
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 26 February 2019
Sec. Antimicrobials, Resistance and Chemotherapy
Volume 10 - 2019 | https://doi.org/10.3389/fmicb.2019.00337
This article is part of the Research Topic Discovery of novel plant-derived compounds with antibacterial actions against antibiotic-resistant bacteria View all 24 articles
 Javier Espinoza1,2*
Javier Espinoza1,2* Alejandro Urzúa3*
Alejandro Urzúa3* Loreto Sanhueza4*Mariana Walter5Paola Fincheira2,6Patricia Muñoz4Leonora Mendoza7
Loreto Sanhueza4*Mariana Walter5Paola Fincheira2,6Patricia Muñoz4Leonora Mendoza7 Marcela Wilkens8
Marcela Wilkens8Staphylococcus aureus is a serious human pathogen that is highly adaptive to environmental conditions and rapidly develops antibiotic resistance. The use of efflux pumps to reduce antibiotic concentrations at the intracellular level is one of the main mechanisms by which bacteria develop antibiotic resistance. The management of efflux pumps, specifically NorA, which is expressed by S. aureus strains, is a valuable strategy for restoring susceptibility in strains resistant to antibacterial agents. In recent years, many studies have focused on searching for natural substances that can reverse efflux pump-mediated resistance in S. aureus. Extracts and compounds obtained from plants can be efficient efflux pump inhibitors (EPIs) and represent a potentially patient-friendly strategy for controlling S. aureus. In the present study, we evaluated the ability of essential oils, petroleum ether extracts, dichloromethane extract (DCME) and six compounds isolated from the heartwood of Pilgerodendron uviferum (Cupressaceae) and two synthetic derivatives to inhibit efflux in NorA pumps in the following three S. aureus strains: K2378, which overexpressed the norA gene (norA++), K1902 (norA-deleted, ΔnorA) and the parental strain, NCTC 8325-4. Efflux activity was evaluated using a fluorometric method that measured the accumulation of the universal efflux pump substrate ethidium bromide (EtBr). Only DCME and the compounds 15-copaenol and epi-cubenol inhibited EtBr efflux by K2378. Even the lowest concentration of 15-copaenol exhibited a stronger inhibitory effect than carbonyl cyanide m-chlorophenyl hydrazone on EtBr efflux by K2378. 15-copaenal only showed inhibition of EtBr efflux in K2378 cells at 125 μg/mL, but not superior to the control inhibitor and 15-copaenyl acetate exerted no intrinsic EPI activity against K2378. Fractional inhibitory concentration index (FICI) values obtained in the checkerboard assays, indicated that all combinations between DCME, epi-cubenol and 15-copaenol, and tested antibiotics showed a synergistic effect in wild type, norA++ and ΔnorA strains. Moreover, those were not toxic for the HeLa cell line at concentrations in which the synergistic effect and inhibitory activity of efflux pumps was determined. Other extracts and compounds obtained from P. uviferum did not display EtBr efflux-inhibiting activity against the evaluated S. aureus strains.
Antibiotic resistance is one of the most pressing health problems worldwide due to the continuous appearance of antibiotic-resistant bacterial strains. Antibiotics decrease or attack bacterial pathogens by altering the functions of the bacterial cell wall, proteins, nucleic acids and metabolic pathways. Antibiotic resistance can occur via several mechanisms: (i) the modification of the active site of the target resulting in reduced drug-binding efficiency, (ii) the direct destruction or modification of the antibiotic drug by enzymes belonging to the microorganism, and (iii) the expression of efflux pumps (Walsh, 2000; Khan and Khan, 2016). Overall, these mechanisms can limit the effectiveness of antibacterial drugs, causing substantial clinical problems. This has increased the cost of treating patients and make it difficult or sometimes even impossible (Astolfi et al., 2017). The bacteria that cause the most major clinical problems are Klebsiella and Enterobacter species, Enterococcus faecium, Clostridium difficile, Acinetobacter baumannii, Pseudomonas aeruginosa, Escherichia coli, and S. aureus (Marinelli and Tomazs, 2010; Khan and Khan, 2016).
S. aureus is a human bacterial pathogen that belongs to the genus Staphylococcus and is characterized by Gram-positive cocci capable of living under aerobic to facultative anaerobic conditions (Reddy et al., 2017). Morphologically, they have a diameter ranging from 0.5 to 1.5 μm and can be found singly or in pairs, tetrads, short chains, and irregular grape-like clusters. Additionally, they are non-motile, non-spore-forming and occasionally may contain a polysaccharide capsule. With regard for their biochemical characteristics, S. aureus secretes coagulases, ferments mannitol to lactic acid, and reduces nitrates to nitrites (Karmakar et al., 2016; Reddy et al., 2017). These bacteria are normally found in human microflora at multiple body sites, such as the skin, perineum, nares, axillae, and gastrointestinal tract, in which approximately 18 strains can share a benign or commensal relationship with the host. Nevertheless, under pathogenic conditions, these bacteria are very difficult to treat due to their strong ability to produce virulence factors, rapidly colonize the host and develop drug resistance (Akanbi et al., 2017). In addition, S. aureus can assume a pathogenic lifestyle in the host cell after entering via a medical implantation or syringe inoculation under hospital conditions (Hiramatsu et al., 2014).
S. aureus is often implicated in a wide array of community and hospital infections, including cutaneous and soft tissue infections, bacteremia, septic arthritis, osteomyelitis and necrotizing pneumonia (Tong et al., 2015; Costa et al., 2016; Reddy et al., 2017). Due to indiscriminate use and its recurrent adaptation to clinical treatments over the last 50 years (Hiramatsu et al., 2014), S. aureus is always among the first bacterial species to develop resistance to multiple antibiotics (multiple drug resistance, MDR) (Thurlow et al., 2012), including resistance to β-lactamics, aminoglycosides, macrolides, lincosamides, fluoroquinolones (FQs), chloramphenicol, sulfonamides, streptomycin, and tetracycline (Schindler et al., 2013).
The mechanisms described in S. aureus that contribute to multidrug resistance include (i) alteration of the target at which the drug exerts its activity, (ii) mutations and alterations in cell wall and cellular membrane components that reduce drug access, and (iii) the derepression of chromosomal components that encode multidrug resistance efflux pumps (Foster, 2017). Efflux pumps are transmembrane proteins with the capacity to expel or exchange toxic compounds from bacterial cells, thus allowing bacterium survival. They are a self-defense mechanism by which the antibiotic is expulsed from the cell, predisposing the organism to high target-resistance (Jo and Ahn, 2016). The pumps are classified according to their characteristics into five families, the most studied of which are NorA, TetK, and MrsA (dos Santos et al., 2018). Analyses of the S. aureus genome have determined that a large number of MDR efflux pumps are encoded by its genes; however, it has been shown that NorA is its predominant efflux pump (Handzlik et al., 2013). It has also been reported that the norA gene, which encodes the carrier protein NorA, is overexpressed in 43% of S. aureus strains (Astolfi et al., 2017). NorA belongs to the major facilitator superfamily (MFS) of efflux pumps, which are composed of approximately 400 amino acids that are putatively arranged into 12 membrane-spanning helices, with a cytoplasmic loop between the helices (Santos-Costa et al., 2013). This efflux pump uses the proton motive force to energize the transport of antimicrobial compounds through the membrane via an antiport mechanism. NorA has the capability to efflux diverse drugs, such as hydrophilic fluoroquinolone antibiotics, dyes, such as acridine and ethidium bromide (EtBr); and biocides (Van Bambeke et al., 2006; Zimmermann et al., 2017).
Thus, the management of efflux pumps, specifically NorA in S. aureus, presents a valuable strategy for activating viable antibacterial targets with the aim of restoring susceptibility to resistant strains to antibacterial agents.
In recent years, many studies have focused on the search for NorA inhibitors of S. aureus of synthetic origin such as riparin derivatives (Costa et al., 2016) and 2-phenylquinoline derivatives (Felicetti et al., 2018). Moreover, natural substances derivates from plants have been able to inhibit NorA, such as 5′-methoxy-hydnocarpin isolated from Berberis fremontii (Stermitz et al., 2000), baicalein isolated from Scutellaria baicalensis (Chan et al., 2011), biochanin A and crysoplenetin isolated from Artemisia annua (Tegos et al., 2011), and boeravinone B isolated from the roots of Boerhavia diffusa (Singh et al., 2017), among others.
Extracts and compounds obtained from plants, such as efflux pump inhibitors (EPIs), have been shown to exert efficient activity and therefore constitute a friendly strategy for controlling S. aureus. Therefore, the present study focuses on the ability of essential oils (EOs), extracts and compounds isolated from the heartwood of Pilgerodendron uviferum (D. Don) Florin (Cupressaceae) and synthetic derivatives to inhibit ejection via the NorA efflux pump in a modified strain of S. aureus, norA++, which overexpresses the genes that encode the NorA efflux pump.
P. uviferum is a conifer native to Chile and Argentina that is known to be highly resistant to attack by microorganisms and insects. It has been postulated that its resistance is related to its chemical composition (Solis et al., 2004; Donoso et al., 2008). Therefore, this tree may be a potential source of natural products that can act as inhibitors of the S. aureus NorA efflux pump.
Essential oil, light petroleum ether extract (PEE), dichloromethane extract (DCME), natural sesquiterpenes, (-)-trans-calamenene (1), cadalene (2), (-)-cubenol (3), (-)-epi-cubenol (4), (-)-torreyol (5), and 15-copaenol (6) were obtained from the heartwood of P. uviferum. All samples were obtained and analyzed according to methods described in previous studies (Espinoza et al., 2016, 2018).
15-copaenyl acetate (6AC) was prepared according to Oyarzún and Garbarino (1988), with modifications. Compound 6 (60 mg, 0.273 mmol) was diluted in CH2Cl2 and treated with acetic anhydride (2.5 mL) and pyridine (0.5 mL) in a 10 mL round-bottom flask at room temperature for 48 h. Then, the reaction mixture was successively extracted with 5% HCl (3 × 5 mL), 5% NaHCO3 (3 × 5 mL), and H2O (2 × 5 mL). The organic layer was dried with anhydrous sodium sulfate, filtered and evaporated to dryness using a rotatory evaporator to yield a colorless oil corresponding to 15-copaenyl acetate (6AC) (50.6 mg, 0.193 mmol, 70.8%) (Oyarzún and Garbarino, 1988). The NMR data is displayed in Table 1.
