 Suwellen S. D. de Azevedo1†
Suwellen S. D. de Azevedo1† Fernanda H. Côrtes1†
Fernanda H. Côrtes1† Edson Delatorre1‡
Edson Delatorre1‡ Marcelo Ribeiro-Alves2
Marcelo Ribeiro-Alves2 Brenda Hoagland2
Brenda Hoagland2 Beatriz Grinsztejn2Valdilea G. Veloso2
Beatriz Grinsztejn2Valdilea G. Veloso2 Mariza G. Morgado1
Mariza G. Morgado1 Gonzalo Bello1*
Gonzalo Bello1*- 1Laboratório de AIDS & Imunologia Molecular, Instituto Oswaldo Cruz, Fundação Oswaldo Cruz, Rio de Janeiro, Brazil
- 2Laboratório de Pesquisa Clínica em DST e AIDS, Instituto Nacional de Infectologia Evandro Chagas, Fundação Oswaldo Cruz, Rio de Janeiro, Brazil
Elite controllers (EC) are able to control HIV-1 replication to extremely low levels (<50 HIV-1 RNA copies/mL) in the absence of antiretroviral therapy. However, some EC experience CD4+ T cell loss and/or lose their ability to control HIV-1 over the course of infection. High levels of HIV-1 env proviral diversity, activated T cells and proinflammatory cytokines were pointed out as relevant biomarkers for detection of EC at risk of virologic/immunologic progression. The aim of this study was to assess the importance of proviral diversity as a prognostic marker of virologic and/or immunologic progression in EC. To this end, we analyzed plasma viremia, total HIV DNA levels, T cells dynamics, and activation/inflammatory biomarkers in EC with low (ECLD = 4) and high (ECHD = 6) HIV-1 env diversity. None of ECLD and ECHD subjects displayed evidence of immunologic progression (decrease in absolute and percentage of CD4+ T cells) and only one ECHD subject presented virologic progression (≥2 consecutive viral loads measurements above the detection limit) 2–5 years after determination of proviral env diversity. Despite differences in proviral genetic diversity, the ECLD and ECHD subgroups displayed comparable levels of total cell-associated HIV DNA, activated CD8+ T (CD38+HLA-DR+) cells and plasmatic inflammatory biomarkers (IP-10, IL-18, RANTES, PDGF-AA, and CTACK). These results indicate that the genetic diversity of the HIV-1 proviral reservoir is not a surrogate marker of residual viral replication, immune activation or inflammation, nor an accurate biomarker for the prediction of virologic breakthrough or CD4+ T cells loss in EC.
Introduction
A rare subset of HIV-1–infected individuals, termed elite controllers (EC), is able to naturally suppress viral replication to levels <50 HIV-1 RNA copies/mL during chronic infection in absence of antiretroviral therapy (ART) (Deeks and Walker, 2007). Despite the extraordinary ability to naturally suppress HIV-1 viremia, a proportion of EC exhibit a CD4+ T cell counts decline (immunologic progression) and/or lose their ability to control HIV (virologic progression) over the course of infection (Andrade et al., 2008; Okulicz et al., 2009; Sajadi et al., 2009; Boufassa et al., 2011; Yang et al., 2012; Olson et al., 2014; Leon et al., 2016; Chereau et al., 2017). Identify those EC at risk of CD4+ T cell loss and/or of breakthrough viremia may help to guide the selection of individuals that may benefit from ART initiation.
Some EC subjects exhibit abnormally high levels of systemic T cell activation and inflammation that may contribute to both CD4+ T cell counts loss and breakthrough viremia (Andrade et al., 2008; Hunt et al., 2008; Noel et al., 2014, 2015b; Pernas et al., 2017). Virologic progression has been also associated with the increase of intermittent viremia episodes, total HIV DNA and HIV proviral diversity in the blood (Noel et al., 2015b; Pernas et al., 2017). High levels of CD8+ T cell activation and interferon gamma-induced protein 10 (IP-10) were pointed out as the most discriminant biomarkers for detection of EC at risk of immunologic progression (Noel et al., 2014, 2015b); while levels of HIV env diversity and plasma pro-inflammatory cytokines CCL5/RANTES (Pernas et al., 2017) and Galectin-3-binding protein (Rodríguez-Gallego et al., 2018) were described as useful baseline markers to predict virologic progression in EC 1 year before the loss of HIV control.
A recent study conducted by our group demonstrated two divergent patterns of intra-host proviral diversity in a cohort of Brazilian EC (de Azevedo et al., 2017). A subgroup of EC displayed highly homogeneous proviral quasispecies (mean env genetic diversity < 2%), consistent with the maintenance of the viral reservoir by clonal expansion of long-lived HIV-infected memory CD4+ T cells. The other subgroup of EC showed more diverse proviral populations (mean env genetic diversity ≥ 2%), consistent with residual evolution and continuous reseeding of the proviral reservoir. The precise association between these divergent patterns of intra-host env proviral diversity and the subsequent HIV-1 disease progression in our EC cohort was not addressed before.
