 Philippe Colson1
Philippe Colson1 Anthony Levasseur1
Anthony Levasseur1 Bernard La Scola1
Bernard La Scola1 Vikas Sharma1,2
Vikas Sharma1,2 Arshan Nasir3,4
Arshan Nasir3,4 Pierre Pontarotti1,2
Pierre Pontarotti1,2 Gustavo Caetano-Anollés3
Gustavo Caetano-Anollés3 Didier Raoult1*
Didier Raoult1*- 1Aix-Marseille Université, Institut de Recherche pour le Développement (IRD), Assistance Publique – Hôpitaux de Marseille (AP-HM); Microbes, Evolution, Phylogeny and Infection (MEΦI); Institut Hospitalo-Universitaire (IHU) – Méditerranée Infection, Marseille, France
- 2Centre National de la Recherche Scientifique, Marseille, France
- 3Evolutionary Bioinformatics Laboratory, Department of Crop Sciences, University of Illinois Urbana-Champaign, Urbana, IL, United States
- 4Department of Biosciences, COMSATS University Islamabad, Islamabad, Pakistan
Giant viruses of amoebae were discovered in 2003. Since then, their diversity has greatly expanded. They were suggested to form a fourth branch of life, collectively named ‘TRUC’ (for “Things Resisting Uncompleted Classifications”) alongside Bacteria, Archaea, and Eukarya. Their origin and ancestrality remain controversial. Here, we specify the evolution and definition of giant viruses. Phylogenetic and phenetic analyses of informational gene repertoires of giant viruses and selected bacteria, archaea and eukaryota were performed, including structural phylogenomics based on protein structural domains grouped into 289 universal fold superfamilies (FSFs). Hierarchical clustering analysis was performed based on a binary presence/absence matrix constructed using 727 informational COGs from cellular organisms. The presence/absence of ‘universal’ FSF domains was used to generate an unrooted maximum parsimony phylogenomic tree. Comparison of the gene content of a giant virus with those of a bacterium, an archaeon, and a eukaryote with small genomes was also performed. Overall, both cladistic analyses based on gene sequences of very central and ancient proteins and on highly conserved protein fold structures as well as phenetic analyses were congruent regarding the delineation of a fourth branch of microbes comprised by giant viruses. Giant viruses appeared as a basal group in the tree of all proteomes. A pangenome and core genome determined for Rickettsia bellii (bacteria), Methanomassiliicoccus luminyensis (archaeon), Encephalitozoon intestinalis (eukaryote), and Tupanvirus (giant virus) showed a substantial proportion of Tupanvirus genes that overlap with those of the cellular microbes. In addition, a substantial genome mosaicism was observed, with 51, 11, 8, and 0.2% of Tupanvirus genes best matching with viruses, eukaryota, bacteria, and archaea, respectively. Finally, we found that genes themselves may be subject to lateral sequence transfers. In summary, our data highlight the quantum leap between classical and giant viruses. Phylogenetic and phyletic analyses and the study of protein fold superfamilies confirm previous evidence of the existence of a fourth TRUC of life that includes giant viruses, and highlight its ancestrality and mosaicism. They also point out that best evolutionary representations for giant viruses and cellular microorganisms are rhizomes, and that sequence transfers rather than gene transfers have to be considered.
Introduction
Since the Mimivirus discovery in 2003, dozens of giant viruses that infect Acanthamoeba spp. or Vermamoeba vermiformis have been isolated from various environmental samples, and more recently from animals including humans (La Scola et al., 2003; Raoult et al., 2004; Colson et al., 2017a). Currently, families Mimiviridae (La Scola et al., 2005) and Marseilleviridae (Boyer et al., 2009; Colson et al., 2013b) and isolates that represent new putative families of giant viruses of amoebae, including pandoraviruses (Philippe et al., 2013), pithoviruses (Legendre et al., 2015), faustoviruses (Reteno et al., 2015), Mollivirus (Legendre et al., 2015), Kaumoebavirus (Bajrai et al., 2016), cedratviruses (Andreani et al., 2016), Pacmanvirus (Andreani et al., 2017), and Orpheovirus (Andreani et al., 2018) have been described (Colson et al., 2017b). These giant viruses of amoebae exhibit unique phenotypic and genotypic characteristics that differentiate them from ‘traditional’ viruses and bring them close to small microbes (Lwoff, 1957; Colson et al., 2017a).
These viruses were linked through phylogenomic analyses to poxviruses, asfarviruses, ascoviruses, iridoviruses, and phycodnaviruses (formerly the largest viral representatives), which were grouped in 2001 in a superfamily named nucleocytoplasmic large DNA viruses (NCLDVs) (Iyer et al., 2001, 2006; Raoult et al., 2004). NCLDVs and giant viruses of amoebae were reported to share a putative ancient common ancestor harboring about 50 conserved core genes responsible for key viral functions (Yutin et al., 2009; Koonin and Yutin, 2010; Yutin and Koonin, 2012). Together with a common virion architecture and common major biological features including reproduction within cytoplasmic factories, this contributed to propose reclassifying NCLDVs, mimiviruses and marseilleviruses in a new viral order named Megavirales (Colson et al., 2013a).