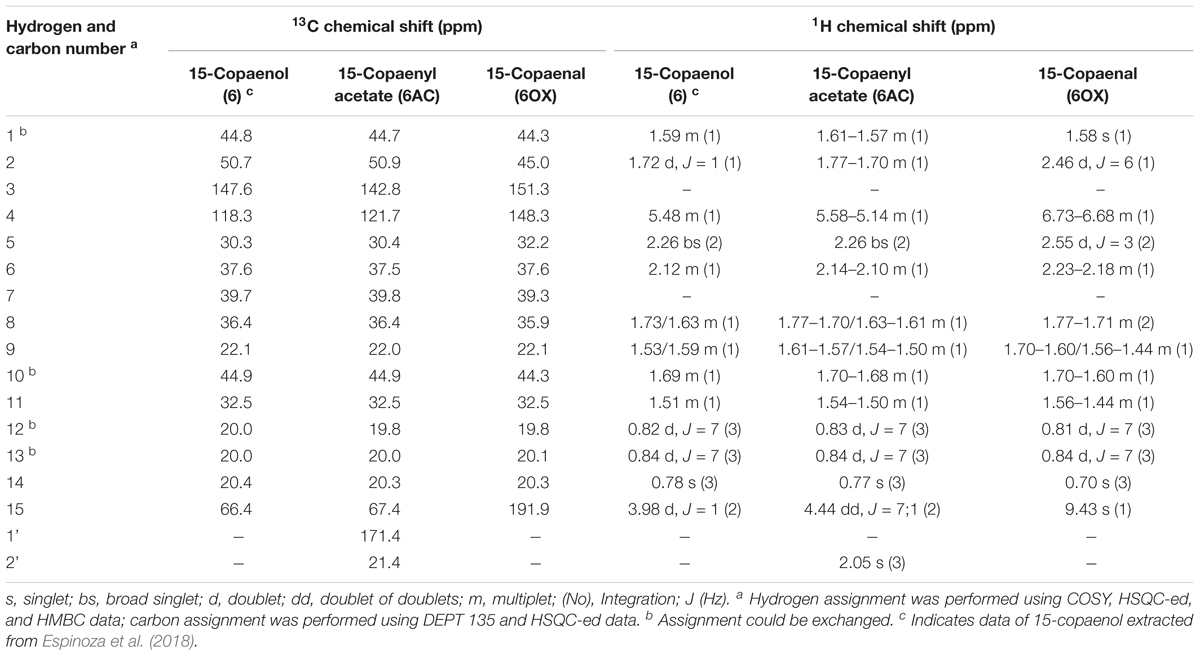
Table 1. 13C (100 MHz, CDCl3) and 1H-NMR (400 MHz, CDCl3) data of 15-copaenol, 15-copaenyl acetate, and 15-copaenal.
Chromium (VI) oxide (CrO3) (3.5 g, ACS reagent, ≥98%, Sigma-Aldrich, United States) was diluted with H2O (5 mL). The mixture was cooled on ice, and 98% H2SO4 (3 mL) (JTBaker, Avantor, United States) was added at a constant drip. Then, the mixture was diluted with H2O (10 mL) and stirred to obtain a solution corresponding to Jones reagent (18 mL) (Meinwald et al., 1973).
15-copaenal (6OX) was prepared according to Harding et al. (1975), with modifications. Compound 6 (60 mg, 0.273 mmol) was diluted in acetone (2.8 mL) and then added to a 10 mL round-bottom flask. The mixture was cooled to 0–5°C in an ice bath, and the Jones reagent was added at a constant drip until the colorless reaction mixture took on an orange color. The mixture was diluted with H2O (5.0 mL) and shaken for 10 min. Then, the reaction mixture was extracted with CH2Cl2 (3 × 5.0 mL). The organic layer was cleaned with an NaCl-saturated solution (3 × 5.0 mL) and evaporated to dryness using a rotatory evaporator. The oily residue was chromatographed on a silica gel column, yielding a pale-yellow oil corresponding to 15-copaenal (6OX) (39.6 mg, 0.182 mmol, 66.6%) (Weyerstahl et al., 1996). The NMR data is displayed in Table 1.
The heartwood EO, PEE and DCME and natural sesquiterpenes (1–6) were analyzed according to previously described methods (Espinoza et al., 2016, 2018). The synthetic derivatives 15-copaenyl acetate (6AC) and 15-copaenal (6OX) were analyzed by ATR-FT-IR and 1H- and 13C-NMR. The FT-IR spectra were recorded on a Perkin Elmer Spectrum 65 spectrometer. Both the 1H- and 13C-NMR spectra were recorded on a Bruker 400 Ultra Shield spectrometer (Bruker-Biospin GmbH, Germany) with the 5 mm NMR tube, using CDCl3 (99.8% D, contains 0.03% (v/v) TMS, Sigma-Aldrich, United States). 1H-NMR spectra were acquired using Bruker pulse program with the following settings: flip angle of 30°; relaxation delay (d1) = 0 s, size of fid = 32768, number of scans = 32, spectral width = 14.983 ppm, acquisition time = 5.4657 s, requested probe temperature = 300.0 K. 13C-NMR spectra were acquired using Bruker pulse program with the following settings: flip angle of 30°; relaxation delay (d11) = 30 ms and (d12) = 20 μs, size of fid = 16384, number of scans = 24166, spectral width = 240.049 ppm, acquisition time = 0.6783 s, requested probe temperature = 300.0 K. All data acquisition and processing were done with MestreNova 6.0.2 (Mestrelab Research, S.L., Santiago de Compostela, Spain).
The S. aureus strains used in this study were an NCTC 8325-4 parental strain, a K1902 mutant (ΔnorA), and a K2378 mutant that overexpressed norA (norA++) from a multicopy plasmid (Felicetti et al., 2018). This strain was produced by cloning norA and its promoter into plasmid pCU1 and then introducing the construct into SA-K1902 (Augustin et al., 1992). The bacterial strains were donated by Dr. Glenn Kaatz of the Division of Infectious Diseases, School of Medicine, Wayne State University, United States, and were stored at the Laboratorio de Microbiología Básica y Aplicada, Universidad de Santiago de Chile. Bacterial cultures were grown in Mueller Hinton (MH) broth (Sigma Aldrich, United States) and Luria-Bertani (LB) (Sigma-Aldrich, United States) medium at 37°C with agitation (220 rpm). S. aureus K1902 and S. aureus K2378 cultures were supplemented with 10 μg/mL chloramphenicol.
The minimal inhibitory concentration (MIC) was defined as the lowest concentration of a compound for which no growth was observed. The MIC determination assays were performed as established by the Clinical and Laboratory Standards Institute [CLSI] (2005) guidelines. The twofold standard micro broth dilution method was performed in 96-well plates. Briefly, bacterial strains were incubated overnight in 3 mL of MH broth at 37°C with shaking at 220 rpm. The bacterial cultures were then diluted in phosphate-buffered solution (PBS) to McFarland 0.5 (1 × 108 CFU/mL). To each well, 188 μL of MH broth, 10 μL of the EO, extract or compound (7.81–1,000 μg/mL), and 2 μL of a bacterial suspension were added to McFarland 0.5 to achieve a final volume of 200 μL. In addition, some wells were used as solvent and sterility controls. The plates were incubated at 37°C for 24 h, and the optical density was measured at 600 nm (OD600) in an Elisa lector (Thermo Scientific Multiskan FC Model, Waltham, MA, United States). The MICs of EtBr, a universal substrate of expulsion pumps, and carbonyl cyanide m-chlorophenyl hydrazone (CCCP) were used as the controls for EPIs and determined by the same method. All results are expressed as μg/mL, and experiments were performed in triplicate in three individual experiments.
The effects of extracts and compounds obtained from P. uviferum on the accumulation of EtBr was evaluated in the S. aureus NCTC 8325-4 (wild type), K2378 (norA++) and K1902 (ΔnorA) strains. Sub-inhibitory concentrations (½, ¼, and ⅛ of the MIC) of the study extracts and compounds were used in all experiments, and CCCP was used as an inhibitor control. The assay was carried out according to Paixão et al. (2009). Briefly, the S. aureus strains were grown in 10 mL of LB medium until they reached a mid-log phase, which corresponded to an OD600 of 0.6. Then, the bacteria were centrifuged at 10,786 × g for 3 min, the pellet was washed twice with the same volume of PBS, and the OD600 of the cellular suspension was adjusted to 0.3. Glucose was added to the cellular suspension to achieve a final concentration of 0.6% (v/v), and aliquots of 95 μL were transferred to 0.2 mL wells in 96-well microtiter plates. Aliquots containing 0.4 μL of EtBr or 0.2 μL of CCCP were then added to obtain final concentrations of 1.0 μg/mL and 1.25 μg/mL of EtBr and CCCP, respectively. Fluorescence (530/580 nm excitation/detection) was monitored by a microplate reader (BioTek Synergy HT Model, Winooski, VT, United States) at 37°C every 5 min for a period of 1 h. The effects of the extracts and compounds on the accumulation of EtBr was determined under conditions that optimized efflux (i.e., the presence of glucose and incubation at 37°C). All experiments were performed in triplicate.
The synergy assays between DCME extract or pure compounds, epi-cubenol and 15-copaenol in combination with antibiotics (ciprofloxacin and oxacillin) against S. aureus strains (wild type, norA++, ΔnorA) were evaluated by the checkerboard method described by Motyl et al. (2005), with minor modifications. Briefly, seven serial, twofold dilutions of extract or pure compounds and antibiotics were prepared. In a 96-well plate, 25 μL of each dilution of DCME, epi-cubenol or 15-copaenol was added in each vertical row, and 25 μL of antibiotic dilution was added in each horizontal row. Both, first horizontal and vertical rows were left with only one agent, and the following rows contained a fixed amount of one agent and increasing concentrations of the second agent. In the selection of the range of concentrations, the MICs obtained for each tested agent and tested S. aureus strains studied were considered. The extract concentrations used ranged from 1.95 to 125 μg/mL, 0.008 to 2 μg/mL for ciprofloxacin and 25 to 0.05 μg/mL for oxacillin. To each well, 100 μL of MH broth and 10 μL bacterial suspension to McFarland 0.5 (1 × 108 CFU/mL) were added. The plates were incubated at 37°C for 24 h and measured at 600 nm in Elisa lector (Thermos Labsystems Multiskan FC Model). All tests were performed in triplicate, in three different experiments. Fractional inhibitory concentrations (FICs) were calculated using the formula FICsample = (MIC extract + antibiotic/MIC sample) or FICantibiotic = (MIC sample + antibiotic/MIC antibiotic). The FIC index (FICI; Garvey et al., 2011) for each combination was calculated by the sum of both FIC values, and results were interpreted as follows: FICI ≤ 0.5 synergic effect, 0.5 < FICI ≤ 4 additive effect and FICI > 4 antagonistic effect (Garvey et al., 2011).
In order to determine the cytotoxic potential of DCME, epi-cubenol (4) and 15-copaenol (6) (active extract and compounds against efflux pumps in S. aureus), the cervical cancer cell line HeLa (ATCC CCL-2) was used.
The HeLa cell line was grown in monolayers in Dulbecco’s modified medium (DMEM; Corning, NY, United States) with bovine fetal serum 10% (FBS) supplemented with 100 U/mL penicillin and 100 μg/mL streptomycin (Corning Eagle, United States) in a modified Thermo Scientific air incubator (5% CO2 at 37°C). The medium was renewed every 2 days to reach 80% confluence. Then, the cells were transferred to T75 flasks, grown to reach 80% confluence, and finally, the culture was divided into sterile 24-well plates.
Approximately, 10,000 cells per well in a 24-well plate were seeded in 100 μL of DMEM. The cells were treated separately with eight different concentrations of extract and pure compounds (125–0.5 μg/mL). Simultaneously, growth controls were performed, which consisted of cells incubated with culture medium alone and with 4 μL methanol as solvent control. Finally, treated HeLa cells and the respective controls were incubated for 24 h in 5% CO2 at 37°C. All tests were carried out in triplicate. Cell viability was determined by counting live and dead cells after staining with 0.5 μM calcein-AM and 2 μM propidium iodide, for 30 min at room temperature, in darkness. Calcein-AM is a live scoreboard and propidium iodide intercalates into DNA being dead cells cell marker. Cells were visualized at 100× magnification on a Nikon microscope equipped Daipot inverted fluorescent and phase contrast optics. Cells were counted in a phase contrast microscope equipped with fluorescence using the following filters: calcein-AM 510–560 nm (excitation) and 590 nm (emission), propidium iodide 536 nm (excitation) and 617 nm (emission).
Data were analyzed by one- and two-way ANOVA test and two-sample T-test using Statistix 10 (Analytical Software, PO Box 12185, Tallahassee, FL, United States).
