 Imen Nouioui1*
Imen Nouioui1* Carlos Cortés-albayay1
Carlos Cortés-albayay1 Lorena Carro2
Lorena Carro2 Jean Franco Castro3
Jean Franco Castro3 Maher Gtari4
Maher Gtari4 Faten Ghodhbane-Gtari4,5
Faten Ghodhbane-Gtari4,5 Hans-Peter Klenk1
Hans-Peter Klenk1 Louis S. Tisa6
Louis S. Tisa6 Vartul Sangal7
Vartul Sangal7 Michael Goodfellow1
Michael Goodfellow1- 1School of Natural and Environmental Sciences, Faculty of Science, Agriculture and Engineering, Newcastle University, Newcastle upon Tyne, United Kingdom
- 2Microbiology and Genetics Department, Universidad de Salamanca, Salamanca, Spain
- 3The Chilean Collection of Microbial Genetic Resources (CChRGM), Instituto de Investigaciones Agropecuarias (INIA) – Quilamapu, Chillán, Chile
- 4Institut National des Sciences Appliquées et de Technologie, Université de Carthage Centre Urbain Nord, Tunis, Tunisia
- 5Laboratoire Microorganismes et Biomolécules Actives, Faculté de Sciences de Tunis, Université de Tunis El Manar, Tunis, Tunisia
- 6Department of Molecular, Cellular and Biomedical Sciences, University of New Hampshire, Durham, NH, United States
- 7Faculty of Health and Life Sciences, Northumbria University, Newcastle upon Tyne, United Kingdom
This study was designed to determine the plant growth promoting (PGP) potential of members of the genus Frankia. To this end, the genomes of 21 representative strains were examined for genes associated directly or indirectly with plant growth. All of the Frankia genomes contained genes that encoded for products associated with the biosynthesis of auxins [indole-3-glycerol phosphate synthases, anthranilate phosphoribosyltransferases (trpD), anthranilate synthases, and aminases (trpA and B)], cytokinins (11 well-conserved genes within the predicted biosynthetic gene cluster), siderophores, and nitrogenases (nif operon except for atypical Frankia) as well as genes that modulate the effects of biotic and abiotic environmental stress (e.g., alkyl hydroperoxide reductases, aquaporin Z, heat shock proteins). In contrast, other genes were associated with strains assigned to one or more of four host-specific clusters. The genes encoding for phosphate solubilization (e.g., low-affinity inorganic phosphate transporters) and lytic enzymes (e.g., cellulases) were found in Frankia cluster 1 genomes, while other genes were found only in cluster 3 genomes (e.g., alkaline phosphatases, extracellular endoglucanases, pectate lyases) or cluster 4 and subcluster 1c genomes (e.g., NAD(P) transhydrogenase genes). Genes encoding for chitinases were found only in the genomes of the type strains of Frankia casuarinae, F. inefficax, F. irregularis, and F. saprophytica. In short, these in silico genome analyses provide an insight into the PGP abilities of Frankia strains of known taxonomic provenance. This is the first study designed to establish the underlying genetic basis of cytokinin production in Frankia strains. Also, the discovery of additional genes in the biosynthetic gene cluster involved in cytokinin production opens up the prospect that Frankia may have novel molecular mechanisms for cytokinin biosynthesis.
Introduction
Actinobacteria classified in the genus Frankia (Brunchorst, 1886) are well known for their ability to induce nitrogen-fixing nodules in over 200 species of dicotyledonous (actinorhizal) plants representing eight angiosperm families (Normand et al., 2014). Mutualistic associations with Frankia strains allow actinorhizal plants to colonize extreme habitats, such as arid, nutritionally poor saline soils (Karthikeyan et al., 2009; Ngom et al., 2016b; Oshone et al., 2017). Until recently, the prospect of selecting Frankia strains for targeted growth promotion of actinorhizal plants in marginal soils was bedeviled by the difficulty of growing these slow-growing bacteria and by the poor state of their systematics. These obstacles have been addressed by the application of better methods for cultivating Frankia strains and by marked improvements in their systematics, mainly due to the application of genome sequence-based taxonomic procedures (Nouioui et al., 2016). The genus currently encompasses 11 validly named species: Frankia alni (Nouioui et al., 2016), the type species, Frankia asymbiotica (Nouioui et al., 2017c), Frankia canadensis (Normand et al., 2018), Frankia casuarinae (Nouioui et al., 2016), Frankia coriariae (Gtari et al., 2015; Nouioui et al., 2017a), Frankia discariae (Nouioui et al., 2017d), Frankia elaeagni (Gtari et al., 2004; Nouioui et al., 2016), Frankia inefficax (Nouioui et al., 2017a), Frankia irregularis (Nouioui et al., 2018b), Frankia saprophytica (Nouioui et al., 2018a), and Frankia torreyi (Nouioui et al., 2019), with the prospect of more to come in the near future (Tisa et al., 2016).
Frankia type strains can be assigned to four clusters with distinct host ranges (Normand et al., 2007; Tisa et al., 2013). Cluster 1 strains nodulate host plants classified in the families Betulaceae, Casuarinaceae (apart from Gymnostoma), and Myricaceae and can be further divided into three subgroups; those assigned to subcluster 1a infect Alnus–Myrica species, subcluster 1b strains, such as strain ARgP5 (Normand et al., 2018), infect Alnus and Myricaceae species while subcluster 1c includes Frankia strains that infect Allocasuarina and Casuarina species (Normand et al., 1996). In turn, cluster 2 strains are associated with plants classified in the families Coriariaceae, Datiscaceae, and Rosaceae and the type genus Ceanothus of the family Rhamnaceae while those in cluster 3 infect host plants belonging to the families Elaeagnaceae, Myricaceae, and Rhamnaceae (except Ceanothus); the genus Gymnostoma and occasionally Alnus species. The fourth cluster encompasses strains isolated from actinorhizal nodules that are unable to either infect or re-establish effective nodulation in the plant from which they were isolated.
Frankia genome sequences generated from representatives of the four clusters are providing valuable insights into the biological properties of members of the genus Frankia (Tisa et al., 2013, 2016), including their potential as a source of novel bioactive compounds (Udwary et al., 2011; Ogasawara et al., 2015) and as biocontrol agents (Gopinathan, 1995). It is particularly interesting that the sizes of Frankia genomes correlate with both host specificity and biogeographic distribution (Normand et al., 2007; Tisa et al., 2013). Further improvements in Frankia systematics and the use of genomic data open up the prospect of selecting specific mutualistic associations between Frankia strains and their hosts for bioremediation (Richards et al., 2002; Diagne et al., 2013, 2015; Rehan et al., 2014a, b, 2015; Baker et al., 2015; Furnholm et al., 2017), notably for saline soils (Sasakawa, 2003; Ngom et al., 2016a; Oshone et al., 2017) and in enhancing the fertility of marginal land (Schwencke and Carú, 2001; Benson and Dawson, 2007; Ngom et al., 2016b).
Plant-growth-promoting bacteria (PGPB) are of interest in sustainable agricultural research and their beneficial effects on plants have been commercially exploited (Gonzalez et al., 2015). In contrast, relatively little is known about the plant growth promoting (PGP) properties of Frankia strains though some have been found to solubilize inorganic phosphate (Sayed et al., 2002) and to synthesize plant hormones (Hirsch et al., 1997; Péret et al., 2007) and siderophores (Boyer et al., 1999; Haansuu et al., 1999; Tisa et al., 2016). However, the improvements in Frankia systematics and the availability of full-genome sequences provide an opportunity to establish the distribution of PGP genes within the genomes of members of the genus and thereby their prospective roles in bioremediation. In the present study, the distribution of PGP genes within the genomes of representative Frankia strains was undertaken with particular reference to those associated with the synthesis of plant hormones, siderophores, and the regulation of phosphate metabolism.
Materials and Methods
Genome Sequences
Table 1 lists the source, host plant specificity, and genome accession numbers of 21 representative Frankia strains, including the type strains of F. alni, F. asymbiotica, F casuarinae, F. coriariae, F discariae, F. elaeagni, F. inefficax, F. irregularis, F. saprophytica, and F. torryei. The following seven type strains were included as outgroups: Acidothermus cellulolyticus 11BT, Blastococcus saxobsidens DD2T, Geodermatophilus obscurus G-20T, Kineococcus radiotolerans ATCC BAA-149T, Modestobacter marinus BC501, Nakamurella multipartita DSM 44233T, and Sporichthya polymorpha DSM 43042T. All of the genome sequences of these strains were obtained from GenBank (accession numbers: CP000481, FO117623, CP001867, CP000750, FO203431, CP001737, and AQZX00000000, respectively).
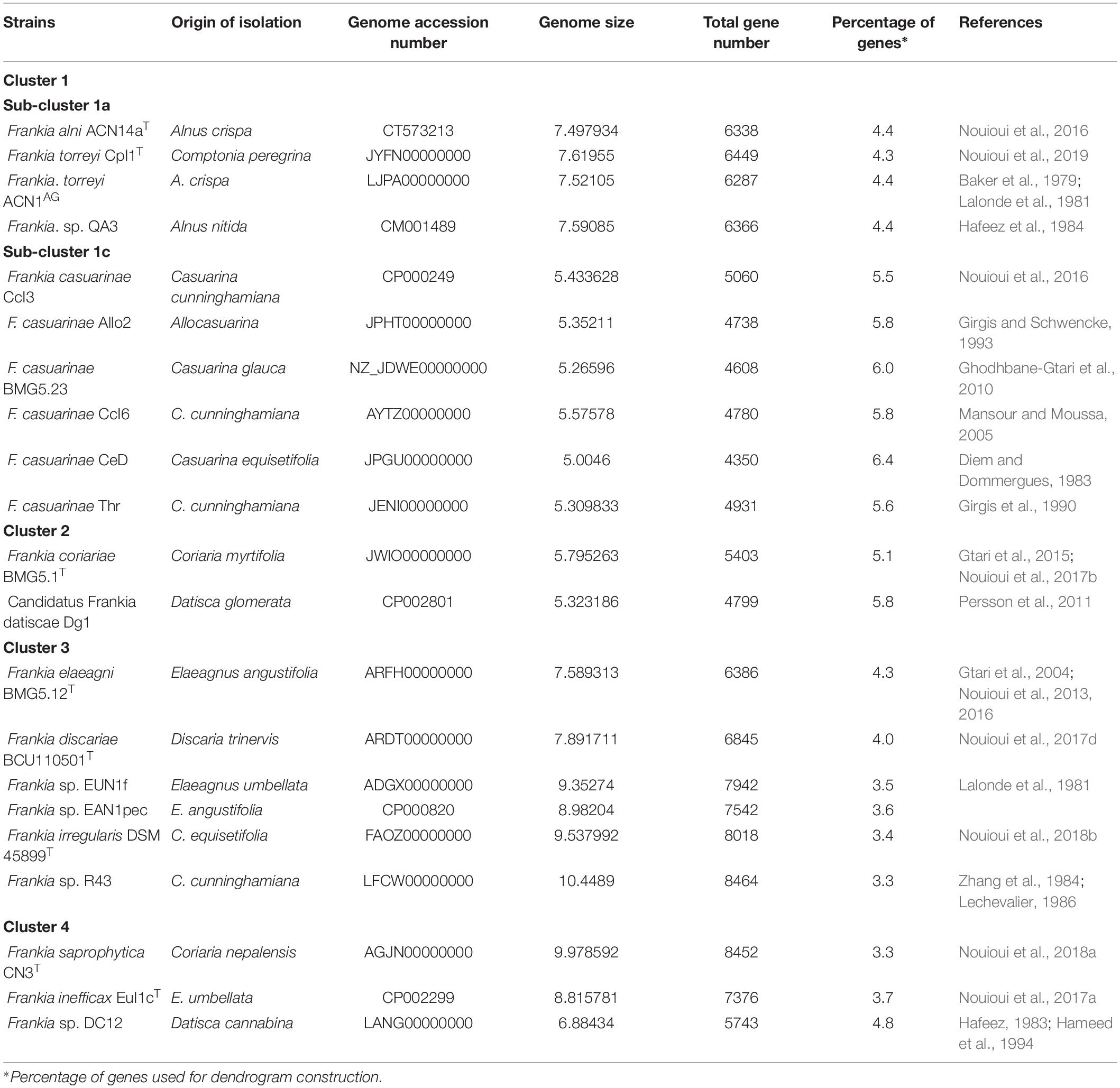
Table 1. Origin of Frankia strains and their genomic features.
In silico Screening of PGP Genes
The genomes of the 21 Frankia strains were annotated using the Rapid Annotation Subsystem Technology server (RAST) (Aziz et al., 2008, 2012). The distribution of PGP genes in the genomes was determined using the SEED server (Overbeek et al., 2014) with a focus on genes encoding for nitrogen fixation, phosphate solubilization, plant hormones, siderophores, lytic enzymes, and those modulating the effect of environmental stress. The gene clusters of the nitrogenase complex (nif) and cytokinins were manually mapped and annotated using ARTEMIS (Berriman and Rutherford, 2003). Each ORF was screened based on an analysis of the GC frame plot of the reading-frames for each of the protein coding sequences (Bibb et al., 1984) and protein domains confirmed after comparison with those available in the Conserved Domains Database (CDD) of NCBI (Marchler-Bauer et al., 2015).
