 Rocio Trastoy Pena1†
Rocio Trastoy Pena1† Lucia Blasco1†
Lucia Blasco1† Antón Ambroa1
Antón Ambroa1 Bertha González-Pedrajo2
Bertha González-Pedrajo2 Laura Fernández-García1
Laura Fernández-García1 Maria López1
Maria López1 Ines Bleriot1
Ines Bleriot1 German Bou1
German Bou1 Rodolfo García-Contreras3*†
Rodolfo García-Contreras3*† Thomas Keith Wood4
Thomas Keith Wood4 Maria Tomás1*†
Maria Tomás1*†- 1Deapartamento de Microbiología y Parasitología, Complejo Hospitalario Universitario A Coruña (CHUAC), Instituto de Investigación Biomédica (INIBIC), Universidad de A Coruña (UDC), A Coruña, Spain
- 2Departamento de Genética Molecular, Instituto de Fisiología Celular, Universidad Nacional Autónoma de México, Mexico City, Mexico
- 3Departamento de Microbiología y Parasitología, Facultad de Medicina, Universidad Nacional Autónoma de México, Mexico City, Mexico
- 4Department of Chemical Engineering, Pennsylvania State University, University Park, PA, United States
Quorum sensing (QS) is a communication mechanism between bacteria that allows specific processes to be controlled, such as biofilm formation, virulence factor expression, production of secondary metabolites and stress adaptation mechanisms such as bacterial competition systems including secretion systems (SS). These SS have an important role in bacterial communication. SS are ubiquitous; they are present in both Gram-negative and Gram-positive bacteria and in Mycobacterium sp. To date, 8 types of SS have been described (T1SS, T2SS, T3SS, T4SS, T5SS, T6SS, T7SS, and T9SS). They have global functions such as the transport of proteases, lipases, adhesins, heme-binding proteins, and amidases, and specific functions such as the synthesis of proteins in host cells, adaptation to the environment, the secretion of effectors to establish an infectious niche, transfer, absorption and release of DNA, translocation of effector proteins or DNA and autotransporter secretion. All of these functions can contribute to virulence and pathogenesis. In this review, we describe the known types of SS and discuss the ones that have been shown to be regulated by QS. Due to the large amount of information about this topic in some pathogens, we focus mainly on Pseudomonas aeruginosa and Vibrio spp.
Introduction
Microorganisms coexist in competitive environments with other species, and they must develop different survival strategies to compete for space, nutrients and ecological niches. Bacteria have developed several molecular mechanisms that enable them to survive under stress conditions in different environments. The general stress response (RpoS) (Battesti et al., 2011), tolerance to reactive oxygen species (ROS) (Zhao and Drlica, 2014; Van den Bergh et al., 2017), energy metabolism (cytochrome bd complex) (Korshunov and Imlay, 2010) and Tau metabolism (Javaux et al., 2007), drug efflux pumps (Blanco et al., 2016), SOS response (Baharoglu and Mazel, 2014), (p)ppGpp signaling under starvation conditions (Hauryliuk et al., 2015), toxin-antitoxin (TA) systems (Wood et al., 2013) and quorum sensing (QS), which we will discuss in detail in this review, are the main molecular mechanisms of tolerance and bacterial persistence (Harms et al., 2016; Trastoy et al., 2018).
Quorum sensing acts by monitoring cell density through chemical signals that allow communication between bacteria in order to regulate the expression of genes involved in virulence, competition, pathogenicity and resistance (Nealson et al., 1970; Hawver et al., 2016; Paul et al., 2018). In general, QS systems are species-dependent and contribute to processes such as cell maintenance, biofilm formation and horizontal gene transfer. QS also plays a role in other events involving the synchronization of the whole population such as antibiotic production (Abisado et al., 2018), natural competence (Shanker and Federle, 2017), sporulation (Rai et al., 2015) and the expression of secretion systems (SS). In this review, we will focus on the relationship between QS networks and SS in two important bacterial pathogens Pseudomonas aeruginosa and Vibrio spp.
QS Network
To explain the structure and functioning of the QS network, we will focus on Gram-negative bacteria, in which the signaling pathways are better described. In general terms, QS systems are composed of synthase proteins that produce QS signals, QS signals, and response regulators that bind QS signals and reprogram gene expression (Ng and Bassler, 2009). N-acyl homoserine lactones (AHLs) are the most common QS signals in Gram-negative bacteria (Geske et al., 2008). Other QS signals include autoinducer-2 (AI-2) in Vibrio harveyi (Surette et al., 1999), PQS (Pseudomonas quinolone signal) (Pesci et al., 1999), DSF (diffusible signaling factor) in Xanthomonas campestris (Barber et al., 1997), indole in Escherichia coli (Lee and Lee, 2010), and PAME (hydroxyl-palmitic acid methyl ester) in Ralstonia solanacearum (Flavier et al., 1997). The LuxI/LuxR QS system of Vibrio fischeri is the prototypical model system for Gram-negative bacteria (Engebrecht et al., 1983; Engebrecht and Silverman, 1984). Homologs of luxI (which encode synthase proteins) and luxR (which encode response regulators) are present in many bacteria (Case et al., 2008). AHL signals are produced inside the cell and most of them are transported freely to the local environment. When the concentration of AHL reaches a certain level outside of the cell, the molecule re-enters the cell (or binds surface receptors) and binds/activates the LuxR-type receptor to alter gene expression. AHL signals with small structural differences are involved in the process of gene regulation (Fuqua et al., 1994; Whiteley et al., 2017; Paul et al., 2018).
Pseudomonas aeruginosa possesses three well-known QS systems: LasI/LasR, RhlI/RhlR, and PQS (Pseudomonas quinolone signal)/PqsR (MvfR). The Las system consists of LasI, a synthase protein which produces the AHL N-(3-oxododecanoyl)-L-homoserine lactone (3O-C12-HSL), and LasR, the transcriptional regulator (Seed et al., 1995; Stintzi et al., 1998; Kariminik et al., 2017). Likewise, the RhlI/RhlR system produces the N-hexanoyl-L-homoserine lactone (C4-HSL) signal and the RhlR transcriptional regulator. Finally, the PQS system comprises 2-heptyl-3-hydroxy-4(1H)-quinolone (PQS signal) and the PqsR (MvfR) receptor (Xiao et al., 2006; Jimenez et al., 2012). In 2016, James and collaborators, analyzed the role of a new binding receptor for PQS signals, i.e., MexG, an inner membrane protein of the mexGHI-opmD operon and a component of a resistance-nodulation-cell division (RND) efflux pump (Hodgkinson et al., 2016).
Quorum quenching (QQ) enzymes have also been shown to be important in the functioning of QS systems (Zhang and Dong, 2004; Dong et al., 2007; Bzdrenga et al., 2017). Our research group has recently described a new QQ enzyme (AidA) which participates in the QS network in Acinetobacter baumannii clinical strains (Lopez et al., 2017b, 2018).
Secretion Systems
Bacterial pathogens secrete proteins through their cell membranes in a fundamental process that enables them to attack other microorganisms, evade the host immune system, produce tissue damage and invade the host cells. Secreted proteins can act as virulence factors that generate toxic products to the host cells and may also facilitate adhesion to these cells. Translocation of proteins across the phospholipid membranes is carried out by several types of SS (Green and Mecsas, 2016). SS play a significant role in bacterial communication. To date, 8 types of SS (T1SS, T2SS, T3SS, T4SS, T5SS, T6SS, T7SS, and T9SS) have been made defined on their structure, composition and activity (Figure 1). These differences can be attributed to the differences between Gram-negative and Gram-positive bacteria (Desvaux et al., 2009; Sato et al., 2010; Costa et al., 2015). The characteristics of each type of SS are described in detail below.
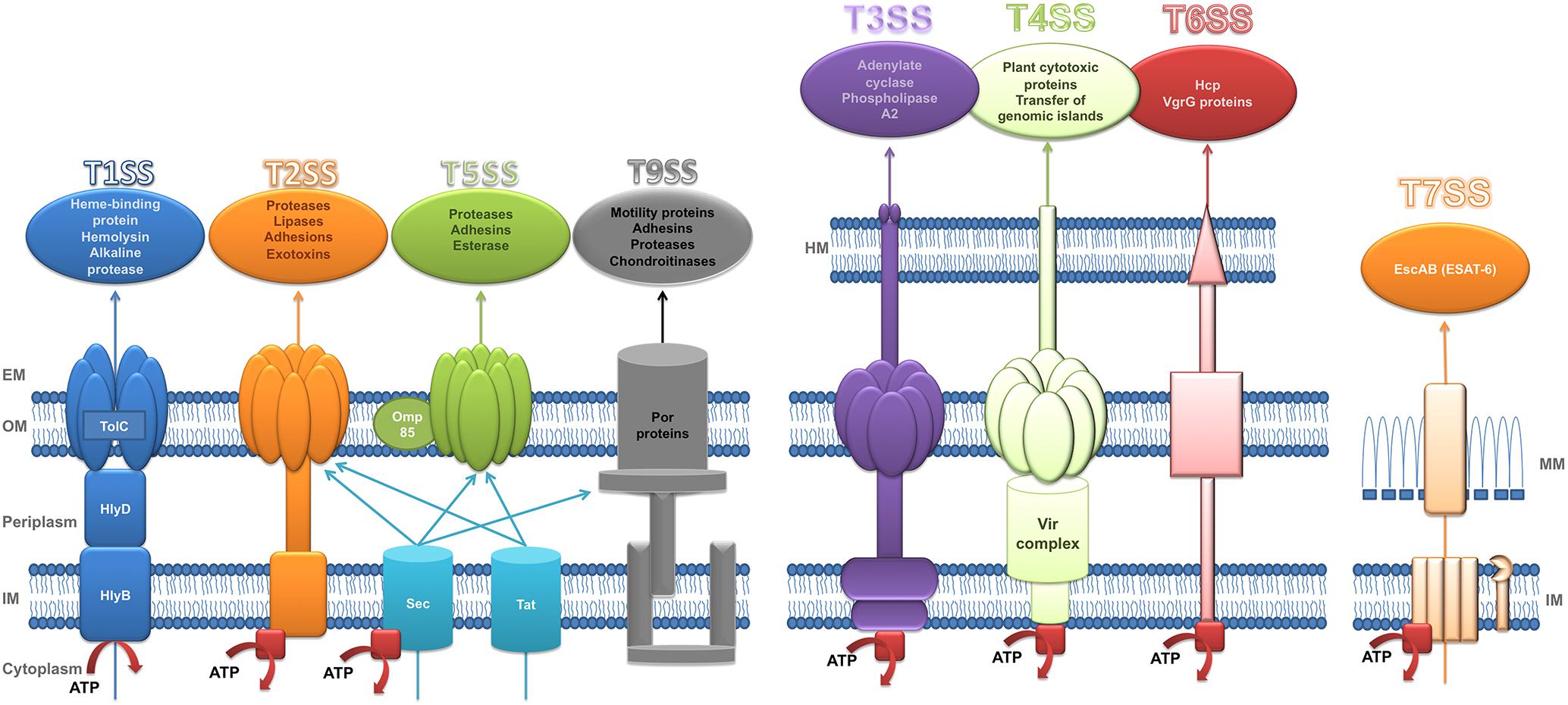
Figure 1. Structure of secretion systems. Schematic representation of secretory systems: type I (T1SS), type II (T2SS), type V (T5SS), type IX (T9SS), type III (T3SS), type IV (T4SS), type VI (T6SS), and type VII (T7SS). The type I pathway is exemplified by hemolysin A (HlyA) secretion in E. coli where TolC, HlyD, and HlyB are the three components which constitute the channel to transport HlyA to extracellular space. Sec (general secretion route) and Tat (twin-arginine translocation pathway) transfer the substrates of T2SS and T5SS across the inner membrane. Sec also participates in the transport of T9SS substrates across de inner membrane. T9SS is also called Por Secretion System (PoSS). T4SS is represented by VirB/D system of Agrobacterium tumefaciens. The T7SS is based on a system in Mycobacteria. Red squares represent the ATPases. HM, host membrane; EM, extracellular medium; OM, outer membrane; IM, inner membrane; MM, mycomembrane. Substrates secreted by each secretory system are included in a circle in the top of the figure. Adapted from Tseng et al. (2009).
T1SS
The type I secretion system is widely distributed in Gram-negative bacteria such as P. aeruginosa, Salmonella enterica, Neisseria meningitidis, and E. coli (Thomas et al., 2014).
The type I secretion system (T1SS), which has three structural elements (ABC transporter protein, a membrane fusion protein and an outer membrane factor), can transfer substrates across both bacterial membranes in Gram negative bacteria in a one-step process (Green and Mecsas, 2016). T1SS uses proteins as substrates, e.g., proteases and lipases of different sizes and with different functions; these proteins have a C-terminal uncleaved secretion signal which is recognized by the ABC transporter protein to form the translocation complex (Delepelaire, 2004; Kanonenberg et al., 2013).
There are two systems described so far that regulate the expression and secretion of substrates of T1SS, the Has system of S. marcescens and P. aeruginosa, and the hemolysins of Vibrio cholerae, N. meningitidis and in particular of uropathogenic E. coli (Thomas et al., 2014).