The compositions of EO and of PEE and DCME isolated from P. uviferum were determined according to Espinoza et al. (2016, 2018), respectively. According to those studies, the isolated EO contained three monoterpenes (0.43%) and 17 sesquiterpenes (86.62%), with (-)-torreyol (5) (24.16%), (-)-cubenol (3) (22.64%), 15-copaenol (6) (15.46%), and δ-cadinene (10.81%) the most abundant components (Espinoza et al., 2016). Regarding the extracts, 19 sesquiterpenes (73.53%) and 6 diterpenes (8.90%) present in the PEE and 17 sesquiterpenes (28.57%) and 5 diterpenes (47.97%) present in the DCME. The sesquiterpenes torreyol (5), cubenol (3), and 15-copaenol (6) were the most abundant compounds in both the PEE and DCME extracts (Espinoza et al., 2018). Sesquiterpenes were obtained from the EO and both of the extracts of P. uviferum heartwood using chromatographic methods and characterized by optical rotation, gas chromatography-mass spectrometry (GC/MS), Fourier-transform infrared spectroscopy (FT-IR), and 1D and 2D nuclear magnetic resonance (NMR) experiments. The following sesquiterpenes were identified: (-)-trans-calamenene (1), cadalene (2), (-)-cubenol (3), (-)-epi-cubenol (4), (-)-torreyol (5), and (-)-15-copaenol (6) (Figure 1).
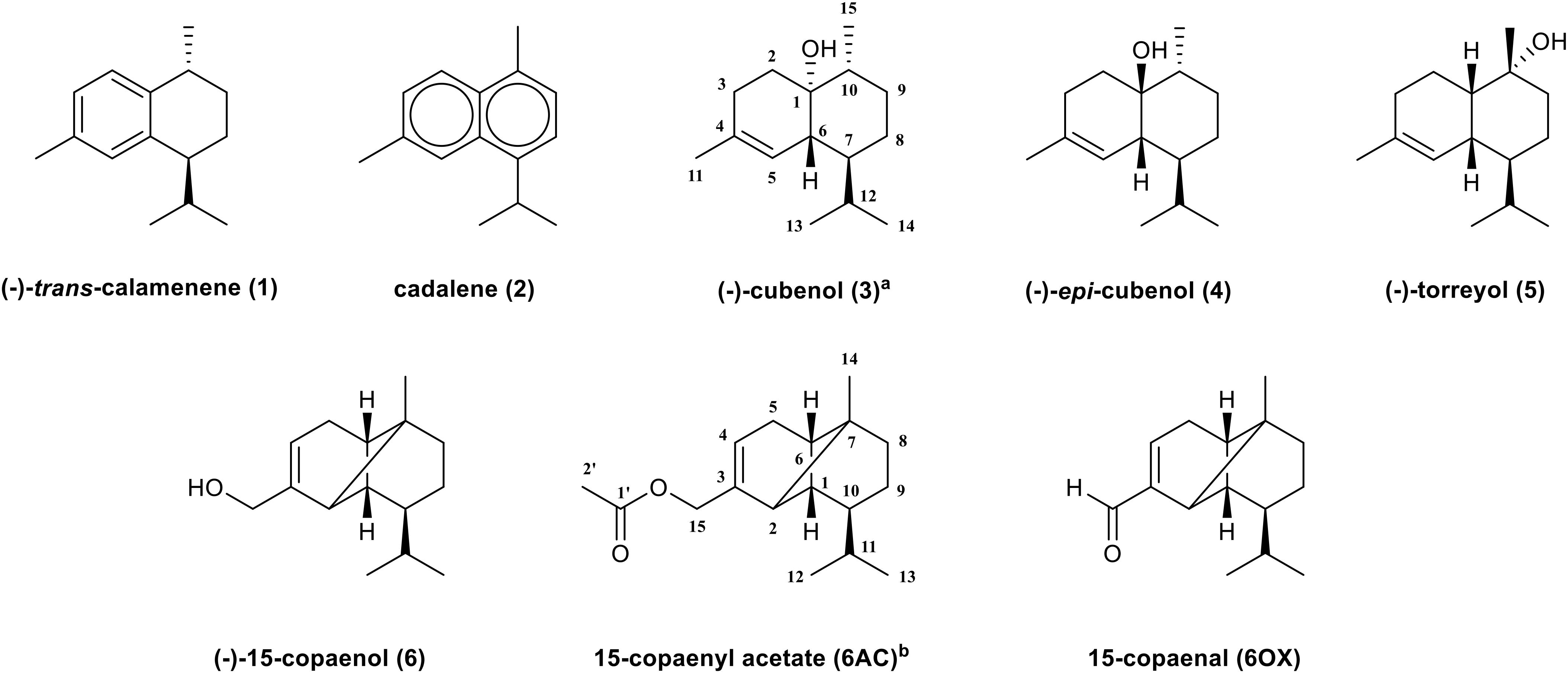
Figure 1. Sesquiterpenes isolated from Pilgerodendron uviferum and synthetic derivates of 15-copaenol. a Numbering belongs to compounds 1–5. b Numbering belongs to 15-copaenol, 6AC, and 6OX.
In MIC determination assays, 15-copaenol (6) had the lowest MIC value between all tested sesquiterpenes. Moreover, in an antibacterial assay against S. aureus (ATCC 6538p) and Bacillus cereus (ATCC 10876) (Data not published), 15-copaenol exerted the major bacterial growth inhibition. In this context, the structural requirements for antibacterial activity of some terpenoids has been reported. A hydrophobic moiety, consisting of a substituted decalin skeleton, and a hydrophilic region possessing one hydrogen-bond-donor group were the key structural requirements (Urzúa et al., 2008). Then, in order to evaluate the influence of the primary alcohol group in the antibacterial activity of copaene-type sesquiterpenes, hemi-synthetic modifications of 15-copaenol (6) were performed. Thus, 15-copaenol (6) was treated with acetic anhydride in pyridine and yielded 50.6 mg of a colorless oil. The 15-copaenol (6) was also treated with Jones reagent, and the product was chromatographically purified to yield 39.6 mg of a pale-yellow oil. Both synthetic derivatives were analyzed by attenuated total reflectance (ATR)-FT-IR and 1H- and 13C-NMR (Table 1). The IR spectrum of the acetylated derivate showed very strong CH valence vibration frequencies ranging from 2955 to 2872 cm-1; these corresponded to the aliphatic part of the molecule. Additionally, a characteristic tri-substituted C–C double bond stretching vibration valence was observed at 1677 cm-1. C–O stretching, C–C(=O)–O asymmetric stretching, and C–C–O asymmetric stretching were observed at 1,740, 1,236, and 1,046 cm-1, respectively, and were assigned to ester group vibrations. In contrast to 15-copaenol (6), the acetylated compound produced two additional signals in the 13C-NMR spectrum, that corresponded to a quaternary carbon (C-1’) and a methyl carbon (C-2’) at 171.4 and 21.4 ppm, respectively. The C-2’ signal showed a HSQC correlation with a singlet (3H) at 2.05 ppm (H-2’). The IR and NMR spectra indicated the substitution of a hydroxyl group in C-15 of 15-copaenol (6) by an acetyl group (Table 1).
The IR spectrum of the oxidized derivate showed the expected CH valence vibrations and tri-substituted C–C double bond stretching vibration valences from 2,956 to 2,872 cm-1 and 1,682 cm-1, respectively. Additionally, it showed a carbonyl group band at 1,707 cm-1 that corresponded to the C–O stretching of α,β-unsaturated aldehyde. Regarding the NMR analyses, compared with 15-copaenol (6), the methylene group signals from the primary alcohol of 15-copaenol (6) were absent in the oxidized derivate 1H and 13C-NMR spectrums. However, the C-15 signal at 191.9 ppm was observed in the 13C-NMR spectra. The C-15 signal had an HSQC correlation with an unshielded singlet (1H) at 9.43 ppm (H-15) in the 1H-NMR spectra. The IR and NMR spectra indicated hydroxyl group oxidation had occurred at C-15 of 15-copaenol (6) to yield an aldehyde group (Table 1). According to the IR and NMR analyses, the synthetic derivatives were identified as 15-copaenyl acetate (6AC), previously synthetized by Oyarzún and Garbarino (1988), and 15-copaenal (6OX), previously synthetized using a different methodology, by Weyerstahl et al. (1996) (Figure 1).
The MICs of all extracts and compounds were evaluated in the NCTC 8325-4 (wild type), K2378 (norA++), and K1902 (ΔnorA) S. aureus strains. The MICs of the samples are shown in Table 2. In general, MIC values were highest in K2378 (norA++) and lowest in K1902 (ΔnorA). In the extracts, PEE was the most active, producing MIC values of 31.3 μg/mL for all of the tested S. aureus strains. It was followed by EO and DCME, which produced higher values. Regarding the isolated compounds, 15-copaenol (6) was the most active compound against all evaluated strains, with MIC values of 31.3 μg/mL in S. aureus wild type and K2378 (norA++), respectively, and 62.5 μg/mL in K1902 (ΔnorA). It was followed by epi-cubenol (4) and cubenol (3), which produced values of 31.3 and 62.5 μg/mL, respectively, in wild type S. aureus, 62.5 μg/mL in K1902 (ΔnorA), and 125 μg/mL in K2378 (norA++). The other compounds produced no intrinsic antibacterial activity against the S. aureus strains (i.e., MIC > 32 mg/L) (Caspar et al., 2015). We found that torreyol (5) was inactive against all evaluated strains, similar to the result reported by Solis et al. (2004), who found that 15-copaenol (6) was more active than cubenol (3) and torreyol (5) against S. aureus (ATCC6538p) and Bacillus subtilis (ATCC6633).
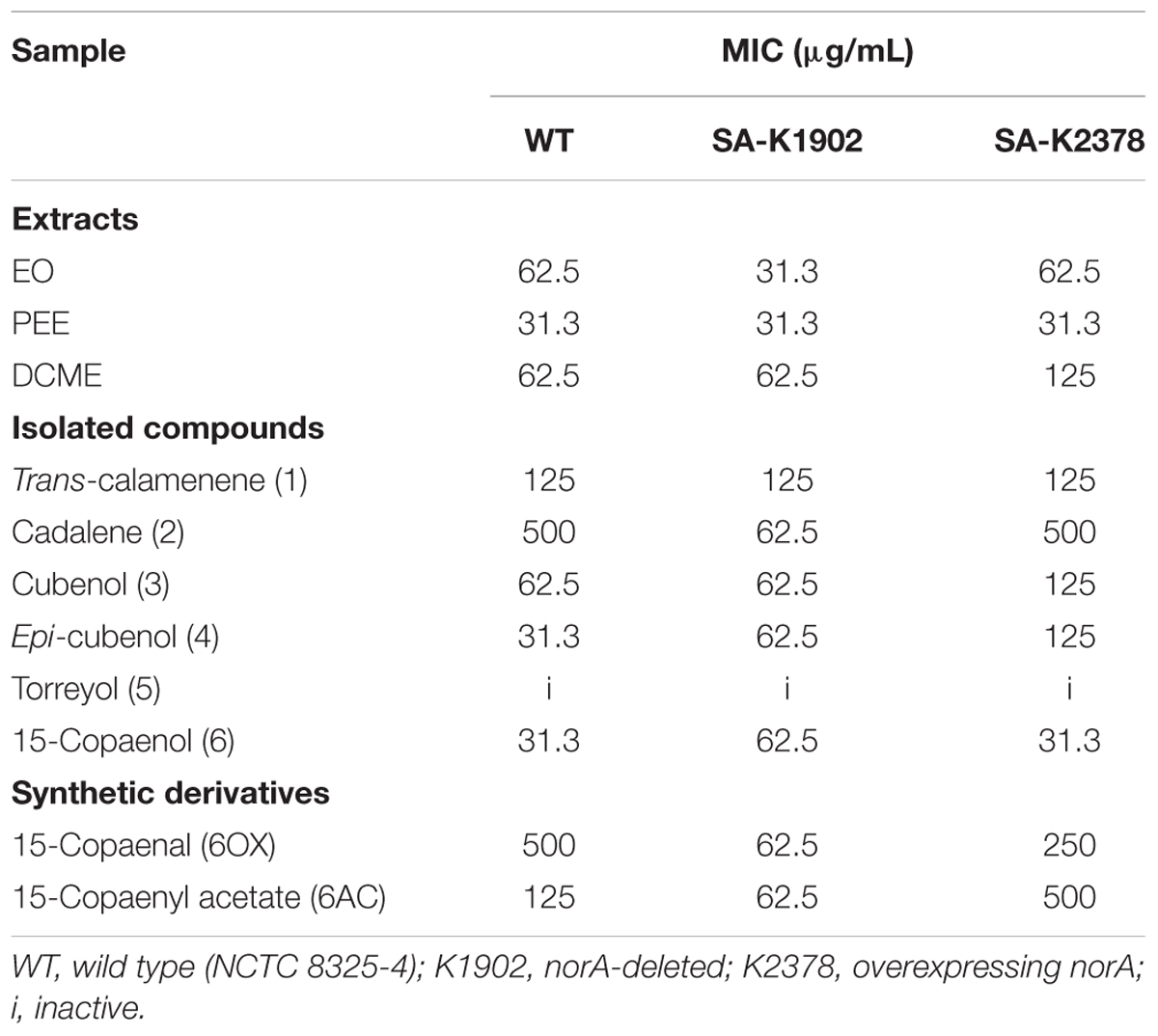
Table 2. MICs of essential oils (EOs), petroleum ether extracts (PEEs), dichloromethane extract (DCME), isolated sesquiterpenes, and synthetic derivates from P. uviferum on S. aureus strains.
