 Jun-Gyu Park1
Jun-Gyu Park1 Ginés Ávila-Pérez1
Ginés Ávila-Pérez1 Ferralita Madere1
Ferralita Madere1 Thomas A. Hilimire1
Thomas A. Hilimire1 Aitor Nogales2
Aitor Nogales2 Fernando Almazán3
Fernando Almazán3 Luis Martínez-Sobrido1*
Luis Martínez-Sobrido1*- 1Department of Microbiology and Immunology, University of Rochester Medical Center, Rochester, NY, United States
- 2Center for Animal Health Research, INIA–CISA, Madrid, Spain
- 3Department of Molecular and Cell Biology, Centro Nacional de Biotecnología (CNB–CSIC), Campus Universidad Autónoma de Madrid, Cantoblanco, Madrid, Spain
Zika virus (ZIKV) is one of the recently emerging vector-borne viruses in humans and is responsible for severe congenital abnormalities such as microcephaly in the Western Hemisphere. Currently, only a few vaccine candidates and therapeutic drugs are being developed for the treatment of ZIKV infections, and as of yet none are commercially available. The polyanionic aromatic compound aurintricarboxylic acid (ATA) has been shown to have a broad-spectrum antimicrobial and antiviral activity. In this study, we evaluated ATA as a potential antiviral drug against ZIKV replication. The antiviral activity of ATA against ZIKV replication in vitro showed median inhibitory concentrations (IC50) of 13.87 ± 1.09 μM and 33.33 ± 1.13 μM in Vero and A549 cells, respectively; without showing any cytotoxic effect in both cell lines (median cytotoxic concentration (CC50) > 1,000 μM). Moreover, ATA protected both cell types from ZIKV-induced cytopathic effect (CPE) and apoptosis in a time- and concentration-dependent manner. In addition, pre-treatment of Vero cells with ATA for up to 72 h also resulted in effective suppression of ZIKV replication with similar IC50. Importantly, the inhibitory effect of ATA on ZIKV infection was effective against strains of the African and Asian/American lineages, indicating that this inhibitory effect was not strain dependent. Overall, these results demonstrate that ATA has potent inhibitory activity against ZIKV replication and may be considered as a potential anti-ZIKV therapy for future clinical evaluation.
Introduction
Zika virus (ZIKV) belongs to the genus Flavivirus within the Flaviviridae family. ZIKV is an enveloped positive sense single-stranded RNA virus with a genome size of ∼10.7 kb that encodes a single polyprotein, which is post-translationally processed by cellular and viral proteases into three structural (capsid, C; pre-membrane, prM; and envelope, E) and seven non-structural (NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5) proteins (Tripathi et al., 2017; Avila-Perez et al., 2018).
Zika virus was initially isolated from Uganda in 1947 and viral infections only occurred sporadically in Africa and Asia until 2007. ZIKV appeared explosively as the first large-scale outbreak occurred in the Yap island in 2007 and French Polynesia in 2013 (Weaver et al., 2016). Most recently, in 2015, the first local transmission of ZIKV was found in territories of Latin America and the Caribbean, resulting in up to 1.3 million of ZIKV infection suspected cases (Tang et al., 2016; Tripathi et al., 2017).
Like other members of the Flaviviridae family, such as yellow fever virus (YFV), Dengue virus (DENV), Japanese encephalitis virus (JEV), and West Nile virus (WNV), ZIKV is commonly transmitted by the bite of infected Aedes mosquitos, but it can also be transmitted vertically from mother to child, through sexual contact, and in rare cases from blood transfusions (Lessler et al., 2016; Fink et al., 2018). Upon infection, ZIKV can be shed in blood, urine, semen, saliva, amniotic fluid, breast milk, and cerebrospinal fluid (Nayak et al., 2016; Colt et al., 2017; Nazerai et al., 2019). Most people (75∼80%) infected with ZIKV are asymptomatic or have mild symptoms such as fever, rash, joint pain, and conjunctivitis that can last for several days to a week (Fink et al., 2018). In rare cases, people with symptoms may have neurological Guillain-Barré syndrome complications (Oehler et al., 2014; Rivera-Concepcion et al., 2018; Nazerai et al., 2019). In the case of pregnant women, ZIKV infection can lead to microcephaly and other fetal complications as occurred during the large-scale ZIKV outbreak in Brazil in 2015 (Lessler et al., 2016). Because of the significant outbreaks in South, Central, and North America, ZIKV was declared a Public Health concern by the World Health Organization (WHO) in February 2016 (Lazear and Diamond, 2016; Ramos da Silva and Gao, 2016; Weaver et al., 2016; Tripathi et al., 2017).
There are several vaccines and antiviral drugs currently under development for the prevention or treatment of ZIKV infection (Abbink et al., 2016; Larocca et al., 2016; Shan et al., 2017; Fink et al., 2018). DNA-based (Abbink et al., 2016; Larocca et al., 2016), inactivated (Abbink et al., 2016; Larocca et al., 2016; Shan et al., 2017), live-attenuated and mRNA (Richner et al., 2017) vaccines have been proposed for the prophylactic treatment of ZIKV infections. On the other hand, arbidol (ARB) (Fink et al., 2018; Haviernik et al., 2018), bortezomib, mycophenolic acid, daptomycin (Barrows et al., 2016), obatoclax, saliphenylhalamide, gemcitabine (Kuivanen et al., 2017), emetine (Yang et al., 2018), and sofosbuvir (Bullard-Feibelman et al., 2017) have been proposed for the therapeutic treatment of ZIKV infection. Despite these tremendous efforts, there is currently no Food and Drug Administration (FDA)-approved vaccines and/or anti-viral drugs available for the treatment of ZIKV infection. Since vaccination takes at least 2 weeks to several months to show protective effects against ZIKV infection, vaccination is probably not the most appropriate prophylactic method for those who are traveling to areas where ZIKV is epidemic, endemic, or have already been infected. Moreover, vaccination may cause an important issue, such as antibody-dependent enhancement (ADE) (Bardina et al., 2017; Priyamvada et al., 2017). ADE, which has been extensively described in DENV (Priyamvada et al., 2017), is a phenomenon where preexisting antibodies facilitate binding and infection during subsequent exposure to infectious viruses, instead of neutralizing them, resulting in exacerbation of clinical signs (Bardina et al., 2017; Priyamvada et al., 2017). Because of the structural similarities between DENV and ZIKV, DENV immunity–linked ADE of ZIKV infection has also been reported (Bardina et al., 2017; Priyamvada et al., 2017). Since vaccination for ZIKV could lead to DENV ADE, antivirals could represent a better choice for the control of ZIKV infection.
Aurintricarboxylic acid (ATA), a polyanionic aromatic compound, has been shown to have inhibitory properties against several bacteria and viruses including, among others, Yersinia pestis (Liang et al., 2003), Cryptosporidium parvum (Klein et al., 2008), human immunodeficient virus (HIV) (Mitra et al., 1996; De Clercq, 2005), hepatitis C virus (HCV) (Chen et al., 2009; Mukherjee et al., 2012; Shadrick et al., 2013), Vaccinia virus (Myskiw et al., 2007), influenza virus (Hung et al., 2009), Enterovirus 71 (Hung et al., 2010) and severe acute respiratory syndrome coronaviruses (SARS-CoV) (He et al., 2004). Mechanistic studies have suggested that ATA has the ability to modulate various cellular enzymes such as activators of the Janus kinase 2 (JAK2) and signal transducer and activator of transcription 5 (STAT5) families (Rui et al., 1998), inhibitors of nucleases (Shadrick et al., 2013), glucose-6-phosphate dehydrogenase (Bina-Stein and Tritton, 1976), and topoisomerase II proteins (Catchpoole and Stewart, 1994; Benchokroun et al., 1995) as well as the enzymatic activity of the Vaccinia virus AH1L phosphatase (Smee et al., 2010). However, to date, the ability of ATA to inhibit ZIKV infection has not been evaluated. Herein, we investigated ATA as a plausible prophylactic and therapeutic candidate against ZIKV infection. Our results demonstrate that ATA has a potent and effective antiviral activity against ZIKV in pre- and post-infection settings, including broadly antiviral activity against strains of the African and American/Asian lineages with no toxicity up to 1,000 μM in cultured cells. These data support the feasibility of implementing ATA for the treatment of ZIKV infection.
Materials and Methods
Cell Lines and Viruses
African green monkey kidney epithelial Vero (ATCC CCL-81) and human adenocarcinoma alveolar basal epithelial A549 (ATCC CCL-185) cells were maintained in Dulbecco’s modified Eagle’s medium (DMEM; Mediatech, Inc.) supplemented with 5% fetal bovine serum (FBS) and 1% PSG (100 U/ml penicillin, 100 μg/ml streptomycin, and 2 mM L-glutamine) at 37°C in a 5% CO2 atmosphere.
