 1). The construct was then transformed into E. coli BL21(DE3) and transformants were tested for activity by performing a PCL plate clearing assay (Figure 6). A halo of clearing, which is indicative of PCL degradation, was observed following 1 day of incubation (Figure 6A), with the activity subsequently increasing after 2, 3, and 4 days (Figures 6B–D, respectively). The negative control with E. coli BL21(DE3) containing the pET-20b(+) plasmid without the insert did not show any PCL-degrading activity (data not shown).
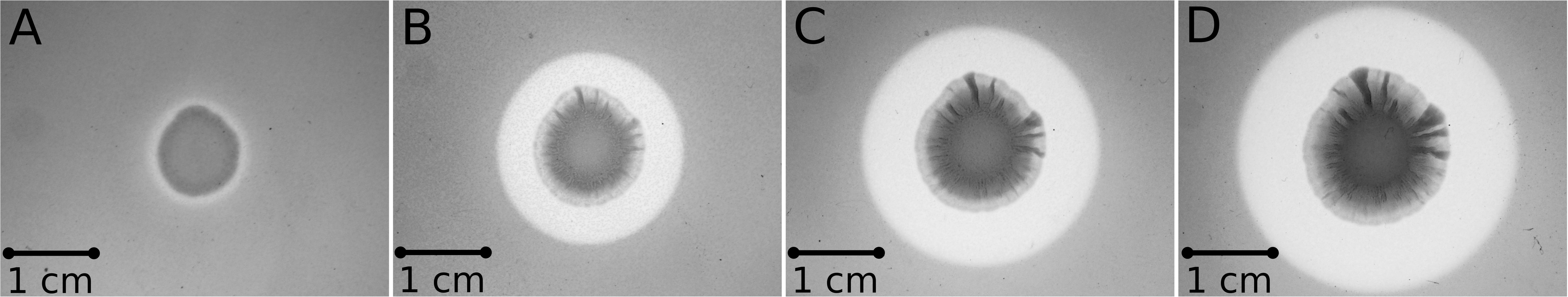
Figure 6. PCL plate clearing assay with the E. coli BL21(DE3)(pET20b:SM14est) heterologous host, incubated at 28°C after (A) 1 day; (B) 2 days; (C) 3 days; (D) 4 days.
Another interesting observation was that the E. coli host successfully exported the heterologously expressed SM14est enzyme, when the native Streptomyces sp. SM14 signal peptide sequence was present in the expression construct (Figures 1, 6). The signal peptide sequence was predicted to consist of the first 25 amino acids of the protein sequence, with a cleavage site probability of 0.9316, and to belong to the general secretory (Sec) pathway, with a likelihood value of 0.9608. In heterologous protein expression systems involving E. coli, successful secretion and maintenance of the native protein confirmation can sometimes be challenging, and in this case potentially so, due to the fact the SM14est protein originates from such a distant host, i.e., a Streptomyces isolate (Freudl, 2018). Several Streptomyces genes encoding different enzymes have previously been heterologously expressed in E. coli (Spasic et al., 2018) – including a xylanase from S. mexicanus HY-14 JQ943651 (Kim et al., 2014); a laccase from S. coelicolor (Sherif et al., 2013); a protease from S. koyangensis (Ben Elhoul et al., 2015); a glucose isomerase from Streptomyces sp. SK (Ben Hlima et al., 2013); and an esterase from S. lividans (Wang et al., 2016). In these cases however, unlike with the heterologous expression of the SM14est in E. coli, the native signal sequence was not employed in the expression constructs.
A number of different signal peptide sequences have previously been employed to ensure the secretion of PETase and PETase-like enzymes from E. coli. A PET carboxylesterase from Thermobifida fusca has been expressed and secreted from E. coli using a pelB leader sequence (Oeser et al., 2010), while a PET hydrolase has also previously been expressed and secreted from Bacillus subtilis using a native PETase signal peptide (SPPETase) (Huang et al., 2018). In addition, a Sec-dependent signal sequence from E. coli has also recently been used to express the IsPETase, resulting in the production of the extracellular enzyme using E. coli (Seo et al., 2019). The successful secretion of heterologously expressed PETases is important not only for their subsequent purification, and biochemical characterization (Han et al., 2017; Joo et al., 2018; Liu et al., 2018), but also for their potential development as recombinant PET-degrading microbes with utility in environmental remediation strategies. Therefore, it is interesting to note that we were able to efficiently heterologous express the PETase-like SM14est enzyme from Streptomyces sp. in E. coli, without the requirement to change its native signal peptide sequence, with extracellular synthetic polyester-degrading activity being observed in a PCL plate clearing assay. To our knowledge this is the first report of a PETase-like enzyme being identified in a marine sponge-derived Streptomyces sp. isolate, and we believe that the PETase-like SM14est enzyme will help provide further insights into our current knowledge of this important class of synthetic polyester-degrading enzymes.
