
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Microbiol. , 03 May 2019
Sec. Virology
Volume 10 - 2019 | https://doi.org/10.3389/fmicb.2019.00873
 María José Altamirano-Lagos1
María José Altamirano-Lagos1 Fabián E. Díaz1
Fabián E. Díaz1 Miguel Andrés Mansilla1
Miguel Andrés Mansilla1 Daniela Rivera-Pérez1
Daniela Rivera-Pérez1 Daniel Soto2
Daniel Soto2 Jodi L. McGill3
Jodi L. McGill3 Abel E. Vasquez2,4
Abel E. Vasquez2,4 Alexis M. Kalergis1,5*
Alexis M. Kalergis1,5*The human respiratory syncytial virus (hRSV) is the main etiologic agent of severe lower respiratory tract infections that affect young children throughout the world, associated with significant morbidity and mortality, becoming a serious public health problem globally. Up to date, no licensed vaccines are available to prevent severe hRSV-induced disease, and the generation of safe-effective vaccines has been a challenging task, requiring constant biomedical research aimed to overcome this ailment. Among the difficulties presented by the study of this pathogen, it arises the fact that there is no single animal model that resembles all aspects of the human pathology, which is due to the specificity that this pathogen has for the human host. Thus, for the study of hRSV, different animal models might be employed, depending on the goal of the study. Of all the existing models, the murine model has been the most frequent model of choice for biomedical studies worldwide and has been of great importance at contributing to the development and understanding of vaccines and therapies against hRSV. The most notable use of the murine model is that it is very useful as a first approach in the development of vaccines or therapies such as monoclonal antibodies, suggesting in this way the direction that research could have in other preclinical models that have higher maintenance costs and more complex requirements in its management. However, several additional different models for studying hRSV, such as other rodents, mustelids, ruminants, and non-human primates, have been explored, offering advantages over the murine model. In this review, we discuss the various applications of animal models to the study of hRSV-induced disease and the advantages and disadvantages of each model, highlighting the potential of each model to elucidate different features of the pathology caused by the hRSV infection.
Human respiratory syncytial virus (hRSV) is an enveloped, single-stranded, and negative-sense RNA virus belonging to order Mononegavirales, family Pneumoviridae, genus Orthopneumovirus, species Human orthopneumovirus (Hacking and Hull, 2002; Borchers et al., 2013; Afonso et al., 2016; Snoeck et al., 2018). This virus is a human pathogen that causes a major burden in public health, both in developing and in industrialized countries (Simoes, 2003; Zang et al., 2015; Kuhdari et al., 2018). Noteworthy, hRSV is the leading cause of acute respiratory infection in newborns and of severe lower tract respiratory disease (LTRD) in children, with an estimation of 33.8 million of RSV-associated acute LTRD episodes in children less than 5 years old in 2005 (Nair et al., 2010). Estimations indicate that this virus is responsible for up to 3.4 million of hospital admission due to severe acute LTRD (Nair et al., 2010) and constitutes the leading cause of acute bronchiolitis and subsequent hospital admissions in industrialized countries (Bush and Thomson, 2007). Importantly, this virus is an important cause of mortality in young children in developing countries. In 2015, it was estimated that 59,600 hospitalized infants younger than 5 years old have died from hRSV-related LTRD worldwide (Shi et al., 2017; Scheltema et al., 2018).
Several attempts to develop safe and protective vaccines for the high-risk groups have been ineffective, and currently, there is no licensed vaccine for this pathogen (Hurwitz, 2011). Therefore, there is an urgent need for the development of a hRSV vaccine. In addition, the efficacy of the single licensed therapeutic option remains controversial, raising interest in the development of alternative therapeutic approaches against this pathogen (Canziani et al., 2012; Ispas et al., 2015; Muñoz-Durango et al., 2018; Simon et al., 2018). Therefore, the implementation of functional animal models for studying this virus has emerged as a critical and indispensable aspect underlying the development of immunotherapies and vaccines against hRSV (Hurwitz, 2011). For this reason, the development of different animal models for studying several aspects of hRSV has been very important and is still a field where research is focused. Since no animal model reflects all aspects of this viral infection and disease (Taylor, 2017), several models have been used in the study of hRSV, ranging from rodents and small mammals to large animals and non-human primates. This results from high specificity of hRSV for the human host, lacking an animal reservoir in nature (Collins and Graham, 2008). This feature has greatly hindered the development of an exclusive animal model, and therefore, the choice of the more suitable animal model required for each researcher will depend strongly on the aspect of the infection that needs to be studied and the investigative hypothesis proposed (Jorquera et al., 2016). The most commonly used animals have been rodents, such as mice (Graham et al., 1988; Bueno et al., 2008) and cotton rats (Prince et al., 1978, 1983; Sawada and Nakayama, 2016); ruminants (Elvander, 1996; Woolums et al., 1999, 2004; Meyerholz et al., 2004; Derscheid and Ackermann, 2012; Ackermann, 2014); and non-human primates (Kakuk et al., 1993; Szentiks et al., 2009), but at the present, the diversification of animal models is a requirement for addressing the diverse problematics of this viral infection and the development of vaccines and treatments. For this reason, the objective of this article is to review the several animal models used and their applications and to discuss their pros and cons. Finally, and based on the current information, recommendations of use are made, besides highlighting the role of the use of the murine model as a first approximation in the recent preclinical studies of new vaccines and antiviral treatments.
Mainly with the objective of vaccine efficacy and safety testing, several species of non-human primates (NHPs) have been used as animal models for hRSV infection, including chimpanzees (Pan troglodytes) (Szentiks et al., 2009), African green monkeys (Chlorocebus sabaeus), macaques (Macaca spp.) (Simoes et al., 1999; De Swart et al., 2002; Patton et al., 2015), and owl monkeys (Aotus spp.) (Prince et al., 1979). This group of species shares several anatomic and physiologic similarities to humans, but its application as an animal model of infection is challenging due to economical, technical, and ethical issues. Only chimpanzees are fully permissive to hRSV infection (Belshe et al., 1977; Crowe et al., 1993; Teng et al., 2000) and develop almost exclusively upper respiratory tract disease (URTD) symptoms in experimental studies.
hRSV was isolated for the first time not from humans, but from a group of chimpanzees naturally infected by the virus, which displayed signs of URTD (Morris et al., 1956). The experimental infection on this species has shown varying results, but high viral replication has been reported with viral doses lower than 104 plaque-forming units (PFUs; Belshe et al., 1977). Viral replication measured in nasopharyngeal and tracheal samples indicated that peak viral loads reached were 1–2 order of magnitude higher than the initial inoculum, and persistence of virus was shown to last 6–10 days post-infection (DPI; Belshe et al., 1977; Crowe et al., 1993; Teng et al., 2000). Although infected chimpanzees show clear URTD signs (Blount et al., 1956), no low respiratory tract disease (LTRD) has been reported under experimental conditions; therefore, human clinical disease condition is not completely replicated in this species. However, naturally infected chimpanzees with fatal LTRD have been reported, including viral antigen presence in lungs and histopathological changes including high neutrophil infiltration, edema, and the deposition of hyaline membranes (Belshe et al., 1977; Szentiks et al., 2009; Unwin et al., 2013). Natural co-infections with Streptococcus pneumoniae have also been reported and may be enhancing those naturally occurring infections (Szentiks et al., 2009; Nguyen et al., 2015).
Applications of this NHP model have included the evaluation of protective efficacy and safety of live-attenuated and recombinant vaccine candidates (Richardson et al., 1978; Collins et al., 1990; Crowe et al., 1993; Teng et al., 2000). Similar body temperatures between chimpanzees and humans have allowed studies with candidate temperature-sensitive vaccines (Council, 2011). Vaccine candidates consisting of the recombinant expression of F and G glycoproteins by modified vaccinia viruses have shown to induce low levels of neutralizing antibodies (NAs) and protection to viral challenge in chimpanzees (Collins et al., 1990; Crowe et al., 1993). On the contrary, temperature-sensitive mutant hRSV vaccines that are highly restricted at replication but highly immunogenic in chimpanzees have induced protective responses against hRSV infection in this species, although protective mechanisms have not been defined in detail (Richardson et al., 1978; Crowe et al., 1995). Permissive replication of hRSV is the most remarkable advantage of this model, being the unique species reflecting the viral replication in humans (Belshe et al., 1977; Crowe et al., 1993). Anatomical, physiological, and genetic similarities also represent a unique advantage of the use of this model.
However, several disadvantages and limitations apply to this model, and the appearance of several other useful models has diminished the utilization of chimpanzees. The economical, logistical, and ethical cost of research on chimpanzees is extremely high, and therefore, this NHP is not commonly available to research groups (Tardif et al., 2013). Published studies to date mostly yielded inconclusive results due to small sample sizes and genetic heterogeneity of animals. Lack of inbred strains, immunological tools, and reagents are also an important restriction to the use of this model (Bem et al., 2011; Jorquera et al., 2016). Finally, specialized housing, handling, and caring of this species are mandatory (Tardif et al., 2013).
The National Research Council has established that this model may be necessary exclusively for testing the candidate vaccines, based on the suitability of this model to serve as surrogate model for seronegative infants (Council, 2011).
Permissiveness of species within Macaca spp. to hRSV infection is low, with high dose intranasal (IN) or intratracheal (IT) inocula reaching low or moderate viral replication capacity, despite the age of individuals (Simoes et al., 1999; De Swart et al., 2002; McArthur-Vaughan and Gershwin, 2002; Grunwald et al., 2014; Grandin et al., 2015). Although clinical disease is rare, a report has shown evidence for mild respiratory disease with fever and increased respiratory rate in juvenile rhesus macaques associated to bronchiolitis, bronchitis, and interstitial pneumonia (McArthur-Vaughan and Gershwin, 2002). The virus has been re-isolated from lungs but at low viral titers (Simoes et al., 1999; McArthur-Vaughan and Gershwin, 2002). Studies using the formalin-inactivated hRSV vaccine (FI-hRSV) have shown enhanced disease with cellular peribronchiolar, perivascular infiltration, and high viral titers, but that enhanced response might be affected by immune responses to non-viral antigens present in the viral challenge preparations (Ponnuraj et al., 2001; De Swart et al., 2002). A vaccine candidate based on the Fusion (F) protein of hRSV adjuvanted with GLA-SE was evaluated in cynomolgus macaques and induced a robust humoral and Th1-biased cellular immunity (Patton et al., 2015). Further, lack of protection and evidence of enhanced disease were observed in infant macaques after immunization with a modified vaccinia virus Ankara vector expressing the hRSV F and G proteins (de Waal et al., 2004). Also, a heterologous adenovirus vector vaccine with the F subunit of hRSV have been evaluated in rhesus macaques (Grunwald et al., 2014). A mucosal booster immunization showed to be effective at reducing viral replication, with an immune response that included a high and persistent expansion of CD4+ and CD8+ T cells in the lower respiratory tract mucosal sites. These data suggest that this vaccine scheme is effective for eliciting protective immune responses at mucosal effector sites in this animal species (Grunwald et al., 2014). The pros and cons for using this animal model are equivalent to those previously mentioned for chimpanzees.
African green monkeys (AGMs) are less susceptible to hRSV infection than chimpanzees (Kakuk et al., 1993), often lack clinical manifestations after hRSV infection, and develop only minor histopathological changes (Kakuk et al., 1993; Ispas et al., 2015). Conversely, the viral load constitutes a more reliable disease parameter in this animal model (Kakuk et al., 1993; Eyles et al., 2013; Ispas et al., 2015). Mild signs, including rhinorrhea, coughing, sneezing, and wheezing, have been described in AGMs following IN and IT administration of the hRSV A2 strain AGMs. Further, viral shedding in the oropharynx has been detected for 8 days in these animals (Kakuk et al., 1993). Studies on AGMs have focused on vaccine candidate testing, including recombinant vaccines (Kakuk et al., 1993; Tang et al., 2004; Jones et al., 2012; Wang et al., 2017) and adjuvanted subunit vaccines (Eyles et al., 2013). Vaccines that had shown efficacy in rodent challenge studies have failed to elicit appropriate responses in AGMs, which could be due to the immunological response itself or to the different experimental design relative to the vaccination and challenge (Taylor, 2017). These observations underscore the difficulty of translating results from rodent species to primates (Eyles et al., 2013). Additionally, antiviral effects of TMC353121, a fusion inhibitor, have been successfully tested in AGMs as a preclinical model (Ispas et al., 2015). The first primate model for FI-hRSV-induced enhanced respiratory disease was developed on this species, with airway and parenchymal inflammatory changes in the vaccinated animals, which could not be evidenced by viral titers (Kakuk et al., 1993). Although less expensive than chimpanzees, utilization of this species has most of the general disadvantages in which NHP models display, as those previously discussed.
Only a few studies have explored the use of this species as an animal model for hRSV infection (Prince et al., 1979; Koff et al., 1983; Hemming et al., 1995). Prince and collaborators have reported that an IN inoculation of 104 PFU of hRSV can result in high viral titers, rhinorrhea, and the development of low levels of neutralizing antibody (NA) virus titers appearing at 14 days post-infection and reaching a peak at 28 days post-infection. Viral infection was observed until 8 to 17 days (Prince et al., 1979; Koff et al., 1983). Antibody-dependent cellular cytotoxicity has been studied in owl monkeys against human epithelial HEp-2 cells infected with hRSV, with peripheral mononuclear blood cells in the presence of hRSV antibodies causing lysis of infected HEp-2 cells in an in vitro assay (Koff et al., 1983). Reduction of viral shedding from nose and trachea has been reported after intravenous administration of human immunoglobulin (IVIG) in owl monkeys that previously received an intratracheal (IT) hRSV challenge (Hemming et al., 1995). Response to vaccination in this species has been scarcely studied; a single dose of recombinant vaccinia virus expressing the F or G glycoprotein of hRSV has been reported to be effective in controlling hRSV-induced LRT (Olmsted et al., 1988).
The order Rodentia includes a variety of small mammals that have been widely used as models of human pathogens in immunological and infection studies, including vaccine testing studies, and elucidation of immunopathogenic mechanisms (Andersen and Winter, 2017). Despite differences at the infective cycle developed by hRSV infection, permissiveness for viral replication, and host immune system differences between mice and humans, the laboratory mouse model remains to date as one of the most prevalent animal models used in hRSV studies. However, the use of the cotton rat has allowed the modeling of several aspects on this infection that the mouse model fails to resemble, including viral replication permissiveness, which is improved in cotton rats. These two species are systematically used nowadays and have allowed researchers to make significant progress in vaccine and immunotherapy development. Further, other rodent models have also been utilized, as described later.
The laboratory mouse (Mus musculus) is the most frequently used animal as a model in the field of biomedical sciences and is the model of choice in immunological and infection research, as well as for the evaluation of vaccines and therapies worldwide (Lee et al., 2012; Ehret et al., 2017). Although the development of vaccines and new therapeutic approaches against hRSV has been a challenge for scientists to date (Salazar et al., 2017; Mazur et al., 2018), mouse models have been very important to achieve past and recent advances in preclinical tests of new vaccines and antiviral treatments. Further, these models have contributed significantly to elucidate their mechanisms of action (Hurwitz, 2011; Heylen et al., 2017). Noteworthy, the clinical symptomatology of the hRSV disease after mouse infection has certain differences when compared to human disease. Besides, clinical differences are also observed between different strains of mice (Openshaw and Tregoning, 2005). Another feature of the murine model for hRSV infection is that these animals must be challenged with large doses (~107 PFU) to detect viral replication in the lungs (Graham et al., 1988). Among mice strains available, the BALB/c mouse is commonly used to study the immunopathology caused by hRSV infection, since it is semi-permissive to lung viral replication. The disease manifests with piloerection, reduced activity, and weight loss (Graham et al., 1988; Kong et al., 2005). Furthermore, moderate bronchiolitis is observed in these mice (Bueno et al., 2008). Importantly, loss of body weight within the first 3 days of infection and infiltration of lungs with neutrophils are two well-established disease parameters to assess protection against this serious disease (Bueno et al., 2008).