Paraiba/2015 ZIKV isolate was kindly provided by Stephen Dewhurst (Department of Microbiology and Immunology, University of Rochester). Uganda/1947 (MR_766 strain, Catalog No. NR-50065) and Nigeria/1968 (IbH 30656 strain, Catalog No. NR-50066) ZIKV isolates were obtained from the Biodefense and Emerging Infections Research Resources Repository (BEI Resources). Puerto Rico/2015 (PRVABC59 strain) and French Polynesia/2013 ZIKV isolates were kindly provided from the Centers for Disease Control and Prevention (CDC). Virus stocks were propagated in Vero cells and titrated by plaque assay as previously described (Marquez-Jurado et al., 2018).
Compounds
Aurintricarboxylic acid (Catalog No. A1895) and Arbidol (ARB, Catalog No. SLM0860) were purchased from Sigma-Aldrich, MO, United States. Both compounds were prepared at 100 mM stock solution dissolved in dimethyl sulfoxide (DMSO) and kept at -20°C until experimental use. Each drug was diluted into infectious media (DMEM 2% FBS, 1% PSG) for the described experiments, where the maximum DMSO concentration was 0.1%.
Cell Viability Assay
Cell viability in Vero and A549 cells was measured using the CellTiter 96 Non-Radioactive Cell Proliferation assay (Promega) following the manufacturer’s instructions. Briefly, confluent Vero or A549 cells (96-well plate format, 5 × 104 cells/well, triplicates) were treated with 100 μl of DMEM containing serially diluted (twofold dilutions, starting concentration of 1,000 μM) chemicals or 0.1% DMSO (vehicle control). Plates were incubated at 37°C in a 5% CO2 atmosphere for 36 or 72 h. Samples were treated with 15 μl of Dye Solution and incubated at 37°C in a 5% CO2 atmosphere for 4 h. Next, cells were treated with 100 μl of Solubilization Solution/Stop Mix and absorbance at 570 nm was measured using a Vmax kinetic microplate reader (Molecular Devices, Waltham, MA, United States). Viability of compound-treated cells was calculated as a percentage relative to values obtained with DMSO-treated cells. Non-linear regression curves and the median cytotoxic concentration (CC50) were calculated using GraphPad Prism software version 8.0.
Microplaque Reduction Assay and Immunostaining
Confluent monolayers (96-plate format, 5 × 104 cells/well, triplicates) of Vero cells were infected with 25 plaque forming units (PFU)/well of Paraiba/2015, Uganda/1947, Nigeria/1968, Puerto Rico/2015, and French Polynesia/2013 ZIKV strains at 37°C in infection media. After 1 h of adsorption, virus inoculum was removed and cells were washed three times with infection media before adding fresh infection media containing 1% microcrystalline cellulose (Avicel, Sigma-Aldrich) and the indicated concentration of compounds, or 0.1% DMSO as vehicle control. In case of pre-treatment experiments, the cell monolayers were treated with the indicated concentration of compound, or 0.1% DMSO, for the indicated times before ZIKV infection. Infected cells were incubated at 37°C for 36–60 h, depending on virus strains. For immunostaining, cells were fixed with 4% paraformaldehyde for 1 h, washed three times with phosphate buffered saline (PBS) and permeabilized with 0.2% Triton X-100 for 10 min at room temperature. Then, the plates were blocked with 1.25% bovine serum albumin (BSA) in PBS (blocking solution) for 1 h at room temperature, followed by incubation with 1 μg/ml of the pan-flavivirus envelop (E) protein monoclonal antibody 4G2 (ATCC, Catalog No. VR-1852) diluted in blocking solution for 1 h at 37°C. After incubation with the primary antibody, cells were washed three times with PBS and developed with the Vectastain ABC kit and the DAB Peroxidase Substrate kit (Vector Laboratory, Inc., CA, United States) according to the manufacturers’ instructions. Stained plaques were analyzed using the CTL ImmunoSpot plate reader and counting software (Cellular Technology Limited, Cleveland, OH, United States). Virus titers were calculated as PFU/ml (Nogales et al., 2015). Non-linear regression curves and the median inhibitory concentration (IC50) were determined as described above.
Virus Growth Kinetics
Confluent monolayers (24-well plate format, 2.5 × 105 cells/well, triplicates) of Vero or A549 cells were infected (multiplicity of infection, MOI, 0.1) with Paraiba/2015 diluted in infection media for 1 h at room temperature. After viral absorption, cells were incubated with infection media containing the indicated concentrations (250, 25, 2.5, and 0 μM) of ATA. At 12, 24, 48, and 72 h post-infection (h p.i.), tissue culture supernatants were collected and titrated on Vero cells by immunostaining as described previously (Marquez-Jurado et al., 2018).
Apoptosis Assay
Levels of apoptosis were measured using the Caspase-Glo® 3/7 Assay (Promega, WI, United States) following the manufacturer’s instruction. Briefly, Vero and A549 cells (24-well plate format, 2.5 × 105 cells/well, triplicates) were infected with ZIKV Paraiba/2015 (MOI of 0.1) and, at the indicated times post-infection, cells and tissue culture supernatants were collected and centrifuged. Twenty five microliters of supernatants were mixed with 25 μl of Caspase-3/7 reagent using a plate shaker, incubated at room temperature for 1 h, and luminescence at 570 nm was measured using a SpectraMax iD5 (Molecular Devices, Waltham, MA, United States) following the manufacturer’s instructions.
Statistical Analysis
Two-way ANOVA was used to evaluate significant differences. Data are expressed as the mean ± standard deviation (SD) of at least three independent experiments in triplicates using Microsoft Excel software. Value were considered statistically significant when ∗p < 0.0332, ∗∗p < 0.0021, ∗∗∗p < 0.0002, ∗∗∗∗p < 0.0001. All data were analyzed with Prism software version 8.00 (GraphPad Software, CA, United States). CC50 and IC50 were determined using sigmoidal dose response curves (GraphPad Software, CA, United States). The selective index (SI) of each compound was calculated by dividing the CC50 with the IC50.
Results
Analysis of ATA Toxicity in Vero and A549 Cells
Before examining the inhibitory effect of ATA (Figure 1) against ZIKV infection, we first determined the CC50 of ATA on Vero and A549 cells (Figure 2). For this, we treated both cell lines with serial (twofold) dilutions of ATA and measured cell viability at 36 and 72 h post-treatment. As an internal control for these studies, we used ARB, a drug that has been previously described to have antiviral activity against ZIKV in Vero (Haviernik et al., 2018) and A549 (Fink et al., 2018) cells. We did not observe any toxicity with ATA in Vero (Figure 2A) or A549 (Figure 2B) cells at 36 or 72 h post-treatment, even at the highest concentration tested (1,000 μM), while ARB showed CC50 values of 74.71 ± 1.09 or 59.37 ± 1.10 μM in Vero (Figure 2C) and 114.6 ± 1.08 or 91.0 ± 1.08 μM in A549 (Figure 2D) cells (Table 1) at 36 or 72 h post-treatment, respectively.
Figure 1. Aurintricarboxylic acid (ATA) structure. Molecular weight = 441.329 g/mol. Compound ID in PubChem: 2259.