Conclusion
Plastics such as the PET have been commonly used in storage materials and in synthetic fabrics, and their resistance to biodegradation has resulted in their accumulation in terrestrial and marine ecosystems at an alarming rate. In an attempt to alleviate this problem, much recent scientific interest has focused on the enzymatic hydrolysis of these types of synthetic polyesters, including PET. While a number of PETase and PETase-like enzymes have been identified and biochemically characterized, there is still much to be learned about this class of enzymes. In addition, more information on their structure, activity, and how widespread they are distributed in nature is required; and if they can ultimately be improved using genetic and protein engineering and applied in bioremediation strategies on an industrial scale.
Although the Streptomyces genus is well-studied with respect to the production of bioactive compounds, less is known about their potential to produce enzymes with synthetic polyester-degradation activities. In this study, based on an in silico screening approach, we were able to identify a PETase-like enzyme, namely SM14est, with synthetic polyester-degrading activity, which was isolated from the marine sponge-derived strain Streptomyces sp. SM14, with enzyme activity being confirmed in vitro with the heterologous expression of the protein in E. coli using PCL plate clearing assays. Importantly, an active heterologously expressed SM14est protein was secreted from E. coli with the native Streptomyces SM14est signal peptide sequence. This will facilitate not only the future biochemical characterization of the protein, but also its potential utility in other bioremediation-based applications targeting synthetic polyesters.
Data Availability Statement
The raw data supporting the conclusions of this manuscript will be made available by the authors, without undue reservation, to any qualified researcher.
Author Contributions
EA, AC, SJ, and AD conceived and designed the experiments. EA and AC performed the experiments. EA, AC, and AD analyzed the data. SJ and AD contributed reagents, materials, and analysis tools. EA and AD wrote the manuscript.
Funding
EA acknowledges the funding support of the Brazilian National Council for Scientific and Technological Development (CNPq). This project was supported by research grants from the Marine Institute through the NMBLI project (Grant-Aid Agreement PBA/MB/16/01), Marine Biotechnology ERA/NET, NEPTUNA project (contract no. PBA/MB/15/02), and Science Foundation Ireland (SSPC-2, 12/RC/2275).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2019.02187/full#supplementary-material
Footnotes
References
Almagro Armenteros, J. J., Tsirigos, K. D., Sønderby, C. K., Petersen, T. N., Winther, O., Brunak, S., et al. (2019). SignalP 5.0 improves signal peptide predictions using deep neural networks. Nat. Biotechnol. 37, 420–423. doi: 10.1038/s41587-019-0036-z
Almeida, E. L., Carillo Rincon, A. F., Jackson, S. A., and Dobson, A. D. W. (2019). Comparative genomics of the marine sponge-derived Streptomyces spp. isolates SM17 and SM18 with their closest terrestrial relatives provides novel insights into environmental niche adaptations and secondary metabolite biosynthesis potential. Front. Microbiol. 10:1713. doi: 10.3389/fmicb.2019.01713
Almeida, E. L., Margassery, L. M., Kennedy, J., and Dobson, A. D. W. (2018). Draft genome sequence of the antimycin-producing bacterium Streptomyces sp. strain sm8, isolated from the marine sponge haliclona simulans. Genome Announc. 6:e01535-17. doi: 10.1128/genomeA.01535-17
Altschul, S. F., Gish, W., Miller, W., Myers, E. W., and Lipman, D. J. (1990). Basic local alignment search tool. J. Mol. Biol. 215, 403–410. doi: 10.1016/S0022-2836(05)80360-2
Awaja, F., and Pavel, D. (2005). Recycling of PET. Eur. Polym. J. 41, 1453–1477. doi: 10.1016/J.EURPOLYMJ.2005.02.005
Balcão, V. M., and Vila, M. M. (2015). Structural and functional stabilization of protein entities: state-of-the-art. Adv. Drug Deliv. Rev. 93, 25–41. doi: 10.1016/j.addr.2014.10.005
