
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 12 December 2018
Sec. Extreme Microbiology
Volume 9 - 2018 | https://doi.org/10.3389/fmicb.2018.03059
 Stephan Christel1*
Stephan Christel1* Malte Herold2
Malte Herold2 Sören Bellenberg1,3
Sören Bellenberg1,3 Antoine Buetti-Dinh4,5Mohamed El Hajjami6
Antoine Buetti-Dinh4,5Mohamed El Hajjami6 Igor V. Pivkin4,5
Igor V. Pivkin4,5 Wolfgang Sand3,7,8
Wolfgang Sand3,7,8 Paul Wilmes2Ansgar Poetsch6,9
Paul Wilmes2Ansgar Poetsch6,9 Mario Vera10,11
Mario Vera10,11 Mark Dopson1
Mark Dopson1Bioleaching is an emerging technology, describing the microbially assisted dissolution of sulfidic ores that provides a more environmentally friendly alternative to many traditional metal extraction methods, such as roasting or smelting. Industrial interest is steadily increasing and today, circa 15–20% of the world’s copper production can be traced back to this method. However, bioleaching of the world’s most abundant copper mineral chalcopyrite suffers from low dissolution rates, often attributed to passivating layers, which need to be overcome to use this technology to its full potential. To prevent these passivating layers from forming, leaching needs to occur at a low oxidation/reduction potential (ORP), but chemical redox control in bioleaching heaps is difficult and costly. As an alternative, selected weak iron-oxidizers could be employed that are incapable of scavenging exceedingly low concentrations of iron and therefore, raise the ORP just above the onset of bioleaching, but not high enough to allow for the occurrence of passivation. In this study, we report that microbial iron oxidation by Sulfobacillus thermosulfidooxidans meets these specifications. Chalcopyrite concentrate bioleaching experiments with S. thermosulfidooxidans as the sole iron oxidizer exhibited significantly lower redox potentials and higher release of copper compared to communities containing the strong iron oxidizer Leptospirillum ferriphilum. Transcriptomic response to single and co-culture of these two iron oxidizers was studied and revealed a greatly decreased number of mRNA transcripts ascribed to iron oxidation in S. thermosulfidooxidans when cultured in the presence of L. ferriphilum. This allowed for the identification of genes potentially responsible for S. thermosulfidooxidans’ weaker iron oxidation to be studied in the future, as well as underlined the need for new mechanisms to control the microbial population in bioleaching heaps.
Biomining is a sustainable process for metal extraction from sulfidic ores that has been studied by researchers around the globe since its emergence in the early 1950s (Temple and Colmer, 1951; Bryner and Jameson, 1958). In the recent decades and with its industrial application in mind, understanding of this natural process has significantly improved (Rohwerder et al., 2003; Watling, 2006; Vera et al., 2013; Jerez, 2017) and today, biomining is defined to be the microbial promoted oxidation of insoluble metal sulfides to acid soluble sulfates. In a technique termed bioleaching, this is undertaken to solubilize and recover metals of interest that form part of the metal sulfide mineral matrix. Ferrous iron (Fe2+)-oxidizing acidophilic microorganisms are responsible for the regeneration of the chemical oxidant ferric iron (Fe3+), which in turn attacks the sulfidic mineral, and breaks its covalent bonds. This releases ferrous iron plus any other contained metals and completes the catalytic cycle (Vera et al., 2013). While having initial economic disadvantages, mainly attributed to the long lag phase after construction of a bioleaching heap, biomining technologies are commonly considered more environmentally friendly than most conventional methods (Johnson, 2014). Today, increasing amounts of metals are extracted or processed by biomining technologies in many countries that include Chile, Australia, and South Africa, with the bioleaching of secondary copper sulfides accounting for an estimated 15–20% of the world wide copper production (Brierley and Brierley, 2013).
Bioleaching of primary copper minerals, such as the world’s most abundant copper mineral chalcopyrite (CuFeS2), remains challenging and suffers from slow dissolution rates. This is often attributed to the formation of passivation layers on the mineral surface (Cordoba et al., 2009; Wang et al., 2016), but it has also been argued that the semiconductor properties of chalcopyrite itself could be responsible (Crundwell, 2015). To date, extensive efforts to elucidate the exact nature of chalcopyrite passivation have not been successful (Khoshkhoo et al., 2014a, 2017). Despite this, strategies have been discovered to diminish the passivating effect, including bioleaching at high temperatures and low redox potentials (Li et al., 2013; Panda et al., 2015). Due to their large concentrations, the oxidation/reduction potential (ORP) in bioleaching systems is predominantly determined by the Fe3+/Fe2+ redox couple, whereby high concentrations of Fe3+ indicate high potentials. At low redox potentials and in the presence of millimolar concentrations of Fe2+ and Cu2+, chalcopyrite is suggested to be transformed into the secondary copper sulfide chalcocite (Cu2S) which is more readily oxidized by the Fe3+ provided by microbial action (Hiroyoshi et al., 2013). Methods to control the redox potential of the leaching solution include the addition of chemical reductants (Zhao et al., 2017) or limitation of oxygen (Third et al., 2002; Spolaore et al., 2009; Gericke et al., 2010). However, the technical realization of such methods in a large industrial bioheap with gradients of, e.g., temperature, oxygen concentration, and substrates has not been accomplished. Many studies have investigated the optimal microbial consortia in bioleaching operations (Rawlings and Johnson, 2007), usually focusing on the need to efficiently oxidize Fe2+ that drives the redox potential above that optimal for chalcopyrite dissolution (Hiroyoshi et al., 2013). In contrast, little attention has been paid to the possibility of controlling the redox potential of a bioleaching system by influencing the ratio of ferric to ferrous iron via suitable iron-oxidizing microbes (Masaki et al., 2018).
A large range of acidophile microbes have the capability to oxidize Fe2+ to gain energy under acidic conditions (Hedrich et al., 2011) and are therefore applicable in biomining operations. Among those are members of the Acidithiobacillus, Acidimicrobium, Acidiferrobacter, Sulfobacillus, and Ferroplasma genera (reviewed in Quatrini and Johnson, 2016). In bioleaching systems, one of the most abundant iron oxidizers is the moderately thermophilic, chemolithoautotroph Leptospirillum ferriphilum (Penev and Karamanev, 2010; Christel et al., 2017). This moderate thermophile solely derives its energy from the oxidation of ferrous iron (Coram and Rawlings, 2002) and is capable of doing so at very low Fe2+ ion concentrations and redox potentials as high as 700 mV vs. Ag/AgCl (Rawlings et al., 1999), giving it a significant advantage over other species. Another iron-oxidizer commonly found in acidic, sulfur rich environments is the moderately thermophilic Sulfobacillus thermosulfidooxidans (Karavaiko et al., 2005) that in contrast to L. ferriphilum, is unable to scavenge exceedingly scarce ferrous iron and is therefore considered a “weak” iron oxidizer in this study. In addition to Fe2+, S. thermosulfidooxidans is capable of oxidizing inorganic sulfur compounds (ISCs) and can utilize organic molecules to meet its carbon demands (Tsaplina et al., 2000). ISC oxidation is an important process in bioleaching heaps to remove excess sulfur compounds (Dopson and Lindstrom, 1999) and generate the necessary acidity, which is otherwise consumed by gangue minerals in low grade ores (Baldi et al., 1991; Dopson et al., 2009). Often, this role is fulfilled by obligate ISC-oxidizing species, such as the mesophile Acidithiobacillus thiooxidans or moderately thermophile A. caldus (Hallberg and Lindstrom, 1994).
In this study, we hypothesized that by inoculation of chalcopyrite ore with suitable iron-oxidizing bacteria the redox potential of the leachate in the initial phase of bioleaching experiments can be controlled. By these means, the redox potential can be maintained close to the optimum range. The initial rate of chalcopyrite dissolution is enhanced and thereby increases the amount of released copper. The applicability of this approach to industrial bioleaching operations is discussed.
Chalcopyrite was provided by Boliden AB (Sweden) and originates from the Aitik copper mine (N 67° 4′ 24″, E 20° 57′ 51″), where traces of bornite (Cu5FeS4) are frequently found accompanying this mineral. The flotation concentrate used in this study was revealed to be of high purity (>98%) by aqua regia digestion and elemental analysis and contained 29.5% copper (Supplementary File 1). For bioleaching experiments, the concentrate was sieved to obtain the size fraction between 50 and 100 μm, and subsequently washed in three volumes of 0.1 M EDTA in 0.4 M NaOH for 10 min under stirring to remove iron and copper compounds originating from weathering of the mineral. Elemental sulfur was then removed from the surfaces by three iterations of washing with one volume of acetone. Finally, the mineral was dried at 60°C overnight and then sterilized at 120°C for 10 h under a nitrogen atmosphere to ensure no changes in the mineral structure occurred.