The origin and ancestrality of giant viruses has remained controversial. From the onset, when the Mimivirus genome was sequenced in 2004, a phylogeny based on seven concatenated universally conserved genes showed that Mimivirus branched near the origin of the eukaryotic branch, and it was suggested that giant viruses comprised a fourth additional branch in the Tree of Life, alongside Bacteria, Archaea, and Eukarya (Raoult et al., 2004). This hypothesis was thereafter strengthened by both cladistic and phenetic analyses based on informational genes, including those implicated in nucleotide biosynthesis, transcription and translation (Boyer et al., 2010). The hypothesis of the existence of a fourth branch of microbes prompted to define the ‘TRUCs,’ which is an acronym for “Things Resisting Uncompleted Classifications” (Raoult, 2013, 2014). This term was coined because the definition of domains of life by C.R. Woese was based on ribosomal genes that are absent in giant viruses. This proposal of a fourth branch of life comprised by giant viruses has remained controversial and a subject of debate among virologists and evolutionary biologists. Some phylogenetic analyses were deemed to suggest complex patterns of evolutionary relationships for different informational proteins from giant viruses, which even questioned the monophyly of NCLDVs (Yutin and Koonin, 2012; Yutin et al., 2014). A high level of mosaicism has been highlighted for the genomes of giant viruses of amoebae, which was related to sequence transfers with organisms belonging to the three cellular domains of Life (Raoult et al., 2004; Boyer et al., 2009). A substantial gene flow has been also described in NCLDVs including in coccolithoviruses (Wilson et al., 2009; Nissimov et al., 2017). It was suspected that lateral gene transfers blurred phylogenies based on genes shared by giant viruses and cellular organisms (Moreira and Lopez-Garcia, 2009). Several phylogenetic reconstructions in which giant viruses branch within eukaryotes were published (Moreira and Lopez-Garcia, 2009, 2015; Williams et al., 2011), and it was put forward that the universally conserved genes used in phylogeny reconstructions might have been acquired by giant viruses from their proto-eukaryotic hosts (Moreira and Lopez-Garcia, 2009; Yutin et al., 2014). The interpretation of some phylogenies was also that modern giant viruses might originate from smaller NCLDVs (Yutin and Koonin, 2013; Yutin et al., 2014). Conversely, it was proposed that giant viruses might derive from ancestral cellular genomes by reductive evolution (Legendre et al., 2012). Besides, phylogenetic reconstructions supporting the fourth TRUC hypothesis triggered methodological criticisms arguing that they were distorted by long-branch attraction and technical issues, and divergences in their interpretation. However, alternative phylogenies were not accurate either regarding the phylogeny of Archaea, Bacteria, or Eukarya (Williams et al., 2011; Moreira and Lopez-Garcia, 2015). A four-branch topology was also obtained by reconstructing phylogenies that describe the evolution of proteomes and protein domain structures (Nasir et al., 2012; Nasir and Caetano-Anollés, 2015). The genomic and structural diversity embedded in giant virus proteomes was found similar to that of proteomes of cellular organisms with parasitic lifestyles. Beyond, other phylogenies based on RNA polymerase suggested the presence in metagenomes of sequences related to giant virus relatives (Wu et al., 2011; Sharma et al., 2014). As a synthesis, it was deemed that more work is needed on Megavirales phylogenies to clarify if these viruses are monophyletic or have different evolutionary histories (Forterre and Gaia, 2016). Here, we specify the definition of giant viruses, highlight their mosaicism at the genome, structure and sequence level, and strengthen the evidence for their ancestrality and the existence of a fourth TRUC of microbes.
Materials and Methods
Definition of Giant Viruses
We collected and reviewed current knowledge on giant viruses from articles gathered from the NCBI PubMed database and from Google Scholar using as keywords “giant virus”; Megavirales; mimivir∗; marseillevir∗; pandoravir∗; pithovir∗; faustovir∗; mollivirus; cedratvirus; kaumoebavirus; pacmanvirus; virophage; transpoviron. We then compared the phenotypic and genotypic features of these viruses with those used as criteria to define classical viruses and those that are hallmark features of cellular organisms. The list of those criteria is presented in Table 1.
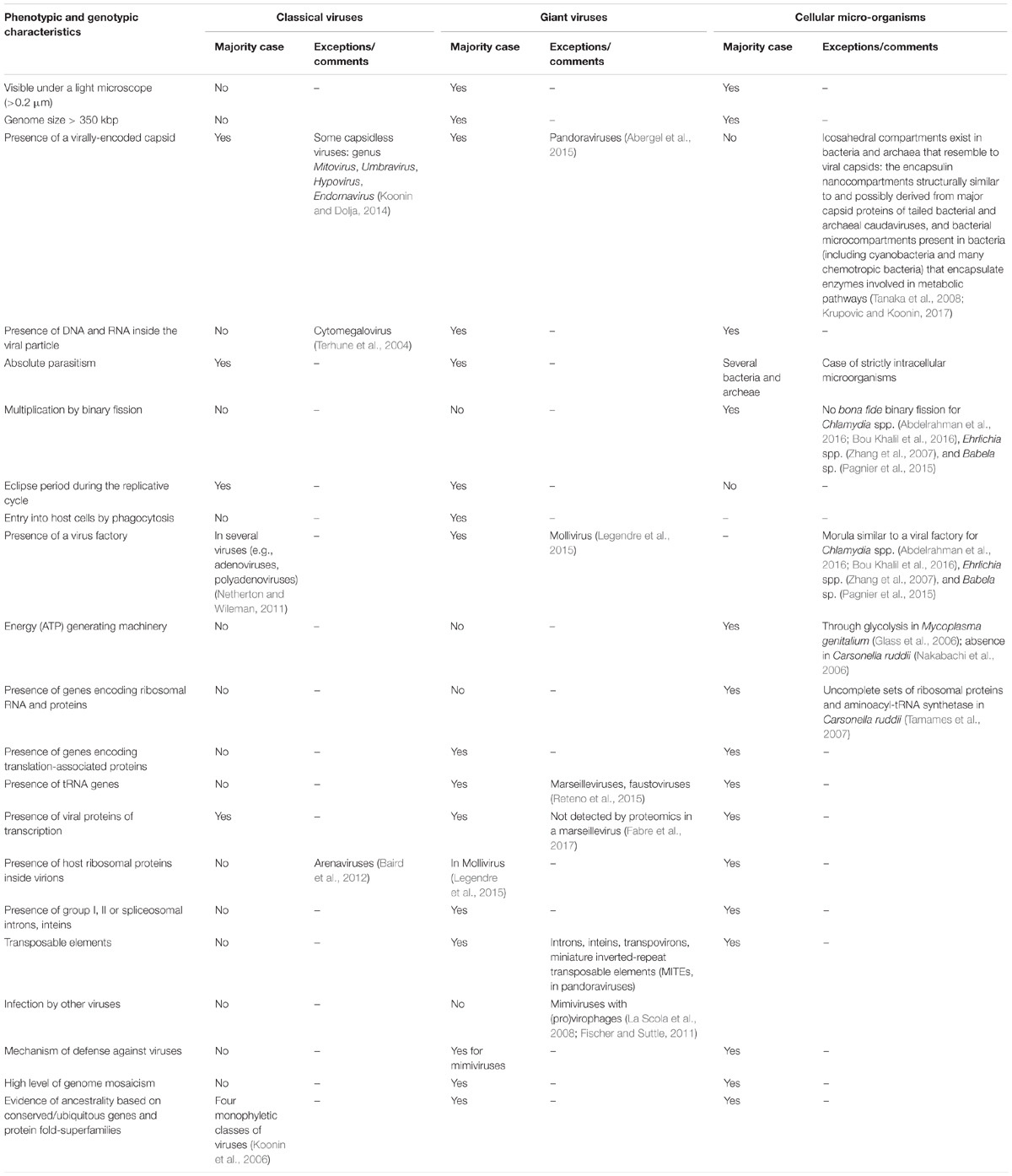
TABLE 1. Comparison of major features used as criteria to define classical viruses with those of giant viruses and to hallmark features of cellular microbes.
Protein Structure Assignment to Viral and Cellular Proteomes