Ben Elhoul, M., Zaraî Jaouadi, N., Rekik, H., Bejar, W., Boulkour Touioui, S., Hmidi, M., et al. (2015). A novel detergent-stable solvent-tolerant serine thiol alkaline protease from Streptomyces koyangensis TN650. Int. J. Biol. Macromol. 79, 871–882. doi: 10.1016/j.ijbiomac.2015.06.006
Ben Hlima, H., Bejar, S., Riguet, J., Haser, R., and Aghajari, N. (2013). Identification of critical residues for the activity and thermostability of Streptomyces sp SK glucose isomerase. Appl. Microbiol. Biotechnol. 97, 9715–9726. doi: 10.1007/s00253-013-4784-2
Benkert, P., Biasini, M., and Schwede, T. (2011). Toward the estimation of the absolute quality of individual protein structure models. Bioinformatics 27, 343–350. doi: 10.1093/bioinformatics/btq662
Benson, D. A., Cavanaugh, M., Clark, K., Karsch-Mizrachi, I., Ostell, J., Pruitt, K. D., et al. (2018). GenBank. Nucleic Acids Res. 46, D41–D47. doi: 10.1093/nar/gkx1094
Bertoni, M., Kiefer, F., Biasini, M., Bordoli, L., and Schwede, T. (2017). Modeling protein quaternary structure of homo- and hetero-oligomers beyond binary interactions by homology. Sci. Rep. 7:10480. doi: 10.1038/s41598-017-09654-8
Bianchetti, C. M., Harmann, C. H., Takasuka, T. E., Hura, G. L., Dyer, K., and Fox, B. G. (2013). Fusion of dioxygenase and lignin-binding domains in a novel secreted enzyme from cellulolytic Streptomyces sp. SirexAA-E. J. Biol. Chem. 288, 18574–18587. doi: 10.1074/jbc.M113.475848
Bienert, S., Waterhouse, A., de Beer, T. A. P., Tauriello, G., Studer, G., Bordoli, L., et al. (2017). The SWISS-MODEL repository-new features and functionality. Nucleic Acids Res. 45, D313–D319. doi: 10.1093/nar/gkw1132
Calabia, B. P., and Tokiwa, Y. (2004). Microbial degradation of poly (d-3-hydroxybutyrate) by a new Thermophilic Streptomyces isolate. Biotechnol. Lett. 26, 15–19. doi: 10.1023/B:BILE.0000009453.81444.51
Camacho, C., Coulouris, G., Avagyan, V., Ma, N., Papadopoulos, J., Bealer, K., et al. (2009). BLAST+: architecture and applications. BMC Bioinform. 10:421. doi: 10.1186/1471-2105-10-421
Chen, C.-C., Han, X., Ko, T.-P., Liu, W., and Guo, R.-T. (2018). Structural studies reveal the molecular mechanism of PETase. FEBS J. 285, 3717–3723. doi: 10.1111/febs.14612
Chen, S., Su, L., Billig, S., Zimmermann, W., Chen, J., and Wu, J. (2010). Biochemical characterization of the cutinases from Thermobifida fusca. J. Mol. Catal. B Enzym 63, 121–127. doi: 10.1016/J.MOLCATB.2010.01.001
Chertkov, O., Sikorski, J., Nolan, M., Lapidus, A., Lucas, S., Del Rio, T. G., et al. (2011). Complete genome sequence of Thermomonospora curvata type strain (B9). Stand. Genomic Sci. 4, 13–22. doi: 10.4056/sigs.1453580
Chevrette, M. G., Carlson, C. M., Ortega, H. E., Thomas, C., Ananiev, G. E., Barns, K. J., et al. (2019). The antimicrobial potential of Streptomyces from insect microbiomes. Nat. Commun. 10:516. doi: 10.1038/s41467-019-08438-0
Chua, T.-K., Tseng, M., and Yang, M.-K. (2013). Degradation of Poly (ε-caprolactone) by thermophilic Streptomyces thermoviolaceus subsp. thermoviolaceus 76T-2. AMB Express 3:8. doi: 10.1186/2191-0855-3-8
Danso, D., Schmeisser, C., Chow, J., Zimmermann, W., Wei, R., Leggewie, C., et al. (2018). New insights into the function and global distribution of polyethylene terephthalate (pet)-degrading bacteria and enzymes in marine and terrestrial metagenomes. Appl. Environ. Microbiol. 84:e02773-17. doi: 10.1128/AEM.02773-17
Dharmaraj, S. (2010). Marine Streptomyces as a novel source of bioactive substances. World J. Microbiol. Biotechnol. 26, 2123–2139. doi: 10.1007/s11274-010-0415-6
Drzyzga, O., and Prieto, A. (2019). Plastic waste management, a matter for the “community”. Microb. Biotechnol. 12, 66–68. doi: 10.1111/1751-7915.13328
Edgar, R. C. (2004). MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 32, 1792–1797. doi: 10.1093/nar/gkh340
England, J. L., and Haran, G. (2011). Role of solvation effects in protein denaturation: from thermodynamics to single molecules and back. Annu. Rev. Phys. Chem. 62, 257–277. doi: 10.1146/annurev-physchem-032210-103531
Food and Agriculture Organization of the United Nations (2017). Sponges and Their Role in The Marine Environment. Available at: http://www.fao.org/3/a-i7775e.pdf (accessed July 31, 2019).