T2SS
The type II secretion system (T2SS), which is conserved in most Gram negative bacteria, is responsible for secreting folded proteins from the periplasm. These proteins are first transported through the IM by the general secretory (Sec) or twin-arginine translocation (Tat) pathways, and then secreted from the periplasm into the extracellular medium by the T2SS (Nivaskumar and Francetic, 2014; Green and Mecsas, 2016).
The Sec pathway consists of three structural parts: a protein targeting component, a motor protein and a membrane integrated conducting channel called SecYEG translocase. This mechanism transports unfolded proteins with a hydrophobic sequence at the N-terminus. Moreover, the secreted protein either remains in the periplasm or is transported to the extracellular space. The proteins may contain a SecB-specific signal sequence for transport to the periplasm or the extracellular milieu; however, if it has the signal recognition particle (SRP) signal it can follow the SRP pathway and remain in the inner membrane (Green and Mecsas, 2016; Tsirigotaki et al., 2017).
By contrast, the Tat secretion pathway consists of 2–3 subunits, TatA and TatB, which form a unique multifunctional protein in Gram-positive bacteria, and TatC. This mechanism translocates folded proteins with a twin-arginine motif. In Gram-positive bacteria, most proteins are transported out of the cell, while in Gram-negative bacteria the protein can remain in the periplasm or it can be translocated to the extracellular space by the T2SS (Patel et al., 2014; Green and Mecsas, 2016).
The T2SS, a complex structure composed of 15 proteins, named general secretion pathway proteins (Gsp) in E. coli (Korotkov et al., 2012), Eps in V. cholera (Abendroth et al., 2009; Sloup et al., 2017) and Xcp in P. aeruginosa (Filloux et al., 1998; Robert et al., 2005), has a wide range of substrates with diverse functions, although all share one feature, an N-terminal signal which enables them pass to the periplasm via the Sec or Tat secretion mechanisms (Nivaskumar and Francetic, 2014; Green and Mecsas, 2016).
The main function of the T2SS is to acquire nutrients (Nivaskumar and Francetic, 2014). It is responsible for secreting numerous exoproteins, most of which are hydrolytic enzymes and other proteins such as toxins, adhesins and cytochromes that have various roles in respiration, biofilm formation and motility (Nivaskumar and Francetic, 2014). The T2SS has been described in various environmental strains and also human pathogens such as V. cholera (Overbye et al., 1993), P. aeruginosa, Aeromonas sp. and enterotoxigenic Escherichia coli (ETEC) (Nivaskumar and Francetic, 2014).
T3SS
The type III secretion system (T3SS) or injectisome, is a double-membrane-embedded apparatus found in multiple pathogenic Gram-negative bacteria such as Salmonella spp., Yersinia spp., enteropathogenic and enterohemorrhagic E. coli, Shigella spp. and Pseudomonas spp. (Cornelis, 2006; Gaytan et al., 2016; Deng et al., 2017). This complex nanomachine promotes the transfer of virulence proteins called effectors from the bacterial cytoplasm into the eukaryotic cell in a single step (Galan and Waksman, 2018).
The T3SS is composed of approximately 25 proteins assembled in three main structures: the basal body, a set of rings spanning the two membranes of the bacterium; a hollow needle-shaped component through which the semi-unfolded effectors are transported (these first two structures are collectively called “needle complex”); and the translocon, made up of a hydrophilic protein that serves as a scaffold for forming a translocation pore, constituted by two hydrophobic proteins, which is inserted into the host cell membrane and through which effectors are directly translocated. A unique set of effectors is delivered by each pathogen, which subverts specific host-cell signaling pathways to allow bacterial colonization (Izore et al., 2011; Notti and Stebbins, 2016; Deng et al., 2017).
The export apparatus associated with the basal body is formed by five poly topic inner membrane proteins that are essential for substrate secretion. This protein complex, together with a cytoplasmic sorting platform and the ATPase complex are responsible for substrate recruitment and classification, and for energizing the secretion process enabling chaperone-effector dissociation and protein unfolding for initial entry into the T3SS central channel that serves as the secretion pathway. These components are highly conserved between different T3SS systems and with the flagella, which is evolutionarily related to the injectisome (Abby and Rocha, 2012; Galan and Waksman, 2018; Lara-Tejero and Galan, 2019).
Several effectors of T3SS have been described such as ExoS, ExoT, ExoU, and ExoY in P. aeruginosa; Tir and EspE in E. coli and YopE, YopH, YopM, YopJ/P, YopO/YpkA, and YopT in Yersinia sp. (Cornelis and Van Gijsegem, 2000).
T4SS
The type IV secretion system family is found in Gram-negative and Gram-positive bacteria as well as in Archaea. T4SS is the most cosmopolitan secretion system and differs from other SS as it is able to transfer DNA in addition to proteins (Cascales and Christie, 2003). More specifically, T4SS is capable of performing contact-dependent secretion of effector molecules into eukaryotic cells, conjugative transfer of mobile DNA elements and also exchange of DNA without any contact with the outside of the cell (Green and Mecsas, 2016; Grohmann et al., 2018). T4SS can be divided on the basis of its functionality into two subfamilies: conjugation systems and effector translocators. Conjugation systems are responsible for the transfer of antibiotic resistance genes and virulence determinants among bacteria. The effector translocators introduce virulence factors into the host cell (Christie, 2016). However, in Gram-negative bacteria T4SS has been divided into two different subfamilies: IVA and IVB. The E. coli conjugation apparatuses and VirB/D system of Agrobacterium tumefaciens are the models used to study the structure of type IVA of T4SS (Grohmann et al., 2018). The VirB/D apparatus consists of 12 proteins which form a complex envelope-spanning structure that facilitate the translocation function. Two of these proteins, VirB2 and VirB5, make up the pilus, while another three proteins act as ATPases, and VirB1 is a lytic transglycosylase (Costa et al., 2015; Green and Mecsas, 2016). The Legionella pneumophila Dot/Icm (Defective for organelle trafficking/Intracellular multiplication) system is the model used to study the IVB subfamily of T4SS (Nagai and Kubori, 2011; Grohmann et al., 2018).
T5SS
The type V secretion system is unique because its substrates transport themselves across the outer membrane. The substrates use the Sec translocase to pass through the inner membrane to the periplasm space. Various different types of T5SS have been identified: autotransporters (T5aSS), two-partner passenger-translocators (T5bSS), trimeric autotransporters (T5cSS), hybrid autotransporters (T5dSS) and inverted autotransporters (T5eSS) (Henderson et al., 2004; Leo et al., 2012; Rojas-Lopez et al., 2017). In general, the T5SS transports proteins across the asymmetric outer membrane (OM) that contains lipopolysaccharides, through their own C-terminal translocation domain that inserts into the OM as a β-barrel to complete the secretion of the N-terminal passenger domain via the barrel pore. Several periplasmic chaperones also participate in transport through the OM, specifically the β-barrel assembly machinery (BAM complex) and the translocation and assembly module (TAM complex) facilitate protein secretion (Rojas-Lopez et al., 2017).
A T5SS has been described in human pathogens such as Bordetella pertussis and Haemophilus influenzae, which have two-partner SS and uropathogenic E. coli, which has chaperone-usher systems (Costa et al., 2015; Green and Mecsas, 2016).
YadA of Yersinia enterocolitica and SadA of Salmonella are T5SS type c (Leo et al., 2012). Intimin of E. coli and invasin of enteropathogenic Yersinia spp. are type Ve SS (Leo et al., 2012).
A self-transporter (T5aSS) (Wilhelm et al., 2007) and three T5bSS: LepA /LepB system (Kida et al., 2008), the CupB system (Ruer et al., 2008) and PdtA/PdtB system (Faure et al., 2014), have been reported in P. aeruginosa. In B. cenocepacia, four T5SS (Holden et al., 2009) have been found, two with pertactin domains and two with haemagglutinin autotransporters; this last type is also present in S. maltophilia (Ryan et al., 2009).
T6SS
The type VI secretion system is widely represented in Gram-negative bacteria (Coulthurst, 2013; Gallique et al., 2017b). T6SS is an integrated secretion device within the membrane and it transfers substrates, which are toxic effectors to eukaryotic (Pukatzki et al., 2007) and prokaryotic cells (Russell et al., 2014). It plays a crucial role in the pathogenesis and competition among bacteria (Ho et al., 2014; Zoued et al., 2014; Costa et al., 2015; Gallique et al., 2017a). The origin of T6SS is related to bacteriophages (Leiman et al., 2009). T6SS is a huge apparatus and consists of 13 core components organized into a trans-membrane complex, a baseplate-like structure at the cytoplasmic face of the inner membrane, and a sheathed inner tube, which is the effector delivery module that is ejected to the target cell. The tube-sheath complex is assembled from the baseplate in the cytoplasm and the hollow tube is built from hexamers of the hemolysin co-regulated protein (Hcp). The sheath contracts and pushes the tube with the associated effectors into targeted cells, using a puncturing mechanism similar to the one used by the contractile tails of phages (Russell et al., 2011, 2014; Cianfanelli et al., 2016; Green and Mecsas, 2016; Galan and Waksman, 2018).
T7SS
Type VII secretory system has been described in some Gram-positive bacteria such as Staphylococcus aureus and in species of Mycobacterium and Corynebacterium. This SS was reported for the first time in 2003 in Mycobacterium tuberculosis and it was called ESX-1 (Stanley et al., 2003), which is an important virulence factor in M. tuberculosis. To date, five T7SS have been identified in Mycobacterium sp. but the transport mechanisms across the mycobacterial membrane are almost unknown (Costa et al., 2015; Ates et al., 2016; Green and Mecsas, 2016).
Most of the substrates of T7SS belong to EscAB clan which includes six protein families: Esx, PE, PPE, LXG, DUF2563, and DUF2580. ESAT-6 is a M. tuberculosis protein which belongs to Esx family and which is secreted with EsxB (CFP-10) (Ates et al., 2016).
T9SS
The type IX secretion system (T9SS) or Por secretion system (PorSS) is the most recently discovered system (Lasica et al., 2017). Its function is to transport molecules across the outer membrane. Its substrates must include a Sec signal, which allows transfer of proteins through the inner membrane with the aid of the Sec system. The T9SS system has been described in almost all members of the phylum Bacteroidetes, but it has mainly been studied in oral pathogens such as Porphyromonas gingivalis and Tannerella forsythia. In P. gigivalis, the T9SS system consists of 16 proteins with structural and functional activity, and another two proteins involved in the regulation of the transport process (Sato et al., 2010; Lasica et al., 2017).
Regulation of Secretion Systems by Quorum Sensing Networks (Table 1)
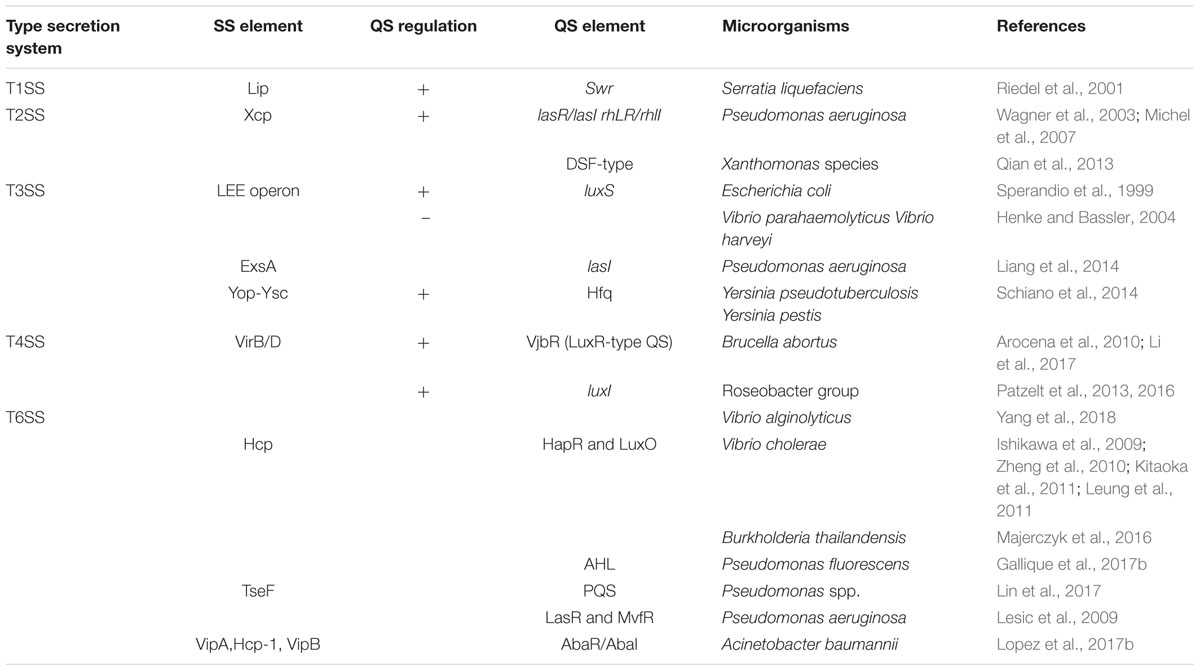
Table 1. Pathogens and QS elements related to secretion systems.
Pseudomonas aeruginosa
T1SS
Transcriptional studies in P. aeruginosa suggest that in this bacterium T1SS is positively regulated by QS, since the expression of its effector, the alkaline protease AprA, depends on QS. In addition, the genes of the AprA inhibitor aprI and the structural genes aprDEF also appear to be positively regulated by QS (Hentzer et al., 2003; Schuster et al., 2003; Wagner et al., 2003).
