
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Microbiol. , 06 August 2019
Sec. Antimicrobials, Resistance and Chemotherapy
Volume 10 - 2019 | https://doi.org/10.3389/fmicb.2019.01690
This article is part of the Research Topic Antimicrobials and Anticancers of Bacterial Origins View all 11 articles
 Gisele Rodrigues1
Gisele Rodrigues1 Gislaine Greice Oliveira Silva2
Gislaine Greice Oliveira Silva2 Danieli Fernanda Buccini2
Danieli Fernanda Buccini2 Harry Morales Duque1
Harry Morales Duque1 Simoni Campos Dias1,3
Simoni Campos Dias1,3 Octávio Luiz Franco1,2*
Octávio Luiz Franco1,2*In recent decades, cancer and multidrug resistance have become a worldwide problem, resulting in high morbidity and mortality. Some infectious agents like Streptococcus pneumoniae, Stomatococcus mucilaginous, Staphylococcus spp., E. coli. Klebsiella spp., Pseudomonas aeruginosa, Candida spp., Helicobacter pylori, hepatitis B and C, and human papillomaviruses (HPV) have been associated with the development of cancer. Chemotherapy, radiotherapy and antibiotics are the conventional treatment for cancer and infectious disease. This treatment causes damage in healthy cells and tissues, and usually triggers systemic side-effects, as well as drug resistance. Therefore, the search for new treatments is urgent, in order to improve efficacy and also reduce side-effects. Proteins and peptides originating from bacteria can thus be a promising alternative to conventional treatments used nowadays against cancer and infectious disease. These molecules have demonstrated specific activity against cancer cells and bacterial infection; indeed, proteins and peptides can be considered as future antimicrobial and anticancer drugs. In this context, this review will focus on the desirable characteristics of proteins and peptides from bacterial sources that demonstrated activity against microbial infections and cancer, as well as their efficacy in vitro and in vivo.
In recent years, global health authorities have had to deal with two significant problems: the alarming number of people suffering from cancer and the rise of antimicrobial resistance (AMR). Cancer is the second most prevalent cause of death worldwide (O’Brien-Simpson et al., 2018; Shoombuatong et al., 2018). According to the World Health Organization (WHO) and the International Agency of Research on Cancer [IARC] (2018) in there were about 18.1 million new cases of cancer and 9.6 million deaths. The estimate for AMR is that 700,000 die annually worldwide, and the annual number of deaths is likely to increase to 10 million by 2050 (Arias and Murray, 2009; World-Health-Organisation [WHO], 2018; Ghosh et al., 2019).
Additionally, 16.1% of newly diagnosed cancers may be attributable to infections [National Cancer Institute – Epidemiology and Genomic Research Program1 (accessed March 12, 2019)]. Oncologic patients are more susceptible to infectious complications caused by Streptococcus pneumoniae, Stomatococcus mucilaginous, Staphylococcus spp., E. coli Klebsiella spp., Pseudomonas aeruginosa, Helicobacter pylori, and Candida spp. (Zorina and Styche, 2015; Rolston, 2017). Another concern related to cancer patients are infections caused by viruses such as hepatitis B and C, and human papillomaviruses (HPV) (Vedham et al., 2014; Rolston, 2017).
In fact, patients with a chronic infection induced by Staphylococcus aureus, Klebsiella pneumoniae, Acinetobacter baumanii, Pseudomonas aeruginosa, and Enterobacter species have been shown to have greater susceptibility to cancer development, as a result of their precarious immune system (Felício et al., 2017; Rolston, 2017). At the same time, antibiotics are used to prevent microbial infection in post-cancer surgery, and after chemotherapy or radiotherapy (Thundimadathil, 2012; Gaspar et al., 2013; Felício et al., 2017; Leite et al., 2018; O’Brien-Simpson et al., 2018; Shoombuatong et al., 2018). Conventional cancer treatments do not act on specific targets, such as malignant cells, resulting in severe side effects for patients, and these may contribute to the selection of cells that are resistant to antibiotics and anticancer drugs (Vedham et al., 2014; Zorina and Styche, 2015).
As a result, the development of a new class of molecules with selectivity and specificity against microbial infection and cancer is essential (Gaspar et al., 2013; Felício et al., 2017; Shoombuatong et al., 2018). Bacteria have an arsenal of proteins and peptides with both antibacterial and antitumoral activity, which can be explored in the search for these new compounds (Karpinski and Adamczak, 2018).
Among these promising molecules are toxins, immunotoxins (Jain, 2001; Gorgal et al., 2012), enzymes (Chakrabarty et al., 2014), bacteriocins (which are part of the same group as peptides) and a vast range of proteins (Karpinski and Adamczak, 2018). Antimicrobial peptides (AMPs) with anticancer activity can be classified according to the spectrum of their activity on tumor cells, and they are divided into two main categories: (i) peptides that show potent activity against bacteria and tumor cells, without causing damage to mammalian cells; (ii) peptides that are toxic to cancer cells, bacteria and healthy cells (Hoskin and Ramamoorthy, 2008; Bandala et al., 2013).
Therefore, this review will focus on the desirable characteristics of proteins and peptides originating from bacteria that demonstrated activity against microbial infections and cancer, as well as their efficacy in clinical trials, and will discuss future prospects.
Some proteins and peptides exhibit antimicrobial and anticancer activities. Therefore, bacteria use indirect and direct strategies to compete and survive. Host bacteria can show antimicrobial activity indirectly, by host immune system modulation (Belmadi et al., 2018). Alternatively, host bacteria can act directly by expressing proteins and peptides that are secreted to the extracellular environment and that target other bacteria (Cascales et al., 2007). Likewise, these proteins and peptides present variable structures linked to their activity, and these characteristics make their classification difficult (Daw and Falkiner, 1996; Vasilchenko and Valyshev, 2019). These structural diversities are represented in Figure 1, by toxins like colicin and pycin, and peptides represented by nisin, pediocin and plantaricin (Jain, 2001; Karpinski and Adamczak, 2018). Colicins are formed by three domains, the N-terminal, central and C- terminal, which act in membrane translocation, binding receptor and activity domain, respectively (Cascales et al., 2007). Pyocins are composed of four domains: domain I represents the receptor binding at the N-terminal, domain II has no defined function yet, domain III is responsible for translocation across the outer membrane and domain IV is responsible for DNase activity at the C-terminal end (Michel-Briand and Baysse, 2002). Nisin has two variants, A and Z, and the only difference between them is the change of His27 by Asn. They interact with the membrane surface in the C-terminal moiety (Lins et al., 1999). Pediocins interact with the target-cell surface in the N-terminal domain, and the C-terminal domain penetrates the membrane. Indeed, for pedicins, the domain is the major specificity determinant (Fimland et al., 2005). Plantaricin can present three variable forms, with 26-residue peptide and two N-terminal forms containing 23 and 22 residues; these forms result from a 48-residue precursor encoded by the plnA gene. Besides that, the amphiphilic nature of plnA can induced pore formation in cell membrane (Kaur and Kaur, 2015) (Figure 1).
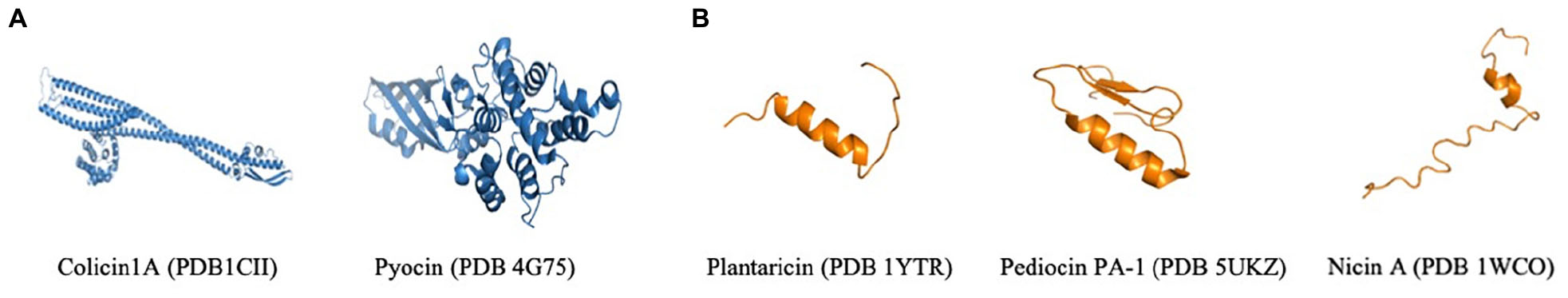
Figure 1. An overview of different structures of bacteriocins, proteins (A) and peptides (B) with dual anticancer and antimicrobial activity. All of these structures are available on the PDB.
In addition, several strategies could be designed to try to combat antimicrobial infection and carry out cancer therapy using proteins and peptides (Shilova et al., 2018). For example, proteins and peptides can be combined with conventional drugs (Liberio et al., 2013; Felício et al., 2017; Leite et al., 2018). They can be used as a heterologous compound infusion with other proteins or peptides that help in site-directed activity (Kawakami et al., 2006; Leshem and Pastan, 2019). It is possible to coat or conjugate proteins and peptides with polymers, such as polyethylene glycol (PEG) (Kelly et al., 2016). Another strategy is rational design for these molecules, because it then becomes possible to substitute naturally occurring amino acids with unnatural ones (Gordon et al., 2005; Uggerhøj et al., 2015).
In the various ways described above, these molecules have shown different applications and modes of action against antimicrobial infections and cancer. Nevertheless, in this study we focus on bacteriocins that have been previously characterized and/or synthesized with dual activity (Figure 2).
Figure 2. Distribution of described anticancer and antimicrobial proteins and peptides.
Bacteriocins make up a class of molecules (proteins and peptides) synthesized by ribosomes in several Gram-positive and negative bacteria. In 1925, the first bacteriocin isolated from E. coli discovered by Gratia (1925), was identified and later named colicin. Since then, several bacteriocins have been discovered in a variety of bacteria (Kaur and Kaur, 2015). Cationic peptides belonging to the bacteriocin class may be associated with biological functions in bacteria and may assist in the inhibition and elimination of possible competing microorganisms in their natural environment (Nes and Holo, 2000).