Three bacterial acidophile species were used in this study, L. ferriphilum DSM 14647T, S. thermosulfidooxidans DSM 9293T, and A. caldus DSM 8584T. Prior to the bioleaching experiments, cells were maintained in three separate axenic continuous cultures so that the cells were under the same growth state when all experiments were inoculated. The continuous cultures were maintained at 38°C, fed with MAC medium (Mackintosh, 1978), and electron donor added in the form of 100 mM ferrous sulfate (L. ferriphilum) or 5 mM potassium tetrathionate (S. thermosulfidooxidans and A. caldus). The continuous culture vessels, all tubing, plus MAC medium were autoclaved while the ferrous sulfate and potassium tetrathionate were sterile filtered (0.2 μm pore size, cellulose acetate filter, PALL).
Bioleaching experiments were conducted in quadruplets in 250 mL Erlenmeyer flasks. 100 mL MAC medium (adjusted to pH 1.8 by addition of sulfuric acid) was supplemented with 2% (wt/vol) chalcopyrite concentrate and inoculated with combinations of the three bacterial species (107 cells per mL per species), obtained by centrifugation from the continuous cultures (12,500 ×g, 20 min) followed by cell counting using a Neubauer Improved counting chamber. Combinations included three single, three binary, and one tertiary combination, plus one sterile control. Cultures were incubated at 38 ± 2°C under slow shaking (120 rpm). Experiments were terminated 14 days after the first onset of microbial oxidation of ferrous iron as indicated by a redox potential >400 mV vs. Ag/AgCl, resulting in total incubation times ranging from 14 to 20 days.
Experiments were analyzed for pH (pHenomenal® 221, VWR), redox potential (Ag/AgCl with 3 M KCl; InLab® Redox-L, Mettler-Toledo), ferrous iron, total dissolved sulfur, elemental sulfur, as well as total iron and copper concentration in the leach liquor. Ferrous iron concentration was assessed by titration of its 1,10-phenanthroline complex (Walden et al., 1933; Dopson and Lindstrom, 1999). In short, 200 μL of bioleaching sample was centrifuged for 5 min at 16,000 ×g. Supernatant was mixed with the same volume of 15 mM 1,10-phenanthroline in aqueous 5 mM FeSO4, added to 1 mL of 1 M H2SO4 and subsequently titrated from orange to blue with 1 mM CeSO4. Total soluble and elemental sulfur were measured by photospectrometric measurement of thiocyanate complexes obtained by cyanolysis (Kelly et al., 1969) from the supernatant and pellet of a bioleaching sample, respectively. For total soluble sulfur, 500 μL sample was centrifuged for 5 min at 16,000 ×g and the supernatant mixed with 100 μL 0.5 M NaCN. After 10 min of incubation at room temperature, 500 μL of phosphate buffer (pH 7.2) and 100 μl 50 mM CuSO4 was added, followed by 30 min of incubation at room temperature. Then, complexes were formed by addition of 400 μL 1.5 M FeNO3 in aqueous 4 M HClO4 and distilled H2O to a volume of 2 mL. The reaction was then measured at 460 nm against a calibration curve of thiocyanate treated in the same way. For measurement of elemental sulfur, the mineral pellet of the same sample was dissolved in 2 mL of absolute acetone. 200 μL of this solution was then processed as described in the total soluble sulfur analysis, except that no CuSO4 was added, and the calibration curve was prepared using elemental sulfur dissolved in acetone. Total released, i.e., dissolved plus precipitated iron and copper concentrations were obtained by adding 1.8 mL of 5 M HCl to 200 μL of bioleaching slurry, followed by incubation at 65°C for 30 min. Then, the supernatant of this digest was diluted in 0.1 M HCl appropriate for measurement by atomic absorption spectroscopy (AAS) using a Perkin Elmer AAnalyst 400.
After 14 days of active bioleaching time (defined by a redox potential above 400 mV vs. Ag/AgCl), experiments were sampled for nucleic acid extraction. The flasks were left to settle for 5 min before removing 75 mL supernatant to be immediately mixed with an equal volume of sterile, ice-cold MAC medium. Then, the sample was centrifuged at 12,500 ×g for 20 min at 4°C. The resulting cell pellet was washed twice by resuspending in 10 and 2 mL of sterile, ice-cold MAC, respectively, and then frozen in liquid nitrogen. Cell pellets were subjected to biomolecular extractions according to a previously published method (Roume et al., 2013), skipping the metabolite extraction step. In short, cell pellets were lysed by cryo-milling and bead-beating followed by spin column based isolation of biomolecules with the Allprep kit (Qiagen, Belgium). Ribosomal RNA was depleted with the Ribo-Zero rRNA Removal Kit for bacteria (Illumina, United States). rRNA-depleted RNA for nine samples was stored at -80°C before shipping on dry ice to Science for Life Laboratory (Stockholm, Sweden) for sequencing.
Library preparation was performed with the Illumina TruSeq Stranded mRNA kit. Paired-end sequencing was performed on two HiSeq 2500 lanes resulting in on average 94 million reads per sample with length of 126 bp and GC% of 54% (Supplementary Table 1). Raw reads were filtered with Trimmomatic v0.32 (Bolger et al., 2014), TrueSeq3-PE adapter sequences were removed using the following parameters: seed mismatch:2; palindrome clip:30; simple clip:10; leading:20; trailing:20; sliding window: 1:3; minlen: 40; maxinfo: 40:0.5. Filtered reads were mapped onto a concatenation of the three reference genomes (A. caldus DSM 8584: GCF_000175575.2; S. thermosulfidooxidans DSM 9293: GCF_900176145.1; L. ferriphilum DSM 14647: GCF_900198525.1) with Bowtie-2 v2.3.2 (Langmead and Salzberg, 2012) with default parameters. Reads mapping to protein coding sequences were counted with the FeatureCounts program of the subread package v1.5.1 (Liao et al., 2014) with the –s 2 parameter accounting for strandedness. Read counts were then normalized and compared per organism with a custom R-script using the DESeq2 package v1.16.1 (Love et al., 2014) in R v3.4.4. Normalization was adapted from scripts provided in a previous publication (Klingenberg and Meinicke, 2017).
Raw sequencing reads are available from ENA SRA under study accession PRJEB27534. Scripts used in the analysis of the sequencing data can be accessed under the following link: https://git-r3lab.uni.lu/malte.herold/RNAseq_LF_ST_redox. Lists of genes relevant for analysis were generated by manual curation of the reference genome annotations and from previous publications (Janosch et al., 2015; Christel et al., 2017).
Bioleaching of chalcopyrite was tested with single, binary, and tertiary combinations of the three model species (A. caldus, L. ferriphilum, and S. thermosulfidooxidans), plus uninoculated controls to investigate the effect of species composition on redox potential and copper release (Figure 1 and Supplementary Figure 1). To aid comprehension, these combinations will be abbreviated using the initial letter of the included species (e.g., ‘ASL’ for the tertiary combination containing all species or ‘LS’ for the binary combination of L. ferriphilum and S. thermosulfidooxidans, etc.).