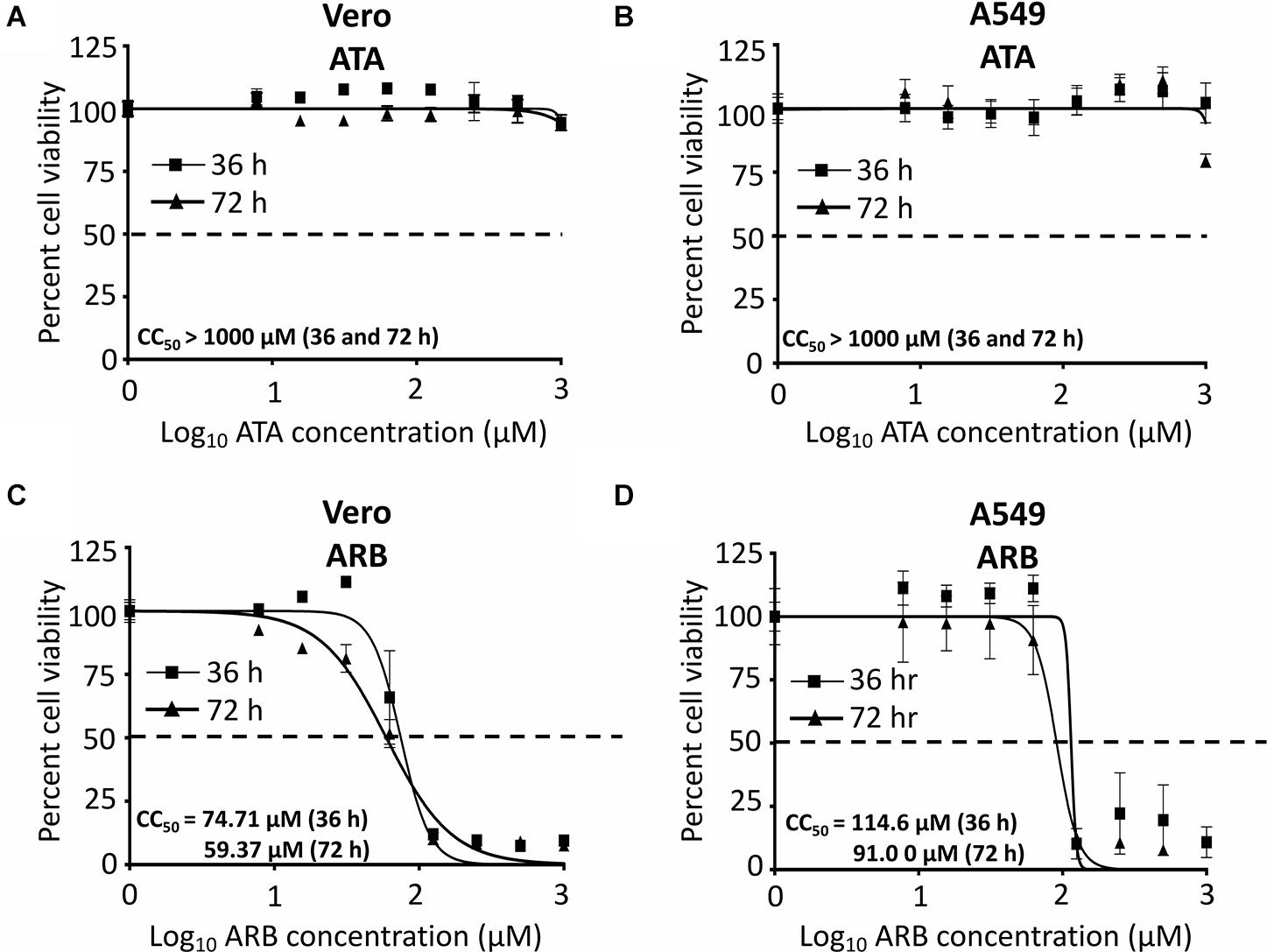
Figure 2. Cytotoxicity of ATA: Vero (A,C) or A549 (B,D) cells (96-well plate format, 5 × 104 cells/well, triplicates) were treated with the indicated doses (twofolds dilutions, starting concentration 1,000 μM) of ATA (A,B) or ARB (C,D). Cell proliferation assays were performed at 36 h (triangle) and 72 h (square) post-treatment and the CC50 for each compound was calculated at both 36 and 72 h after treatment. Dotted line indicates the 50% toxicity. Data was expressed as mean and SD from three independent experiments conducted in triplicates.
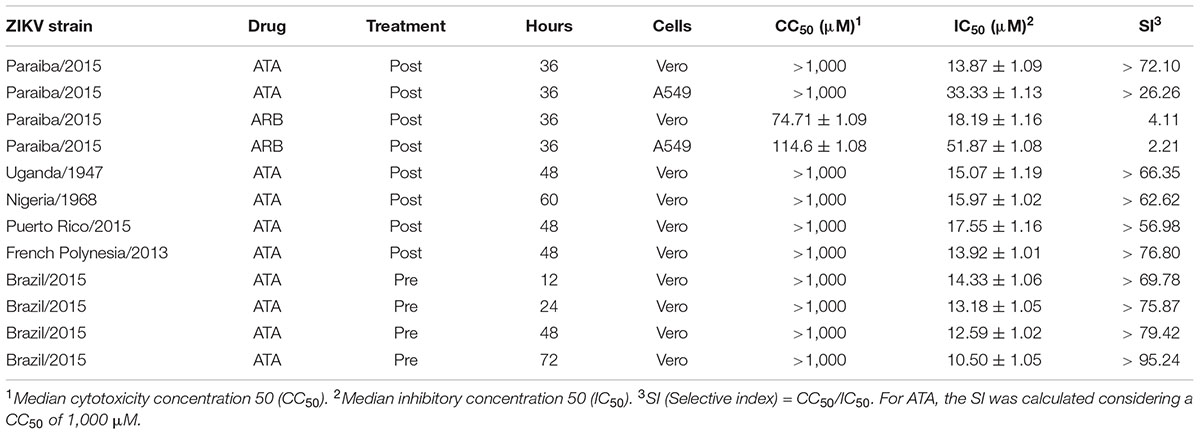
Table 1. Inhibition of ZIKV replication with ATA.
Inhibitory Effect of ATA on ZIKV Replication
To determine the IC50 of ATA, Vero and A549 cells were infected with 25 PFU/well of Paraiba/2015 and after 1 h of viral absorption, virus inoculum was replaced with infection media with twofold serial dilutions (starting concentration of 1,000 μM) of ATA or ARB (Figure 3) and the IC50 calculated as described in the Section “Materials and Methods.” Although the IC50 of ATA (Figure 3A) and ARB (Figure 3C) in Vero cells were similar (13.87 ± 1.09 μM and 18.19 ± 1.6 μM, respectively), the selective index (SI, CC50/IC50) of ATA (>72.10) was significantly higher than that of ARB (4.11) (Table 1). Likewise, the IC50 of ATA (Figure 3B) and ARB (Figure 3D) in A549 cells were similar but with clearly different SI values (>26.26 for ATA and 2.21 for ARB) (Table 1). Notably the CC50, IC50, and SI of ARB were similar to those previously described in the literature in these cell lines (Fink et al., 2018; Haviernik et al., 2018). These data suggest that ATA exhibited an effective inhibition of ZIKV infection with limited toxicity and SI values better than those previously described for ARB.
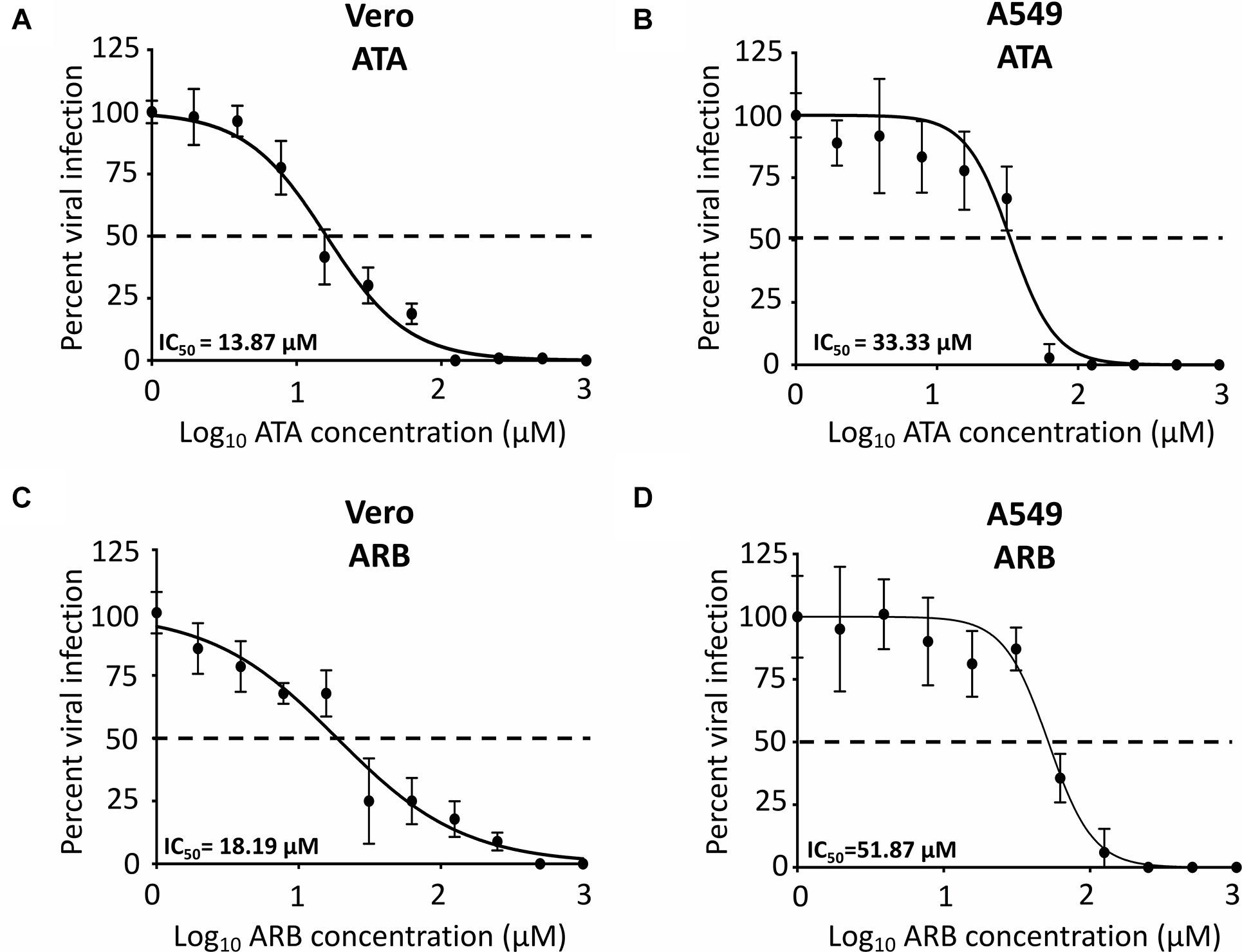
Figure 3. Inhibition of ZIKV Paraiba/2015 infection by ATA: Vero (A,C) or A549 (B,D) cells (96-well plate format, 5 × 104 cells/well, triplicates) were infected with 25 PFU of Paraiba/2015. After 1 h of viral adsorption, the indicated concentrations (twofolds dilutions, starting concentration 1,000 μM) of ATA (A,B) or ARB (C,D) were added to 100 μl of infection media containing 1% Avicel. At 36 h p.i., infected cells were fixed for virus titration by immunostaining assay. Dotted line indicates 50% inhibition. Data was expressed as mean and SD from three independent experiments conducted in triplicates.