Here, we evaluated the importance of HIV-1 env proviral diversity as a possible prognostic marker of immunologic and/or virologic progression in our EC cohort by analyzing the plasma viremia, total cell-associated HIV DNA levels, CD4+ T cells dynamics, CD8+ T cell activation and inflammatory biomarkers over infection course in EC harboring proviral quasispecies with low (ECLD = 4) and high (ECHD = 6) env diversity.
Materials and Methods
Study Subjects
The EC was defined as HIV-1–infected subjects with the most (≥70%) plasma viral load (VL) determinations under the detection limit (<50–80 copies/mL) in absence of ART as described previously (de Azevedo et al., 2017). Then, they were divided into two groups based on the proviral genetic diversity: (1) ECLD for those harboring proviral quasispecies with a mean env genetic diversity < 2% (n = 4); and (2) ECHD for those harboring proviral quasispecies with a mean env genetic diversity ≥ 2% (n = 6). All EC subjects were followed at least by 2 years after HIV-1 genetic diversity evaluation. A group of viremic controllers (VC) that displayed most (≥70%) VL determinations between 51 and 2,000 copies/mL (n = 8) was included as control. Immunologic progression was defined as a statistically significant decline in both absolute and percentage of CD4+ T cells (Burcham et al., 1991; Hulgan et al., 2007). Virologic progression in EC was defined as ≥ 2 consecutive detectable VL within 1 year (Pernas et al., 2017).
CD4+ T Cell Counts, Plasmatic Viral Load and Total Cell-Associated HIV-1 DNA Load Measurements
CD4+ T cells counts and plasma HIV-1 RNA VL were determined as described previously (de Azevedo et al., 2017). Total DNA was extracted from PBMC (1 × 107 cells) using the QIAamp DNA Mini Kit (Qiagen, Germany) and total cell-associated HIV-1 DNA load was quantified using the Generic HIV®DNA cell Kit (Biocentric, France), following the manufacturer’s recommendations. Results were reported either as actual numbers of HIV DNA copies/106 cells or as the threshold value of detection.
Analyses of Proviral Genetic Diversity
Proviral env sequences from PBMC were performed by single genome amplification (SGA) followed by analysis of viral diversity using conditions previously described (de Azevedo et al., 2017).
Markers of T Cell Activation and Inflammation
To quantify the expression of CD38 and HLA-DR on CD8+ T cells, cryopreserved PBMC were thawed and stained with the following antibodies: anti-CD3 APC-H7, anti-CD4 PECF594, anti-CD8 APC, anti-CD38 BB515, and anti-HLA-DR PE (BD Biosciences, United States), and acquired using a BD FACSAria IIu Flow Cytometer (BD Biosciences, United States). The Fixable Viability Stain 450 (FVS 450-BD Biosciences, United States) was used to exclude non-viable cells. Flow cytometric analyses were performed with FlowJo v.10.0.7 (Tree Star Inc., Ashland, OR, United States). Plasmatic levels (pg/mL) of IP-10, IL-18, RANTES, CTACK, and PDGF-AA were measured using the human Magnetic Luminex Performance Assay (R&D systems, United States), following manufacturer’s instruction and the analyses were performed on a Luminex 200 System (Luminex, United States).
Statistical Analysis
The Mann–Whitney U-test was used to compare data between ECLD and ECHD independent groups. The Wilcoxon signed-rank test was used to compare data between pre- and pos-diversity evaluation within either ECLD or ECHD. Correlations between markers cumulative measures and HIV-1 DNA diversity were assessed by the Spearman correlation coefficient. The slopes of subgroups of T cells were calculated for each one by linear regression analysis fitted by generalized least squares and with an autocorrelation structure of first order in respect to the time after HIV diagnosis. Tests were two-sided, and P ≤ 0.05 were considered as significant. Graphics and statistical analyses were performed using either GraphPad v6 (Prism Software, United States) or R (R Foundation for Statistical Computing, Austria) software.