Phylogenomic Analyses
The core genome of the Frankia strains was calculated using the default setting of BPGA 1.3 (Chaudhari et al., 2016) which identified 279 genes. The concatenated protein sequences of the core genes were aligned using MAFFT v7.300b (Katoh and Standley, 2013) and poorly aligned regions and missing data from the concatenated protein sequence alignments were removed using GBLOCKS (Castresana, 2000). The best-fit substitution model, LG+F+I+G4 was identified by ModelFinder (Kalyaanamoorthy et al., 2017) within the IQ-Tree algorithm (Nguyen et al., 2015), which was used to construct a maximum-likelihood dendrogram with 100,000 ultrafast bootstrap iterations and SH-like approximate likelihood ratio tests (Minh et al., 2013) from the resulting alignment.
Results and Discussion
Phylogenomic Diversity
The Frankia strains were assigned to four distinct clusters that were sharply separated from representatives of the seven related genera (Figure 1). Strains assigned to clusters 1 and 3 were found to have high genetic variability. Cluster 1 encompasses ten strains six of which were assigned to subcluster 1c, belonged to F. casuarinae (Gtari et al., 2019) while subcluster 1a was composed of four strains associated with Alnus–Comptonia–Myrica; the latter were assigned to three subgroups which enclosed F. alni ACN14aT, Frankia sp. QA3, and strains of F. torreyi. In turn, Frankia strain ACN1AG has been classified as F. torreyi (Gtari et al., 2019). The topology of subclusters 1a and 1c is in line with that of the MLSA phylogenetic tree of Pozzi et al. (2018) where members of subcluster 1c, which show low genetic diversity, diverge from those of subcluster 1a while those a cluster 2 form a deep rooted evolutionary group. Cluster 3 strains were assigned to four subgroups containing (a) Frankia sp. EAN1pec and F. discariae BCU110501T; (b) F. elaeagni BMG5.12T; (c) Frankia sp. EUN1f; and (d) F. irregularis DSM 45899T and Frankia sp. R43; all of the strains within this cluster form distinct species (Gtari et al., 2019). The overall group structures are highly supported and consistent with those represented by Pozzi et al. (2018). Minor differences in the topology of clusters 1 and 2 compared to the phylogenomic tree provided by Tisa et al. (2016), which was based on 1421 genes, are due to the diversity added by addition of more Frankia genomes and those of the related genera which reduced the core genome to 279 genes.
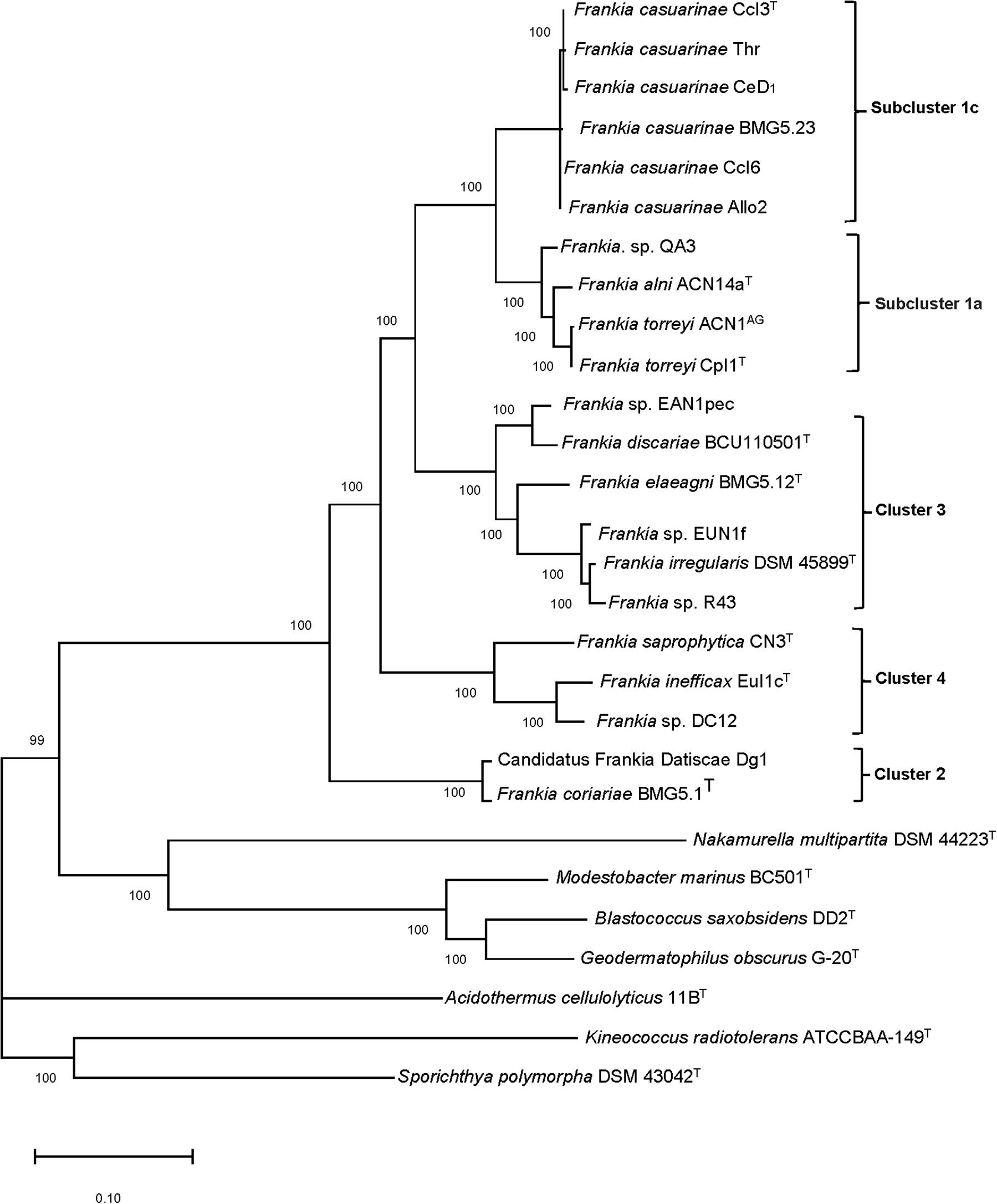
Figure 1. Maximum-likelihood dendrogram, based on 279 core proteins found among these genomes, with 100,000 ultrafast bootstrap iterations showing relationships between Frankia strains assigned to clusters.
The Frankia strains classified in subclusters 1a and 1c showed genome sizes of 5–7.6 Mb and 5.0–5.4 Mb with gene numbers of 6287–6449 and 4350–5060, respectively. In turn, clusters 2 and 3 had genome sizes of 5.0–5.8 Mb and 7.5–10.4 Mb with 4799–5403 and 6845–8464 coding sequences, respectively. Frankia strains associated with cluster 4 had genome sizes within the range of 6.8–9.9 Mb with total gene numbers of 5743–8452 (Table 1). The genome sizes were found to be related to host specificity, found in previous studies (Normand et al., 2007; Tisa et al., 2016).
Direct Mechanisms
Free-living and symbiotic bacteria use direct and indirect mechanisms to promote and protect plant growth. The beneficial effect of Frankia strains in promoting plant growth has been the subject of several studies (Prat, 1989; Steele et al., 1989). Direct mechanisms include nitrogen fixation, phosphate solubilization, enhancement of mineral uptake, and phytohormone production.
Resource acquisition
Nitrogen fixation
Nitrogen is an essential element of most biomolecules that are crucial for life. It is available in the atmosphere as dinitrogen (N2) and can be converted into a plant-usable form through the activities of free-living diazotrophic microorganisms and mutualistic bacteria (Cleveland et al., 1999; Reed et al., 2011). These processes have important ecological and economical roles in sustainable agriculture.
The oxygen-labile enzyme, nitrogenase, converts atmospheric N2 into NH3. Microbes have developed different strategies to protect nitrogenase enzymes from oxygen inactivation. Free-living Frankia strains are able to fix atmospheric nitrogen independent of their host plant in specific cell structures named vesicles (Berry et al., 1993). Frankia vesicles, which contain nitrogenases, are surrounded by a lipid barrier that allows the enzyme to reduce dinitrogen (N2) to ammonium () (Berry et al., 1993). Frankia strains in mutualistic associations with host plants are able to fix up to 300 N2 kg/hectare/year (Shantharam and Mattoo, 1997).
Nitrogenase complexes are composed of two major components: the first, the catalytic part of nitrogenase contains a Fe–Mo cofactor and P clusters (two iron-sulfur clusters) which are encoded by the structural genes nifD and nifK while component II is a nitrogenase reductase that comprises a Fe–S protein encoded by nifH (Dean et al., 1993; Hu et al., 2008). The nif operons consist of three structural genes (nifH, nifD, and nifK) and several accessorial genes such as nifV, nifE, nifN, nifX, nifW, nifZ, nifB, nifU, and nifS (Oh et al., 2003). Accessory genes have different roles in the maturation of inactive products, molecular scaffolds, and electron transport systems within nitrogenase complexes (Dos Santos et al., 2004). Three additional genes have been found within nitrogenase complexes: orA and orB genes encode for ferredoxin oxidoreductase alpha and beta units, respectively, while fdxI encodes for a ferredoxin (Souza et al., 2010). However, little is known about the distribution and organization of genes in the nif operons of Frankia strains (Oh et al., 2012).
In the present study, nif operons were found in the genomes of Frankia strains classified in clusters 1, 2, and 3 (Figure 1 and Supplementary Table S1). All of the accessory nif genes mentioned above, including nifHDK, were present in the genomes of F. alni ACN14aT, F. casuarinae CCI3T, F. coriariae BMG5.1T, and F. elaeagni BMG5.12T (Figure 1). In addition, nifV genes were found in all of the Frankia genomes though in the case of F. elaeagni BMG5.12T it was located 4.4 Mb downstream from the nif operon (Figure 2). In turn, nifV genes are considered to be essential for the activity of nitrogenase complexes because they encode for a homocitrate synthase that catalyzes the condensation of acetyl-CoA and α-ketoglutarate to homocitrate which is used as an organic component of the FeMo cofactor (Oh et al., 2003). However, the homocitrate synthase amino acid sequences of Frankia cluster 1 strains (26%), 2 (29%), and 3 (29%) are not closely related to those involved in the lysine biosynthesis pathways of yeasts and fungi. The alignment of homocitrate synthase amino acid sequences of Frankia strains with those of Saccharomyces cerevisiae showed low identity values between 26 and 29%. It is also interesting that nifENX genes were clustered within the nif operon without any intergenic space (Figure 2). In addition, two orfs (1 and 2), which encode for the protein domains DUF269 and DUF68 with unknown function, were located between the nifX and nifW genes in all of the Frankia genomes (Figure 2). Finally, orA, orB, and fdxI genes were found in the genomes of all of the Frankia strains, as shown in Figure 2. However, the location of these genes was found to vary in the nif operon of F. coriariae BMG5.1T, here the orAB genes were located at the beginning of the operon upstream of nifV while fdxI was located approximately 0.9 Mb downstream of nifS (Figure 2).