As demonstrated above, proteins and peptides have variable structures (Vasilchenko and Valyshev, 2019). Regarding classification, which is a challenge, it is known that 80% of the bacteriocins are of cationic and amphipathic nature, due mainly to the excess of residues of lysine or arginine amino acids (Nes and Holo, 2000; Hammami et al., 2010). These characteristics of bacteriocins may be linked to their efficacy in combating cancer cells, possibly by the interaction of these cationic molecules with the negative surface charge of the cancerous cell membrane (Saito et al., 1979; Baindara et al., 2015). A visible example of such characteristics can be observed in lantipeptides, a class I bacteriocin, which can form pores, resembling cationic antimicrobial peptides (cAMPs) (Smith and Hillman, 2008). In addition, some bacteriocins produced by Gram-positive bacteria can resemble the defensins, AMPs produced by eukaryotic cells, which have membrane-permeabilizing characteristics (Singh et al., 2015).
Another relevant feature of bacteriocins is their low toxicity when in contact with mammalian cells. Due to these characteristics, bacterial strains producing bacteriocins have been implemented in foods as probiotics (Cutter and Siragusa, 1998). Their low toxicity has already been demonstrated in studies, for example, a bacteriocin named laterosporulin did not show hemolysis in the presence of erythrocytes, even at concentrations exceeding the minimum inhibitory concentration (MIC) values (Singh et al., 2015). The same can be observed in studies with penisin, a class IA lantibiotic, which in addition to not being hemolytic showed no cytotoxic activity against mouse macrophages (RAW) (Baindara et al., 2015). These characteristics are important from the point of view of the development of new therapies, since their affinity with tumor cells and low toxicity show that the bacteriocins are excellent candidates for the treatment of cancer, besides presenting antibacterial activities (Chakrabarty et al., 2014; Yang et al., 2014; Kaur and Kaur, 2015; Karpinski and Adamczak, 2018).
In this section, we will address the issue of protein and peptide bacteriocins isolated from bacteria. Only those molecules with a molecular mass of 10 kDa or less will be classified as peptides. Among the bacteriocins studied to date, some have been submitted to tests with tumor cells. Table 1 lists the bacteriocins that have been evaluated so far and have demonstrated activity against bacterial and cancer cells (Figure 3). These activities are divided into membrane adsorption (Figure 3A) and conformational change (Figure 3B). Membrane adsorption is represented by carpet model (in which the formation of membrane micellar structures occurs in an area with high peptide densities); barrel-stave model (protein and peptides form a pore in a perpendicular orientation to the membrane surface and interact with the phospholipid acyl chains); toroidal pore model (the contact of protein and peptides with the phospholipid head groups during the pore formations causes expanding membrane curvature); disordered toroidal pore (which promotes the internal curvature of lipid molecules for pore formation with few peptides; this process occurs in randomic form); non-bilayer intermediate (the peptide aggregation on the membrane surface activates the formation of the intermediate bilayer, causing membrane disruption, allowing the peptide to be translocated into the internal leaflet); membrane thinning/thickening (originates with peptides aligned parallel to the surface of the membrane, provoking thinning or thickening due to hydrophilic characteristics associated with the lipid bilayer) (Lohner, 2009; Haney et al., 2010; Nguyen et al., 2011; Gaspar et al., 2013; Gabernet et al., 2016). The conformational charge is represented by anion carrier (this model acts directly on the membrane through lipid isolation or addresses oxidized phospholipids, or even acts as a small anion carrier through the membrane); charge lipid cluster (cationic peptides interact with clustering of anionic lipids in the membrane region, and this interaction allows pore formation); electroporation (occurs subtly, when proteins or peptides bind small anions across the bilayer, inducing the efflux and changes in the membrane potential); non-lytic membrane depolarization (proteins and peptides induce charge modification, cause membrane instability, and allow the proteins and peptides to translocate across the cytoplasmic membrane); and oxidized lipid targeting [stimulating the formation of cellular reactive oxygen species (ROS) causing oxidative cellular damage, developing metabolites that can be mutagenic, cytotoxic and also promote cellular aging and apoptosis] (Gifford et al., 2005; Epand and Epand, 2011; Nguyen et al., 2011; Hasegawa et al., 2012; Gaspar et al., 2013; Gabernet et al., 2016; Alexander et al., 2019).
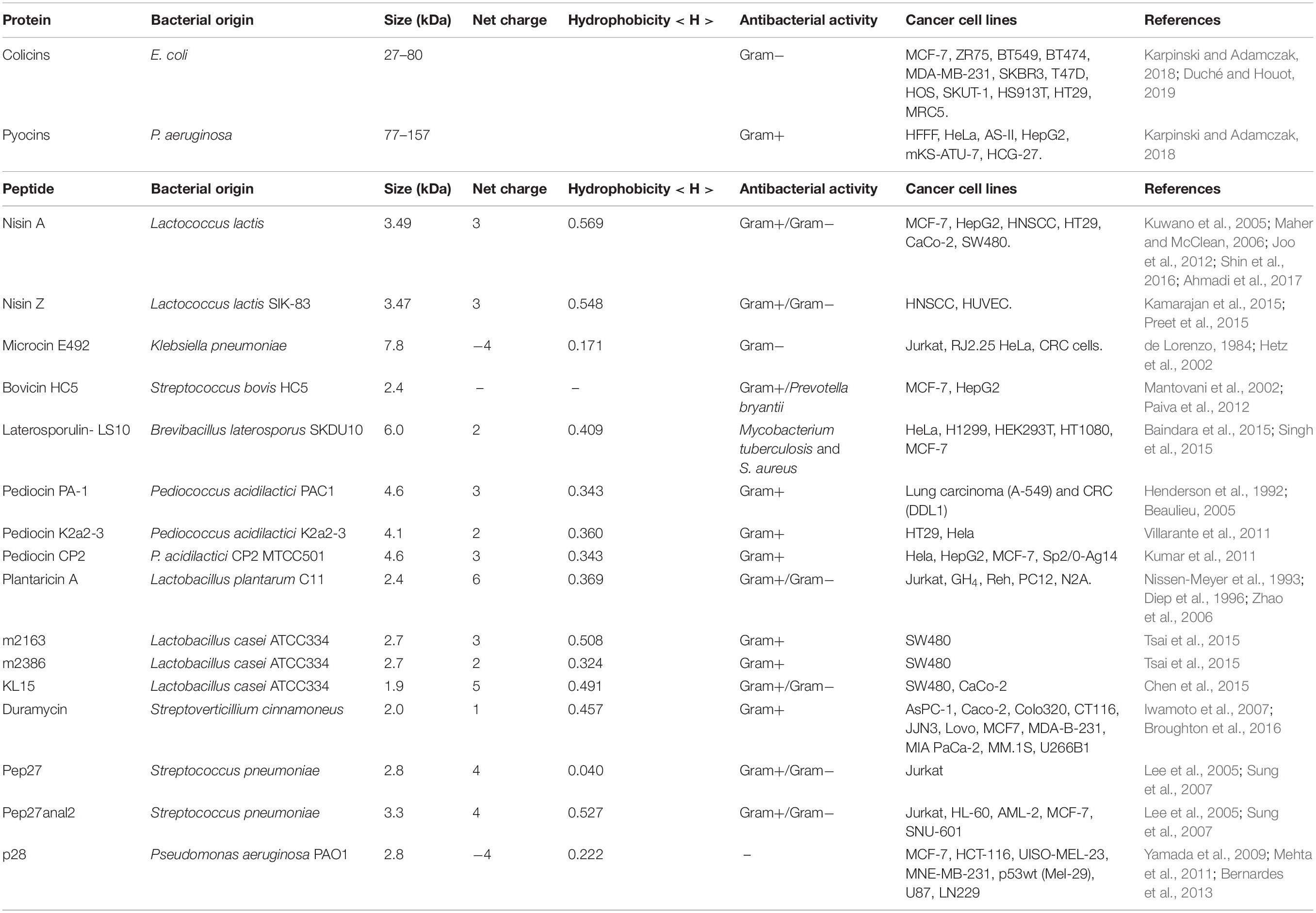
Table 1. Anticancer and antimicrobial activity of proteins and peptides from bacterial origins.