T2SS
Three T2SS systems, the Xcp, Hxc and Txc systems, have been described in P. aeruginosa. The first of these, Xcp, secretes the QS regulated virulence factors elastase A and B (LasA and LasB) as well as the exotoxin A (ExoA) and it is itself positively regulated by QS (Figure 2). Accordingly, recently it was demonstrated by ChIPseq analysis that MvfR (the receptor of the PQS autoinducer) is able to directly bind xcpQ-xcpP-xcpR regions and this is related to their induction in the presence of MvfR (Maura et al., 2016).
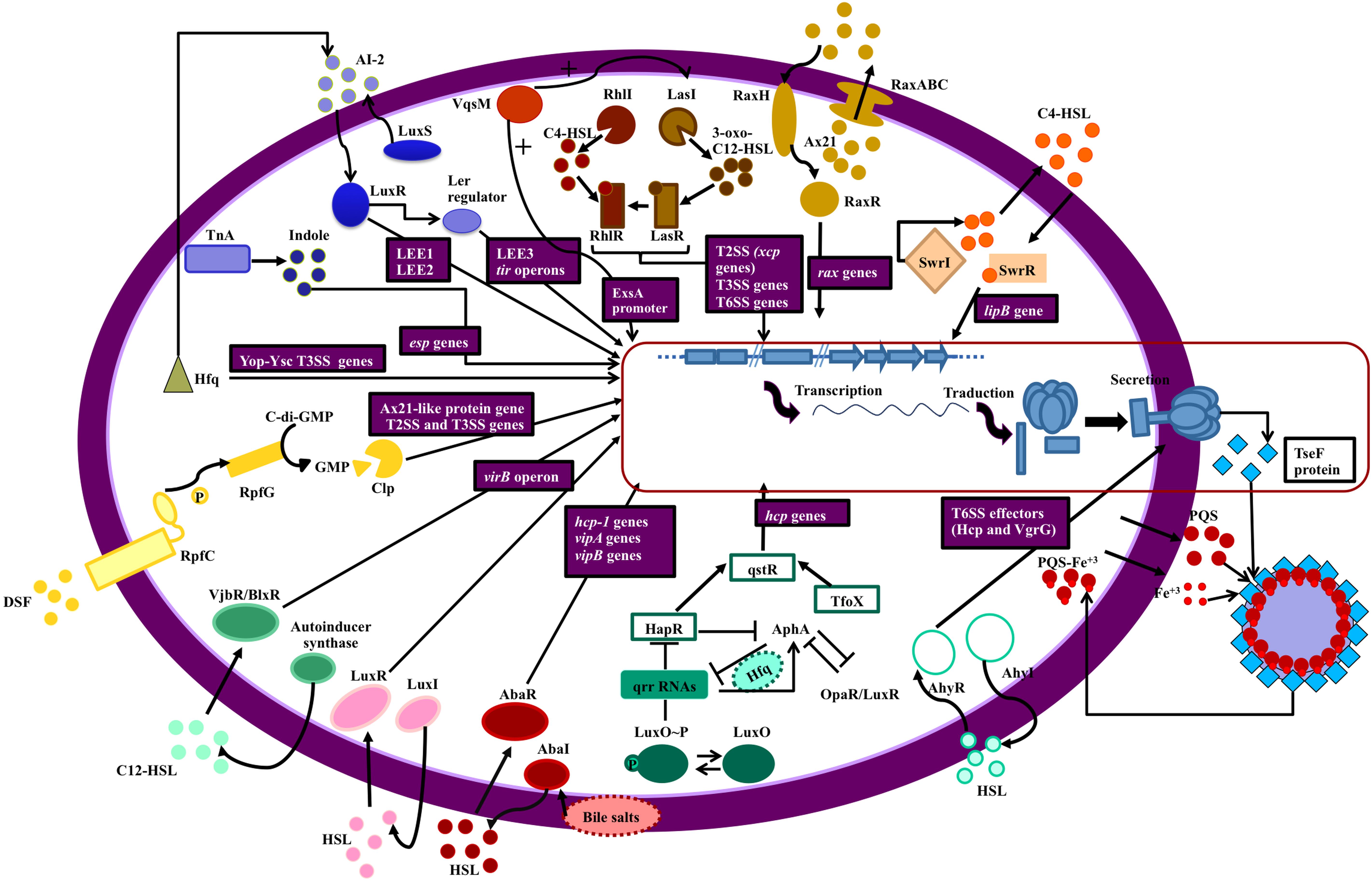
Figure 2. Secretion systems and QS network elements. The figure shows the relationship between QS networks and expression of secretion systems (blue squares). The genes regulated by QS are in purple boxes. Each QS network is represented by a different color. Starting at the top right of the figure: The swr QS system of S. liquefaciens controls the lipB genes of the T1SS (orange); Ax21 (QS effector) and QS system Rax regulate RaxABC TOSS (T1SS) in gram negative bacteria (ochre); QS (RhlIR and LasIR) regulates expression of T2SS, T3SS, and T6SS in P. aeruginosa (brown); VqsM (an AraC family transcription factor) interacts with the LasIR and ExsA promoters (a master regulator of T3SS) in P. aeruginosa (dark orange); T2SS is regulated by LuxS/LuxI/AI-2 QS in E. coli and indole production by TnA (tryptophanase) regulates esp genes expression (T2SS) in this bacterium (blue); in Yersinia sp. the Hfq chaperone is connected with QS (AI-2) and regulates the Yop-Ysc type III secretion system (T3SS) (green); in Xanthomonas sp. T2SS and T3SS are regulated by DSF (diffusible signal factor) which is a quorum sensing signal (yellow); T4SS (virB operon) is regulated by VjbR (LuxR like protein) and LuxI in Brucella (turquoise) and Roseobacter (pink), respectively; a connection between Acinetobacter baumanii QS (AbaI/AbaR, controlled by bile salts) and T6SS has been established (maroon); in Vibrio sp. there is a complex network which relates QS (LuxO/HapR/TfoX) with T6SS (aquamarine); AhyRI (a QS network) in Aeromonas sp. and P. atrosepticum is involved in Hcp and VgrG secretion (sky blue) and finally, iron is transported across the cell membrane accompanied by PQS, a quorum sensing signal in P. aeruginosa, and this process depend on Tse a substrate of T6SS, which binds to OMVs (outer membrane vesicles) containing PQS- Fe3+.
The second T2SS, Hxc, is regulated by the availability of phosphate and secretes LapA a low-molecular weight alkaline phosphatase (Wagner et al., 2003; Michel et al., 2007). Two genes, xphA and xqhA, which encode the PaQa subunit of the Xcp functional hybrid system, have been described. These genes, which are located outside the xcp locus, are regulated by environmental conditions but not by QS, in contrast to what occurs with the rest of the Xcp system (Michel et al., 2007, 2011). In contrast to the first two systems, the third system Txc has just recently been described and so far only identified in a region of genome plasticity of the strain PA7; it is regulated by a two component system (TtsSR) and secretes the chitin binding protein CpbE (Cadoret et al., 2014).
T3SS
Current evidence suggest that as in Vibrio spp., QS in P. aeruginosa negatively regulates the expression of T3SS, specifically the RhlI/RhlR system, as transcription of the T3SS genes and secretion of ExoS increase significantly in a rhlI mutant and return to basal levels on the addition of exogenous C4-HSL (Bleves et al., 2005; Kong et al., 2009; Figure 2). In agreement, the expression of exoS is also negatively regulated by QS, specifically by the RhlI/RhlR system, as well as by the stationary phase sigma factor RpoS (Hogardt et al., 2004).
The fact that the T3SS genes do not appear to be repressed by QS in some global transcriptomic studies with mutants may be explained by the presence of high calcium concentrations in the media, or by the lack of resolution of DNA microarrays (Hentzer et al., 2003; Schuster et al., 2003). More striking is the fact that some QS inhibitors like 6-gingerol and coumarin inhibit rather than increase the expression of T3SS (Zhang et al., 2018). Nevertheless, these studies were done in the presence of high calcium, and QS-independent inhibition of T3SS has not been ruled out. Moreover, a recent study in the PA01 strain, using a lasR rhlR double mutant, demonstrated that it remains virulent in a murine abscess model, despite that it does not produce QS-dependent virulence factors and that the secretion of ExoT and ExoS is fully functional in this mutant. Hence the authors hypothesized that T3SS is the cause of the remaining virulence (Soto-Aceves et al., 2019).
The P. aeruginosa QS network and its T3SS are also related by the fact that VqsM, an AraC-family transcription factor, binds to both the promoter region of lasI and the promoter of exsA, which encodes a master regulator of the T3SS, regulating both mechanisms (Liang et al., 2014; Figure 2).
T6SS
The T6SS is involved in iron transport, and a connection has been observed between T6SS and QS through the TseF protein, which is a substrate of T6SS and interacts with PQS (Lin et al., 2017; Figure 2).
In P. aeruginosa, three loci which encode T6SS have been found to be regulated by QS proteins (LasR and MvfR) (Lesic et al., 2009). Expression of the second loci, H2-T6SS, is regulated by the Las and Rhl QS systems in PAO1 strains (Sana et al., 2012; Figure 2) and by the direct binding of MvfR in PA14 (Maura et al., 2016).
Vibrio sp.
T2SS
The formation of biofilms has multifactorial regulation in V. cholerae as in other pathogens. The QS network controls directly biofilm production which is related to type II secretion system in V. cholerae (Teschler et al., 2015). Several proteins such as RbmA, RbmC and Bap1, which are involved in the formation of biofilms, are transported by T2SS. In addition, mutant strains with inactivated T2SS have reduced biofilm formation (Johnson et al., 2014; Teschler et al., 2015).
T3SS
In V. parahaemolyticus and V. harveyi (unlike in E. coli), both the HAI-1 and AI-2 QS systems inhibit the expression of T3SS genes (Henke and Bassler, 2004). QS also represses T3SS during V. harveyi infections of gnotobiotic brine shrimp (Ruwandeepika et al., 2015). Waters et al. (2010) have described the regulatory pathway by which QS controls T3SS. At low cell density when LuxR is repressed, which entails the derepression of two promoters of the exsBA operon and the exsA operon, ExsA activates the expression of genes that encode the structural proteins of the type III secretion system. However, when the cell density is high, LuxR directly represses transcription of the PB promoter, preventing the production of ExsA and consequently decreasing the expression of structural genes of T3SS (Waters et al., 2010; Ball et al., 2017). OpaR inhibits the T3SS1 in V. parahaemolyticus which is the most important factor in its cytotoxicity (Gode-Potratz and McCarter, 2011).
T6SS
Several researchers have demonstrated the regulation of T6SS by QS networks in Vibrio spp. We present the main findings in this field here. In V. alginolyticus, activation of T6SS and the QS network has been found to be coordinated by the serine/threonine kinase PpkA cascade (Yang et al., 2018). PpkA2 is autophosphorylation and it transfers the phosphate group to VstR. Phosphorylated VstR promotes the expression of both of the T6SS in V. alginolyticus through the inhibition of LuxO activity, which acts to impede the expression of LuxR, a promoter of the T6SS. LuxR inhibits the expression of the first T6SS (T6SS1) or promotes the expression of the second T6SS (T6SS2) (Yang et al., 2018).
At low cell population density, LuxO is phosphorylated, which activates the expression of specific small regulatory RNAs (sRNAs) in conjunction with alternative sigma factor σ54 (Sheng et al., 2012). sRNAs inhibit the expression of LuxR with the help of RNA chaperone Hfq (Liu et al., 2011). However, at high cell population density, LuxO is dephosphorylated turning off the transcription of the sRNAs and allowing the translation of LuxR (Waters and Bassler, 2005; Milton, 2006). Sheng et al. (2012) also demonstrated that the expression of the hcp T6SS gene is growth phase-dependent and the QS regulators controls the haemolysin co-regulated protein, which is one of the main proteins of the T6SS functioning as an effector of the system and/or an effector binding protein (Figure 2). The phosphatase PppA also acts on the QS (modulating the transcription of LuxR) and the expression and secretion of hcp1 and hcp2 (Sheng et al., 2013). It is important to highlight that PppA permits the cross-talk between the two T6SS in V. alginolyticus (Sheng et al., 2013).
Rpo N (σ54) collaborates with QS in the regulation of T6SS genes. It is involved in the regulation of the expression of hcp and vgrG3 operons that encode T6SS secreted molecules, but does not control the genes that encode the structural and sheath components of T6SS (Ishikawa et al., 2009; Dong and Mekalanos, 2012).