The effects of different sub-inhibitory concentrations of extracts and compounds isolated from P. uviferum on EtBr accumulation were next evaluated in S. aureus wild type (NCTC 8325-4), K2378 (norA++) and K1902 (ΔnorA) strains. Experiments were carried out using EtBr concentrations that did not affect cell viability. The accumulation of EtBr inside the bacterial cells was increased by the presence of an EPI, such as CCCP, an EPI known to work by acting as a protonophore (Lowrence et al., 2016). Compounds that inhibit the efflux of EtBr results in higher residual fluorescence, whereas non-EPI compounds/untreated cells extrude EtBr, resulting in lower residual fluorescence. Accordingly, only the DCME and 15-copaenol (6) and epi-cubenol (4) acted as potential NorA EPIs to induce a higher level of fluorescence over time than was observed for the EtBr curve obtained in untreated K2378 (norA++) cells (Figures 2A–C). In particular, 15-copaenol (6) inhibited EtBr efflux in K2378 (norA++) cells at all evaluated concentrations (3.9–15.6 μg/mL). Even at the lowest concentration, it displayed a higher level of efflux inhibition than was observed for CCCP (Figure 2C), indicating that it could be indeed a highly potent efflux inhibitor in S. aureus (Lowrence et al., 2016). Despite this result, the synthetic derivative, 15-copaenal (6OX) only showed inhibition of EtBr efflux in K2378 (norA++) cells at 125 μg/mL, but not superior to the control inhibitor (CCCP) (Figure 2D) and 15-copaenyl acetate (6AC) showed no intrinsic EPI activity against the K2378 strain (Figure 2E). The DCME inhibited EtBr efflux in K2378 (norA++) cells at all evaluated concentrations (15.6–62.5 μg/mL) (Figure 2A), but it did not produce better efflux inhibition than was observed for CCCP. Epi-cubenol (4) inhibited EtBr efflux in the K2378 (norA++) strain only at the highest evaluated concentration (62.5 μg/mL) (Figure 2B); in those experiments, it produced an effect similar to that of CCCP.
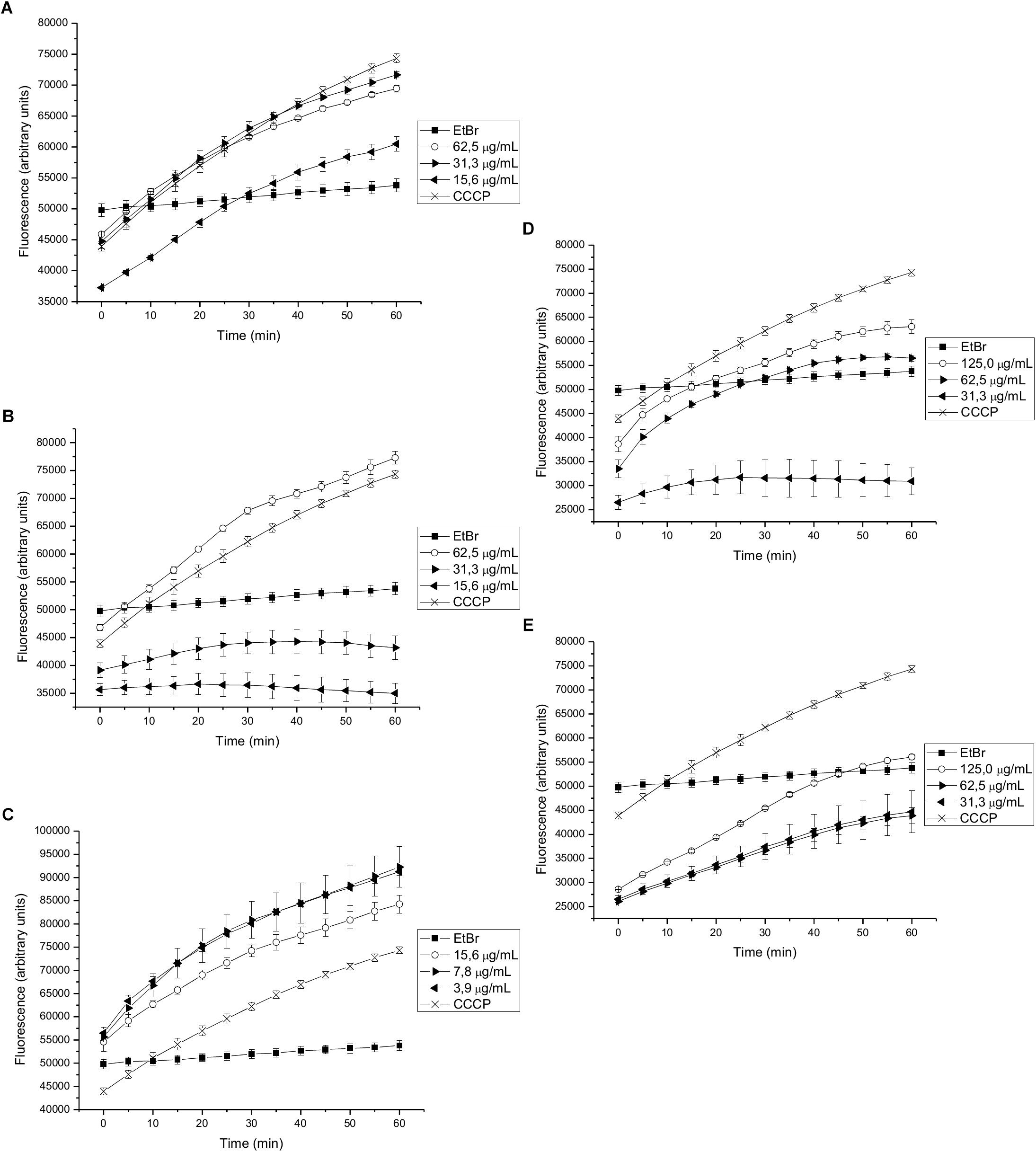
Figure 2. Effect on the intracellular accumulation of ethidium bromide of (A) DCME extract, (B) epi-cubenol (4), (C) 15-copaenol (6), (D) 15-copaenal (6OX), and (E) 15-copaenyl acetate (6AC) in K2378 (norA++) strain. The bacteria were exposed to 2.0 μg/mL of EtBr at 37°C for a period of 1 h in the presence of glucose. CCCP (1.25 μg/mL) was used as inhibitor control.
We found that 15-copaenol (6) also inhibited EtBr efflux in wild type S. aureus (NCTC 8325-4) at all evaluated concentrations (15.6–3.9 μg/mL) (Figure 3C), and its synthetic derivatives, 6OX and 6AC, inhibited EtBr efflux at 125–250 μg/mL (Figure 3D) and 15.6–62.5 μg/mL (Figure 3E), respectively. DCME inhibited EtBr efflux in wild type S. aureus when added at 15.6 and 31.3 μg/mL (Figure 3A), and epi-cubenol (4) inhibited EtBr efflux, better than CCCP, at concentrations of 3.9 and 7.8 μg/mL) (Figure 3B). Additionally, EO and PEE extract inhibited EtBr efflux in wild type S. aureus (NCTC 8325-4) (Supplementary Figures 2A,B). Finally, 15-copaenol (6) and EO were the only samples that inhibited EtBr efflux in the K1902 (ΔnorA) strain (at 31.3 μg/mL and 7.8 μg/mL, respectively) (Figure 4C and Supplementary Figure 3A). Other extracts and compounds obtained from P. uviferum did not display EtBr efflux-inhibiting activity in the wild type (NCTC 8325-4), K2378 (norA++), and K1902 (ΔnorA) (Figures 4A,B,D,E) strains of S. aureus (Supplementary Figures 1–3).
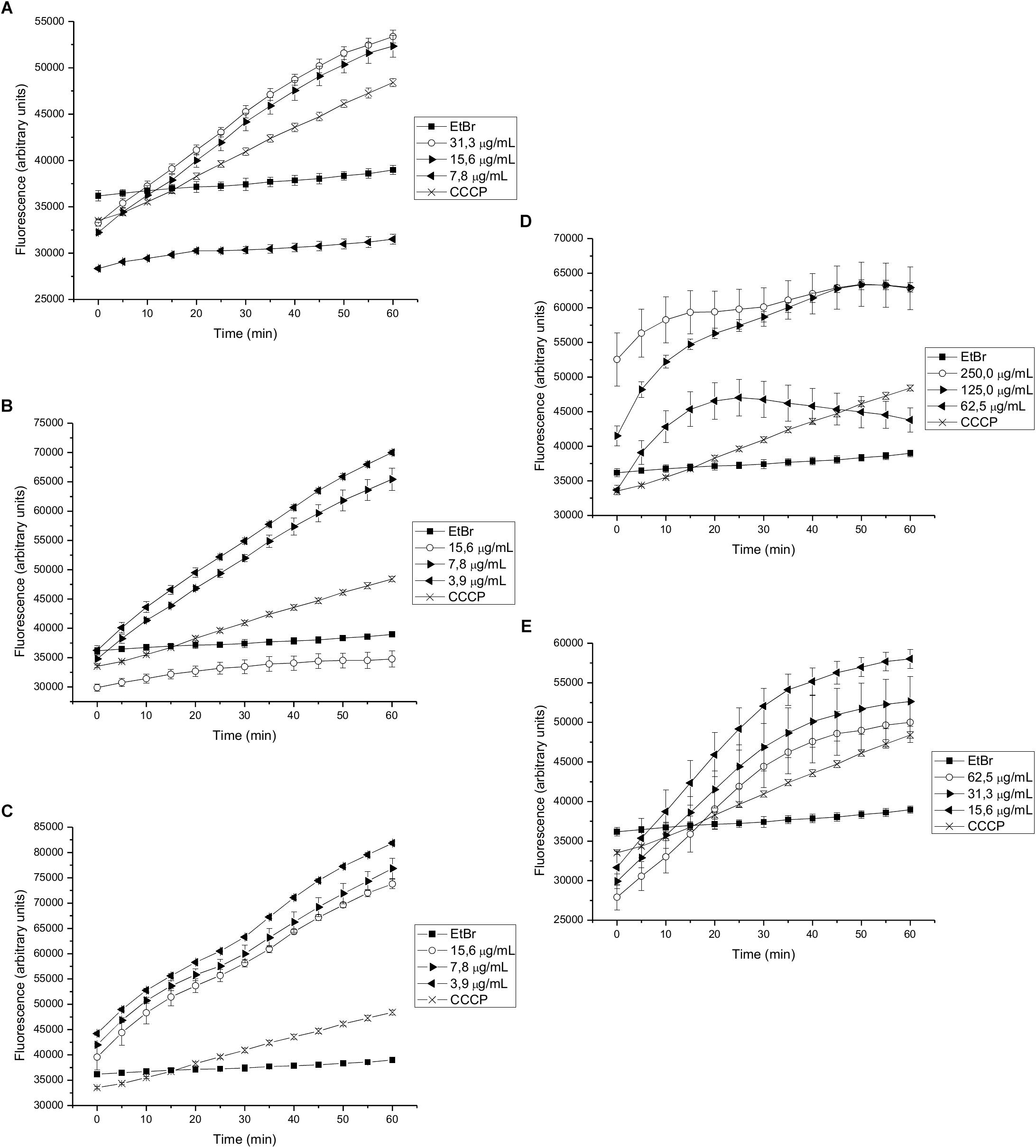
Figure 3. Effect on the intracellular accumulation of ethidium bromide of (A) DCME extract, (B) epi-cubenol (4), (C) 15-copaenol (6), (D) 15-copaenal (6OX), and (E) 15-copaenyl acetate (6AC) in S. aureus wild-type (NCTC 8325-4). The bacteria were exposed to 2.0 μg/mL of EtBr at 37°C for a period of 1 h in the presence of glucose. CCCP (1.25 μg/mL) was used as inhibitor control.