Protein sequences from completely-sequenced proteomes of 80 Megavirales were scanned against the library of hidden Markov models (HMMs) of structural recognition maintained by the SUPERFAMILY database for structure assignment at an E-value cutoff of < 0.0001 (Gough et al., 2001; Gough and Chothia, 2002). The SUPERFAMILY HMMs represent proteins of known three-dimensional (3D) structures and assign each detected occurrence of protein domain into fold superfamilies (FSFs), as defined by the Structural Classification of Proteins (ver. 1.75) database (Andreeva et al., 2008). FSFs are collections of one or more protein families that show recognizable 3D structural and functional similarities, but not necessarily sequence identities, that are indicative of common origin. Thus, FSFs represent highly dissimilar protein domains at the sequence level that have evolved via divergence from a common structure and can still be recognized based on the presence of that conserved structural core by HMMs trained to detect remote homologies. Because of the fast mutation rates of viral genes, it sometimes becomes impossible to generate meaningful global sequence alignments when considering viral and cellular genes together in data matrices. The fast mutation rates, especially when considering proteins separated by large evolutionary distances and involving distantly related taxa, lead to alignment inaccuracies and large number of gaps. In contrast, protein structure evolves at least 3 to 10 times slower than molecular sequences (Illergard et al., 2009) and hence provides an alternative to study the deep evolutionary history of cells and viruses (Nasir et al., 2012; Nasir and Caetano-Anollés, 2015). In parallel, FSF assignments for a total of 102 cellular organisms including an equal number of archaea, bacteria, and eukaryota were retrieved from a previous work during which a total of 1,797 distinct FSF domains had been detected (E-value < 0.0001) (Nasir and Caetano-Anollés, 2015).
Structural Phylogenomics
Using an in-house Python script, we generated a data matrix containing 182 rows (proteomes from 34 archaea, 34 bacteria, 34 eukaryota, and 80 Megavirales members) and 289 columns (FSFs) containing presence/absence information for ‘universal’ FSFs. ‘Universal’ FSFs, by definition, included FSFs that were detected in at least one proteome each from archaea, bacteria, eukaryota, and a Megavirales member. In other words, FSFs unique to one of these four groups (e.g., bacteria-specific FSFs) or shared by 2-to-3 groups of cellular organisms and/or viruses (e.g., FSFs detected in archaea, bacteria, and viruses but not eukaryota) were excluded from our definition of universal FSFs (see (Nasir et al., 2015) for details on FSF groups in cellular organisms and viruses). This data matrix containing 182 proteomes and 289 universal FSFs was imported into the PAUP (ver. 4.0b10) software (Swofford, 2002) for phylogenomic tree reconstruction. Proteomes were treated as taxa and FSFs as characters. Presence/absence of FSFs (represented by 1 and 0, respectively) were used as distinct character states to distinguish taxa. Maximum parsimony method was set as optimality criterion to reconstruct the most parsimonious unrooted phylogenomic tree describing the evolution of sampled proteomes based on the presence/absence of 286 parsimony informative FSF characters. The unrooted reconstructed tree was rooted a posteriori by the branch resulting in minimum increase in overall tree length using the Lundberg method (Lundberg, 1972; see Nasir et al., 2017; Caetano-Anollés et al., 2018 for description and review of rooting methodology). The reliability of the phylogenetic splits was evaluated by running 1,000 bootstraps. Separately, we performed principal coordinate analysis (PCoA) on the same data matrix and plotted the 182 sampled viral and cellular proteomes into 3D space. Proteomes are composed of FSF domains of different evolutionary and geological ages. From a previously reconstructed tree of domains (ToD) (Nasir and Caetano-Anollés, 2015), we retrieved the relative evolutionary ages for each of the 289 universal FSFs. The relative scale reflects the distance of each node (FSF domain) from the root of the ToD and ranges from 0 (closer to the root, most ancient) to 1 (most recent). The node distance (nd) value thus describes a clock-like behavior for the evolution of FSF domains and has previously been linked to the geological record (Wang et al., 2011). Euclidean distance was used to plot proteome dissimilarity based on the 1-nd transformation of the nd scale for each FSF domain in every proteome, as previously (Nasir and Caetano-Anollés, 2015). Since the PCoA is centered around nd variable derived from an evolutionary tree, we refer to this method as evo-PCoA. The evo-PCoA thus projects proteome dissimilarity into 3D space based on differences in the evolutionary ages of components of each proteome. XLSTAT plugin was added to Microsoft Excel for generation of PCoA.