Results
There were no statistically significant differences between ECLD and ECHD groups in terms of epidemiological (age, sex, and HIV-1 transmission), immunologic (absolute CD4+ and CD8+ T cells counts, %CD4 and CD4/CD8 ratio), genetic (HLA-B∗57/27 status) and virologic (plasma HIV-1 RNA, total HIV-1 DNA loads, and Hepatitis C status) characteristics at proviral diversity determination point or throughout the follow-up (Supplementary Table S1). Longitudinal analysis revealed no evidence of immunologic progression in our EC cohort (Figure 1). Subjects that showed a significant (P < 0.05) decrease in absolute CD4+ T cells counts over time (EC19LD, EC30HD, and EC39HD) maintained CD4+ T cells counts > 500 cells/mm3 and stable percentage of CD4+ T cells > 30%. Subjects that displayed significant (P < 0.01) decline in the percentage of CD4+ T cells (EC11LD and EC42HD) maintained stable or increasing numbers of CD4+ T cells counts over time. All ECLD and ECHD subjects kept persistent virologic control for 2–5 years after determination of proviral env diversity, with exception of subject EC42HD that displayed ≥2 consecutive VL measurements above the detection limit (59–97 copies/mL) in a 1-year period and thus lost the elite virologic control profile during follow-up (Figure 1 and Supplementary Table S2).
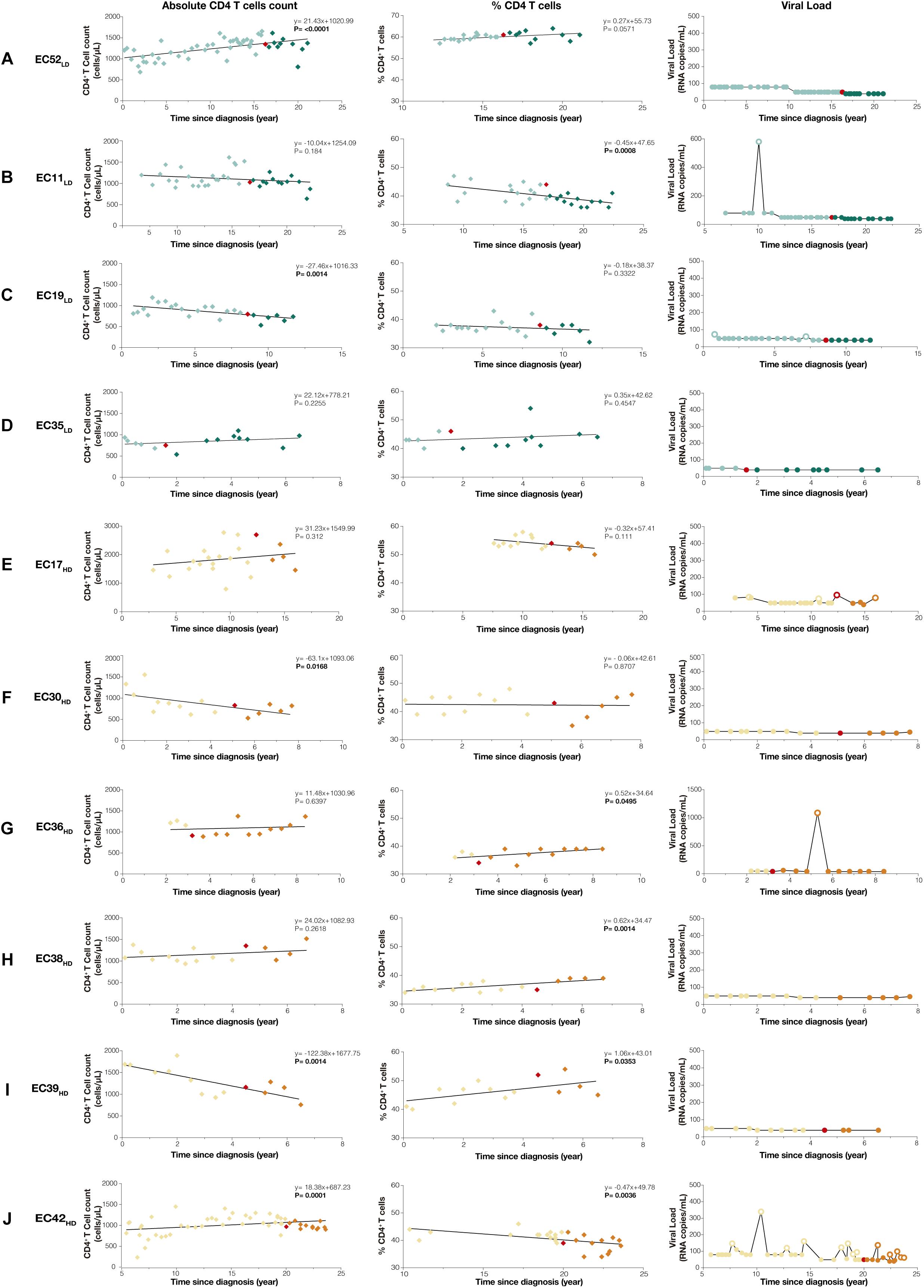
Figure 1. Temporal dynamics of CD4+ T cell counts and plasma viremia in ECLD (A–D) and ECHD (E–J) subgroups. Absolute CD4+ T cells counts (cells/μL), percentages of CD4+ T cell (%CD4) and plasma RNA viral load measurements (copies/mL) since HIV diagnosis are shown on the left, middle and right columns, respectively. The slope of CD4+ T cells and %CD4 overtime, calculated by linear regression analysis fitted by generalized least squares and with an autocorrelation structure of first order in respect to the time after HIV diagnosis, are shown with their respective P-values in the upper right corner of each graph. The red symbols (diamonds or circles) indicate the time point selected for analysis of env proviral diversity, while light and dark colored symbols indicate pre- and post-period of env proviral diversity determination, respectively. RNA viral load measurements below and above the detection limit were represented by fill and empty circles, respectively. ECLD and ECHD subjects were represented by green and orange symbols, respectively.