We also observed that ZIKV replication was completely inhibited at a concentration of 250 μM of ATA in Vero (Figure 4A) and A549 (Figure 4B) cells while 2.5 μM and 25 μM concentrations of ATA showed partial viral inhibition in Vero and A549 cells (Figures 4A,B), respectively, demonstrating a dose-dependent inhibition of viral replication in both cell lines.
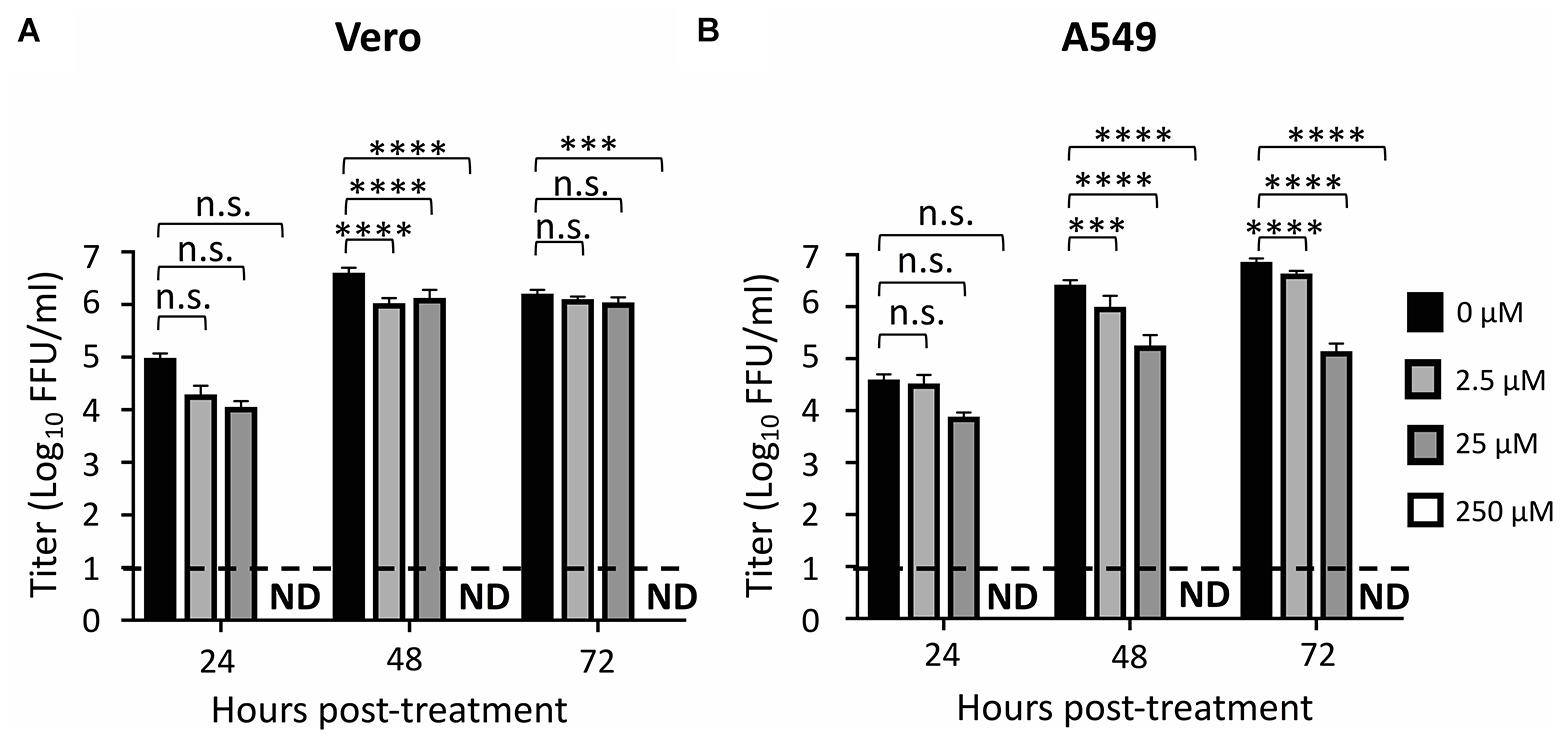
Figure 4. Aurintricarboxylic acid inhibition of ZIKV replication: Vero (A) and A549 (B) cells (24-well plate format, 2.5 × 105 cells/well, triplicates) were infected (MOI 0.1) with Paraiba/2015. Tissue culture supernatants were collected at 24, 48, and 72 h p.i., and viral titer were calculated by immunostaining (fluorescent forming units, FFU/ml). Dotted line indicates the limit of detection (20 FFU/ml). Data was expressed as mean and standard deviations (SD) from three independent experiments conducted in triplicates. Statistical analysis was conducted by two-way ANOVA, ∗p < 0.0332, ∗∗p < 0.0021, ∗∗∗p < 0.0002, ∗∗∗∗p < 0.0001, or no significance (n.s.).
ATA Protects Cells From ZIKV-Induced Cell Death
We next evaluated the ability of ATA to protect cells from the cytopathic effect (CPE) induced during ZIKV infection (Figure 5). To that end, Vero and A549 cells were infected (MOI 0.1) with Paraiba/2015 and, after 1 h of viral absorption, cells were treated with 0, 2.5, 25, and 250 μM of ATA. At 48 h p.i., cells were observed under a light microscope for evaluation of their morphology and CPE (Figure 5A). As expected from our previous results, 25 μM and more clearly 250 μM of ATA were able to prevent ZIKV-induced CPE in both cell lines (Figure 5A). To quantify the ability of ATA to prevent ZIKV-induced apoptosis, tissue culture supernatants from ZIKV-infected Vero and A549 cells were harvested at 24, 48, and 72 h p.i. to measure the level of apoptotic signal as determined by caspase 3 and 7 activities (Figure 5B). ZIKV-infected cells showed increased caspase 3 and 7 levels up to eightfolds in Vero cells and up to 4.2-folds in A549 cells compared to mock-infected cells (Figure 5B). Levels of caspase 3 and 7 activation were dose-dependently reduced by ATA with 250 μM of ATA showed only 0.4- and 1.7-fold induction as compared to mock-infected Vero and A549 cells, respectively (Figure 5B).