Collection of Orthologous Sequences From Viruses
Analysis was performed as described in previous works (Boyer et al., 2010; Sharma et al., 2014). The genes used in the present study were identified from clusters of orthologous groups of proteins (COGs) involved in nucleotide transport and metabolism and information storage and processing (i.e., categories F, J, A, K, L, and B). These genes comprise proteins that are the most conserved between cellular organisms and viruses (Boyer et al., 2010). They notably include three genes conserved among previously identified Megavirales representatives and in faustoviruses, and that encode DNA-dependent RNA polymerase subunits 1 (RNAP1) and 2 (RNAP2), and family B DNA polymerase (DNApol). Viral orthologs for these three genes were retrieved with the OrthoMCL program (Li et al., 2003) from the gene complements of 317 viral genomes harboring > 100 genes downloaded from the NCBI sequence databases1, and orthologs from nine faustovirus genomes (Benamar et al., 2016) and Mollivirus sibericum (Legendre et al., 2015) were added to this sequence set (Supplementary Table S1).
Collection of Orthologous Sequences From Cellular Organisms
Informational gene homologs from cellular organisms (maximum number: 20,000) were retrieved from the NCBI GenBank non-redundant (nr) protein sequence database by stand-alone BLAST searches with viral sequences as query, using default parameters except for the maximum target number limit, set to 20,000 (Altschul et al., 1990). Homologous sequences were selected from representative species that diverged approximately 500 million years ago using TimeTree (Hedges et al., 2006; Sharma et al., 2014). BLASTp results were filtered by taxon identifiers, selected sequences were downloaded using their GenBank identifier, and duplicates were removed by clustering with the CD-HIT suite, as previously described (Sharma et al., 2014, 2015b).
Multiple Sequence Alignments and Phylogeny Reconstructions
Sequences (Supplementary Table S2) were aligned with the MUSCLE software (Edgar, 2004) and alignments were manually curated. Phylogeny reconstructions were performed using FastTree (Price et al., 2010) with the Maximum Likelihood method, and the CAT 20 model that analyses the alignment site by site and reduces long branch attraction artifacts (Lartillot et al., 2007). Then, trees were visualized using FigTree2. Confidence values were determined by the Shimodaira-Hasegawa (SH) test using FastTree (Price et al., 2010).
Comparison of Informational Genes Repertoires
Hierarchical clustering was performed with the Pearson distance method and the TM4 multi-package software, as previously described (Sharma et al., 2015a,b). This analysis relied on the comparison of the presence/absence patterns of 726 COGs involved in nucleotide transport and metabolism and information storage and processing in the gene contents of viruses and of selected bacterial, archaeal, and eukaryotic representatives (Sharma et al., 2015a,b). Viral orthologs were identified through BLASTp searches using these 726 COGs. BLAST searches were performed with default parameters, except for the maximum target number limit, set to 20,000.
Comparison of Gene Repertoires From a Representative of Each of the Three Cellular Domains of Life and From a Giant Virus, and Construction of the Rhizome of Genomes and Genes
Comparison of the gene contents was performed for three members of cellular domains that were selected because they harbor small genomes and are intracellular parasites [namely Encephalitozoon intestinalis (an eukaryote) (Corradi et al., 2010), Methanomassiliicoccus luminyensis (an archaeon) (Gorlas et al., 2012), Rickettsia bellii (a bacterium) (Ogata et al., 2006)], and for Tupanvirus soda lake (Abrahao et al., 2018), a recently described giant virus that was selected here because it has a particularly large gene content and harbors the largest set of translation components among giant viruses. This comparison used the ProteinOrtho v5 tool with 1e-3, 20 and 30% as thresholds for e-value, amino acid identity, and coverage of aligned sequences, respectively (Lechner et al., 2011). In addition, best BLASTp hits against the NCBI GenBank protein sequence database were obtained for these four organisms. The “rhizomes” of the genomes were built using the Circos tool3. Rhizomes consist in a representation of the genome evolution and mosaicism that takes into consideration the fact that genes from this genome as well as intragenic sequences do not have the same evolutionary history, and can result from exchanges, fusions, recombination, degradation, or de novo creation (Raoult, 2010). Rhizomes, which are devoid of a center, were proposed as a better paradigm of genetic evolution than trees (Deleuze and Guattari, 1976; Raoult, 2010). Rhizomes built here show in a single figure, for all the genes from a given virus or cellular organisms, the taxonomy of their best BLASTp hits that represent putative donors or acceptors involved in sequence transfers, as well as the ORFans (sequences devoid of homolog in databases). Furthermore, a rhizome of genes was also determined for the genes encoding a methionyl-tRNA synthetase shared by the four organisms, by performing BLASTp searches with fragments obtained from this gene by cutting its amino acid sequence into 40 amino acid-long fragments that overlapped with a sliding window of 20 amino acids.