It has been reported that cell-mediated immunity decreases as individuals get older. Therefore, studying CD8+ T-lymphocyte populations in different age groups is a relevant topic to address. (Fulton et al., 2013; Mosquera et al., 2014). The aged BALB/c mice generate a weak primary response of CD8+ T cells specific for hRSV in the lung, which is associated with a delay in viral clearance. When evaluating the maximum magnitudes of the responses of virus-specific CD8+ T cells in lung and airways, significant decreases have been seen in aged BALB/c mice when compared with young BALB/c mice (Fulton et al., 2013). Infected aged BALB/c mice develop severe inflammatory changes with a predominance of neutrophils and lymphocytes, in addition to diffuse alveolar damage 4 days post-infection (Mosquera et al., 2014). It has been shown that both the kinetics and the magnitude of antiviral gene expression decrease as a result of advanced age. In addition to the delay in cytokine signaling and the induction of the receptor pattern recognition, it has been found that TLR 7/8 signaling is altered in alveolar macrophages in elderly mice, and this shows that there are inherent differences in response to hRSV infection in pneumonia models of young BALB/c mice versus the model of aged BALB/c mice (Wong et al., 2014). Experimentally, it has been described that a senescence prone SAM-P1 mouse strain (H-2 K), which shares the genetic background of AKR/J mice, is more susceptible to infection by hRSV, and shows a deficient CD8+ T cell response, as well as a lower gamma-interferon (INF-γ) production, which contrasts to high interleukin (IL)-4 production (Liu and Kimura, 2007). Noteworthy, this animal model allows for the generation of humanized mice or mice with a human immune system (HIS mice; Sharma et al., 2016), which has been also used to evaluate vaccine efficacy (Sharma et al., 2016). During the first 3 days of infection, the HIS mice lost more weight and eliminated hRSV faster than did NOD-scid gamma mice (NSG mice). These mice were generated by the introduction of an adeno-associated virus serotype 9 (AAV9) vector carrying human cytokine genes into highly immunodeficient NOD-scid gamma (NSG). The pathological characteristics induced by hRSV infection in HIS mice include peribronchiolar inflammation, neutrophil predominance in the bronchoalveolar lavage fluid, and enhanced production of mucus in the respiratory tract (Sharma et al., 2016). In addition, the mouse model has been extensively used to perform preclinical studies for vaccine and therapies for this virus (Hurwitz, 2011). Recently, vaccine efficacy studies were performed in mice to evaluate safety and immunogenicity of a vaccine consisting of a recombinant strain of bacillus Calmette-Guérin (rBCG) expressing the nucleoprotein antigen of hRSV (Cautivo et al., 2010; Céspedes et al., 2017). A single immunization with the rBCG vaccine elicited protective Th1 type immunity against hRSV in mice, preventing weight loss due to infection, as well as reducing viral replication and disease severity in these animals (Céspedes et al., 2017). Vaccination with rBCG promotes the elimination of the virus at the pulmonary level and prevents development of interstitial pneumonia without evident adverse effects (Céspedes et al., 2017). Moreover, subunit vaccines against hRSV based on a novel genome replication-deficient Sendai virus (SeV) vector expressing the F protein as a genetically stable antigen have been studied (Wiegand et al., 2017). After IN or intramuscular (IM) immunization of BALB/c mice, a robust hRSV-specific immune response was induced, consisting of serum IgG and neutralizing antibodies, as well as cytotoxic T cells. Further, IN immunization was also able to stimulate hRSV-specific mucosal IgA in the upper and lower respiratory tract of these animals (Wiegand et al., 2017). The mouse is also a useful model for the study of basic aspects of mechanisms related to the pathogenesis of asthma as a consequence of hRSV infection (Peebles and Graham, 2005). It is well established that the mouse model has multiple comparative advantages in relation to other animals, such as genetic homogeneity between consanguineous animals; availability of humanized, knockout, and transgenic stains; very good reproductive performance; ease of maintenance in confined spaces; non-complex nutritional requirements; and wide variety of reagents to study the immune response. Additionally, the similarities between the mouse and human immunological response, as well as the docile nature of the laboratory mice and non-complex management, make this species one of the most used for studies relative to infections and immunity worldwide (Mestas and Hughes, 2004; Drake, 2013; Sharma et al., 2016).
The cotton rat (Sigmodon hispidus) is currently among the most extensively used animal models of hRSV and other human infectious respiratory diseases, in part due to the semi-permissive replication of several respiratory viral agents and the developing industry of genetic and immunological tools applicable on this model. Up to date, this model has been successfully used for several studies in hRSV, including antibody prophylaxis (Ottolini et al., 2002), vaccine testing (Widjojoatmodjo et al., 2010; Garlapati et al., 2012; Kamphuis et al., 2013; Rostad et al., 2016; Stobart et al., 2016; Widjaja et al., 2016; Fuentes et al., 2017), FI-RSV enhanced respiratory disease (Prince et al., 1986; Sawada and Nakayama, 2016; Widjaja et al., 2016), maternally induced immunity (Prince et al., 1983; Blanco et al., 2015), and susceptibility of high-risk human groups (Boukhvalova and Blanco, 2013). In fact, the main advantage of this model is its greater permissiveness to hRSV infection than inbred mice, other rodents, and animals in general, reaching a replication that is 50–1,000 fold higher than mouse strains (Prince et al., 1979). Infection with hRSV undergoes active replication in lungs and nasal tissue, lasting about 6–9 days, respectively. However, tracheal replication reaches only low titers (Prince et al., 1978). Histopathological examinations have revealed mild lesions as early as day 2, including bronchitis, bronchiolitis, and exudative rhinitis (Prince et al., 1978). Interestingly, with higher doses (106 PFU), cotton rats develop alveolitis, peribronchiolitis, and interstitial pneumonitis, therefore reflecting a direct relationship between viral replication, histopathology, and the infectious dose inoculated (Prince et al., 1978). Although the quality and timing of pathological changes observed in cotton rats after hRSV infection share several features with the natural human hRSV infection, bronchiolitis is notoriously less severe in cotton rats (Prince et al., 1978, 1986). The absence of clinical signs, such as cough, rhinorrhea, and fever, constitutes a disadvantage that should be considered for certain studies. Although cotton rats remain susceptible through the whole life, viral replication and persistence are major in infant animals. Reinfection of this species leads to an inflammatory response in lungs that is characterized by the absence of viral production due to an abortive viral replication (Prince et al., 1979; Boukhvalova et al., 2007a), which is paralleled by an early upregulation of interferon response and expression of interferon-inducible MX genes (Pletneva et al., 2008). The presence of MX genes is noteworthy, since Mx1 and Mx2 are functionally absent in inbred mouse strains C57BL/6, BALB/c, and CBA/J, and is an important component of the antiviral innate response (Staeheli et al., 1993). In terms of the immune response to experimental infection, the arachidonic acid pathway is upregulated after cotton rat infection with hRSV and has been associated to the severity of disease (Richardson et al., 2005). Cytokine synthesis in lung after hRSV infection reveals an increase in IFN-γ, IL-10, IL-6, CCL-2, and growth-regulated oncogene, reaching a peak at day 4 after infection in infant cotton rats, which is delayed to day 6 in aged individuals (Boukhvalova et al., 2007b).
Response to repetitive exposures to hRSV has been evaluated in cotton rats, showing that during the first infection there is an absence of neutralizing antibody production, which results in a lack of protection to a second infection. During a second infection, a neutralizing antibody response and a CD8+IFN-γ+ T cell response take place, which seem to be nevertheless unable to clear the virus from the lungs (Yamaji et al., 2016).
The study of maternal transferred immunity is one of the earliest applications for this model (Blanco et al., 2015). Both colostrum feeding and transplacental immunity can provide protection in lungs and nasal tissue in infant cotton rats gestated by immunized mothers challenged with hRSV prior to litter birth. Unfortunately, this passively transferred protection is transient and significantly reduced after 4 weeks from birth (Prince et al., 1983; Blanco et al., 2015). This protective effect is correlated to serum neutralizing antibodies, with titers >1:380 being protective in infant cotton rats, which is similar to human infant passively-acquired immunity, which results in reduced susceptibility to infection with antibody titers equal or higher to 1:400 (Prince et al., 1983, 1985). However, the lack of correlation in some studied animals suggests that other immune mediators might be contributing to maternally transmitted protective immunity (Prince et al., 1983). Interestingly, a strong correlation between hRSV-specific neutralizing antibody titers in cotton rat mothers and their litters has been described, and the protection provided by those antibodies appears to be inversely associated to cytokine expression in lungs (Blanco et al., 2015). Also, the maternally acquired humoral immunity is responsible for suppressing vaccine immunogenicity in infant cotton rats (Prince et al., 1979, 1982), thereby highlighting the difficulties of developing a vaccine for human infants.
The mentioned correlation between antibody levels as a way of providing protection in cotton rats and human infants led to several studies (Saez-Llorens et al., 1998; Johnson et al., 1999), and to the development of a commercial antibody formulation for prophylactic use in human infants in high risk of severe hRSV disease (Olchanski et al., 2018). The ability of predicting the success of immunoprophylaxis in infants and of assessing the dose of antibody required to elicit protective serum levels is one of the most remarkable milestones of this animal model (Prince et al., 1985; Group, 1998). It is important to mention that those studies advanced to clinical phases without the need of intermediate studies in NHPs and are currently available from commercial use in patients (Niewiesk and Prince, 2002). On the other hand, research on immunotherapeutic use of antibodies against hRSV has been conducted, but has yielded dissimilar results. Several antibodies tested have been useful for controlling viral replication but not for ameliorating lung pathology and disease severity (Rodriguez et al., 1997; Malley et al., 1998; Prince et al., 2000). More recently, a novel Ig formulation, RI-002, was successfully tested in cotton rats for treating immunocompromised patients. This formulation, containing high quantities of hRSV-specific neutralizing antibodies, inhibited the prolonged hRSV replication, which is typical in immunocompromised rats (Johnson et al., 1982) and reduced dissemination of replicative virus. Pulmonary interstitial inflammation and epithelial hyperplasia were also reduced by RI-002 administration, therefore providing evidence of a beneficial therapy for hRSV disease outcome in immunocompromised cotton rats (Boukhvalova et al., 2016) results that should be further investigated on its ability to reduce disease symptoms in humans.
Research on the pathology of vaccine-enhanced respiratory disease, described decades ago in human infants, also finds a suitable model in cotton rats. Knowledge on the mechanisms of this manifestation is critical for future studies on vaccine development. FI-hRSV vaccinated cotton rats subsequently challenged with hRSV present an increased pulmonary pathology, with severe alveolitis as a hallmark of histopathological changes, as well as a rise in neutrophil and lymphocyte infiltration (Prince et al., 1986). Additionally, association of Th2 polarization as a known mechanism of vaccine-enhanced respiratory disease has been studied in this model in which an upregulation of Th2 cytokines (IL-4, IL-10, IL-13, and CCL5) has been observed (Boukhvalova et al., 2006; Sawada and Nakayama, 2016). However, an increase in Th1 chemokines and cytokines has also been reported, and it has been suggested that those pro-inflammatory cytokines and chemokines may play a role in enhanced pulmonary inflammation (Prince et al., 2001; Boukhvalova et al., 2006). Immunodominance of post-fusion-specific site I of F protein elicits weak neutralizing antibody responses against that site in FI-RSV-immunized cotton rats and an absence of an efficient neutralizing response by other antibodies (Widjaja et al., 2016). This situation contrasts with immunization response to experimentally infected or vaccinated cotton rats, which showed a high virus neutralizing capacity from pre-fusion specific antibodies binding antigenic site Ø and other parts of the F protein, and may account for the mechanism on FI-hRSV-enhanced respiratory disease (Widjaja et al., 2016). Noteworthy, limitations of some studies performed in cotton rats have been revealed, including observations on the role of non-viral products in vaccine or challenge inoculated medium as drivers of alveolitis, which appears to be mediated chiefly by T cell specific responses to non-viral antigens, having the hRSV antigens a limited role as co-factors in the inflammation and disease manifestation (Shaw et al., 2013).
Despite limitations, cotton rats have been the main model in research on development of efficient and secure vaccine candidates, and several vaccine candidates have been recently tested in this model including subunit vaccines (Garlapati et al., 2012), recombinant vaccines (Widjojoatmodjo et al., 2010, 2015; Stobart et al., 2016; Fuentes et al., 2017), VLPs and virosomes with viral subunits (Kamphuis et al., 2013; Cullen et al., 2015), as well as live attenuated vaccines (Luongo et al., 2013; Stobart et al., 2016), therefore confirming the relevance of this animal model for those studies. However, prediction capacity of this model on protective responses after vaccination in humans is still unknown, and further investigations are required to elucidate this question.
Increased susceptibility to severe hRSV infection occurring in immunosuppressed infants and older people has been successfully modeled in the cotton rat model. Late clearance of virus is classical in immunosuppressed cotton rats, therefore impacting in disease pathogenesis (Ottolini et al., 1999). Also, this model has been used to test prophylactic and therapeutic schemes in immunosuppressed rats, reflecting the need of multiple administrations of Ig to diminish viral replication (Ottolini et al., 1999; Boukhvalova et al., 2016). Unlike mice, infant cotton rats permit a greater viral replication with a greater persistence in URT than 4-week-age rats and mount a less efficient NA response than adult animals (Prince et al., 1978). In elderly, immunosenescent cotton rats (>6-month age), prolonged viral persistence is present (Curtis et al., 2002), and peak expression of cytokines is delayed in older rats (>6 month age) than young age rats (<2-month age) (Boukhvalova et al., 2007b).
Technical advantages on the use of this model include the availability of commercial and laboratory-owned inbred strains, as well as the increasing quantity of immunological reagents and assays, which, added to similarities between cotton rats and human innate immune system (Boukhvalova and Blanco, 2013), make this species an interesting and useful model for pathogenesis and immune responses to infection. On the other hand, the lack of clinical manifestations after hRSV infection limits the use of this model in this field. Additionally, the natural progression of infection from the URT to the LRT taking place in humans is not observed in this species (Boukhvalova and Blanco, 2013). For laboratories looking forward to including this animal model, it must be noted that handling, maintenance, and breeding of this species are different from mice strains, demanding specific facilities and protocols. Handling might be difficult to inexperienced scientists and students, and these animals may bite if not held properly. Also, blood withdrawal and drug administration are different; the retro-orbital plexus is the best site for blood collection, which requires training of personal for ensuring animal welfare (Niewiesk and Prince, 2002).
Beyond inbred mice and cotton rats, several other rodents have been used for hRSV studies, including vaccine efficacy studies. However, to date, robust and consistent data from established research lines are lacking, and the application of these studies to vaccine development and testing is very limited. The use of these other rodent models in general and the scope of the findings are limited by the disadvantages of being not fully permissive for hRSV replication, the use of outbred animals, and the lack of immunological reagents and inbred animals. However, recent observations highlight the importance of expanding the animal model repertoire.
Experimental infections of hamsters have been reported in 3-week-age hamsters inoculated with high hRSV doses (104.6–106.5 PFU), which allow for viral replication in URT and LRT but lacking histopathological and clinical manifestations of lung compromise (Wright et al., 1970). Remarkably, Syrian hamsters (Mesocricetus auratus) have been used for the development and preclinical evaluation of live attenuated recombinant human parainfluenza virus 1 vectors expressing the hRSV F protein. This attenuated vaccine has provided protection in these rodents against an IN challenge on day 30 post immunization, with a dose of 106 PFU, protection that well correlated with the production of serum NAs against hRSV (Mackow et al., 2015).
Studies on Chinchillas (Chinchilla lanigera) have shown that this species is semi-permissive for hRSV infection, but only in the upper airways following high-dose intranasal challenge (1x106 – 1x107 PFU). It has been proposed as a suitable model for studies of hRSV infection in upper airways, including the role of hRSV infection in the development of otitis media, based on observations of viral replication in nasal cavities and Eustachian tubes (Gitiban et al., 2005; Grieves et al., 2010), as well as a model for the understanding of mucosal immunity responses (McGillivary et al., 2013); however, this model has not been employed enough to draw robust conclusions of its application.