Longitudinal analyses of the cumulative mean CD4+ T cell counts (cCD4+ T cell) and VL (cVL) in the pre- and post-periods of the proviral diversity assay date revealed no statistically significant differences between ECLD and ECHD subgroups (Figures 2A–D) nor within each subgroup between time periods (Supplementary Figure S1). Similarly, cross-sectional analyses of total cell-associated HIV DNA levels, cellular immune activation and plasma inflammatory biomarkers also did not distinguish both EC subgroups (Figures 2E–K). The EC42HD subject, who had putative virologic progression after env diversity assay, presented most of the parameters within the range of other EC, except for cVL (pre) for which he exhibited the highest value of the group (Figure 2). The env proviral diversity was not significantly correlated with any of the virologic or immunologic parameter evaluated when EC were analyzed separately (Supplementary Figure S2). However, %CD8+CD38+HLA-DR+ T cells (P < 0.0001), total HIV DNA load (P = 0.003), and IP-10 (P = 0.03), were significantly higher in VC compared with EC (Supplementary Figure S3) and significant positive correlations with env proviral diversity (cVL, IP-10, and IL-18; Supplementary Figure S4A) and cVL (total HIV DNA load, %CD8+CD38+HLA-DR+ T cells, IP-10 and IL-18; Supplementary Figure S4B) were detected when HIV controllers (EC and VC) were taken as a whole.
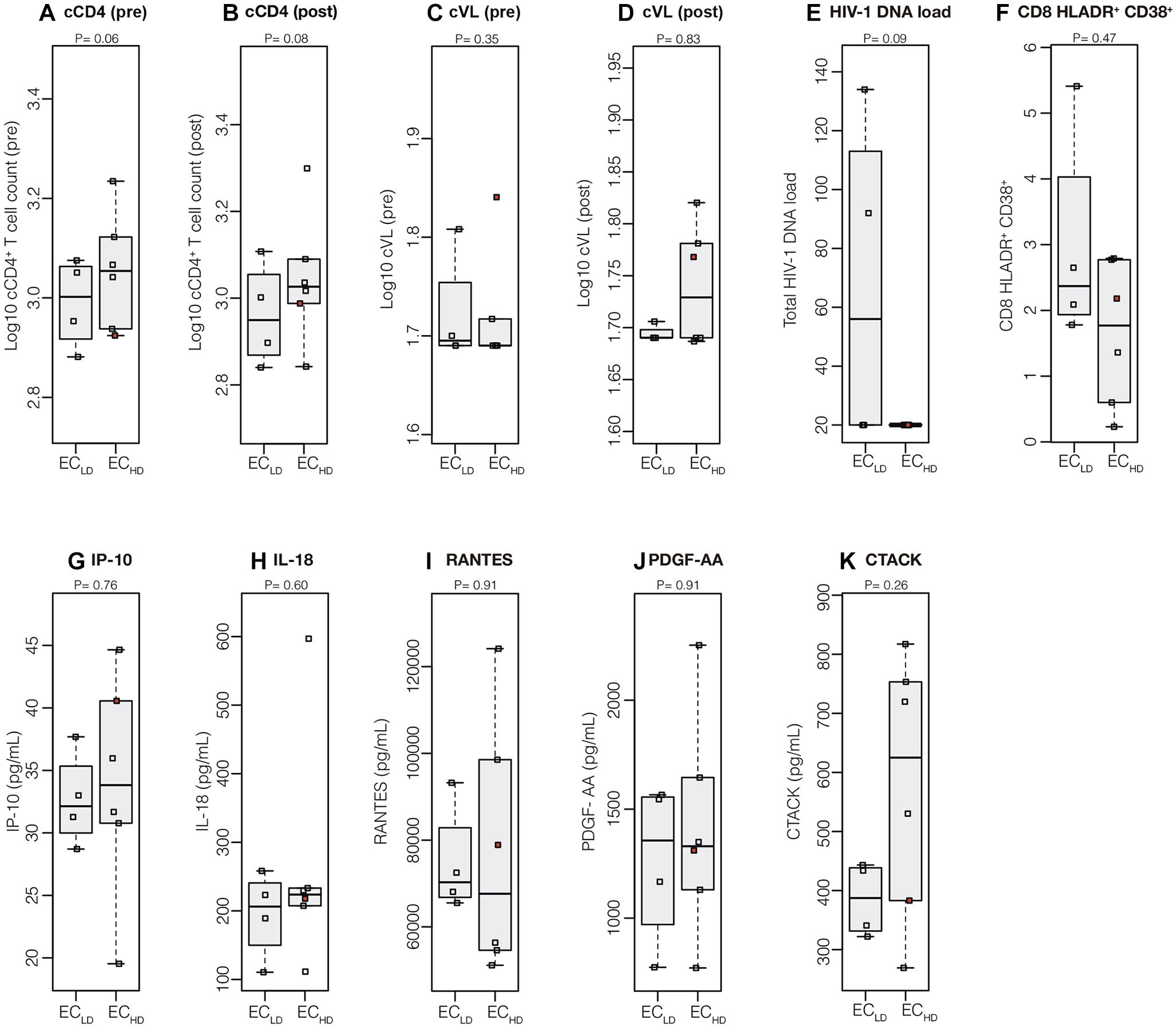
Figure 2. Virologic and immunologic progression markers in ECLD and ECHD subgroups. Mean (log10-transformed) cumulative measures of CD4+ T cell counts (A,B) and VL (C,D) in the pre- and post-periods respect to the point of determination of env proviral diversity and the total cell-associated HIV-1 DNA load (E), frequency of activated CD8+ T cells (F), and plasmatic levels of IP-10 (G), IL-18 (H), RANTES (I), PDGF-AA (J), and CTACK (K) at the env proviral diversity point were compared between EC subgroups. Central black solid bars and boxplots represent sample medians and interquartile, respectively. The red point in each graph indicate the values obtained for the subject who had putative virologic progression (EC42HD). P-values were obtained with Wilcoxon signed-rank tests.
Discussion