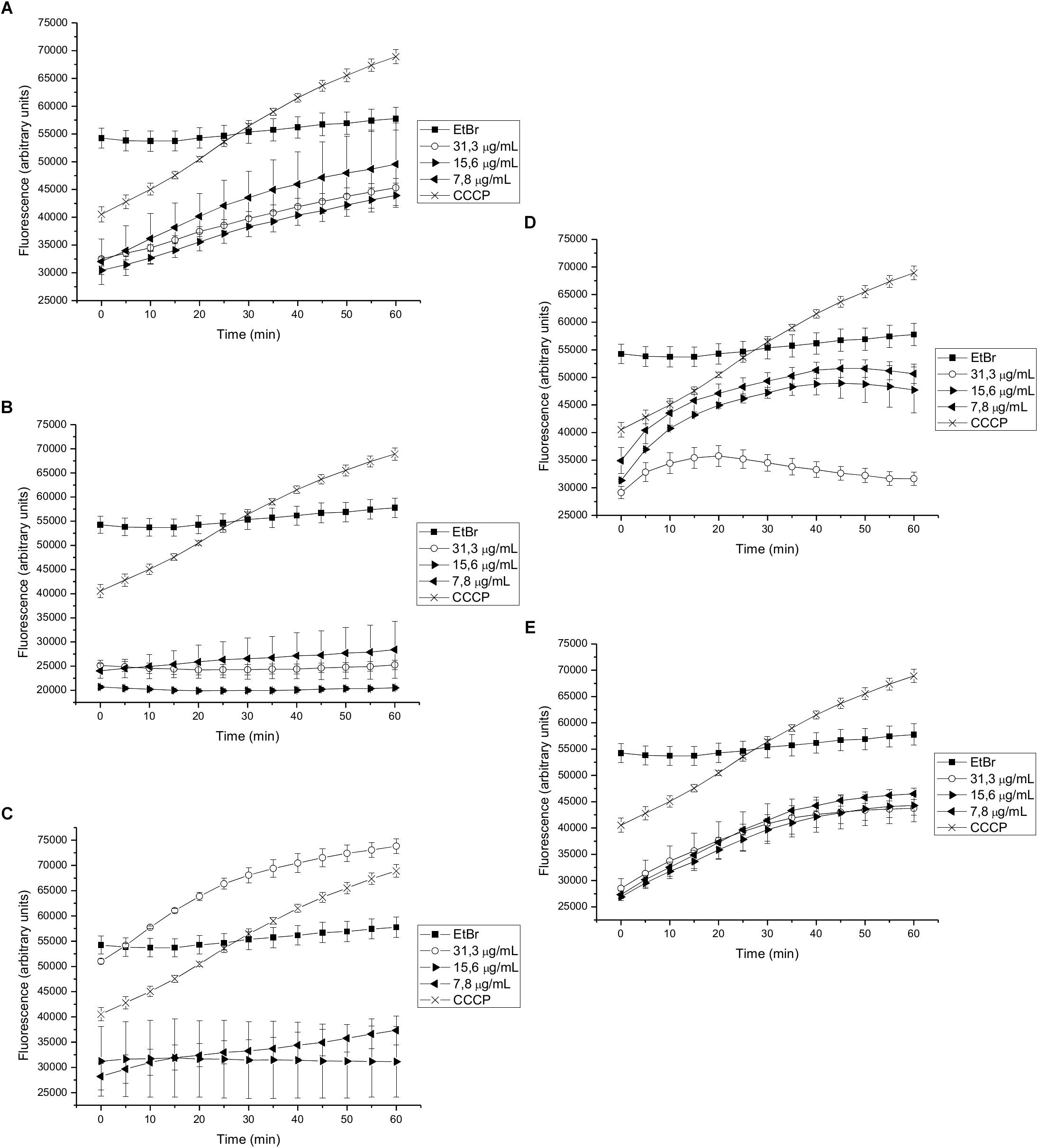
Figure 4. Effect on the intracellular accumulation of ethidium bromide of (A) DCME extract, (B) epi-cubenol (4), (C) 15-copaenol (6), (D) 15-copaenal (6OX), and (E) 15-copaenyl acetate (6AC) in K1902 (ΔnorA) strain. The bacteria were exposed to 2.0 μg/mL of EtBr at 37°C for a period of 1h in the presence of glucose. CCCP (1.25 μg/mL) was used as inhibitor control.
The results of synergism are shown in Tables 3, 4. The MICs of DCME, epi-cubenol (4) and 15-copaenol (6) varied in a range of 31.3–125 μg/mL in all strains studied. In general, the MIC values of ciprofloxacin were highest in norA++ (K2378) strain (2 μg/mL) in comparison with ΔnorA (K1902) strain (0.5 μg/mL), showing ciprofloxacin is substrate of NorA efflux pump. The MIC values of oxacillin did not show significative differences in strains studied (12.5–25 μg/mL). Additionally, was determined that all combinations between DCME extract or pure compounds [epi-cubenol (4) or 15-copaenol (6)] with ciprofloxacin or oxacillin decreased MICs values of both antibiotics in tested S. aureus strains (Table 3).
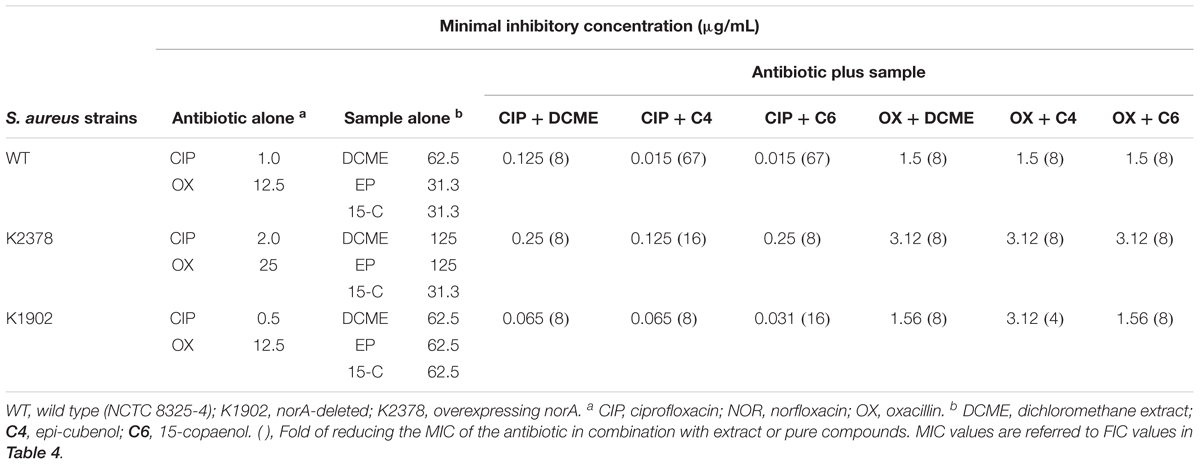
Table 3. Minimal inhibitory concentration of extract and isolated compounds from P. uviferum, and antibiotics and mixtures against S. aureus strains.
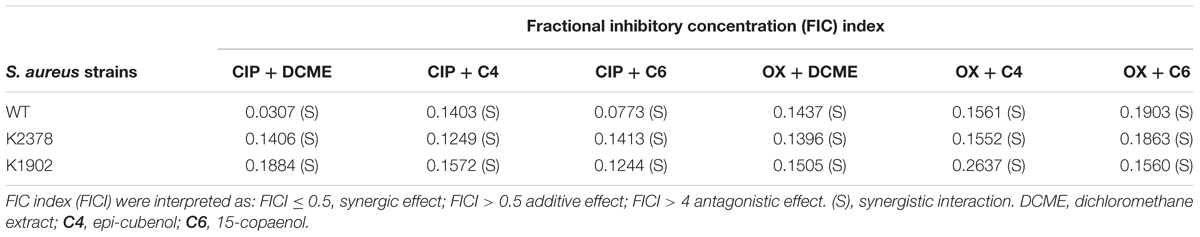
Table 4. Fractional inhibitory concentration index (FICI) of different combination between extract or isolated compounds of P. uviferum with antibiotics in S. aureus strains.
The largest reductions in the MIC value of ciprofloxacin were observed when it was mixed with epi-cubenol (4) and 15-copaenol (6), reaching 67-fold reduction of MIC values in wild type strain. While the combinations between DCME, epi-cubenol (4), 15-copaenol (6) and oxacillin reduced the MIC of this antibiotic 4–16 times fold. Fractional inhibitory concentration index (FICI) values obtained in the checkerboard assays were in the range of 0.0307–0.2637, indicating that all combinations between DCME, epi-cubenol (4) or 15-copaenol (6) and tested antibiotics have a synergistic effect (FICI ≤ 0.5) in wild type, norA++ and ΔnorA strains (Table 4).
Toxicity of DCME extract, epi-cubenol (4) and 15-copaenol (6) was evaluated due to the synergism they showed with ciprofloxacin and oxacillin, reducing the MIC of the antibiotics until 67-fold in the NCTC-8325-4 strain. In addition, DCME, epi-cubenol (4) and 15-copaenol (6) showed activity against efflux pumps in S. aureus.
The toxicity evaluation of DCME, epi-cubenol (4) and 15-copaenol (6) on HeLa cells is shown in Figure 5. When HeLa cell was treated with different concentrations of DCME (125–0.98 μg/mL; it was started from the MIC), the cell viability decreased in 7% compared to the control (98% cell viability), with cell viability percentages in a range of 92.3–91.1 in all concentrations tested. Whereas, when the HeLa cells were treated with the highest concentrations of epi-cubenol (4) (125 and 62.5 μg/mL), the viability percentages were reduced to 0.9 and 79.6%, respectively, compared to the control (98.3%). Nevertheless, low concentrations (31.3–1 μg/mL) increases cell viability up to 93.8%. 15-Copaenol showed the greatest toxicity at 62.5 μg/mL (MIC value) and 31.3 μg/mL, and the cell viability decreased by 0.7% and 2%, respectively. By decreasing the concentration to 15.6 and 3.9 μg/mL, the cell viability increased up 94.1%. Finally, using 15-copaenol (6) in concentrations of 1.95–0.49 μg/mL, the viability of HeLa cells increased, respectively, to 96.3% and 98%, significantly equal to the control.