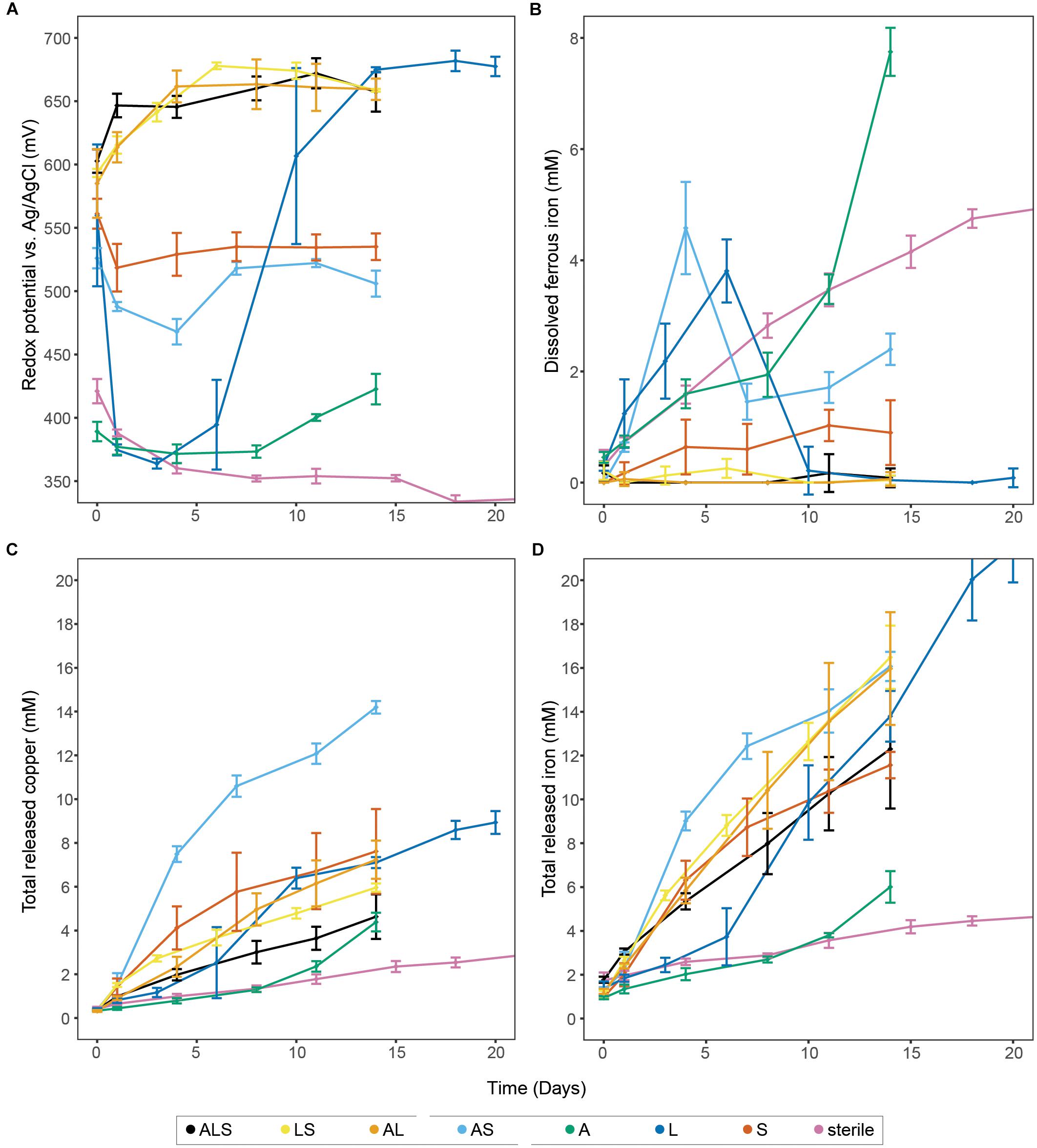
Figure 1. Bioleaching of chalcopyrite concentrate with single species, binary, and tertiary combinations of the three studied model species, plus uninoculated control. The panels show redox potential (A), dissolved Fe2+ (B), and total released copper and iron (C,D, respectively). Data points represent means ± SD (n = 4). Abbreviations in the legend denote: A, A. caldus, L; L. ferriphilum; and S, S. thermosulfidooxidans.
Physical and chemical analysis (Figure 1 and Supplementary Figure 1) of the uninoculated controls showed a redox potential of circa 310–330 mV (vs. Ag/AgCl) after stabilization, while the Fe2+ concentration steadily increased until plateauing at 5.7 ± 0.2 mM in later stages of the experiment (i.e., day 32, late stage data not shown). The abiotic leaching released a small amount of metal (4.2 ± 0.3 mM Fe and 2.4 ± 0.3 mM Cu after 15 days) from the chalcopyrite by proton attack and/or a small concentration of Fe3+ present on the mineral or in the medium. The same behavior was observed in the experiment inoculated exclusively with A. caldus, where the redox potential remained at circa 370 mV (i.e., well below the 400 mV perceived to mark the onset of bioleaching) until day 12 when likely environmental bacteria that survived the sterilization process on the mineral became active and commenced iron oxidation. Metal release was only marginally higher than from uninoculated controls, but showed a slight acceleration after the redox increase and reached 6.0 ± 0.7 mM Fe and 4.4 ± 0.4 mM Cu after 14 days. As sulfur compounds released from the mineral matrix are debated to be involved in formation of passivating layers (Khoshkhoo et al., 2014a,b), it is of importance to note that experiments containing, e.g., A. caldus exhibited examples of ISC degradation (Supplementary Figure 1), which is suggested to aid chalcopyrite dissolution (Spolaore et al., 2011). However, in the timeframe investigated during the leaching experiments, significant accumulation of soluble ISCs and elemental sulfur was also not observed in the ‘L’ combination excluding the sulfur oxidizers or in the sterile controls (Supplementary Figure 1). Beside the removal of passivating layers, ISC oxidation has the important role of maintaining low pH conditions in bioleaching heaps (Dopson and Lindstrom, 1999). Lacking sulfur oxidation potential in the microbial community can lead to a continuous rise in pH as observed in the sterile controls, or a potentially detrimental pH peak before the precipitation of ferric hydroxides and sulfates increases net acidity again (Ma et al., 2018; Supplemental Figure 1). This has implications for both the biology and engineering of bioleaching heaps, as many biomining species are sensitive to high pH, and accumulation of precipitates affect retention time of the leachate (Fagan et al., 2014). Additionally, pH induced precipitation of ferric iron alters the leachates ORP further affecting mineral dissolution.
Experiments including inoculation with iron-oxidizing bacteria allowed for transformation of Fe2+ to Fe3+ and therefore the redox potential rose up to, e.g., 682 ± 8 mV in the case of ‘L’. Accordingly, iron and copper release from all such experiments was significantly higher than from uninoculated controls and ‘A’; e.g., reaching the highest Fe concentration of 21.6 ± 1.7 mM in ‘L’ and highest copper concentration of 14.2 ± 0.3 mM in ‘AS’ (Figure 1).
The tertiary combination ‘ALS’ was outperformed by all binary combinations (Figure 1). This indicated that, in contrast to the currently accepted paradigm of inoculation of bioleaching applications with a broad mixture of biomining organisms, a well-chosen and defined mixture of microorganisms could benefit leaching efforts in the early stages of a bioleaching heap. Furthermore, the different combinations showed very distinct ORP profiles that, based on the present iron oxidizer(s), fell into one of two groups. All combinations containing L. ferriphilum had redox potentials between 650 and 680 mV compared to combinations in which it was excluded (i.e., ‘AS’ and ‘S’, showing ORPs below 550 mV). To confirm the lower redox potential in bioleaching cultures without L. ferriphilum, the ‘AS’ combination was repeated seven times with the redox potential reaching a maximum of 593 ± 15 mV (Figure 2A). Previous studies report that low redox potentials are favorable for chalcopyrite bioleaching (Third et al., 2002; Hiroyoshi et al., 2013). In accordance with that, in this study the redox potential of the leaching experiments correlated positively with the ratio of released iron/copper (Figure 2B). As dissolution of pure chalcopyrite is theoretically characterized by a 1:1 ratio of released iron to copper, this confirms the preferential oxidation of this copper mineral, or the transiently produced chalcocite, over associated copper-deficient minerals, such as pyrite, at low redox potentials. Our data independently confirms a study by Masaki et al. (2018), in which microbial redox control was also attempted, likewise by members of the Sulfobacilli, i.e., S. sibiricus and S. acidophilus. Using iron-oxidizing bacteria in bioleaching processes, which raise the ORP only minimally over the threshold for the onset of leaching therefore appears possible and could benefit the performance of chalcopyrite bioleaching processes.