Results and Discussion
Phylogenetic Analyses of Protein Structural Domains of Viral and Cellular Proteomes
A total of ∼1,200 folds, ∼2,000 superfamilies, and ∼5,000 families of structural domains encompass the entire evolutionary and functional diversity of the protein world. The history of these folds, superfamilies and families has been traced with phylogenomic methods by studying the entire repertoires of proteins (proteomes), beginning with a study of a small set of 32 completely sequenced genomes (Caetano-Anollés and Caetano-Anollés, 2003) and continuing with a recent extended analysis of thousands of viral and cellular genomes (Nasir and Caetano-Anollés, 2015). Timelines of domain history could be calibrated with a molecular clock that relates them to the geological record (Wang et al., 2011). The timelines showed that the oldest domain families harbored ‘Rossmann-like’ α/β/α-layered and bundle structures typical of globular proteins, followed by barrel structures typical of membrane and metabolic proteins (Caetano-Anollés et al., 2012). The oldest of these structures are predominant in membrane-associated proteins, suggesting a very early onset of cellular structure. Their link to metabolism, but not translation, also suggests the late development of the genetic code and the late appearance of the ribosome (Harish and Caetano-Anollés, 2012; Caetano-Anollés et al., 2013). Remarkably, the late arrival of modern genetics ∼3 billion years (Gy) ago signals the end of a period responsible for the primordial cellular origin of viruses, clearly evident by the fact that the oldest superfamilies are common to cells and viruses (Nasir and Caetano-Anollés, 2015). In addition, these data also indicated that RNA polymerases are more ancient than the ribosome. Such diversification occurred prior to the appearance of the cellular domains of life.
A previous phylogenomic data-driven analysis of proteomes confirmed the early cellular origin of viruses and the rise of viral RNA proteomes followed by that of DNA viruses and Megavirales representatives (Nasir and Caetano-Anollés, 2015). Here we focused on the evolutionary relationship of Megavirales and cellular organisms. Out of all possible FSF domains (Figure 1), we selected 289 that were universal, i.e., that were shared by viruses and cellular organisms. We then used this set to build a phylogeny of proteomes (Figure 2). Megavirales representatives appear as a basal group in the tree of proteomes, which is consistent with results from sequence analyses performed here and previously (Boyer et al., 2010; Sharma et al., 2014). The subgroup that was closest to cellular organisms was family Mimiviridae, followed by family Phycodnaviridae and then groups comprised by family Marseilleviridae and by faustoviruses, mollivirus, and pandoraviruses. Similar phylogenetic patterms were revealed when we used multidimensional scaling approaches to explore the temporal space of ages of individual structural domains in proteomes (Figure 3). We found distinct temporal clouds of proteomes for viruses and organisms belonging to Archaea, Bacteria, and Eukarya. The Mimiviridae group was clearly dissected from the main viral cloud, which was temporally closer to cellular proteomes, suggesting their late appearance in viral evolution. Again, the family Phycodnaviridae appeared between the family Mimiviridae and the rest of the viral cloud. In terms of the proportions of FSFs detected in giant viral groups, asfarviruses have a proteome that is more similar to that of faustovirus, which is consistent with phylogenetic analysis of sequences. However, when considering raw number, mimiviruses have more FSFs in common with faustovirus. Finally, when plotting phylogenetic indices measuring the levels of homoplasy of the MP tree reconstruction (corresponding to Figure 2) against age of the phylogenetic character (fold superfamily), high retention indices, especially for lower nd values (oldest domains), indicated excellent fit of characters to the phylogeny (Figure 4). Homoplasy indicates the level of independent gain of characters in lineages and is a good indicator of deviations from vertical inheritance (Farris, 1983). The levels of homoplasy were moderate for protein folds, showing that the vertical signals override the horizontal signals.