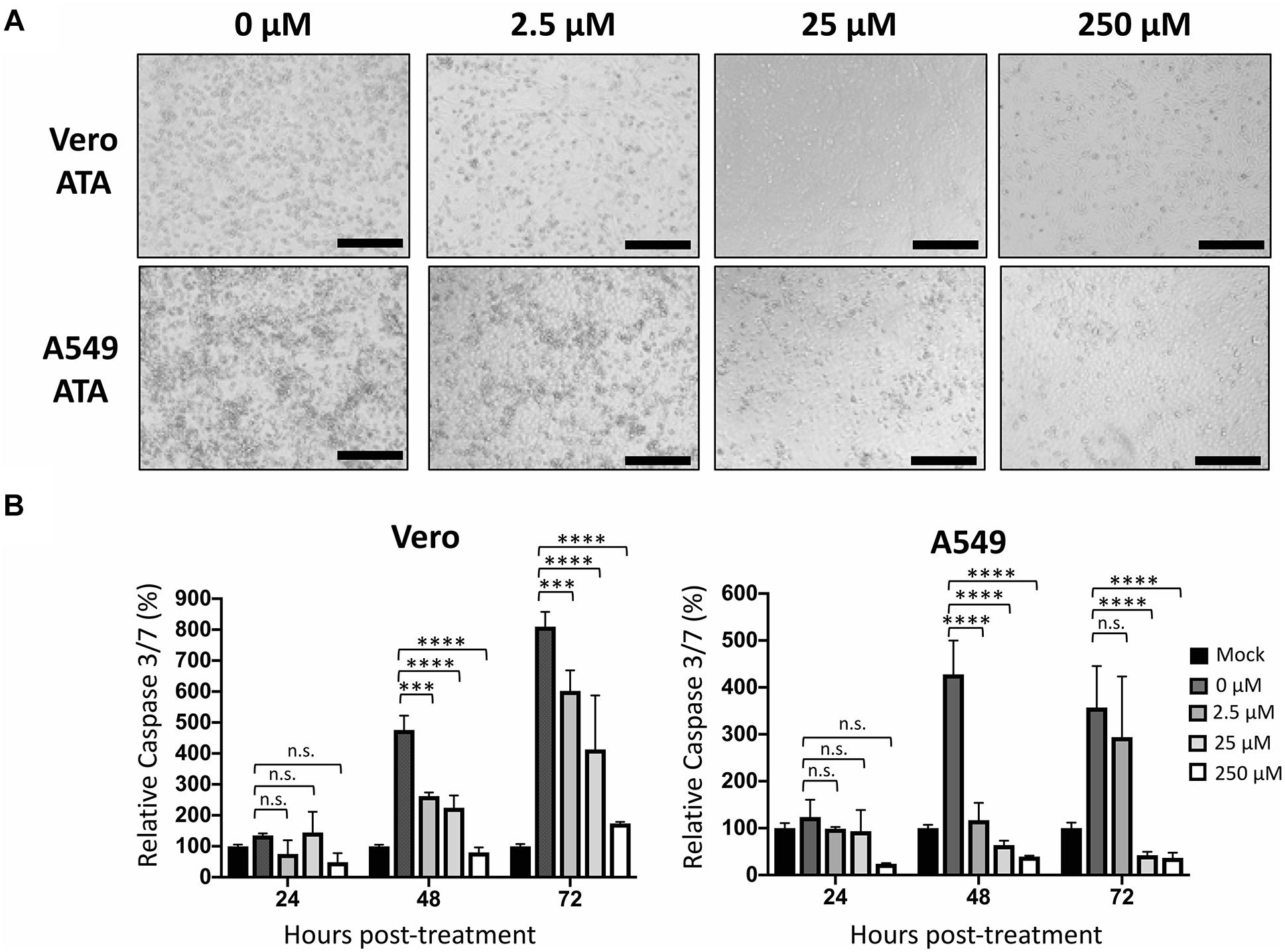
Figure 5. Aurintricarboxylic acid protects Vero and A549 cells from ZIKV-induced cell death: Vero and A549 cells (24-well plate format, 2.5 × 105 cells/well, triplicates) were infected (MOI 0.1) with Paraiba/2015. After 1 h viral adsorption, cells were treated with the indicated concentrations (250, 25, 2.5, and 0 μM) of ATA. At 48 h p.i., cells were observed and imaged under an optical microscope. Scale bar = 100 μm. (A) Caspase 3/7 levels were measured in the tissue culture supernatants at 24, 48, and 72 h p.i. (B) Data of each time point was compared to mock-infected control cells and expressed as mean of relative percentage and SD from three independent experiments conducted in triplicates. Statistical analyses were conducted by two-way ANOVA, ∗p < 0.0332, ∗∗p < 0.0021, ∗∗∗p < 0.0002, ∗∗∗∗p < 0.0001, or no significance (n.s.).
ATA Inhibition of Representative African and Asia/American ZIKV Strains
We next determined whether ATA is able to inhibit both ancestor African (Uganda/1947 and Nigeria/1968) and contemporary Asian/American (Puerto Rico/2015 and French Polynesia/2013) ZIKV lineage strains using our microplaque reduction assay (Figure 6). We observed similar IC50 values of ATA with Uganda/1947 (Figure 6A, IC50 = 15.07 ± 1.19 μM), Nigeria/1968 (Figure 6B, IC50 = 15.97 ± 1.02 μM), Puerto Rico/2015 (Figure 6C, IC50 = 17.55 ± 1.16 μM), and French Polynesia/2013 (Figure 6D, IC50 = 13.92 ± 1.01 μM), compared to those observed with Paraiba/2015 (Figure 3 and Table 1), demonstrating the broad antiviral activity of ATA against different ZIKV strains, regardless of the year and place of isolation.
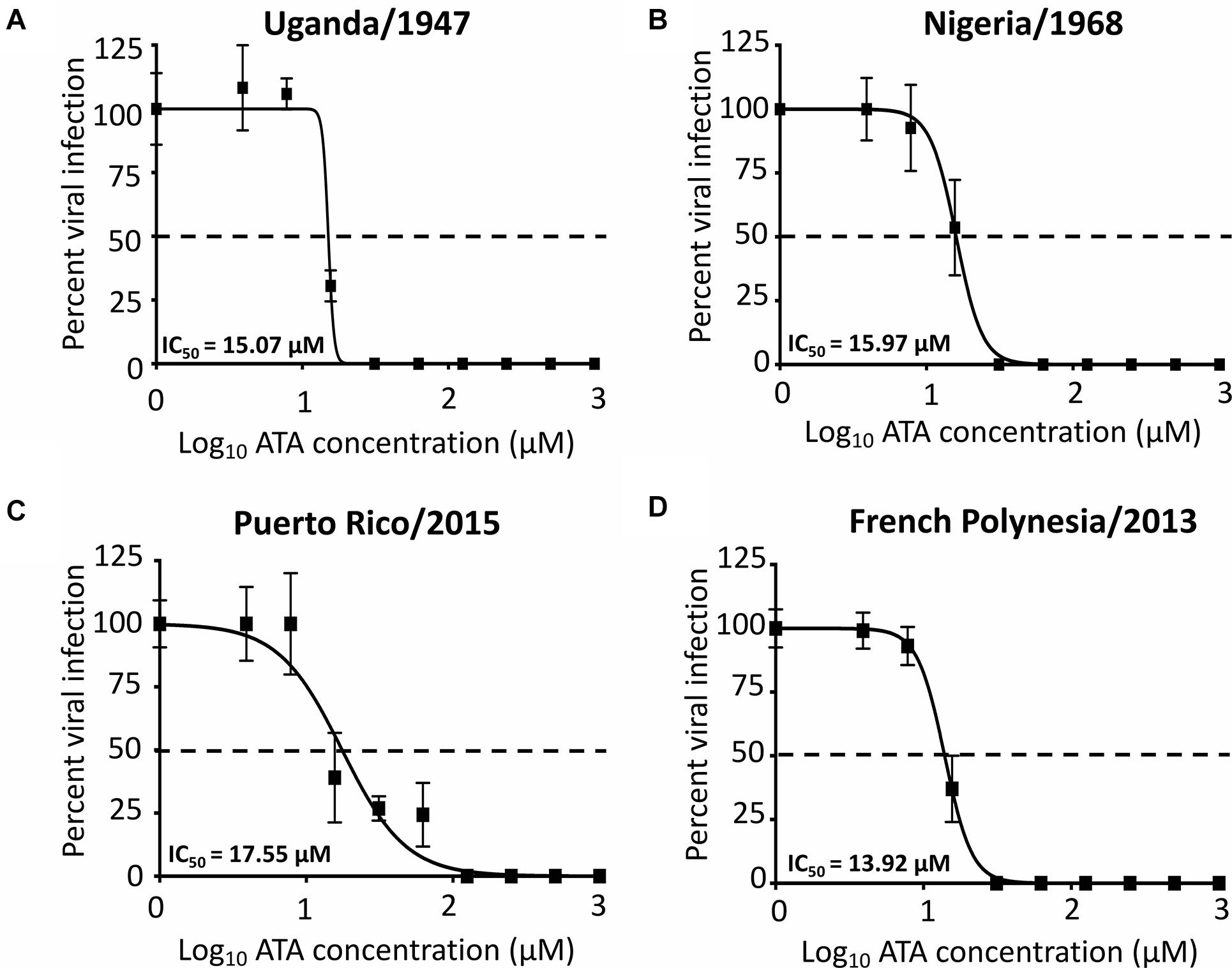
Figure 6. Aurintricarboxylic acid inhibition of African and Asian/American ZIKV strains: Vero cells (96-well plates, 5 × 104 cells/well, triplicates) were infected with 25 PFU of Uganda/1947 (A), Nigeria/1968 (B), Puerto Rico/2015 (C), and French Polynesia/2013 (D) ZIKV strains. After 1 h of viral adsorption, the indicated concentrations (twofolds dilutions, starting concentration 1,000 μM) of ATA were added to 100 μl of infection media containing 1% of Avicel. Infected cells were fixed for virus titration by immunostaining assay at 36–60 h p.i., depending on the ZIKV strains. Dotted line indicates 50% inhibition. Data was expressed as mean and SD from three independent experiments conducted in triplicates.
Pre-treatment of ATA Inhibits ZIKV Replication
To demonstrate the feasibility of using ATA for the prevention of ZIKV infection, important for travelers to regions where ZIKV is endemic, we next evaluated whether pre-treatment with ATA results in inhibition of ZIKV replication (Figure 7). To that end, Vero cells were pre-treated with ATA for 12 (Figure 7A), 24 (Figure 7B), 48 (Figure 7C), or 72 (Figure 7D) h prior to infection (MOI 0.1) with Paraiba/2015. Pre-treatment with ATA for 12–72 h before ZIKV infection resulted in similar IC50 values (14.33 ± 1.06 μM, 13.18 ± 1.05 μM, 12.59 ± 1.02 μM, and 10.50 ± 1.05 μM; respectively) demonstrating that ATA is stable and able to prevent ZIKV infection even when administered 3 days previous to viral infection (Figure 7 and Table 1).