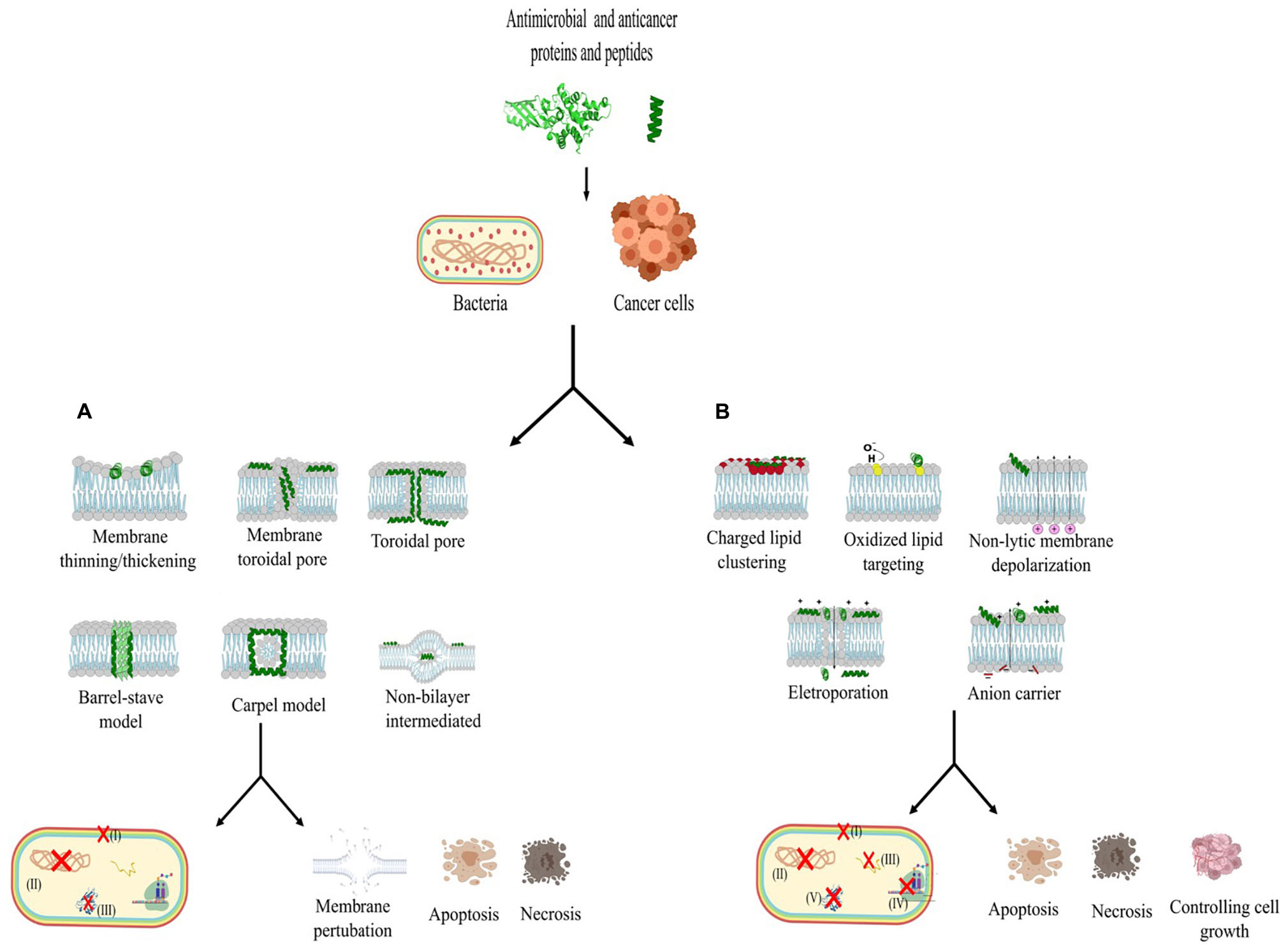
Figure 3. Different mechanisms of action of proteins and peptides with anticancer and antibacterial activity. (A) This section represents the protein and peptide interaction with the carpet model, barrel-stave model, toroidal pore, disorder toroidal pore, non-bilayer intermediate and membrane thinning/thickening, which are mechanisms that act in the conformation of the external membrane (I), target the inhibition of phospholipase, lipid II and LPS permeabilization. In the inner membrane, these mechanisms act in DNA synthesis (II) and inhibition of fold protein (III). (B) This section represents the action of proteins and peptides interaction the other mechanisms described above, such as anion carrier, charged lipid clustering, electroporation, non-lytic membrane depolarization, and oxidized lipid targeting, act in the destabilization of membrane components (I): in addition, they act in the inner membrane by interrupting DNA (II), RNA(III), and protein syntheses (IV) and (V). All these mechanisms can cause bacterial death and, in cancer cells, can cause disruption of membrane, apoptosis, necrosis, and control of angiogenesis. The model described in (A) and (B) acting in bacteria and cancer cells.
There are a small number of proteins classified as bacteriocins, including colicin, pyocin, pesticin, butyricin, and megacin (Konisky, 1982). These proteins are used by bacterial species for intraspecies competition (Di Masi et al., 1973; Chai et al., 1982; Michel-Briand and Baysse, 2002; Spector et al., 2010; Yang et al., 2014). However, only colicin and pyocin have been reported in the literature as presenting dual activity.
Colicins are proteins produced by E. coli, and they present a diversity of molecules (E1-3, K, A, L, B, Ia, Ib, V, D, and M) with varied masses between 27 and 80 kDa (Konisky, 1982). The structure of colicin proteins presents three domains; these are the receptor-binding domain responsible for target interaction, the N-terminal domain that mediates interaction with transporters, and the cytotoxic domain that allows antibacterial activity (Vasilchenko and Valyshev, 2019). Colicins act by binding to the outer membrane of integral membrane protein receptors, transporting the colicin to the inner membrane through Tol complex, inducing membrane depolarization, and degradation of DNA, ribosomal RNA, or tRNA (Duché and Houot, 2019) (Figure 3B). Colicins show activity on a wide bacterial range (Duché and Houot, 2019), and they also showed cytotoxic activity on breast carcinoma, fibrosarcoma, leiomyosarcoma, osteosarcoma, and colon carcinoma (Bandala et al., 2013; Karpinski and Adamczak, 2018). In this sense, a study using the different isolates of colicins, E1, E6, E7, K, and M, was evaluated against E. coli strains from patients with bacteraemia of urinary tract origin. The colicin isolates were tested in concentrations of 1, 10, and 100 μg/ml. The results showed that colicin E7 was able to kill 87% of the strains tested. The results showed that colicin E7 at 10 μg. ml–1 was able to kill 87% of strains tested. The inhibitory activity of combinations of colicins E7, K, M and E1, E6, E7, K effectively inhibited the growth of pathogenic E. coli strains (Budič et al., 2011). Furthermore, colicin presents anticancer activity. This activity was elucidated by Chumchalova and Šmarda (2003), in a study that used four colicins (A, E1, U, E3) against 11 cancer cell lines. They detected anticancer activity in most of cell lines tested. Although fibrosarcoma HS913T has demonstrated 50% inhibition with colicin E1, it was also shown to be sensitive to colicin A and U. Only colicin E3 did not exhibit significant inhibition activity against tested cancer cells. Another study demonstrated the efficiency of colicin E3 against murine leukemia P388cells. It tested colicin E3 in final concentrations of 0.4 mg. ml–1, 0.8 mg. ml–1, 1.6 mg. ml–1, and 3.2 mg. ml–1 for 0, 24, 48 and 72-h cultivations at 37°C. The results demonstrated the colicin E3 was able to inhibit the proliferation of murine leukemia cells in a time- and dose-dependent manner (Fuska et al., 1979).
Pyocins originate from P. aeruginosa (Ghequire and De Mot, 2018); their size varies between 77 and 157 amino acid residues (Ghequire and De Mot, 2014). Three different types of pyocins are described, namely the R, F, and S type. The R-type pyocins connect to the outer membrane receptor and induce the depolarization of the cytoplasmic membrane in relation to pore formation (Figure 3A). S-type or colicin-like pyocins show endonuclease activity when placed at the C-terminal end, and this activity induces cell death by interrupting DNA, RNA and protein syntheses (Michel-Briand and Baysse, 2002) (Figure 3B). These proteins act on Gram-negative bacteria (McCaughey et al., 2016), and demonstrated activity against embryonal ovary carcinoma, human hepatocellular carcinoma, and cervical adenocarcinoma (Karpinski and Adamczak, 2018).
Thus, Ling et al. (2010) studied the pyocin S5 in different concentrations (3.5, 14, 56, and 225 μg. ml–1) against seven clinical P. aeruginosa isolates (DWW3, InA, InB, In3, In4, In7, and In8). Their results indicated that isolate DWW3 is most sensitive in an inhibitory concentration of 12.6l μg.ml–1, and the other isolates were killed in a concentration of 225 μg. ml–1 isolates (Ling et al., 2010). Additionally, R-type pyocins were tested in a lethal mouse peritonitis model. The researchers infected female mice (Charles River Laboratories) with 0.5. ml–1 of inoculum of P. aeruginosa strain 13s per mouse, in concentrations of 106 to 107 CFU. ml–1. The treatment with 0.1. ml–1 of pyocins was administered intraperitoneally (i.p.) and intravenously (i.v.). At 24 and 48 h after treatment inoculation, blood and spleen samples were evaluated. The result showed that i.p. and i.v. dose responses were similar, and pyocin killed approximately 99.99% of the bacteria in the blood and spleen samples. This study suggested that R-type pyocins could be an effective therapy (Scholl and Martin, 2008).
A different test using purified pyocin S2 and partially purified pyocin from P. aeruginosa 42A was evaluated against human tumor cell lines, HepG2 and Im9, and the normal human cell line HFFF (Human Fetal Foreskin Fibroblast). Pyocin S2 and partially purified pyocin were tested at concentrations of 6.25, 12.25, 25, and 50 U. ml–1, for 5 days. The results showed cell line Im9 was more sensitive than HepG2, and maximum growth inhibition of 80% was observed at the maximum pyocin concentration of 50 U. ml–1 after 5 days (Abdi-Ali et al., 2004). According to the examples cited above, the colicins and pyocins demonstrated efficacious antimicrobial and anticancer activity.
Nisin can be secreted by Lactococcus lactis subspecies lactis and has a low molecular mass of 3.49 kDa with 34 amino acid residues. This polycyclic peptide belongs to a class of lantibiotics, which have unusual amino acids such as lanthionines (Moll et al., 1997; de Arauz et al., 2009; Shin et al., 2016). Nisin exhibits stability at different temperatures and has potent activity against different bacteria, including pathogenic strains, acting primarily on Gram-positive bacteria (Jack et al., 1995; Severina et al., 1998). In view of this, nisins are classified as broad-spectrum peptides, presenting activity in different classes of bacteria (Shin et al., 2016). In addition, nisins share some similarities with pore-forming AMPs, such as positive net charge and amphipathicity. These peptides may exhibit different mechanisms of activity that may involve interaction with membranes facilitated by lipid II binding (Christ et al., 2007). This can lead to the formation of pores in the membrane (Moll et al., 1997) (Figure 3). Due to its low toxicity, this molecule has been used for a long time in food preservation (Gharsallaoui et al., 2016). In addition to its antibacterial activity, nisin and its natural variants have been shown to be effective in combating cancerous cells (Joo et al., 2012; Kamarajan et al., 2015).
In this sense, several studies have been carried out to demonstrate the efficiency of nisin against microorganisms. One of these studies purified nisin Z, which exhibits antimicrobial activity against S. aureus and E. coli. However, the test using nisin Z and 100 mM NaCl demonstrated activity just for S. aureus (Kuwano et al., 2005). Another study described the nisin activity against cariogenic microorganisms (Streptococcus spp., Lactobacillus spp., Actinomyces spp.). According to the researchers, electron microscopy showed that nisin exerted bactericidal activity by forming small pores on the surface of cells (Tong et al., 2010) (Figure 3A).
In addition, we described studies using nisin with anticancer activity. Joo et al. (2012) demonstrated that nisin decreases head and neck squamous cell carcinoma (HNSCC) tumorigenesis in vitro and in vivo by inducing increased cell apoptosis and decreased cell proliferation. For the in vitro test, the researchers used a concentration of 5, 10, 20, 40, and 80 μg.ml–1 nisin against three different HNSCC cell lines, and after 24 h of treatment they observed increased levels of DNA fragmentation or apoptosis. In vivo tests were evaluated with a 150 mg/kg dose of nisin administered over the course of 3 weeks. They observed that mice that had received nisin treatment exhibited statistically significant reduced tumor volumes compared with control. These effects are associated with the activation of CHAC1, broadening calcium influxes and inducing cell cycle arrest (Joo et al., 2012).