Guinea pig (Cavia porcellus) is another rodent that is semi-permissible for hRSV infection, as observed in a single study to date. Acute bronchiolitis, including bronchiolar epithelial necrosis, and either mononuclear and polymorphonuclear (PMN) lymphocyte infiltrates, have been observed at 6 days post-infection, with a remission at 14 days post-infection, in one-month age outbred guinea pigs, intranasally infected with 4 × 103 PFU. This was accompanied by the absence of clinical signs. The histopathological lesions at 6 days post-infection were paralleled with low viral titers, indicative of limited viral replication (Hegele et al., 1993). Although these observations support the use of this model for the understanding of acute hRSV infections, it has not been extensively studied, and the very limited availability of inbred strains and immunological reagents restricts its use. Noteworthy, an advantage on the use of this model is that guinea pig is more similar to humans than other small animal models in terms of physiology and immune system responses, which has been observed in several studies reviewed elsewhere (Padilla-Carlin et al., 2008).
Ferrets (Mustela putorius furo) are small carnivores that belong to the Mustelidae family, which have been used for the study of different infectious diseases, including those hRSV infection. Their small size, and similarities shared with humans at an anatomical and physiopathological level are key advantages of the use of this species for investigation of infectious diseases.
In 1976, it was shown that ferrets allow for viral replication in nasal tissue of all age animals (Prince and Porter, 1976), but replication in lung tissue was observed only in infant ferrets; therefore, it was proposed as an interesting model for the age dependence studies on severe hRSV infection (Byrd and Prince, 1997). However, a recent study further highlighted the advantages of ferret models, showing that adult ferrets are highly susceptible to hRSV infection, since IT infection of immunocompetent individuals resulted in productive virus replication in the upper and lower respiratory tract, with the presence of viral antigens in tracheal and bronchial epithelial cells, reaching viral loads in throat, trachea, and lungs with similar order of magnitude in comparison to cotton rats (Stittelaar et al., 2016). This seems to be a major advantage of this model, along with the ease of handling the animals. Interestingly, recent viral interference studies have been carried out in ferrets to elucidate mechanisms that explain separate seasonal peak incidence of influenza and hRSV, showing that replication of a pandemic influenza virus strain (A[H1N1]pdm09) prevents subsequent infection with laboratory hRSV strains by antigen-independent mechanisms (Chan et al., 2018). Besides, studies on immunocompromised ferrets have shown hRSV replication in bronchiolar epithelial cells and reduced/delayed viral clearance (de Waal et al., 2018). This would allow the development of a specific animal model for a certain population at risk, such as immunocompromised patients, who experience severe pulmonary manifestations after hRSV infection (de Waal et al., 2018). Remarkably, viral transmission studies in immunocompetent or immunocompromised animals have been recently carried out, which suggest the suitability of ferrets in studying interventions aimed to limit hRSV transmission (Chan et al., 2017; de Waal et al., 2018).
Some disadvantages regarding the use of this model include the lack of clinical manifestations, the requirement of a more specialized husbandry facility and caging system, and the limited availability of immunological reagents and genetically modified mutants for immunological investigation. Also, the limited development of inbred ferrets restricts the application for mechanistic studies (Enkirch and Von Messling, 2015; Stittelaar et al., 2016).
Sheep (Ovis aries) are susceptible to natural infection by ovine respiratory syncytial virus and bovine respiratory syncytial virus (bRSV; Masot et al., 1995, 1996). The last mentioned is responsible for severe infection in preterm lambs, with an age-dependent clearance of the virus, being this preterm lamb model a useful model of severe hRSV disease in the preterm infants (Meyerholz et al., 2004), and also in exploring disease mechanisms, therapeutic regimens, and risk factors (Derscheid and Ackermann, 2012). Lambs are also susceptible to experimental infection by high doses of hRSV, which results in URTD and LTRD (Olivier et al., 2009; Sow et al., 2011; Larios Mora et al., 2015). Intratracheal inoculation of lambs with 108 PFU of the A2 strain induces a pulmonary pathology that shares similarities to pathology observed after human infant hRSV infection, including bronchiolitis with neutrophil infiltration, mild peribronchiolar interstitial neumonia. A peak of viral replication in airway epithelium occurs at 6 days post-infection, with virus clearance at 14 days post-infection (Olivier et al., 2009; Sow et al., 2011). Nebulization of 7.6 × 107 PFU of Memphis 37 hRSV strain results in increased expiratory effort in lambs at 4, 6, and 8 days post-infection, a viral titer peak in bronchoalveolar lavage fluid (BALF) at 3 days post-infection and viral antigen in lung tissue, both reduced at 8 days post-infection. Further, infected animals develop histopathologic lesions including bronchitis, bronchiolitis, necrosis, and hyperplasia of epithelia, peribronchial lymphocyte infiltration, and syncytial cells, resembling those described for lambs and infants (Larios Mora et al., 2015).
Lamb respiratory tract allows for a more suitable comparison with the respiratory tract of human infants than rodent model, in terms of development, structure, susceptibility to hRSV strains, pathology, immune response, and clinical manifestations (Plopper et al., 1983; Lindgren et al., 1996; Derscheid and Ackermann, 2012). Similar developmental and structural features include the size of the nasal cavity and airways, the presence of airway submucosal glands that express lactoperoxidase, prenatal alveolar development, airway branching patterns, percentage of Club cells, and development of type II cell (Scheerlinck et al., 2008; Derscheid and Ackermann, 2012; Ackermann, 2014). Additionally, lamb model offers the possibility to evaluate pulmonary functions using the same principles and techniques that are applicable for children and adults (Kirschvink and Reinhold, 2008). These advantages, along with the reported findings, establish the preterm and neonatal lamb as a model with key features that mimics hRSV infection in preterm and neonatal infants.
Additionally, induction of resistance to hRSV infection and development of hRSV-specific antibodies by neonatal and maternal immunization has been recently studied in the sheep model. Immunization of neonates with hRSV F protein in adjuvant provides effective immunization even in the presence of maternal antibodies (MatAbs; Garg et al., 2015). Moreover, pregnant ewes immunized with the same vaccine resulted in transfer of MatAbs to the newborn lambs through the colostrum. Those newborn lambs receiving MatAbs challenged with hRSV at 3 days of age showed a reduction of 70% in viral load, less lung pathology, and higher virus neutralizing titers (VNTs) than in control animals without passive immunization. This newborn lamb model of hRSV is therefore suitable for studies on maternal immunization and evaluation of the ability and safety of a vaccine to induce MatAbs and protection following hRSV challenge (Garg et al., 2016).
Studies on FI-RSV-vaccinated lambs revealed that the immune response triggered by this immunization reduced RSV titers in bronchoalveolar lavage fluids and lungs, as well as histopathology scores, but increased peribronchiolar and perivascular lymphocyte infiltration as compared to lambs undergoing either an acute RSV infection or naïve controls. Furthermore, no evident disease exacerbation was observed (Derscheid et al., 2013). These observations suggest that the lamb FI-RSV infection differs from what has been observed in children presenting FI-RSV enhanced disease (Kim et al., 1969; Prince et al., 2001). However, further studies need to be performed to understand the altered response of lambs to FI-RSV vaccination and to establish the scope of this model.
Technical advantages regarding the use of the lamb model include the ease of repeated sampling and manipulation, a long lifespan that facilitates realization and interpretation of studies on elder population. Unfortunately, there are some significant disadvantages regarding the use of the lamb model, including limited availability of molecular tools for immunologic and genetic studies, and additional efforts and requirements in housing and handling of this species (Bem et al., 2011; Gerdts et al., 2015).
Pneumoviruses (family Paramyxoviridae, subfamily Pneumovirinae) include several pathogens of veterinary concern, including avian metapneumovirus, bovine respiratory syncytial virus (bRSV), ovine and caprine RSVs, pneumonia virus of mice (PVM), and canine pneumovirus (Easton et al., 2004; Renshaw et al., 2011). A pneumoviral infection in its natural host is associated with a fully permissive replication and a persistent inflammatory response. Interest in using some of those natural host pneumoviral infections as a model of hRSV infection has increased, since this approach of study offers several advantages over less permissive models, which are discussed later.
The bRSV infection in cattle (Bos primigenius taurus) occurs naturally and is one of the main pathogens causing pneumonia in the cattle industry along with other viral agents such as parainfluenza type 3. In addition, bRSV infection predisposes animals to secondary bacterial infections with agents such as Mannheimia haemolytica, Pasteurella multocida, Histophilus somni, and Mycoplasma bovis, producing a synergism between viral and bacterial agents known as bovine respiratory disease (van der Sluijs et al., 2010; Ellis, 2017). Therefore, bRSV infection has been extensively studied with the objective of reducing its major health and economic burden. Vaccines for bRSV are commercially available, showing variable efficacy in disease reduction (Bem et al., 2011; Ellis, 2017). Cattle can also be experimentally infected by hRSV passaged in bovine cell lines (Thomas et al., 1984). Interestingly, antigenic similarities, epidemiology, and viral pathogenesis have increased interest in studying bRSV infection in calves as an animal model of hRSV infection. Calves reproduce many of the clinical signs associated with hRSV infection in human infants, with clinical signs such as fever, rhinorrhea, cough, serous or mucopurulent nasal discharge, abnormal pulmonary auscultation, tachypnea, and hypoxia (Sacco et al., 2012a, 2015; Taylor, 2013). After bRSV infection, an incubation period of 2–5 days is followed by a range of clinical signs, from asymptomatic to URTD and severe LTRD. The severity of disease is age dependent; most severe disease occurs in young calves in the first 1–6 months of life; however, older animals are also susceptible when subjected to stressful conditions (Elvander, 1996; Hägglund and Valarcher, 2016). Experimental infection of calves results in macroscopic and microscopic lesions resembling natural infection. Lung pathology peaks at 7–8 days post-infection and includes lung consolidation, atelectasis, and sporadic deposition of hyaline membranes, with microscopic lesions of bronchointerstitial pneumonia, epithelial necrosis, alveolitis, occasional syncytial cells, debris obstruction of lumen of the bronchi and bronchioles, and extensive apoptosis of lung epithelial cells (van der Poel et al., 1996; Woolums et al., 1999; Valarcher and Taylor, 2007; Blodorn et al., 2015).
Increased levels of pro-inflammatory cytokines and chemokines, including CCL5, CCL3, IL-8, IL-6, IL-12, TNF-α, and IFN-γ, are observed after bRSV infection, as summarized from in vitro studies on dendritic cells (Werling et al., 2002) and in vivo calf studies that analyzed lung samples (Antonis et al., 2010; Sacco et al., 2012b). Calves recovering from bRSV infection demonstrate significant neutrophil accumulation in the BALF – as seen in children with hRSV bronchiolitis – as well as a predominance of CD8+ T cells (Taylor et al., 1989; McInnes et al., 1999; Antonis et al., 2010). Neutrophils have a role in clearance of virus that is excreted to the lumen of the respiratory tract (Viuff et al., 2002). On the other hand, recent proteomic analyses performed in bRSV-infected calves support an immunopathologic role for neutrophils in RSV disease, demonstrating a positive correlation between disease severity and neutrophil presence in alveolar septa, as well as upregulation of neutrophil-associated proteins in the BALF during infection. Increased neutrophil degranulation and diminished anti-oxidant activity were also observed in calves with bRSV disease. Noteworthy, several histones were associated with the disease, and citrullinated histone 3, an indicator of neutrophil extracellular traps (NET), was only present in unimmunized, infected animals (Hägglund et al., 2017). Considering previous studies that identified NET formation in the lungs and airways of RSV infected children presenting with severe LTRD (Cortjens et al., 2016), these results suggest that dysregulated neutrophil responses and NET activity contribute to severe immunopathology leading to LTRD after RSV infection (Afonso et al., 2016; Hägglund et al., 2017) and evidence the advantages of using a bovine model to study neutrophil-related pathological mechanisms.
Results from several studies suggest that cellular responses are crucial for clearance of bRSV. CD8+ T cells are the predominant lymphocyte subpopulation in the respiratory tract of calves recovering from bRSV infection (McInnes et al., 1999), and their appearance in the lungs on days 7–10 post-infection coincides with viral clearance (Gaddum et al., 1996; McInnes et al., 1999). Additionally, CD8+ depletion in calves results in increased severity of pneumonic consolidation and prolonged nasopharyngeal excretion of the virus (Taylor et al., 1995). Interestingly, however, rapid viral clearance has been observed in bRSV-infected calves that might be related to apoptotic mechanisms rather than T cell cytotoxicity. Apoptosis of the bronchial epithelium and phagocytosis of bRSV antigen-containing apoptotic cells were evident in the lungs at 6 days post-infection, prior to a significant recruitment of CD8+ cells (Viuff et al., 2002).
γδ T cells have been implicated in disease pathogenesis and the development of airway hyperreactivity following hRSV infection in humans and rodent models (Aoyagi et al., 2003; Dodd et al., 2009; Huang et al., 2015); however, it has been difficult to study their role in the disease due to their rare frequency in circulation. By contrast, γδ T cells comprise around 40% of the circulating lymphocyte pool in ruminants (Mackay and Hein, 1989), making the bovine an important model for studying the role of non-conventional T cells in bRSV infection. Bovine γδ T cells produce multiple proinflammatory cytokines and chemokines in response to in vitro and in vivo bRSV infection (McGill et al., 2013) and are a major source of IL-17 in in vitro bRSV antigen recall assays (McGill et al., 2016). However, their importance in this infection remains unclear, as depletion of γδ T cells does not significantly alter the course of bRSV disease (Taylor et al., 1995).
The calf model is useful for the evaluation of several aspects of vaccine development. The calf is scalable in size to human infants and is a tractable model of the neonatal immune system. Furthermore, vaccine development for bRSV faces many of the same challenges as vaccine development for hRSV, including the need to immunize infant populations and to induce long-lived and balanced immune responses, often in the face of maternal or preexisting immunity. The calf is also useful for studying vaccine-enhanced respiratory disease, as this phenomenon has been observed following natural bRSV infection of Belgian White Blue calves 3–4 months after vaccination, with fatal outcome (Kimman et al., 1989). Studies designed to experimentally induce FI-RSV enhanced respiratory disease in calves have shown mixed results regarding the onset and development of pulmonary disease (West et al., 1999; Antonis et al., 2003; Kalina et al., 2004). The differences in disease presentation may be attributable to different vaccine antigen doses, levels of serum antibodies at the time of vaccination, and the time of subsequent challenge (Taylor, 2017).
Several vaccines have been developed and tested in calves, including recombinant bRSV vaccines, modified live viruses, DNA and subunit vaccines, immunostimulating complexes, and novel adjuvants, such as CpG oligodeoxynucleotides (Oumouna et al., 2005; Letellier et al., 2008; Ellis et al., 2010; Riffault et al., 2010; Hagglund et al., 2011; Kavanagh et al., 2013; Woolums et al., 2013; Blodorn et al., 2014; Taylor et al., 2014, 2015). The calf model has also been used in preclinical safety and immunogenicity studies for vaccine candidates containing conserved proteins between both viruses, leading to phase I clinical trials in healthy human adults (Green et al., 2015; Taylor et al., 2015). The recent description of the pre-fusion form of the hRSV F protein has changed our understanding of hRSV antigenicity (McLellan et al., 2013a,b). Similarly, the recent description of the pre-fusion form of the F protein of bRSV (Zhang et al., 2017) has led to major advancements in bRSV vaccine development. Immunization of seronegative calves with a recombinant pre-fusion F protein in Montanide, an oil-in-water adjuvant, induces neutralizing antibody responses nearly 100-fold greater than similar vaccination with post-fusion form of the F protein and induces sterilizing immunity to bRSV challenge (Zhang et al., 2017). Vaccination with recombinant pre-fusion F was also recently shown to enhance neutralizing antibody responses in adult, seropositive cows (Steff et al., 2017).