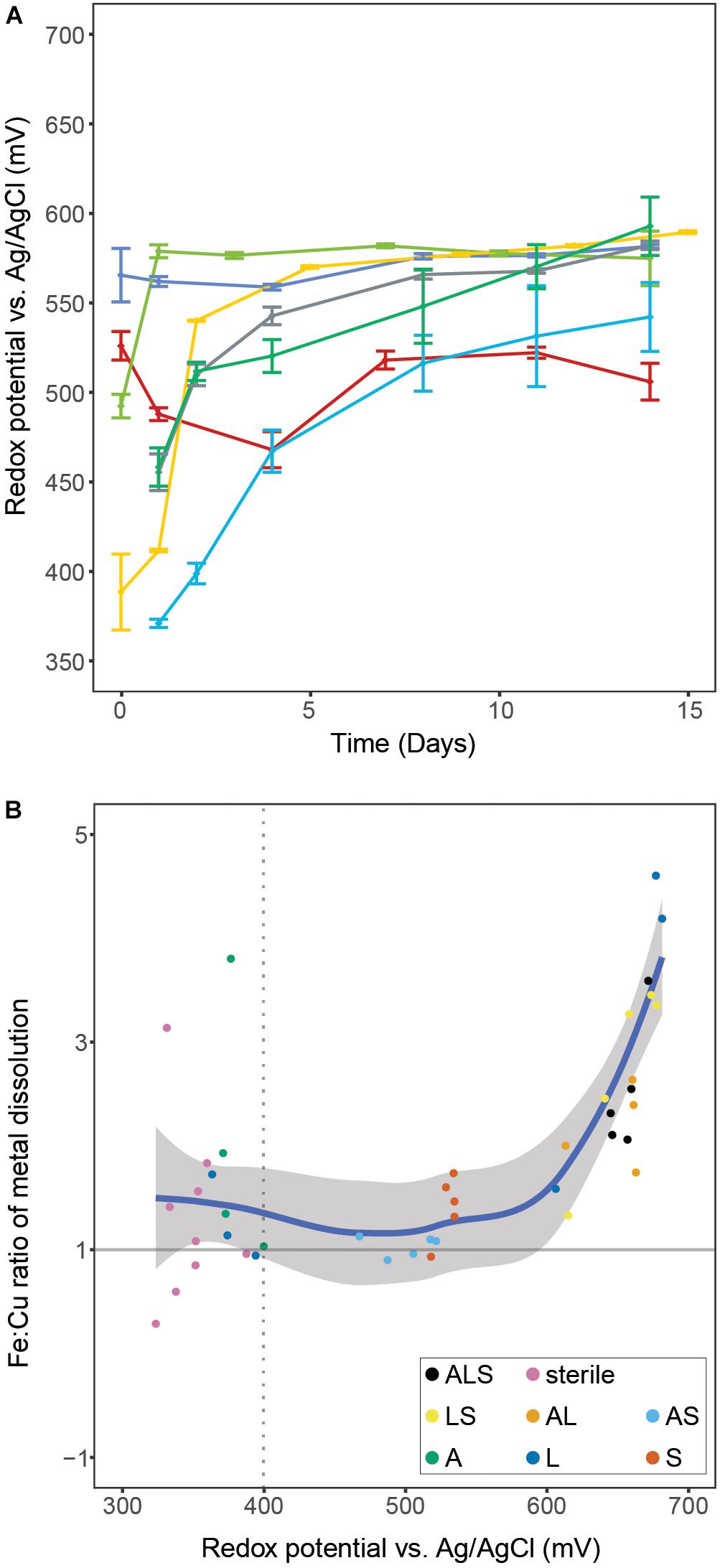
Figure 2. Panel A shows redox potentials remaining below 600 mV during seven independent experiments containing only S. thermosulfidooxidans for microbial iron oxidation. Data points represent means ± SD (n = 4). Panel B illustrates the correlation of the ratio of released iron:copper vs. redox potential during bioleaching of chalcopyrite concentrate with various combinations of the three model species. The ratio was calculated by dividing the amounts of the two metals that were released between two consecutive sampling points during the leaching experiments. The regression was calculated using the LOESS method with 95% confidence interval marked by the shaded area. The dotted line denotes the onset of microbial iron oxidation indicated by a redox potential above 400 mV. Abbreviations in the legend denote: A, A. caldus; L, L. ferriphilum; and S, S. thermosulfidooxidans.
Multiple reasons for the induction of diverse redox potentials by different species are conceivable. First and foremost, the effect could be explained by the effectivity and/or affinity of the respective species’ iron oxidation system. Species with a low affinity to ferrous iron, or inferior capability to oxidize it, should in theory maintain lower redox potentials. Additionally, high concentrations of Fe3+ ions are known to inhibit iron oxidation differently in different species (Rawlings et al., 1999), which could also contribute to this effect. Fluorescence microscopy examination of chalcopyrite grains in our bioleaching experiments revealed another difference that could contribute to the observed effect. L. ferriphilum showed significantly higher rates of colonization of mineral grains compared to S. thermosulfidooxidans (Bellenberg et al., 2018; Supplementary Figure 2). Attachment to metal sulfides is considered important for bioleaching, since in the so called ‘contact mechanism’, mineral-attached microbes concentrate Fe3+ in their EPS, effectively locally increasing the ORP at the microbe-mineral interface compared to the rest of the medium. Low levels of cell attachment on chalcopyrite mineral grains support the idea that a non-contact mechanism is observed for S. thermosulfidooxidans. Consequently, ferric ions diluted in the bulk medium maintain a more homogeneous redox environment. Unfortunately, Masaki et al. (2018) did not report on attachment rates and this hypothesis remains to be tested.
Additional factors affecting the iron oxidation efficiency could be the chosen experimental conditions. All used strains were adapted to grow at a temperature of 38°C prior to the bioleaching experiments, initially chosen as it is low enough to serve as an estimation of the temperature in a starting bioleaching heap (Leahy et al., 2007). At the same time, it is close to the optimum for L. ferriphilum that has a much narrower temperature range for growth while allowing reasonable growth for the other two species. Testing of S. thermosulfidooxidans iron oxidation efficiency near its growth optimum may become necessary, however, a raise in temperature has previously been reported to reduce leachate ORP in stirred reactors, and was connected to the exclusion of L. ferriphilum and retention of S. thermosulfidooxidans at a temperature of 48°C (Hedrich et al., 2018). Despite temperature control not being feasible in bioleaching heaps, this indicates that the low redox potential induced by S. thermosulfidooxidans in our experiments is not solely an effect of its growth at temperatures lower than its optimum. Overall, in the background of the other strain specific differences in experiments that run for prolonged periods of time, we consider the temperature influence as negligible, as the microbe’s growth capabilities are limited in the first place by the solubility of the mineral concentrates rather than temperature. This is strongly related to the acid-solubility of the mineral and to the iron oxidation capabilities of the respective single and mixed cultures. This is shown by all the iron being in the ferric form when leaching is efficient, i.e., the cells are limited by the availability of ferrous iron. Similar considerations were made for other parameters such as pH and carbon dioxide concentrations.
Owing to its heterogeneity and complexity, control of both chemical and biological parameters in a bioleaching heap is challenging (Petersen, 2016). Biologically, a major cause of this challenge is that due to implied costs, the mineral cannot be sterilized, and environmental bacteria will be present and thrive in the heap, competing with the inoculated ‘strategic’ microorganisms (Martinez et al., 2015). Procedures will have to be developed in order to fully exploit the potential of selected microorganisms, e.g., by continuously inoculating the heap with these strategic organisms through the irrigation system. In future studies, efforts should therefore be made to identify species with both low Fe2+ scavenging capabilities and increasingly high optimal growth temperatures.
In any case, manipulation of leachate, mineral, or other components of a bioleaching heap will naturally increase running costs. Further studies and ultimately large scale testing are needed to validate the viability of such approaches.
In an attempt to elucidate the biological background for the difference in redox potential, both iron-oxidizing model species’ transcriptomic response toward each other was investigated (i.e., ‘ASL’ vs. ‘AL’ for effect of S. thermosulfidooxidans on L. ferriphilum, and ‘ASL’ vs. ‘AS’ for the vice versa effect; Figure 3). The entirety of ecological interactions between the involved species is beyond the scope of this study and instead, this section concentrates on gene products related to energy metabolism, iron-, and sulfur oxidation as previously reported (Mangold et al., 2011; Guo et al., 2014; Christel et al., 2017; Supplementary Tables 1, 2).