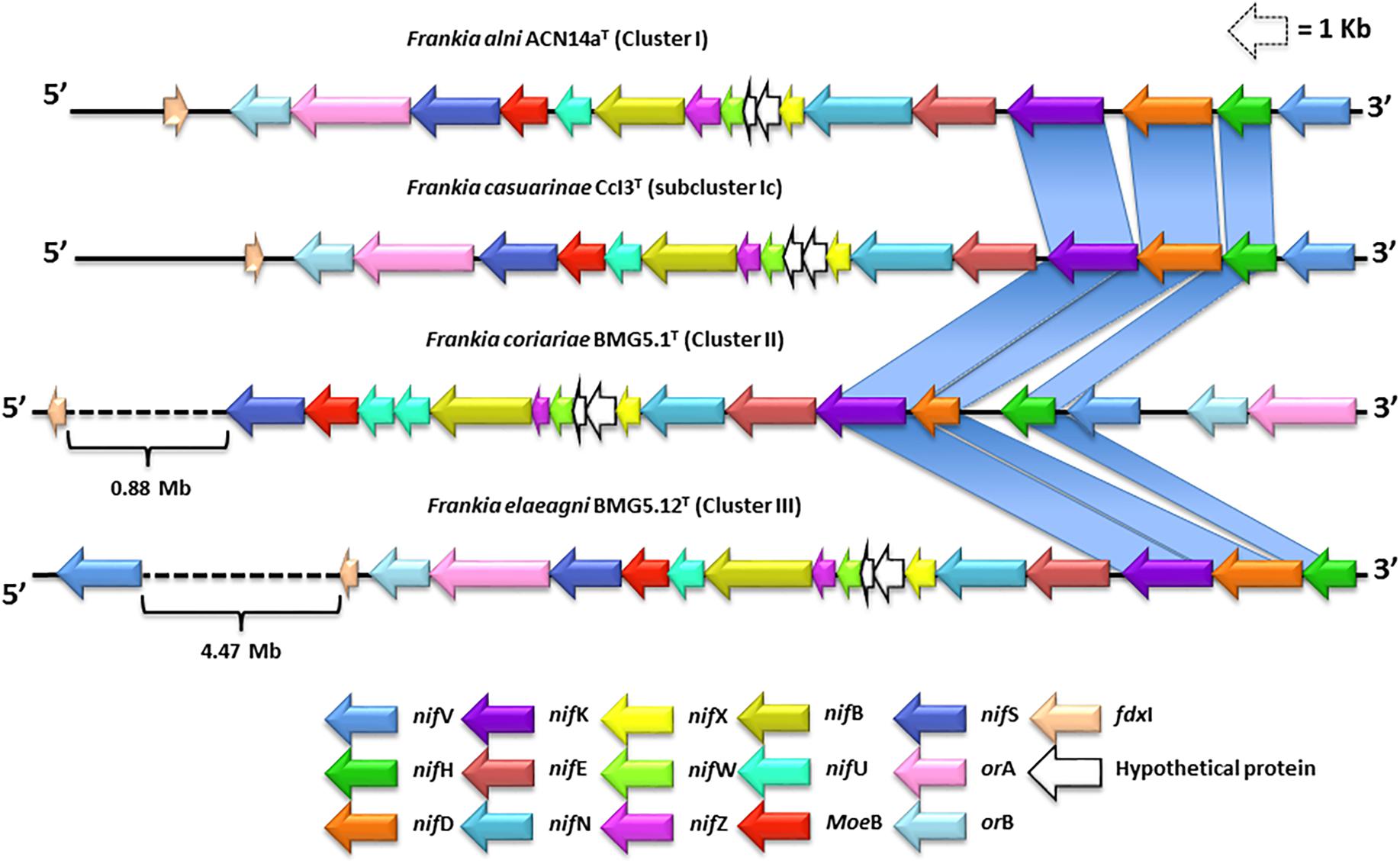
Figure 2. Genome mapping and comparative analysis of nif operons in strains representing Frankia subclusters 1a, 1c, 2, and 3. The nif operons are mainly composed of 18 well-conserved genes which enclose the structural nifHDK genes. The operon also contains the accessory genes, nifV, nifE, nifN, nifX, nifW, nifZ, nifB, nifU, and nifS.
Phosphate solubilization
Phosphorus (P) is an essential element in many biological processes including plant growth and, after nitrogen, is considered to be one of the most important elements limiting crop growth (Tak et al., 2012). Phosphate solubilizing microorganisms (PSM) are able to increase the bioavailability of P for plants by solubilizing inorganic phosphate (Zhu et al., 2011). To this end, microorganisms can release P from organic compounds either enzymatically (Rossolini et al., 1998) or by producing molecules, such as hydroxyl ions, CO2, organic acids, protons, and siderophores that solubilize inorganic phosphate (Rodriguez and Fraga, 1999; Sharma et al., 2013). The most effective PSM belong to the genera Bacillus, Enterobacter, Flavobacterium, Micrococcus, and Rhizobium and to the fungal taxa Aspergillus and Penicillium (Whitelaw, 2000). In contrast, little is known about the ability of representative Frankia strains to solubilize inorganic phosphate.
In the present study, the genomes of most of the Frankia strains were shown to contain an alkaline phosphatase gene (Supplementary Table S2) known to hydrolyze phosphomonoesters and catalyze the transfer of phosphoryl groups to alcohol in the presence of certain phosphate acceptors (Coleman, 1992). It seems likely that this gene is involved in mutualistic relationships between Frankia strains and their host plants by exchanging nutrients in a similar way to that suggested for arbuscular mycorrhizal associations (Aono et al., 2004). In addition, low-affinity inorganic phosphate transporter genes were found in the genomes of the Frankia strains belonging to cluster 1 and F. asymbiotica M16386T (cluster 4). The alignment of amino acid sequences of the low-affinity inorganic phosphate transporter genes of Frankia showed identity values between 90.0 and 99.7% between Frankia strains of cluster 1 and 82.0% with strain M16386T. BLAST results of the alignment of amino acid sequences of low-affinity inorganic phosphate transporter genes of Frankia showed that they are closely related to those found in the genome sequences of other actinobacteria.
Phytohormones
Phytohormones have a crucial role in the growth, development, and the differentiation of plant tissues (Carro and Nouioui, 2017). The best-known ones are indole-3-acetic acid (IAA), cytokinins, ethylene (ET), and gibberellins; the levels of these hormones in plants can be regulated directly by soil microorganisms that synthesize these compounds.
IAA
It has been shown that PGP bacteria may have more than one biosynthetic pathway for the synthesis of hormones such as IAA (Mano and Nemoto, 2012). The latter can be synthesized via indole-3-acetamide (IAM) in phytopathogenic bacteria; the overproduction of IAA leads to the formation of plant tumors (Jameson, 2000). IAA can also be synthesized through the indole-3-pyruvic acid (IPA) pathway, directly by tryptophane (Trp) side chain oxidase (TSO) or through the indole-3-acetonitrile (IAN)/indole-3-acetaldoxime (IAOx) pathway (Glick, 2015). It is likely that L-tryptophan can be converted to IAM by tryptophan-2-monooxygenase that is encoded by the aux1 gene, IAM is then transformed to IAA by IAM hydrolase following the expression of the aux2 gene (Mano and Nemoto, 2012).
Several Frankia strains have been shown to produce auxins (Wheeler et al., 1984; Perrine-Walker et al., 2010) that are involved in Frankia–host plant interactions; decreased concentrations of auxins were found to have a negative effect on root nodule formation in Casuarina glauca (Hammad et al., 2003; Péret et al., 2007). The genomes of the type strains of F. alni, F. casuarinae, and F. elaeagni contain genes that have been seen to be involved in the IPA and phenyl pyruvate IAA biosynthetic pathways (Perrine-Walker et al., 2010) while the type strain of F. discariae has been found to produce IAA and gibberellins in vitro analyses (Solans et al., 2011).
In the present study, the genomes of all of the Frankia strains were shown to have genes that encode for indole-3-glycerol phosphate synthase which is considered to be a branch point of IAA in the tryptophan biosynthetic pathway in plants (Ouyang et al., 2000; Supplementary Table S3). This pathway requires the involvement of the gene products anthranilate phosphoribosyltransferase (trpD), anthranilate synthase, and aminase component (trpA and B) (Lambrecht and Downs, 2013) all of which were detected in the Frankia genomes.
Cytokinins
Cytokinins promote cell division and have growth regulatory functions in plants (Skoog and Armstrong, 1970). In general, they are formed by an adenine nucleotide together with an isoprene, modified isoprene, or aromatic side chain linked to a N6 amino group of adenine (Wong et al., 2015). These chemical structures are precursors to five types of cytokinins: trans-zeatin (tz), kinetin (K), N6-[2-isopentyl]adenine (iP), N6-benzyladenine (BA), and N6-isopentyladenosine (iPR) (Pertry et al., 2009).
The biosynthesis of cytokinins in plants and bacteria starts with the key intermediary dimethylallyl pyrophosphate (DMAPP), this isomerized form of isopentenyl pyrophosphate (IPP) is synthesized in the last step of the mevalonate pathway by isopentenyl-diphosphate delta isomerase (IDI) (Nett et al., 2017). In plants, an isopentenyl group from DMAPP is transferred to the N6 of ATP/ADP (Kakimoto, 2001) while bacteria start off with AMP, which is converted to an intermediary N6-isopentenyladenosine monophosphate (i6AMP) by isopentenyltransferase (ipt). i6AMP is the main enzyme responsible for the synthesis and expression of different variants of cytokinins (Kamínek et al., 1997); it is dephosphorylated to N6-iPR, the first active cytokinin, and is subsequently transformed to the second active cytokine, N6-iP, following an additional deribosylation step. In addition, i6AMP can be hydroxylated to generate the intermediary trans-zeatin riboside-5′-monophosphate (tZMP) which is subsequently dephosphorylated to produce trans-zeatin riboside (tZR) that undergoes deribosylation to yield the active cytokinin tz (Haberer and Kieber, 2002; Kakimoto, 2003; Sakakibara, 2006; Tarkowski et al., 2009; Frébort et al., 2011).
The ipt gene is common in the genomes of plant symbiotic bacteria, as exemplified by Agrobacterium tumefaciens where it is found in the T-region of the “Ti” plasmid which mediates infection in host plants while the homologous gene “tzs” is found near the vir-region on the same plasmid (Mok et al., 2000). Similarly, in Rhodococcus fascians D188T, a homologous gene fasD has been detected in the fas operon located on the pFiD188 plasmid which is involved in cytokinin biosynthesis and infection (Pertry et al., 2009, 2010).
Little is known about the ability of Frankia strains to produce cytokinins though Frankia strain HFPArI3 synthesizes iPR (Stevens and Berry, 1988). However, there is no clear evidence of the genetic mechanisms involved in the biosynthesis of cytokinins within Frankia strains. In the present study, genome mapping of cytokinin gene clusters in the nine strains that represented the Frankia clusters showed that they were composed of 11 highly conserved genes (Figure 3). Two of the genes were associated with the production of ipt and (dimethylallyl) adenosine tRNA methylthiotransferase (damt) (Figure 3) which are involved in the catalysis of the 2-methylthiolated derivative 2-methylthio-isopentenyladenosine (2MeSiPR) (Pertry et al., 2009). An additional gene in this putative cytokinin biosynthetic cluster encodes for a protein domain corresponding to a phosphodiesterase (PDE) that may be involved in the dephosphorylation of i6AMP to iPR. Most of the putative cytokinin biosynthetic gene clusters displayed two genes that encode for recombinase A (recA) and its regulator (RecX) which are involved in DNA exchange and homologous recombination (Roca and Cox, 1990; Kowalczykowski et al., 1994). A third gene located at the end of the gene clusters (Figure 3) encodes for a lysine-motif (LysM), a small protein domain found in bacteria and eukaryotes that is involved in signaling functions for plant–bacteria recognition during bacterial infections (Willmann and Nurnberger, 2012). These preliminary results not only provide a starting point for understanding cytokinin biosynthetic mechanisms in representatives of the genus Frankia but may also provide an insight into the process by which frankiae infect host plants.
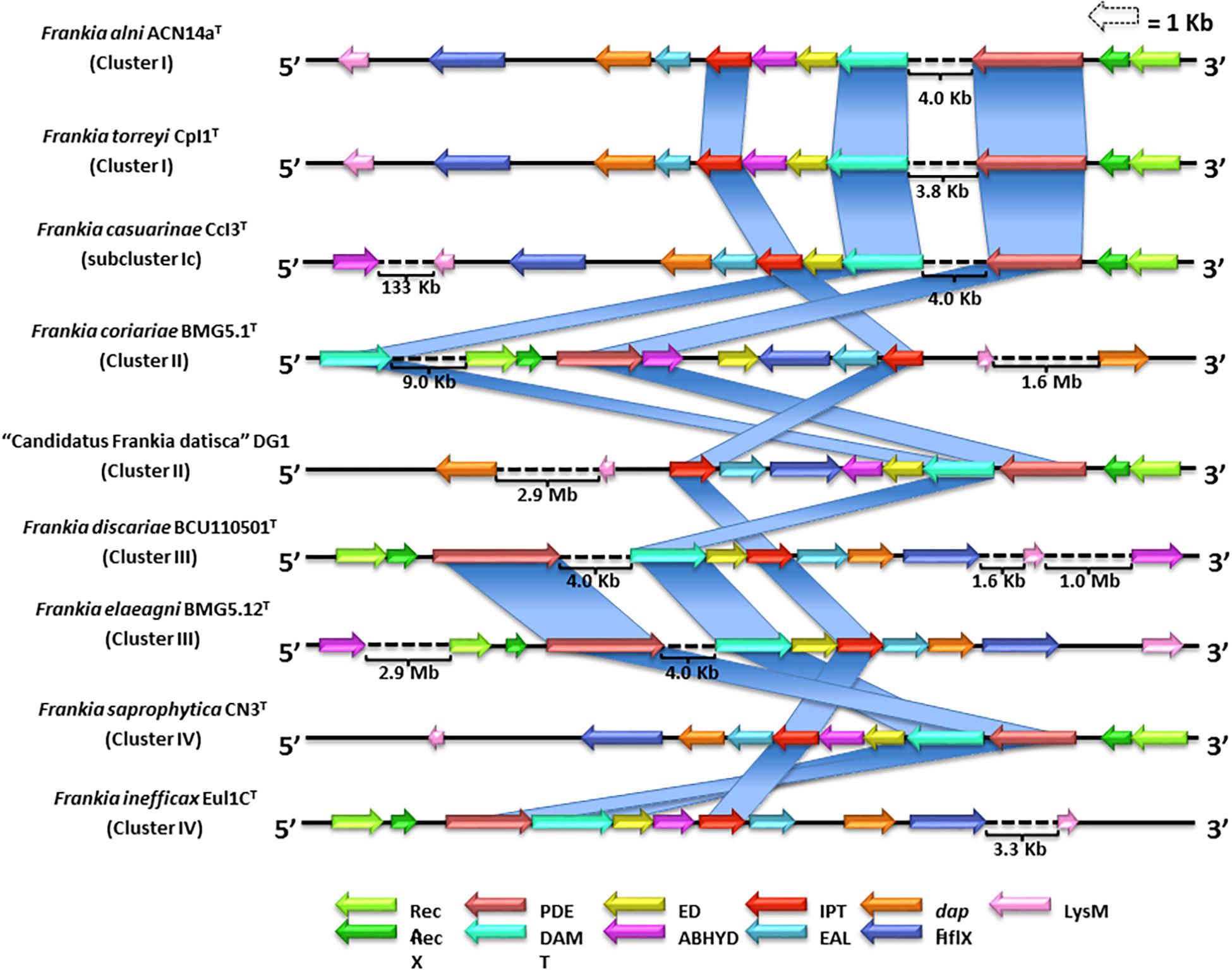
Figure 3. Genome mapping and comparative analysis of the putative cytokinin biosynthetic gene cluster for nine representative strains classified in the genus Frankia. Eleven well-conserved genes encode for: ispentenyltransferase (ipt); (dimethylallyl) adenosine tRNA methylthiotransferase (DAMT); phosphodiesterase (PDE); recombinase A (RecA) and its regulator (RecX); a lysin-motif (LysM); a pimeloyl-ACP methyl ester carboxylesterase (ABHYD); a cyclic diguanylate phosphodiesterase (EAL), a GTPase protein domain (HflX), an extradiol dioxygenase class III protein domain (ED), and a diaminopimelate epimerase (dapF).