Recent studies have also shown that the bovine RSV model constitutes a recommendable in vivo system to study therapeutic compounds aimed to control hRSV replication and airway obstruction. It has been reported that administration of GS1, a fusion inhibitor, to infected calves, provides therapeutic improvement in clinical manifestations, as well as attenuation of lung pathology and viral load (Jordan et al., 2015). More recently, Cortjens and collaborators have demonstrated that therapeutic administration of local dornase alfa reduces NET formation and airway occlusion in 4-week-old bRSV infected calves, highlighting the relevance of targeting NETs to treat severe airway obstruction induced by bRSV (Cortjens et al., 2018). Housing and handling of cattle require more expertise and specialized housing facilities; however, for veterinary personnel accustomed to working with large animals, this model has the advantage of offering a rapid and repeated sampling of the URT, in comparison to mice. Similarities in bovine and human palatine and nasopharyngeal tonsils (Rebelatto et al., 2000) provide an additional sampling zone for cattle and mark a difference from rodent models, which lack this tissue (Velin et al., 1997). Airway submucosal glands are common in humans and cattle. It has been suggested that inflammatory processes in lung traduce in only a brief detrimental effect in mice, because of the relatively large airway lumen and lack of mucosal gland (Irvin and Bates, 2003). Studies on lung function are more easily performed in calves, and the similarities in structure between calves and humans permit a more credible interpretation of those tests (Kirschvink and Reinhold, 2008). These features, added to the natural replication of the virus and clinical manifestations in calves, make this model highly relevant for studies of clinical lung disease by hRSV. The availability of reagents and tools for immunological, genetic, and molecular studies in cattle is reduced when compared to mice. However, an important number of immunological reagents, including monoclonal antibodies to cell surface molecules, recombinant cytokines and chemokines, and cross-reactive anti-human mAbs, is currently available for the cattle (Sacco et al., 2015).
The pneumonia virus of mice (PVM) was first detected in mouse lungs (Horsfall and Hahn, 1940). This virus usually causes natural infections in mouse colonies, especially in immunodeficient mice. In immunocompetent mice, infections are short lived and mostly asymptomatic (Homberger and Thomann, 1994).
Previously described as a rodent-specific pathogen (Parker and Richter, 1982), its host range remains largely unknown, but has been reported as a cause of disease in hedgehogs (Madarame et al., 2014). PVM is the only pneumovirus that is full permissible in the rodent model, with experimental low dose infections leading to clinical disease (Cook et al., 1998; Bonville et al., 2006). C57BL/6 mice show a higher resistance to infection than BALB/c mice when challenged with lethal and sub-lethal doses, probably due to the increased ability of C57BL/6 mice to control viral replication and the immune response elicited by PVM (Watkiss et al., 2013). However, higher protective responses have been observed in a second infection challenge in C57BL/6 mice, several weeks after a previous IN inoculation with 300 PFU of PVM-15 strain (Shrivastava et al., 2015). This protective response appears to be related to an innate pro-inflammatory response and IgA in the lungs after the first inoculation, leading to a higher capacity of generating VNT during the second challenge (Shrivastava et al., 2015). Virus replication capacity is high and occurs in alveolar and bronchial epithelial cells. BALF analyses post-infection have shown an increase in eosinophil and neutrophil infiltration that progresses to an inflammatory state with almost 100% of neutrophil component (Domachowske et al., 2000). This inflammatory state induces local production of proinflammatory mediators including CCL3, CXCL2, and CCL2 (Bonville et al., 2006) consistent with those detected in lung and nasal washes in severe hRSV disease in human infants (Domachowske et al., 2004). High morbidity and mortality are associated to this infection, with clinical signs including difficult breathing, and cyanosis. Histopathological analyses show alveolar epithelial cell apoptosis, bronchial, epithelial necrosis, multifocal acute alveolitis, intraalveolar edema, multifocal hemorrhage, and granulocytic infiltration (Cook et al., 1998). A correlation between pathologic abnormalities and clinical signs has been reported with an IN inoculation of 120 PFU, showing a virus clearance at 10 days post-infection (Cook et al., 1998). A CD8+ T cell response with high levels of TNF-α and IFN-γ in a cytokine storm context has a major role in the pathology induced by this infection, as it has been described in C57BL/6 mice at 5 days post-infection (Walsh et al., 2014). The role of these cells in virus clearance and lung pathology was previously seen in other models of hRSV infection in mice (Cannon et al., 1988; Munoz et al., 1991). However, there are clear differences between PVM and RSV models; acute inflammatory responses to PVM infection vary in anage-dependent form, with scarce recruitment of PMN cells and inflammatory mediators in neonatal mice when compared to 3- or 4-week-age mice (Bonville et al., 2010).
Human respiratory syncytial virus vaccine development studies have been made on this model (Maunder et al., 2015; Martinez et al., 2016) including a model of FI-PVM enhanced respiratory disease, mediated by a Th2-biased response and eosinophil infiltration in lung (Percopo et al., 2009). Most recently, a replication deficient recombinant human adenovirus serotype 5 expressing the M or N protein of PVM pathogenic strain J3666, provided protection in mice, mediated mainly by CD8+ T cells (Maunder et al., 2015). However, there is still a limited number of studies in this field. Moreover, immunological and pathological studies are scarce too, and differences in mice and virus strain and infective doses have resulted in different clinical outcomes; therefore, drawing conclusions on this topic requires further studies. An advantage of rodent models in general is the availability of reagents and tools for immunological studies, including gene knockout mice facilitate mechanistic studies of PVM disease (Moore and Peebles, 2006) making this model useful for elucidation of inflammatory mechanisms associated with pneumovirus infection (Dyer et al., 2012).
Disadvantages of the use of this model include the high genetic distance between PVM and hRSV, which traduces in gene products with low aminoacidic sequence identity, ranging from 10 to 60%, and lack of direct cross-reactivity (Krempl et al., 2005; Dyer et al., 2012). Also, inherent to every mouse model of human pathogens, there are several differences in immune response between species and lung anatomy. Therefore, extrapolations of this model to hRSV studies should be made carefully.
The contribution of animal models to the study of human respiratory syncytial virus has been a fundamental part for the development of vaccines and alternative therapies. Each animal model reviewed in this document presents advantages or disadvantages (Table 1) highlighting the importance of the correct choice of the model according to the hypothesis to be evaluated or the aspect of infection that requires investigation (Figure 1).
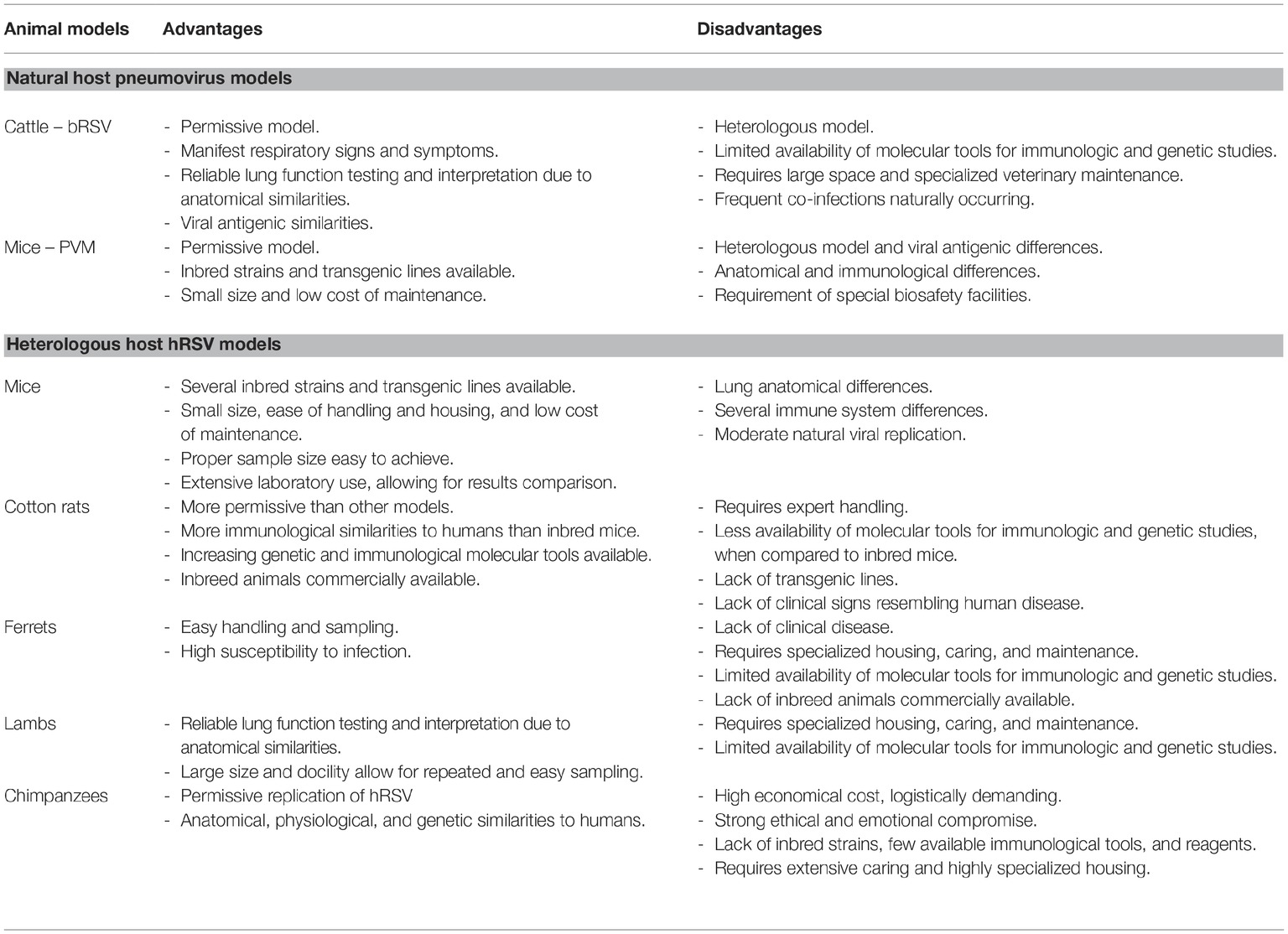
Table 1. Advantages and disadvantages of different animal species suitable for modeling different aspects of hRSV-induced disease.
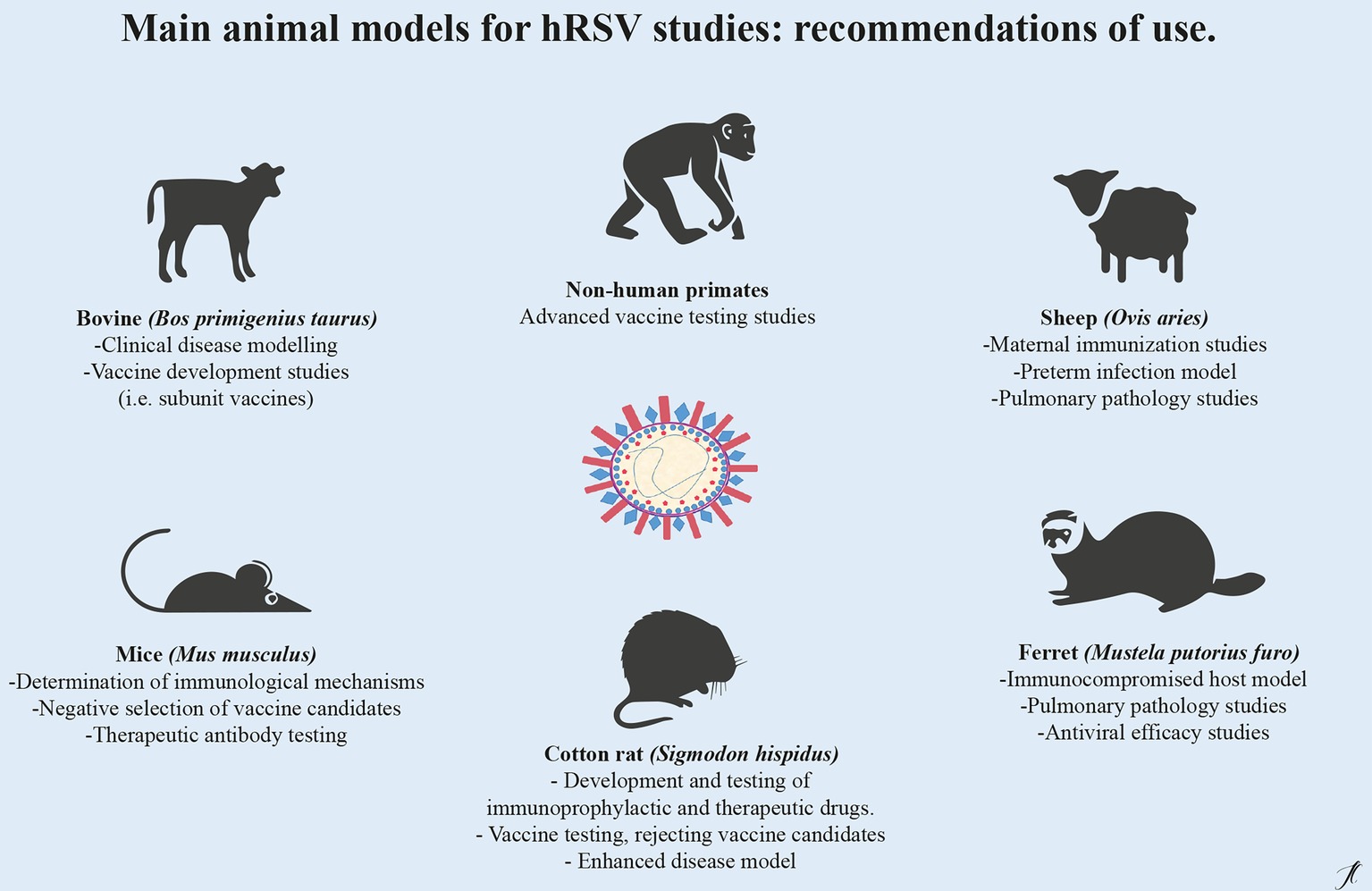
Figure 1. Main animal models for hRSV preclinical studies: recommendations for use. For the study of respiratory syncytial virus, there are different animal models of infection, since there is not a single animal that replicates in an integral way the human pathology and the response to treatment and vaccine candidates. The choice of the model will depend on the investigative hypothesis and should considerate the advantages and disadvantages inherent to every model.
Within the complexity of the natural development of hRSV infection, we emphazise that there is no animal model that recapitulates morbidity and mortality trends, since high viral doses are needed to establish infection and/or manifest disease, as is the case of mice or cotton rats. Among other models of hRSV infection, we highlight the potential of ruminant models to understand pathology aspects of hRSV infection, since calves display a physiopathology very similar to infected human infants, showing very similar histopathological lesions. In case of non-human primates, it is noteworthy that they allow for viral replication with doses lower than those used in other models such as rodents, being a more permissive model for the study of hRSV. However, its use should be restricted exclusively to vaccine testing studies, due to ethical and technical issues. The antecedents reviewed here lead us to conclude that each animal model for the study of hRSV is fundamental for the understanding of the viral pathogenesis and all of them have to a greater or lesser extent to the development of biomedical tools against this agent. Expanding the availability of sophisticated models might be crucial to overcome information gaps and developing safe, effective therapies such as monoclonal antibodies or vaccines. To date, within all existing preclinical models, the murine model is the first preclinical approach for the development of new drugs, being a versatile tool that allows sophisticated immunological studies. Even considering its limitations, this model allow us to advance towards in other animal models that have more complex and expensive requirements in their management, maintenance, and infrastructure, as described in this work.
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
This work was supported by Comisión Nacional de Investigación Científica y Tecnológica (CONICYT) no. 21151028; doctorate grant no. 21170620; FONDECYT grant nos. 1150862 and 1190830; and the Millennium Institute on Immunology and Immunotherapy (P09/016-F).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
AK is a Helen C. Levitt Visiting Professor at the Department of Microbiology and Immunology of the University of Iowa. The authors thank María Trinidad Celis-Dooner for her valuable help in image edition.