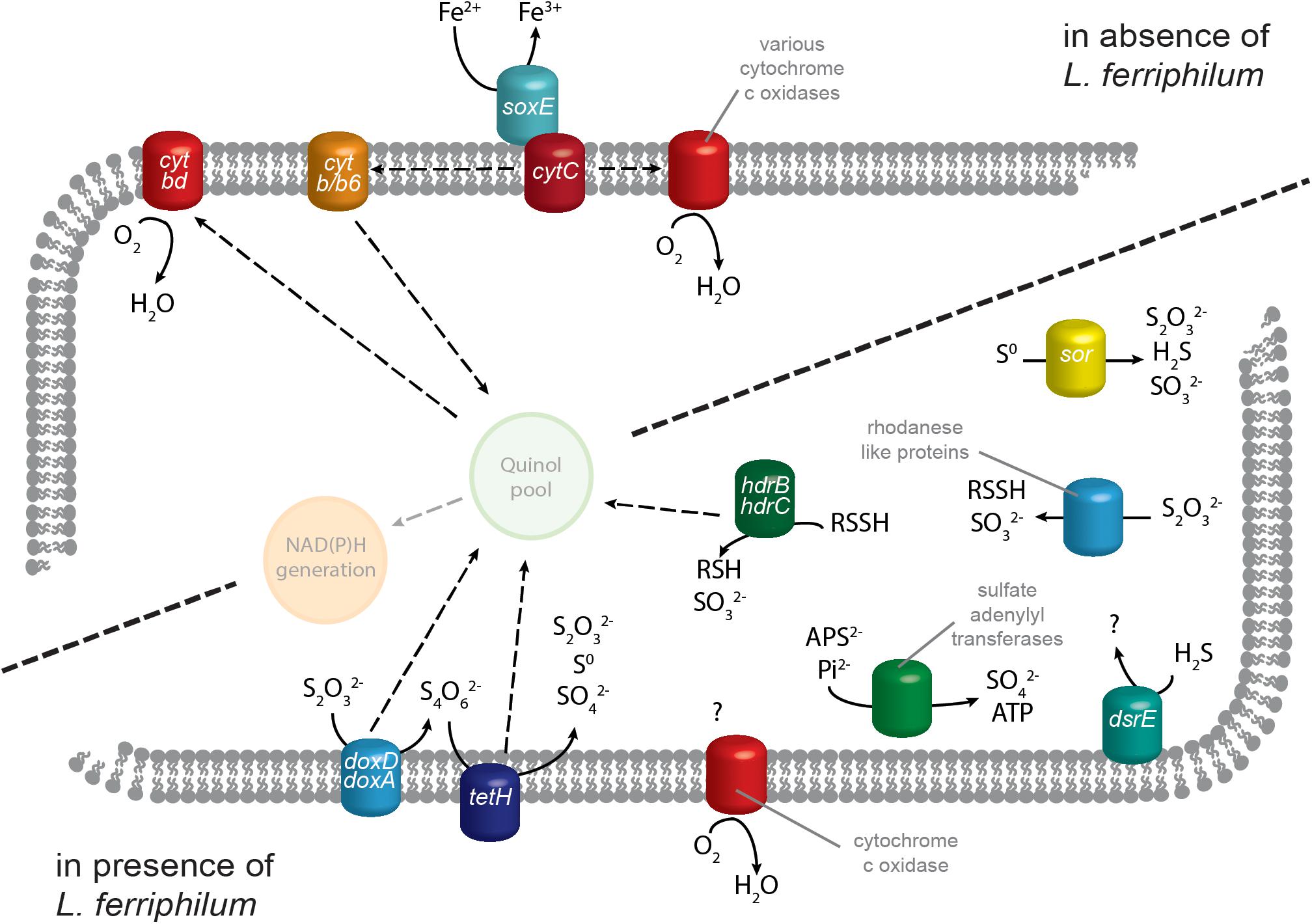
Figure 3. Proposed model of S. thermosulfidooxidans transcript regulation of genes related to energy generation. Iron oxidation systems and electron transport by cytochromes has a greater number of RNA transcripts in the absence of the strong iron oxidizer L. ferriphilum. In its presence, S. thermosulfidooxidans instead has higher transcript numbers for genes contributing to ISC oxidation plus one cytochrome c oxidase complex. Quinone pool and NAD(P)H generation are depicted translucently for comprehension, but corresponding genes were not analyzed in this study. Solid arrows represent metabolic reactions while dashed arrows indicate the relocation of electrons.
In bioleaching co-culture, L. ferriphilum remained remarkably unaffected by the presence of S. thermosulfidooxidans. Over its entire genome (2486 genes), only 36 genes showed significant differential expression in response to S. thermosulfidooxidans (‘AL’ vs. ‘ALS’; p ≤ 0.05; data not shown). Among the 26 genes attributed to iron oxidation and electron transport, merely three cbb3-type cytochrome c oxidase subunits (LFTS_01396, _02094, and _02276) exhibited significantly increased transcript numbers in the presence of S. thermosulfidooxidans, all of which have log2-fold changes below 1.5 (Supplementary Table 2). No genes involved in iron oxidation or electron transport had significantly higher numbers of RNA transcripts in the absence of S. thermosulfidooxidans.
Acidithiobacillus caldus exhibited 508 significantly differentially expressed genes (of 2885) depending on the iron oxidizer present (‘AL’ vs. ‘AS’; p ≤ 0.05; data not shown). Of these, 246 showed increased transcript counts during co-culture with S. thermosulfidooxidans. Significant changes related to energy acquisition included a strong increase in relative RNA transcript counts for a large portion of the sulfur oxidizing soxABXYZ and likely associated cytochrome c biogenesis protein genes (Supplementary Table 3). This was surprising as, although S. thermosulfidooxidans is capable of sulfur oxidation, due to its preference for iron oxidation in the absence of stronger iron oxidizers (see following sections), this species should not be in direct competition with A. caldus in this case. This may potentially be explained if A. caldus maintains a high level of sulfur oxidation proteins in the presence of other potential sulfur oxidizing microbes so as not to be outcompeted. In contrast, the presence of L. ferriphilum appeared to enhance transcription of cytochrome bd ubiquinol oxidase subunits I and II (Supplementary Table 3). In addition to its respiratory function, cytochrome bd complex has been reported to increase resistance against oxidative stress in Escherichia coli (Lindqvist et al., 2000) and could fulfill a similar function in the high ORP conditions induced by L. ferriphilum.
Sulfobacillus thermosulfidooxidans gene transcript numbers exhibited great variation depending on presence of L. ferriphilum. Of its 3805 identified genes, 828 showed significant differential expression (‘AS’ vs. ‘ALS’; p ≤ 0.05; data not shown). Among the 83 selected genes involved in iron oxidation, electron transport, and sulfur oxidation, 55 had significantly greater or lower RNA transcripts (Table 1 and Supplementary Table 4). Large variation was observed in genes related to iron oxidation. In contrast to, e.g., some members of the genus Acidithiobacillus, Sulfobacilli genomes lack the common iron oxidation protein rusticyanin (Guo et al., 2014). Instead, Sulfobacilli are suggested to utilize sulfocyanin, which is also found in the archaeal iron oxidizers of the genus Ferroplasma (Dopson et al., 2005). In the presence of L. ferriphilum, S. thermosulfidooxidans strongly decreased transcript numbers attributed to two of the five soxE genes coding for this protein (Sulth_0453 and _2749). Additionally, the vast majority of identified cytochromes of all types exhibited decreased transcript counts, along with corresponding biogenesis proteins and quinol oxidases (Table 1 and Supplementary Table 4). The strong downregulation of electron chain components that were likely linked to iron oxidation in S. thermosulfidooxidans could be explained by the chemical data reported in the previous section. In cultures containing both iron oxidizers, the concentration of available ferrous iron was below the detection limit and likely too low for utilization by S. thermosulfidooxidans. This may be attributed to L. ferriphilum being able to scavenge Fe2+ at concentrations far below S. thermosulfidooxidans’ capabilities and at large Fe3+ concentrations that exceed its inhibition limits (Rawlings et al., 1999).
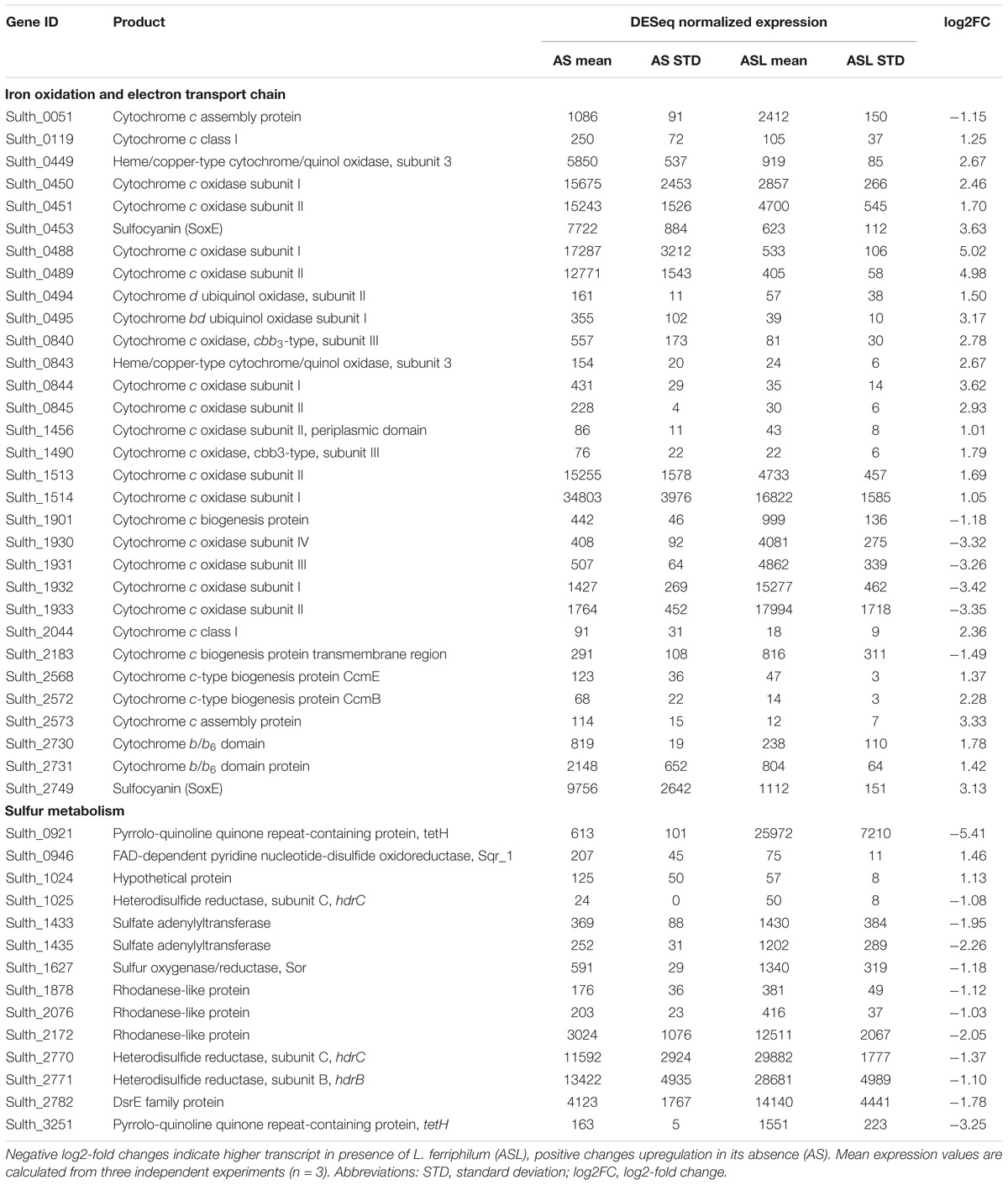
Table 1. Excerpt of Supplementary Table 1 showing significant (|log2FC| ≥ 1.0, p ≤ 0.05) differential expression of S. thermosulfidooxidans genes related to iron and sulfur oxidation as well as electron transport.