Another group also tested the efficiency of nisin Z for the treatment of HNSCC in vitro and in vivo. They used nisin Z at different concentrations (0, 100, 400, and 800 μg.ml–1) against normal-human umbilical vein endothelial cells (HUVECs) in an in vitro test. In in vivo tests, they used an oral cancer floor-of-mouth mouse model. The treatment started after confirmation that tumors were established, using a control group that was given water (equal volume/control), and the treatment groups were treated with nisin, at two different concentrations (400 and 800 mg/kg per day) by oral gavage for 3 weeks. The researchers observed that the in vitro test demonstrated notably increasing levels of apoptosis when compared to cells treated with control medium. Furthermore, the concentrations of nisin tested in vivo significantly decreased the tumor volumes (13.5 and 88.5 mm3 for nisin ZP 800 mg/kg, nisin ZP 400 mg/kg, respectively) compared to controls (232.8 mm3). The authors concluded that nisin is an alternative therapy for HNSCC, exhibiting HNSCC cell apoptosis, suppressing HNSCC cell proliferation, inhibiting angiogenesis, HNSCC and tumorigenesis in vivo (Kamarajan et al., 2015). Different studies demonstrated the capacity of nisin to reduce, control and increase the apoptosis of distinct types of tumor cells (Preet et al., 2015; Ahmadi et al., 2017).
In addition, bovicin HC5 is a peptide with a strong similarity to the structure and function of nisin. This peptide can cause cell membrane disruption through pore formation and by modifying cellular potassium efflux (Figure 3A). It is a lantibiotic secreted by Streptococcus bovis HC5 and presents about 2.4 kDa of molecular mass. It can be considered as an AMP with a broad spectrum of activity on Gram-positive and negative bacteria. It is considered a stable molecule at high temperatures and low pH, but it may undergo loss of activity in the presence of pronase E and trypsins (Mantovani et al., 2002). Bovicin HC5 has already demonstrated anticancer activity against human adenocarcinoma cells (MCF-7) and hepatocellular carcinoma (HepG2) (Paiva et al., 2012).
Bacteria belonging to the Enterobacteriaceae family are responsible for the production of microcin. These peptides can have a molecular mass up to 10 kDa, and they present activity on different strains of pathogenic bacteria, such as Salmonella, Enterobacter, Klebsiella, Escherichia, and Citrobacter (de Lorenzo, 1984). The microcins are classified in two classes: class I is represented by peptides with molecular masses <5 kDa, being codified by cluster genes located either on plasmids or on the chromosome, like microcins B17, C (or C7, C51 or C7/C51), and J25.6. Class II is represented by higher molecular mass peptides ranging from 5- to 10-kDa. Additionally, they are subdivided into two subclasses, IIa and IIb. Class IIa presents three plasmid-encoded peptides, microcins L, V, 6 and N. They do not have post-translational modifications and can present two, one, and no disulfide bond(s), respectively. Class IIb is described as linear microcins with post-translational modification represented by microcins E492, M, H47, and presumably I47 and G492 (Rebuffat, 2013). The microcin act taking advantage of iron-siderophore receptors (FepA, Cir, Fiu). After receptor interaction, microcin was translocated though the TonB-ExbB-ExbD inner-membrane protein complex (Morin et al., 2011; Rebuffat, 2013). Thus, microcin induces depolarization of the cytoplasmic membrane, reaching a specific molecular target (de Lorenzo, 1984; de Lorenzo and Pugsley, 1985; Lagos et al., 1993; Hetz et al., 2002; Morin et al., 2011; Rebuffat, 2013) (Figure 3A).
The efficiency of microcin was studied by Yu et al. (2018). They tested microcin (MccJ25) against the ETEC K88 (serotype O149:K91, K88ac) strain, which is a pathogen related with human infants and neonatal diarrhea. They used MccJ25 in 0.125 to 256 μg/mL concentrations. The results demonstrated the efficacy of MccJ25 at a concentration of 0.25 μg. ml–1 against ETEC K88. They observed that MccJ25 was not cytotoxic, and that is was also able to protect the intestine against ETEC K88-induced damage and inflammatory response. According to the authors, MccJ25 can be used as a novel prophylactic agent to reduce pathogenic infection in animals, food or humans.
Furthermore, the efficacy of microcin with in vitro tumor cells was described by Hetz et al. (2002). They studied microcin E492 produced by Klebsiella pneumoniae strain RYC492 with 7,887 Da. This peptide was tested against some human tumor cells such as Jurkat, RJ2.25, colorectal carcinoma (CRC) and HeLa to observe the capacity to induce apoptosis. The authors tested microcin E492 at different concentrations of 5, 10, and 20 μg/ml. The result demonstrated that at a concentration of 5 μg. ml–1 microcin induced tumor cells to apoptosis, and at concentrations of 10 and 20 μg. ml–1 necrosis was observed (Hetz et al., 2002).
Laterosporulins (LS) are peptides with low molecular masses (approximately 5.6 kDa) that present a strong similarity to defensins (Singh et al., 2015). This class of peptides exhibits amphiphilic helical structure, which permits laterosporulin to insert itself into the membrane of the target cell, inducing depolarization and death (Kaur and Kaur, 2015) (Figure 3A). These peptides were isolated from different strains, namely GI-9 and SKDU10, from Brevibacillus sp. and have showed potent antibacterial activity against several pathogens (Singh et al., 2015). One example is laterosporulin 10 (LS10) isolated from Brevibacillus SKDU10, with 6 kDa, which was tested at different concentrations of 4, 8, and 20 μM against Staphylococcus aureus and Mycobacterium tuberculosis (MtbH37Rv). The result showed that LS10 inhibited the growth of S. aureus with LD50 of 4.0 μM and M. tuberculosis (Mtb H37Rv) with LD50 of 0.5 μM. In addition, microscopic studies demonstrated that LS10 acts on S. aureus cell membrane and the Mtb H37Rv strain by disrupting cellular metabolic homeostasis. LS10 was able to alter the membrane of the Mtb H37Rv strain, which has a thick lipid layer (Baindara et al., 2016). Moreover, regarding LS10 antitumor activity, this peptide was tested in the concentrations 1–20 μM against HeLa, H1299, HEK293T, HT1080, MCF-7, and RWPE-1 cells. The results demonstrated that LS10 showed activity against diverse cancer cells like MCF-7, HEK293T, HT1080, HeLa, and H1299 in low concentrations (10 μM), but failed against RWPE-1 cells. Moreover, the LS10 at a concentration of 2.5 μM induced apoptosis (Baindara et al., 2018).
Pediocins originate from bacteria that produce lactic acid, mainly species of Pediococcus (Kumar et al., 2011). A variety of pediocins have been identified so far, namely, pediocin CP-2, F, K1, AcH, AcM SJ-1, and L50, some of which are cited in an extensive review by Porto et al. (2017). They can be considered as small plasmid-encoded cationic AMPs (>5 kDa), with high stability at a variety of temperatures and pHs. However, they may undergo actions of different proteolytic enzymes such as trypsin, α-trypsin, pepsin, papain, and proteases (Kumar et al., 2011). Their N-terminal region contains the conserved Y-G-N-G-V/L “pediocin box” motif and two conserved cysteine residues that are joined by a disulfide bridge, which forms a three-stranded antiparallel beta-sheet (Papagianni and Anastasiadou, 2009). Pediocins contain a conserved N-terminal region folded by disulfide bonds, and this domain mediates binding of class IIa bacteriocins to the target cell membrane. In contrast, the C-terminal region forms a hairpin that is able to penetrate the target cell membrane hydrophobic region, thereby mediating leakage through the membrane (Fimland et al., 2005; Drider et al., 2006) (Figure 3A). Pediocins have showed antimicrobial activity. According to Bédard et al. (2018), they synthesized pediocin PA-1 and demonstrated that it is a potent inhibitor of Listeria monocytogenes (MIC = 6.8 nM), similar to that produced naturally by Pediococcus acidilactici. Pediocin PA-1 was also tested against different strains of Carnobacterium divergens ATCC 35677 (MIC = 1.9 nM), Leuconostoc mesenteroides ATCC 23386 (MIC = 1.9 nM), Listeria seeligeri ATCC 35967 MIC = 4.7 nM), Clostridium perfringens AAC 1–222 (MIC = 37.8 nM), Clostridium perfringens AAC 1–223 (MIC = 75.7 nM), Listeria murrayi ATCC 25401 (MIC = 151.4 nM), and Lactobacillus plantarum ATCC 8014 (MIC = 605.5 nM). Studies have shown the antitumor activity of some pediocins, as exemplified by PA-1, a pediocin produced by P. acidilactici PAC1. In the presence of human lung carcinoma cells and colorectal adenocarcinoma, PA-1 inhibited the growth of these cells. Pediocin PA-1 isolated from P. acidilactici K2a2-3 has also been shown to be cytotoxic to human colon adenocarcinoma HT29 and HeLa cells, but the mechanism of cytotoxicity has not been studied (Villarante et al., 2011). As also observed, pediocin CP2 produced by P. acidilactici CP2 MTCC501 has antitumor activity on HEPg2, HeLa and MCF-7 human cancer cells (Kumar et al., 2011).