Ackermann, M. R. (2014). Lamb model of respiratory syncytial virus-associated lung disease: insights to pathogenesis and novel treatments. ILAR J. 55, 4–15. doi: 10.1093/ilar/ilu003
Afonso, C. L., Amarasinghe, G. K., Banyai, K., Bao, Y., Basler, C. F., Bavari, S., et al. (2016). Taxonomy of the order Mononegavirales: update 2016. Arch. Virol. 161, 2351–2360. doi: 10.1007/s00705-016-2880-1
Andersen, M. L., and Winter, L. M. (2017). Animal models in biological and biomedical research-experimental and ethical concerns. An. Acad. Bras. Cienc. 91(Suppl. 1). doi: 10.1590/0001-3765201720170238
Antonis, A. F., de Jong, M. C., van der Poel, W. H., van der Most, R. G., Stockhofe-Zurwieden, N., Kimman, T., et al. (2010). Age-dependent differences in the pathogenesis of bovine respiratory syncytial virus infections related to the development of natural immunocompetence. J. Gen. Virol. 91, 2497–2506. doi: 10.1099/vir.0.020842-0
Antonis, A. F., Schrijver, R. S., Daus, F., Steverink, P. J., Stockhofe, N., Hensen, E. J., et al. (2003). Vaccine-induced immunopathology during bovine respiratory syncytial virus infection: exploring the parameters of pathogenesis. J. Virol. 77, 12067–12073. doi: 10.1128/JVI.77.22.12067-12073.2003
Aoyagi, M., Shimojo, N., Sekine, K., Nishimuta, T., and Kohno, Y. (2003). Respiratory syncytial virus infection suppresses IFN-gamma production of gammadelta T cells. Clin. Exp. Immunol. 131, 312–317. doi: 10.1046/j.1365-2249.2003.02062.x
Belshe, R. B., Richardson, L. S., London, W. T., Sly, D. L., Lorfeld, J. H., Camargo, E., et al. (1977). Experimental respiratory syncytial virus infection of four species of primates. J. Med. Virol. 1, 157–162. doi: 10.1002/jmv.1890010302
Bem, R. A., Domachowske, J. B., and Rosenberg, H. F. (2011). Animal models of human respiratory syncytial virus disease. Am. J. Physiol. Lung Cell. Mol. Physiol. 301, L148–L156. doi: 10.1152/ajplung.00065.2011
Blanco, J. C. G., Pletneva, L. M., Oue, R. O., Patel, M. C., and Boukhvalova, M. S. (2015). Maternal transfer of RSV immunity in cotton rats vaccinated during pregnancy. Vaccine 33, 5371–5379. doi: 10.1016/j.vaccine.2015.08.071
Blodorn, K., Hagglund, S., Fix, J., Dubuquoy, C., Makabi-Panzu, B., Thom, M., et al. (2014). Vaccine safety and efficacy evaluation of a recombinant bovine respiratory syncytial virus (BRSV) with deletion of the SH gene and subunit vaccines based on recombinant human RSV proteins: N-nanorings, P and M2-1, in calves with maternal antibodies. PLoS One 9:e100392. doi: 10.1371/journal.pone.0100392
Blodorn, K., Hagglund, S., Gavier-Widen, D., Eleouet, J. F., Riffault, S., Pringle, J., et al. (2015). A bovine respiratory syncytial virus model with high clinical expression in calves with specific passive immunity. BMC Vet. Res. 11:76. doi: 10.1186/s12917-015-0389-6
Blount, R. E. Jr., Morris, J. A., and Savage, R. E. (1956). Recovery of cytopathogenic agent from chimpanzees with coryza. Proc. Soc. Exp. Biol. Med. 92, 544–549. doi: 10.3181/00379727-92-22538
Bonville, C. A., Bennett, N. J., Koehnlein, M., Haines, D. M., Ellis, J. A., DelVecchio, A. M., et al. (2006). Respiratory dysfunction and proinflammatory chemokines in the pneumonia virus of mice (PVM) model of viral bronchiolitis. Virology 349, 87–95. doi: 10.1016/j.virol.2006.02.017
Bonville, C. A., Ptaschinski, C., Percopo, C. M., Rosenberg, H. F., and Domachowske, J. B. (2010). Inflammatory responses to acute pneumovirus infection in neonatal mice. Virol. J. 7, 320–320. doi: 10.1186/1743-422X-7-320
Borchers, A. T., Chang, C., Gershwin, M. E., and Gershwin, L. J. (2013). Respiratory syncytial virus—a comprehensive review. Clin. Rev. Allergy Immunol. 45, 331–379. doi: 10.1007/s12016-013-8368-9
Boukhvalova, M. S., and Blanco, J. C. G. (2013). “The cotton rat Sigmodon hispidus model of respiratory syncytial virus infection” in Challenges and opportunities for respiratory syncytial virus vaccines. eds. L. J. Anderson, and B. S. Graham (Berlin, Heidelberg: Springer), 347–358.
Boukhvalova, M., Blanco, J., Falsey, A., and Mond, J. (2016). Treatment with novel RSV Ig RI-002 controls viral replication and reduces pulmonary damage in immunocompromised Sigmodon hispidus. Bone Marrow Transplant. 51, 119–126. doi: 10.1038/bmt.2015.212
Boukhvalova, M. S., Prince, G. A., and Blanco, J. C. (2007a). Respiratory syncytial virus infects and abortively replicates in the lungs in spite of preexisting immunity. J. Virol. 81, 9443–9450. doi: 10.1128/JVI.00102-07
Boukhvalova, M. S., Prince, G. A., Soroush, L., Harrigan, D. C., Vogel, S. N., and Blanco, J. C. (2006). The TLR4 agonist, monophosphoryl lipid A, attenuates the cytokine storm associated with respiratory syncytial virus vaccine-enhanced disease. Vaccine 24, 5027–5035. doi: 10.1016/j.vaccine.2006.03.064
Boukhvalova, M. S., Yim, K. C., Kuhn, K. H., Hemming, J. P., Prince, G. A., Porter, D. D., et al. (2007b). Age-related differences in pulmonary cytokine response to respiratory syncytial virus infection: modulation by anti-inflammatory and antiviral treatment. J. Infect. Dis. 195, 511–518. doi: 10.1086/510628
Bueno, S. M., Gonzalez, P. A., Cautivo, K. M., Mora, J. E., Leiva, E. D., Tobar, H. E., et al. (2008). Protective T cell immunity against respiratory syncytial virus is efficiently induced by recombinant BCG. Proc. Natl. Acad. Sci. USA 105, 20822–20827. doi: 10.1073/pnas.0806244105
Bush, A., and Thomson, A. H. (2007). Acute bronchiolitis. Br. Med. J. 335, 1037–1041. doi: 10.1136/bmj.39374.600081.AD
Byrd, L. G., and Prince, G. A. (1997). Animal models of respiratory syncytial virus infection. Clin. Infect. Dis. 25, 1363–1368. doi: 10.1086/516152
Cannon, M. J., Openshaw, P. J., and Askonas, B. A. (1988). Cytotoxic T cells clear virus but augment lung pathology in mice infected with respiratory syncytial virus. J. Exp. Med. 168, 1163–1168. doi: 10.1084/jem.168.3.1163
Canziani, G. A., Melero, J. A., and Lacy, E. R. (2012). Characterization of neutralizing affinity-matured human respiratory syncytial virus F binding antibodies in the sub-picomolar affinity range. J. Mol. Recognit. 25, 136–146. doi: 10.1002/jmr.2149
Cautivo, K. M., Bueno, S. M., Cortes, C. M., Wozniak, A., Riedel, C. A., and Kalergis, A. M. (2010). Efficient lung recruitment of respiratory syncytial virus-specific Th1 cells induced by recombinant Bacillus Calmette-Guérin promotes virus clearance and protects from infection. J. Immunol. 185, 7633–7645. doi: 10.4049/jimmunol.0903452
Céspedes, P. F., Rey-Jurado, E., Espinoza, J. A., Rivera, C. A., Canedo-Marroquín, G., Bueno, S. M., et al. (2017). A single, low dose of a cGMP recombinant BCG vaccine elicits protective T cell immunity against the human respiratory syncytial virus infection and prevents lung pathology in mice. Vaccine 35, 757–766. doi: 10.1016/j.vaccine.2016.12.048
Chan, K. F., Carolan, L. A., Druce, J., Chappell, K., Watterson, D., Young, P., et al. (2017). Pathogenesis, humoral immune responses and transmission between co-housed animals in a ferret model of human RSV infection. J. Virol. 92:e01322–17. doi: 10.1128/JVI.01322-17
Chan, K. F., Carolan, L. A., Korenkov, D., Druce, J., McCaw, J., Reading, P. C., et al. (2018). Investigating viral interference between influenza a virus and human respiratory syncytial virus in a ferret model of infection. J. Infect. Dis. 218, 406–417. doi: 10.1093/infdis/jiy184
Collins, P. L., and Graham, B. S. (2008). Viral and host factors in human respiratory syncytial virus pathogenesis. J. Virol. 82, 2040–2055. doi: 10.1128/JVI.01625-07
Collins, P. L., Purcell, R. H., London, W. T., Lawrence, L. A., Chanock, R. M., and Murphy, B. R. (1990). Evaluation in chimpanzees of vaccinia virus recombinants that express the surface glycoproteins of human respiratory syncytial virus. Vaccine 8, 164–168. doi: 10.1016/0264-410X(90)90141-8
Cook, P. M., Eglin, R. P., and Easton, A. J. (1998). Pathogenesis of pneumovirus infections in mice: detection of pneumonia virus of mice and human respiratory syncytial virus mRNA in lungs of infected mice by in situ hybridization. J. Gen. Virol. 79, 2411–2417. doi: 10.1099/0022-1317-79-10-2411
Cortjens, B., De Boer, O. J., De Jong, R., Antonis, A. F., Sabogal Piñeros, Y. S., Lutter, R., et al. (2016). Neutrophil extracellular traps cause airway obstruction during respiratory syncytial virus disease. J. Pathol. 238, 401–411. doi: 10.1002/path.4660
Cortjens, B., de Jong, R., Bonsing, J. G., Van Woensel, J. B., Antonis, A. F., and Bem, R. A. (2018). Local dornase alfa treatment reduces NETs-induced airway obstruction during severe RSV infection. Thorax 73, 578–580. doi: 10.1136/thoraxjnl-2017-210289
Crowe, J. E. Jr., Bui, P. T., Siber, G. R., Elkins, W. R., Chanock, R. M., and Murphy, B. R. (1995). Cold-passaged, temperature-sensitive mutants of human respiratory syncytial virus (RSV) are highly attenuated, immunogenic, and protective in seronegative chimpanzees, even when RSV antibodies are infused shortly before immunization. Vaccine 13, 847–855. doi: 10.1016/0264-410X(94)00074-W
Crowe, J. E. Jr., Collins, P. L., London, W. T., Chanock, R. M., and Murphy, B. R. (1993). A comparison in chimpanzees of the immunogenicity and efficacy of live attenuated respiratory syncytial virus (RSV) temperature-sensitive mutant vaccines and vaccinia virus recombinants that express the surface glycoproteins of RSV. Vaccine 11, 1395–1404. doi: 10.1016/0264-410X(93)90168-W
Cullen, L. M., Schmidt, M. R., Kenward, S. A., Woodland, R. T., and Morrison, T. G. (2015). Murine immune responses to virus-like particle associated pre-and post-fusion forms of the respiratory syncytial virus F protein. J. Virol. 89, 6835–6847. doi: 10.1128/JVI.00384-15
Curtis, S. J., Ottolini, M. G., Porter, D. D., and Prince, G. A. (2002). Age-dependent replication of respiratory syncytial virus in the cotton rat. Exp. Biol. Med. 227, 799–802. doi: 10.1177/153537020222700912
De Swart, R. L., Kuiken, T., Timmerman, H. H., Van Amerongen, G., Van Den Hoogen, B. G., Vos, H. W., et al. (2002). Immunization of macaques with formalin-inactivated respiratory syncytial virus (RSV) induces interleukin-13-associated hypersensitivity to subsequent RSV infection. J. Virol. 76, 11561–11569. doi: 10.1128/JVI.76.22.11561-11569.2002
de Waal, L., Smits, S. L., Veldhuis Kroeze, E. J. B., van Amerongen, G., Pohl, M. O., Osterhaus, A. D. M. E., et al. (2018). Transmission of human respiratory syncytial virus in the immunocompromised ferret model. Viruses 10:18. doi: 10.3390/v10010018
de Waal, L., Wyatt, L. S., Yüksel, S., van Amerongen, G., Moss, B., Niesters, H. G. M., et al. (2004). Vaccination of infant macaques with a recombinant modified vaccinia virus Ankara expressing the respiratory syncytial virus F and G genes does not predispose for immunopathology. Vaccine 22, 923–926. doi: 10.1016/j.vaccine.2003.10.010
Derscheid, R. J., and Ackermann, M. R. (2012). Perinatal lamb model of respiratory syncytial virus (RSV) infection. Viruses 4, 2359–2378. doi: 10.3390/v4102359
Derscheid, R. J., Gallup, J. M., Knudson, C. J., Varga, S. M., Grosz, D. D., van Geelen, A., et al. (2013). Effects of formalin-inactivated respiratory syncytial virus (FI-RSV) in the perinatal lamb model of RSV. PLoS One 8:e81472. doi: 10.1371/journal.pone.0081472
Dodd, J., Riffault, S., Kodituwakku, J. S., Hayday, A. C., and Openshaw, P. J. (2009). Pulmonary V gamma 4+ gamma delta T cells have proinflammatory and antiviral effects in viral lung disease. J. Immunol. 182, 1174–1181. doi: 10.4049/jimmunol.182.2.1174
Domachowske, J. B., Bonville, C. A., Dyer, K. D., Easton, A. J., and Rosenberg, H. F. (2000). Pulmonary eosinophilia and production of MIP-1alpha are prominent responses to infection with pneumonia virus of mice. Cell. Immunol. 200, 98–104. doi: 10.1006/cimm.2000.1620
Domachowske, J. B., Bonville, C. A., and Rosenberg, H. F. (2004). Animal models for studying respiratory syncytial virus infection and its long term effects on lung function. Pediatr. Infect. Dis. J. 23(Suppl. 11), S228–S234.
Drake, A. C. (2013). Of mice and men: what rodent models don’t tell us. Cell. Mol. Immunol. 10, 284–285. doi: 10.1038/cmi.2013.21
Dyer, K. D., Garcia-Crespo, K. E., Glineur, S., Domachowske, J. B., and Rosenberg, H. F. (2012). The pneumonia virus of mice (PVM) model of acute respiratory infection. Viruses 4, 3494–3510. doi: 10.3390/v4123494
Easton, A. J., Domachowske, J. B., and Rosenberg, H. F. (2004). Animal pneumoviruses: molecular genetics and pathogenesis. Clin. Microbiol. Rev. 17, 390–412. doi: 10.1128/CMR.17.2.390-412.2004
Ehret, T., Torelli, F., Klotz, C., Pedersen, A. B., and Seeber, F. (2017). Translational rodent models for research on parasitic protozoa—a review of confounders and possibilities. Front. Cell. Infect. Microbiol. 7:238. doi: 10.3389/fcimb.2017.00238
Ellis, J. A. (2017). How efficacious are vaccines against bovine respiratory syncytial virus in cattle? Vet. Microbiol. 206, 59–68. doi: 10.1016/j.vetmic.2016.11.030
Ellis, J. A., Gow, S. P., and Goji, N. (2010). Response to experimentally induced infection with bovine respiratory syncytial virus following intranasal vaccination of seropositive and seronegative calves. J. Am. Vet. Med. Assoc. 236, 991–999. doi: 10.2460/javma.236.9.991
Elvander, M. (1996). Severe respiratory disease in dairy cows caused by infection with bovine respiratory syncytial virus. Vet. Rec. 138, 101–105. doi: 10.1136/vr.138.5.101
Enkirch, T., and Von Messling, V. (2015). Ferret models of viral pathogenesis. Virology 479, 259–270. doi: 10.1016/j.virol.2015.03.017
Eyles, J. E., Johnson, J. E., Megati, S., Roopchand, V., Cockle, P. J., Weeratna, R., et al. (2013). Nonreplicating vaccines can protect african green monkeys from the memphis 37 strain of respiratory syncytial virus. J. Infect. Dis. 208, 319–329. doi: 10.1093/infdis/jit169
Fuentes, S., Klenow, L., Golding, H., and Khurana, S. (2017). Preclinical evaluation of bacterially produced RSV-G protein vaccine: strong protection against RSV challenge in cotton rat model. Sci. Rep. 7:42428. doi: 10.1038/srep42428
Fulton, R. B., Weiss, K. A., Pewe, L. L., Harty, J. T., and Varga, S. M. (2013). Aged mice exhibit a severely diminished CD8 T cell response following respiratory syncytial virus infection. J. virol. 87, 12694–12700.