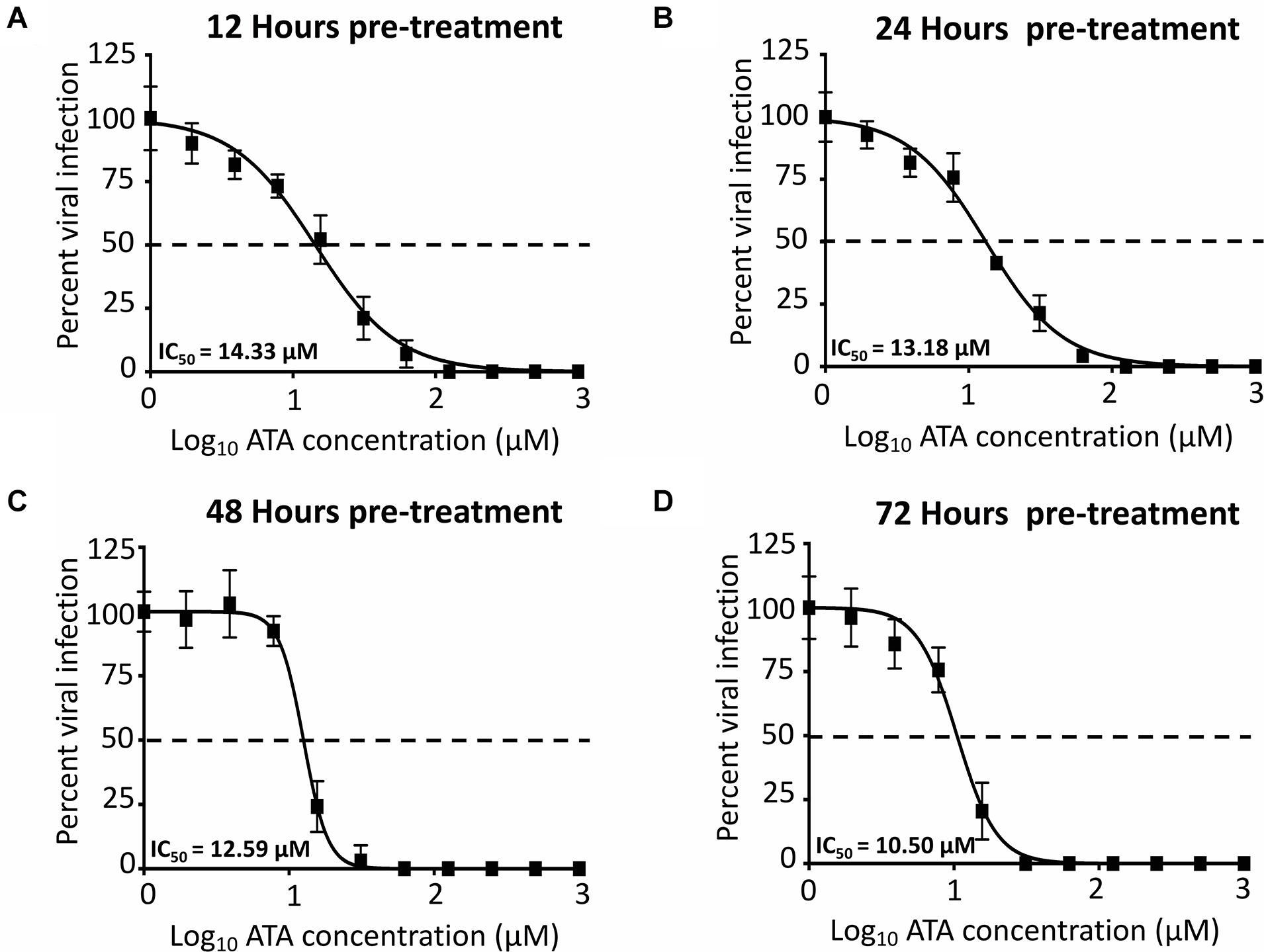
Figure 7. Inhibition of ZIKV by pre-treatment with ATA: Vero cells (96-well plates, 5 × 104 cells/well, triplicates) were pre-treated with the indicated twofold dilution concentrations of ATA (starting concentration 1,000 μM) for 12 (A), 24 (B), 48 (C), and 72 (D) h before infection (25 PFU) with ZIKV Paraiba/2015. At 36 h p.i., infected cells were fixed for virus titration by immunostaining. Dotted line indicates 50% inhibition. Data was expressed as mean and SD from three independent experiments conducted in triplicates.
Discussion
The recent outbreak of ZIKV accompanied with severe pathology, including microcephaly in newborns, prompted many researchers to develop prophylactic vaccines and to identify therapeutic drugs against ZIKV infection (Abbink et al., 2016; Larocca et al., 2016; Lessler et al., 2016; Shan et al., 2017; Fink et al., 2018). Currently, there are no commercially available vaccines and/or antiviral therapies for the treatment of ZIKV infection. Therefore, there is an urgent medical need for the development of effective counter measurements to control ZIKV infection.
In this study, we demonstrated that ATA (Figure 1) has limited toxicity (Figure 2) and an effective and dose-dependent antiviral activity against ZIKV infection (Figures 3, 4) in both monkey kidney epithelial Vero and human alveolar A549 cells. Notably, ATA can prevent ZIKV-induced CPE and apoptosis in both cell lines (Figure 5) and has broad anti-viral activity against representative ZIKV strains from the African (Uganda/1947 and Nigeria/1968) and the Asian/American (Puerto Rico/2015 and French Polynesia/2013) lineages (Figure 6). Moreover, ATA can also prevent ZIKV infection even when administered 3 days before infection (Figure 7).
Aurintricarboxylic acid is a polyanionic aromatic compound that structurally relates to suramin (Balzarini et al., 1986) and is believed to influence over 100 host and viral enzymes (Shadrick et al., 2013). Although the exact mechanism by which ATA inhibits ZIKV infection was not identified in this study, there are several plausible mechanisms on ZIKV inhibition mediated by ATA, including the targeting of viral and cellular proteins. In terms of inhibiting viral proteins, ATA could bind to ZIKV NS3 helicase and prevent its binding to either ATP or nucleic acids, as previously described for HCV (Mukherjee et al., 2012; Shadrick et al., 2013). Likewise, ATA could inhibit the ZIKV RNA-dependent RNA polymerase (RdRp) NS5 protein, as described for HCV (Chen et al., 2009; Mukherjee et al., 2012; Shadrick et al., 2013) and enterovirus 71 (Hung et al., 2010). Similarly, ATA could inhibit the methyltransferase activity of NS5 involved in mRNA capping processes, as previously described for other Flaviviruses (DENV and YFV) (Milani et al., 2009; Garcia et al., 2017). Because of the structural similarities between DENV and ZIKV NS5 proteins, it is feasible that, similar to DENV, ATA binds to NS5 to inhibit ZIKV infection (Garcia et al., 2017). Moreover, it is possible that ATA targets and has inhibitory activities against one or more of the viral proteins described above.
In terms of targeting cellular proteins important for the efficient replication of ZIKV, it has been previously described that ATA has anti-apoptotic properties in a variety of cells (Chen et al., 2002). It is possible that the anti-apoptotic activity of ATA protects against ZIKV-induced cell death, as demonstrated in this study (Figure 5). Notably, it has been recently shown that ZIKV infection induced apoptosis through caspase 3 and 9 in A549 cells and through caspase 3 in neonatal mice brain (Huang et al., 2016; Frumence et al., 2016). These results suggest that inhibition of ZIKV replication results in a decrease in the level of apoptotic cells and that the anti-apoptotic effect of ATA affects ZIKV replication. Further research is guaranteed to yield a better understanding of the antiviral activity of ATA on ZIKV infection, and other viruses, before the use of ATA as an antiviral drug.
During January 2015 to February 2016, a total of 116 residents from 33 states in the United States were diagnosed with ZIKV infection (Armstrong et al., 2016). Out of 115 patients, 110 (96%) traveled to areas of active ZIKV transmission before the infection and five (4%) did not travel but reported sexual contact with a traveler who had a symptomatic illness (Armstrong et al., 2016). For these reasons, preventive efforts are required prior to travel to areas of active ZIKV transmission. In this study, cells pretreated with ATA for up to 72 h prior to infection with ZIKV showed similar IC50 than those in post-treatment settings, potentially suggesting that ATA might target a cellular protein required for ZIKV replication or that the concentration and stability of ATA in pre-treated cells is sufficient to inhibit ZIKV infection, or both. Nevertheless, these results demonstrate the feasibility of using ATA for the prophylactic treatment of viral infection, including those traveling to areas where ZIKV is endemic. Moreover, due the broad inhibition effect of ATA against others viruses and parasites (Liang et al., 2003; He et al., 2004; De Clercq, 2005; Myskiw et al., 2007; Klein et al., 2008; Chen et al., 2009; Hung et al., 2009, 2010; Mukherjee et al., 2012; Shadrick et al., 2013) that are present in ZIKV endemic areas, treatment with ATA could be used for the broad prevention of DENV, YFV (Milani et al., 2009; Shadrick et al., 2013; Garcia et al., 2017), HCV (Chen et al., 2009; Mukherjee et al., 2012; Shadrick et al., 2013), and parasitic infestation (Cryptosporidium parvum) (Klein et al., 2008) for people traveling to these endemic regions. Moreover, the broad spectrum antiviral activity of ATA against different African and Asian/American ZIKV strains further guarantees the feasibility of implementing ATA to prevent ZIKV infection to travelers around the world.