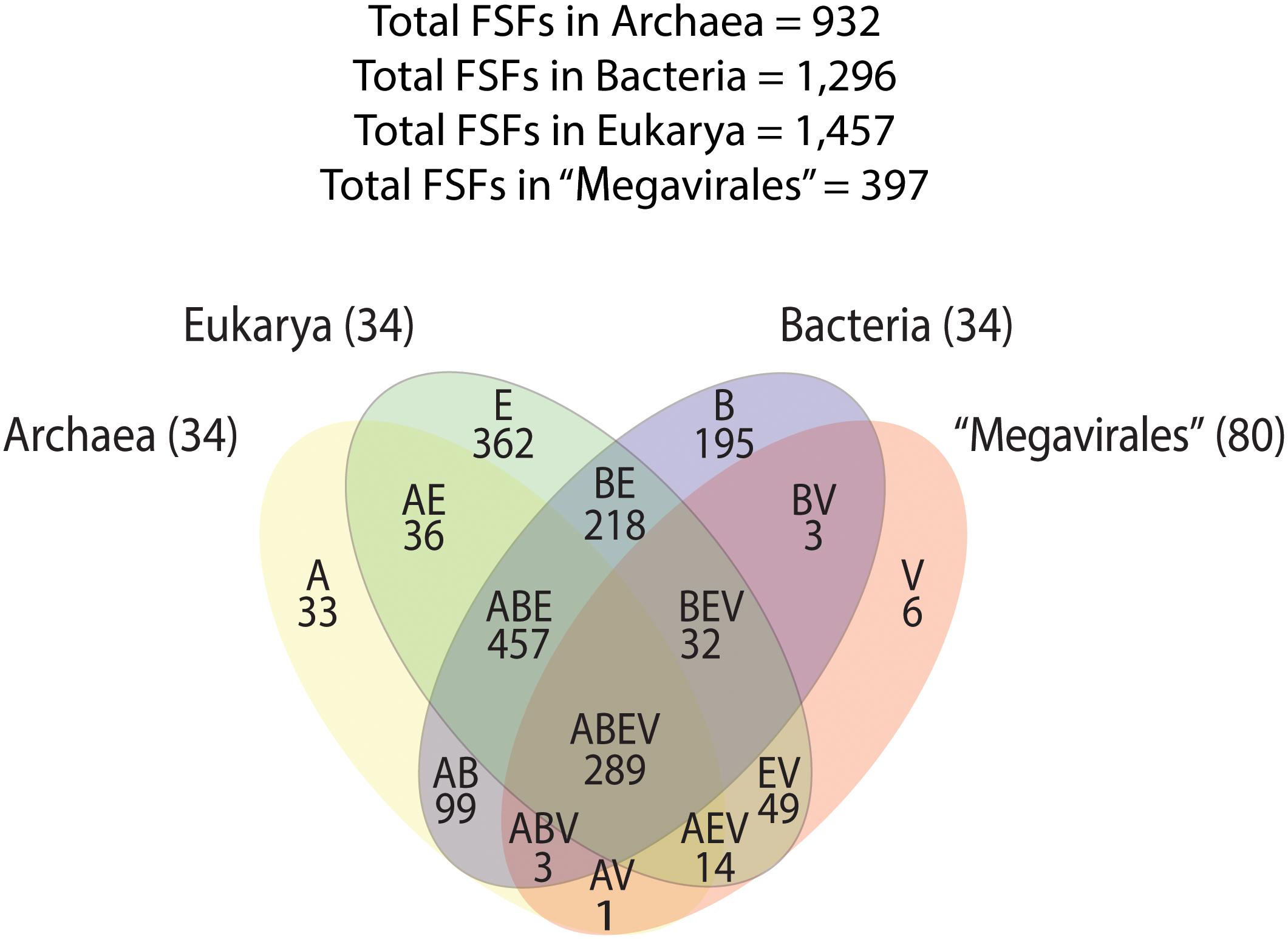
FIGURE 1. Venn diagram displaying FSF distribution and sharing patterns among Archaea, Bacteria, Eukarya, and Megavirales. A, Archaea; B, Bacteria; E, Eukarya; FSF, fold superfamilies; V, viruses.
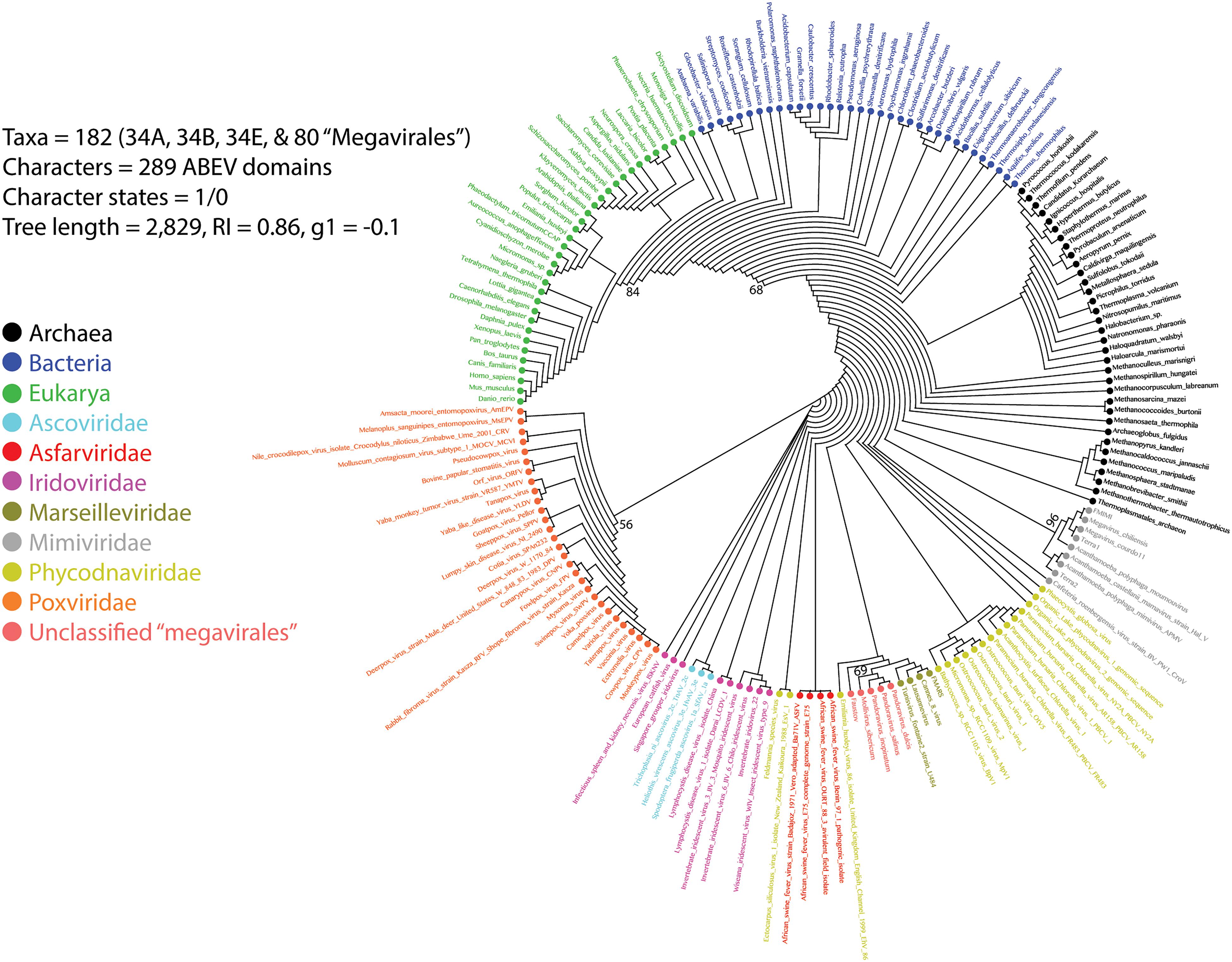
FIGURE 2. Phylogeny of proteomes describing the evolution of 182 proteomes randomly sampled from cellular organisms and viruses. The universal Tree of Life is rooted using Weston’s generality criterion. The 102 cellular proteomes are from Nasir and Caetano-Anollés (2015).