Gaddum, R. M., Cook, R. S., Thomas, L. H., and Taylor, G. (1996). Primary cytotoxic T-cell responses to bovine respiratory syncytial virus in calves. Immunology 88, 421–427.
Garg, R., Latimer, L., Gerdts, V., Potter, A., and van Drunen Littel-van den Hurk, S. (2015). The respiratory syncytial virus fusion protein formulated with a novel combination adjuvant induces balanced immune responses in lambs with maternal antibodies. Vaccine 33, 1338–1344. doi: 10.1016/j.vaccine.2015.01.041
Garg, R., Latimer, L., Wang, Y., Simko, E., Gerdts, V., Potter, A., et al. (2016). Maternal immunization with respiratory syncytial virus fusion protein formulated with a novel combination adjuvant provides protection from RSV in newborn lambs. Vaccine 34, 261–269. doi: 10.1016/j.vaccine.2015.11.029
Garlapati, S., Garg, R., Brownlie, R., Latimer, L., Simko, E., Hancock, R., et al. (2012). Enhanced immune responses and protection by vaccination with respiratory syncytial virus fusion protein formulated with CpG oligodeoxynucleotide and innate defense regulator peptide in polyphosphazene microparticles. Vaccine 30, 5206–5214. doi: 10.1016/j.vaccine.2012.06.011
Gerdts, V., Wilson, H. L., Meurens, F., van Drunen Littel-van den Hurk, S., Wilson, D., Walker, S., et al. (2015). Large animal models for vaccine development and testing. ILAR J. 56, 53–62. doi: 10.1093/ilar/ilv009
Gitiban, N., Jurcisek, J. A., Harris, R. H., Mertz, S. E., Durbin, R. K., Bakaletz, L. O., et al. (2005). Chinchilla and murine models of upper respiratory tract infections with respiratory syncytial virus. J. Virol. 79, 6035–6042. doi: 10.1128/JVI.79.10.6035-6042.2005
Graham, B. S., Perkins, M. D., Wright, P. F., and Karzon, D. T. (1988). Primary respiratory syncytial virus infection in mice. J. Med. Virol. 26, 153–162. doi: 10.1002/jmv.1890260207
Grandin, C., Lucas-Hourani, M., Clavel, M., Taborik, F., Vabret, A., Tangy, F., et al. (2015). Evidence for an intranasal immune response to human respiratory syncytial virus infection in cynomolgus macaques. J. Gen. Virol. 96, 782–792. doi: 10.1099/vir.0.000039
Green, C. A., Scarselli, E., Sande, C. J., Thompson, A. J., de Lara, C. M., Taylor, K. S., et al. (2015). Chimpanzee adenovirus- and MVA-vectored respiratory syncytial virus vaccine is safe and immunogenic in adults. Sci. Transl. Med. 7:300ra126. doi: 10.1126/scitranslmed.aac5745
Grieves, J. L., Jurcisek, J. A., Quist, B., Durbin, R. K., Peeples, M. E., Durbin, J. E., et al. (2010). Mapping the anatomy of respiratory syncytial virus infection of the upper airways in Chinchillas (Chinchilla lanigera). Comp. Med. 60, 225–232.
Group, I.-R.S (1998). Palivizumab, a humanized respiratory syncytial virus monoclonal antibody, reduces hospitalization from respiratory syncytial virus infection in high-risk infants. Pediatrics 102, 531–537. doi: 10.1542/peds.102.3.531
Grunwald, T., Tenbusch, M., Schulte, R., Raue, K., Wolf, H., Hannaman, D., et al. (2014). Novel vaccine regimen elicits strong airway immune responses and control of respiratory syncytial virus in non-human primates. J. Virol. 88, 3997–4007. doi: 10.1128/JVI.02736-13
Hacking, D., and Hull, J. (2002). Respiratory syncytial virus—viral biology and the host response. J. Infect. 45, 18–24. doi: 10.1053/jinf.2002.1015
Hägglund, S., Blodörn, K., Näslund, K., Vargmar, K., Lind, S. B., Mi, J., et al. (2017). Proteome analysis of bronchoalveolar lavage from calves infected with bovine respiratory syncytial virus—Insights in pathogenesis and perspectives for new treatments. PLoS One 12:e0186594. doi: 10.1371/journal.pone.0186594
Hagglund, S., Hu, K., Vargmar, K., Pore, L., Olofson, A. S., Blodorn, K., et al. (2011). Bovine respiratory syncytial virus ISCOMs-immunity, protection and safety in young conventional calves. Vaccine 29, 8719–8730. doi: 10.1016/j.vaccine.2011.07.146
Hägglund, S., and Valarcher, J. (2016). Bovine respiratory syncytial virus. Mol. Detect. Animal Viral Pathog. 413–420. doi: 10.1201/b19719-51
Hegele, R., Robinson, P., Gonzalez, S., and Hogg, J. (1993). Production of acute bronchiolitis in guinea-pigs by human respiratory syncytial virus. Eur. Respir. J. 6, 1324–1331.
Hemming, V. G., Prince, G. A., Groothuis, J. R., and Siber, G. R. (1995). Hyperimmune globulins in prevention and treatment of respiratory syncytial virus infections. Clin. Microbiol. Rev. 8, 22–33. doi: 10.1128/CMR.8.1.22
Heylen, E., Neyts, J., and Jochmans, D. (2017). Drug candidates and model systems in respiratory syncytial virus antiviral drug discovery. Biochem. Pharmacol. 127, 1–12. doi: 10.1016/j.bcp.2016.09.014
Homberger, F. R., and Thomann, P. E. (1994). Transmission of murine viruses and mycoplasma in laboratory mouse colonies with respect to housing conditions. Lab. Anim. 28, 113–120. doi: 10.1258/002367794780745263
Horsfall, F. L., and Hahn, R. G. (1940). A latent virus in normal mice capable of producing pneumonia in its natural host. J. Exp. Med. 71, 391–408. doi: 10.1084/jem.71.3.391
Huang, H., Saravia, J., You, D., Shaw, A. J., and Cormier, S. A. (2015). Impaired gamma delta T cell-derived IL-17A and inflammasome activation during early respiratory syncytial virus infection in infants. Immunol. Cell Biol. 93, 126–135. doi: 10.1038/icb.2014.79
Hurwitz, J. L. (2011). Respiratory syncytial virus vaccine development. Expert Rev. Vaccines 10, 1415–1433. doi: 10.1586/erv.11.120
Irvin, C. G., and Bates, J. H. (2003). Measuring the lung function in the mouse: the challenge of size. Respir. Res. 4:4. doi: 10.1186/rr199
Ispas, G., Koul, A., Verbeeck, J., Sheehan, J., Sanders-Beer, B., Roymans, D., et al. (2015). Antiviral activity of TMC353121, a respiratory syncytial virus (RSV) fusion inhibitor, in a non-human primate model. PLoS One 10:e0126959. doi: 10.1371/journal.pone.0126959
Johnson, S., Griego, S. D., Pfarr, D. S., Doyle, M. L., Woods, R., Carlin, D., et al. (1999). A direct comparison of the activities of two humanized respiratory syncytial virus monoclonal antibodies: MEDI-493 and RSHZl9. J. Infect. Dis. 180, 35–40. doi: 10.1086/314846
Johnson, R., Prince, G., Suffin, S., Horswood, R., and Chanock, R. (1982). Respiratory syncytial virus infection in cyclophosphamide-treated cotton rats. Infect. Immun. 37, 369–373.
Jones, B. G., Sealy, R. E., Rudraraju, R., Traina-Dorge, V. L., Finneyfrock, B., Cook, A., et al. (2012). Sendai virus-based RSV vaccine protects African green monkeys from RSV infection. Vaccine 30, 959–968. doi: 10.1016/j.vaccine.2011.11.046
Jordan, R., Shao, M., Mackman, R. L., Perron, M., Cihlar, T., Lewis, S. A., et al. (2015). antiviral efficacy of a respiratory syncytial virus (RSV) fusion inhibitor in a bovine model of RSV infection. Antimicrob. Agents Chemother. 59, 4889–4900. doi: 10.1128/aac.00487-15
Jorquera, P. A., Anderson, L., and Tripp, R. A. (2016). Understanding respiratory syncytial virus (RSV) vaccine development and aspects of disease pathogenesis. Expert. Rev. Vaccines 15, 173–187. doi: 10.1586/14760584.2016.1115353
Kakuk, T. J., Soike, K., Brideau, R. J., Zaya, R. M., Cole, S. L., Zhang, J.-Y., et al. (1993). A human respiratory syncytial virus (RSV) primate model of enhanced pulmonary pathology induced with a formalin-inactivated RSV vaccine but not a recombinant FG subunit vaccine. J. Infect. Dis. 167, 553–561. doi: 10.1093/infdis/167.3.553
Kalina, W. V., Woolums, A. R., Berghaus, R. D., and Gershwin, L. J. (2004). Formalin-inactivated bovine RSV vaccine enhances a Th2 mediated immune response in infected cattle. Vaccine 22, 1465–1474. doi: 10.1016/j.vaccine.2003.10.024
Kamphuis, T., Stegmann, T., Meijerhof, T., Wilschut, J., and de Haan, A. (2013). A virosomal respiratory syncytial virus vaccine adjuvanted with monophosphoryl lipid A provides protection against viral challenge without priming for enhanced disease in cotton rats. Influenza Other Respir. Viruses 7, 1227–1236. doi: 10.1111/irv.12112
Kavanagh, O. V., Adair, B. M., Welsh, M., and Earley, B. (2013). Immunogenetic responses in calves to intranasal delivery of bovine respiratory syncytial virus (BRSV) epitopes encapsulated in poly (DL-lactide-co-glycolide) microparticles. Res. Vet. Sci. 95, 786–793. doi: 10.1016/j.rvsc.2013.06.023
Kim, H. W., Canchola, J. G., Brandt, C. D., Pyles, G., Chanock, R. M., Jensen, K., et al. (1969). Respiratory syncytial virus disease in infants despite prior administration of antigenic inactivated vaccine. Am. J. Epidemiol. 89, 422–434. doi: 10.1093/oxfordjournals.aje.a120955
Kimman, T. G., Sol, J., Westenbrink, F., and Strayer, P. J. (1989). A severe outbreak of respiratory tract disease associated with bovine respiratory syncytial virus probably enhanced by vaccination with modified live vaccine. Vet. Q. 11, 250–253. doi: 10.1080/01652176.1989.9694231
Kirschvink, N., and Reinhold, P. (2008). Use of alternative animals as asthma models. Curr. Drug Targets 9, 470–484. doi: 10.2174/138945008784533525
Koff, W. C., Caplan, F. R., Case, S., and Halstead, S. B. (1983). Cell-mediated immune response to respiratory syncytial virus infection in owl monkeys. Clin. Exp. Immunol. 53, 272–280.
Kong, X., Hellermann, G. R., Patton, G., Kumar, M., Behera, A., Randall, T. S., et al. (2005). An immunocompromised BALB/c mouse model for respiratory syncytial virus infection. Virol. J. 2:3. doi: 10.1186/1743-422X-2-3
Krempl, C. D., Lamirande, E. W., and Collins, P. L. (2005). Complete sequence of the RNA genome of pneumonia virus of mice (PVM). Virus Genes 30, 237–249. doi: 10.1007/s11262-004-5631-4
Kuhdari, P., Brosio, F., Malaventura, C., Stefanati, A., Orsi, A., Icardi, G., et al. (2018). Human respiratory syncytial virus and hospitalization in young children in Italy. Ital. J. Pediatr. 44:50. doi: 10.1186/s13052-018-0492-y
Larios Mora, A., Detalle, L., Van Geelen, A., Davis, M. S., Stohr, T., Gallup, J. M., et al. (2015). Kinetics of respiratory syncytial virus (RSV) memphis strain 37 (M37) infection in the respiratory tract of newborn lambs as an RSV infection model for human infants. PLoS One 10:e0143580. doi: 10.1371/journal.pone.0143580
Lee, B. H., Gauna, A. E., Pauley, K. M., Park, Y.-J., and Cha, S. (2012). Animal models in autoimmune diseases: lessons learned from mouse models for Sjögren’s syndrome. Clin. Rev. Allergy Immunol. 42, 35–44. doi: 10.1007/s12016-011-8288-5
Letellier, C., Boxus, M., Rosar, L., Toussaint, J. F., Walravens, K., Roels, S., et al. (2008). Vaccination of calves using the BRSV nucleocapsid protein in a DNA prime-protein boost strategy stimulates cell-mediated immunity and protects the lungs against BRSV replication and pathology. Vaccine 26, 4840–4848. doi: 10.1016/j.vaccine.2008.06.100
Lindgren, C., Lin, J., Graham, B. S., Gray, M. E., Parker, R. A., and Sundell, H. W. (1996). Respiratory syncytial virus infection enhances the response to laryngeal chemostimulation and inhibits arousal from sleep in young lambs. Acta Paediatr. 85, 789–797. doi: 10.1111/j.1651-2227.1996.tb14153.x
Liu, B., and Kimura, Y. (2007). Local immune response to respiratory syncytial virus infection is diminished in senescence-accelerated mice. J. Gen. Virol. 88, 2552–2558. doi: 10.1099/vir.0.83089-0
Luongo, C., Winter, C. C., Collins, P. L., and Buchholz, U. J. (2013). Respiratory syncytial virus modified by deletions of the NS2 gene and amino acid S1313 of the L polymerase protein is a temperature sensitive live-attenuated vaccine candidate that is phenotypically stable at physiological temperature. J. Virol. 87, 1985–1996. doi: 10.1128/JVI.02769-12
Mackay, C. R., and Hein, W. R. (1989). A large proportion of bovine T cells express the gamma delta T cell receptor and show a distinct tissue distribution and surface phenotype. Int. Immunol. 1, 540–545. doi: 10.1093/intimm/1.5.540
Mackow, N., Carambot, E. A., Liang, B., Surman, S., Lingemann, M., Yang, L., et al. (2015). Attenuated human parainfluenza virus type 1 (HPIV1) expressing the fusion F glycoprotein of human respiratory syncytial virus (RSV) as a bivalent HPIV1/RSV vaccine. J. Virol. 89, 10319–10332. doi: 10.1128/JVI.01380-15
Madarame, H., Ogihara, K., Kimura, M., Nagai, M., Omatsu, T., Ochiai, H., et al. (2014). Detection of a pneumonia virus of mice (PVM) in an African hedgehog (Atelerix albiventris) with suspected wobbly hedgehog syndrome (WHS). Vet. Microbiol. 173, 136–140. doi: 10.1016/j.vetmic.2014.07.012
Malley, R., DeVincenzo, J., Ramilo, O., Dennehy, P. H., Meissner, H. C., Gruber, W. C., et al. (1998). Reduction of respiratory syncytial virus (RSV) in tracheal aspirates in intubated infants by use of humanized monoclonal antibody to RSV F protein. J. Infect. Dis. 178, 1555–1561. doi: 10.1086/314523
Martinez, E. C., Garg, R., Shrivastava, P., Gomis, S., and van Drunen Littel-van den Hurk, S. (2016). Intranasal treatment with a novel immunomodulator mediates innate immune protection against lethal pneumonia virus of mice. Antivir. Res. 135, 108–119. doi: 10.1016/j.antiviral.2016.10.008
Masot, A. J., Gazquez, A., Regodon, S., Franco, A., and Redondo, E. (1995). Lesions in lambs experimentally infected with bovine respiratory syncytial virus. Histol. Histopathol. 10, 71–77.
Masot, A. J., Gomez-Tejedor, C., Gomez, L., Gazquez, A., and Redondo, E. (1996). Pathological study of experimentally induced bovine respiratory syncytial viral infection in lambs. Zentralbl. Veterinarmed. B 43, 233–243.