Freudl, R. (2018). Signal peptides for recombinant protein secretion in bacterial expression systems. Microb. Cell Fact. 17:52. doi: 10.1186/s12934-018-0901-3
Geyer, R., Jambeck, J. R., and Law, K. L. (2017). Production, use, and fate of all plastics ever made. Sci. Adv. 3:e1700782. doi: 10.1126/sciadv.1700782
Godefroy, N., Le Goff, E., Martinand-Mari, C., Belkhir, K., Vacelet, J., and Baghdiguian, S. (2019). Sponge digestive system diversity and evolution: filter feeding to carnivory. Cell Tissue Res. doi: 10.1007/s00441-019-03032-8 [Epub ahead of print].
Guex, N., Peitsch, M. C., and Schwede, T. (2009). Automated comparative protein structure modeling with SWISS-MODEL and swiss-pdbviewer: a historical perspective. Electrophoresis 30, S162–S173. doi: 10.1002/elps.200900140
Han, X., Liu, W., Huang, J.-W., Ma, J., Zheng, Y., Ko, T.-P., et al. (2017). Structural insight into catalytic mechanism of PET hydrolase. Nat. Commun. 8:2106. doi: 10.1038/s41467-017-02255-z
Hantani, Y., Imamura, H., Yamamoto, T., Senga, A., Yamagami, Y., Kato, M., et al. (2018). Functional characterizations of polyethylene terephthalate-degrading cutinase-like enzyme Cut190 mutants using bis(2-hydroxyethyl) terephthalate as the model substrate. AIMS Biophys. 5, 290–302. doi: 10.3934/biophy.2018.4.290
Hassan, S. S., Anjum, K., Abbas, S. Q., Akhter, N., Shagufta, B. I., Shah, S. A., et al. (2017). Emerging biopharmaceuticals from marine actinobacteria. Environ. Toxicol. Pharmacol. 49, 34–47. doi: 10.1016/j.etap.2016.11.015
Hegde, K., and Veeranki, V. D. (2013). Production optimization and characterization of recombinant cutinases from Thermobifida fusca sp. NRRL B-8184. Appl. Biochem. Biotechnol. 170, 654–675. doi: 10.1007/s12010-013-0219-x
Herrero Acero, E., Ribitsch, D., Steinkellner, G., Gruber, K., Greimel, K., Eiteljoerg, I., et al. (2011). Enzymatic surface hydrolysis of PET: effect of structural diversity on kinetic properties of cutinases from Thermobifida. Macromolecules 44, 4632–4640. doi: 10.1021/ma200949p
Hotelier, T., Renault, L., Cousin, X., Negre, V., Marchot, P., and Chatonnet, A. (2004). ESTHER, the database of the α/β-hydrolase fold superfamily of proteins. Nucleic Acids Res. 32, 145–147. doi: 10.1093/nar/gkh141
Hsu, K. J., Tseng, M., Don, T. M., and Yang, M. K. (2012). Biodegradation of poly (β-hydroxybutyrate) by a novel isolate of Streptomyces bangladeshensis 77T-4. Bot. Stud. 53, 307–313.