There are a few more studies in V. cholerae related to this topic than other species. Two QS autoinducers, CAI-I (cholerae autoinducer) and AI-2 (autoinducer-2), co-operate to control the gene expression depending on the cell density (Ng and Bassler, 2009). Two enzymes are necessary for the biosynthesis of these autoinducers: CqsA and LuxS, respectively (Schauder et al., 2001; Miller et al., 2002; Chen et al., 2002; Higgins et al., 2007). These signal molecules are detected by two sensor kinases, LuxQ (sensor of CAI-I) and CqsS (sensor of AI-2). Both pathways merge on LuxU, a phosphotranfer protein. At low cell density (LCD), the two sensor kinases phosphorylate LuxU due to the absence of their respective autoinducers. There are two histidine kinases which also contribute to the phosphorylation of LuxU: VpsS and CqsR (Jung et al., 2015). Then, LuxU transfers the phosphorylate group to a DNA-binding response regulator protein called LuxO. Phosphorylated LuxO activates the expression of sRNA molecules (known as qrr1-4) when the cell density is low thanks to the interaction with the alternative sigma factor σ54 (Freeman et al., 2000; Lenz et al., 2004). In conjunction with the RNA-binding protein Hfq, LuxO represses the expression of HapR (Lenz et al., 2004), a TetR-family global transcriptional regulator which acts on QstR (Tsou et al., 2009; Shao and Bassler, 2014; Watve et al., 2015; Figure 2). HapR is accumulated when the cell density is high (Lenz et al., 2004) because LuxO is not phosphorylated and transcription of the sRNAs is blocked. QstR is a master regulator of the T6SS belonging to the LuxR-type family of regulators (Jaskólska et al., 2018). QstR binds to the promoter region of the T6SS cluster inducing the expression of the genes. The regulation of the T6SS by cAMP-CRP pathway is not clear, but it is possible that it influences T6SS genes through regulation of QS and chitin-induced competency (Liang et al., 2007; Blokesch, 2012). It is known that CRP positively regulates T6SS (Ishikawa et al., 2009). Apart from the activation of QstR via QS, it is also regulated by chitin and arabinose (Lo Scrudato and Blokesch, 2012, 2013).
The expression of the three T6SS gene clusters in V. cholerae requires TfoX, CytR, HapR, and QstR for the highest level of expression (Watve et al., 2015). CytR and TfoX are required for the expression of the T6SS genes but their regulatory effects are only mediated by QstR (Figure 2).
Other Pathogens
T1SS
The swr QS system, which controls swarming motility, regulates the Lip secretion system, a T1SS responsible for the secretion of lipases, metalloproteases and S-layer proteins in Serratia liquefaciens MG1 (Riedel et al., 2001). The swr QS system consists of SwrI, which synthesizes C4-HSL, and SwrR, which regulates gene transcription after binding the diffusible signal C4-HSL. QS-mediated regulation of lipB, which encodes the LipB exporter, was demonstrated in swrI mutants with luxAB insertions, in which the level of secreted proteins was lower (Riedel et al., 2001; Figure 2). Other relationships between T1SS and QS have also been observed. The rice pathogen recognition XA21 receptor recognizes a sulphated peptide (axYS22) derived from the Ax21 protein (activator of XA21-mediated immunity) and confers resistance to Xanthomonas oryzae strains. Ax21 may have a key biological role because it is conserved in Xanthomonas spp., Xylella fastidiosa, and Stenotrophomonas maltophilia. Ax21 requires RaxABC TOSS (type I secretion system) for secretion and activity. The expression of rax genes which encode T1SS has been demonstrated to be QS-dependent due to the cell-density dependency (Han et al., 2011). These data indicate that Ax21 could have a role as a signaling molecule and a direct relationship between the QS network and T1SS is established (Figure 2; Lee et al., 2006).
T2SS
In Xanthomonas species, QS is mediated by the diffusible signal factor (DSF). A proteomic analysis conducted in 2013 revealed 33 proteins that are controlled by DSF. Their putative functions are associated with QS and include cellular processes, intermediary metabolism, oxidative adaption, macromolecule metabolism, cell-structure, protein catabolism, and hypothetical functions (Qian et al., 2013). In this study, it was observed that three genes encoding T2SS-dependent proteins and one gene which encodes Ax21 (activator of XA21-mediated immunity)-like protein are regulated by QS and are essential for virulence-associated functions, including extracellular protease, cell motility, antioxidative ability, extracellular polysaccharide biosynthesis (EPS), colonization, and biofilm (Qian et al., 2013; Figure 2).
T3SS
The relationship between QS and T3SS in E. coli was first demonstrated by Sperandio et al. (1999), who showed that expression of the locus of enterocyte effacement (LEE) operons that encode the T3SS is activated by QS in both enterohemorrhagic (EHEC) and enteropathogenic (EPEC) E. coli due to transcriptional control of the LEE operons by LuxS, which directly activates the LEE1 and LEE2 operons and indirectly activates (via the Ler regulator) the LEE3 and tir operons (Figure 2). These researchers proposed that activation of the T3SS by the AI-2 autoinducer synthesized by commensal E. coli resident in the large intestine could explain the high infectivity of E. coli O157: H7, which has an infectious dose of about 50 bacterial cells (Sperandio et al., 1999).
The major virulence factors of EHEC and EPEC are intimin (T5eSS), Tir (the receptor for intimin) and the three secreted proteins EspA, EspB and EspD. T3SS functions in the secretion of the Tir and Esp proteins. The LuxR-type response regulator SdiA negatively regulates the expression of EspD and intimin in the same bacterium, indicating multifactorial regulation of the T3SS by bacterial QS signals (Kanamaru et al., 2000).
Indole, which is produced by tryptophanase (TnA) in enteric bacteria and reaches high concentrations in the gut, is another signaling molecule that influences expression of T3SS in E. coli (Lee et al., 2007, 2008). Indole increases the production and secretion of the translocators EspA and EspB in EHEC O157:H7 (Hirakawa et al., 2009; Figure 2); hence, indole promotes the development of attaching and effacing (A/E) lesions in HeLa cells.
The involvement of the RNA chaperone protein Hfq, which also participates in QS, in T3SS expression was demonstrated in Yersinia pseudotuberculosis and Yersinia pestis (Schiano et al., 2014; Figure 2). Moreover, Schiano et al., 2014 have demonstrated the regulation of T3SS by QS through virulence regulators LcrF and YmoA in Y. pseudotuberculosis (Amy, 2018).
In Aeromonas hydrophila, an unique QS system, encoded in ahyR/ahyI loci, has been described (Vilches et al., 2009; Garde et al., 2010). Vilches et al. (2009) have used the A. hydrophila AH-3 strain to study the T3SS regulation. AH-3: ahyI and AH-3: ahyR mutants have reduced activity of the aopN-aopB promoter (promoter of T3SS components) compared to the wild-type strain (Figure 2). So they concluded that QS could be involved in the positive regulation of the production of the T3SS component in the AH-3 strain (Vilches et al., 2009).
T4SS
In Brucella abortus, there is a clear relationship between the QS network and T4SS. For the virB operon, which encodes the T4SS regulated by VjbR, a LuxR-type QS is responsible for the virulence characteristics of B. abortus (Li et al., 2017). The virB operon is responsible for establishing the replicative niche of the bacterium once it enters the host cell. The T4SS in B. abortus, as in other bacteria, translocates effector proteins into the host cell to avoid the immune defense mechanisms, making it one of the two main virulence factors for Brucella. Arocena et al. (2010) described the binding site of VjbR to the virB operon (Li et al., 2017). Otherwise, the conjugation process between two members of the Roseobacter group mediated by T4SS, encoded in RepABC-type plasmids, is controlled by the QS network. This was demonstrated by construction of luxI mutant and the addition of external long chain AHLs, which restored the phenotype (Patzelt et al., 2013, 2016; Figure 2).
T6SS
Quorum sensing has been reported to control expression of T6SS toxin-immunity systems in Burkholderia thailandensis. Moreover, a new role for T6SS in constraining the proliferation of QS mutants has been described in B. thailandensis (Majerczyk et al., 2016). Interestingly, it has been observed that T6SS effectors function as cell-to-cell signals in a Pseudomonas fluorescens MFE01 strain lacking the AHL QS pathway (Gallique et al., 2017b).
In A. hydrophila, Hcp and VgrG- two of the “core” proteins and also effectors of the T6SS- secretion have been suggested to be regulated by the AhyRI QS regulon (Khajanchi et al., 2009; Figure 2). Finally, our research group has described the association between T6SS machinery and the activation of the QS system by bile salts in A. baumannii clinical strains (Lopez et al., 2017a; Figure 2).
T7SS
As with other bacteria, Mycobacterium spp regulate biofilm formation by QS (Virmani et al., 2018). The second messenger c-di-cGMP, an intracellular signaling molecule, coordinates biofilm production and QS signaling (Sharma et al., 2014). Both M. tuberculosis (Kulka et al., 2012) and diverse species of non-tuberculous mycobacteria (M. smegmatis, M. marinum, M. fortuitum, M. chelonae, M. ulcerans, M. abscessus, M. avium, and M. bovis) produce biofilm depending on certain environmental conditions such as the availability of nutrients or the pH of the medium (Hall-Stoodley et al., 1998; Bardouniotis et al., 2003; Ojha et al., 2005; Marsollier et al., 2007; Johansen et al., 2009; Rhoades et al., 2009).
In the recent work of Lai et al. (2018) it was demonstrated that the espE, espF, espG, and espH genes, located in the T7SS ESX-1 operon, are crucial for sliding motility and biofilm formation in M. marinum. Esp proteins, which regulate substrate transport, are involved also in virulence. This paper clearly demonstrates the role of M. marinum T7SS in the production of biofilm which, as already mentioned, is related to QS (Lai et al., 2018).
The T7SS of S. aureus, a virulence factors export machinery, plays a key role in the promotion of bacterial survival and long-term persistence of subpopulations of staphylococci. The expression of T7SS is regulated by the bacterial interaction with host tissues (Lopez et al., 2017c) mediated by the secondary sigma factor (σB) (Schulthess et al., 2012). Schulthess et al. (2012) reported that the repression of esxA by σB is due to the transcription of sarA induced by σB, which leads to a strong repression of esxA. The activation of the esxA transcript, on the other hand, is stimulated by arlR, the response regulator of the ArlRS two-component system, SpoVG, a σ-dependent element, and the Agr quorum detection system (Schulthess et al., 2012). Agr QS system is composed by AIP (self-activating peptide), the inducer ligand of AgrC which is the receptor of the agr signal. In the case of the QS Agr system, the effector of global gene regulation is an important regulatory RNA, RNAIII (Novick and Geisinger, 2008).
T9SS
Moreover, an important relationship between T9SS and biofilm formation has been observed in periodontopathogenic pathogens such as Capnocytophaga ochracea, Porphyromonas spp., Fusobacterium spp. and Prevotella spp. (Kita et al., 2016). In the study by Kita et al. (2016), the participation of T9SS in the formation of biofilm of C. ochracea is demonstrated. The formation of biofilm of C. ochracea is crucial for the development of dental plaque and the same happens with other periodontal pathogens, in which it has also been seen that genes related to T9SS are present. Therefore, the components of the T9SS could be potential targets to inhibit the formation of biofilm and thus avoid the formation of dental plaque (McBride and Zhu, 2013; Kita et al., 2016). However, in depth analysis of the relationship between T9SS and QS network in different pathogens is required.
Discussion
To date, the T1SS, T2SS, T3SS, T4SS, T6SS, T7SS, and T9SS SS have been found to have important relationships with QS networks. The involvement of the T1SS system (Lip B which is part of the Lip exporter) in the QS network (swr quorum system) of S. liquefaciens MG1 has been investigated (Riedel et al., 2001). In P. aeruginosa, two QS systems (lasR/lasI and rhLR/rhlI) are linked to T2SS system by microarrays and proteomic studies (Chapon-Herve et al., 1997; Wagner et al., 2003; Michel et al., 2007), and DSF-type systems are also linked to T2SS in Xanthomonas species through proteome analysis (Qian et al., 2013). The QS signal AI-2 has been associated with a T3SS system in E. coli (Sperandio et al., 1999) and Vibrio spp. (Henke and Bassler, 2004). Moreover, this T3SS system has been related to QS proteins in another two pathogens, P. aeruginosa and Yersinia spp. (Liang et al., 2014; Schiano et al., 2014).
Several T4SS (virB operon) are controlled by VjbR protein which is a LuxR-type quorum-sensing regulator in B. abortus (Arocena et al., 2010; Li et al., 2017). Moreover, in the Roseobacter group, the conjugation of plasmids, which encode T4SS, is QS-controlled and the QS system may detect a broad range of long-chain AHLs at the cell surface (Patzelt et al., 2013, 2016).
There is a wealth of information relating the T6SS to QS in pathogens such as Vibrio spp. For example, Hcp and VasH from the T6SS system in V. cholerae are involved in QS (Ishikawa et al., 2009; Zheng et al., 2010; Kitaoka et al., 2011; Leung et al., 2011; Yang et al., 2018). For Pseudomonas spp,. there are numerous works where the different T6SS are regulated by QS networks (Lesic et al., 2009; Gallique et al., 2017b; Lin et al., 2017). In other pathogens as Burkholderia thailandensis (Majerczyk et al., 2016), and in A. baumannii, the relationship between QS and SS has begun to be studied (Lopez et al., 2017a).
In M. marinum, the relationship between biofilm formation, which is tightly connected with QS, and T7SS, has been demonstrated (Lai et al., 2018). Also in S. aureus, the Agr QS network has been related to T7SS (Schulthess et al., 2012). An important relationship between T9SS and biofilm formation has been observed in periodontopathogenic pathogens (Kita et al., 2016). Finally, although the involvement of T5SS secretion system in virulence, motility and competence is well-known, these systems and their association with QS must be studied in greater depth in order to clarify their roles.