Maunder, H. E., Taylor, G., Leppard, K. N., and Easton, A. J. (2015). Intranasal immunisation with recombinant adenovirus vaccines protects against a lethal challenge with pneumonia virus of mice. Vaccine 33, 6641–6649. doi: 10.1016/j.vaccine.2015.10.105
Mazur, N. I., Higgins, D., Nunes, M. C., Melero, J. A., Langedijk, A. C., Horsley, N., et al. (2018). The respiratory syncytial virus vaccine landscape: lessons from the graveyard and promising candidates. Lancet Infect. Dis. 18, PE295–PE311. doi: 10.1016/S1473-3099(18)30292-5
McArthur-Vaughan, K., and Gershwin, L. (2002). A rhesus monkey model of respiratory syncytial virus infection. J. Med. Primatol. 31, 61–73. doi: 10.1034/j.1600-0684.2002.01006.x
McGill, J. L., Nonnecke, B. J., Lippolis, J. D., Reinhardt, T. A., and Sacco, R. E. (2013). Differential chemokine and cytokine production by neonatal bovine gammadelta T-cell subsets in response to viral toll-like receptor agonists and in vivo respiratory syncytial virus infection. Immunology 139, 227–244. doi: 10.1111/imm.12075
McGill, J. L., Rusk, R. A., Guerra-Maupome, M., Briggs, R. E., and Sacco, R. E. (2016). Bovine gamma delta t cells contribute to exacerbated IL-17 production in response to co-infection with bovine RSV and Mannheimia haemolytica. PLoS One 11:e0151083. doi: 10.1371/journal.pone.0151083
McGillivary, G., Jordan, Z. B., Peeples, M. E., and Bakaletz, L. O. (2013). Replication of respiratory syncytial virus is inhibited by the host defense molecule viperin. J. Innate Immun. 5, 60–71. doi: 10.1159/000342473
McInnes, E., Sopp, P., Howard, C. J., and Taylor, G. (1999). Phenotypic analysis of local cellular responses in calves infected with bovine respiratory syncytial virus. Immunology 96, 396–403.
McLellan, J. S., Chen, M., Joyce, M. G., Sastry, M., Stewart-Jones, G. B., Yang, Y., et al. (2013a). Structure-based design of a fusion glycoprotein vaccine for respiratory syncytial virus. Science 342, 592–598. doi: 10.1126/science.1243283
McLellan, J. S., Chen, M., Leung, S., Graepel, K. W., Du, X., Yang, Y., et al. (2013b). Structure of RSV fusion glycoprotein trimer bound to a prefusion-specific neutralizing antibody. Science 340, 1113–1117. doi: 10.1126/science.1234914
Mestas, J., and Hughes, C. C. W. (2004). Of mice and not men: differences between mouse and human immunology. J. Immunol. 172:2731. doi: 10.4049/jimmunol.172.5.2731
Meyerholz, D. K., Grubor, B., Fach, S. J., Sacco, R. E., Lehmkuhl, H. D., Gallup, J. M., et al. (2004). Reduced clearance of respiratory syncytial virus infection in a preterm lamb model. Microbes Infect. 6, 1312–1319. doi: 10.1016/j.micinf.2004.08.006
Medicine, I. O., and Council, N. R. (2011). Chimpanzees in biomedical and behavioral research: Assessing the necessity. (Washington, DC: National Academies Press). doi: 10.17226/13257
Moore, M. L., and Peebles, R. S. Jr. (2006). Respiratory syncytial virus disease mechanisms implicated by human, animal model, and in vitro data facilitate vaccine strategies and new therapeutics. Pharmacol. Ther. 112, 405–424. doi: 10.1016/j.pharmthera.2006.04.008
Morris, J., Blount, R. Jr., and Savage, R. (1956). Recovery of cytopathogenic agent from chimpanzees with goryza. Proc. Soc. Exp. Biol. Med. 92, 544–549. doi: 10.3181/00379727-92-22538
Mosquera, R. A., Stark, J. M., Atkins, C. L., Colasurdo, G. N., Chevalier, J., Samuels, C. L., et al. (2014). Functional and immune response to respiratory syncytial virus infection in aged BALB/c mice: a search for genes determining disease severity. Expert. Lung Res. 40, 40–49.
Munoz, J. L., McCarthy, C. A., Clark, M. E., and Hall, C. B. (1991). Respiratory syncytial virus infection in C57BL/6 mice: clearance of virus from the lungs with virus-specific cytotoxic T cells. J. Virol. 65, 4494–4497.
Muñoz-Durango, N., Pizarro-Ortega, M. S., Rey-Jurado, E., Díaz, F. E., Bueno, S. M., and Kalergis, A. M. (2018). Patterns of antibody response during natural hRSV infection: insights for the development of new antibody-based therapies. Expert Opin. Investig. Drugs 27, 721–731. doi: 10.1080/13543784.2018.1511699
Nair, H., Nokes, D. J., Gessner, B. D., Dherani, M., Madhi, S. A., Singleton, R. J., et al. (2010). Global burden of acute lower respiratory infections due to respiratory syncytial virus in young children: a systematic review and meta-analysis. Lancet 375, 1545–1555. doi: 10.1016/S0140-6736(10)60206-1
Nguyen, D. T., Louwen, R., Elberse, K., van Amerongen, G., Yüksel, S., Luijendijk, A., et al. (2015). Streptococcus pneumoniae enhances human respiratory syncytial virus infection in vitro and in vivo. PLoS One 10:e0127098. doi: 10.1371/journal.pone.0127098
Niewiesk, S., and Prince, G. (2002). Diversifying animal models: the use of hispid cotton rats (Sigmodon hispidus) in infectious diseases. Lab. Anim. 36, 357–372. doi: 10.1258/002367702320389026
Olchanski, N., Hansen, R. N., Pope, E., D’Cruz, B., Fergie, J., Goldstein, M., et al. (2018). Palivizumab prophylaxis for respiratory syncytial virus: examining the evidence around value. Open Forum Infect. Dis. 5:ofy031. doi: 10.1093/ofid/ofy031
Olivier, A., Gallup, J., de Macedo, M. M., Varga, S. M., and Ackermann, M. (2009). Human respiratory syncytial virus A2 strain replicates and induces innate immune responses by respiratory epithelia of neonatal lambs. Int. J. Exp. Pathol. 90, 431–438. doi: 10.1111/j.1365-2613.2009.00643.x
Olmsted, R. A., Buller, R. M. L., Collins, P. L., London, W. T., Beeler, J. A., Prince, G. A., et al. (1988). Evaluation in non-human primates of the safety, immunogenicity and efficacy of recombinant vaccinia viruses expressing the F or G glycoprotein of respiratory syncytial virus. Vaccine 6, 519–524. doi: 10.1016/0264-410X(88)90104-1
Openshaw, P. J., and Tregoning, J. S. (2005). Immune responses and disease enhancement during respiratory syncytial virus infection. Clin. Microbiol. Rev. 18, 541–555. doi: 10.1128/CMR.18.3.541-555.2005
Ottolini, M. G., Curtis, S. R., Mathews, A., Ottolini, S. R., and Prince, G. A. (2002). Palivizumab is highly effective in suppressing respiratory syncytial virus in an immunosuppressed animal model. Bone Marrow Transplant. 29, 117–120. doi: 10.1038/sj.bmt.1703326
Ottolini, M., Porter, D., Hemming, V., Zimmerman, M., Schwab, N., and Prince, G. (1999). Effectiveness of RSVIG prophylaxis and therapy of respiratory syncytial virus in an immunosuppressed animal model. Bone Marrow Transplant. 24, 41–45. doi: 10.1038/sj.bmt.1701813
Oumouna, M., Mapletoft, J. W., Karvonen, B. C., Babiuk, L. A., and van Drunen Littel-van den Hurk, S. (2005). Formulation with CpG oligodeoxynucleotides prevents induction of pulmonary immunopathology following priming with formalin-inactivated or commercial killed bovine respiratory syncytial virus vaccine. J. Virol. 79, 2024–2032. doi: 10.1128/JVI.79.4.2024-2032.2005
Padilla-Carlin, D. J., McMurray, D. N., and Hickey, A. J. (2008). The guinea pig as a model of infectious diseases. Comp. Med. 58, 324–340.
Parker, J. C., and Richter, C. B. (1982). “Chapter 8—viral diseases of the respiratory system” in Diseases. eds. H. L. Foster, J. D. Small, and J. G. Fox (Academic Press), 109–158.
Patton, K., Aslam, S., Shambaugh, C., Lin, R., Heeke, D., Frantz, C., et al. (2015). Enhanced immunogenicity of a respiratory syncytial virus (RSV) F subunit vaccine formulated with the adjuvant GLA-SE in cynomolgus macaques. Vaccine 33, 4472–4478. doi: 10.1016/j.vaccine.2015.07.025
Peebles, R. S., and Graham, B. S. (2005). Pathogenesis of respiratory syncytial virus infection in the murine model. Proc. Am. Thorac. Soc. 2, 110–115. doi: 10.1513/pats.200501-002AW
Percopo, C. M., Qiu, Z., Phipps, S., Foster, P. S., Domachowske, J. B., and Rosenberg, H. F. (2009). Pulmonary eosinophils and their role in immunopathologic responses to formalin-inactivated pneumonia virus of mice. J. Immunol. 183, 604–612. doi: 10.4049/jimmunol.0802270
Pletneva, L. M., Haller, O., Porter, D. D., Prince, G. A., and Blanco, J. C. (2008). Induction of type I interferons and interferon-inducible Mx genes during respiratory syncytial virus infection and reinfection in cotton rats. J. Gen. Virol. 89, 261–270. doi: 10.1099/vir.0.83294-0
Plopper, C. G., Mariassy, A. T., and Lollini, L. O. (1983). Structure as revealed by airway dissection. A comparison of mammalian lungs. Am. Rev. Respir. Dis. 128, S4–S7. doi: 10.1164/arrd.1983.128.2P2.S4
Ponnuraj, E. M., Hayward, A. R., Raj, A., Wilson, H., and Simoes, E. A. (2001). Increased replication of respiratory syncytial virus (RSV) in pulmonary infiltrates is associated with enhanced histopathological disease in bonnet monkeys (Macaca radiata) pre-immunized with a formalin-inactivated RSV vaccine. J. Gen. Virol. 82, 2663–2674. doi: 10.1099/0022-1317-82-11-2663
Prince, G. A., Curtis, S. J., Yim, K. C., and Porter, D. D. (2001). Vaccine-enhanced respiratory syncytial virus disease in cotton rats following immunization with Lot 100 or a newly prepared reference vaccine. J. Gen. Virol. 82, 2881–2888. doi: 10.1099/0022-1317-82-12-2881
Prince, G. A., Horswood, R. L., Camargo, E., Koenig, D., and Chanock, R. M. (1983). Mechanisms of immunity to respiratory syncytial virus in cotton rats. Infect. Immun. 42, 81–87.
Prince, G. A., Horswood, R. L., Camargo, E., Suffin, S. C., and Chanock, R. M. (1982). Parenteral immunization with live respiratory syncytial virus is blocked in seropositive cotton rats. Infect. Immun. 37, 1074–1078.
Prince, G. A., Horswood, R. L., and Chanock, R. M. (1985). Quantitative aspects of passive immunity to respiratory syncytial virus infection in infant cotton rats. J. Virol. 55, 517–520.
Prince, G. A., Jenson, A., Hemming, V., Murphy, B., Walsh, E., Horswood, R., et al. (1986). Enhancement of respiratory syncytial virus pulmonary pathology in cotton rats by prior intramuscular inoculation of formalin-inactiva ted virus. J. Virol. 57, 721–728.
Prince, G. A., Jenson, A. B., Horswood, R. L., Camargo, E., and Chanock, R. M. (1978). The pathogenesis of respiratory syncytial virus infection in cotton rats. Am. J. Pathol. 93, 771–791.
Prince, G. A., Mathews, A., Curtis, S. J., and Porter, D. D. (2000). Treatment of respiratory syncytial virus bronchiolitis and pneumonia in a cotton rat model with systemically administered monoclonal antibody (palivizumab) and glucocorticosteroid. J. Infect. Dis. 182, 1326–1330. doi: 10.1086/315894
Prince, G. A., and Porter, D. D. (1976). The pathogenesis of respiratory syncytial virus infection in infant ferrets. Am. J. Pathol. 82, 339–352.
Prince, G. A., Suffin, S. C., Prevar, D. A., Camargo, E., Sly, D. L., London, W. T., et al. (1979). Respiratory syncytial virus infection in owl monkeys: viral shedding, immunological response, and associated illness caused by wild-type virus and two temperature-sensitive mutants. Infect. Immun. 26, 1009–1013.
Rebelatto, M. C., Mead, C., and HogenEsch, H. (2000). Lymphocyte populations and adhesion molecule expression in bovine tonsils. Vet. Immunol. Immunopathol. 73, 15–29. doi: 10.1016/S0165-2427(99)00149-X
Renshaw, R., Laverack, M., Zylich, N., Glaser, A., and Dubovi, E. (2011). Genomic analysis of a pneumovirus isolated from dogs with acute respiratory disease. Vet. Microbiol. 150, 88–95. doi: 10.1016/j.vetmic.2011.01.013
Richardson, L. S., Belshe, R. B., London, W. T., Sly, D. L., Prevar, D. A., Camargo, E., et al. (1978). Evaluation of five temperature-sensitive mutants of respiratory syncytial virus in primates: I. Viral shedding, immunologic response, and associated illness. J. Med. Virol. 3, 91–100. doi: 10.1002/jmv.1890030202
Richardson, J. Y., Ottolini, M. G., Pletneva, L., Boukhvalova, M., Zhang, S., Vogel, S. N., et al. (2005). Respiratory syncytial virus (RSV) infection induces cyclooxygenase 2: a potential target for RSV therapy. J. Immunol. 174, 4356–4364. doi: 10.4049/jimmunol.174.7.4356
Riffault, S., Meyer, G., Deplanche, M., Dubuquoy, C., Durand, G., Soulestin, M., et al. (2010). A new subunit vaccine based on nucleoprotein nanoparticles confers partial clinical and virological protection in calves against bovine respiratory syncytial virus. Vaccine 28, 3722–3734. doi: 10.1016/j.vaccine.2010.03.008
Rodriguez, W. J., Gruber, W. C., Welliver, R. C., Groothuis, J. R., Simoes, E. A., Meissner, H. C., et al. (1997). Respiratory syncytial virus (RSV) immune globulin intravenous therapy for RSV lower respiratory tract infection in infants and young children at high risk for severe RSV infections. Pediatrics 99, 454–461. doi: 10.1542/peds.99.3.454
Rostad, C. A., Stobart, C. C., Gilbert, B. E., Pickles, R. J., Hotard, A. L., Meng, J., et al. (2016). A recombinant respiratory syncytial virus vaccine candidate attenuated by a low-fusion f protein is immunogenic and protective against challenge in cotton rats. J. Virol. 90, 7508–7518. doi: 10.1128/JVI.00012-16
Sacco, R. E., Durbin, R. K., and Durbin, J. E. (2015). Animal models of respiratory syncytial virus infection and disease. Curr. Opin. Virol. 13, 117–122. doi: 10.1016/j.coviro.2015.06.003
Sacco, R. E., McGill, J. L., Palmer, M. V., Lippolis, J. D., Reinhardt, T. A., and Nonnecke, B. J. (2012a). Neonatal calf infection with respiratory syncytial virus: drawing parallels to the disease in human infants. Viruses 4, 3731–3753. doi: 10.3390/v4123731
Sacco, R. E., Nonnecke, B. J., Palmer, M. V., Waters, W. R., Lippolis, J. D., and Reinhardt, T. A. (2012b). Differential expression of cytokines in response to respiratory syncytial virus infection of calves with high or low circulating 25-hydroxyvitamin D3. PLoS One 7:e33074. doi: 10.1371/journal.pone.0033074
Saez-Llorens, X., Castano, E., Null, D., Steichen, J., Sanchez, P. J., Ramilo, O., et al. (1998). Safety and pharmacokinetics of an intramuscular humanized monoclonal antibody to respiratory syncytial virus in premature infants and infants with bronchopulmonary dysplasia. The MEDI-493 Study Group. Pediatr. Infect. Dis. J. 17, 787–791. doi: 10.1097/00006454-199809000-00007
Salazar, G., Zhang, N., Fu, T.-M., and An, Z. (2017). Antibody therapies for the prevention and treatment of viral infections. NPJ Vaccines 2:19. doi: 10.1038/s41541-017-0019-3
Sawada, A., and Nakayama, T. (2016). Experimental animal model for analyzing immunobiological responses following vaccination with formalin-inactivated respiratory syncytial virus. Microbiol. Immunol. 60, 234–242. doi: 10.1111/1348-0421.12365
Scheerlinck, J. P., Snibson, K. J., Bowles, V. M., and Sutton, P. (2008). Biomedical applications of sheep models: from asthma to vaccines. Trends Biotechnol. 26, 259–266. doi: 10.1016/j.tibtech.2008.02.002
Scheltema, N. M., Kavelaars, X. M., Thorburn, K., Hennus, M. P., van Woensel, J. B., van der Ent, C. K., et al. (2018). Potential impact of maternal vaccination on life-threatening respiratory syncytial virus infection during infancy. Vaccine. 36, 4693–4700. doi: 10.1016/j.vaccine.2018.06.021
Sharma, A., Wu, W., Sung, B., Huang, J., Tsao, T., Li, X., et al. (2016). Respiratory syncytial virus (RSV) pulmonary infection in humanized mice induces human anti-RSV immune responses and pathology. J. Virol. 90, 5068–5074. doi: 10.1128/JVI.00259-16
Shaw, C. A., Galarneau, J.-R., Bowenkamp, K. E., Swanson, K. A., Palmer, G. A., Palladino, G., et al. (2013). The role of non-viral antigens in the cotton rat model of respiratory syncytial virus vaccine-enhanced disease. Vaccine 31, 306–312. doi: 10.1016/j.vaccine.2012.11.006
Shi, T., McAllister, D. A., O'Brien, K. L., Simoes, E. A. F., Madhi, S. A., Gessner, B. D., et al. (2017). Global, regional, and national disease burden estimates of acute lower respiratory infections due to respiratory syncytial virus in young children in 2015: a systematic review and modelling study. Lancet 390, 946–958. doi: 10.1016/S0140-6736(17)30938-8
Shrivastava, P., Atanley, E., Sarkar, I., Watkiss, E., Gomis, S., and van Drunen Littel-van den Hurk, S. (2015). Blunted inflammatory and mucosal IgA responses to pneumonia virus of mice in C57BL/6 neonates are correlated to reduced protective immunity upon re-infection as elderly mice. Virology 485, 233–243. doi: 10.1016/j.virol.2015.07.019
Simoes, E. A. (2003). Environmental and demographic risk factors for respiratory syncytial virus lower respiratory tract disease. J. Pediatr. 143, 118–126. doi: 10.1067/S0022-3476(03)00511-0
Simoes, E. A., Hayward, A. R., Ponnuraj, E. M., Straumanis, J. P., Stenmark, K. R., Wilson, H. L., et al. (1999). Respiratory syncytial virus infects the Bonnet monkey, Macaca radiata. Pediatr. Dev. Pathol. 2, 316–326.