Other genes observed in the putative cytokinin biosynthetic cluster encode for protein domains corresponding to genes that express for pimeloyl-ACP methyl ester carboxylesterase (ABHYD), a cyclic diguanylate phosphodiesterase (EAL); a GTPase protein domain (HflX); an extradiol dioxygenase (ED) class III protein domain; a PDE that may be involved in the dephosphorylation of i6AMP to form iPR (Levy et al., 2011); and a diaminopimelate epimerase (DapF) (Figure 3). At present, these are insufficient data to confirm the function of these genes in cytokinin biosynthesis. Clearly, further studies are required to determine the roles of these genes and the molecular mechanisms involved in cytokinin biosynthesis.
Ethylene
The positive effect of this gaseous hormone on plant growth (e.g., seed germination, formation of leaves, flowers, and fruits) is well known (Abeles et al., 1992; Bleecker and Kende, 2000). An increase in the production of ET in plants is a sign of biotic and abiotic stress (e.g., high salinity, increased temperature, insect predation, drought, flooding, presence of toxic compounds) which may lead to enhanced survival of plants or may trigger senescence when the stress persists and ethylene (ET) production is high (Glick, 2012). Methionine is the starting point for ET biosynthesis in plants via S-adenosyl-L-methionine (SAM) which is converted to 1-aminocyclopropane-1-carboxylic acid (ACC) in the presence of ACC synthase (ACS); ACC oxidase (ACCO) has a role in releasing ET and cyanide (converted to β-cyanoalanine to avoid toxicity in plants) (Yang and Hoffman, 1984). In diverse bacteria and fungi (e.g., Escherichia coli, Cryptococcus albidus), ET is synthesized through the oxidation of a transaminated derivative of methionine namely 2-keto-methylthiobutyric acid (KMBA) as well as by a lack of ammonia. In Pseudomonas syringae and Penicillium digitatum, the ET biosynthesis pathway calls for two substrates α-ketoglutarate and arginine which are calalyzed by an ET-forming enzyme (Eckert et al., 2014). In PGP rhizobacteria, ACC deaminase inhibits toxicity caused by high levels of ET in plants, it regulates ET levels by converting ACC produced by the plant to ammonia and α-ketobutyrate (Glick, 1995; GLick et al., 1998). Moreover, it has been shown that ACC deaminase has a significant role in the stimulation of the elongation of plant roots by PGP rhizobacteria. In this context, it is interesting that the genomes of all of the Frankia strains, apart from the F. casuarinae strains, contained genes associated with ACC deaminase (Supplementary Table S3).
Indirect Mechanisms
Plant growth promoting microorganisms also support the growth of plants by modulating environmental biotic and abiotic stress. They are able to either decrease, neutralize, or prevent infection of plants by phytopathogenic bacteria and fungi either by producing lytic enzymes or antibiotics (Singh and Jha, 2015; Gouda et al., 2018). These processes also support the growth of the plants under abiotic stress caused by drought, salinity, and extreme temperature (Akhgar et al., 2014).
Lytic enzymes
One of the defense strategies bacteria use against phytopathogenic fungi involves the production of hydrolytic enzymes such as cellulases, chitinases, glucanases, lipases, lysozymes, and proteases (Neeraja et al., 2010; Maksimov et al., 2011), as well as by other lytic compounds such as lactic acid. The most abundant insoluble polymer in nature, after cellulose, is chitin which can be hydrolyzed by chitinases ChiA, ChiB, and ChiC to N-N′-diacetylchitobiose which is converted to N-acetylglucosamine by N-acetylglucosaminidases. The genomes of F. casuarinae CcI3T, F. inefficax EuI1cT, F. irregularis DSM 45899T, and F. saprophytica CN3T were found to contain genes which encode for chitinases whereas genes associated with cellulase production were only detected in the genomes of the type strains of F. alni, F. torreyi, and Frankia sp. ACN1AG (Supplementary Table S4).
Three types of cellulases, endoglucanases (EC3.2.1.4), exoglucanases (EC3.2.1.91), and β-glucosidases (EC3.2.1.21), belonging to the glycosyl hydrolase family have been described. These enzymes, which are present in microorganisms isolated from diverse ecological niches (Lynd et al., 2002), transform cellulose to glucose. They are also active against phytopathogenic fungi since they hydrolyze β-1,3-glucan, the principal component of fungal cell walls, and indirectly stimulate plant defenses by releasing immune elicitors from the cell walls (Lynd et al., 2002).
The genomes for all of the Frankia strains, apart from those of the type strains of F. casuarinae, F. inefficax and F. irregularis, were shown to contain a gene encoding for an endoglucanase that has an important role in initiating cellulose hydrolysis (Supplementary Table S4; Cohen et al., 2005). In addition, the cluster 3 strains and the type strain of F. saprophytica (cluster 4) were found to have the capacity to produce an extracellular endoglucanase which has been detected in Paenibacillus polymyxa BEb-40 (Gastelum-Arellanez et al., 2014) and used in industry to breakdown lignocellulose (Supplementary Table S4). Furthermore, a gene encoding for the type III effector hrpW hairpin, known to induce hypersensitivity responses in plants (Charkowski et al., 1998) and previously detected in plant-related actinobacteria (Carro et al., 2018), was detected in the genomes of F. elaeagni BMG5.12T and F. saprophytica CN3T (Supplementary Table S4). Gene pl, which encodes for pectate lyase (PL), was detected in the genomes of F. saprophytica CN3T and Frankia strains assigned to cluster 3. This gene has been found in pathogenic bacteria and is known to degrade host tissues, a process in line with its role in the maceration and soft rotting of plant tissues (Marín-Rodríguez et al., 2002). Since the gene hrpW is associated with PA production, it seems likely that genes hrpW and pa are involved in the initiation of Frankia–host plant interactions.
Siderophores
Iron is an essential element for all organisms, including microorganisms. Bacteria and fungi produce siderophores in response to iron limitation (Saha et al., 2016). Consequently, these Fe3+ chelators have an important role in the survival of bacteria, including pathogens, by scavenging iron from iron-binding proteins produced by their hosts (Wandersman and Delepelaire, 2004). The genomes of the F. casuarina strains were shown to harbor a gene that encodes for 2-amino-3,7-dideoxy-D-threo-hept-6-ulosonate synthase (aroA′) which is involved in the shikimate pathway (Supplementary Table S3). Chorismate synthase (CS), chorismate mutase (CM), and shikimate synthase are known to be fundamental in catalyzing the aromatic amino acid (AAA) biosynthetic pathway which is necessary for the production of specialized metabolites essential for plant growth (Helmstaedt et al., 2001; Sasso et al., 2004). The AAA, chorismate, is considered to be an intermediate compound from which catecholate siderophore is synthesized, a reaction that involves a series of enzymes (Walsh et al., 1990). The expression of siderophore genes is regulated by an iron-binding repressor protein, a ferric uptake regulator (Fur) (Escolar et al., 1999), which is common in Gram-negative and AT-rich Gram-positive bacteria; the genome of the GC-rich actinobacterium, Corynebacterium diphtheriae, contains a diphtheria toxin repressor (dtxR) which is essential for siderophore-dependant iron uptake (Qian et al., 2002). Several siderophores have been described in actinobacteria, such as desferrioxamine (G, B, and E), tsukubachelin, and oxachelin, which are characteristic of Streptomyces species (Challis and Hopwood, 2003). In addition, catecholic and hydroxamate moieties have been detected in 44% of soil actinobacteria (Nakouti et al., 2012) while heterobactin has only been reported from Nocardia and Rhodococcus strains (Lee et al., 2012; Wang et al., 2014).
The genomes of all of the Frankia strains showed some variation in the distribution of genes involved in the production of siderophores though siderophore biosynthesis non-ribosomal peptide synthetase modules were found in all of the Frankia genomes; siderophore biosynthesis proteins, related to a monooxygenase and to diaminobutyrate–2-oxoglutarate amino transferase, were present in all of the Frankia genomes except those of the cluster 4 strains (Supplementary Table S5).
Stress genes
Bacteria have developed several ways of coping with environmental stress. In this context, they produce three types of hemoglobin proteins: truncated hemoglobins (trHbo), hemoglobins (Hbos), and flavohemoglobins (flavoHbo), in response to oxygen limitation, oxidative and nitrosative stress. Frankia strains produce two of these hemoglobins: Hbo and flavoHbo. There are two types of trHbo, namely HboO and HboN, which act as scavengers of O2 and NO, respectively (Frey and Kallio, 2003; Supplementary Table S6) while flavoHbo is involved in the nitric dioxygenase reaction by detoxifying NO and protects bacteria from several noxious nitrogen compounds (Frey and Kallio, 2003). The genomes of several Frankia strains express for trHBo- and flavoHbo-associated products that may protect them from nitrosative stress and increase their respiration rates in low-oxygen environments (Beckwith et al., 2002; Tjepkema et al., 2002; Niemann et al., 2005; Niemann and Tisa, 2008). These genes are expressed when host plants are infected followed by the liberation of free radical oxygen and nitric oxide which act as plant defense mechanisms (Niemann and Tisa, 2008).
In this present investigation, the genomes of Frankia strains classified in subcluster 1a and some representatives of subcluster 1c (strains CcI3, CeD, and BMG5.23) and cluster 3 (strains EUN1f and R43) were shown to carry the hmpX gene which encodes for a flavohemoglobin involved in nitrosative stress (Supplementary Table S6). In addition, hboN and hboO genes were detected in the genomes of F. alni ACN14aT, F. torreyi CpI1T (subcluster 1a), F. casuarinae CcI3T (subcluster 1c), F. discariae BCU110501T, F. elaeagni BMG5.12T (cluster 3), and F. inefficax Eul1CT (cluster 4). Interestingly, only the genomes of F. coriariae BMG5.1T, candidatus Frankia datiscae Dg1 (cluster 2), and F. saprophytica CN3T (cluster 4) contained the hboO gene which is involved in hypoxic stress. All of these results are in good agreement with those from previous studies (Beckwith et al., 2002; Niemann et al., 2005; Niemann and Tisa, 2008).
The presence of such putative stress genes in Frankia strains was expected since they are known for their ability to survive in harsh environments, including nutrient poor soils (Karthikeyan et al., 2009; Oshone et al., 2017). Further, the genomes of all of the Frankia strains carried a common set of genes, notably ones associated with the production of alkyl hydroperoxide reductase (ahp) which is involved in hydrogen peroxide stress (Bsat et al., 1996) and in the defense of DNA against oxidative damage (Jacobson et al., 1989); the peroxide stress regulator perR, which is related to the FUR family; redox-sensitive transcriptional regulators (rex and sox) (Wietzke and Bahl, 2012) that have a role in oxidative stress protection; rubrerythrin (rbr), which encodes for a peroxidase and has a role in the protection of nitrogenase from oxygen in cyanobacteria (Zhao et al., 2007); aquaporin Z (aqpZ), which is associated with drought stress and cold (cspA and C) and heat shock (grpE) and chaperon proteins (dnaJ and K) that are involved in heat shock responses (Paek and Walker, 1987; Ellis and Hemmingsen, 1989) and the zinc uptake regulator protein (zur) which helps to protect bacteria against oxidative stress (Smith et al., 2009).