Huang, X., Cao, L., Qin, Z., Li, S., Kong, W., and Liu, Y. (2018). Tat-independent secretion of polyethylene terephthalate hydrolase PETase in Bacillus subtilis 168 mediated by its native signal peptide. J. Agric. Food Chem. 66, 13217–13227. doi: 10.1021/acs.jafc.8b05038
Hwang, K. S., Kim, H. U., Charusanti, P., Palsson, B. Ø, and Lee, S. Y. (2014). Systems biology and biotechnology of Streptomyces species for the production of secondary metabolites. Biotechnol. Adv. 32, 255–268. doi: 10.1016/j.biotechadv.2013.10.008
Ian, E., Malko, D. B., Sekurova, O. N., Bredholt, H., Rückert, C., Borisova, M. E., et al. (2014). Genomics of sponge-associated Streptomyces spp. closely related to streptomyces albus j1074: insights into marine adaptation and secondary metabolite biosynthesis potential. PLoS One 9:e96719. doi: 10.1371/journal.pone.0096719
Irwin, J. J., and Shoichet, B. K. (2005). ZINC - A free database of commercially available compounds for virtual screening. J. Chem. Inf. Model. 45, 177–182. doi: 10.1021/ci049714+
Irwin, J. J., Sterling, T., Mysinger, M. M., Bolstad, E. S., and Coleman, R. G. (2012). ZINC: a free tool to discover chemistry for biology. J. Chem. Inf. Model. 52, 1757–1768. doi: 10.1021/ci3001277
Jackson, S., Crossman, L., Almeida, E., Margassery, L., Kennedy, J., and Dobson, A. (2018). Diverse and abundant secondary metabolism biosynthetic gene clusters in the genomes of marine sponge derived Streptomyces spp. Isolates. Mar. Drugs 16, 67. doi: 10.3390/md16020067
Jambeck, J. R., Geyer, R., Wilcox, C., Siegler, T. R., Perryman, M., Andrady, A., et al. (2015). Plastic waste inputs from land into the ocean. Science 347, 768–771. doi: 10.1126/science.1260352
Jamieson, A. J., Brooks, L. S. R., Reid, W. D. K., Piertney, S. B., Narayanaswamy, B. E., and Linley, T. D. (2019). Microplastics and synthetic particles ingested by deep-sea amphipods in six of the deepest marine ecosystems on Earth. R. Soc. Open Sci. 6:180667. doi: 10.1098/rsos.180667
Jin, J., Yang, X., Liu, T., Xiao, H., Wang, G., Zhou, M., et al. (2018). Fluostatins M–Q Featuring a 6-5-6-6 ring skeleton and high oxidized a-rings from marine Streptomyces sp. PKU-MA00045. Mar. Drugs 16:87. doi: 10.3390/md16030087
Joo, S., Cho, I. J., Seo, H., Son, H. F., Sagong, H.-Y., Shin, T. J., et al. (2018). Structural insight into molecular mechanism of poly (ethylene terephthalate) degradation. Nat. Commun. 9:382. doi: 10.1038/s41467-018-02881-1
Kawai, F., Kawabata, T., and Oda, M. (2019). Current knowledge on enzymatic PET degradation and its possible application to waste stream management and other fields. Appl. Microbiol. Biotechnol. 103, 4253–4268. doi: 10.1007/s00253-019-09717-y
Kawai, F., Oda, M., Tamashiro, T., Waku, T., Tanaka, N., Yamamoto, M., et al. (2014). A novel Ca2+-activated, thermostabilized polyesterase capable of hydrolyzing polyethylene terephthalate from Saccharomonospora viridis AHK190. Appl. Microbiol. Biotechnol. 98, 10053–10064. doi: 10.1007/s00253-014-5860-y
Kennedy, J., Baker, P., Piper, C., Cotter, P. D., Walsh, M., Mooij, M. J., et al. (2009). Isolation and analysis of bacteria with antimicrobial activities from the marine sponge Haliclona simulans collected from irish waters. Mar. Biotechnol. 11, 384–396. doi: 10.1007/s10126-008-9154-1
Kim, D. Y., Shin, D.-H., Jung, S., Lee, J. S., Cho, H.-Y., Bae, K. S., et al. (2014). Biocatalytic properties and substrate-binding ability of a modular GH10 β-1,4-xylanase from an insect-symbiotic bacterium, Streptomyces mexicanus HY-14. J. Microbiol. 52, 863–870. doi: 10.1007/s12275-014-4390-8
Kumar, S., Stecher, G., Li, M., Knyaz, C., and Tamura, K. (2018). MEGA X: molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 35, 1547–1549. doi: 10.1093/molbev/msy096
Lebreton, L., and Andrady, A. (2019). Future scenarios of global plastic waste generation and disposal. Palgrave Commun. 5:6. doi: 10.1057/s41599-018-0212-7
Liu, B., He, L., Wang, L., Li, T., Li, C., Liu, H., et al. (2018). Protein crystallography and site-direct mutagenesis analysis of the poly (ethylene terephthalate) hydrolase petase from Ideonella sakaiensis. Chem. Biol. Chem. 19, 1471–1475. doi: 10.1002/cbic.201800097
Lumry, R., and Eyring, H. (1954). Conformation changes of proteins. J. Phys. Chem. 58, 110–120. doi: 10.1021/j150512a005
Mabrouk, M. M., and Sabry, S. A. (2001). Degradation of poly (3-hydroxybutyrate) and its copolymer poly (3-hydroxybutyrate-co-3-hydroxyvalerate) by a marine Streptomyces sp. SNG9. Microbiol. Res. 156, 323–335.