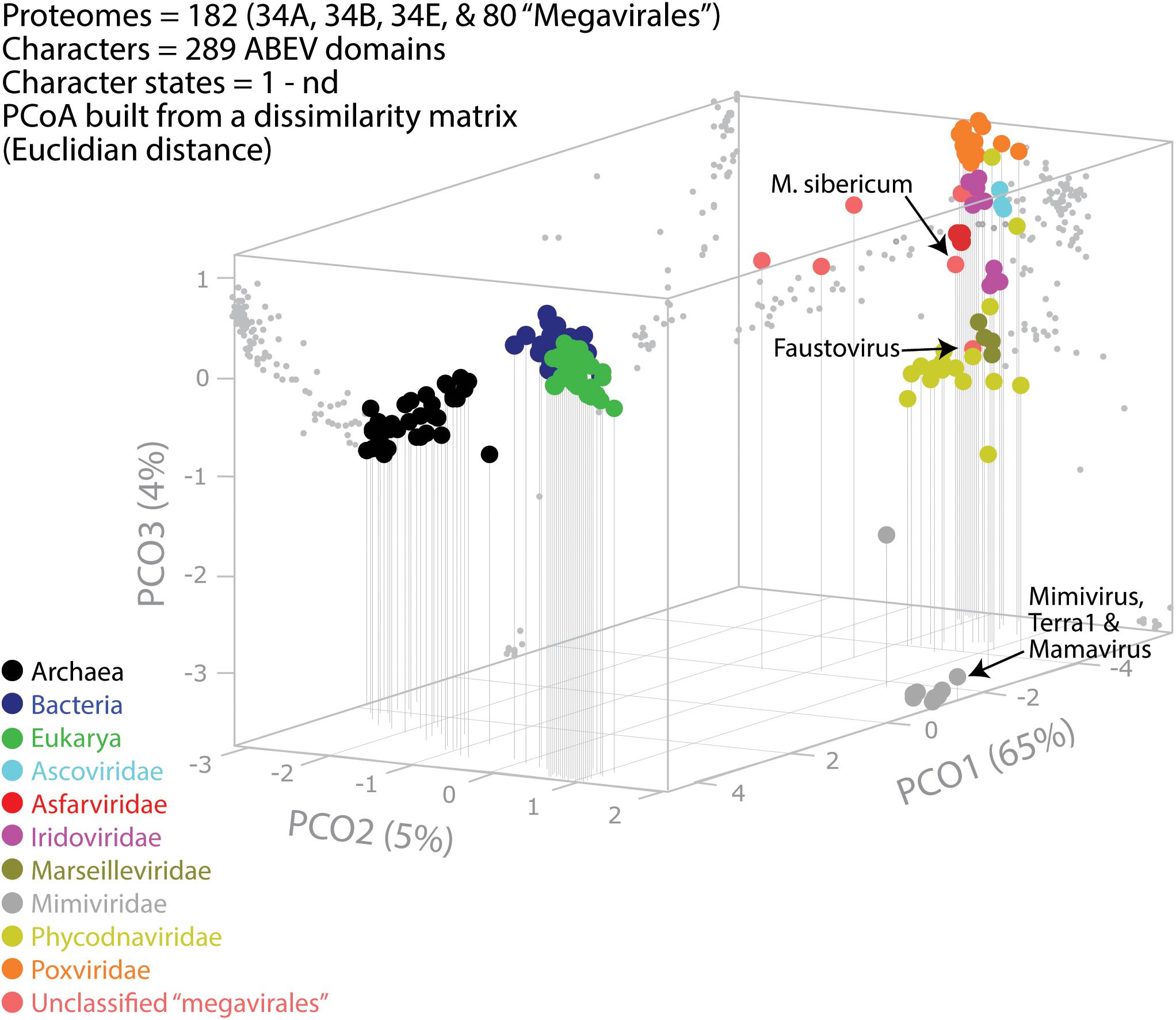
FIGURE 3. Evolutionary principal coordinate (evoPCO) analysis plot portrays in its first three axes the evolutionary distances between cellular and viral proteomes. The percentage of variability explained by each coordinate is given in parentheses on each axis. Data points of the 3-dimensional scatter plot describing temporal clouds are mapped onto projections planes and connected with vertical leading drop lines along the PCO3 axis. The list of whole coordinate information for building the PCoA plot of this figure is provided in Supplementary Table S3.