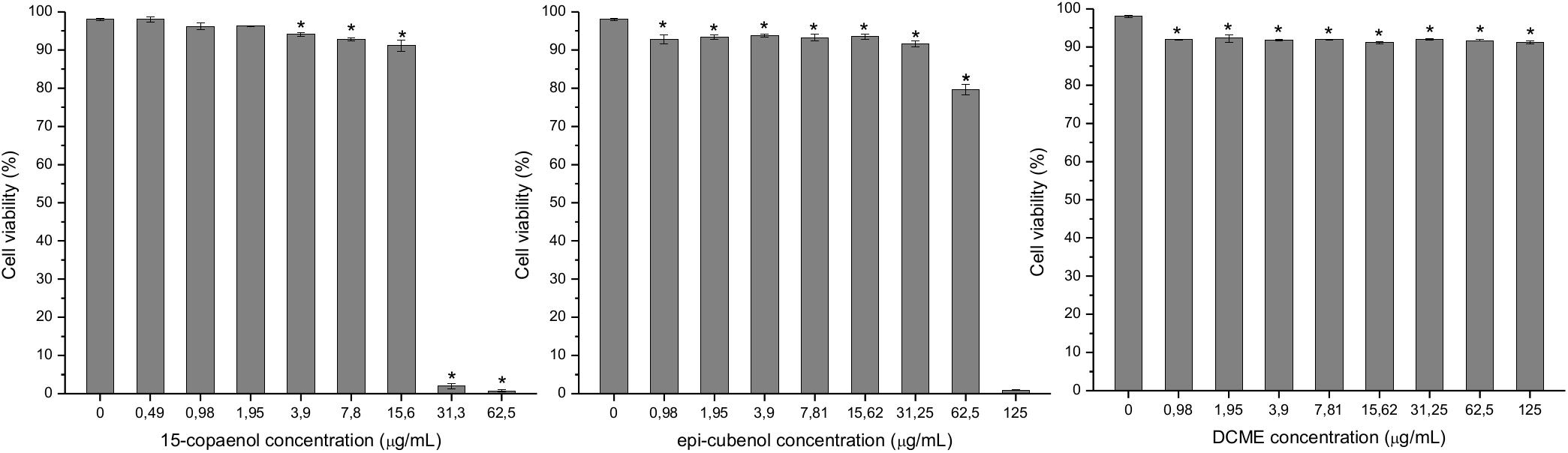
Figure 5. Effects of 15-copaenol, epi-cubenol, and DCME extract on HeLa cell viability. The HeLa cells were incubated with different concentration of samples in a range of 125–0.5 μg/mL. Results are the mean ± SE, n = 3. The asterisk indicates significantly different from the control at the same level (P < 0.05).
Extracts and compounds were evaluated as bacterial growth inhibitors against wild type, norA-deleted, and norA-overexpressing strains of S. aureus. According to Duarte et al. (2007); Wang et al. (2008), and Oliveira et al. (2013) plant extracts can be classified as antibacterial agents according to their MIC values; MIC < 500 μg/mL indicate strong inhibitors, MIC between 600 and 1,500 μg/mL indicate moderate inhibitors, and MIC > 1,600 μg/mL indicate weak inhibitors. Therefore, the EO and extracts from P. uviferum heartwood can be classified as strong inhibitors showing MIC range of 31.3–500 μg/mL in all evaluated S. aureus strains. Additionally, the analyses showed that the MIC values were the lowest in the K1902 (ΔnorA) strain. Wild type and K2378 (overexpressing norA) S. aureus possessed high levels of antimicrobial resistance because they were capable of reducing the intracellular antibacterial agent concentration via efflux, which was partly mediated by NorA pumps. These pumps were absent in K1902 norA-deleted (ΔnorA) cells.
In this study, the efflux pump activity was evaluated using a fluorometric method that measured the accumulation of the universal efflux pump substrate EtBr. Overexpressing efflux pumps is an important mechanism by which bacteria can evade the effects of antimicrobial agents. The management of efflux pumps is a valuable strategy to restore susceptibility in strains resistant to antibacterial agents. In this context, DCME and the compounds 15-copaenol (6) and epi-cubenol (4) acted as NorA EPIs, as shown by the finding that they produced an higher level of fluorescence over time than was observed for the EtBr curve obtained in untreated S. aureus strains. Moreover, 15-copaenol (6) was the only sample that was able to increase the EtBr accumulation in the K1902 (ΔnorA) strain. Because the norA gene was suppressed in the K1902 (ΔnorA) strain, K1902 cells cannot express the NorA efflux pump. However, other authors have demonstrated that K1902 cells were able to express other efflux pumps for which EtBr is a substrate, such as MepA, TetK, and MrsA (Jang, 2016). It should to be noted that, 15-copaenol (6) was able to increase the EtBr accumulation at all the used concentrations in wild type and K2378 S. aureus strains while it was able to increase the EtBr accumulation in K1902 only at 31.5 μg/mL. These results can be explained using the cytotoxicity experiments. 15-Copaenol (6) at 31.5 μg/mL was toxic for HeLa cells (Figure 5). Moreover, MIC of 15-copaenol (6) was 62.5 μg/mL in K1902 strain, but, according with cytotoxicity assays (Figure 5), clearly, at 31.5 μg/mL starts to damage membrane of K1902 cells, thus producing an increase in EtBr accumulation. Therefore, it suggests that 15-copaenol (6) at low concentrations likely could inhibit NorA efflux pumps and other efflux pumps, but at high concentrations (≥31.5 μg/mL) 15-copaenol (6) shows a non-specific effect (such as membrane depolarization) thereby resulting toxic for S. aureus strains: 8325-4, K2378 and K1902.
By following a structure-activity analysis, the EtBr accumulation observed in the K2378 (norA++) strain indicated that the hydroxylated compound 15-copaenol (6) exerted a stronger inhibitory effect than was found for the tertiary sesquiterpene alcohols epi-cubenol (4) and cubenol (3) and the sesquiterpene hydrocarbons trans-calamenene (1) and cadalene (2). Among the tertiary alcohols, epi-cubenol (4), which has a C1-α-OH, was more active than the C1-epimer, cubenol (3), which was inactive at all evaluated concentrations. Additionally, when the primary hydroxyl group of 15-copaenol (6) was replaced with an aldehyde or an ester group, resulting in the synthetic derivatives 6OX and 6AC, respectively, the modified derivatives did not act like a NorA EPI, indicating that the primary hydroxyl group is essential to generating the EtBr efflux inhibition of copaene-type sesquiterpenes in S. aureus K2378 cells. Consistent with these results, Vieira et al. (1991) observed a decrease in the antibacterial activity of 15-copaenol (6) when it was oxidized at C11 and C8 to aldehyde and ketone groups, respectively. Moreover, the MIC values obtained in this study (Table 2) are consistent with the results of the structure-antibacterial activity analyses performed in wild type and K2378 S. aureus. However, when we studied growth inhibition in the K2378 strain, we found that the MIC values for cubenol (3) and epi-cubenol (4) were the same. These data indicate that the C1-tertiary hydroxyl configuration was not involved in antibacterial activity. This analysis represents an initial approach to determining the structural characteristics of the cadalene-type sesquiterpenes that act to induce NorA efflux pump inhibition in S. aureus strains.
The results obtained in synergism assays, are coincident with those previous in vitro studies which showed a synergistic effect between plant extracts and/or isolated compounds with different classes of antibiotics against sensitive and multidrug resistant pathogenic strains. It has also been determined that these combinations reduced the minimum inhibitory concentration of antibiotics in different S. aureus strains (Chung et al., 2011; Langeveld et al., 2014; Bag and Chattopadhyay, 2015; Dong et al., 2017). In addition, Magi et al. (2015), described that carvacrol, the major component of the EOs of Origanum and Thymus plants act in synergism with erythromycin against 21 of 32 clinical erythromycin-resistant Streptococci, with reduction of the erythromycin MIC up to 2,048-fold.
Hemaiswarya et al. (2008); Wagner and Ulrich-Merzenich (2009), and Wagner (2011) have described four mechanisms by how antibiotic synergism with phytoextracts occurs: (a) multi objective effect, which is based on the fact that the action of the phytoextract affect several cellular targets, such as enzymes, receptors and substrates, among others; (b) pharmacokinetic effects based on the fact that components of the phytoextract help the solubility; (c) elimination or neutralization of toxic or adversely acting substances by one agent that has been added to an extract; (d) the interaction of the components of the phytoextract with the resistance mechanisms of the bacteria. Our results showed that DCME extract, epi-cubenol (4) and 15-copaenol (6) can act in synergy with antibiotics with different mechanisms of action. The ciprofloxacin (a fluoroquinolone) has as cellular target the DNA gyrase, enzyme involved in the synthesis of DNA, while oxacillin (a β-lactam) is involved in the inhibition of the peptidoglycan synthesis (synthesis of the cell wall). Then, these antibiotics in combination with extracts and pure compounds resulting in a multi-objective effect. However, the synergistic effect can be produced by more than one mechanism, for example a multi-objective effect and the inhibition of bacterial resistance mechanisms such as efflux pumps.
Moreover, according to ISO 10993-5 (2009), DCME extract, epi-cubenol (4), 15-copaenol (6) were not toxic for the HeLa cell line at concentrations ≤125 μg/mL, ≤62.5 μg/mL, and ≤15.6 μg/mL, respectively, in which the synergistic effect and inhibitory activity of efflux pumps was determined in S. aureus strains. However, cytotoxicity studies of sesquiterpenes are scarce. (+)-Torreyol (5) has showed low cytotoxicity (Yap et al., 2017), and cubenol (3) and cadalene (2) are not cytotoxic (Kouamé et al., 2013; Jiangseubchatveera et al., 2015).
Other terpenes are also capable of producing efflux pump inhibition. Oluwatuyi et al. (2004) demonstrated that the abietane diterpenes, carnosic acid and carnosol, which were isolated from Rosmarinus officinalis, had modest EPI activity. Carnosic acid decreased the MIC values of erythromycin against RN4220 (pUL5054; msrA++) by eightfold and caused a twofold decreased in the EtBr MIC against the SA-1199B strain of S. aureus, which overexpresses the norA gene that encodes the NorA MDR efflux pump (Sun et al., 2016), and that of tetracycline against the XU212 strain of S. aureus, which possesses the TetK tetracycline efflux protein (Sun et al., 2016). However, carnosic acid inhibited EtBr efflux in SA-1199B cells, suggesting that it has weak NorA-inhibitory activity. Carnosol was less active than carnosic acid, but when used in combination with tetracycline, it produced a fourfold decrease in the MIC against XU212 cells. The authors mentioned that high diterpene lipophilicity is a unique common characteristic of these factors and concluded that lipophilicity was essential to efflux pump activity in multi-resistant Gram-positive bacteria. However, in the present study, we found that the lipophilic sesquiterpenes trans-calamenene (1) and cadalene (2) were not NorA EPIs and that these isolates resulted in the highest MIC values in all of the evaluated strains. This finding suggests that other structural factors are needed to produce NorA efflux pump inhibition.