The peptide plantaracin (Pln) is produced by different strains of Lactobacillus plantarum (C11, WCFS1, V90), showing low molecular mass (∼2.4 kDa) (Diep et al., 1996). The amphiphilic nature of the Pln peptide (class IIb), could facilitate the formation of membrane channels (Nissen-Meyer et al., 1993) (Figure 3A). In fact, Pln has already been shown to permeate eukaryotic cells, but demonstrates affinity to negatively charged membranes and exhibits strong interaction with glycolate membrane proteins (Sand et al., 2013). Some works have demonstrated a broad-spectrum activity of these peptides on different bacterial strains (Nissen-Meyer et al., 1993; Diep et al., 1996). A recent study demonstrated the antimicrobial activity of plantaricins (Pln) A, E, F, J, and K against Staphylococcus epidermidis. The plantaricins alone were tested at a concentration of 0.097 to 50 μM, and the plantaricins in association with the antibiotics were tested at concentrations12.5 and 6.25 μM. The results showed that S. epidermidis was more susceptible to PlnEF than PlnJK, with MIC 12.5 and 25 μM, respectively. PlnE, F, J and K inhibited bacterial growth, and PlnEF and PlnJK, at 25 and 50 μM, caused rapid bacterial lysis (data not shown). In addition, PlnA (50 μM) alone repressed bacterial growth. Pln in combination with low concentrations of antibiotics displayed antimicrobial activity against S. epidermidis. According to results, Pln in combination with antibiotics in low concentration was efficient against S. epidermidis and exhibited strong potential to treat infections (Selegård et al., 2019). Pln also demonstrated activity on cancer cells (Zhao et al., 2006; Sand et al., 2010, 2013). This activity was shown in a study by Sand et al. (2010), in which they studied the effect of synthesized PlnA against normal human B and T cells, Reh cells (from human B cell leukemia), and Jurkat cells (from human T cell leukemia). The cell types were tested at a concentration of 10–100 μM PlnA. The results showed that all cells were affected by PlnA, but at low concentrations (10 μM) this did not demonstrate a strong effect. The mechanism of action was seen to be membrane permeabilization, leading to apoptosis along with necrosis.
Duramycin is a type of lantibiotic produced by streptomycetes. This tetracyclic peptide is synthesized by ribosomes and exhibits post-translational changes, as well as possessing antimicrobial activity. It consists of 19 amino acid residues corresponding to a molecular mass of ∼2 kDa (Phoenix et al., 2015; Broughton et al., 2016). The post-translational changes undergone by duramycin, such as the enzymatic addition of three thioether bonds, besides increasing the proteolytic stability of this molecule, also provide selectivity and binding to phosphatidylethanolamine (PE) present on the membrane of various cell types, including Gram-positive and negative bacteria (Iwamoto et al., 2007). The interaction of duramycin with target cells can trigger plasmatic membrane imbalance, affecting membrane integrity and influencing the ion transportation mediated by pore formation on the surface of the cell membrane (Sheth et al., 1992; Oliynyk et al., 2010) (Figure 3A).
Other effects of duramycin on the plasma membrane have been shown, such as an inhibitory effect on plasma membrane ATPase activity (Nakamura and Racker, 1984), an increase in cell membrane permeability, and inhibition of Na+-K+-ATPase in the cellular plasma membranes of Ehrlich ascites tumor cells (Nakamura and Racker, 1984). Due to these characteristics, the antineoplastic capacity of duramycin has been studied. Since the membrane surface of some cancer cells is positive for PE, the effectiveness of duramycin on these cells was visible, decreasing the proliferation of tumor cells and inducing apoptosis. In accordance with a study conducted by Broughton et al. (2016), it was shown that about 11 cancer cell lines (AsPC-1, Caco-2, Colo320, HCT116, JJN3, Lovo, MCF-7, MDA-MB-231, MIA PaCa-2, MM. 1S, and U266B1) expressed PE on the surface. Furthermore, cell death from necrosis in these cancer cells and the release of Ca 2+ calcium ions were identified, depending on the time of exposure as well as the concentration of duramycin. Other findings such as morphological changes and influx of iodide have also been reported (Broughton et al., 2016).
Pep27anal2 contains 27 amino acid residues and has a molecular mass of 3.3 kDa. This peptide is an analog of pep27, produced by Streptococcus pneumoniae (Sung et al., 2007). Therefore, pep27 showed MIC 12.5 for Gram-positive and Gram-negative bacteria without a hemolytic effect on human erythrocytes (Sung et al., 2007). Pep27anal2 has a higher number of hydrophobic residues compared to the native peptide pep27. This hydrophobic characteristic of pep27anal2 may be related to the interaction with cell membranes and possibly the anticancer activity that this peptide has demonstrated (Figure 3A) (Lee et al., 2005). It demonstrated activity against leukemia cancer cells (Jurkat, HL-60, AML-2), breast cancer (MCF-7) and gastric cancer (SNU-601). Besides the ability to permeate cancer cells, data indicate that the mechanism responsible for cytotoxicity in neoplastic cells arises from the induction of apoptosis of caspase-free and cytochrome-C. In addition, electron microscopy revealed that pep27anal2 induced the morphological features of apoptosis in Jurkat cells, and showed cytoplasmic condensation, cell shrinkage, loss of plasma membrane microvilli, condensed or fragmented nuclei, and the formation of membrane vesicles (Lee et al., 2005). Considering its potent activity against cancer cells, pep27anal2 is a potential candidate for antineoplastic therapy (Lee et al., 2005).
Based on genomic analyses of Lactobacillus casei ATCC334, some DNA sequences responsible for the expression of the antimicrobial peptides m2163 and m2386 were identified (Tsai et al., 2015). These peptides demonstrated activity on different lactobacillus strains as well as species of Listeria sp. Furthermore, m2163 and m2386 showed effective activity on SW480 cancer cells, acting on the cell membrane and then penetrating the cell cytoplasm to induce apoptosis (Figure 3B) (Tsai et al., 2015). These bacteriocins, m2163 and m2386, were the sources of inspiration for the development of the KL15 antimicrobial peptide through in silico modifications in their sequences. KL15, besides having potent antibacterial activity on pathogenic bacteria such as Enterococcus, Staphylococcus, Bacillus, Escherichia, and Listeria, presented anticancer activity against SW480 and CaCo-2 human adenocarcinoma cells (Chen et al., 2015). Moreover, KL15 (50 μg. mL–1) has been shown to be able to permeate the membranes of SW480 cells, resulting in the formation of porous structures, resulting in necrotic cell death. However, the 150 μg. mL–1 dose of KL15 showed cytotoxicity on human normal mammary epithelial cells H184B5F5/M10 (Chen et al., 2015).
As mentioned above, cancer and chronic infections are the predominant causes of death worldwide. The conventional treatment for these problems generates resistance against multiple drugs. Moreover, conventional treatments are not efficient and effective, inducing serious side effects in patients (Liu et al., 2015; Felício et al., 2017; Leite et al., 2018).
Thus, a new class of molecules needs to be developed and used to provide a more targeted therapy, by exploiting more specific interactions between the drugs and their targets (Gaspar et al., 2013; Felício et al., 2017; Wang et al., 2017; Leite et al., 2018; Shoombuatong et al., 2018). To this end, bacteria have an arsenal of proteins and peptides with both antibacterial and antitumoral activity, which can be explored in the search for these new compounds (Karpinski and Adamczak, 2018).
Various approaches have been discussed here, demonstrating the properties of proteins and peptides derived from bacteria as an alternative strategy for cancer treatment. Among the proteins and peptides that act against bacterial and cancer cells, several stand out. Colicins act on a wide range of bacteria (Duché and Houot, 2019), and have also demonstrated activity against breast carcinoma, fibrosarcoma, leiomyosarcoma, osteosarcoma, and colon carcinoma (Karpinski and Adamczak, 2018). Another promising group is the nisins, which act on Gram-positive bacteria (Jack et al., 1995; Severina et al., 1998), and some nisin variations have demonstrated activity against Gram-negative bacteria (Kuwano et al., 2005), besides potent activity on HNSCC, reducing tumorigenesis (Joo et al., 2012).
Proteins and peptides have been studied for some years, but recently the number of publications and in vivo tests have increased. This has led to the rising number of proteins and peptides approved for use in medical practice. In addition, it is to be expected that in upcoming years these molecules may replace conventional treatments. Thus, it is necessary to improve some properties of these molecules in order to decrease or eliminate cytotoxic effects and increase the specificity of the targeting. Besides that, to expand the use of proteins and peptides it will be important to combine these molecules with conventional drugs. This can reduce costs per treatment, besides decreasing the resistance problem. Therefore, proteins and peptides derived from bacteria with dual activity are an important alternative to current treatments against infections and cancer, reducing side effects and curbing the rise of resistant bacteria.
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
This work was supported by grants from Fundação de Apoio à Pesquisa do Distrito Federal (FAPDF), Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES), Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), and Fundação de Apoio ao Desenvolvimento do Ensino, Ciência e Tecnologia do Estado de Mato Grosso do Sul (FUNDECT), Brazil.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Abdi-Ali, A., Worobec, E. A., Deezagi, A., and Malekzadeh, F. (2004). Cytotoxic effects of pyocin S2 produced by Pseudomonas aeruginosa on the growth of three human cell lines. Can. J. Microbiol. 50, 375–381. doi: 10.1139/w04-019
Ahmadi, S., Ghollasi, M., and Hosseini, H. M. (2017). The apoptotic impact of nisin as a potent bacteriocin on the colon cancer cells. Microb. Pathog. 111, 193–197. doi: 10.1016/j.micpath.2017.08.037
Alexander, J. L., Thompson, Z., Yu, Z., and Cowan, J. A. (2019). Cu-ATCUN derivatives of Sub5 exhibit enhanced antimicrobial activity via multiple modes of action. ACS Chem. Biol. 14, 449–458. doi: 10.1021/acschembio.8b01087
Arias, C. A., and Murray, B. E. (2009). Antibiotic-resistant bugs in the 21st century—a clinical super-challenge. N. Engl. J. Med. 360, 439–443. doi: 10.1056/nejmp0804651
Baindara, P., Chaudhry, V., Mittal, G., Liao, L. M., Matos, C. O., Khatri, N., et al. (2015). Characterization of the antimicrobial peptide penisin, a class ia novel lantibiotic from Paenibacillus sp. Strain A3. Antimicrob. Agents Chemother. 60, 580–591. doi: 10.1128/AAC.01813-15
Baindara, P., Korpole, S., and Grove, V. (2018). Bacteriocins: perspective for the development of novel anticancer drugs. Appl. Microbiol. Biotechnol. 102, 10393–10408. doi: 10.1007/s00253-018-9420-8
Baindara, P., Singh, N., Ranjan, M., Nallabelli, N., Chaudhry, V., Pathania, G. L., et al. (2016). Laterosporulin10: a novel defensin like class IId bacteriocin from Brevibacillus sp. strain SKDU10 with inhibitory activity against microbial pathogens. Microbiology 162, 1286–1299. doi: 10.1099/mic.0.000316
Bandala, C., Perez-Santos, J. L. M., Lara-Padilla, E., Delgado, L., and Anaya-Ruiz, M. (2013). Effect of botulinum toxin A on proliferation and apoptosis in the T47D breast cancer cell line. Asian Pac. J. Cancer Prev. 14, 891–894. doi: 10.7314/apjcp.2013.14.2.891
Beaulieu, L. (2005). Production, Purification et Caracterisation de la Pediocine PA-1 Naturelle et de ses Formes Recombinantes: Contribution a la Mise en Evidence d’une Nouvelle Activite Biologique. Quebec, QC: Universite Laval.