Taking P. aeruginosa as a reference point, the positive effect of QS in the expression of T1SS and T2SS could be related to the fact that this organism secretes exoproducts that are public goods (proteases and lipases); hence, it is better to produce and secrete these compounds when a high cell density is reached, since these products are costly and the benefits associated to their production are higher at high cell densities. Similarly, the T6SS, which is involved in killing competitors by contact, will be more efficient at high cell densities since the probability of finding target bacteria is higher. In contrast, the T3SS appears to be negatively regulated by QS, and this may be related to its role in an acute infection and its “inhibition by QS” may be a way to facilitate the transition to a chronic infection state. In addition to its well established role in infections, T3SS has a broader ecological role suggested by its role in killing biofilm associated Acanthamoeba castellanii amoeba (Matz et al., 2008). Furthermore, it was recently demonstrated that T3SS is susceptible of cheating by mutants that do not produce it, allowing their establishment in infections (Czechowska et al., 2014); hence, the selective forces that act over T3SS are complex.
Therefore, research into the relationship between QS and SS must be further developed in order to better understand human infections.
Author Contributions
RTP, LB, AA, BG-P, LF-G, ML, IB, and GB developed the redaction of the manuscript, figures and table. RG-C, TW, and MT designed the review, assigned writing tasks to co-authors, contributed to writing and proofread the final version.
Funding
This study was funded by grant PI16/01163 awarded to MT within the State Plan for R+D+I 2013–2016 (National Plan for Scientific Research, Technological Development and Innovation 2008–2011) and co-financed by the ISCIII-Deputy General Directorate for Evaluation and Promotion of Research – European Regional Development Fund “A way of making Europe” and Instituto de Salud Carlos III FEDER, Spanish Network for the Research in Infectious Diseases (REIPI, RD16/0016/0001 and RD16/0016/0006) and by the Study Group on Mechanisms of Action and Resistance to Antimicrobials, GEMARA (SEIMC, http://www.seimc.org/). MT was financially supported by the Miguel Servet Research Programme (SERGAS and ISCIII). RTP and LF-G were financially supported by, respectively, a post-speciality grant awarded by the Fundación Novoa Santos (CHUAC-SERGAS, Galicia) and predoctoral fellowship from the Xunta de Galicia (GAIN, Axencia de Innovación). RG-C research is supported by the grant PAPIIT-UNAM number IN214218. BG-P was supported by grants PAPIIT-UNAM IN209617 and CONACYT 284081.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Abby, S. S., and Rocha, E. P. (2012). The non-flagellar type III secretion system evolved from the bacterial flagellum and diversified into host-cell adapted systems. PLoS Genet. 8:e1002983. doi: 10.1371/journal.pgen.1002983
Abendroth, J., Mitchell, D. D., Korotkov, K. V., Johnson, T. L., Kreger, A., Sandkvist, M., et al. (2009). The three-dimensional structure of the cytoplasmic domains of EpsF from the type 2 secretion system of Vibrio cholerae. J. Struct. Biol. 166, 303–315. doi: 10.1016/j.jsb.2009.03.009
Abisado, R. G., Benomar, S., Klaus, J. R., Dandekar, A. A., and Chandler, J. R. (2018). Bacterial quorum sensing and microbial community interactions. mBio 9:e02331-17. doi: 10.1128/mBio.02331-17
Amy, S. (2018). Unraveling the Regulatory Relationship between Quorum Sensing and the Type III Secretion System in Yersinia pseudotuberculosis. Ph.D. thesis, University of Nottingham, Nottingham.
Arocena, G. M., Sieira, R., Comerci, D. J., and Ugalde, R. A. (2010). Identification of the quorum-sensing target DNA sequence and N-Acyl homoserine lactone responsiveness of the Brucella abortus virB promoter. J. Bacteriol. 192, 3434–3440. doi: 10.1128/JB.00232-10
Ates, L. S., Houben, E. N., and Bitter, W. (2016). Type VII secretion: a highly versatile secretion system. Microbiol. Spectr. 4. doi: 10.1128/microbiolspec.VMBF-0011-2015
Baharoglu, Z., and Mazel, D. (2014). SOS, the formidable strategy of bacteria against aggressions. FEMS Microbiol. Rev. 38, 1126–1145. doi: 10.1111/1574-6976.12077
Ball, A. S., Chaparian, R. R., and van Kessel, J. C. (2017). Quorum sensing gene regulation by LuxR/HapR master regulators in vibrios. J. Bacteriol. 199:e00105-17. doi: 10.1128/JB.00105-17
Barber, C. E., Tang, J. L., Feng, J. X., Pan, M. Q., Wilson, T. J., Slater, H., et al. (1997). A novel regulatory system required for pathogenicity of Xanthomonas campestris is mediated by a small diffusible signal molecule. Mol. Microbiol. 24, 555–566. doi: 10.1046/j.1365-2958.1997.3721736.x
Bardouniotis, E., Ceri, H., and Olson, M. E. (2003). Biofilm formation and biocide susceptibility testing of Mycobacterium fortuitum and Mycobacterium marinum. Curr. Microbiol. 46, 28–32. doi: 10.1007/s00284-002-3796-4
Battesti, A., Majdalani, N., and Gottesman, S. (2011). The RpoS-mediated general stress response in Escherichia coli. Annu. Rev. Microbiol. 65, 189–213. doi: 10.1146/annurev-micro-090110-102946
Blanco, P., Hernando-Amado, S., Reales-Calderon, J. A., Corona, F., Lira, F., Alcalde-Rico, M., et al. (2016). Bacterial multidrug efflux pumps: much more than antibiotic resistance determinants. Microorganisms 4:E14. doi: 10.3390/microorganisms4010014
Bleves, S., Soscia, C., Nogueira-Orlandi, P., Lazdunski, A., and Filloux, A. (2005). Quorum sensing negatively controls type III secretion regulon expression in Pseudomonas aeruginosa PAO1. J. Bacteriol. 187, 3898–3902. doi: 10.1128/JB.187.11.3898-3902.2005
Blokesch, M. (2012). Chitin colonization, chitin degradation and chitin-induced natural competence of Vibrio cholerae are subject to catabolite repression. Environ. Microbiol. 14, 1898–1912. doi: 10.1111/j.1462-2920.2011.02689.x
Bzdrenga, J., Daudé, D., Rémy, B., Jacquet, P., Plener, L., Elias, M., et al. (2017). Biotechnological applications of quorum quenching enzymes. Chem. Biol. Interact. 267, 104–115. doi: 10.1016/j.cbi.2016.05.028
Cadoret, F., Ball, G., Douzi, B., and Voulhoux, R. (2014). Txc, a new type II secretion system of Pseudomonas aeruginosa strain PA7, is regulated by the TtsS/TtsR two-component system and directs specific secretion of the CbpE chitin-binding protein. J. Bacteriol. 196, 2376–2386. doi: 10.1128/JB.01563-14
Cascales, E., and Christie, P. J. (2003). The versatile bacterial type IV secretion systems. Nat. Rev. Microbiol. 1, 137–149. doi: 10.1038/nrmicro753
Case, R. J., Labbate, M., and Kjelleberg, S. (2008). AHL-driven quorum-sensing circuits: their frequency and function among the Proteobacteria. ISME J. 2, 345–349. doi: 10.1038/ismej.2008.13
Chapon-Herve, V., Akrim, M., Latifi, A., Williams, P., Lazdunski, A., and Bally, M. (1997). Regulation of the xcp secretion pathway by multiple quorum-sensing modulons in Pseudomonas aeruginosa. Mol. Microbiol. 24, 1169–1178. doi: 10.1046/j.1365-2958.1997.4271794.x
Chen, X., Schauder, S., Potier, N., Van Dorsselaer, A., Pelczer, I., Bassler, B. L., et al. (2002). Structural identification of a bacterial quorum-sensing signal containing boron. Nature 415, 545–549. doi: 10.1038/415545a
Christie, P. J. (2016). The mosaic type IV secretion systems. Ecosal Plus 7. doi: 10.1128/ecosalplus.ESP-0020-2015
Cianfanelli, F. R., Monlezun, L., and Coulthurst, S. J. (2016). Aim, load, fire: the type VI secretion system, a bacterial nanoweapon. Trends Microbiol. 24, 51–62. doi: 10.1016/j.tim.2015.10.005
Cornelis, G. R. (2006). The type III secretion injectisome. Nat. Rev. Microbiol. 4, 811–825. doi: 10.1038/nrmicro1526
Cornelis, G. R., and Van Gijsegem, F. (2000). Assembly and function of type III secretory systems. Annu. Rev. Microbiol. 54, 735–774. doi: 10.1146/annurev.micro.54.1.735
Costa, T. R., Felisberto-Rodrigues, C., Meir, A., Prevost, M. S., Redzej, A., Trokter, M., et al. (2015). Secretion systems in Gram-negative bacteria: structural and mechanistic insights. Nat. Rev. Microbiol. 13, 343–359. doi: 10.1038/nrmicro3456
Coulthurst, S. J. (2013). The Type VI secretion system - a widespread and versatile cell targeting system. Res. Microbiol. 164, 640–654. doi: 10.1016/j.resmic.2013.03.017
Czechowska, K., McKeithen-Mead, S., Al Moussawi, K., and Kazmierczak, B. I. (2014). Cheating by type 3 secretion system-negative Pseudomonas aeruginosa during pulmonary infection. Proc. Natl. Acad. Sci. U.S.A. 111, 7801–7806. doi: 10.1073/pnas.1400782111
Delepelaire, P. (2004). Type I secretion in gram-negative bacteria. Biochim. Biophys. Acta 1694, 149–161. doi: 10.1016/j.bbamcr.2004.05.001
Deng, W., Marshall, N. C., Rowland, J. L., McCoy, J. M., Worrall, L. J., Santos, A. S., et al. (2017). Assembly, structure, function and regulation of type III secretion systems. Nat. Rev. Microbiol. 15, 323–337. doi: 10.1038/nrmicro.2017.20
Desvaux, M., Hebraud, M., Talon, R., and Henderson, I. R. (2009). Secretion and subcellular localizations of bacterial proteins: a semantic awareness issue. Trends Microbiol. 17, 139–145. doi: 10.1016/j.tim.2009.01.004
Dong, T. G., and Mekalanos, J. J. (2012). Characterization of the RpoN regulon reveals differential regulation of T6SS and new flagellar operons in Vibrio cholerae O37 strain V52. Nucleic Acids Res. 40, 7766–7775. doi: 10.1093/nar/gks567
Dong, Y. H., Wang, L. Y., and Zhang, L. H. (2007). Quorum-quenching microbial infections: mechanisms and implications. Philos. Trans. R. Soc. Lond. B Biol. Sci. 362, 1201–1211. doi: 10.1098/rstb.2007.2045
Engebrecht, J., Nealson, K., and Silverman, M. (1983). Bacterial bioluminescence: isolation and genetic analysis of functions from Vibrio fischeri. Cell 32, 773–781. doi: 10.1016/0092-8674(83)90063-6