Although ATA has been amply evaluated in vitro, only few studies have assessed the activity of ATA in vivo, including its use as a curative agent against thrombosis (Strony et al., 1990), apoptosis (Roberts-Lewis et al., 1993; Heiduschka and Thanos, 2000), parasite infestations (Klein et al., 2008), bacterial (Y. pestis) (Liang et al., 2003), and Vaccinia virus (Smee et al., 2010) infections. In the case of Vaccinia virus, ATA did not protect mice from a lethal challenge at a dose of 30 mg/kg/day (Smee et al., 2010). Further studies are needed to evaluate the anti-viral activity of ATA in vivo for the treatment of viral infections, including ZIKV.
Our studies show limited toxicity, if any, of ATA in cultured cells, including human A549 cells. The lack of knowledge about the use of ATA in pregnant women requires future additional safety tests, including studies using validated animal models of ZIKV infection, before using ATA for the treatment of ZIKV infection during pregnancy. Based on the effectiveness of ATA against ZIKV infection (SI = 72.1 in Vero cells and 26.26 in A549 cells) as compared to other previously described drugs, including emetine (SI = 9.84 in SNB-19 cells and 2.88 in ENV+ cells) (Yang et al., 2018), obatoclax [SI = 65 in human retinal pigment epithelial (RPE) cells] (Kuivanen et al., 2017), saliphenylhalamide (SI > 200 in RPE cells) (Kuivanen et al., 2017), gemcitabine (SI > 1,000 in RPE cells) (Kuivanen et al., 2017), sofosbuvir (SI > 52.63 in Huh-7 cells and 90.9 in Jar cells) (Bullard-Feibelman et al., 2017) and ARB (SI = 4.11 in Vero cells and 2.21 in A549 cells) (Fink et al., 2018; Haviernik et al., 2018) and this study (ATA, SI = 56.98–95.24 in Vero cells, and 26.26 in A549 cells), it is possible that ATA represents one of the most reasonable options of the treatment of ZIKV infection.
Author Contributions
J-GP, GÁ-P, and FM conceived, designed, and performed the experiments. TH, AN, FA, and LM-S provided the reagents, suggestions, and resources. FA and LM-S provided the funds. J-GP analyzed the data and wrote the initial manuscript draft. GÁ-P, FM, TH, AN, FA, and LM-S edited the original draft.
Funding
This work was supported in part by the National Institutes of Health (NIH) grant 1R21AI130500 to LM-S and FA.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
References
Abbink, P., Larocca, R. A., De La Barrera, R. A., Bricault, C. A., Moseley, E. T., Boyd, M., et al. (2016). Protective efficacy of multiple vaccine platforms against Zika virus challenge in rhesus monkeys. Science 353, 1129–1132. doi: 10.1126/science.aah6157
Armstrong, P., Hennessey, M., Adams, M., Cherry, C., Chiu, S., Harrist, A., et al. (2016). Travel-associated zika virus disease cases among u.s. residents–united states, january 2015-february 2016. MMWR 65, 286–289. doi: 10.15585/mmwr.mm6511e1
Avila-Perez, G., Nogales, A., Martin, V., Almazan, F., and Martinez-Sobrido, L. (2018). Reverse genetic approaches for the generation of recombinant zika virus. Viruses 10:E597. doi: 10.3390/v10110597
Balzarini, J., Mitsuya, H., De Clercq, E., and Broder, S. (1986). Aurintricarboxylic acid and evans blue represent two different classes of anionic compounds which selectively inhibit the cytopathogenicity of human T-cell lymphotropic virus type III/lymphadenopathy-associated virus. Biochem. Biophys. Res. Commun. 136, 64–71.
Bardina, S. V., Bunduc, P., Tripathi, S., Duehr, J., Frere, J. J., Brown, J. A., et al. (2017). Enhancement of zika virus pathogenesis by preexisting antiflavivirus immunity. Science 356, 175–180. doi: 10.1126/science.aal4365
Barrows, N. J., Campos, R. K., Powell, S. T., Prasanth, K. R., Schott-Lerner, G., Soto-Acosta, R., et al. (2016). A screen of FDA-approved drugs for inhibitors of zika virus infection. Cell Host Microbe 20, 259–270. doi: 10.1016/j.chom.2016.07.004
Benchokroun, Y., Couprie, J., and Larsen, A. K. (1995). Aurintricarboxylic acid, a putative inhibitor of apoptosis, is a potent inhibitor of DNA topoisomerase II in vitro and in Chinese hamster fibrosarcoma cells. Biochem. Pharmacol. 49, 305–313.
Bina-Stein, M., and Tritton, T. R. (1976). Aurintricarboxylic acid is a nonspecific enzyme inhibitor. Mol. Pharmacol. 12, 191–193.
Bullard-Feibelman, K. M., Govero, J., Zhu, Z., Salazar, V., Veselinovic, M., Diamond, M. S., et al. (2017). The FDA-approved drug sofosbuvir inhibits zika virus infection. Antiviral Res. 137, 134–140. doi: 10.1016/j.antiviral.2016.11.023
Catchpoole, D. R., and Stewart, B. W. (1994). Inhibition of topoisomerase II by aurintricarboxylic acid: implications for mechanisms of apoptosis. Anticancer Res. 14, 853–856.
Chen, C. W., Chao, Y., Chang, Y. H., Hsu, M. J., and Lin, W. W. (2002). Inhibition of cytokine-induced JAK-STAT signalling pathways by an endonuclease inhibitor aurintricarboxylic acid. Br. J. Pharmacol. 137, 1011–1020. doi: 10.1038/sj.bjp.0704955
Chen, Y., Bopda-Waffo, A., Basu, A., Krishnan, R., Silberstein, E., Taylor, D. R., et al. (2009). Characterization of aurintricarboxylic acid as a potent hepatitis C virus replicase inhibitor. Antivir. Chem. Chemother. 20, 19–36. doi: 10.3851/IMP1286
Colt, S., Garcia-Casal, M. N., Pena-Rosas, J. P., Finkelstein, J. L., Rayco-Solon, P., Weise Prinzo, Z. C., et al. (2017). Transmission of Zika virus through breast milk and other breastfeeding-related bodily-fluids: a systematic review. PLoS Negl. Trop. Dis. 11:e0005528. doi: 10.1371/journal.pntd.0005528
De Clercq, E. (2005). Emerging anti-HIV drugs. Expert Opin. Emerg. Drugs 10, 241–273. doi: 10.1517/14728214.10.2.241
Fink, S. L., Vojtech, L., Wagoner, J., Slivinski, N. S. J., Jackson, K. J., Wang, R., et al. (2018). The antiviral drug arbidol inhibits Zika virus. Sci. Rep. 8:8989. doi: 10.1038/s41598-018-27224-4
Frumence, E., Roche, M., Krejbich-Trotot, P., El-Kalamouni, C., Nativel, B., Rondeau, P., et al. (2016). The South Pacific epidemic strain of Zika virus replicates efficiently in human epithelial A549 cells leading to IFN-beta production and apoptosis induction. Virology 493, 217–226. doi: 10.1016/j.virol.2016.03.006
Garcia, L. L., Padilla, L., and Castano, J. C. (2017). Inhibitors compounds of the flavivirus replication process. Virol. J. 14:95. doi: 10.1186/s12985-017-0761-1
Haviernik, J., Stefanik, M., Fojtikova, M., Kali, S., Tordo, N., Rudolf, I., et al. (2018). Arbidol (Umifenovir): a broad-spectrum antiviral drug that inhibits medically important arthropod-borne flaviviruses. Viruses 10:E184. doi: 10.3390/v10040184
He, R., Adonov, A., Traykova-Adonova, M., Cao, J., Cutts, T., Grudesky, E., et al. (2004). Potent and selective inhibition of SARS coronavirus replication by aurintricarboxylic acid. Biochem. Biophys. Res. Commun. 320, 1199–1203. doi: 10.1016/j.bbrc.2004.06.076
Heiduschka, P., and Thanos, S. (2000). Aurintricarboxylic acid promotes survival and regeneration of axotomised retinal ganglion cells in vivo. Neuropharmacology 39, 889–902.