Bédard, F., Hammami, R., Zirah, S., Rebuffat, S., Fliss, I., and Biron, E. (2018). Synthesis, antimicrobial activity and conformational analysis of the class IIa bacteriocin pediocin PA-1 and analogs thereof. Sci. Rep. 8:9029. doi: 10.1038/s41598-018-27225-3
Belmadi, N., Wu, Y., and Touqui, L. (2018). Immuno-modulatory functions of the type-3 secretion system and impacts on the pulmonary host defense: a role for ExoS of Pseudomonas aeruginosa in cystic fibrosis. Toxicon 143, 68–73. doi: 10.1016/j.toxicon.2018.01.004
Bernardes, N., Chakrabarty, A. M., and Fialho, A. M. (2013). Engineering of bacterial strains and their products for cancer therapy. Appl. Microbiol. Biotechnol. 97, 5189–5199. doi: 10.1007/s00253-013-4926-6
Broughton, L. J., Crow, C., Maraveyas, A., and Madden, L. A. (2016). Duramycin-induced calcium release in cancer cells. Anticancer Drugs 27, 173–182. doi: 10.1097/CAD.0000000000000313
Budič, M., Rijavec, M., Petkovšek, Ž, and Žgur-Bertok, D. (2011). Escherichia coli bacteriocins: antimicrobial efficacy and prevalence among isolates from patients with bacteraemia. PLoS One 6:e28769. doi: 10.1371/journal.pone.0028769
Cascales, E., Buchanan, S. K., Duché, D., Kleanthous, C., Lloubes, R., Postle, K., et al. (2007). Colicin biology. Microbiol. Mol. Biol. Rev. 71, 158–229. doi: 10.1128/MMBR.00036-06
Chakrabarty, A. M., Bernardes, N., and Fialho, A. M. (2014). Bacterial proteins and peptides in cancer therapy: today and tomorrow. Bioengineered 5, 234–242. doi: 10.4161/bioe.29266
Chai, T., Wu, V., and Foulds, J. (1982). Colicin A receptor: role of two Escherichia coli outer membrane proteins (OmpF protein and btuB gene product) and lipopolysaccharide. J. Bacteriol. 151, 983–988.
Chen, Y.-C., Tsai, T.-L., Ye, X.-H., and Lin, T.-H. (2015). Anti-proliferative effect on a colon adenocarcinoma cell line exerted by a membrane disrupting antimicrobial peptide KL15. Cancer Biol. Ther. 16, 1172–1183. doi: 10.1080/15384047.2015.1056407
Chumchalova, J., and Šmarda, J. (2003). Human tumor cells are selectively inhibited by colicins. Folia Microbiol. 48, 111–115. doi: 10.1007/BF02931286
Cutter, C. N., and Siragusa, G. R. (1998). Incorporation of nisin into a meat binding system to inhibit bacteria on beef surfaces. Lett. Appl. Microbiol. 27, 19–23. doi: 10.1046/j.1472-765X.1998.00381.x
Daw, M. A., and Falkiner, F. R. (1996). Bacteriocins: nature, function and structure. Micron 27, 467–479. doi: 10.1016/s0968-4328(96)00028-5
de Arauz, L. J., Jozala, A. F., Mazzola, P. G., and Penna, T. C. V. (2009). Nisin biotechnological production and application: a review. Trends Food Sci. Technol. 20, 146–154. doi: 10.1016/j.tifs.2009.01.056
de Lorenzo, V. (1984). Isolation and characterization of microcin E 492 from Klebsiella pneumoniae. Arch. Microbiol. 139, 72–75. doi: 10.1007/BF00692715
de Lorenzo, V., and Pugsley, A. P. (1985). Microcin E492, a low-molecular-weight peptide antibiotic which causes depolarization of the Escherichia coli cytoplasmic membrane. Antimicrob. Agents Chemother. 27, 666–669. doi: 10.1128/aac.27.4.666
Di Masi, D. R., White, J. C., Schnaitman, C. A., and Bradbeer, C. (1973). Transport of vitamin B12 in Escherichia coli : common receptor sites for vitamin B12 and the E colicins on the outer membrane of the cell envelope. J. Bacteriol. 115, 506–513.
Diep, D. B., Håvarstein, L. S., and Nes, I. F. (1996). Characterization of the locus responsible for the bacteriocin production in Lactobacillus plantarum C11. J. Bacteriol. 178, 4472–4483. doi: 10.1128/jb.178.15.4472-4483.1996
Drider, D., Fimland, G., Hechard, Y., McMullen, L. M., and Prevost, H. (2006). The continuing story of class IIa bacteriocins. Microbiol. Mol. Biol. Rev. 70, 564–582. doi: 10.1128/MMBR.00016-05
Duché, D., and Houot, L. (2019). Similarities and differences between colicin and filamentous phage uptake by bacterial cells. EcoSal Plus 8, doi: 10.1128/ecosalplus.ESP-0030-2018
Epand, R. M., and Epand, R. F. (2011). Bacterial membrane lipids in the action of antimicrobial agents. J. Pept. Sci. 17, 298–305. doi: 10.1002/psc.1319
Felício, M. R., Silva, O. N., Goncçalves, S., Santos, N. C., and Franco, O. L. (2017). Peptides with dual antimicrobial and anticancer activities. Front. Chem. 5:5. doi: 10.3389/fchem.2017.00005
Fimland, G., Johnsen, L., Dalhus, B., and Nissen-Meyer, J. (2005). Pediocin-like antimicrobial peptides (class IIa bacteriocins) and their immunity proteins: biosynthesis, structure, and mode of action. J. Pept. Sci. 11, 688–696. doi: 10.1002/psc.699
Fuska, J., Fusková, A., Šmarda, J., and Mach, J. (1979). Effect of colicin E3 on leukemia cells P 388 in vitro. Experientia 35, 406–407. doi: 10.1007/bf01964380
Gabernet, G., Müller, A. T., Hiss, J. A., and Schneider, G. (2016). Membranolytic anticancer peptides. MedChemComm 7, 2232–2245. doi: 10.1039/c6md00376a
Gaspar, D., Veiga, A. S., and Castanho, M. A. (2013). From antimicrobial to anticancer peptides. A review. Front. Microbiol. 4:294. doi: 10.3389/fmicb.2013.00294
Gharsallaoui, A., Oulahal, N., Joly, C., and Degraeve, P. (2016). Nisin as a food preservative: part 1: physicochemical properties, antimicrobial activity, and main uses. Critic. Rev. Food Sci. Nutr. 56, 1262–1274. doi: 10.1080/10408398.2013.763765
Ghequire, M. G., and De Mot, R. (2014). Ribosomally encoded antibacterial proteins and peptides from Pseudomonas. FEMS Microbiol. Rev. 38, 523–568. doi: 10.1111/1574-6976.12079
Ghequire, M. G., and De Mot, R. (2018). Turning over a new leaf: bacteriocins going green. Trends Microbiol. 26, 1–2. doi: 10.1016/j.tim.2017.11.001
Ghosh, C., Sarkar, P., Issa, R., and Haldar, J. (2019). Alternatives to conventional antibiotics in the Era of antimicrobial resistance. Trends Microbiol. 27, 323–338. doi: 10.1016/j.tim.2018.12.010
Gifford, J. L., Hunter, H. N., and Vogel, H. J. (2005). Lactoferricin. Cell. Mol. Life Sci. 62:2588.
Gordon, Y. J., Romanowski, E. G., and McDermott, A. M. (2005). A review of antimicrobial peptides and their therapeutic potential as anti-infective drugs. Curr. Eye Res. 30, 505–515. doi: 10.1080/02713680590968637
Gorgal, T., Charrua, A., Silva, J. F., Avelino, A., Dinis, P., and Cruz, F. (2012). Expression of apoptosis-regulating genes in the rat prostate following botulinum toxin type a injection. BMC Urol. 12:1. doi: 10.1186/1471-2490-12-1
Gratia, A. (1925). Sur un remarquable example d’antagonisme entre deux souches de colibacille. Compt. Rend. Soc. Biol. 93, 1040–1042.
Hammami, R., Zouhir, A., Le Lay, C., Hamida, J. B., and Fliss, I. (2010). BACTIBASE second release: a database and tool platform for bacteriocin characterization. BMC Microbiol. 10:22. doi: 10.1186/1471-2180-10-22
Haney, E. F., Nathoo, S., Vogel, H. J., and Prenner, E. J. (2010). Induction of non-lamellar lipid phases by antimicrobial peptides: a potential link to mode of action. Chem. Phys. Lipids 163, 82–93. doi: 10.1016/j.chemphyslip.2009.09.002
Hasegawa, Y., Shimizu, T., Takahashi, N., and Okada, Y. (2012). The apoptotic volume decrease is an upstream event of MAP kinase activation during staurosporine-induced apoptosis in HeLa cells. Int. J. Mol. Sci. 13, 9363–9379. doi: 10.3390/ijms13079363
Henderson, J. T., Chopko, A. L., and van Wassenaar, P. D. (1992). Purification and primary structure of pediocin PA-1 produced by Pediococcus acidilactici PAC-1.0. Arch. Biochem. Biophys. 295, 5–12. doi: 10.1016/0003-9861(92)90480-K
Hetz, C., Bono, M. R., Barros, L. F., and Lagos, R. (2002). Microcin E492, a channel-forming bacteriocin from Klebsiella pneumoniae, induces apoptosis in some human cell lines. Proc. Natl. Acad. Sci. U.S.A. 99, 2696–2701. doi: 10.1073/pnas.052709699
Hoskin, D. W., and Ramamoorthy, A. (2008). Studies on anticancer activities of antimicrobial peptides. Biochim. Biophys. Acta 1778, 357–375. doi: 10.1016/j.bbamem.2007.11.008
International Agency of Research on Cancer [IARC] (2018). World Cancer Report. Geneva: World-Health-Organisation.