Genes encoding for L-proline glycine betaine binding ABC transporter proteins (proX and V) play a crucial role in resistance to osmotic stress in Gram-negative bacteria, such as Sinorhizobium meliloti (Le Rudulier and Bernard, 1986) were found in all of the Frankia genomes, apart from those of subcluster1c and cluster 2 strains (Supplementary Table S6). This finding is consistent with the observation of Oshone et al. (2017) who noted the absence of sarcosine oxidase (SO) genes in F. casuarinae strains.
All of the Frankia genomes were found to contain a range of genes associated with DNA repair systems, as exemplified by exconuclease ABC (uvr operon) and formamidopyrimidine-DNA glycosylase (Gly1) which are responsible for the oxidation of purines of damaged DNA (Supplementary Table S6). Similarly, all of the genomes harbored genes that encode for enzymes involved in photosynthesis, such as phytoene synthase (crtB) and octaprenyl diphosphate synthase (ispB) (Supplementary Table S6). Genes associated with carotenoid biosynthesis (e.g., β-carotene ketolase) were detected in the genomes of some of the Frankia strains belonging to clusters 1 and 4 (Supplementary Table S6). Carotenoids have a crucial role in preventing photooxidative damage (Howitt and Pogson, 2006) and are considered to be precursors of abscisic acid, a phytohormone involved in the control of water retention and some other stress responses (Koornneef, 1986). Further, the genomes of the F. casuarinae strains and those of the representatives of cluster 4 contained the NAD(P) transhydrogenase gene (Supplementary Table S6), which is involved in the reduction of glutathione, an antioxidant that has an important role in preventing damage to cellular components caused by reactive oxygen species (Pompella et al., 2003).
In addition to the ability of Frankia strains to solubilize and convert insoluble phosphate to bioavailable forms, some of them are able to modulate the lack of phosphate in natural environments. In this context, several genes that encode for inducible phosphate starvation (psi), and which belong to the PHO regulon (Hsieh and Wanner, 2010), are involved in organic phosphate solubilization and uptake by either enhancing the ability of cells to efficiently use limited sources of phosphate or to provide access to other sources of phosphate (Antelmann et al., 2000). The genomes of all of the Frankia strains were found to contain phoA, phoB, phoH, phoR, phoU, phy (phytase), tag, ushA (nucleotidase), and ptsABCS genes (Supplementary Table S2). The phoA and phoB genes encode for alkaline phosphatase while phoD expresses for PDE/alkaline phosphatase D which has a role in teichoic acid turnover in the cell wall in Bacillus subtilis (Eder et al., 1996); the pstS gene belongs to the pstSACB1B2 operon which is involved in phosphate transport (Eymann et al., 1996; Qi et al., 1997).
The alkaline phosphatase genes identified in Frankia strains have amino acid sequence similarities of 53–58% and are similar to those found in some actinobacterial species. However, the alignment and comparison of alkaline phosphatase of Frankia strains to PhoA, PhoC, and PhoD proteins of Streptomyces coelicolor showed identity values between 41.9–47, 41.9–54.8, and 26.5–28.5%, respectively, and 35.0–46.4% with the phoA gene from Streptomyces griseus. The alkaline phosphatase of F. elaeagni BMG5.12T showed an amino acid sequence identify value of 58.3% with the phoC gene. These results show that the alkaline phosphatases of Frankia strains are quite specific and are not closely related to the well-studied ones of the cited Streptomyces species.
Overview, Significance, and Future Studies
Frankia strains are well known for their ability to form nitrogen-fixing nodules in actinorhizal plants and to promote plant growth. Genome mining of representative Frankia strains representing the four host infection groups not only show that the genetic machinery of their nitrogenase complexes are conserved but also highlighted the presence of 11 conserved genes (ipt, damt, recA, recX, lysM, eal, hflX, ed, dapF, pde, and abhyd) in the putative cytokinin biosynthetic gene cluster; the presence of the LysM domain and recombinase genes indicates that the cytokinin cluster may also be involved in the ability of Frankia strains to infect their hosts plant. In addition, the genomes of all of the Frankia strains were shown to be equipped with genes associated with the synthesis and production of phytohormones and contained genes functionally linked to inorganic phosphate solubilization and siderophore production. Moreover, the genomes of all the representative strains carried a set of universal genes the products of which are involved in modulating the effects of abiotic and biotic environmental stress. Consequently, it can be concluded that Frankia strains should be seen as potential substitutes for chemical fertilizers and thereby may prove to have an important role in the improving ecosystem quality. However, further work is required to understand the PGP mechanisms of frankiae before they can be developed for use in sustainable agriculture.
Data Availability
The datasets analyzed for this study can be found in the National Center for Biotechnology Information: https://www.ncbi.nlm.nih.gov/.
Author Contributions
IN conceived the project and performed the genome mining analyses while IN and MiG developed the concepts. VS carried out the phylogenomic analyses and interpreted the results together with IN, MiG, LT, and H-PK. CC-A, LC, JFC, H-PK, FG-G, MaG, LT, and VS played roles in analyzing the data and in interpreting the results. IN and MiG wrote the manuscript. All the authors approved the final version.
Funding
This project was supported by the School of Natural and Environmental Sciences (Newcastle University, United Kingdom). IN is grateful to Newcastle University for a postdoctoral fellowship. LC thanks Newcastle and Salamanca Universities for postdoctoral fellowships. JFC was supported by the Instituto de Investigaciones Agropecuarias project no. 502612–70. LT was supported by the USDA National Institute of Food and Agriculture (Hatch 022821), the Plant Health and Production and Plant Products Program (grant no. 2015-67014-22849/project accession no. 1005242) from the USDA National Institute of Food and Agriculture, and the College of Life Science and Agriculture at the University of New Hampshire-Durham.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2019.01457/full#supplementary-material
References
Abeles, F. B., Morgan, P. W., and Saltveit, M. E. J. (1992). Ethylene in Plant Biology. New York, NY: Academic Press.
Akhgar, R., Arzanlou, M., Bakker, P. A. H. M., and Hamidpour, M. (2014). Characterization of 1-aminocyclopropane-1-carboxylate (ACC) deaminase-containing Pseudomonas sp. in the rhizosphere of salt-stressed canola. Pedosphere 24, 461–468. doi: 10.1016/s1002-0160(14)60032-1
Antelmann, H., Scharf, C., and Hecker, M. (2000). Phosphate starvation-inducible proteins of Bacillus subtilis: proteomics and transcriptional analysis. J. Bacteriol. 182, 4478–4490. doi: 10.1128/jb.182.16.4478-4490.2000
Aono, T., Maldonado-Mendoza, I. E., Dewbre, G. R., Harrison, M. J., and Saito, M. (2004). Expression of alkaline phosphatase genes in arbuscular mycorrhizas. New Phytol. 162, 525–534. doi: 10.1016/j.fgb.2013.02.009
Aziz, R. K., Bartels, D., Best, A. A., Dejongh, M., Disz, T., Edwards, R. A., et al. (2008). The RAST server: rapid annotations using subsystems technology. BMC Genomics 9:75. doi: 10.1186/1471-2164-9-75
Aziz, R. K., Devoid, S., Disz, T., Edwards, R. A., Henry, C. S., Olsen, G. J., et al. (2012). SEED servers: high-performance access to the SEED genomes, annotations, and metabolic models. PLoS One 7:e48053. doi: 10.1371/journal.pone.0048053
Baker, D., Torrey, J. G., and Kidd, G. H. (1979). Isolation by sucrose-density fractionation and cultivation in vitro of actinomycetes from nitrogen-fixing root nodules. Nature 281, 76–78. doi: 10.1038/281076a0
Baker, E., Tang, Y., Chu, F., and Tisa, L. S. (2015). Molecular responses of Frankia sp. strain QA3 to naphthalene. Can. J. Microbiol. 61, 281–292. doi: 10.1139/cjm-2014-0786
Beckwith, J., Tjepkema, J. D., Cashon, R. E., Schwintzer, C. R., and Tisa, L. S. (2002). Hemoglobin in five genetically diverse Frankia strains. Can. J. Microbiol. 48, 1048–1055. doi: 10.1139/w02-106
Benson, D., and Dawson, F. (2007). Recent advances in the biogeography and genecology of symbiotic Frankia and its host plants. Physiol. Plant 130, 318–330. doi: 10.1111/j.1399-3054.2007.00934.x
Berriman, M., and Rutherford, K. (2003). Viewing and annotating sequence data with Artemis. Brief. Bioinform. 4, 124–132. doi: 10.1093/bib/4.2.124
Berry, A. M., Harriott, O. T., Moreau, R. A., Osman, S. F., Benson, D. R., and Jones, A. D. (1993). Hopanoid lipids compose the Frankia vesicle envelope, presumptive barrier of oxygen diffusion to nitrogenase. Microbiology 90, 6091–6094. doi: 10.1073/pnas.90.13.6091
Bibb, M. J., Findlay, P. R., and Johnson, M. W. (1984). The relationship between base composition and codon usage in bacterial genes and its use for the simple and reliable identification of protein-coding sequences. Gene 30, 157–166. doi: 10.1016/0378-1119(84)90116-1
Bleecker, A. B., and Kende, H. (2000). Ethylene: a gaseous signal molecule in plants. Annu. Rev. Cell. Dev. Biol. 16, 1–18. doi: 10.1146/annurev.cellbio.16.1.1
Boyer, G. L., Kane, S. A., Alexander, J. A., and Aronson, D. B. (1999). Siderophore formation in iron-limited cultures of Frankia sp. strain 52065 and Frankia sp. strain CeSI5. Can. J. Bot. 77, 1316–1320. doi: 10.1139/cjb-77-9-1316
Brunchorst, J. (1886). Über einige Wurzelanschwellungen, besonders diejenigen von Alnus und den Elaeagnaceen. Unters. Bot. Inst. Tübingen. 2, 151–177.
Bsat, N., Chen, L., and Helmann, J. D. (1996). Mutation of the Bacillus subtilis alkyl hydroperoxide reductase (ahpCF) operon reveals compensatory interactions among hydrogen peroxide stress genes. J. Bacteriol. 178, 6579–6586. doi: 10.1128/jb.178.22.6579-6586.1996
Carro, L., and Nouioui, I. (2017). Taxonomy and systematics of plant probiotic bacteria in the genomic era. AIMS Microbiol. 3, 383–412. doi: 10.3934/microbiol.2017.3.383
Carro, L., Nouioui, I., Sangal, V., Meier-Kolthoff, J. P., Trujillo, M. E., and Montero-Calasanz, M. D. C. (2018). Genome-based classification of micromonosporae with a focus on their biotechnological and ecological potential. Sci. Rep. 8:525. doi: 10.1038/s41598-017-17392-0
Castresana, J. (2000). Selection of conserved blocks from multiple alignments for their use in phylogenetic analysis. Mol. Biol. Evol. 17, 540–552. doi: 10.1093/oxfordjournals.molbev.a026334
Challis, G. L., and Hopwood, D. L. (2003). Synergy and contingency as driving forces for the evolution of multiple secondary metabolite production by Streptomyces species. PNAS 100, 14555–14561. doi: 10.1073/pnas.1934677100
Charkowski, A. O., Alfano, J. R., Preston, G., Yuan, J., He, S. Y., and Collmer, A. (1998). The Pseudomonas syringae pv. tomato HrpW protein has domains similar to harpins and pectate lyases and can elicit the plant hypersensitive response and bind to pectate. J. Bacteriol. 180, 5211–5217.
Chaudhari, N. M., Gupta, V. K., and Dutta, C. (2016). BPGA- an ultra-fast pan-genome analysis pipeline. Sci.Rep. 6:24373. doi: 10.1038/srep24373
Cleveland, C. C., Townsend, A. R., Schimel, D. S., Fisher, H., Howarth, R. W., Hedin, L. O., et al. (1999). Global patterns of terrestrial biological nitrogen (N2 fixation in natural ecosystems. Glob. Biogeochem. Cycles 13, 623–645. doi: 10.1111/plb.12082
Cohen, R., Suzuki, M. R., and Hammel, K. E. (2005). Processive endoglucanase active in crystalline cellulose hydrolysis by the brown rot basidiomycete Gloeophyllum trabeum. Appl. Environ. Microbiol. 71, 2412–2417. doi: 10.1128/aem.71.5.2412-2417.2005
Coleman, J. E. (1992). Structure and mechanism of alkaline phosphatase. Annu. Rev. Biophys. Biomol. Struct. 21, 441–483. doi: 10.1146/annurev.biophys.21.1.441
Dean, D. R., Bolin, J. T., and Zheng, L. (1993). Nitrogenase metalloclusters: structures, organization, and synthesis. J. Bacteriol. 175, 6737–6744. doi: 10.1128/jb.175.21.6737-6744.1993
Diagne, N., Arumugam, K., Ngom, M., Nambiar-Veetil, M., Franche, C., Narayanan, K. K., et al. (2013). Use of Frankia and actinorhizal plants for degraded lands reclamation. Biomed. Res. Int. 2013:948258. doi: 10.1155/2013/948258
Diagne, N., Ngom, M., Djighaly, P. I., Ngom, D., Ndour, B., Cissokho, M., et al. (2015). “Remediation of heavy-metal-contaminated soils and enhancement of their fertility with actinorhizal plants,” in Heavy Metal Contamination of Soils, Soil Biology, eds I. Sherameti and A. VarmaA (Switzerland: Springer International Publishing), 355–366. doi: 10.1007/978-3-319-14526-6_19
Diem, H. G., and Dommergues, Y. (1983). The isolation of Frankia from nodules of Casuarina. Can. J. Bot. 61, 2822–2825. doi: 10.1139/b83-313
Dos Santos, P. C., Dean, D. R., Hu, Y., and Ribbe, M. W. (2004). Formation and insertion of the nitrogenase iron-molybdenum cofactor. Chem. Rev. Argent. Microbiol. 104, 1159–1173.