Engebrecht, J., and Silverman, M. (1984). Identification of genes and gene products necessary for bacterial bioluminescence. Proc. Natl. Acad. Sci. U.S.A. 81, 4154–4158. doi: 10.1073/pnas.81.13.4154
Faure, L. M., Garvis, S., de Bentzmann, S., and Bigot, S. (2014). Characterization of a novel two-partner secretion system implicated in the virulence of Pseudomonas aeruginosa. Microbiology 160(Pt 9), 1940–1952. doi: 10.1099/mic.0.079616-0
Filloux, A., Michel, G., and Bally, M. (1998). GSP-dependent protein secretion in gram-negative bacteria: the Xcp system of Pseudomonas aeruginosa. FEMS Microbiol. Rev. 22, 177–198. doi: 10.1111/j.1574-6976.1998.tb00366.x
Flavier, A. B., Clough, S. J., Schell, M. A., and Denny, T. P. (1997). Identification of 3-hydroxypalmitic acid methyl ester as a novel autoregulator controlling virulence in Ralstonia solanacearum. Mol. Microbiol. 26, 251–259. doi: 10.1046/j.1365-2958.1997.5661945.x
Freeman, J. A., Lilley, B. N., and Bassler, B. L. (2000). A genetic analysis of the functions of LuxN: a two-component hybrid sensor kinase that regulates quorum sensing in Vibrio harveyi. Mol. Microbil. 35, 139–149. doi: 10.1046/j.1365-2958.2000.01684.x
Fuqua, W. C., Winans, S. C., and Greenberg, E. P. (1994). Quorum sensing in bacteria: the LuxR-LuxI family of cell density-responsive transcriptional regulators. J. Bacteriol. 176, 269–275. doi: 10.1128/jb.176.2.269-275.1994
Galan, J. E., and Waksman, G. (2018). Protein-injection machines in bacteria. Cell 172, 1306–1318. doi: 10.1016/j.cell.2018.01.034
Gallique, M., Bouteiller, M., and Merieau, A. (2017a). The type VI secretion system: a dynamic system for bacterial communication? Front. Microbiol. 8:1454. doi: 10.3389/fmicb.2017.01454
Gallique, M., Decoin, V., Barbey, C., Rosay, T., Feuilloley, M. G., Orange, N., et al. (2017b). Contribution of the Pseudomonas fluorescens MFE01 type VI secretion system to biofilm formation. PLoS One 12:e0170770. doi: 10.1371/journal.pone.0170770
Garde, C., Bjarnsholt, T., Givskov, M., Jakobsen, T. H., Hentzer, M., Claussen, A., et al. (2010). Quorum sensing regulation in Aeromonas hydrophila. J. Mol. Biol. 396, 849–857. doi: 10.1016/j.jmb.2010.01.002
Gaytan, M. O., Martinez-Santos, V. I., Soto, E., and Gonzalez-Pedrajo, B. (2016). Type three secretion system in attaching and effacing pathogens. Front. Cell. Infect. Microbiol. 6:129. doi: 10.3389/fcimb.2016.00129
Geske, G. D., O’Neill, J. C., and Blackwell, H. E. (2008). Expanding dialogues: from natural autoinducers to non-natural analogues that modulate quorum sensing in Gram-negative bacteria. Chem. Soc. Rev. 37, 1432–1447. doi: 10.1039/b703021p
Gode-Potratz, C. J., and McCarter, L. L. (2011). Quorum sensing and silencing in Vibrio parahaemolyticus. J. Bacteriol. 193, 4224–4237. doi: 10.1128/JB.00432-11
Green, E. R., and Mecsas, J. (2016). Bacterial secretion systems: an overview. Microbiol. Spectr. 4. doi: 10.1128/microbiolspec.VMBF-0012-2015
Grohmann, E., Christie, P. J., Waksman, G., and Backert, S. (2018). Type IV secretion in Gram-negative and Gram-positive bacteria. Mol. Microbiol. 107, 455–471. doi: 10.1111/mmi.13896
Hall-Stoodley, L., Keevil, C. W., and Lappin-Scott, H. M. (1998). Mycobacterium fortuitum and Mycobacterium chelonae biofilm formation under high and low nutrient conditions. J. Appl. Microbiol. 85(Suppl. 1), 60s–69s. doi: 10.1111/j.1365-2672.1998.tb05284.x
Han, S. W., Sriariyanun, M., Lee, S. W., Sharma, M., Bahar, O., Bower, Z., et al. (2011). Small protein-mediated quorum sensing in a Gram-negative bacterium. PLoS One 6:e29192. doi: 10.1371/journal.pone.0029192
Harms, A., Maisonneuve, E., and Gerdes, K. (2016). Mechanisms of bacterial persistence during stress and antibiotic exposure. Science 354:aaf4268. doi: 10.1126/science.aaf4268
Hauryliuk, V., Atkinson, G. C., Murakami, K. S., Tenson, T., and Gerdes, K. (2015). Recent functional insights into the role of (p)ppGpp in bacterial physiology. Nat. Rev. Microbiol. 13, 298–309. doi: 10.1038/nrmicro3448
Hawver, L. A., Jung, S. A., and Ng, W. L. (2016). Specificity and complexity in bacterial quorum-sensing systems. FEMS Microbiol. Rev. 40, 738–752. doi: 10.1093/femsre/fuw014
Henderson, I. R., Navarro-Garcia, F., Desvaux, M., Fernandez, R. C., and Ala’Aldeen, D. (2004). Type V protein secretion pathway: the autotransporter story. Microbiol. Mol. Biol. Rev. 68, 692–744. doi: 10.1128/MMBR.68.4.692-744.2004
Henke, J. M., and Bassler, B. L. (2004). Quorum sensing regulates type III secretion in Vibrio harveyi and Vibrio parahaemolyticus. J. Bacteriol. 186, 3794–3805. doi: 10.1128/JB.186.12.3794-3805.2004
Hentzer, M., Wu, H., Andersen, J. B., Riedel, K., Rasmussen, T. B., Bagge, N., et al. (2003). Attenuation of Pseudomonas aeruginosa virulence by quorum sensing inhibitors. EMBO J. 22, 3803–3815. doi: 10.1093/emboj/cdg366
Higgins, D. A., Pomianek, M. E., Kraml, C. M., Taylor, R. K., Semmelhack, M. F., and Bassler, B. L. (2007). The major Vibrio cholerae autoinducer and its role in virulence factor production. Nature 450, 883–886. doi: 10.1038/nature06284
Hirakawa, H., Kodama, T., Takumi-Kobayashi, A., Honda, T., and Yamaguchi, A. (2009). Secreted indole serves as a signal for expression of type III secretion system translocators in enterohaemorrhagic Escherichia coli O157:H7. Microbiology 155(Pt 2), 541–550. doi: 10.1099/mic.0.020420-0
Ho, B. T., Dong, T. G., and Mekalanos, J. J. (2014). A view to a kill: the bacterial type VI secretion system. Cell Host Microbe 15, 9–21. doi: 10.1016/j.chom.2013.11.008
Hodgkinson, J. T., Gross, J., Baker, Y. R., Spring, D. R., and Welch, M. (2016). A new Pseudomonas quinolone signal (PQS) binding partner: MexG. Chem. Sci. 7, 2553–2562. doi: 10.1039/c5sc04197j
Hogardt, M., Roeder, M., Schreff, A. M., Eberl, L., and Heesemann, J. (2004). Expression of Pseudomonas aeruginosa exoS is controlled by quorum sensing and RpoS. Microbiology 150(Pt 4), 843–851. doi: 10.1099/mic.0.26703-0
Holden, M. T., Seth-Smith, H. M., Crossman, L. C., Sebaihia, M., Bentley, S. D., Cerdeno-Tarraga, A. M., et al. (2009). The genome of Burkholderia cenocepacia J2315, an epidemic pathogen of cystic fibrosis patients. J. Bacteriol. 191, 261–277. doi: 10.1128/JB.01230-08
Ishikawa, T., Rompikuntal, P. K., Lindmark, B., Milton, D. L., and Wai, S. N. (2009). Quorum sensing regulation of the two hcp alleles in Vibrio cholerae O1 strains. PLoS One 4:e6734. doi: 10.1371/journal.pone.0006734
Izore, T., Job, V., and Dessen, A. (2011). Biogenesis, regulation, and targeting of the type III secretion system. Structure 19, 603–612. doi: 10.1016/j.str.2011.03.015
Jaskólska, M., Stutzmann, S., Stoudmann, C., and Blokesch, M. (2018). QstR-dependent regulation of natural competence and type VI secretion in Vibrio cholerae. Nucleic Acids Res. 46, 10619–10634. doi: 10.1093/nar/gky717
Javaux, C., Joris, B., and De Witte, P. (2007). Functional characteristics of TauA binding protein from TauABC Escherichia coli system. Protein J. 26, 231–238. doi: 10.1007/s10930-006-9064-x
Jimenez, P. N., Koch, G., Thompson, J. A., Xavier, K. B., Cool, R. H., and Quax, W. J. (2012). The multiple signaling systems regulating virulence in Pseudomonas aeruginosa. Microbiol. Mol. Biol. Rev. 76, 46–65. doi: 10.1128/MMBR.05007-11
Johansen, T. B., Agdestein, A., Olsen, I., Nilsen, S. F., Holstad, G., and Djonne, B. (2009). Biofilm formation by Mycobacterium avium isolates originating from humans, swine and birds. BMC Microbiol. 9:159. doi: 10.1186/1471-2180-9-159
Johnson, T. L., Fong, J. C., Rule, C., Rogers, A., Yildiz, F. H., and Sandkvist, M. (2014). The Type II secretion system delivers matrix proteins for biofilm formation by Vibrio cholerae. J. Bacteriol. 196, 4245–4252. doi: 10.1128/JB.01944-14
Jung, S. A., Chapman, C. A., and Ng, W. L. (2015). Quadruple quorum-sensing inputs control Vibrio cholerae virulence and maintain system robustness. PLoS Pathog. 11:e1004837. doi: 10.1371/journal.ppat.1004837
Kanamaru, K., Tatsuno, I., Tobe, T., and Sasakawa, C. (2000). SdiA, an Escherichia coli homologue of quorum-sensing regulators, controls the expression of virulence factors in enterohaemorrhagic Escherichia coli O157:H7. Mol. Microbiol. 38, 805–816. doi: 10.1046/j.1365-2958.2000.02171.x
Kanonenberg, K., Schwarz, C. K., and Schmitt, L. (2013). Type I secretion systems - a story of appendices. Res. Microbiol. 164, 596–604. doi: 10.1016/j.resmic.2013.03.011
Kariminik, A., Baseri-Salehi, M., and Kheirkhah, B. (2017). Pseudomonas aeruginosa quorum sensing modulates immune responses: an updated review article. Immunol. Lett. 190, 1–6. doi: 10.1016/j.imlet.2017.07.002
Khajanchi, B. K., Sha, J., Kozlova, E. V., Erova, T. E., Suarez, G., Sierra, J. C., et al. (2009). N-acylhomoserine lactones involved in quorum sensing control the type VI secretion system, biofilm formation, protease production, and in vivo virulence in a clinical isolate of Aeromonas hydrophila. Microbiology 155(Pt 11), 3518–3531. doi: 10.1099/mic.0.031575-0
Kida, Y., Higashimoto, Y., Inoue, H., Shimizu, T., and Kuwano, K. (2008). A novel secreted protease from Pseudomonas aeruginosa activates NF-kappaB through protease-activated receptors. Cell Microbiol. 10, 1491–1504. doi: 10.1111/j.1462-5822.2008.01142.x
Kita, D., Shibata, S., Kikuchi, Y., Kokubu, E., Nakayama, K., Saito, A., et al. (2016). Involvement of the type IX secretion system in Capnocytophaga ochracea gliding motility and biofilm formation. Appl. Environ. Microbiol. 82, 1756–1766. doi: 10.1128/AEM.03452-15
Kitaoka, M., Miyata, S. T., Brooks, T. M., Unterweger, D., and Pukatzki, S. (2011). VasH is a transcriptional regulator of the type VI secretion system functional in endemic and pandemic Vibrio cholerae. J. Bacteriol. 193, 6471–6482. doi: 10.1128/JB.05414-11
Kong, W., Liang, H., Shen, L., and Duan, K. (2009). [Regulation of type III secretion system by Rhl and PQS quorum sensing systems in Pseudomonas aeruginosa]. Wei Sheng Wu Xue Bao 49, 1158–1164.