In a previous study, we identified two subgroups of EC (ECLD and ECHD) in a cohort of Brazilian subjects with divergent patterns of intra-host proviral diversity (de Azevedo et al., 2017). Our results comparing the clinical and epidemiologic aspects of the EC subgroups revealed no difference between them. Moreover, biomarkers previously associated with the risk of immunologic and/or virologic progression (CD4+ and CD8+ T cells counts, %CD4, CD8+ T cell activation, inflammation, HIV-1 RNA and DNA loads) in EC were not significantly different either before or after the proviral diversity assay among EC subsets.
The combined use of absolute and percentage CD4+ T cells trends over time reinforce the absence of true immunologic damage in our EC cohort. Subjects with a decrease in absolute CD4+ T cell counts showed stable (EC19LD and EC30HD) or increasing (EC39HD) %CD4+ T cells, indicating that reduction was not specific to CD4+ T cells (Supplementary Figure S5). Subjects with a significant decrease in the %CD4+ T cells displayed stable (EC11LD) or even increasing (EC42HD) absolute CD4+ T cells counts. Furthermore, all ECLD and ECHD subjects maintained CD4+ T cell counts > 500 cells/mm3 and percentage of CD4+ T cells > 30% during follow-up, supporting a low risk for disease progression (Burcham et al., 1991; Hulgan et al., 2007). The only subject with a virologic breakthrough (EC42HD) maintained VL in the very low range (59–97 copies/mL) and it could be argued that he did not display a true virologic progression (>2,000 copies/mL) (Noel et al., 2015b; Chereau et al., 2017). These data show that immunologic and/or virologic progression was rare in both ECLD and ECHD groups and reinforce the need to unify criteria for the definition of true progression in EC.
A recent study described higher levels of HIV env diversity and proinflammatory cytokines (RANTES, PDGF-AA, and CTACK) in EC that lost virologic control 1 year later, compared with EC that maintained persistent virologic control (Pernas et al., 2017). Our analyses did not reveal significant differences in the levels of RANTES, PDGF-AA, and CTACK between ECLD and ECHD subgroups. Similarly, levels of CD8+ T cell activation and IP-10, previously pointed out as biomarkers for detection of EC at risk of immunologic progression (Noel et al., 2014; Côrtes et al., 2018), were also not significantly different between the ECLD and ECHD groups. Noteworthy, the only EC subject who had allegedly virologic progression in our cohort (EC42HD) displayed similar levels of immune activation and inflammatory biomarkers than subjects with persistent elite virologic control.