Simon, A., Gehrmann, S., Wagenpfeil, G., and Wagenpfeil, S. (2018). Use of palivizumab in Germany – report from the German Synagis™ Registry 2009–2016. Klin. Padiatr. doi: 10.1055/a-0595-7771
Snoeck, C. J., Ponghsavath, V., Luetteke, N., Kaufmann, S., Sausy, A., Samountry, B., et al. (2018). Etiology of viral respiratory infections in Northern Lao PDR. J. Med. Virol. 90, 1553–1558. doi: 10.1002/jmv.25237
Sow, F. B., Gallup, J. M., Olivier, A., Krishnan, S., Patera, A. C., Suzich, J., et al. (2011). Respiratory syncytial virus is associated with an inflammatory response in lungs and architectural remodeling of lung-draining lymph nodes of newborn lambs. Am. J. Physiol. Lung Cell. Mol. Physiol. 300, L12–L24. doi: 10.1152/ajplung.00169.2010
Staeheli, P., Pitossi, F., and Pavlovic, J. (1993). Mx proteins: GTPases with antiviral activity. Trends Cell Biol. 3, 268–272. doi: 10.1016/0962-8924(93)90055-6
Steff, A. M., Monroe, J., Friedrich, K., Chandramouli, S., Nguyen, T. L., Tian, S., et al. (2017). Pre-fusion RSV F strongly boosts pre-fusion specific neutralizing responses in cattle pre-exposed to bovine RSV. Nat. Commun. 8:1085. doi: 10.1038/s41467-017-01092-4
Stittelaar, K. J., de Waal, L., van Amerongen, G., Veldhuis Kroeze, E. J., Fraaij, P. L., van Baalen, C. A., et al. (2016). Ferrets as a novel animal model for studying human respiratory syncytial virus infections in immunocompetent and immunocompromised hosts. Viruses 8:168. doi: 10.3390/v8060168
Stobart, C. C., Rostad, C. A., Ke, Z., Dillard, R. S., Hampton, C. M., Strauss, J. D., et al. (2016). A live RSV vaccine with engineered thermostability is immunogenic in cotton rats despite high attenuation. Nat. Commun. 7:13916. doi: 10.1038/ncomms13916
Szentiks, C. A., Köndgen, S., Silinski, S., Speck, S., and Leendertz, F. H. (2009). Lethal pneumonia in a captive juvenile chimpanzee (Pan troglodytes) due to human-transmitted human respiratory syncytial virus (HRSV) and infection with Streptococcus pneumoniae. J. Med. Primatol. 38, 236–240. doi: 10.1111/j.1600-0684.2009.00346.x
Tang, R. S., MacPhail, M., Schickli, J. H., Kaur, J., Robinson, C. L., Lawlor, H. A., et al. (2004). Parainfluenza virus type 3 expressing the native or soluble fusion (F) protein of respiratory syncytial virus (RSV) confers protection from RSV infection in African green monkeys. J. Virol. 78, 11198–11207. doi: 10.1128/JVI.78.20.11198-11207.2004
Tardif, S. D., Coleman, K., Hobbs, T. R., and Lutz, C. (2013). IACUC review of nonhuman primate research. ILAR J. 54, 234–245. doi: 10.1093/ilar/ilt040
Taylor, G. (2013). “Bovine model of respiratory syncytial virus infection” in Challenges and opportunities for respiratory syncytial virus vaccines. eds. L. J. Anderson, and B. S. Graham (Berlin, Heidelberg: Springer), 327–345.
Taylor, G. (2017). Animal models of respiratory syncytial virus infection. Vaccine 35, 469–480. doi: 10.1016/j.vaccine.2016.11.054
Taylor, G., Thom, M., Capone, S., Pierantoni, A., Guzman, E., Herbert, R., et al. (2015). Efficacy of a virus-vectored vaccine against human and bovine respiratory syncytial virus infections. Sci. Transl. Med. 7:300ra127. doi: 10.1126/scitranslmed.aac5757
Taylor, G., Thomas, L. H., and Stott, E. J. (1989). Effect of vaccination on cell populations in lung washes from calves after infection with respiratory syncytial virus. Res. Vet. Sci. 47, 231–235. doi: 10.1016/S0034-5288(18)31211-6
Taylor, G., Thomas, L. H., Wyld, S. G., Furze, J., Sopp, P., and Howard, C. J. (1995). Role of T-lymphocyte subsets in recovery from respiratory syncytial virus infection in calves. J. Virol. 69, 6658–6664.
Taylor, G., Wyld, S., Valarcher, J. F., Guzman, E., Thom, M., Widdison, S., et al. (2014). Recombinant bovine respiratory syncytial virus with deletion of the SH gene induces increased apoptosis and pro-inflammatory cytokines in vitro, and is attenuated and induces protective immunity in calves. J. Gen. Virol. 95, 1244–1254. doi: 10.1099/vir.0.064931-0
Teng, M. N., Whitehead, S. S., Bermingham, A., Claire, M. S., Elkins, W. R., Murphy, B. R., et al. (2000). Recombinant respiratory syncytial virus that does not express the NS1 or M2-2 protein is highly attenuated and immunogenic in chimpanzees. J. Virol. 74, 9317–9321. doi: 10.1128/JVI.74.19.9317-9321.2000
Thomas, L. H., Stott, E. J., Collins, A. P., Crouch, S., and Jebbett, J. (1984). Infection of gnotobiotic calves with a bovine and human isolate of respiratory syncytial virus. Modification of the response by dexamethasone. Arch. Virol. 79, 67–77. doi: 10.1007/BF01314304
Unwin, S., Chatterton, J., and Chantrey, J. (2013). Management of severe respiratory tract disease caused by human respiratory syncytial virus and Streptococcus pneumoniae in captive chimpanzees (Pan troglodytes). J. Zoo Wildl. Med. 44, 105–115. doi: 10.1638/1042-7260-44.1.105
Valarcher, J. F., and Taylor, G. (2007). Bovine respiratory syncytial virus infection. Vet. Res. 38, 153–180. doi: 10.1051/vetres:2006053
van der Poel, W. H. M., Schrijver, R. S., Middel, W. G. J., Kramps, J. A., Brand, A., and Van Oirschot, J. T. (1996). Experimental reproduction of respiratory disease in calves with non-cell-culture-passaged bovine respiratory syncytial virus. Vet. Q. 18, 81–86. doi: 10.1080/01652176.1996.9694622
van der Sluijs, M. T. W., Kuhn, E. M., and Makoschey, B. (2010). A single vaccination with an inactivated bovine respiratory syncytial virus vaccine primes the cellular immune response in calves with maternal antibody. BMC Vet. Res. 6, 2–2. doi: 10.1186/1746-6148-6-2
Velin, D., Fotopoulos, G., Luthi, F., and Kraehenbuhl, J.-P. (1997). The nasal-associated lymphoid tissue of adult mice acts as an entry site for the mouse mammary tumor retrovirus. J. Exp. Med. 185, 1871–1876. doi: 10.1084/jem.185.10.1871
Viuff, B., Tjornehoj, K., Larsen, L. E., Rontved, C. M., Uttenthal, A., Ronsholt, L., et al. (2002). Replication and clearance of respiratory syncytial virus: apoptosis is an important pathway of virus clearance after experimental infection with bovine respiratory syncytial virus. Am. J. Pathol. 161, 2195–2207. doi: 10.1016/s0002-9440(10)64496-3
Walsh, K. B., Teijaro, J. R., Brock, L. G., Fremgen, D. M., Collins, P. L., Rosen, H., et al. (2014). Animal model of respiratory syncytial virus: CD8+ T cells cause a cytokine storm that is chemically tractable by sphingosine-1-phosphate 1 receptor agonist therapy. J. Virol. 88, 6281–6293. doi: 10.1128/jvi.00464-14
Wang, D., Phan, S., DiStefano, D. J., Citron, M. P., Callahan, C. L., Indrawati, L., et al. (2017). A single dose recombinant PIV5 expressing respiratory syncytial virus F and G proteins protected cotton rats and African green monkeys from RSV challenge. J. Virol. 91:e00066–17. doi: 10.1128/JVI.00066-17
Watkiss, E. R., Shrivastava, P., Arsic, N., Gomis, S., and van Drunen Littel-van den Hurk, S. (2013). Innate and adaptive immune response to pneumonia virus of mice in a resistant and a susceptible mouse strain. Viruses 5, 295–320. doi: 10.3390/v5010295
Werling, D., Koss, M., Howard, C. J., Taylor, G., Langhans, W., and Hope, J. C. (2002). Role of bovine chemokines produced by dendritic cells in respiratory syncytial virus-induced T cell proliferation. Vet. Immunol. Immunopathol. 87, 225–233. doi: 10.1016/S0165-2427(02)00086-7
West, K., Petrie, L., Haines, D. M., Konoby, C., Clark, E. G., Martin, K., et al. (1999). The effect of formalin-inactivated vaccine on respiratory disease associated with bovine respiratory syncytial virus infection in calves. Vaccine 17, 809–820. doi: 10.1016/S0264-410X(98)00265-5
Widjaja, I., Wicht, O., Luytjes, W., Leenhouts, K., Rottier, P. J. M., van Kuppeveld, F. J. M., et al. (2016). Characterization of epitope-specific anti-respiratory syncytial virus (anti-RSV) antibody responses after natural infection and after vaccination with formalin-inactivated RSV. J. Virol. 90, 5965–5977. doi: 10.1128/JVI.00235-16
Widjojoatmodjo, M. N., Boes, J., van Bers, M., van Remmerden, Y., Roholl, P. J., and Luytjes, W. (2010). A highly attenuated recombinant human respiratory syncytial virus lacking the G protein induces long-lasting protection in cotton rats. Virol. J. 7:114. doi: 10.1186/1743-422X-7-114
Widjojoatmodjo, M. N., Bogaert, L., Meek, B., Zahn, R., Vellinga, J., Custers, J., et al. (2015). Recombinant low-seroprevalent adenoviral vectors Ad26 and Ad35 expressing the respiratory syncytial virus (RSV) fusion protein induce protective immunity against RSV infection in cotton rats. Vaccine 33, 5406–5414. doi: 10.1016/j.vaccine.2015.08.056
Wiegand, M. A., Gori-Savellini, G., Gandolfo, C., Papa, G., Kaufmann, C., Felder, E., et al. (2017). A respiratory syncytial virus vaccine vectored by a stable chimeric and replication-deficient sendai virus protects mice without inducing enhanced disease. J. Virol. 91, e02298–e02216. doi: 10.1128/JVI.02298-16
Wong, T. M., Boyapalle, S., Sampayo, V., Nguyen, H. D., Bedi, R., Kamath, S. G., et al. (2014). Respiratory syncytial virus (RSV) infection in elderly mice results in altered antiviral gene expression and enhanced pathology. PLoS One 9, e88764–e88764. doi: 10.1371/journal.pone.0088764
Woolums, A. R. L., Anderson, M., Gunther, R., Schelegle, E., Danielle, R., Singer, R., et al. (1999). Evaluation of severe disease induced by aerosol inoculation of calves with bovine respiratory syncytial virus. Am. J. Vet. Res. 60, 473–480.
Woolums, A. R., Berghaus, R. D., Berghaus, L. J., Ellis, R. W., Pence, M. E., Saliki, J. T., et al. (2013). Effect of calf age and administration route of initial multivalent modified-live virus vaccine on humoral and cell-mediated immune responses following subsequent administration of a booster vaccination at weaning in beef calves. Am. J. Vet. Res. 74, 343–354. doi: 10.2460/ajvr.74.2.343
Woolums, A. R., Brown, C. C., Brown, J. C. Jr., Cole, D. J., Scott, M. A., Williams, S. M., et al. (2004). Effects of a single intranasal dose of modified-live bovine respiratory syncytial virus vaccine on resistance to subsequent viral challenge in calves. Am. J. Vet. Res. 65, 363–372. doi: 10.2460/ajvr.2004.65.363
Wright, P. F., Woodend, W. G., and Chanock, R. M. (1970). Temperature-sensitive mutants of respiratory syncytial virus: in-vivo studies in hamsters. J. Infect. Dis. 122, 501–512. doi: 10.1093/infdis/122.6.501
Yamaji, Y., Yasui, Y., and Nakayama, T. (2016). Development of acquired immunity following repeated respiratory syncytial virus infections in cotton rats. PLoS One 11:e0155777. doi: 10.1371/journal.pone.0155777
Zang, N., Zhuang, J., Deng, Y., Yang, Z., Ye, Z., Xie, X., et al. (2015). Pulmonary C fibers modulate MMP-12 production via PAR2 and are involved in the long-term airway inflammation and airway Hyperresponsiveness induced by respiratory Syncytial virus infection. J. Virol. 90, 2536–2543. doi: 10.1007/s12013-015-0682-9
Keywords: human respiratory syncytial virus, bovine respiratory syncytial virus, lower respiratory tract infections, rodent model, non-human primate model
Citation: Altamirano-Lagos MJ, Díaz FE, Mansilla MA, Rivera-Pérez D, Soto D, McGill JL, Vasquez AE and Kalergis AM (2019) Current Animal Models for Understanding the Pathology Caused by the Respiratory Syncytial Virus. Front. Microbiol. 10:873. doi: 10.3389/fmicb.2019.00873
Edited by:
Hirokazu Kimura, Gunma Paz University, JapanReviewed by:
Geraldine Taylor, Pirbright Institute (BBSRC), United KingdomCopyright © 2019 Altamirano-Lagos, Díaz, Mansilla, Rivera-Pérez, Soto, McGill, Vasquez and Kalergis. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alexis M. Kalergis, YWthbGVyZ2lzQGJpby5wdWMuY2w=; YWthbGVyZ2lzQGljbG91ZC5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.