Manteca, Á, and Yagüe, P. (2019). “Streptomyces as a source of antimicrobials: novel approaches to activate cryptic secondary metabolite pathways,” in Antimicrobials, Antibiotic Resistance, Antibiofilm Strategies and Activity Methods, ed. S. Kırmusaoğlu (London: IntechOpen).
Molitor, R., Bollinger, A., Kubicki, S., Loeschcke, A., Jaeger, K.-E., and Thies, S. (2019). Agar plate-based screening methods for the identification of polyester hydrolysis by Pseudomonas species. Microb. Biotech. 0, 1–11. doi: 10.1111/1751-7915.13418
Murphy, C. A., Cameron, J. A., Huang, S. J., and Vinopal, R. T. (1996). Fusarium polycaprolactone depolymerase is cutinase. Appl. Environ. Microbiol. 62, 456–460.
Nardini, M., and Dijkstra, B. W. (1999). α/β Hydrolase fold enzymes: the family keeps growing. Curr. Opin. Struct. Biol. 9, 732–737. doi: 10.1016/S0959-440X(99)00037-8
Nawaz, A., Hasan, F., and Shah, A. A. (2015). Degradation of poly (ε-caprolactone) (PCL) by a newly isolated Brevundimonas sp. strain MRL-AN1 from soil. FEMS Microbiol. Lett. 362, 1–7. doi: 10.1093/femsle/fnu004
Nishida, H., Suzuki, S., and Tokiwa, Y. (1998). Distribution of Poly (β-propiolactone) aerobic degrading microorganisms in different environments. J. Polym. Environ. 6, 43–58. doi: 10.1023/A:1022878512743
Nishida, H., and Tokiwa, Y. (1993). Distribution of poly (β -hydroxybutyrate) and poly (ε -caprolactone) aerobic degrading microorganisms in different environments. J. Environ. Polym. Degrad. 1, 227–233. doi: 10.1007/BF01458031
Nyyssölä, A., Pihlajaniemi, V., Järvinen, R., Mikander, S., Kontkanen, H., Kruus, K., et al. (2013). Screening of microbes for novel acidic cutinases and cloning and expression of an acidic cutinase from Aspergillus niger CBS 513.88. Enzyme Microb. Technol. 52, 272–278. doi: 10.1016/j.enzmictec.2013.01.005
Oeser, T., Wei, R., Baumgarten, T., Billig, S., Föllner, C., and Zimmermann, W. (2010). High level expression of a hydrophobic poly(ethylene terephthalate)-hydrolyzing carboxylesterase from Thermobifida fusca KW3 in Escherichia coli BL21(DE3). J. Biotechnol. 146, 100–104. doi: 10.1016/j.jbiotec.2010.02.006
Ohnishi, Y., Ishikawa, J., Hara, H., Suzuki, H., Ikenoya, M., Ikeda, H., et al. (2008). Genome sequence of the streptomycin-producing microorganism Streptomyces griseus IFO 13350. J. Bacteriol. 190, 4050–4060. doi: 10.1128/JB.00204-08
Ollis, D. L., Cheah, E., Cygler, M., Dijkstra, B., Frolow, F., Franken, S. M., et al. (1992). The α/β hydrolase fold. Protein Eng. Des. Sel. 5, 197–211. doi: 10.1093/protein/5.3.197
Pettersen, E. F., Goddard, T. D., Huang, C. C., Couch, G. S., Greenblatt, D. M., Meng, E. C., et al. (2004). UCSF Chimera–A visualization system for exploratory research and analysis. J. Comput. Chem. 25, 1605–1612. doi: 10.1002/jcc.20084
PlasticsEurope (2018). Plastics – the Facts 2018. PlasticsEurope. Available at: https://www.plasticseurope.org/en/resources/publications/619-plastics-facts-2018 (accessed May 14, 2019).