These results support that some EC are able to maintain stable CD4+ T cells and persistent control of viral replication for several years (>2–5 years) in the setting of high proviral diversity. Although HIV-1 diversity and other surrogate markers of viral replication (total HIV-1 DNA load, CD8+ T cell activation, and IP-10) were positively correlated among each other and with the cVL when EC and VC were taken as a whole, consistent with previous findings (Bello et al., 2005; Sajadi et al., 2007; Groves et al., 2012; Côrtes et al., 2015; Noel et al., 2015a; Platten et al., 2016; Canouï et al., 2017; Tarancon-Diez et al., 2018); we did not detect any significant association between the env proviral diversity and those biomarkers when EC were analyzed separately. Hence, the genetic diversity of the HIV-1 proviral reservoir in many EC is probably not driven by continuous residual viral replication (de Azevedo et al., 2017), which may explain why ECHD does not seem to display higher risk of immunologic or virologic progression in compared with ECLD in our cohort. Those EC for which diversity of HIV-1 proviral reservoir truly reflects continuous viral replication and persistent inflammation are probably the only ones at risk of immunologic or virologic progression.
The main limitation of this study was the small number of EC individuals analyzed, which may have resulted in the apparent absence of significant differences in various markers evaluated between EC subgroups. This limitation was partially counterbalanced by the well recorded long period of follow-up of these extremely rare group of patients and the very rigorous classification criteria used to ensure that only EC individuals with a long-lasting HIV-1 control profile were included in our cohort (Walker and Yu, 2013). Further studies comprising a larger number of individuals are needed to complement our findings and to define the most suitable combination of biomarkers necessary to predict immunologic and/or virologic progression in long-term EC.
Conclusion
Our data suggest that genetic diversity of the HIV-1 proviral reservoir is not a surrogate marker of residual viral replication, immune activation or inflammation, nor an accurate biomarker for the prediction of virologic breakthrough or CD4+ T cells loss in EC. Most EC in our cohort maintained a persistent control of viremia and stable CD4+ T cells for up to 5 years after determination of HIV-1 quasispecies composition, irrespective of proviral genetic diversity. Understand the mechanisms leading to the divergent patterns of intra-host viral diversity in EC is of paramount importance to determine the potential impact of such divergent patterns on the long-term natural control of HIV-1 infection and their relevance for clinical management of EC.
Data Availability
All datasets generated for this study are included in the manuscript and/or the Supplementary Files.
Ethics Statement
All participants provided written informed consent and the ethical committee of Instituto Nacional de Infectologia Evandro Chagas (INI-Fiocruz) approved the study (CAAE 1717.0.000.009-07).
Author Contributions
GB conceived and designed the study and supervised the experiments. SA and FC conducted the experiments and analyzed the data. ED performed the quantification of total cell-associated HIV-1 DNA, its analysis, and provided intellectual input. MR-A analyzed the data. BH, BG, and VV conducted the patient recruitment and follow-up. MM contributed to the study design and provided intellectual input. SA, FC, and GB wrote the first draft. All authors assisted with the writing and approved the final manuscript.
Funding
This work was supported by the Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro – FAPERJ (Grant No. E-26/110.123/2014) and the Conselho Nacional de Desenvolvimento Científico e Tecnológico – CNPq (Grant No. 401220/2016-8). SA was supported by funding from Conselho Nacional de Desenvolvimento Científico e Tecnológico – CNPq and Fundação de Amparo à Pesquisa do Estado do Rio de Janeiro – FAPERJ. ED was financed by a Postdoctoral fellowship from the “Programa Nacional de Pós-Doutorado (PNPD)” by the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior – Brasil (CAPES) – Finance Code 001. GB was supported by fundings from CNPq (Grant No. 302317/2017-1) and FAPERJ (Grant No. E-26/202.896/2018).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank the patients, who participated in the study, as well as all the technical staff involved in the clinical follow-up of these patients. We also thank Leila Botelho Rodrigues da Silva and Marcelle Bral de Mello from Laboratório Tecnologia e Diagnóstico – Biomanguinhos/FIOCRUZ for sharing the Luminex 200 System facility.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2019.00673/full#supplementary-material.