Korotkov, K. V., Sandkvist, M., and Hol, W. G. (2012). The type II secretion system: biogenesis, molecular architecture and mechanism. Nat. Rev. Microbiol. 10, 336–351. doi: 10.1038/nrmicro2762
Korshunov, S., and Imlay, J. A. (2010). Two sources of endogenous hydrogen peroxide in Escherichia coli. Mol. Microbiol. 75, 1389–1401. doi: 10.1111/j.1365-2958.2010.07059.x
Kulka, K., Hatfull, G., and Ojha, A. K. (2012). Growth of Mycobacterium tuberculosis biofilms. J. Vis. Exp. 60:3820. doi: 10.3791/3820
Lai, L. Y., Lin, T. L., Chen, Y. Y., Hsieh, P. F., and Wang, J. T. (2018). Role of the Mycobacterium marinum ESX-1 secretion system in sliding motility and biofilm formation. Front. Microbiol. 9:1160. doi: 10.3389/fmicb.2018.01160
Lara-Tejero, M., and Galan, J. E. (2019). The injectisome, a complex nanomachine for protein injection into mammalian cells. Ecosal Plus 8. doi: 10.1128/ecosalplus.ESP-0039-2018
Lasica, A. M., Ksiazek, M., Madej, M., and Potempa, J. (2017). The type IX secretion system (T9SS): highlights and recent insights into its structure and function. Front. Cell. Infect. Microbiol. 7:215. doi: 10.3389/fcimb.2017.00215
Lee, J., Bansal, T., Jayaraman, A., Bentley, W. E., and Wood, T. K. (2007). Enterohemorrhagic Escherichia coli biofilms are inhibited by 7-hydroxyindole and stimulated by isatin. Appl. Environ. Microbiol. 73, 4100–4109. doi: 10.1128/AEM.00360-07
Lee, J., Zhang, X-S., Hegde, M., Bentley, W. E., Jayaraman, A., and Wood, T. K. (2008). Indole cell signaling occurs primarily at low temperatures in Escherichia coli. ISME J. 2, 1007–1023. doi: 10.1038/ismej.2008.54
Lee, J. H., and Lee, J. (2010). Indole as an intercellular signal in microbial communities. FEMS Microbiol. Rev. 34, 426–444. doi: 10.1111/j.1574-6976.2009.00204.x
Lee, S. W., Han, S. W., Bartley, L. E., and Ronald, P. C. (2006). From the academy: colloquium review. Unique characteristics of Xanthomonas oryzae pv. oryzae AvrXa21 and implications for plant innate immunity. Proc. Natl. Acad. Sci. U.S.A. 103, 18395–18400. doi: 10.1073/pnas.0605508103
Leiman, P. G., Basler, M., Ramagopal, U. A., Bonanno, J. B., Sauder, J. M., Pukatzki, S., et al. (2009). Type VI secretion apparatus and phage tail-associated protein complexes share a common evolutionary origin. Proc. Natl. Acad. Sci. U.S.A. 106, 4154–4159. doi: 10.1073/pnas.0813360106
Lenz, D. H., Mok, K. C., Lilley, B. N., Kulkarni, R. V., Wingreen, N. S., and Bassler, B. L. (2004). The small RNA chaperone Hfq and multiple small RNAs control quorum sensing in Vibrio harveyi and Vibrio cholerae. Cell 118, 69–82. doi: 10.1016/j.cell.2004.06.009
Leo, J. C., Grin, I., and Linke, D. (2012). Type V secretion: mechanism(s) of autotransport through the bacterial outer membrane. Philos. Trans. R. Soc. Lond. B Biol. Sci. 367, 1088–1101. doi: 10.1098/rstb.2011.0208
Lesic, B., Starkey, M., He, J., Hazan, R., and Rahme, L. G. (2009). Quorum sensing differentially regulates Pseudomonas aeruginosa type VI secretion locus I and homologous loci II and III, which are required for pathogenesis. Microbiology 155(Pt 9), 2845–2855. doi: 10.1099/mic.0.029082-0
Leung, K. Y., Siame, B. A., Snowball, H., and Mok, Y. K. (2011). Type VI secretion regulation: crosstalk and intracellular communication. Curr. Opin. Microbiol. 14, 9–15. doi: 10.1016/j.mib.2010.09.017
Li, P., Tian, M., Bao, Y., Hu, H., Liu, J., Yin, Y., et al. (2017). Brucella rough mutant induce macrophage death via activating IRE1alpha pathway of endoplasmic reticulum stress by enhanced T4SS secretion. Front. Cell. Infect. Microbiol. 7:422. doi: 10.3389/fcimb.2017.00422
Liang, H., Deng, X., Li, X., Ye, Y., and Wu, M. (2014). Molecular mechanisms of master regulator VqsM mediating quorum-sensing and antibiotic resistance in Pseudomonas aeruginosa. Nucleic Acids Res. 42, 10307–10320. doi: 10.1093/nar/gku586
Liang, W., Pascual-Montano, A., Silva, A. J., and Benitez, J. A. (2007). The cyclic AMP receptor protein modulates quorum sensing, motility and multiple genes that affect intestinal colonization in Vibrio cholerae. Microbiology 153(Pt 9), 2964–2975. doi: 10.1099/mic.0.2007/006668-0
Lin, J., Zhang, W., Cheng, J., Yang, X., Zhu, K., Wang, Y., et al. (2017). A Pseudomonas T6SS effector recruits PQS-containing outer membrane vesicles for iron acquisition. Nat. Commun. 8:14888. doi: 10.1038/ncomms14888
Liu, H., Wang, Q., Liu, Q., Cao, X., Shi, C., and Zhang, Y. (2011). Roles of Hfq in the stress adaptation and virulence in fish pathogen Vibrio alginolyticus and its potential application as a target for live attenuated vaccine. Appl. Microbiol. Biotechnol. 91, 353–364. doi: 10.1007/s00253-011-3286-3
Lo Scrudato, M., and Blokesch, M. (2012). The regulatory network of natural competence and transformation of Vibrio cholerae. PLoS Genet. 8:e1002778. doi: 10.1371/journal.pgen.1002778
Lo Scrudato, M., and Blokesch, M. (2013). A transcriptional regulator linking quorum sensing and chitin induction to render Vibrio cholerae naturally transformable. Nucleic Acids Res. 41, 3644–3658. doi: 10.1093/nar/gkt041
Lopez, M., Blasco, L., Gato, E., Perez, A., Fernandez-Garcia, L., Martinez-Martinez, L., et al. (2017a). Response to bile salts in clinical strains of Acinetobacter baumannii lacking the AdeABC efflux pump: virulence associated with quorum sensing. Front. Cell. Infect. Microbiol. 7:143. doi: 10.3389/fcimb.2017.00143
Lopez, M., Mayer, C., Fernandez-Garcia, L., Blasco, L., Muras, A., Ruiz, F. M., et al. (2017b). Quorum sensing network in clinical strains of A. baumannii: aidA is a new quorum quenching enzyme. PLoS One 12:e0174454. doi: 10.1371/journal.pone.0174454
Lopez, M. S., Tan, I. S., Yan, D., Kang, J., McCreary, M., Modrusan, Z., et al. (2017c). Host-derived fatty acids activate type VII secretion in Staphylococcus aureus. Proc. Natl. Acad. Sci. U.S.A. 114, 11223–11228. doi: 10.1073/pnas.1700627114
Lopez, M., Rueda, A., Florido, J. P., Blasco, L., Fernandez-Garcia, L., Trastoy, R., et al. (2018). Evolution of the Quorum network and the mobilome (plasmids and bacteriophages) in clinical strains of Acinetobacter baumannii during a decade. Sci. Rep. 8:2523. doi: 10.1038/s41598-018-20847-7
Majerczyk, C., Schneider, E., and Greenberg, E. P. (2016). Quorum sensing control of Type VI secretion factors restricts the proliferation of quorum-sensing mutants. eLife 5:e14712. doi: 10.7554/eLife.14712
Marsollier, L., Brodin, P., Jackson, M., Kordulakova, J., Tafelmeyer, P., Carbonnelle, E., et al. (2007). Impact of Mycobacterium ulcerans biofilm on transmissibility to ecological niches and Buruli ulcer pathogenesis. PLoS Pathog. 3:e62. doi: 10.1371/journal.ppat.0030062
Matz, C., Moreno, A. M., Alhede, M., Manefield, M., Hauser, A. R., Givskov, M., et al. (2008). Pseudomonas aeruginosa uses type III secretion system to kill biofilm-associated amoebae. ISME J. 2, 843–852. doi: 10.1038/ismej.2008.47
Maura, D., Hazan, R., Kitao, T., Ballok, A. E., and Rahme, L. G. (2016). Evidence for direct control of virulence and defense gene circuits by the Pseudomonas aeruginosa quorum sensing regulator, MvfR. Sci. Rep. 6:34083. doi: 10.1038/srep34083
McBride, M. J., and Zhu, Y. (2013). Gliding motility and Por secretion system genes are widespread among members of the phylum bacteroidetes. J. Bacteriol. 195, 270–278. doi: 10.1128/JB.01962-12
Michel, G. P., Aguzzi, A., Ball, G., Soscia, C., Bleves, S., and Voulhoux, R. (2011). Role of fimV in type II secretion system-dependent protein secretion of Pseudomonas aeruginosa on solid medium. Microbiology 157(Pt 7), 1945–1954. doi: 10.1099/mic.0.045849-0
Michel, G. P., Durand, E., and Filloux, A. (2007). XphA/XqhA, a novel GspCD subunit for type II secretion in Pseudomonas aeruginosa. J. Bacteriol. 189, 3776–3783. doi: 10.1128/JB.00205-07
Miller, M. B., Skorupski, K., Lenz, D. H., Taylor, R. K., and Bassler, B. L. (2002). Parallel quorum sensing systems converge to regulate virulence in Vibrio cholerae. Cell 110, 303–314. doi: 10.1016/S0092-8674(02)00829-2
Milton, D. L. (2006). Quorum sensing in vibrios: complexity for diversification. Int. J. Med. Microbiol. 296, 61–71. doi: 10.1016/j.ijmm.2006.01.044
Nagai, H., and Kubori, T. (2011). Type IVB secretion systems of Legionella and other gram-negative bacteria. Front. Microbiol. 2:136. doi: 10.3389/fmicb.2011.00136
Nealson, K. H., Platt, T., and Hastings, J. W. (1970). Cellular control of the synthesis and activity of the bacterial luminescent system. J. Bacteriol. 104, 313–322.
Ng, W. L., and Bassler, B. L. (2009). Bacterial quorum-sensing network architectures. Annu. Rev. Genet. 43, 197–222. doi: 10.1146/annurev-genet-102108-134304
Nivaskumar, M., and Francetic, O. (2014). Type II secretion system: a magic beanstalk or a protein escalator. Biochim. Biophys. Acta 1843, 1568–1577. doi: 10.1016/j.bbamcr.2013.12.020
Notti, R. Q., and Stebbins, C. E. (2016). The structure and function of type III secretion systems. Microbiol. Spectr. 4. doi: 10.1128/microbiolspec.VMBF-0004-2015
Novick, R. P., and Geisinger, E. (2008). Quorum sensing in staphylococci. Annu. Rev. Genet. 42, 541–564. doi: 10.1146/annurev.genet.42.110807.091640
Ojha, A., Anand, M., Bhatt, A., Kremer, L., Jacobs, W. R. Jr., and Hatfull, G. F. (2005). GroEL1: a dedicated chaperone involved in mycolic acid biosynthesis during biofilm formation in mycobacteria. Cell 123, 861–873. doi: 10.1016/j.cell.2005.09.012
Overbye, L. J., Sandkvist, M., and Bagdasarian, M. (1993). Genes required for extracellular secretion of enterotoxin are clustered in Vibrio cholerae. Gene 132, 101–106. doi: 10.1016/0378-1119(93)90520-D
Patel, R., Smith, S. M., and Robinson, C. (2014). Protein transport by the bacterial Tat pathway. Biochim. Biophys. Acta 1843, 1620–1628. doi: 10.1016/j.bbamcr.2014.02.013
Patzelt, D., Michael, V., Pauker, O., Ebert, M., Tielen, P., Jahn, D., et al. (2016). Gene flow across genus barriers - conjugation of Dinoroseobacter shibae’s 191-kb killer plasmid into Phaeobacter inhibens and AHL-mediated expression of type IV secretion systems. Front. Microbiol. 7:742. doi: 10.3389/fmicb.2016.00742
Patzelt, D., Wang, H., Buchholz, I., Rohde, M., Grobe, L., Pradella, S., et al. (2013). You are what you talk: quorum sensing induces individual morphologies and cell division modes in Dinoroseobacter shibae. ISME J. 7, 2274–2286. doi: 10.1038/ismej.2013.107
Paul, D., Gopal, J., Kumar, M., and Manikandan, M. (2018). Nature to the natural rescue: silencing microbial chats. Chem. Biol. Interact. 280, 86–98. doi: 10.1016/j.cbi.2017.12.018
Pesci, E. C., Milbank, J. B., Pearson, J. P., McKnight, S., Kende, A. S., Greenberg, E. P., et al. (1999). Quinolone signaling in the cell-to-cell communication system of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. U.S.A. 96, 11229–11234. doi: 10.1073/pnas.96.20.11229
Pukatzki, S., Ma, A. T., Revel, A. T., Sturtevant, D., and Mekalanos, J. J. (2007). Type VI secretion system translocates a phage tail spike-like protein into target cells where it cross-links actin. Proc. Natl. Acad. Sci. U.S.A. 104, 15508–15513. doi: 10.1073/pnas.0706532104
Qian, G., Zhou, Y., Zhao, Y., Song, Z., Wang, S., Fan, J., et al. (2013). Proteomic analysis reveals novel extracellular virulence-associated proteins and functions regulated by the diffusible signal factor (DSF) in Xanthomonas oryzae pv. oryzicola. J. Proteome Res. 12, 3327–3341. doi: 10.1021/pr4001543
Rai, N., Rai, R., and Kareenhalli, V. (2015). “Quorum sensing in competence and sporulation”, in Quorum Sensing vs Quorum Qenching: A Battle with No End in Sight, ed. K. Vipin Chandra (New Delhi: Springer), 61–64.