Huang, W. C., Abraham, R., Shim, B. S., Choe, H., and Page, D. T. (2016). Zika virus infection during the period of maximal brain growth causes microcephaly and corticospinal neuron apoptosis in wild type mice. Sci. Rep. 6:34793. doi: 10.1038/srep34793
Hung, H. C., Chen, T. C., Fang, M. Y., Yen, K. J., Shih, S. R., Hsu, J. T., et al. (2010). Inhibition of enterovirus 71 replication and the viral 3D polymerase by aurintricarboxylic acid. J. Antimicrob. Chemother. 65, 676–683. doi: 10.1093/jac/dkp502
Hung, H. C., Tseng, C. P., Yang, J. M., Ju, Y. W., Tseng, S. N., Chen, Y. F., et al. (2009). Aurintricarboxylic acid inhibits influenza virus neuraminidase. Antiviral Res. 81, 123–131. doi: 10.1016/j.antiviral.2008.10.006
Klein, P., Cirioni, O., Giacometti, A., and Scalise, G. (2008). In vitro and in vivo activity of aurintricarboxylic acid preparations against Cryptosporidium parvum. J. Antimicrob. Chemother. 62, 1101–1104. doi: 10.1093/jac/dkn303
Kuivanen, S., Bespalov, M. M., Nandania, J., Ianevski, A., Velagapudi, V., De Brabander, J. K., et al. (2017). Obatoclax, saliphenylhalamide and gemcitabine inhibit Zika virus infection in vitro and differentially affect cellular signaling, transcription and metabolism. Antiviral Res. 139, 117–128. doi: 10.1016/j.antiviral.2016.12.022
Larocca, R. A., Abbink, P., Peron, J. P., Zanotto, P. M., Iampietro, M. J., Badamchi-Zadeh, A., et al. (2016). Vaccine protection against Zika virus from Brazil. Nature 536, 474–478. doi: 10.1038/nature18952
Lazear, H. M., and Diamond, M. S. (2016). Zika virus: new clinical syndromes and its emergence in the western hemisphere. J. Virol. 90, 4864–4875. doi: 10.1128/JVI.00252-16
Lessler, J., Chaisson, L. H., Kucirka, L. M., Bi, Q., Grantz, K., Salje, H., et al. (2016). Assessing the global threat from Zika virus. Science 353:aaf8160. doi: 10.1126/science.aaf8160
Liang, F., Huang, Z., Lee, S. Y., Liang, J., Ivanov, M. I., Alonso, A., et al. (2003). Aurintricarboxylic acid blocks in vitro and in vivo activity of YopH, an essential virulent factor of Yersinia pestis, the agent of plague. J. Biol. Chem. 278, 41734–41741. doi: 10.1074/jbc.M307152200
Marquez-Jurado, S., Nogales, A., Avila-Perez, G., Iborra, F. J., Martinez-Sobrido, L., and Almazan, F. (2018). An alanine-to-valine substitution in the residue 175 of zika virus NS2A protein affects viral rna synthesis and attenuates the virus in vivo. Viruses 10:547. doi: 10.3390/v10100547
Milani, M., Mastrangelo, E., Bollati, M., Selisko, B., Decroly, E., Bouvet, M., et al. (2009). Flaviviral methyltransferase/RNA interaction: structural basis for enzyme inhibition. Antiviral Res. 83, 28–34. doi: 10.1016/j.antiviral.2009.03.001
Mitra, D., Steiner, M., Lynch, D. H., Staiano-Coico, L., and Laurence, J. (1996). HIV-1 upregulates Fas ligand expression in CD4+ T cells in vitro and in vivo: association with Fas-mediated apoptosis and modulation by aurintricarboxylic acid. Immunology 87, 581–585. doi: 10.1046/j.1365-2567.1996.510589.x
Mukherjee, S., Hanson, A. M., Shadrick, W. R., Ndjomou, J., Sweeney, N. L., Hernandez, J. J., et al. (2012). Identification and analysis of hepatitis C virus NS3 helicase inhibitors using nucleic acid binding assays. Nucleic Acids Res. 40, 8607–8621. doi: 10.1093/nar/gks623
Myskiw, C., Deschambault, Y., Jefferies, K., He, R., and Cao, J. (2007). Aurintricarboxylic acid inhibits the early stage of vaccinia virus replication by targeting both cellular and viral factors. J. Virol. 81, 3027–3032. doi: 10.1128/JVI.02531-06
Nayak, S., Lei, J., Pekosz, A., Klein, S., and Burd, I. (2016). Pathogenesis and molecular mechanisms of zika virus. Semin. Reprod. Med. 34, 266–272. doi: 10.1055/s-0036-1592071
Nazerai, L., Pravsgaard Christensen, J., and Randrup Thomsen, A. (2019). A ’furry-tale’ of zika virus infection: what have we learned from animal models? Viruses 11:E29. doi: 10.3390/v11010029
Nogales, A., Baker, S. F., and Martinez-Sobrido, L. (2015). Replication-competent influenza a viruses expressing a red fluorescent protein. Virology 476, 206–216. doi: 10.1016/j.virol.2014.12.006
Oehler, E., Watrin, L., Larre, P., Leparc-Goffart, I., Lastere, S., Valour, F., et al. (2014). Zika virus infection complicated by guillain-barre syndrome–case report, french polynesia, december 2013. Euro Surveill. 19:20720.
Priyamvada, L., Suthar, M. S., Ahmed, R., and Wrammert, J. (2017). Humoral immune responses against zika virus infection and the importance of preexisting flavivirus immunity. J. Infect. Dis. 216, S906–S911. doi: 10.1093/infdis/jix513
Ramos da Silva, S., and Gao, S. J. (2016). Zika virus: an update on epidemiology, pathology, molecular biology, and animal model. J. Med. Virol. 88, 1291–1296. doi: 10.1002/jmv.24563
Richner, J. M., Himansu, S., Dowd, K. A., Butler, S. L., Salazar, V., Fox, J. M., et al. (2017). Modified mRNA vaccines protect against zika virus infection. Cell 169:176. doi: 10.1016/j.cell.2017.03.016
Rivera-Concepcion, J. R., Betancourt, J. P., Cerra, J., and Reyes, E. (2018). The zika virus: an association to guillain-barre syndrome in the united states - a case report. P. R. Health Sci. J. 37, S93–S95.
Roberts-Lewis, J. M., Marcy, V. R., Zhao, Y., Vaught, J. L., Siman, R., and Lewis, M. E. (1993). Aurintricarboxylic acid protects hippocampal neurons from NMDA- and ischemia-induced toxicity in vivo. J. Neurochem. 61, 378–381.
Rui, H., Xu, J., Mehta, S., Fang, H., Williams, J., Dong, F., et al. (1998). Activation of the Jak2-Stat5 signaling pathway in Nb2 lymphoma cells by an anti-apoptotic agent, aurintricarboxylic acid. J. Biol. Chem. 273, 28–32.
Shadrick, W. R., Mukherjee, S., Hanson, A. M., Sweeney, N. L., and Frick, D. N. (2013). Aurintricarboxylic acid modulates the affinity of hepatitis C virus NS3 helicase for both nucleic acid and ATP. Biochemistry 52, 6151–6159. doi: 10.1021/bi4006495
Shan, C., Muruato, A. E., Nunes, B. T. D., Luo, H., Xie, X., Medeiros, D. B. A., et al. (2017). A live-attenuated Zika virus vaccine candidate induces sterilizing immunity in mouse models. Nat. Med. 23, 763–767. doi: 10.1038/nm.4322
Smee, D. F., Hurst, B. L., and Wong, M. H. (2010). Lack of efficacy of aurintricarboxylic acid and ethacrynic acid against vaccinia virus respiratory infections in mice. Antivir. Chem. Chemother. 20, 201–205. doi: 10.3851/IMP1480
Strony, J., Phillips, M., Brands, D., Moake, J., and Adelman, B. (1990). Aurintricarboxylic acid in a canine model of coronary artery thrombosis. Circulation 81, 1106–1114.
Tang, H., Hammack, C., Ogden, S. C., Wen, Z., Qian, X., Li, Y., et al. (2016). Zika virus infects human cortical neural progenitors and attenuates their growth. Cell Stem Cell 18, 587–590. doi: 10.1016/j.stem.2016.02.016
Tripathi, S., Balasubramaniam, V. R., Brown, J. A., Mena, I., Grant, A., Bardina, S. V., et al. (2017). A novel Zika virus mouse model reveals strain specific differences in virus pathogenesis and host inflammatory immune responses. PLoS Pathog. 13:e1006258. doi: 10.1371/journal.ppat.1006258
Weaver, S. C., Costa, F., Garcia-Blanco, M. A., Ko, A. I., Ribeiro, G. S., Saade, G., et al. (2016). Zika virus: History, emergence, biology, and prospects for control. Antiviral Res. 130, 69–80. doi: 10.1016/j.antiviral.2016.03.010
Keywords: Flavivirus, Zika virus, aurintricarboxylic acid, antivirals, prophylactic, therapeutic, drug treatment
Citation: Park J-G, Ávila-Pérez G, Madere F, Hilimire TA, Nogales A, Almazán F and Martínez-Sobrido L (2019) Potent Inhibition of Zika Virus Replication by Aurintricarboxylic Acid. Front. Microbiol. 10:718. doi: 10.3389/fmicb.2019.00718
Received: 05 February 2019; Accepted: 21 March 2019;
Published: 12 April 2019.
Edited by:
Slobodan Paessler, The University of Texas Medical Branch at Galveston, United StatesReviewed by:
Sanja Glisic, University of Belgrade, SerbiaRichard A. Bowen, Colorado State University, United States
Copyright © 2019 Park, Ávila-Pérez, Madere, Hilimire, Nogales, Almazán and Martínez-Sobrido. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Luis Martínez-Sobrido, bHVpc19tYXJ0aW5lekB1cm1jLnJvY2hlc3Rlci5lZHU=