References
Andrade, A., Bailey, J. R., Xu, J., Philp, F. H., Quinn, T. C., Williams, T. M., et al. (2008). CD4 + T cell depletion in an untreated HIV type 1–infected human leukocyte antigen–B∗5801–positive patient with an undetectable viral load. Clin. Infect. Dis. 46, e78–e82. doi: 10.1086/529387
Bello, G., Casado, C., Sandonis, V., Alonso-Nieto, M., Vicario, J. L., García, S., et al. (2005). A subset of human immunodeficiency virus type 1 long-term non-progressors is characterized by the unique presence of ancestral sequences in the viral population. J. Gen. Virol. 86, 355–364. doi: 10.1099/vir.0.80410-0
Boufassa, F., Saez-Cirion, A., Lechenadec, J., Zucman, D., Avettand-Fenoel, V., Venet, A., et al. (2011). CD4 dynamics over a 15 year-period among HIV controllers enrolled in the ANRS French observatory. PLoS One 6:e18726. doi: 10.1371/journal.pone.0018726
Burcham, J., Marmor, M., Dubin, N., Tindall, B., Cooper, D. A., Berry, G., et al. (1991). CD4% is the best predictor of development of AIDS in a cohort of HIV-infected homosexual men. Aids 5, 365–372. doi: 10.1097/00002030-199104000-00002
Canouï, E., Lécuroux, C., Avettand-Fenoël, V., Gousset, M., Rouzioux, C., Saez-Cirion, A., et al. (2017). A subset of extreme human immunodeficiency virus (HIV) controllers is characterized by a small HIV blood reservoir and a weak T-cell activation level. Open Forum Infect. Dis. 4, 1–8. doi: 10.1093/ofid/ofx064
Chereau, F., Madec, Y., Sabin, C., Obel, N., Ruiz-Mateos, E., Chrysos, G., et al. (2017). Impact of CD4 and CD8 dynamics and viral rebounds on loss of virological control in HIV controllers. PLoS One 12:e0173893. doi: 10.1371/journal.pone.0173893
Côrtes, F. H., de Paula, H. H. S., Bello, G., Ribeiro-Alves, M., de Azevedo, S. S. D., Caetano, D. G., et al. (2018). Plasmatic levels of IL-18, IP-10, and activated CD8+T cells are potential biomarkers to identify HIV-1 elite controllers with a true functional cure profile. Front. Immunol. 9:1576. doi: 10.3389/fimmu.2018.01576
Côrtes, F. H., Passaes, C. P. B., Bello, G., Teixeira, S. L. M., Vorsatz, C., Babic, D., et al. (2015). HIV controllers with different viral load cutoff levels have distinct virologic and immunologic profiles. J. Acquir. Immune Defic. Syndr. 68, 377–385. doi: 10.1097/QAI.0000000000000500
de Azevedo, S. S. D., Caetano, D. G., Côrtes, F. H., Teixeira, S. L. M., Santos Silva, K., Hoagland, B., et al. (2017). Highly divergent patterns of genetic diversity and evolution in proviral quasispecies from HIV controllers. Retrovirology 14, 1–13. doi: 10.1186/s12977-017-0354-5
Deeks, S. G., and Walker, B. D. (2007). Human immunodeficiency virus controllers: mechanisms of durable virus control in the absence of antiretroviral therapy. Immunity 27, 406–416. doi: 10.1016/j.immuni.2007.08.010
Groves, K. C., Bibby, D. F., Clark, D. A., Isaksen, A., Deayton, J. R., and Anderson, J. (2012). Disease progression in HIV-1-infected viremic controllers. J. Acquir. Immune Defic. Syndr. 61, 407–416. doi: 10.1097/QAI.0b013e318269c414
Hulgan, T., Shepherd, B. E., Raffanti, S. P., Fusco, J. S., Beckerman, R., Barkanic, G., et al. (2007). Absolute count and percentage of CD4+ lymphocytes are independent predictors of disease progression in HIV-infected persons initiating highly active antiretroviral therapy. J. Infect. Dis. 195, 425–431. doi: 10.1086/510536
Hunt, P. W., Brenchley, J., McCune, J. M., Roland, M., Page-Shafer, K., et al. (2008). Relationship between T cell Activation and CD4 + T cell count in HIV-seropositive individuals with undetectable plasma HIV RNA levels in the absence of therapy. J. Infect. Dis. 197, 126–133. doi: 10.1086/524143
Leon, A., Perez, I., Ruiz-Mateos, E., Benito, J. M., Leal, M., Lopez-Galindez, C., et al. (2016). Rate and predictors of progression in elite and viremic HIV-1 controllers. Aids 30, 1209–1220. doi: 10.1097/QAD.0000000000001050