Contrary to this overall trend, one gene cluster of S. thermosulfidooxidans encoding cytochrome c oxidase subunits I-IV showed strongly increased transcript counts in the presence of L. ferriphilum (Table 1, Sulth_1930-1933). In addition, two cytochrome c biogenesis proteins (Sulth_1901 and _2183) and one cytochrome c assembly protein (Sulth_0051) exhibited similarly increased transcript numbers. A direct role of cytochromes in iron oxidation has been suggested in an acid mine drainage biofilm and in L. ferrooxidans (Jeans et al., 2008; Blake and Griff, 2012). Therefore, the strong opposite regulation of cytochrome oxidases in S. thermosulfidooxidans raises the question of their potential functional and/or structural differences. It could be possible that the oxidase exhibiting increased transcript counts in the presence of L. ferriphilum indirectly facilitates a higher affinity for Fe2+, or has a lower sensitivity toward oxidative stress induced by accumulating Fe3+. Together with the upregulation of biogenesis and assembly proteins, this could enable S. thermosulfidooxidans to gain some energy from Fe2+ in the presence of a stronger iron oxidizer. Alternatively, the cytochrome c oxidase complex upregulated in the presence of L. ferriphilum could be part of the strong upregulation of genes described in the following section, i.e., reducing oxygen in the final step of sulfur oxidation systems. Nevertheless, as only cytochromes and cytochrome oxidases that are upregulated in absence of L. ferriphilum correlated with higher copper extraction, they may be of greater interest in the context of this study and should be considered in more detail in the future.
Genes coding for known sulfur oxidation proteins exhibited directionally opposite changes in transcript numbers compared to iron oxidation systems. Conceivably, this was to ensure sufficient supply of energy in a Fe2+ deficient environment and the vast majority of S. thermosulfidooxidans genes related to sulfur metabolism (Guo et al., 2014) had significantly higher RNA transcripts in the presence of L. ferriphilum (Table 1 and Supplementary Table 4). This reiterates that in presence of a strong iron oxidizer, S. thermosulfidooxidans’ role lies in the removal of ISCs as discussed by Spolaore et al. (2011). The highest of these log2-fold changes were recorded for two copies of tetrathionate hydrolase gene tetH (Sulth_0921 and _3251) while a third copy (Sulth_1188) exhibited moderately increased transcript counts in the absence of L. ferriphilum. TetH is responsible for the hydrolysis of tetrathionate, an important intermediate in sulfide mineral dissolution. Additionally, thiosulfate can be oxidized by thiosulfate quinone oxidoreductase, encoded by a doxDA homolog. The two encoded copies of this gene exhibited increased transcript counts when in co-culture with L. ferriphilum, although the log2-fold changes were low (Sulth_1989 and 1691). A similar function is suggested for rhodanese-like proteins, which in humans are sulfur transferases involved in the detoxification of cyanide by transformation to thiocyanate in the liver (Nakajima, 2015). Proteins sharing their active domain are also suggested to play a role in microbial sulfur oxidation, in particular of thiosulfate (Valdes et al., 2008). In S. thermosulfidooxidans, seven genes coding for such proteins were present (Table 1 and Supplementary Table 4) and four of them exhibited significantly increased transcript numbers in the presence of L. ferriphilum (Sulth_1878, _2076, _2172, and _3294). The product of their enzymatic reaction can be further oxidized by heterodisulfide reductase, which is encoded in S. thermosulfidooxidans by three sets of genes for its respective subunits HdrBC (Supplementary Table 4). Four Hdr subunit gene loci exhibited significant slightly increased transcript counts in presence of L. ferriphilum (Sulth_1025, _1026, _2770, and _2771). Elemental sulfur oxidation is also relevant in the context of sulfide mineral dissolution, and is conducted by the product of two copies of sulfur oxygenase/reductase gene sor (Janosch et al., 2015). While one copy was only minimally expressed in both conditions (Sulth_1798; indicating a possibly defunct gene), the second exhibited greater transcript numbers (Sulth_1627) and appeared to be enhanced by the presence of L. ferriphilum.
Additional contributions to sulfur oxidation systems include sulfate adenylyltransferase, which is suggested to be involved in sulfite oxidation in A. ferrooxidansT and S. thermosulfidooxidans strain ST (Guo et al., 2014) and is likely to fulfill the same role in S. thermosulfidooxidansT. Similarly, DsrE-family protein (Sulth_2782) has been reported to be associated to oxidative sulfite metabolism (Dahl et al., 2005) and was also found to exhibit increased transcript numbers in presence of L. ferriphilum (Table 1).
In the absence of L. ferriphilum, only a few genes related to sulfur oxidation were significantly enhanced. Of note in this regard is one of three present copies of sulfide quinone reductase gene sqr1 (Sulth_0946), which is responsible for the oxidation of sulfide to elemental sulfur a hypothetical protein within the hdr gene cluster (Sulth_1024), as well as the upper mentioned copy of tetH (Sulth_1188).
During bioleaching of chalcopyrite concentrate, S. thermosulfi-dooxidans but not L. ferriphilum maintained a low redox potential that is favorable for the extraction of copper. We hypothesize that this was due to differences in affinity and/or effectivity of the species’ respective iron oxidation systems, as well as the attachment rate of the microorganisms to the mineral grains. This finding could potentially contribute to overcoming passivation and improving dissolution rates in large-scale chalcopyrite bioleaching. Expression of iron and sulfur oxidation systems in S. thermosulfidooxidans were investigated during bioleaching experiments in presence and absence of L. ferriphilum. Presence of the strong iron oxidizer induced greatly decreased transcript counts attributed to iron oxidation and increased counts for sulfur oxidation. Analysis of this data revealed genes products potentially responsible for the difference in ORP, which should be studied in this regard in the future. Additionally, this study underlines the importance of developing methods to control microbial populations in a bioleaching heap in order to exploit desired properties of selected microorganisms.
SC conducted the laboratory experiments and drafted the manuscript. MH performed bioinformatic analysis of transcript data. SB provided microscopic imaging. SC, SB, AB-D, MEH, IP, WS, PW, AP, MV, and MD were involved in data analysis and biological interpretation of the results. All authors contributed to its preparation.
This project was supported by Vetenskapsrådet (contract 2014-6545), the Luxembourg National Research Fund (FNR, INTER/SYSAPP/14/05), Bundesministerium für Bildung und Forschung (BMBF, 031A600A and B), and The Swiss Initiative in Systems Biology (SystemsX.ch, SysMetEx) under the frame of ERASysAPP. MV acknowledges funding from FONDECYT grant 1161007.