Iwamoto, K., Hayakawa, T., Murate, M., Makino, A., Ito, K., Fujisawa, T., et al. (2007). Curvature-dependent recognition of ethanolamine phospholipids by duramycin and cinnamycin. Biophys. J. 93, 1608–1619. doi: 10.1529/biophysj.106.101584
Jack, R. W., Tagg, J. R., and Ray, B. (1995). Bacteriocins of gram-positive bacteria. Microbiol. Mol. Biol. Rev. 59, 171–200.
Jain, K. K. (2001). Use of bacteria as anticancer agents. Exp. Opin. Biol. Ther. 1, 291–300. doi: 10.1517/14712598.1.2.291
Joo, N. E., Ritchie, K., Kamarajan, P., Miao, D., and Kapila, Y. L. (2012). Nisin, an apoptogenic bacteriocin and food preservative, attenuates HNSCC tumorigenesis via CHAC1. Cancer Med. 1, 295–305. doi: 10.1002/cam4.35
Kamarajan, P., Hayami, T., Matte, B., Liu, Y., Danciu, T., Ramamoorthy, A., et al. (2015). Nisin ZP, a bacteriocin and food preservative, inhibits head and neck cancer tumorigenesis and prolongs survival. PLoS One 10:e0131008. doi: 10.1371/journal.pone.0131008
Karpinski, T. M., and Adamczak, A. (2018). Anticancer activity of bacterial proteins and peptides. Pharmaceutics 10:E54. doi: 10.3390/pharmaceutics10020054
Kaur, S., and Kaur, S. (2015). Bacteriocins as potential anticancer agents. Front. Pharmacol. 6:272. doi: 10.3389/fphar.2015.00272
Kawakami, K., Nakajima, O., Morishita, R., and Nagai, R. (2006). Targeted anticancer immunotoxins and cytotoxic agents with direct killing moieties. Sci. World J. 6, 781–790. doi: 10.1100/tsw.2006.162
Kelly, G. J., Kia, A. F.-A., Hassan, F., O’Grady, S., Morgan, M. P., Creaven, B. S., et al. (2016). Polymeric prodrug combination to exploit the therapeutic potential of antimicrobial peptides against cancer cells. Org. Biomol. Chem. 14, 9278–9286. doi: 10.1039/C6OB01815G
Kominami, K., Nakabayashi, J., Nagai, T., Tsujimura, Y., Chiba, K., Kimura, H., et al. (2012). The molecular mechanism of apoptosis upon caspase-8 activation: quantitative experimental validation of a mathematical model. Biochim. Biophys. Acta Mol. Cell Res. 1823, 1825–1840. doi: 10.1016/j.bbamcr.2012.07.003
Konisky, J. (1982). Colicins and other bacteriocins with established modes of action. Annu. Rev. Microbiol. 36, 125–144. doi: 10.1146/annurev.mi.36.100182.001013
Kumar, B., Balgir, P. P., Kaur, B., and Garg, N. (2011). Cloning and expression of bacteriocins of Pediococcus spp.: a review. Arch. Clin. Microbiol. 2, 1–18. doi: 10.3823/231
Kuwano, K., Tanaka, N., Shimizu, T., Nagatoshi, K., Nou, S., and Sonomoto, K. (2005). Dual antibacterial mechanisms of nisin Z against Gram-positive and Gram-negative bacteria. Int. J. Antimicrob. Agents 26, 396–402. doi: 10.1016/j.ijantimicag.2005.08.010
Lagos, R., Wilkens, M., Vergara, C., Cecchi, X., and Monasterio, O. (1993). Microcin E492 forms ion channels in phospholipid bilayer membranes. FEBS Lett. 321, 145–148. doi: 10.1016/0014-5793(93)80096-d
Lee, D. G., Hahm, K.-S., Park, Y., Kim, H.-Y., Lee, W., Lim, S.-C., et al. (2005). Functional and structural characteristics of anticancer peptide Pep27 analogues. Cancer Cell. Int. 5:21. doi: 10.1186/1475-2867-5-21
Leite, M. L., da Cunha, N. B., and Cost, F. F. (2018). Antimicrobial peptides, nanotechnology, and natural metabolites as novel approaches for cancer treatment. Pharmacol. Ther. 183, 160–176. doi: 10.1016/j.pharmthera.2017.10.010
Leshem, Y., and Pastan, I. (2019). Pseudomonas exotoxin immunotoxins and anti-tumor immunity: from observations at the patient’s bedside to evaluation in preclinical models.Toxins 11:20. doi: 10.3390/toxins11010020
Liberio, M. S., Joanitti, G. A., Fontes, W., and Castro, S. (2013). Anticancer peptides and proteins: a panoramic view. Protein a Peptide Lett. 20, 380–391. doi: 10.2174/092986613805290435
Ling, H., Saeidi, N., Rasouliha, B. H., and Chang, M. W. (2010). A predicted S-type pyocin shows a bactericidal activity against clinical Pseudomonas aeruginosa isolates through membrane damage. FEBS Lett. 584, 3354–3358. doi: 10.1016/j.febslet.2010.06.021
Lins, L., Ducarme, P., Breukink, E., and Brasseur, R. (1999). Computational study of nisin interaction with model membrane. Biochim. Biophys. Acta Biomembr. 1420, 111–120. doi: 10.1016/S0005-2736(99)00090-5
Liu, X., Li, Y., Li, Z., Lan, X., Leung, P. H. M., Li, J., et al. (2015). Mechanism of anticancer effects of antimicrobial peptides. J. Fiber Bioeng. Inform. 8, 25–36. doi: 10.3993/jfbi03201503
Lohner, K. (2009). New strategies for novel antibiotics: peptides targeting bacterial cell. Gen. Physiol. Biophys. 28, 105–116. doi: 10.4149/gpb_2009_02_105
Maher, S., and McClean, S. (2006). Investigation of the cytotoxicity of eukaryotic and prokaryotic antimicrobial peptides in intestinal epithelial cells in vitro. Biochem. Pharmacol. 71, 1289–1298. doi: 10.1016/j.bcp.2006.01.012
Mantovani, H. C., Hu, H., Worobo, R. W., and Russell, J. B. (2002). Bovicin HC5, a bacteriocin from Streptococcus bovis HC5. Microbiology 148, 3347–3352. doi: 10.1099/00221287-148-11-3347
Martarelli, D., Pompei, P., and Mazzoni, G. (2009). Inhibition of adrenocortical carcinoma by diphtheria toxin mutant CRM197. Chemotherapy 55, 425–432. doi: 10.1159/000264689
McCaughey, L. C., Ritchie, N. D., Douce, G. R., Evans, T. J., and Walker, D. (2016). Efficacy of species-specific protein antibiotics in a murine model of acute Pseudomonas aeruginosa lung infection. Sci. Rep. 6:30201. doi: 10.1038/srep30201
Mehta, R. R., Yamada, T., Taylor, B. N., Christov, K., King, M. L., Majumdar, D., et al. (2011). A cell penetrating peptide derived from azurin inhibits angiogenesis and tumor growth by inhibiting phosphorylation of VEGFR-2, FAK and Akt. Angiogenesis 14, 355–369. doi: 10.1007/s10456-011-9220-6
Michel-Briand, Y., and Baysse, C. (2002). The pyocins of Pseudomonas aeruginosa. Biochimie 84, 499–510. doi: 10.1093/oxfordjournals.jbchem.a129929
Moll, G. N., Clark, J., Chan, W. C., Bycroft, B. W., Roberts, G. C., Konings, W. N., et al. (1997). Role of transmembrane pH gradient and membrane binding in nisin pore formation. J. Bacteriol. 179, 135–140. doi: 10.1128/jb.179.1.135-140.1997
Morin, N., Lanneluc, I., Connil, N., Cottenceau, M., Pons, A. M., and Sablé, S. (2011). Mechanism of bactericidal activity of microcin L in Escherichia coli and Salmonella enterica. Antimicrob. Agents Chemother. 55, 997–1007. doi: 10.1128/AAC.01217-10
Nakamura, S., and Racker, E. (1984). Inhibitory effect of duramycin on partial reactions catalyzed by sodium-potassium adenosinetriphosphatase from dog kidney. Biochemistry 23, 385–389. doi: 10.1021/bi00297a031
Nes, I. F., and Holo, H. (2000). Class II antimicrobial peptides from lactic acid bacteria. Pept. Sci. 55, 50–61. doi: 10.1002/1097-0282(2000)55:1<50::aid-bip50>3.0.co;2-3
Nguyen, L. T., Haney, E. F., and Vogel, H. J. (2011). The expanding scope of antimicrobial peptide structures and their modes of action. Trends Biotechnol. 29, 464–472. doi: 10.1016/j.tibtech.2011.05.001
Nissen-Meyer, J., Larsen, A. G., Sletten, K., Daeschel, M., and Nes, I. F. (1993). Purification and characterization of plantaricin A, a Lactobacillus plantarum bacteriocin whose activity depends on the action of two peptides. J. Gen. Microbiol. 139, 1973–1978. doi: 10.1099/00221287-139-9-1973
O’Brien-Simpson, N. M., Hoffmann, R., Chia, C. S. B., and Wade, J. D. (2018). Editorial: antimicrobial and anticancer peptides. Front. Chem. 6:13. doi: 10.3389/fchem.2018.00013
Oliynyk, I., Varelogianni, G., Roomans, G. M., and Johannesson, M. (2010). Effect of duramycin on chloride transport and intracellular calcium concentration in cystic fibrosis and non-cystic fibrosis epithelia. APMIS 118, 982–990. doi: 10.1111/j.1600-0463.2010.02680.x
Paiva, A. D., de Oliveira, M. D., de Paula, S. O., Baracat-Pereira, M. C., Breukink, E., and Mantovani, H. C. (2012). Toxicity of bovicin HC5 against mammalian cell lines and the role of cholesterol in bacteriocin activity. Microbiology 158, 2851–2858. doi: 10.1099/mic.0.062190-0
Papagianni, M., and Anastasiadou, S. (2009). Pediocins: the bacteriocins of Pediococci. Sources, production, properties and applications. Microb. Cell Fact. 8:3. doi: 10.1186/1475-2859-8-3
Phoenix, D. A., Harris, F., Mura, M., and Dennison, S. R. (2015). The increasing role of phosphatidylethanolamine as a lipid receptor in the action of host defence peptides. Prog. Lipid Res. 59, 26–37. doi: 10.1016/j.plipres.2015.02.003
Porto, M. C. W., Kuniyoshi, T. M., Azevedo, P. O. S., Vitolo, M., and Oliveira, R. P. D. S. (2017). Pediococcus spp.: an important genus of lactic acid bacteria and pediocin producers. Biotechnol. Adv. 35, 361–374. doi: 10.1016/j.biotechadv.2017.03.004
Preet, S., Bharati, S., Panjeta, A., Tewari, R., and Rishi, P. (2015). Effect of nisin and doxorubicin on DMBA-induced skin carcinogenesis—a possible adjunct therapy. Tumor Biol. 36, 8301–8308. doi: 10.1007/s13277-015-3571-3
Racker, E., Riegler, C., and Abdel-Ghany, M. (1984). Stimulation of glycolysis by placental polypeptides and inhibition by duramycin. Cancer Res. 44, 1364–1367.