Ragaert, K., Delva, L., and Van Geem, K. (2017). Mechanical and chemical recycling of solid plastic waste. Waste Manag. 69, 24–58. doi: 10.1016/J.WASMAN.2017.07.044
Ribitsch, D., Acero, E. H., Greimel, K., Eiteljoerg, I., Trotscha, E., Freddi, G., et al. (2012a). Characterization of a new cutinase from Thermobifida alba for PET-surface hydrolysis. Biocatal. Biotransform. 30, 2–9. doi: 10.3109/10242422.2012.644435
Ribitsch, D., Herrero Acero, E., Greimel, K., Dellacher, A., Zitzenbacher, S., Marold, A., et al. (2012b). A new esterase from Thermobifida halotolerans hydrolyses polyethylene terephthalate (PET) and polylactic acid (PLA). Polymers 4, 617–629. doi: 10.3390/polym4010617
Ribitsch, D., Hromic, A., Zitzenbacher, S., Zartl, B., Gamerith, C., Pellis, A., et al. (2017). Small cause, large effect: structural characterization of cutinases from Thermobifida cellulosilytica. Biotechnol. Bioeng. 114, 2481–2488. doi: 10.1002/bit.26372
Robert, X., and Gouet, P. (2014). Deciphering key features in protein structures with the new ENDscript server. Nucleic Acids Res. 42, W320–W324. doi: 10.1093/nar/gku316
Sanner, M. F. (1999). Python: a programming language for software integration and development. J. Mol. Graph. Model. 17, 57–61.
Santillo, D., Miller, K., and Johnston, P. (2017). Microplastics as contaminants in commercially important seafood species. Integr. Environ. Assess. Manag. 13, 516–521. doi: 10.1002/ieam.1909
Seemann, T. (2014). Prokka: rapid prokaryotic genome annotation. Bioinformatics 30, 2068–2069. doi: 10.1093/bioinformatics/btu153
Seo, H., Kim, S., Son, H. F., Sagong, H.-Y., Joo, S., and Kim, K.-J. (2019). Production of extracellular PETase from Ideonella sakaiensis using sec-dependent signal peptides in E. coli. Biochem. Biophys. Res. Commun. 508, 250–255. doi: 10.1016/J.BBRC.2018.11.087
Ser, H.-L., Tan, L. T.-H., Law, J. W.-F., Chan, K.-G., Duangjai, A., Saokaew, S., et al. (2017). Focused review: cytotoxic and antioxidant potentials of mangrove-derived Streptomyces. Front. Microbiol. 8:2065. doi: 10.3389/fmicb.2017.02065
Sherif, M., Waung, D., Korbeci, B., Mavisakalyan, V., Flick, R., Brown, G., et al. (2013). Biochemical studies of the multicopper oxidase (small laccase) from Streptomyces coelicolor using bioactive phytochemicals and site-directed mutagenesis. Microb. Biotechnol. 6, 588–597. doi: 10.1111/1751-7915.12068
Shin, S. C., Ahn, D. H., Kim, S. J., Lee, H., Oh, T.-J., Lee, J. E., et al. (2013). Advantages of single-molecule real-time sequencing in high-gc content genomes. PLoS One 8:e68824. doi: 10.1371/journal.pone.0068824
Shivlata, L., and Satyanarayana, T. (2015). Thermophilic and alkaliphilic actinobacteria: biology and potential applications. Front. Microbiol. 6:1014. doi: 10.3389/fmicb.2015.01014
Silva, C., Da, S., Silva, N., Matama, T., Araujo, R., Martins, M., et al. (2011). Engineering Thermobifida fusca cutinase with increased activity on polyester substrates. Biotechnol. J. 6, 1230–1239. doi: 10.1002/biot.201000391
Spasic, J., Mandic, M., Djokic, L., and Nikodinovic-Runic, J. (2018). Streptomyces spp. in the biocatalysis toolbox. Appl. Microbiol. Biotechnol. 102, 3513–3536. doi: 10.1007/s00253-018-8884-x
Street, F. (2019). Deepest Ever Dive Finds Plastic Bag at Bottom of Mariana Trench. CNN. Available at: https://edition.cnn.com/travel/article/victor-vescovo-deepest-dive-pacific/index.html (accessed May 14, 2019).