The authors SC, MH, SB, AB-D, MEH, IP, PW, AP, and MD and/or the institutions in which they are affiliated with, have a patent pending based on the results of the study.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The chalcopyrite concentrate used in this study was kindly provided by Boliden Mineral AB. The authors acknowledge support from Science for Life Laboratory and the National Genomics Infrastructure for providing assistance in massive parallel sequencing and computational infrastructure. Uppsala Multidisciplinary Center for Advanced Computational Science is greatly acknowledged for assistance with massively parallel sequencing and access to the UPPMAX computational infrastructure. Parts of the sequencing analysis presented in this paper were carried out using the HPC facilities of the University of Luxembourg.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.03059/full#supplementary-material
Baldi, F., Bralia, A., Riccobono, F., and Sabatini, G. (1991). Bioleaching of cobalt and zinc from pyrite ore in relation to calcitic gangue content. World J. Microbiol. Biotechnol. 7, 298–308. doi: 10.1007/BF00329395
Bellenberg, S., Buetti-Dinh, A., Galli, V., Ilie, O., Herold, M., Christel, S., et al. (2018). Automated microscopical analysis of metal sulfide colonization by acidophilic microorganisms. Appl. Environ. Microbiol. 84, e01835-18. doi: 10.1128/AEM.01835-18
Blake, R. C. II, and Griff, M. N. (2012). In situ spectroscopy on intact Leptospirillum ferrooxidans reveals that reduced cytochrome 579 is an obligatory intermediate in the aerobic iron respiratory chain. Front. Microbiol. 3:136. doi: 10.3389/fmicb.2012.00136
Bolger, A. M., Lohse, M., and Usadel, B. (2014). Trimmomatic: a flexible trimmer for illumina sequence data. Bioinformatics 30, 2114–2120. doi: 10.1093/bioinformatics/btu170
Brierley, C. L., and Brierley, J. A. (2013). Progress in bioleaching: Part B: applications of microbial processes by the minerals industries. Appl. Microbiol. Biotechnol. 97, 7543–7552. doi: 10.1007/s00253-013-5095-3
Bryner, L. C., and Jameson, A. K. (1958). Microorganisms in leaching sulfide minerals. Appl. Microbiol. 6, 281–287.
Christel, S., Herold, M., Bellenberg, S., El Hajjami, M., Buetti-Dinh, A., Pivkin, I. V., et al. (2017). Multi-omics reveal the lifestyle of the acidophilic, mineral-oxidizing model species Leptospirillum ferriphilum. Appl. Environ. Microbiol. doi: 10.1128/AEM.02091-17 [Epub ahead of print].
Coram, N. J., and Rawlings, D. E. (2002). Molecular relationship between two groups of the genus Leptospirillum and the finding that Leptospirillum ferriphilum sp nov dominates South African commercial biooxidation tanks that operate at 40 degrees C. Appl. Environ. Microbiol. 68, 838–845. doi: 10.1128/Aem.68.2.838-845.2002
Cordoba, E. M., Munoz, J. A., Blazquez, M. L., Gonzalez, F., and Ballester, A. (2009). Passivation of chalcopyrite during its chemical leaching with ferric ion at 68 degrees C. Minerals Eng. 22, 229–235. doi: 10.1016/j.mineng.2008.07.004
Crundwell, F. K. (2015). The semiconductor mechanism of dissolution and the pseudo-passivation of chalcopyrite. Can. Metallurg. Q. 54, 279–288. doi: 10.1179/1879139515y.0000000007
Dahl, C., Engels, S., Pott-Sperling, A. S., Schulte, A., Sander, J., Lubbe, Y., et al. (2005). Novel genes of the dsr gene cluster and evidence for close interaction of dsr proteins during sulfur oxidation in the phototrophic sulfur bacterium Allochromatium vinosum. J. Bacteriol. 187, 1392–1404. doi: 10.1128/JB.187.4.1392-1404.2005
Dopson, M., Baker-Austin, C., and Bond, P. L. (2005). Analysis of differential protein expression during growth states of ferroplasma strains and insights into electron transport for iron oxidation. Microbiology 151(Pt 12), 4127–4137. doi: 10.1099/mic.0.28362-0
Dopson, M., and Lindstrom, E. B. (1999). Potential role of Acidithiobacillus caldus in arsenopyrite bioleaching. Appl. Environ. Microbiol. 65, 36–40.
Dopson, M., Lovgren, L., and Bostrom, D. (2009). Silicate mineral Dissolution in the presence of acidophilic microorganisms: implications for heap bioleaching. Hydrometallurgy 96, 288–293. doi: 10.1016/j.hydromet.2008.11.004
Fagan, M. A., Ngoma, I. E., Chiume, R. A., Minnaar, S., Sederman, A. J., Johns, M. L., et al. (2014). Mri and gravimetric studies of hydrology in drip irrigated heaps and its effect on the propagation of bioleaching micro-organisms. Hydrometallurgy 150, 210–221. doi: 10.1016/j.hydromet.2014.04.022
Gericke, M., Govender, Y., and Pinches, A. (2010). Tank bioleaching of low-grade chalcopyrite concentrates using redox control. Hydrometallurgy 104, 414–419. doi: 10.1016/j.hydromet.2010.02.024
Guo, X., Yin, H., Liang, Y., Hu, Q., Zhou, X., Xiao, Y., et al. (2014). Comparative genome analysis reveals metabolic versatility and environmental adaptations of Sulfobacillus thermosulfidooxidans Strain St. PLoS One 9:e99417. doi: 10.1371/journal.pone.0099417
Hallberg, K. B., and Lindstrom, E. B. (1994). Characterization of Acidithiobacillus caldus Sp. nov., a moderately thermophilic acidophile. Microbiology 140(Pt 12), 3451–3456. doi: 10.1099/13500872-140-12-3451
Hedrich, S., Joulian, C., Graupner, T., Schippers, A., and Guézennec, A.-G. (2018). Enhanced chalcopyrite dissolution in stirred tank reactors by temperature increase during bioleaching. Hydrometallurgy 179, 125–131. doi: 10.1016/j.hydromet.2018.05.018
Hedrich, S., Schlomann, M., and Johnson, D. B. (2011). The iron-oxidizing Proteobacteria. Microbiology 157(Pt 6), 1551–1564. doi: 10.1099/mic.0.045344-0
Hiroyoshi, N., Tsunekawa, M., Okamoto, H., Nakayama, R., and Kuroiwa, S. (2013). Improved chalcopyrite leaching through optimization of redox potential. Can. Metallurg. Q. 47, 253–258. doi: 10.1179/cmq.2008.47.3.253
Janosch, C., Remonsellez, F., Sand, W., and Vera, M. (2015). Sulfur oxygenase reductase (Sor) in the moderately thermoacidophilic leaching bacteria: studies in Sulfobacillus thermosulfidooxidans and Acidithiobacillus caldus. Microorganisms 3, 707–724. doi: 10.3390/microorganisms3040707
Jeans, C., Singer, S. W., Chan, C. S., Verberkmoes, N. C., Shah, M., Hettich, R. L., et al. (2008). Cytochrome 572 is a conspicuous membrane protein with iron oxidation activity purified directly from a natural acidophilic microbial community. ISME J. 2, 542–550. doi: 10.1038/ismej.2008.17
Jerez, C. A. (2017). Biomining of metals: how to access and exploit natural resource sustainably. Microb. Biotechnol. 10, 1191–1193. doi: 10.1111/1751-7915.12792
Johnson, D. B. (2014). Biomining-biotechnologies for extracting and recovering metals from ores and waste materials. Curr. Opin. Biotechnol. 30, 24–31. doi: 10.1016/j.copbio.2014.04.008
Karavaiko, G. I., Bogdanova, T. I., Tourova, T. P., Kondrat’eva, T. F., Tsaplina, I. A., Egorova, M. A., et al. (2005). Reclassification of ‘Sulfobacillus thermosulfidooxidans subsp. Thermotolerans’ strain K1 as Alicyclobacillus tolerans sp. nov. and Sulfobacillus disulfidooxidans dufresne Et Al. 1996 as Alicyclobacillus disulfidooxidans Comb. Nov., and emended description of the genus Alicyclobacillus. Int. J. Syst. Evol. Microbiol. 55(Pt 2), 941–947. doi: 10.1099/ijs.0.63300-0
Kelly, D. P., Chambers, L. A., and Trudinge, P. A. (1969). Cyanolysis and spectrophotometric estimation of trithionate in mixture with thiosulfate and tetrathionate. Anal. Chem. 41, 898–901. doi: 10.1021/ac60276a029
Khoshkhoo, M., Dopson, M., Engstrom, F., and Sandstrom, A. (2017). New insights into the influence of redox potential on chalcopyrite leaching behaviour. Minerals Eng. 100, 9–16. doi: 10.1016/j.mineng.2016.10.003
Khoshkhoo, M., Dopson, M., Shchukarev, A., and Sandstrom, A. (2014a). Chalcopyrite leaching and bioleaching: an x-ray photoelectron spectroscopic (Xps) investigation on the nature of hindered dissolution. Hydrometallurgy 149, 220–227. doi: 10.1016/j.hydromet.2014.08.012