Rhoades, E. R., Archambault, A. S., Greendyke, R., Hsu, F. F., Streeter, C., and Byrd, T. F. (2009). Mycobacterium abscessus Glycopeptidolipids mask underlying cell wall phosphatidyl-myo-inositol mannosides blocking induction of human macrophage TNF-alpha by preventing interaction with TLR2. J. Immunol. 183, 1997–2007. doi: 10.4049/jimmunol.0802181
Riedel, K., Ohnesorg, T., Krogfelt, K. A., Hansen, T. S., Omori, K., Givskov, M., et al. (2001). N-acyl-L-homoserine lactone-mediated regulation of the lip secretion system in Serratia liquefaciens MG1. J. Bacteriol. 183, 1805–1809. doi: 10.1128/JB.183.5.1805-1809.2001
Robert, V., Filloux, A., and Michel, G. P. (2005). Subcomplexes from the Xcp secretion system of Pseudomonas aeruginosa. FEMS Microbiol. Lett. 252, 43–50. doi: 10.1016/j.femsle.2005.08.029
Rojas-Lopez, M., Zorgani, M. A., Kelley, L. A., Bailly, X., Kajava, A. V., Henderson, I. R., et al. (2017). Identification of the autochaperone domain in the type va secretion system (T5aSS): prevalent feature of autotransporters with a beta-helical passenger. Front. Microbiol. 8:2607. doi: 10.3389/fmicb.2017.02607
Ruer, S., Ball, G., Filloux, A., and de Bentzmann, S. (2008). The ‘P-usher’, a novel protein transporter involved in fimbrial assembly and TpsA secretion. EMBO J. 27, 2669–2680. doi: 10.1038/emboj.2008.197
Russell, A. B., Hood, R. D., Bui, N. K., LeRoux, M., Vollmer, W., and Mougous, J. D. (2011). Type VI secretion delivers bacteriolytic effectors to target cells. Nature 475, 343–347. doi: 10.1038/nature10244
Russell, A. B., Peterson, S. B., and Mougous, J. D. (2014). Type VI secretion system effectors: poisons with a purpose. Nat. Rev. Microbiol. 12, 137–148. doi: 10.1038/nrmicro3185
Ruwandeepika, H. A., Karunasagar, I., Bossier, P., and Defoirdt, T. (2015). Expression and quorum sensing regulation of type III secretion system genes of Vibrio harveyi during infection of gnotobiotic brine shrimp. PLoS One 10:e0143935. doi: 10.1371/journal.pone.0143935
Ryan, R. P., Monchy, S., Cardinale, M., Taghavi, S., Crossman, L., Avison, M. B., et al. (2009). The versatility and adaptation of bacteria from the genus Stenotrophomonas. Nat. Rev. Microbiol. 7, 514–525. doi: 10.1038/nrmicro2163
Sana, T. G., Hachani, A., Bucior, I., Soscia, C., Garvis, S., Termine, E., et al. (2012). The second type VI secretion system of Pseudomonas aeruginosa strain PAO1 is regulated by quorum sensing and Fur and modulates internalization in epithelial cells. J. Biol. Chem. 287, 27095–27105. doi: 10.1074/jbc.M112.376368
Sato, K., Naito, M., Yukitake, H., Hirakawa, H., Shoji, M., McBride, M. J., et al. (2010). A protein secretion system linked to bacteroidete gliding motility and pathogenesis. Proc. Natl. Acad. Sci. U.S.A. 107, 276–281. doi: 10.1073/pnas.0912010107
Schauder, S., Shokat, K., Surette, M. G., and Bassler, B. L. (2001). The LuxS family of bacterial autoinducers: biosynthesis of a novel quorum-sensing signal molecule. Mol. Microbiol. 41, 463–476. doi: 10.1046/j.1365-2958.2001.02532.x
Schiano, C. A., Koo, J. T., Schipma, M. J., Caulfield, A. J., Jafari, N., and Lathem, W. W. (2014). Genome-wide analysis of small RNAs expressed by Yersinia pestis identifies a regulator of the Yop-Ysc type III secretion system. J. Bacteriol. 196, 1659–1670. doi: 10.1128/JB.01456-13
Schulthess, B., Bloes, D. A., and Berger-Bachi, B. (2012). Opposing roles of sigmaB and sigmaB-controlled SpoVG in the global regulation of esxA in Staphylococcus aureus. BMC Microbiol. 12:17. doi: 10.1186/1471-2180-12-17
Schuster, M., Lostroh, C. P., Ogi, T., and Greenberg, E. P. (2003). Identification, timing, and signal specificity of Pseudomonas aeruginosa quorum-controlled genes: a transcriptome analysis. J. Bacteriol. 185, 2066–2079. doi: 10.1128/JB.185.7.2066-2079.2003
Seed, P. C., Passador, L., and Iglewski, B. H. (1995). Activation of the Pseudomonas aeruginosa lasI gene by LasR and the Pseudomonas autoinducer PAI: an autoinduction regulatory hierarchy. J. Bacteriol. 177, 654–659. doi: 10.1128/jb.177.3.654-659.1995
Shanker, E., and Federle, M. J. (2017). Quorum sensing regulation of competence and bacteriocins in Streptococcus pneumoniae and mutans. Genes 8:E15. doi: 10.3390/genes8010015
Shao, Y., and Bassler, B. L. (2014). Quorum regulatory small RNAs repress type VI secretion in Vibrio cholerae. Mol. Microbiol. 92, 921–930. doi: 10.1111/mmi.12599
Sharma, I. M., Petchiappan, A., and Chatterji, D. (2014). Quorum sensing and biofilm formation in mycobacteria: role of c-di-GMP and methods to study this second messenger. IUBMB Life 66, 823–834. doi: 10.1002/iub.1339
Sheng, L., Gu, D., Wang, Q., Liu, Q., and Zhang, Y. (2012). Quorum sensing and alternative sigma factor RpoN regulate type VI secretion system I (T6SSVA1) in fish pathogen Vibrio alginolyticus. Arch. Microbiol. 194, 379–390. doi: 10.1007/s00203-011-0780-z
Sheng, L., Lv, Y., Liu, Q., Wang, Q., and Zhang, Y. (2013). Connecting type VI secretion, quorum sensing, and c-di-GMP production in fish pathogen Vibrio alginolyticus through phosphatase PppA. Vet. Microbiol. 162, 652–662. doi: 10.1016/j.vetmic.2012.09.009
Sloup, R. E., Konal, A. E., Severin, G. B., Korir, M. L., Bagdasarian, M. M., Bagdasarian, M., et al. (2017). Cyclic Di-GMP and VpsR induce the expression of type II secretion in Vibrio cholerae. J. Bacteriol. 199:e00106-17. doi: 10.1128/JB.00106-17
Soto-Aceves, M. P., Cocotl-Yanez, M., Merino, E., Castillo-Juarez, I., Cortes-Lopez, H., Gonzalez-Pedrajo, B., et al. (2019). Inactivation of the quorum-sensing transcriptional regulators LasR or RhlR does not suppress the expression of virulence factors and the virulence of Pseudomonas aeruginosa PAO1. Microbiology 165, 425–432. doi: 10.1099/mic.0.000778
Sperandio, V., Mellies, J. L., Nguyen, W., Shin, S., and Kaper, J. B. (1999). Quorum sensing controls expression of the type III secretion gene transcription and protein secretion in enterohemorrhagic and enteropathogenic Escherichia coli. Proc. Natl. Acad. Sci. U.S.A. 96, 15196–15201. doi: 10.1073/pnas.96.26.15196
Stanley, S. A., Raghavan, S., Hwang, W. W., and Cox, J. S. (2003). Acute infection and macrophage subversion by Mycobacterium tuberculosis require a specialized secretion system. Proc. Natl. Acad. Sci. U.S.A. 100, 13001–13006. doi: 10.1073/pnas.2235593100
Stintzi, A., Evans, K., Meyer, J. M., and Poole, K. (1998). Quorum-sensing and siderophore biosynthesis in Pseudomonas aeruginosa: lasR/lasI mutants exhibit reduced pyoverdine biosynthesis. FEMS Microbiol. Lett. 166, 341–345. doi: 10.1111/j.1574-6968.1998.tb13910.x
Surette, M. G., Miller, M. B., and Bassler, B. L. (1999). Quorum sensing in Escherichia coli, Salmonella typhimurium, and Vibrio harveyi: a new family of genes responsible for autoinducer production. Proc. Natl. Acad. Sci. U.S.A. 96, 1639–1644. doi: 10.1073/pnas.96.4.1639
Teschler, J. K., Zamorano-Sanchez, D., Utada, A. S., Warner, C. J., Wong, G. C., Linington, R. G., et al. (2015). Living in the matrix: assembly and control of Vibrio cholerae biofilms. Nat. Rev. Microbiol. 13, 255–268. doi: 10.1038/nrmicro3433
Thomas, S., Holland, I. B., and Schmitt, L. (2014). The Type 1 secretion pathway -the hemolysin system and beyond. Biochim. Biophys. Acta 1843, 1629–1641. doi: 10.1016/j.bbamcr.2013.09.017
Trastoy, R., Manso, T., Fernandez-Garcia, L., Blasco, L., Ambroa, A., Perez Del Molino, M. L., et al. (2018). Mechanisms of bacterial tolerance and persistence in the gastrointestinal and respiratory environments. Clin. Microbiol. Rev. 31:e00023-18. doi: 10.1128/CMR.00023-18
Tseng, T.-T., Tyler, B. M., and Setubal, J. C. (2009). Protein secretion systems in bacterial-host associations, and their description in the Gene Ontology. BMC Microbiol. 9:S2. doi: 10.1186/1471-2180-9-S1-S2
Tsirigotaki, A., De Geyter, J., Sostaric, N., Economou, A., and Karamanou, S. (2017). Protein export through the bacterial Sec pathway. Nat. Rev. Microbiol. 15, 21–36. doi: 10.1038/nrmicro.2016.161
Tsou, A. M., Cai, T., Liu, Z., Zhu, J., and Kulkarni, R. V. (2009). Regulatory targets of quorum sensing in Vibrio cholerae: evidence for two distinct HapR-binding motifs. Nucleic Acids Res. 37, 2747–2756. doi: 10.1093/nar/gkp121
Van den Bergh, B., Fauvart, M., and Michiels, J. (2017). Formation, physiology, ecology, evolution and clinical importance of bacterial persisters. FEMS Microbiol. Rev. 41, 219–251. doi: 10.1093/femsre/fux001
Vilches, S., Jimenez, N., Tomas, J. M., and Merino, S. (2009). Aeromonas hydrophila AH-3 type III secretion system expression and regulatory network. Appl. Environ. Microbiol. 75, 6382–6392. doi: 10.1128/AEM.00222-09
Virmani, R., Hasija, Y., and Singh, Y. (2018). Effect of homocysteine on biofilm formation by mycobacteria. Indian J. Microbiol. 58, 287–293. doi: 10.1007/s12088-018-0739-8
Wagner, V. E., Bushnell, D., Passador, L., Brooks, A. I., and Iglewski, B. H. (2003). Microarray analysis of Pseudomonas aeruginosa quorum-sensing regulons: effects of growth phase and environment. J. Bacteriol. 185, 2080–2095. doi: 10.1128/JB.185.7.2080-2095.2003
Waters, C. M., and Bassler, B. L. (2005). Quorum sensing: cell-to-cell communication in bacteria. Annu. Rev. Cell Dev. Biol. 21, 319–346. doi: 10.1146/annurev.cellbio.21.012704.131001
Waters, C. M., Wu, J. T., Ramsey, M. E., Harris, R. C., and Bassler, B. L. (2010). Control of the type 3 secretion system in Vibrio harveyi by quorum sensing through repression of ExsA. Appl. Environ. Microbiol. 76, 4996–5004. doi: 10.1128/AEM.00886-10
Watve, S. S., Thomas, J., and Hammer, B. K. (2015). CytR is a global positive regulator of competence, type VI secretion, and chitinases in Vibrio cholerae. PLoS One 10:e0138834. doi: 10.1371/journal.pone.0138834
Whiteley, M., Diggle, S. P., and Greenberg, E. P. (2017). Progress in and promise of bacterial quorum sensing research. Nature 551, 313–320. doi: 10.1038/nature24624
Wilhelm, S., Gdynia, A., Tielen, P., Rosenau, F., and Jaeger, K. E. (2007). The autotransporter esterase EstA of Pseudomonas aeruginosa is required for rhamnolipid production, cell motility, and biofilm formation. J. Bacteriol. 189, 6695–6703. doi: 10.1128/JB.00023-07
Wood, T. K., Knabelm, S. J., and Kwan, B. W. (2013). Bacterial persister cell formation and dormancy. Appl. Environ. Microbiol. 79, 7116–7121. doi: 10.1128/AEM.02636-13
Xiao, G., Déziel, E., He, J., Lépine, F., Lesic, B., Castonguay, M. H., et al. (2006). MvfR, a key Pseudomonas aeruginosa pathogenicity LTTR-class regulatory protein, has dual ligands. Mol. Microbiol. 62, 1689–1699. doi: 10.1111/j.1365-2958.2006.05462.x
Yang, Z., Zhou, X., Ma, Y., Zhou, M., Waldor, M. K., Zhang, Y., et al. (2018). Serine/threonine kinase PpkA coordinates the interplay between T6SS2 activation and quorum sensing in the marine pathogen Vibrio alginolyticus. Environ. Microbiol. 20, 903–919. doi: 10.1111/1462-2920.14039
Zhang, L. H., and Dong, Y. H. (2004). Quorum sensing and signal interference: diverse implications. Mol. Microbiol. 53, 1563–1571. doi: 10.1111/j.1365-2958.2004.04234.x
Zhang, Y., Sass, A., Van Acker, H., Wille, J., Verhasselt, B., Van Nieuwerburgh, F., et al. (2018). Coumarin reduces virulence and biofilm formation in Pseudomonas aeruginosa by affecting quorum sensing, type III secretion and C-di-GMP levels. Front. Microbiol. 9:1952. doi: 10.3389/fmicb.2018.01952
Zhao, X., and Drlica, K. (2014). Reactive oxygen species and the bacterial response to lethal stress. Curr. Opin. Microbiol. 21, 1-6. doi: 10.1016/j.mib.2014.06.008
Zheng, J., Shin, O. S., Cameron, D. E., and Mekalanos, J. J. (2010). Quorum sensing and a global regulator TsrA control expression of type VI secretion and virulence in Vibrio cholerae. Proc. Natl. Acad. Sci. U.S.A. 107, 21128–21133. doi: 10.1073/pnas.1014998107
Keywords: quorum, secretion, virulence, motility, competence
Citation: Pena RT, Blasco L, Ambroa A, González-Pedrajo B, Fernández-García L, López M, Bleriot I, Bou G, García-Contreras R, Wood TK and Tomás M (2019) Relationship Between Quorum Sensing and Secretion Systems. Front. Microbiol. 10:1100. doi: 10.3389/fmicb.2019.01100
Received: 20 February 2019; Accepted: 30 April 2019;
Published: 07 June 2019.
Edited by:
Tom Defoirdt, Ghent University, BelgiumReviewed by:
Romé Voulhoux, UMR7255 Laboratoire d’Ingénierie des Systèmes Macromoléculaires (LISM), FranceJin Zhou, Tsinghua University, China
Copyright © 2019 Pena, Blasco, Ambroa, González-Pedrajo, Fernández-García, López, Bleriot, Bou, García-Contreras, Wood and Tomás. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rodolfo García-Contreras, cmdhcmNAYnEudW5hbS5teA==; Maria Tomás, TUEuZGVsLk1hci5Ub21hcy5DYXJtb25hQHNlcmdhcy5lcw==
†These authors have contributed equally to this work