Sulaiman, S., Yamato, S., Kanaya, E., Kim, J.-J., Koga, Y., Takano, K., et al. (2012). Isolation of a novel cutinase homolog with polyethylene terephthalate-degrading activity from leaf-branch compost by using a metagenomic approach. Appl. Environ. Microbiol. 78, 1556–1562. doi: 10.1128/AEM.06725-11
Taylor, M. W., Radax, R., Steger, D., and Wagner, M. (2007). Sponge-associated microorganisms: evolution, ecology, and biotechnological potential. Microbiol. Mol. Biol. Rev. 71, 295–347. doi: 10.1128/MMBR.00040-06
Tokiwa, Y., and Calabia, B. P. (2004). Review degradation of microbial polyesters. Biotechnol. Lett. 26, 1181–1189. doi: 10.1023/B:BILE.0000036599.15302.e5
Trott, O., and Olson, A. J. (2009). AutoDock vina: improving the speed and accuracy of docking with a new scoring function, efficient optimization, and multithreading. J. Comput. Chem. 31, 455–461. doi: 10.1002/jcc.21334
Wang, B., Wang, A., Cao, Z., and Zhu, G. (2016). Characterization of a novel highly thermostable esterase from the gram-positive soil bacterium Streptomyces lividans TK64. Biotechnol. Appl. Biochem. 63, 334–343. doi: 10.1002/bab.1465
Waterhouse, A., Bertoni, M., Bienert, S., Studer, G., Tauriello, G., Gumienny, R., et al. (2018). SWISS-MODEL: homology modelling of protein structures and complexes. Nucleic Acids Res. 46, W296–W303. doi: 10.1093/nar/gky427
Watve, M., Tickoo, R., Jog, M., and Bhole, B. (2001). How many antibiotics are produced by the genus Streptomyces? Arch. Microbiol. 176, 386–390. doi: 10.1007/s002030100345
Wei, R., Oeser, T., Then, J., Kühn, N., Barth, M., Schmidt, J., et al. (2014). Functional characterization and structural modeling of synthetic polyester-degrading hydrolases from Thermomonospora curvata. AMB Express 4:44. doi: 10.1186/s13568-014-0044-9
Wei, R., and Zimmermann, W. (2017a). Biocatalysis as a green route for recycling the recalcitrant plastic polyethylene terephthalate. Microb. Biotechnol. 10, 1302–1307. doi: 10.1111/1751-7915.12714
Wei, R., and Zimmermann, W. (2017b). Microbial enzymes for the recycling of recalcitrant petroleum-based plastics: how far are we? Microb. Biotechnol. 10, 1308–1322. doi: 10.1111/1751-7915.12710
Wei, Y., Swenson, L., Castro, C., Derewenda, U., Minor, W., Arai, H., et al. (1998). Structure of a microbial homologue of mammalian platelet-activating factor acetylhydrolases: Streptomyces exfoliatus lipase at 1.9 å resolution. Structure 6, 511–519. doi: 10.1016/S0969-2126(98)00052-5
Xu, X.-N., Chen, L.-Y., Chen, C., Tang, Y.-J., Bai, F.-W., Su, C., et al. (2018). Genome Mining of the marine actinomycete streptomyces sp. dut11 and discovery of tunicamycins as anti-complement agents. Front. Microbiol. 9:1318. doi: 10.3389/fmicb.2018.01318
Yoshida, S., Hiraga, K., Takehana, T., Taniguchi, I., Yamaji, H., Maeda, Y., et al. (2016). A bacterium that degrades and assimilates poly (ethylene terephthalate). Science 351, 1196–1199. doi: 10.1126/science.aad6359
Keywords: synthetic polyesters, polycaprolactone, PETases, Streptomyces, plastics
Citation: Almeida EL, Carrillo Rincón AF, Jackson SA and Dobson ADW (2019) In silico Screening and Heterologous Expression of a Polyethylene Terephthalate Hydrolase (PETase)-Like Enzyme (SM14est) With Polycaprolactone (PCL)-Degrading Activity, From the Marine Sponge-Derived Strain Streptomyces sp. SM14. Front. Microbiol. 10:2187. doi: 10.3389/fmicb.2019.02187
Received: 04 June 2019; Accepted: 05 September 2019;
Published: 01 October 2019.
Edited by:
Kian Mau Goh, University of Technology, Malaysia, MalaysiaReviewed by:
Giovanna Cristina Varese, University of Turin, ItalyNur Hazlin Hazrin-Chong, National University of Malaysia, Malaysia
Copyright © 2019 Almeida, Carrillo Rincón, Jackson and Dobson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alan D. W. Dobson, YS5kb2Jzb25AdWNjLmll