Khoshkhoo, M., Dopson, M., Shchukarev, A., and Sandstrom, A. (2014b). Electrochemical simulation of redox potential development in bioleaching of a pyritic chalcopyrite concentrate. Hydrometallurgy 144, 7–14. doi: 10.1016/j.hydromet.2013.12.003
Klingenberg, H., and Meinicke, P. (2017). How to normalize metatranscriptomic count data for differential expression analysis. PeerJ 5:e3859. doi: 10.7717/peerj.3859
Langmead, B., and Salzberg, S. L. (2012). Fast gapped-read alignment with bowtie 2. Nat. Methods 9, 357–359. doi: 10.1038/nmeth.1923
Leahy, M. J., Davidson, M. R., and Schwarz, M. P. (2007). A model for heap bioleaching of chalcocite with heat balance: mesophiles and moderate thermophiles. Hydrometallurgy 85, 24–41. doi: 10.1016/j.hydromet.2006.07.004
Li, Y., Kawashima, N., Li, J., Chandra, A. P., and Gerson, A. R. (2013). A Review of the structure, and fundamental mechanisms and kinetics of the leaching of chalcopyrite. Adv. Colloid Interface Sci. 19, 1–32. doi: 10.1016/j.cis.2013.03.004
Liao, Y., Smyth, G. K., and Shi, W. (2014). Featurecounts: an efficient general purpose program for assigning sequence reads to genomic features. Bioinformatics 30, 923–930. doi: 10.1093/bioinformatics/btt656
Lindqvist, A., Membrillo-Hernández, J., Poole, R. K., and Cook, G. M. (2000). Roles of respiratory oxidases in protecting Escherichia Coli K12 from oxidative stress. Antonie Van Leeuwenhoek 78, 23–31. doi: 10.1023/a:1002779201379
Love, M. I., Huber, W., and Anders, S. (2014). Moderated estimation of fold change and dispersion for RNA-Seq data with Deseq2. Genome Biol. 15:550. doi: 10.1186/s13059-014-0550-8
Ma, Y., Wang, H., Song, Y., Wu, Y., and Guo, Z. (2018). The synthesis of secondary iron minerals induced by quartz sand during the bioleaching process improves the dewaterability of municipal sewage sludge. Minerals 8:419. doi: 10.3390/min8100419
Mackintosh, M. E. (1978). Nitrogen-fixation by Thiobacillus ferrooxidans. J. Gen. Microbiol. 105, 215–218. doi: 10.1099/00221287-105-2-215
Mangold, S., Valdes, J., Holmes, D. S., and Dopson, M. (2011). Sulfur metabolism in the extreme acidophile Acidithiobacillus Caldus. Front. Microbiol. 2:17. doi: 10.3389/fmicb.2011.00017
Martinez, P., Vera, M., and Bobadilla-Fazzini, R. A. (2015). Omics on bioleaching: current and future impacts. Appl. Microbiol. Biotechnol. 99, 8337–8350. doi: 10.1007/s00253-015-6903-8
Masaki, Y., Hirajima, T., Sasaki, K., Miki, H., and Okibe, N. (2018). Microbiological redox potential control to improve the efficiency of chalcopyrite bioleaching. Geomicrobiol. J. 35, 648–656. doi: 10.1080/01490451.2018.1443170
Nakajima, T. (2015). Roles of sulfur metabolism and rhodanese in detoxification and anti-oxidative stress functions in the liver: responses to radiation exposure. Med. Sci. Monit. 21, 1721–1725. doi: 10.12659/MSM.893234
Panda, S., Akcil, A., Pradhan, N., and Deveci, H. (2015). Current scenario of chalcopyrite bioleaching: a review on the recent advances to its heap-leach technology. Bioresour. Technol. 196, 694–706. doi: 10.1016/j.biortech.2015.08.064
Penev, K., and Karamanev, D. (2010). Batch kinetics of ferrous iron oxidation by Leptospirillum ferriphilum at moderate to high total iron concentration. Biochem. Eng. J. 50, 54–62. doi: 10.1016/j.bej.2010.03.004
Petersen, J. (2016). Heap leaching as a key technology for recovery of values from low-grade ores – a brief overview. Hydrometallurgy 165, 206–212. doi: 10.1016/j.hydromet.2015.09.001
Quatrini, R., and Johnson, D. B. (eds) (2016). Acidophiles: Life in Extremely Acidic Environments. Poole: Caister Academic Press.
Rawlings, D. E., and Johnson, D. B. (2007). The microbiology of biomining: development and optimization of mineral-oxidizing microbial consortia. Microbiology 153(Pt 2), 315–324. doi: 10.1099/mic.0.2006/001206-0
Rawlings, D. E., Tributsch, H., and Hansford, G. S. (1999). Reasons why ‘leptospirillum’-like species rather than Thiobacillus ferrooxidans are the dominant iron-oxidizing bacteria in many commercial processes for the biooxidation of pyrite and related ores. Microbiology 145(Pt 1), 5–13. doi: 10.1099/13500872-145-1-5
Rohwerder, T., Gehrke, T., Kinzler, K., and Sand, W. (2003). Bioleaching review part a: progress in bioleaching: fundamentals and mechanisms of bacterial metal sulfide oxidation. Appl. Microbiol. Biotechnol. 63, 239–248. doi: 10.1007/s00253-003-1448-7
Roume, H., Muller, E. E., Cordes, T., Renaut, J., Hiller, K., and Wilmes, P. (2013). A biomolecular isolation framework for eco-systems biology. ISME J. 7, 110–121. doi: 10.1038/ismej.2012.72
Spolaore, P., Joulian, C., Gouin, J., Ibáñez, A., Augé, T., Morin, D., et al. (2009). Bioleaching of an organic-rich polymetallic concentrate using stirred-tank technology. Hydrometallurgy 99, 137–143. doi: 10.1016/j.hydromet.2009.07.011
Spolaore, P., Joulian, C., Gouin, J., Morin, D., and d’Hugues, P. (2011). Relationship between bioleaching performance, bacterial community structure and mineralogy in the bioleaching of a copper concentrate in stirred-tank reactors. Appl. Microbiol. Biotechnol. 89, 441–448. doi: 10.1007/s00253-010-2888-5
Temple, K. L., and Colmer, A. R. (1951). The autotrophic oxidation of iron by a new bacterium, Thiobacillus ferrooxidans. J. Bacteriol. 62, 605–611.
Third, K. A., Cord-Ruwisch, R., and Watling, H. R. (2002). Control of the redox potential by oxygen limitation improves bacterial leaching of chalcopyrite. Biotechnol. Bioeng. 78, 433–441. doi: 10.1002/bit.10184
Tsaplina, I. A., Krasil’nikova, E. N., Zakharchuk, L. M., Egorova, M. A., Bogdanova, T. I., and Karavaiko, G. I. (2000). Carbon metabolism in Sulfobacillus thermosulfidooxidans Subsp. Asporogenes, Strain 41. Mikrobiologiia 69,334–340.
Valdes, J., Pedroso, I., Quatrini, R., Dodson, R. J., Tettelin, H., Blake, R., et al. (2008). Acidithiobacillus ferrooxidans metabolism: from genome sequence to industrial applications. BMC Genomics 9:597. doi: 10.1186/1471-2164-9-597
Vera, M., Schippers, A., and Sand, W. (2013). Progress in bioleaching: fundamentals and mechanisms of bacterial metal sulfide oxidation–Part A. Appl. Microbiol. Biotechnol. 97, 7529–7541. doi: 10.1007/s00253-013-4954-2
Walden, G. H., Hammett, L. P., and Chapman, R. P. (1933). Phenanthroline-ferrous ion: a reversible oxidation—reduction indicator of high potential and its use in oxidimetric titrations. J. Am. Chem. Soc. 55, 2649–2654. doi: 10.1021/ja01334a005
Wang, J., Gan, X. W., Zhao, H. B., Hu, M. H., Li, K. Y., Qin, W. Q., et al. (2016). Dissolution and passivation mechanisms of chalcopyrite during bioleaching: Dft calculation, Xps and electrochemistry analysis. Minerals Eng. 98, 264–278. doi: 10.1016/j.mineng.2016.09.008
Watling, H. R. (2006). The bioleaching of sulphide minerals with emphasis on copper sulphides–a review. Hydrometallurgy 84, 81–108. doi: 10.1016/j.hydromet.2006.05.001
Keywords: redox control, microbial, bioleaching, chalcopyrite, iron oxidation, Sulfobacillus, Leptospirillum
Citation: Christel S, Herold M, Bellenberg S, Buetti-Dinh A, El Hajjami M, Pivkin IV, Sand W, Wilmes P, Poetsch A, Vera M and Dopson M (2018) Weak Iron Oxidation by Sulfobacillus thermosulfidooxidans Maintains a Favorable Redox Potential for Chalcopyrite Bioleaching. Front. Microbiol. 9:3059. doi: 10.3389/fmicb.2018.03059
Received: 30 August 2018; Accepted: 27 November 2018;
Published: 12 December 2018.
Edited by:
Masahiro Ito, Toyo University, JapanReviewed by:
Satoshi Wakai, Kobe University, JapanCopyright © 2018 Christel, Herold, Bellenberg, Buetti-Dinh, El Hajjami, Pivkin, Sand, Wilmes, Poetsch, Vera and Dopson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Stephan Christel, c3RlcGhhbi5jaHJpc3RlbEBsbnUuc2U=; bWFpbEBzdGVwaGFuY2hyaXN0ZWwuZGU=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.