Eckert, C., Xu, W., Xiong, W., Lynch, S., Ungerer, J., Tao, L., et al. (2014). Ethylene-forming enzyme and bioethylene production. Biotechnol. Biofuels 7:33. doi: 10.1186/1754-6834-7-33
Eder, S., Shi, L., Jensen, K., Yamane, K., and Hulett, F. M. (1996). A Bacillus subtilis secreted phosphodiesterase/alkaline phosphatase is the product of a Pho regulon gene, phoD. Microbiology 142(Pt 8), 2041–2047. doi: 10.1099/13500872-142-8-2041
Ellis, R. J., and Hemmingsen, S. M. (1989). Molecular chaperones: proteins essential for the biogenesis of some macromolecular structures. Trends Biochem. Sci. 14, 339–342. doi: 10.1016/0968-0004(89)90168-0
Escolar, L., Pe’Rez-Marti’, N. J., and De Lorenzo, V. (1999). Opening the iron box: transcriptional metalloregulation by the fur protein. J. Bacteriol. 181, 6223–6229.
Eymann, C., Mach, H., Harwood, C. R., and Hecker, M. (1996). Phosphate-starvation-inducible proteins in Bacillus subtilis: a two-dimensional gel electrophoresis study. Microbiology 142(Pt 11), 3163–3170. doi: 10.1099/13500872-142-11-3163
Frébort, I., Kowalska, M., Hluska, T., Frébortová, J., and Galuszka, P. (2011). Evolution of cytokinin biosynthesis and degradation. J. Exp. Bot. 62, 2431–2452. doi: 10.1093/jxb/err004
Frey, A. D., and Kallio, P. T. (2003). Bacterial hemoglobins and flavohemoglobins: versatile proteins and their impact on microbiology and biotechnology. FEMS Microbiol. Rev. 27, 525–545. doi: 10.1016/s0168-6445(03)00056-1
Furnholm, T., Rehan, M., Wishart, J., and Tisa, L. S. (2017). Pb(2 Tolerance by Frankia sp. strain EAN1pec involves a surface-binding. Microbiology 163, 472–487. doi: 10.1099/mic.0.000439
Gastelum-Arellanez, A., Paredes-Lopez, O., and Olalde-Portugal, V. (2014). Extracellular endoglucanase activity from Paenibacillus polymyxa BEb-40: production, optimization and enzymatic characterization. World J. Microbiol. Biotechnol. 30, 2953–2965. doi: 10.1007/s11274-014-1723-z
Ghodhbane-Gtari, F., Nouioui, I., Chair, M., Boudabous, A., and Gtari, M. (2010). 16S-23S rRNA intergenic spacer region variability in the genus Frankia. Microb. Ecol. 60, 487–495. doi: 10.1007/s00248-010-9641-6
Girgis, M. G. Z., Ishac, Y. Z., El-Haddad, M., Saleh, E. A., Diem, H. G., and Dommergues, Y. R. (1990). “First report on isolation and culture of effective Casuarina-compatible strains of Frankia from Egypt,” in 2nd International Casuarina Workshop, eds M. H. El-Lakany, J. W. Turnbull, and J. L. Brewbaker (Egypt: American University), 156–164.
Girgis, M. G. Z., and Schwencke, J. (1993). Differentiation of Frankia strains by their electrophoretic patterns of intracellular esterases and aminopeptidases. J. Gen. Microbiol. 139, 2225–2232. doi: 10.1099/00221287-139-9-2225
Glick, B. R. (1995). The enhancement of plant growth by free-living bacteria. Can. J. Microbiol. 41, 109–117. doi: 10.1139/m95-015
Glick, B. R. (2012). Plant growth-promoting bacteria: mechanisms and applications. Scientifica 2012:963401. doi: 10.6064/2012/963401
Glick, B. R. (2015). Beneficial Plant-Bacterial Interactions. Canada: Springer International Publishing.
GLick, B. R., Penrose, D. M., and Li, J. (1998). A model for the lowering of plant ethylene concentrations by plant growth-promoting bacteria. J. Theor. Biol. 190, 63–68. doi: 10.1006/jtbi.1997.0532
Gonzalez, A. J., Larraburu, E. E., and Llorente, B. E. (2015). Azospirillum brasilense increased salt tolerance of Jojoba during in vitro rooting. Ind. Crops Products 76, 41–48. doi: 10.1016/j.indcrop.2015.06.017
Gopinathan, K. (1995). Biological control of Rhizoctonia sp. root rot of Casuarina equisetifolia seedlings by Frankia spp. strains. Biol. Fertil. Soils 20, 221–225. doi: 10.1007/bf00336081
Gouda, S., Kerry, R. G., Das, G., Paramithiotis, S., Shin, H. S., and Patra, J. K. (2018). Revitalization of plant growth promoting rhizobacteria for sustainable development in agriculture. Microbiol. Res. 206, 131–140. doi: 10.1016/j.micres.2017.08.016
Gtari, M., Brusetti, L., Skander, G., Mora, D., Boudabous, A., Daffonchio, D., et al. (2004). Isolation of Elaeagnus-compatible Frankia from soils collected in Tunisia. FEMS Microbiol. Lett. 234, 349–355. doi: 10.1016/j.femsle.2004.04.001
Gtari, M., Ghodhbane-Gtari, F., Nouioui, I., Ktari, A., Hezbri, K., Mimouni, W., et al. (2015). Cultivating the uncultured: growing the recalcitrant cluster-2 Frankia strains. Sci. Rep. 5:13112. doi: 10.1038/srep13112
Gtari, M., Nouioui, I., Sarkar, I., Ghodhbane-Gtari, F., Tisa, L. S., Sen, A., et al. (2019). An update on the taxonomy of the genus Frankia, 174AL. Antonie Van Leeuwenhoek 112, 5–21. doi: 10.1007/s10482-018-1165-y
Haansuu, P., Vuorela, P., and Haahtela, K. (1999). Detection of antimicrobial and 45Ca2(-transport blocking activity in Frankia culture broth extracts. Pharm. Pharmacol. Lett. 9, 1–4.
Haberer, G., and Kieber, J. J. (2002). Cytokinins. New insights into a classic phytohormone. Plant. Physiol. 128, 354–362. doi: 10.1104/pp.128.2.354
Hafeez, F. (1983). Nitrogen Fixation and Nodulation in Datisca cannabina L. and Alnus nitida Endl. Ph.D. thesis, Quaid-e-Azam University, Islamabad.
Hafeez, F., Akkermans, A. D. L., and Chaudhary, A. H. (1984). Morphology, physiology, and infectivity of two Frankia isolates, An1 and An2 from root nodules of Alnus nitida. Plant Soil 78, 45–59. doi: 10.1007/978-94-009-6158-6_6
Hameed, S., Hafeez, F. Y., Mirza, M. S., Malik, K. A., and Akkermans, A. D. L. (1994). Confirmation of an isolate from Datisca cannabina as atypical Frankia strain using PCR amplified 16 rRNA sequence analysis. Pak. J. Bot. 26, 247–251.
Hammad, Y., Nalin, R., Marechal, J., Fiasson, K., Pepin, R., Berry, A. M., et al. (2003). A possible role for phenyl acetic acid (PAA) on Alnus glutinosa nodulation by Frankia. Plant Soil 254, 193–205. doi: 10.1007/978-94-017-1601-7_21
Helmstaedt, K., Krappmann, S., and Braus, G. H. (2001). Allosteric regulation of catalytic activity: Escherichia coli aspartate transcarbamoylase versus yeast chorismate mutase. Microbiol. Mol. Biol. Rev. 65, 404–421. doi: 10.1128/mmbr.65.3.404-421.2001
Hirsch, A., Fang, Y., Asad, S., and Kapulnik, Y. (1997). The role of phytohormones in plant-microbe symbioses. Plant Soil 194, 171–184. doi: 10.1007/978-94-011-5744-5_17
Howitt, C. A., and Pogson, B. J. (2006). Carotenoid accumulation and function in seeds and non-green tissues. Plant. Cell Environ. 29, 435–445. doi: 10.1111/j.1365-3040.2005.01492.x
Hsieh, Y. J., and Wanner, B. L. (2010). Global regulation by the seven-component Pi signaling system. Curr. Opin. Microbiol. 13, 198–203. doi: 10.1016/j.mib.2010.01.014
Hu, Y., Fay, A. W., Lee, C. C., Yoshizawa, J., and Ribbe, M. W. (2008). Assembly of nitrogenase MoFe protein. Biochemistry 47, 3973–3981. doi: 10.1021/bi7025003
Jacobson, F. S., Morgan, R. W., Christman, M. F., and Ames, B. N. (1989). An alkyl hydroperoxide reductase from Salmonella typhimurium involved in the defense of DNA against oxidative damage. Purification and properties. J. Biol. Chem. 264, 1488–1496.
Jameson, P. E. (2000). Cytokinins and auxins in plant-pathogen interactions-an overview. Plant Growth. Reg. 32, 369–380.
Kakimoto, T. (2001). Identification of plant cytokinin biosynthetic enzymes as dimethylallyl diphosphate:ADP/ADP isopentenyltransferases. Plant Cell Physiol. 42, 677–685. doi: 10.1093/pcp/pce112
Kakimoto, T. (2003). Biosynthesis of cytokinins. J. Plant. Res. 116, 233–239. doi: 10.1007/s10265-003-0095-5
Kalyaanamoorthy, S., Minh, B. Q., Wong, T. K. F., von Haeseler, A., and Jermiin, L. S. (2017). ModelFinder: fast model selection for accurate phylogenetic estimates. Nat. Methods 14, 587–589. doi: 10.1038/nmeth.4285
Kamínek, M., Motyka, V., and Vaòková, R. (1997). Regulation of cytokinins content in plant cells. Physiol. Plant 101, 689–700. doi: 10.1034/j.1399-3054.1997.1010404.x
Karthikeyan, A., Deepara, J. B., and Nepolean, P. (2009). Reforestation in bauxite mine spoils with Casuarina equisetifolia frost and beneficial microbes. For. Trees Livelihoods 19, 153–165. doi: 10.1080/14728028.2009.9752661
Katoh, K., and Standley, D. M. (2013). MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol. Biol. Evol. 30, 772–780. doi: 10.1093/molbev/mst010
Koornneef, M. (1986). “Genetic aspects of abscisic acid,” in Plant Gene Research, eds A. D. Blonstein and P. J. King (New York, NY: Springer), 35–54. doi: 10.1007/978-3-7091-6989-6_2
Kowalczykowski, S. C., Dixon, D. A., Eggleston, A. K., Lauder, S. D., and Rehrauer, W. M. (1994). Biochemistry of homologous recombination in Escherichia coli. Microbiol. Rev. 58, 401–465.
Lalonde, M., Calvert, H. E., and Pine, S. (1981). “Isolation and use of Frankia strains in actinorhizae formation,” in Current Perspectives in Nitrogen Fixation, eds A. H. Gibson and W. E. Newton (Canberra: Australian Academy of Science), 296–299.
Lambrecht, J. A., and Downs, D. M. (2013). Anthranilate phosphoribosyl transferase (TrpD) generates phosphoribosylamine for thiamine synthesis from enamines and phosphoribosyl pyrophosphate. ACS Chem. Biol. 8, 242–248. doi: 10.1021/cb300364k
Le Rudulier, D., and Bernard, T. (1986). Salt tolerance in Rhizobium: a possible role for betaines. FEMS Microb. Lett. 39, 67–72. doi: 10.1016/0378-1097(86)90062-5
Lee, J., Postmaster, A., Soon, H. P., Keast, D., and Carson, K. C. (2012). Siderophore production by actinomycetes isolates from two soil sites in Western Australia. Biometal 25, 285–296. doi: 10.1007/s10534-011-9503-9
Levy, I., Horvath, A., Azevedo, M., de Alexandre, R. B., and Stratakis, C. A. (2011). Phosphodiesterase function and endocrine cells: links to human disease and roles in tumor development and treatment. Curr. Opin. Pharmacol 11, 689–697. doi: 10.1016/j.coph.2011.10.003
Lynd, L. R., Weimer, P. J., Van Zyl, W. H., and Pretorius, I. S. (2002). Microbial cellulose utilization: fundamentals and biotechnology. Microbiol. Mol. Biol. Rev. 66, 506–577. doi: 10.1128/mmbr.66.3.506-577.2002
Maksimov, I. V., Abizgil’dina, R. R., and Pusenkova, L. I. (2011). Plant growth promoting microorganisms as alternative to chemical protection from pathogens. Prikl. Biokhim. Mikrobiol. 47, 373–385.