Noel, N., Boufassa, F., Lécuroux, C., Saez-Cirion, A., Bourgeois, C., Dunyach-Remy, C., et al. (2014). Elevated IP10 levels are associated with immune activation and low CD4+ T-cell counts in HIV controller patients. Aids 28, 467–476. doi: 10.1097/QAD.0000000000000174
Noel, N., Lerolle, N., Lécuroux, C., Goujard, C., Venet, A., Saez-Cirion, A., et al. (2015a). Class-modeling analysis reveals T-cell homeostasis disturbances involved in loss of immune control in elite controllers. PLoS One 10:e0131922. doi: 10.1186/s12916-018-1026-6
Noel, N., Lerolle, N., Lécuroux, C., Goujard, C., Venet, A., Saez-Cirion, A., et al. (2015b). Immunologic and virologic progression in HIV controllers: the role of viral “blips” and immune activation in the ANRS CO21 CODEX study. PLoS One 10:e0131922. doi: 10.1371/journal.pone.0131922
Okulicz, J. F., Marconi, V. C., Landrum, M. L., Wegner, S., Weintrob, A., Ganesan, A., et al. (2009). Clinical outcomes of elite controllers, viremic controllers, and long-term nonprogressors in the US department of defense HIV natural history study. J. Infect. Dis. 200, 1714–1723. doi: 10.1086/646609
Olson, A. D., Meyer, L., Prins, M., Thiebaut, R., Gurdasani, D., Guiguet, M., et al. (2014). An evaluation of HIV elite controller definitions within a large seroconverter cohort collaboration. PLoS One 9:e86719. doi: 10.1371/journal.pone.0086719
Pernas, M., Tarancón-Diez, L., Rodríguez-Gallego, E., Gómez, J., Prado, J. G., Casado, C., et al. (2017). Factors leading to the loss of natural elite control of HIV-1 infection. J. Virol. 92, e01805–e01817. doi: 10.1128/JVI.01805-17
Platten, M., Jung, N., Trapp, S., Flossdorf, P., Meyer-Olson, D., Schulze Zur Wiesch, J., et al. (2016). Cytokine and chemokine signature in elite versus viremic controllers infected with HIV. AIDS Res. Hum. Retroviruses 32, 579–587. doi: 10.1089/AID.2015.0226
Rodríguez-Gallego, E., Tarancón-Diez, L., García, F., Del Romero, J., Miguel Benito, J., Alba, V., et al. (2018). Proteomic profile associated with loss of spontaneous HIV-1 elite control. J. Infect. Dis. 219, 867–876. doi: 10.1093/infdis/jiy599
Sajadi, M. M., Constantine, N. T., Mann, D. L., Charurat, M., Dadzan, E., Kadlecik, P., et al. (2009). Epidemiologic characteristics and natural history of HIV-1 natural viral suppressors. J. Acquir. Immune Defic. Syndr. 50, 403–408. doi: 10.1097/QAI.0b013e3181945f1e
Sajadi, M. M., Heredia, A., Le, N., Constantine, N. T., and Redfield, R. R. (2007). HIV-1 natural viral suppressors: control of viral replication in the absence of therapy. Aids 21, 517–519. doi: 10.1097/QAD.0b013e328013d9eb
Tarancon-Diez, L., Dominguez-Molina, B., Viciana, P., Lopez-Cortes, L., and Ruiz-Mateos, E. (2018). Long-term persistent elite HIV-controllers: the right model of functional cure. EBioMedicine 28, 15–16. doi: 10.1016/j.ebiom.2018.01.013
Walker, B. D., and Yu, X. G. (2013). Unravelling the mechanisms of durable control of HIV-1. Nat. Rev. Immunol. 13, 487–498. doi: 10.1038/nri3478
Keywords: elite controllers, HIV proviral diversity, plasma biomarkers, CD4+ T cell loss, breakthrough viremia
Citation: de Azevedo SSD, Côrtes FH, Delatorre E, Ribeiro-Alves M, Hoagland B, Grinsztejn B, Veloso VG, Morgado MG and Bello G (2019) Proviral Quasispecies Diversity Is Not Associated With Virologic Breakthrough or CD4+ T Cell Loss in HIV-1 Elite Controllers. Front. Microbiol. 10:673. doi: 10.3389/fmicb.2019.00673
Received: 14 February 2019; Accepted: 18 March 2019;
Published: 02 April 2019.
Edited by:
Hirofumi Akari, Kyoto University, JapanReviewed by:
Takamasa Ueno, Kumamoto University, JapanMa Angeles Muñoz-Fernandez, Hospital General Universitario Gregorio Marañón, Spain
Copyright © 2019 de Azevedo, Côrtes, Delatorre, Ribeiro-Alves, Hoagland, Grinsztejn, Veloso, Morgado and Bello. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gonzalo Bello Z2JlbGxvYnJAZ21haWwuY29t; Z2JlbGxvQGlvYy5maW9jcnV6LmJy
‡Present address: Edson Delatorre, Laboratório de Genética Molecular de Microrganismos, Instituto Oswaldo Cruz, Fundação Oswaldo Cruz, Rio de Janeiro, Brazil
†These authors have contributed equally to this work