Rebuffat, S. (2013). “Microcins,” in Handbook of Biologically Active Peptides, ed. A. Kastin (Cambridge, MA: Academic Press), 129–137. doi: 10.1016/b978-0-12-385095-9.00020-8
Rolston, K. V. I. (2017). Infections in cancer patients with solid tumors: a review. Infect. Dis. Ther. 6, 69–83. doi: 10.1007/s40121-017-0146-1
Saito, H., Watanabe, T., and Tomioka, H. (1979). Purification, properties, and cytotoxic effect of a bacteriocin from Mycobacterium smegmatis. Antimicrob. Agents Chemother. 15, 504–509. doi: 10.1128/aac.15.4.504
Sand, S. L., Nissen-Meyer, J., Sand, O., and Haug, T. M. (2013). Plantaricin A, a cationic peptide produced by Lactobacillus plantarum, permeabilizes eukaryotic cell membranes by a mechanism dependent on negative surface charge linked to glycosylated membrane proteins. Biochim. Biophys. Acta Biomembr. 1828, 249–259. doi: 10.1016/j.bbamem.2012.11.001
Sand, S. L., Oppegård, C., Ohara, S., Iijima, T., Naderi, S., Blomhoff, H. K., et al. (2010). Plantaricin A, a peptide pheromone produced by Lactobacillus plantarum, permeabilizes the cell membrane of both normal and cancerous lymphocytes and neuronal cells. Peptides 31, 1237–1244. doi: 10.1016/j.peptides.2010.04.010
Scholl, D., and Martin, D. W. (2008). Antibacterial efficacy of R-type pyocins towards Pseudomonas aeruginosa in a murine peritonitis model. Antimicrob. Agents Chemother. 52, 1647–1652. doi: 10.1128/AAC.01479-07
Selegård, R., Musa, A., Nyström, P., Aili, D., Bengtsson, T., and Khalaf, H. (2019). Plantaricins markedly enhance the effects of traditional antibiotics against Staphylococcus epidermidis. Fut. Microbiol. 14, 195–205. doi: 10.2217/fmb-2018-0285
Severina, E., Severin, A., and Tomasz, A. (1998). Antibacterial efficacy of nisin against multidrug-resistant Gram-positive pathogens. J. Antimicrob. Chemother. 41, 341–347. doi: 10.1093/jac/41.3.341
Sheth, T. R., Henderson, R. M., Hladky, S. B., and Cuthbert, A. W. (1992). Ion channel formation by duramycin. Biochim. Biophys. Acta Biomemb. 1107, 179–185. doi: 10.1016/0005-2736(92)90345-M
Shilova, O. N., Shilov, E. S., Lieber, A., and Deyev, S. M. (2018). Disassembling a cancer puzzle: cell junctions and plasma membrane as targets for anticancer therapy. J. Control. Rel. 286, 125–136. doi: 10.1016/j.jconrel.2018.07.030
Shin, J. M., Gwak, J. W., Kamarajan, P., Fenno, J. C., Rickard, A. H., and Kapila, Y. L. (2016). Biomedical applications of Nisin. J. Appl. Microbiol. 120, 1449–1465. doi: 10.1111/jam.13033
Shoombuatong, W., Schaduangrat, N., and Nantasenamat, C. (2018). Unraveling the bioactivity of anticancer peptides as deduced from machine learning. EXCLI J. 17, 734–752. doi: 10.17179/excli2018-1447
Smith, L., and Hillman, J. D. (2008). Therapeutic potential of type A (I) lantibiotics, a group of cationic peptide antibiotics. Curr. Opin. Microbiol. 11, 401–408. doi: 10.1016/j.mib.2008.09.008
Singh, P. K., Solanki, V., Sharma, S., Thakur, K. G., Krishnan, B., and Korpole, S. (2015). The intramolecular disulfide-stapled structure of laterosporulin, a class IId bacteriocin, conceals a human defensin-like structural module. FEBS J. 282, 203–214. doi: 10.1111/febs.13129
Spector, J., Zakharov, S., Lill, Y., Sharma, O., Cramer, W. A., and Ritchie, K. (2010). Mobility of BtuB and OmpF in the Escherichia coli outer membrane: implications for dynamic formation of a translocon complex. Biophys. J. 99, 3880–3886. doi: 10.1016/j.bpj.2010.10.029
Sung, W. S., Park, Y., Choi, C.-H., Hahm, K.-S., and Lee, D. G. (2007). Mode of antibacterial action of a signal peptide, Pep27 from Streptococcus pneumoniae. Biochem. Biophys. Res. Commun. 363, 806–810. doi: 10.1016/j.bbrc.2007.09.041
Thundimadathil, J. (2012). Cancer treatment using peptides: current therapies and future prospects. J. Amino Acids 2012, 1–13. doi: 10.1155/2012/967347
Tong, Z., Dong, L., Zhou, L., Tao, R., and Ni, L. (2010). Nisin inhibits dental caries-associated microorganism in vitro. Peptides 31, 2003–2008. doi: 10.1016/j.peptides.2010.07.016
Tsai, T.-L., Li, A.-C., Chen, Y.-C., Liao, Y.-S., and Lin, T.-H. (2015). Antimicrobial peptide m2163 or m2386 identified from Lactobacillus casei ATCC 334 can trigger apoptosis in the human colorectal cancer cell line SW480. Tumor Biol. 36, 3775–3789. doi: 10.1007/s13277-014-3018-2
Uggerhøj, L. E., Poulsen, T. J., Munk, J. K., Fredborg, M., Sondergaard, T. E., Frimodt-Moller, N., et al. (2015). Rational design of alpha- helical antimicrobial peptides: do’s and don’ts. ChemBioChem 16, 242–253. doi: 10.1002/cbic.201402581
Vasilchenko, A. S., and Valyshev, A. V. (2019). Pore-forming bacteriocins: structural–functional relationships. Arch. Microbiol. 201, 147–154. doi: 10.1007/s00203-018-1610-3
Vedham, V., Divi, R. L., Starks, V. L., and Verma, M. (2014). Multiple infections and cancer: implications in epidemiology. Technol. Cancer Res. Treat. 13, 177–194. doi: 10.7785/tcrt.2012.500366
Villarante, K. I., Elegado, F. B., Iwatani, S., Zendo, T., Sonomoto, K., and de Guzman, E. E. (2011). Purification, characterization and in vitro cytotoxicity of the bacteriocin from Pediococcus acidilactici K2a2-3 against human colon adenocarcinoma (HT29) and human cervical carcinoma (HeLa) cells. World J. Microbiol. Biotechnol. 27, 975–980. doi: 10.1007/s11274-010-0541-1
Wang, L., Dong, C., Li, X., Han, W., and Su, X. (2017). Anticancer potential of bioactive peptides from animal sources. Oncol. Rep. 38, 637–651. doi: 10.3892/or.2017.5778
World-Health-Organisation [WHO] (2018). Cancer: World Cancer Report. Geneva: World-Health-Organisation.
Yamada, T., Mehta, R. R., Lekmine, F., Christov, K., King, M. L., Majumdar, D., et al. (2009). A peptide fragment of azurin induces a p53-mediated cell cycle arrest in human breast cancer cells. Mol. Cancer Ther. 8, 2947–2958. doi: 10.1158/1535-7163.MCT-09-0444
Yang, S. C., Lin, C. H., Sung, C. T., and Fang, J. Y. (2014). Antibacterial activities of bacteriocins: application in foods and pharmaceuticals. Front. Microbiol. 5:241. doi: 10.3389/fmicb.2014.00241
Yu, H., Ding, X., Shang, L., Zeng, X., Liu, H., Li, N., et al. (2018). Protective ability of biogenic antimicrobial peptide microcin J25 against Enterotoxigenic Escherichia Coli -induced intestinal epithelial dysfunction and inflammatory responses IPEC-J2 cells. Front. Cell. Infect. Microbiol. 8:242. doi: 10.3389/fcimb.2018.00242
Zhao, H., Sood, R., Jutila, A., Bose, S., Fimland, G., Nissen-Meyer, J., et al. (2006). Interaction of the antimicrobial peptide pheromone Plantaricin A with model membranes: implications for a novel mechanism of action. Biochim. Biophys. Acta Biomembr. 1758, 1461–1474. doi: 10.1016/j.bbamem.2006.03.037
Keywords: antimicrobial, anticancer, bacteriocin, protein, peptides
Citation: Rodrigues G, Silva GGO, Buccini DF, Duque HM, Dias SC and Franco OL (2019) Bacterial Proteinaceous Compounds With Multiple Activities Toward Cancers and Microbial Infection. Front. Microbiol. 10:1690. doi: 10.3389/fmicb.2019.01690
Received: 14 April 2019; Accepted: 09 July 2019;
Published: 06 August 2019.
Edited by:
Bingyun Li, West Virginia University, United StatesReviewed by:
Ramya Viswanathan, National Institutes of Health (NIH), United StatesCopyright © 2019 Rodrigues, Silva, Buccini, Duque, Dias and Franco. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Octávio Luiz Franco, b2NmcmFuY29AZ21haWwuY29t
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.