Mano, Y., and Nemoto, K. (2012). The pathway of auxin biosynthesis in plants. J. Exp. Bot. 63, 2853–2872. doi: 10.1093/jxb/ers091
Mansour, S. R., and Moussa, L. A. A. (2005). Role of gamma-radiation on spore germination and infectivity of Frankia strains CeI523 and CcI6 isolated from Egyptian Casuarina. Isotope Rad. Res. 37, 1023–1038.
Marchler-Bauer, A., Derbyshire, M. K., Gonzales, N. R., Lu, S., Chitsaz, F., Geer, L. Y., et al. (2015). CDD: NCBI’s conserved domain database. Nucleic Acids. Res. 43, D222–D226. doi: 10.1093/nar/gku1221
Marín-Rodríguez, M. C., Orchard, J., and Seymour, G. B. (2002). Pectate lyases, cell wall degradation and fruit softening. J. Exp. Bot. 53, 2115–2119. doi: 10.1093/jxb/erf089
Minh, B. Q., Nguyen, M. A., and Von Haeseler, A. (2013). Ultrafast approximation for phylogenetic bootstrap. Mol. Biol. Evol. 30, 1188–1195. doi: 10.1093/molbev/mst024
Mok, D. W. S., Martin, R. C., Shan, X., and Mok, C. C. (2000). Genes encoding zeatin o-glycosyltransferases. Plant Growth Regul. 32, 285–287.
Nakouti, I., Sihanonth, P., and Hobbs, G. (2012). A new approach to isolating siderophore-producing actinobacteria. Lett. Appl. Microbiol. 55, 68–72. doi: 10.1111/j.1472-765X.2012.03259.x
Neeraja, C., Anil, K., Purushotham, P., Suma, K., Sarma, P., Moerschbacher, B. M., et al. (2010). Biotechnological approaches to develop bacterial chitinases as a bioshield against fungal diseases of plants. Crit. Rev. Biotechnol. 30, 231–241. doi: 10.3109/07388551.2010.487258
Nett, R. S., Montanares, M., Marcassa, A., Lu, X., Nagel, R., Charles, T. C., et al. (2017). Elucidation of gibberellin biosynthesis in bacteria reveals convergent evolution. Nat. Chem. Biol. 13, 69–74. doi: 10.1038/nchembio.2232
Ngom, M., Gray, K., Diagne, N., Oshone, R., Fardoux, J., Gherbi, H., et al. (2016a). Symbiotic performance of diverse Frankia strains on salt-stressed Casuarina glauca and Casuarina equisetifolia plants. Front. Plant Sci. 7:1331. doi: 10.3389/fpls.2016.01331
Ngom, M., Oshone, R., Diagne, N., Cissoko, M., Svistoonoff, S., Tisa, L. S., et al. (2016b). Tolerance to environmental stress by the nitrogen-fixing actinobacterium Frankia and its role in actinorhizal plants adaptation. Symbiosis 70, 17–29. doi: 10.1007/s13199-016-0396-9
Nguyen, L. T., Schmidt, H. A., Von Haeseler, A., and Minh, B. Q. (2015). IQ-TREE: a fast and effective stochastic algorithm for estimating maximum-likelihood phylogenies. Mol. Biol. Evol. 32, 268–274. doi: 10.1093/molbev/msu300
Niemann, J., and Tisa, L. S. (2008). Nitric oxide and oxygen regulate truncated hemoglobin gene expression in Frankia strain CcI3. J. Bacteriol. 190, 7864–7867. doi: 10.1128/JB.01100-08
Niemann, J. M., Tjepkema, J. D., and Tis, L. S. (2005). Identification of the truncated hemoglobin gene in Frankia. Symbiosis 39, 91–95.
Normand, N., Nouioui, I., Pujic, P., Fournier, P., Dubost, A., Klenk, H.-P., et al. (2018). Frankia canadensis sp. nov., isolated from root nodules of Alnus incana subspecies rugosa growing in Canada. Int. J. Evol. Syst. Microbiol. 68, 3001–3011. doi: 10.1099/ijsem.0.002939
Normand, P., Benson, D. R., Berry, A. M., and Tisa, L. S. (2014). “Family Frankiaceae,” in The Prokaryote–Actinobacteria, eds E. Rosenberg, E. F. Delong, S. Lory, E. Stackebrandt, and F. Thompson (Berlin: Springer-Verlag), 339–356. doi: 10.1007/978-3-642-30138-4_183
Normand, P., Lapierre, P., Tisa, L. S., Gogarten, J. P., Alloisio, N., Bagnarol, E., et al. (2007). Genome characteristics of facultatively symbiotic Frankia sp. strains reflect host range and host plant biogeography. Genome Res. 17, 7–15. doi: 10.1101/gr.5798407
Normand, P., Orso, S., Cournoyer, B., Jeannin, P., Chapelon, C., Dawson, J., et al. (1996). Molecular phylogeny of the genus Frankia and related genera and emendation of the family Frankiaceae. Int. J. Syst. Bacteriol. 46, 1–9. doi: 10.1099/00207713-46-1-1
Nouioui, I., Beauchemin, N., Cantor, M. N., Chen, A., Detter, J. C., Furnholm, T., et al. (2013). Draft genome sequence of Frankia sp. strain BMG5.12, a nitrogen-fixing actinobacterium isolated from Tunisian soils. Gen. Announc. 1:e00468-13. doi: 10.1128/genomeA.00468-13
Nouioui, I., Ghodhbane-Gtari, F., Jando, M., Tisa, L. S., Klenk, H.-P., and Gtari, M. (2019). Frankia torreyi sp. nov., the first actinobacterium of the genus Frankia Brunchorst, 1886, 174AL isolated in axenic culture. Antonie Van Leeuwenhoek 112, 57–65. doi: 10.1007/s10482-018-1131-8
Nouioui, I., Ghodhbane-Gtari, F., Montero-Calasanz, M. D. C., Göker, M., Meier-Kolthoff, J. P., Schumann, P., et al. (2016). Proposal of a type strain for Frankia alni (Woronin 1866) Von Tubeuf 1895, emended description of Frankia alni, and recognition of Frankia casuarinae sp. nov. and Frankia elaeagni sp. nov. Int. J. Syst. Evol. Microbiol. 66, 5201–5210. doi: 10.1099/ijsem.0.001496
Nouioui, I., Ghodhbane-Gtari, F., Klenk, H.-P., and Gtari, M. (2018a). Frankia saprophytica sp. nov. an atypical non-infective (Nod–) and non-nitrogen fixing (Fix–) actinobacterium isolated from Coriaria nepalensis root nodules. Int. J. Syst. Evol. Microbiol. 68, 1090–1095. doi: 10.1099/ijsem.0.002633
Nouioui, I., Ghodhbane-Gtari, F., Rhode, M., Sangal, V., Klenk, H. P., and Gtari, M. (2018b). Frankia irregularis sp. nov., an actinobacterium unable to nodulate its original host, Casuarina equisetifolia, but effectively nodulates members of the actinorhizal Rhamnales. Int. J. Syst. Evol. Microbiol. 68, 2883–2914. doi: 10.1099/ijsem.0.002914
Nouioui, I., Ghodhbane-Gtari, F., Montero-Calasanz, M. D. C., Rohde, M., Tisa, L. S., Gtari, M., et al. (2017a). Frankia inefficax sp. nov., an actinobacterial endophyte inducing ineffective, non nitrogen-fixing, root nodules on its actinorhizal host plants. Antonie Van Leeuwenhoek 110, 313–320. doi: 10.1007/s10482-016-0801-7
Nouioui, I., Ghodhbane-Gtari, F., Rohde, M., Klenk, H.-P., and Gtari, M. (2017b). Frankia coriariae sp. nov., an infective and effective microsymbiont isolated from Coriaria japonica. Int. J. Syst. Evol. Microbiol. 67, 1266–1270. doi: 10.1099/ijsem.0.001797
Nouioui, I., Gueddou, A., Ghodhbane-Gtari, F., Rhode, M., Gtari, M., and Klenk, H.-P. (2017c). Frankia asymbiotica sp. nov., a non-infective actinobacterium isolated from Morella californica root nodule. Int. J. Syst. Evol. Microbiol. 67, 4897–4901. doi: 10.1099/ijsem.0.002153
Nouioui, I., Montero-Calasanz, M. D. C., Ghodhbane-Gtari, F., Rohde, M., Tisa, L. S., Klenk, H.-P., et al. (2017d). Frankia discariae sp. nov.: an infective and effective microsymbiont isolated from the root nodule of Discaria trinervis. Arch. Microbiol. 199, 641–647. doi: 10.1007/s00203-017-1337-6
Ogasawara, Y., Yackley, B. J., Greenberg, J. A., Rogelj, S., and Melancon, C. E. I. I. I. (2015). Expanding our understanding of sequence-function relationships of type II polyketide biosynthetic gene clusters: bioinformatics-guided identification of Frankiamicin A from Frankia sp. EAN1pec. PLoS One 10:e0121505. doi: 10.1371/journal.pone.0121505
Oh, C. J., Kim, H. B., and An, C. S. (2003). Molecular cloning and complementation analysis of nifV gene from Frankia EuIK1 strain. Mol. Cell 15, 27–33.
Oh, C. J., Kim, H. B., Kim, J., Kim, W. J., Lee, H., and An, C. S. (2012). Organization of nif gene cluster in Frankia sp. EuIK1 strain, a symbiont of Elaeagnus umbellata. Arch. Microbiol. 194, 29–34. doi: 10.1007/s00203-011-0732-7
Oshone, R., Ngom, M., Chu, F., Mansour, S., Sy, M. O., Champion, A., et al. (2017). Genomic, transcriptomic, and proteomic approaches towards understanding the molecular mechanisms of salt tolerance in Frankia strains isolated from Casuarina trees. BMC Genomics 18:633. doi: 10.1186/s12864-017-4056-0
Ouyang, J., Shao, X., and Li, J. (2000). Indole-3-glycerol phosphate, a branchpoint of indole-3-acetic acid biosynthesis from the tryptophan biosynthetic pathway in Arabidopsis thaliana. Plant J. 24, 327–333.
Overbeek, R., Olson, R., Pusch, G. D., Olsen, G. J., Davis, J. J., Disz, J., et al. (2014). The SEED and the rapid annotation of microbial genomes using subsystems technology (RAST). Nucleic Acids Res. 42, D206–D214.
Paek, K. H., and Walker, G. C. (1987). Escherichia coli dnaK null mutants are inviable at high temperature. J. Bacteriol. 169, 283–290. doi: 10.1128/jb.169.1.283-290.1987
Péret, B., Swarup, R., Jansen, L., Devos, G., Auguy, F., Collin, M., et al. (2007). Auxin influx activity is associated with Frankia infection during actinorhizal nodule formation in Casuarina glauca. Plant physiol. 144, 1852–1862. doi: 10.1104/pp.107.101337
Perrine-Walker, F., Doumas, P., Lucas, M., Vaissayre, V., Beauchemin, N. J., Band, L. R., et al. (2010). Auxin carriers localization drives auxin accumulation in plant cells infected by Frankia in Casuarina glauca actinorhizal nodules. Plant. Physiol. 154, 1372–1380. doi: 10.1104/pp.110.163394
Persson, T., Benson, D. R., Normand, P., Vanden Heuvel, B., Pujic, P., Chertkov, O., et al. (2011). Genome sequence of “Candidatus Frankia datiscae” Dg1, the uncultured microsymbiont from nitrogen-fixing root nodules of the dicot Datisca glomerata. J. Bacteriol. 193, 7017–7018. doi: 10.1128/JB.06208-11
Pertry, I., Vaclavikova, K., Depuydt, S., Galuszka, P., Spichal, L., Temmerman, W., et al. (2009). Identification of Rhodococcus fascians cytokinins and their modus operandi to reshape the plant. Proc. Nat. Acad. Sci. U.S.A. 106, 929–934. doi: 10.1073/pnas.0811683106
Pertry, I., Vaclavikova, K., Gemrotova, M., Spichal, L., Galuszka, P., Depuydt, S., et al. (2010). Rhodococcus fascians impacts plant development through the dynamic fas-mediated production of a cytokinin mix. Mol. Plant Microbe Interact. 23, 1164–1174. doi: 10.1094/MPMI-23-9-1164
Pompella, A., Visvikis, A., Detata, V., and Casini, A. F. (2003). The changing faces of glutathione, a cellular protagonist. Biochem. Pharmacol. 66, 1499–1503. doi: 10.1016/s0006-2952(03)00504-5
Pozzi, A. C., Bautista-Guerrero, H. H., Abby, S. S., Herrera-Belaroussi, A., Abrouk, D., Normand, P., et al. (2018). Robust Frankia phylogeny, species delineation and intraspecies diversity based on multi-locus sequence analysis (MLSA) and single-locus strain typing (SLST) adapted to a large sample size. Syst. Appl. Microbiol. 41, 311–323. doi: 10.1016/j.syapm.2018.03.002
Prat, D. (1989). Effects of some pure and mixed Frankia strains on seedling growth in different Alnus species. Plant Soil 113, 31–38. doi: 10.1007/bf02181918
Qi, Y., Kobayashi, Y., and Hulett, F. M. (1997). The pst operon of Bacillus subtilis has a phosphate-regulated promoter and is involved in phosphate transport but not in regulation of the pho regulon. J. Bacteriol. 179, 2534–2539. doi: 10.1128/jb.179.8.2534-2539.1997
Qian, Y., Lee, J. H., and Holmes, R. K. (2002). Identification of a DtxR-regulated operon that is essential for siderophore-dependent iron uptake in Corynebacterium diphtheriae. J. Bacteriol. 184, 4846–4856. doi: 10.1128/jb.184.17.4846-4856.2002
Reed, S. C., Cleveland, C. C., and Townsend, A. R. (2011). Functional ecology of free-living nitrogen fixation: a contemporary perpective. Annu. Rev. Ecol. Evol. Syst. 42, 489–512. doi: 10.1146/annurev-ecolsys-102710-145034
Rehan, M., El-Sharkawy, A., El-Keredy, A., and El-Fadly, G. (2015). BIodegradation oF s-triazine compounds using actinobacterium Frankia. Egypt. J. Genet. Cytol. 44, 265–280.