Ferruginol, a diterpene isolated from Chamaecyparis lawsoniana, was demonstrated to have an IC50 of 17 μM in an EtBr efflux-inhibition assay performed using SA-1199B cells (Yusa and Tsuruo, 1989). Both reserpine and ferruginol caused a fourfold reduction in the MIC of tetracycline in XU212 cells. Ferruginol also increased the activity of erythromycin by fourfold against S. aureus RN4220 cells (pUL5054; msrA++), suggesting that these cells exhibit a rarely observed MsrA inhibitory activity (Smith et al., 2007a). It should be noted that ferruginol and other similar compounds, such as 6,7-dehydroferruginol, hinokione, hinokiol, and sugiol, were present in the DCME and PEE isolated from P. uviferum. These compounds could be related to the inhibitory activity of DCME. Totarol, a phenolic diterpene isolated from Chamaecyparis nootkatensis, reduced by eightfold the MIC of EtBr against SA-K3092 (norA++); EtBr efflux was similarly inhibited by reserpine (Smith et al., 2007b). Interestingly, this concentration of totarol mediated an eightfold reduction in the erythromycin MIC in RN4220 cells (pUL5054), demonstrating that it exerts activity against MsrA (Schindler et al., 2013).
Currently, other types of secondary metabolites have also been suggested to act as EPIs in Gram-positive bacteria. Reserpine (an indole alkaloid), flavones, isoflavones, flavolignans, coumarins, galvanic acid (sesquiterpene-coumarin), porphyrins, and chalcones, among others, have been described as EPIs (Markham et al., 1999; Gibbons et al., 2003; Stavri et al., 2007; Smith et al., 2007a; Schindler et al., 2013; Hazra et al., 2018). Unfortunately, the substantial structural heterogeneity of those secondary metabolites has not allowed a correlation analysis of the structural requirements of efflux pump inhibition. In addition, none of these potential EPIs has been approved for clinical use, partly because their inhibitory mechanisms have not been clarified (Sun et al., 2014). Future research should include detailed structure-activity relationship (SAR) studies, which today are very scarce, as the results of such studies could lead researchers to discover new EPIs.
JE, AU, LS, and MWi designed the research. JE and AU analyzed chemical data. LS and MWa performed bioassays. PM performed the cytotoxicity test. LS, MWa, and JE performed statistical analyses. JE, AU, LS, PF, and LM wrote the manuscript. All authors read and approved the final manuscript.
This research was funded by CONICYT-PCHA grant no. 2012-21120365, CONICYT-Postdoctoral FONDECYT grants no. 3180532 and USACH Proyecto Basal USA 1799.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors would like to express their gratitude for CONICYT-PCHA grant no. 2012-21120365 (awarded to JE) and CONICYT-postdoctoral FONDECYT grant no. 3180532 (awarded to JE). JE is grateful to Vicerrectoría de Investigación y Postgrado, Universidad de La Frontera, UFRO; AU is grateful to Proyecto Basal USA 1799, Universidad de Santiago de Chile, USACH; and LS is grateful to Fondo de Desarrollo a la Publicación (FDP), Universidad Mayor 2017–2018; Specifically, the authors would like to thank Dr. Glenn Kaatz of the Division of Infectious Diseases, School of Medicine, Wayne State University for donating the S. aureus strains (NCTC-8325-4, K2378, and K1902).
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2019.00337/full#supplementary-material
Akanbi, O. E., Njom, H. A., Fri, J., Otigbu, A. C., and Clarke, A. M. (2017). Antimicrobial susceptibility of Staphylococcus aureus isolated from recreational waters and beach sand in eastern Cape province of South Africa. Int. J. Environ. Res. Public Health 14:1001. doi: 10.3390/ijerph14091001
Astolfi, A., Felicetti, T., Iraci, N., Manfroni, G., Massari, S., Pietrella, D., et al. (2017). Pharmacophore-based repositioning of approved drugs as novel Staphylococcus aureus NorA efflux pump inhibitors. J. Med. Chem. 60, 1598–1604. doi: 10.1021/acs.jmedchem.6b01439
Augustin, J., Rosenstein, R., Wieland, B., Schneider, U., Schnell, N., Engelke, G., et al. (1992). Genetic analysis of epidermin biosynthetic genes and epidermin-negative mutants of Staphylococcus epidermidis. Eur. J. Biochem. 204, 1149–1154. doi: 10.1111/j.1432-1033.1992.tb16740.x
Bag, A., and Chattopadhyay, R. R. (2015). Evaluation of synergistic antibacterial and antioxidant efficacy of essential oils of spices and herbs in combination. PLoS One 10:e0131321. doi: 10.1371/journal.pone.0131321
Caspar, Y., Jeanty, M., Blu, J., Burchak, O., Le Pihive, E., Maigre, L., et al. (2015). Novel synthetic bis-indolic derivatives with antistaphylococcal activity, including against MRSA and VISA strains. J. Antimicrob. Chemother. 70, 1727–1737. doi: 10.1093/jac/dkv015
Chan, B., Ip, M., Lau, C., Lui, S., Jolivat, C., Ganem- Elbaz, C., et al. (2011). Synergistic effects of baicalein with ciprofloxacin against NorA overexpressed methicillin-resistant Staphylococcus aureus (MRSA) and inhibition of MRSA pyruvate kinase. J. Ethnopharmacol. 137, 767–773. doi: 10.1016/j.jep.2011.06.039
Chung, P. Y., Navaratnam, P., and Chung, L. Y. (2011). Synergistic antimicrobial activity between pentacyclic triterpenoids and antibiotics against Staphylococcus aureus strains. Ann. Clin. Microbiol. Antimicrob. 10, 1–6. doi: 10.1186/1476-0711-10-25
Clinical and Laboratory Standards Institute [CLSI] (2005). Performance Standards for Antimicrobial Susceptibility Testing: Fifteenth Informational Supplement M100-S15. Wayne, PA: CLSI.
Costa, L. M., de Macedo, E. V., Oliveira, F. A., Ferreira, J. H., Gutierrez, S. J., Peláez, W. J., et al. (2016). Inhibition of the NorA efflux pump of Staphylococcus aureus by synthetic riparins. J. Appl. Microbiol. 121, 1312–1322. doi: 10.1111/jam.13258
Dong, C. H., Li, L. X., Cui, Z. H., Chen, S. W., Xiong, Y. Q., Lu, J. Q., et al. (2017). Synergistic effect of pleuromutilins with other antimicrobial agents against Staphylococcus aureus in vitro and in experimental Galleria mellonella model. Front. Pharmacol. 8:553. doi: 10.3389/fphar.2017.00553
Donoso, C. A., Becerra, J., Bittner, M., Elissetche, J. P., Freer, J., Mendoza, R., et al. (2008). Allelochemicals and natural durability in Chilean Cupressaceae heartwoods. Allelopathy J. 21, 119–132.
dos Santos, J. F. S., Tintino, S. R., de Freitas, T. S., Campina, F. F., Menezesc, I. R. D. A., Siqueira-Júniord, J. P., et al. (2018). In vitro e in silico evaluation of the inhibition of Staphylococcus aureus efflux pumps by caffeic and gallic acid. Comp. Immunol. Microbiol. Infect. Dis. 57, 22–28. doi: 10.1016/j.cimid.2018.03.001
Duarte, M. C. T., Leme, E. E., Delarmelina, C., Soares, A. A., Figueira, G. M., and Sartoratto, A. (2007). Activity of essential oils from Brazilian medicinal plants on Escherichia coli. J. Ethnopharmacol. 111, 197–201. doi: 10.1016/j.jep.2006.11.034
Espinoza, J., Urzúa, A., Bardehle, L., Quiroz, A., Echeverría, J., and González-Teuber, M. (2018). Antifeedant effects of essential oil, extracts, and isolated sesquiterpenes from Pilgerodendron uviferum (D. Don) florin heartwood on red clover borer Hylastinus obscurus (Coleoptera: Curculionidae). Molecules 23:1282. doi: 10.3390/molecules23061282
Espinoza, J., Urzúa, A., Tampe, J., Parra, L., and Quiroz, A. (2016). Repellent activity of the essential oil from the heartwood of Pilgerodendron uviferum (D. Don) Florin against Aegorhinus superciliosus (Coleoptera: Curculionidae). Molecules 21:533. doi: 10.3390/molecules21040533
Felicetti, T., Cannalire, R., Pietrella, D., Latacz, G., Lubelska, A., Manfroni, G., et al. (2018). 2-Phenylquinoline S. aureus NorA Efflux Pump Inhibitors: Evaluation of the importance of methoxy group introduction. J. Med. Chem. 61, 7827–7848. doi: 10.1021/acs.jmedchem.8b00791
Foster, T. J. (2017). Antibiotic resistance in Staphylococcus aureus. Current status and future prospects. FEMS Microbiol. Rev. 41, 430–449. doi: 10.1093/femsre/fux007
Garvey, M., Rahman, M., Gibbons, S., and Piddock, L. (2011). Medicinal plants extracts with efflux inhibitory activity against Gram-negative bacteria. Int. J. Antimicrob. Agents 37, 145–151. doi: 10.1016/j.ijantimicag.2010.10.027
Gibbons, S., Oluwatuyi, M., and Kaatz, G. W. (2003). Novel inhibitor of multidrug efflux pumps in Staphylococcus aureus. J. Antimicrob. Chemother. 51, 13–17. doi: 10.1093/jac/dkg044
Handzlik, J., Matys, A., and Kieć-Kononowicz, K. (2013). Recent advances in multi-drug resistance (MDR) efflux pump inhibitors of gram-positive bacteria S. aureus. Antibiotics 2, 28–45. doi: 10.3390/antibiotics2010028
Harding, K. E., May, L. M., and Dick, K. F. (1975). Selective oxidation of allylic alcohols with chromic acid. J. Org. Chem. 40, 1664–1665. doi: 10.1021/jo00899a040
Hazra, B. M., Sutapa, B., Ghosh, S., and Avlani, D. (2018). “Synergistic interaction between plant products and antibiotics against potential pathogenic bacteria,” in Frontiers in Anti-Infective Drug Discovery, eds M. Atta-ur-Rahman, I. Choudhary, and A. N. Rezitis (Emirate of Sharjah: Bentham Science Publishers), 88–152.
Hemaiswarya, S., Kumar, A., and Doble, M. (2008). Synergism between natural products and antibiotics against infectious diseases. Phytomedicine 15, 639–652. doi: 10.1016/j.phymed.2008.06.008
Hiramatsu, K., Katayama, Y., Matsuo, M., Sasaki, T., Morimoto, Y., Sekiguchi, A., et al. (2014). Multi-drug-resistant Staphylococcus aureus and future chemotherapy. J. Infect. Chemother. 20, 593–601. doi: 10.1016/j.jiac.2014.08.001
ISO 10993-5. (2009). Biological Evaluation of Medical Devices. Part 5: Tests for in Vitro Cytotoxicity. Geneva: ISO.
Jang, S. (2016). Multidrug efflux pumps in Staphylococcus aureus and their clinical implications. J. Microbiol. 54, 1–8. doi: 10.1007/s12275-016-5159-z
Jiangseubchatveera, N., Liawruangrath, B., Liawruangrath, S., Korth, J., and Pyne, S. G. (2015). The chemical constituents and biological activities of the essential oil and the extracts from leaves of Gynura divaricata (L.) DC. growing in Thailand. J. Essent. Oil Bear. Plants 18, 543–555. doi: 10.1080/0972060X.2014.935016
Jo, A., and Ahn, J. (2016). Phenotypic and genotypic characterization of multiple antibiotic-resistant Staphylococcus aureus exposed to subinhibitory levels of oxacillin and levofloxacin. BMC Microbiol. 16:170. doi: 10.1186/s12866-016-0791-7
Karmakar, A., Dua, P., and Ghosh, C. (2016). Biochemical and molecular analysis of Staphylococcus aureus clinical isolates from hospitalized patients. Can. J. Infect. Dis. Med. Microbiol. 2016:9041636. doi: 10.1155/2016/9041636
Khan, S., and Khan, A. (2016). Breaking the spell: combating multidrug resistant ìsuperbugs ì. Front. Microbiol. 7:174. doi: 10.3389/fmicb.2016.00174
Kouamé, P. B., Jacques, C., Bedi, G., Silvestre, V., Loquet, D., Barilleì-Nion, S., et al. (2013). Phytochemicals isolated from leaves of Chromolaena odorata: impact on viability and clonogenicity of cancer cell lines. Phytother. Res. 27, 835–840. doi: 10.1002/ptr.4787
Langeveld, W. T., Veldhuizen, E. J., and Burt, S. A. (2014). Synergy between essential oil components and antibiotics: a review. Crit. Rev. Microbiol. 40, 76–94. doi: 10.3109/1040841X.2013.763219
Lowrence, R. C., Raman, T., Makala, H. V., Ulaganathan, V., Subramaniapillai, S. G., Kuppuswamy, A. A., et al. (2016). Dithiazole thione derivative as competitive NorA efflux pump inhibitor to curtail multi drug resistant clinical isolate of MRSA in a zebrafish infection model. Appl. Microbiol. Biotechnol. 100, 9265–9281. doi: 10.1007/s00253-016-7759-2
Magi, G., Marini, E., and Facinelli, B. (2015). Antimicrobial activity of essential oils and carvacrol, and synergy of carvacrol and erythromycin, against clinical, erythromycin-resitant group A Streptococci. Front. Microbiol. 6:165. doi: 10.3389/fmicb.2015.00165
Marinelli, F., and Tomazs, A. (2010). Antimicrobials. Curr. Opin. Microbiol. 13, 547–550. doi: 10.1016/j.mib.2010.09.010
Markham, P. N., Westhaus, E., Klyachko, K., Johnson, M. E., and Neyfakh, A. A. (1999). Multiple novel inhibitors of the NorA multidrug transporter of Staphylococcus aureus. Antimicrob. Agents Chemother. 43, 2404–2408. doi: 10.1128/AAC.43.10.2404
Meinwald, J., Crandall, J., and Hymans, W. E. (1973). Nortricyclanone [Tricyclo[2.2.1.02,6]heptan-3-one]. Org. Syn. Coll. 5:866. doi: 10.15227/orgsyn.045.0077
Motyl, M., Dorso, K., Barret, J., and Giacobbe, R. B. (2005). Basic microbiological techniques for antibacterial drug discovery. Curr. Protoc. Pharmacol. 31, 13A.3.1–13A.3.22.
Oliveira, D. A., Salvador, A. A., Smânia, A. Jr., Smânia, E. F., Maraschin, M., and Ferreira, S. R. (2013). Antimicrobial activity and composition profile of grape (Vitis vinifera) pomace extracts obtained by supercritical fluids. J. Biotechnol. 164, 423–432. doi: 10.1016/j.jbiotec.2012.09.014
Oluwatuyi, M., Kaatz, G. W., and Gibbons, S. (2004). Antibacterial and resistance modifying activity of Rosmarinus officinalis. Phytochemistry 65, 3249–3254. doi: 10.1016/j.phytochem.2004.10.009
Oyarzún, M. L., and Garbarino, J. A. (1988). Sesquiterpenoids from Pilgerodendron uvífera. Phytochemistry 27, 1121–1123. doi: 10.1016/0031-9422(88)80286-3
Paixão, L., Rodrigues, L., Couto, I., Fernandes, P., De Carvalho, C., Monteiro, G. A., et al. (2009). Fluorometric determination of ethidium bromide efflux kinetics in Escherichia coli. J. Biol. Engi. 3, 18–30. doi: 10.1186/1754-1611-3-18
Reddy, P. N., Srirama, K., and Dirisala, V. R. (2017). An update on clinical burden, diagnostic tools, and therapeutic options of Staphylococcus aureus. Infect. Dis. 10:1179916117703999. doi: 10.1177/1179916117703999
Santos-Costa, S., Viveiros, M., Amaral, L., and Couto, I. (2013). Multidrug efflux pumps in Staphylococcus aureus: an update. Open Microbiol. J. 7, 59–71. doi: 10.2174/1874285801307010059
Schindler, B. D., Jacinto, P., and Kaatz, G. W. (2013). Inhibition of drug efflux pumps in Staphylococcus aureus: current status of potentiating existing antibiotics. Fut. Microbiol. 8, 491–507. doi: 10.2217/fmb.13.16
Singh, S., Kalia, N., Joshi, P., Kumar, A., Sharma, P. R., Kumar, A., et al. (2017). Boeravinone B, novel dual inhibitors of NorA bacterial efflux pump of Staphylococcus aureus and human P-glycoprotein, reduces the biofilm formation and intracelular invasión of bacteria. Front. Microbiol. 8:1868. doi: 10.3389/fmicb.2017.01868
Smith, E. C., Kaatz, G. W., Seo, S. M., Wareham, N., Williamson, E. M., and Gibbons, S. (2007a). The phenolic diterpene totarol inhibits multidrug efflux pump activity in Staphylococcus aureus. Antimicrob. Agents Chemother. 51, 4480–4483.
Smith, E. C., Williamson, E. M., Wareham, N., Kaatz, G. W., and Gibbons, S. (2007b). Antibacterials and modulators of bacterial resistance from the immature cones of Chamaecyparis lawsoniana. Phytochemistry 68, 210–217.
Solis, C., Becerra, J., Flores, C., Rebolledo, J., and Silva, M. (2004). Antibacterial and antifungal terpenes from Pilgerodendron uviferum (D. Don) Florin. J. Chil. Chem. Soc. 49, 157–161. doi: 10.4067/S0717-97072004000200010
Stavri, M., Piddock, L., and Gibbons, S. (2007). Bacterial efflux pump inhibitors from natural sources. J. Antimicrob. Chemother. 59, 1247–1260. doi: 10.1093/jac/dkl460
Stermitz, F., Lorenz, P., Tawara, J., Zenewiez, L., and Lewis, K. (2000). Synergy and medicinal plant. Antimicrobial action of berberine potentiated by 5-methoxy hidnocarpin, a multidrug pump inhibitor. Proc. Natl. Acad. Sci. U.S.A. 97, 1433–1437. doi: 10.1073/pnas.030540597
Sun, J., Deng, Z., and Yan, A. (2014). Bacterial multidrug efflux pumps: mechanisms, physiology and pharmacological exploitations. Biochem. Biophys. Res. Commun. 453, 254–267. doi: 10.1016/j.bbrc.2014.05.090
Sun, Z.-L., He, J.-M., Wang, S.-Y., Ma, R., Khondkar, P., Kaatz, G. W., et al. (2016). Benzocyclohexane oxide derivatives and neolignans from Piper betle inhibit efflux-related resistance in Staphylococcus aureus. RSC Adv. 6, 43518–43525. doi: 10.1039/C6RA10199B
Tegos, G., Haynes, M., Strouse, J., Mohiuddin, M., Khan, T., Bologa, C., et al. (2011). Microbial effux pump inhibition: tactis and strategies. Curr. Pharm. Des. 17, 1291–1302. doi: 10.2174/138161211795703726
Thurlow, L., Joshi, G., and Richardson, A. (2012). Virulence strategies of the dominant USA 300 lineage of community associated meticillin resistant Staphylococcus aureus (CA-MRSA). FEMS Immunol. Med. Microbiol. 65, 1–27. doi: 10.1111/j.1574-695X.2012.00937.x
Tong, S. Y. C., Davis, J. S., Eichenberger, E., Holland, T. L., and Fowler, V. G. Jr. (2015). Staphylococcus aureus infections: epidemiology, pathophysiology, clinical manifestations, and management. Clin. Microbiol. Rev. 28, 603–661. doi: 10.1128/CMR.00134-14
Urzúa, A., Rezende, M. C., Mascayano, C., and Vásquez, L. (2008). A Structure-Activity Study of Antibacterial Diterpenoids. Molecules 13, 882–891. doi: 10.3390/molecules13040822
Van Bambeke, F., Pagés, J., and Lee, V. (2006). Inhibitors of bacterial effux pumps as adjuvants in antibiotics treatments and diagnostic tools for detection of resistance by efflux. Recent Pat. Antiinfect. Drug Discov. 1, 157–175. doi: 10.2174/157489106777452692
Vieira, P. C., Himejima, M., and Kubo, I. (1991). Sesquiterpenoids from Brachylaena hutchznszz. J. Nat. Prod. 54, 416–420. doi: 10.1021/np50074a011
Wagner, H. (2011). Synergy research: approaching a new generation of phytopharmaceuticals. Fitoterapia 82, 34–37. doi: 10.1016/j.fitote.2010.11.016
Wagner, H., and Ulrich-Merzenich, G. (2009). Synergy research: approaching a new generation of phytopharmaceuticals. Phytomedicine 16, 97–110. doi: 10.1016/j.phymed.2008.12.018
Walsh, C. (2000). Molecular mechanisms that confer antibacterial drug resistance. Nature 406, 775–781. doi: 10.1038/35021219
Wang, Y. S., He, H. P., Yang, J. H., Di, Y. T., and Hao, X. J. (2008). New monoterpenoid coumarins from Clausena anisum olens. Molecules 13, 931–937. doi: 10.3390/molecules13040931
Weyerstahl, P., Marschall, H., Christiansen, C., and Seelmann, I. (1996). On Some Italicene Derivatives. Liebigs Ann. 1996, 1641–1644. doi: 10.1002/jlac.199619961022
Yap, H.-Y. Y., Muria-Gonzalez, M. J., Kong, B. H., Stubbs, K. A., Tan, C. S., Ng, S. T., et al. (2017). Heterologous expression of cytotoxic sesquiterpenoids from the medicinal mushroom Lignosus rhinocerotis in yeast. Microb Cell Fact. 16:103. doi: 10.1186/s12934-017-0713-x
Yusa, K., and Tsuruo, T. (1989). Reversal mechanism of multidrug resistance by verapamil: direct binding of verapamil to P-glycoprotein on specific sites and transport of verapamil outward across the plasma membrane of K562/ADM cells. Cancer Res. 49, 5002–5006.
Keywords: antibiotic resistance, efflux pump, NorA, efflux pumps inhibitors, sesquiterpenes, 15-copaenol
Citation: Espinoza J, Urzúa A, Sanhueza L, Walter M, Fincheira P, Muñoz P, Mendoza L and Wilkens M (2019) Essential Oil, Extracts, and Sesquiterpenes Obtained From the Heartwood of Pilgerodendron uviferum Act as Potential Inhibitors of the Staphylococcus aureus NorA Multidrug Efflux Pump. Front. Microbiol. 10:337. doi: 10.3389/fmicb.2019.00337
Received: 30 October 2018; Accepted: 08 February 2019;
Published: 26 February 2019.
Edited by:
Ren-You Gan, Shanghai Jiao Tong University, ChinaReviewed by:
Jianhua Wang, Gene Engineering Laboratory, Feed Research Institute (CAAS), ChinaCopyright © 2019 Espinoza, Urzúa, Sanhueza, Walter, Fincheira, Muñoz, Mendoza and Wilkens. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Javier Espinoza, amF2aWVyLmVzcGlub3phQHVmcm9udGVyYS5jbA== Alejandro Urzúa, YWxlamFuZHJvLnVyenVhQHVzYWNoLmNs Loreto Sanhueza, bG9yZXRvLnNhbmh1ZXphQHVtYXlvci5jbA==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.