Rehan, M., Furnholm, T., Finethy, R. H., Chu, F., El-Fadly, G., and Tisa, L. S. (2014a). Copper tolerance in Frankia sp. strain EuI1c involves surface binding and copper transport. Appl. Microbiol. Biotechnol. 98, 8005–8015. doi: 10.1007/s00253-014-5849-6
Rehan, M., Kluge, M., Franzle, S., Kellner, H., Ullrich, R., and Hofrichter, M. (2014b). Degradation of atrazine by Frankia alni ACN14a: gene regulation, dealkylation, and dechlorination. Appl. Microbiol. Biotechnol. 98, 6125–6135. doi: 10.1007/s00253-014-5665-z
Richards, J. W., Krumholz, G. D., Chval, M. S., and Tisa, L. S. (2002). Heavy metal resistance patterns of Frankia strains. Appl. Environ. Microbiol. 68, 923–927. doi: 10.1128/aem.68.2.923-927.2002
Roca, A. I., and Cox, M. M. (1990). The RecA protein: structure and function. Crit. Rev. Biochem. Mol. Biol. 25, 415–456.
Rodriguez, H., and Fraga, R. (1999). Phosphate solubilizing bacteria and their role in plant growth promotion. Biotechnol. Adv. 17, 319–339. doi: 10.1016/s0734-9750(99)00014-2
Rossolini, G. M., Schippa, S., Riccio, M. L., Berlutti, F., Macaskie, L. E., and Thaller, M. C. (1998). Bacterial nonspecific acid phosphohydrolases: physiology, evolution and use as tools in microbial biotechnology. Cell. Mol. Life Sci. 54, 833–850. doi: 10.1007/s000180050212
Saha, M., Sarkar, S., Sarkar, B., Sharma, B. K., Bhattacharjee, S., and Tribedi, P. (2016). Microbial siderophores and their potential applications: a review. Environ. Sci. Pollut. Res. Int. 23, 3984–3999. doi: 10.1007/s11356-015-4294-0
Sakakibara, H. (2006). Cytokinins: activity, biosynthesis, and translocation. Annu. Rev. Plant Biol. 57, 431–449. doi: 10.1146/annurev.arplant.57.032905.105231
Sasakawa, C. T. H. (2003). Salt tolerance of Casuarina equisetifolia and Frankia Ceq1 strain isolated from the root nodules of C. equisetifolia. Soil Sci. Plant Nutr. 49, 215–222. doi: 10.1080/00380768.2003.10410000
Sasso, S., Ramakrishnan, C., Gamper, M., Hilvert, D., and Kast, P. (2004). Characterization of the secreted chorismate mutase from the pathogen Mycobacterium tuberculosis. FEBS J. 272, 375–389. doi: 10.1111/j.1742-4658.2004.04478.x
Sayed, W., El-Sharouny, H., Zahran, H., and Ali, W. (2002). Composition of Casuarina leaf litter and its influence on Frankia-Casuarina symbiosis in soil. Folia. Microbiol. 47, 429–434. doi: 10.1007/bf02818703
Schwencke, J., and Carú, M. (2001). Advances in actinorhizal symbiosis: host plant-Frankia interactions, biology, and applications in arid land reclamation. A review. Arid. Land. Res. Manag. 15, 285–327. doi: 10.1080/153249801753127615
Shantharam, S., and Mattoo, A. K. (1997). Enhancing biological nitrogen fixation: an appraisal of current and alternative technologies for N input into plants. Plant Soil 194, 205–216. doi: 10.1007/978-94-011-5744-5_20
Sharma, S. B., Sayyed, R. Z., Trivedi, M. H., and Gobi, T. A. (2013). Phosphate solubilizing microbes: sustainable approach for managing phosphorus deficiency in agricultural soils. Springerplus 2:587. doi: 10.1186/2193-1801-2-587
Singh, R. P., and Jha, P. N. (2015). Molecular identification and characterization of rhizospheric bacteria for plant growth promoting ability. Int. J. Curr. Biotechnol. 3, 12–18.
Smith, K. F., Bibb, L. A., Schmitt, M. P., and Oram, D. M. (2009). Regulation and activity of a zinc uptake regulator, Zur, in Corynebacterium diphtheriae. J. Bacteriol. 191, 1595–1603. doi: 10.1128/JB.01392-08
Solans, M., Vobis, G., Cassa ìn, F., Luna, V., and Wall, L. G. (2011). Production of phytohormones by root-associated saprophytic actinomycetes isolated from the actinorhizal plant Ochetophila trinervis. World J. Microbiol. Biotechnol. 27, 2195–2202. doi: 10.1007/s11274-011-0685-7
Souza, A. L., Invitti, A. L., Rego, F. G., Monteiro, R. A., Klassen, G., Souza, E. M., et al. (2010). The involvement of the nif-associated ferredoxin-like genes fdxA and fdxN of Herbaspirillum seropedicae in nitrogen fixation. J. Microbiol. 48, 77–83. doi: 10.1007/s12275-009-0077-y
Steele, D. B., Ramirez, K., and Stowers, M. D. (1989). Host plant growth response to inoculation with Frankia. Plant Soil 118, 139–142.
Stevens, G. A., and Berry, A. M. (1988). Cytokinin secretion by Frankia sp. HFP ArI3 in defined medium. Plant Physiol. 87, 15–16. doi: 10.1104/pp.87.1.15
Tak, H., Ahmad, F., Babalola, O. O., and Inam, A. (2012). Growth, photosynthesis and yield of chickpea as influenced by urban wastewater and different levels of phosphorus. Int. J. Plant. Res. 2, 6–13. doi: 10.5923/j.plant.20120202.02
Tarkowski, P., Ge, L., Yong, J. W. H., and Tan, S. N. (2009). Analytical methods for cytokinins. Trend. Anal. Chem. Rev. Argent. Microbiol. 28, 323–335. doi: 10.1016/j.trac.2008.11.010
Tisa, L. S., Beauchemin, N., Gtari, M., Sen, A., and Wall, L. G. (2013). What stories can the Frankia genomes start to tell us? J. Biosci. 38, 719–726. doi: 10.1007/s12038-013-9364-1
Tisa, L. S., Oshone, R., Sarkar, I., Ktari, A., Sen, A., and Gtari, M. (2016). Genomic approaches toward understanding the actinorhizal symbiosis: an update on the status of the Frankia genomes. Symbiosis 70, 5–16. doi: 10.1007/s13199-016-0390-2
Tjepkema, J. D., Cashon, R. E., Beckwith, J., and Schwintzer, C. R. (2002). Hemoglobin in Frankia, a nitrogen-fixing actinomycete. Appl. Environ. Microbiol. 68, 2629–2631. doi: 10.1128/aem.68.5.2629-2631.2002
Udwary, D. W., Gontang, E. A., Jones, A. C., Jones, C. S., Schultz, A. W., Winter, J. M., et al. (2011). Significant natural product biosynthetic potential of actinorhizal symbionts of the genus Frankia, as revealed by comparative genomic and proteomic analyses. Appl. Environ. Microbiol. 77, 3617–3625. doi: 10.1128/AEM.00038-11
Walsh, C. T., Liu, J., Rusnak, F., and Sakaitani, M. (1990). Molecular studies on enzymes in chorismate metabolism and enterobactin biosynthetic pathway. Chem. Rev. 90, 1105–1129. doi: 10.1021/cr00105a003
Wandersman, C., and Delepelaire, P. (2004). Bacterial iron sources: from siderophores to hemophores. Annu. Rev. Microbiol. 58, 611–647. doi: 10.1146/annurev.micro.58.030603.123811
Wang, W., Qiu, Z., Tan, H., and Cao, L. (2014). Siderophore production by actinobacteria. Biometals 27, 623–631. doi: 10.1007/s10534-014-9739-2
Wheeler, C. T., Crozier, A., and Sandberg, G. (1984). The biosynthesis of indole-3-acetic-acid by Frankia. Plant Soil 78, 99–104. doi: 10.1007/978-94-009-6158-6_10
Whitelaw, M. A. (2000). Growth promotion of plants inoculated with phosphate-solubilizing fungi. Adv. Agron. 66, 99–151. doi: 10.1016/s0065-2113(08)60948-7
Wietzke, M., and Bahl, H. (2012). The redox-sensing protein Rex, a transcriptional regulator of solventogenesis in Clostridium acetobutylicum. Appl. Microbiol. Biotechnol. 96, 749–761. doi: 10.1007/s00253-012-4112-2
Willmann, R., and Nurnberger, T. (2012). How plant lysin motif receptors get activated: lessons learned from structural biology. Sci. Signal. 5:e28. doi: 10.1126/scisignal.2003274
Wong, W. S., Tan, S. N., Ge, L., Chen, X., and Yong, J. W. H. (2015). “The Importance of Phytohormones and Microbes in Biofertilizers,” in Bacterial Metabolites in Sustainable Agroecosystem, ed. D. K. Maheshwari (Berlin: Springer), 105–158. doi: 10.1007/978-3-319-24654-3_6
Yang, S. F., and Hoffman, N. E. (1984). Ethylene biosynthesis and its regulation in higher plants. Ann. Rev. Plant Physiol. 35, 155–189. doi: 10.1146/annurev.arplant.35.1.155
Zhang, Z., Lopez, M. F., and Torrey, J. G. (1984). A comparison of cultural characteristics and infectivity of Frankia isolates from root nodules of Casuarina species. Plant Soil 78, 79–90. doi: 10.1007/978-94-009-6158-6_8
Zhao, W., Ye, Z., and Zhao, J. (2007). RbrA, a cyanobacterial rubrerythrin, functions as a FNR-dependent peroxidase in heterocysts in protection of nitrogenase from damage by hydrogen peroxide in Anabaena sp. PCC 7120. Mol. Microbiol. 66, 1219–1230. doi: 10.1111/j.1365-2958.2007.05994.x
Keywords: PGPB, actinobacteria, biotic and abiotic stress, symbiosis, biofertilizers
Citation: Nouioui I, Cortés-albayay C, Carro L, Castro JF, Gtari M, Ghodhbane-Gtari F, Klenk H-P, Tisa LS, Sangal V and Goodfellow M (2019) Genomic Insights Into Plant-Growth-Promoting Potentialities of the Genus Frankia. Front. Microbiol. 10:1457. doi: 10.3389/fmicb.2019.01457
Received: 19 March 2019; Accepted: 11 June 2019;
Published: 04 July 2019.
Edited by:
Camille Eichelberger Granada, Universidade do Vale do Taquari – Univates, BrazilReviewed by:
Chengshu Wang, Shanghai Institutes for Biological Sciences (CAS), ChinaJuan F. Martin, Universidad de León, Spain
Tiago Falcon Lopes, Hospital de Clínicas de Porto Alegre, Brazil
Copyright © 2019 Nouioui, Cortés-albayay, Carro, Castro, Gtari, Ghodhbane-Gtari, Klenk, Tisa, Sangal and Goodfellow. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Imen Nouioui, aW1lbi5ub3Vpb3VpQG5ld2Nhc3RsZS5hYy51aw==; bm91aW91aWltZW5AeWFob28uZnI=