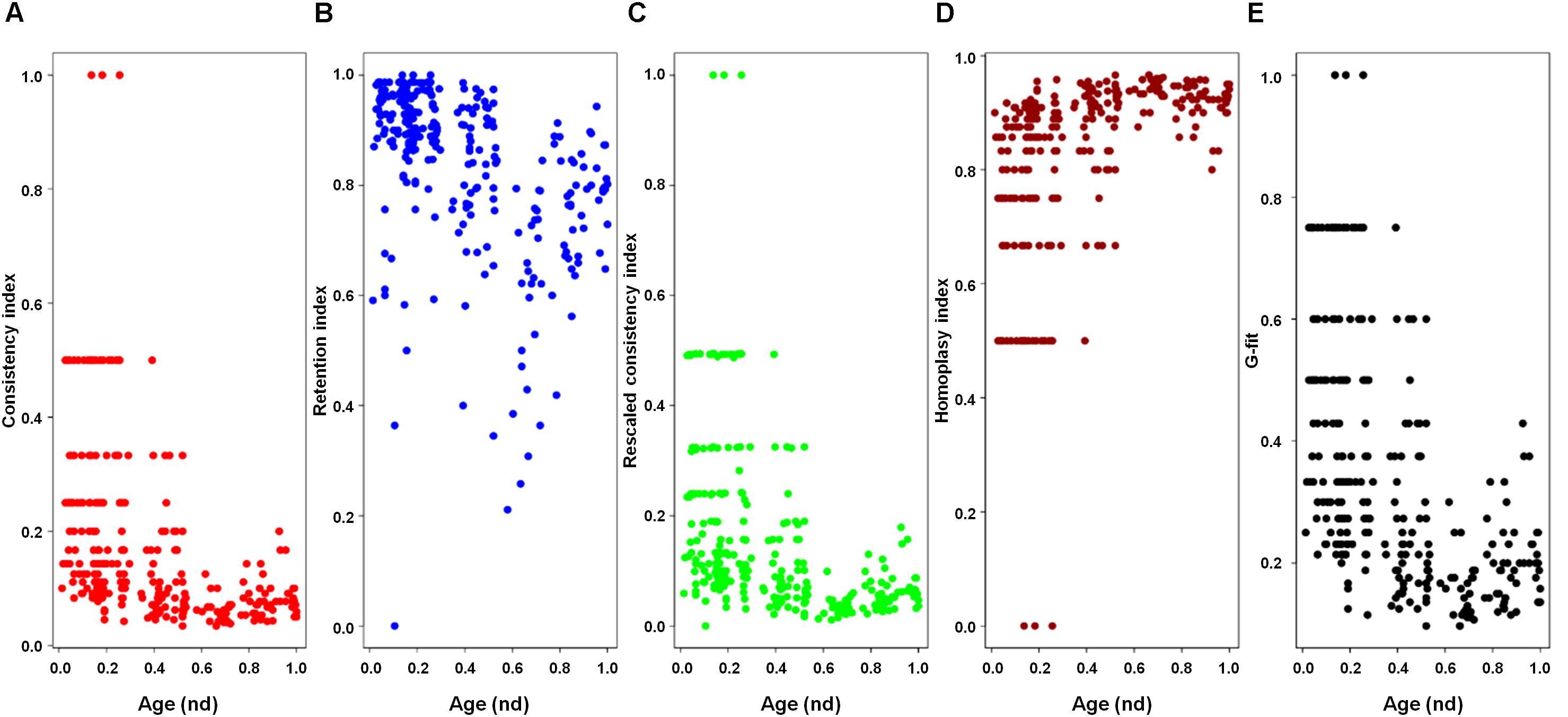
FIGURE 4. Plots of the indices of the phylogenetic tree of proteomes describing the evolution of 182 proteomes randomly sampled from cellular organisms and viruses (corresponding to Figure 2) against the age of the phylogenetic character [fold superfamily (FSF)]. Five measures of the levels of lateral sequence transfers for the maximum parsimony tree reconstruction performed in the present study, namely consistency index (A), retention index (B), rescaled consistency index (C), homoplasy index (D), and G-fit (E), are plotted against the age of the phylogenetic character FSF [measured as node distance (nd) values] for 289 characters (FSF) shared by archaea, bacteria, eukaryota, and viruses. High retention indices, especially for lower nd values (corresponding to older domains), indicates excellent fit of the characters to the phylogeny.
Phylogenetic Analyses of RNA and DNA Polymerases and Phenetic Comparison of Informational COGs
As shown in Figures 5, 6, trees reconstructed using both RNA polymerase subunit sequences (RNAP1 and 2) from members of Megavirales (including recently described giant viruses of amoebae), Bacteria, Archaea, and Eukarya clearly displayed a topology with four branches. The Megavirales group exhibits a considerable genetic diversity. Regarding phylogeny reconstruction based on DNA polymerases present in archaea, eukaryotes and giant viruses, giant viruses are separated into two groups. Faustoviruses and asfarviruses are clustered together and comprise sister branches, apart from other giant viruses that form an independent and strongly supported cluster (Figure 7). Hierarchical clustering analysis was performed based on a binary presence/absence matrix constructed using 727 informational COGs present in 143 representative genomes of cellular organisms from Bacteria, Archaea and Eukarya, and viruses from Megavirales (Figure 8). This phenetic analysis based on informational genes also showed a four-branch topology, Megavirales being a distinct branch alongside Eukarya, Archaea, and Bacteria.
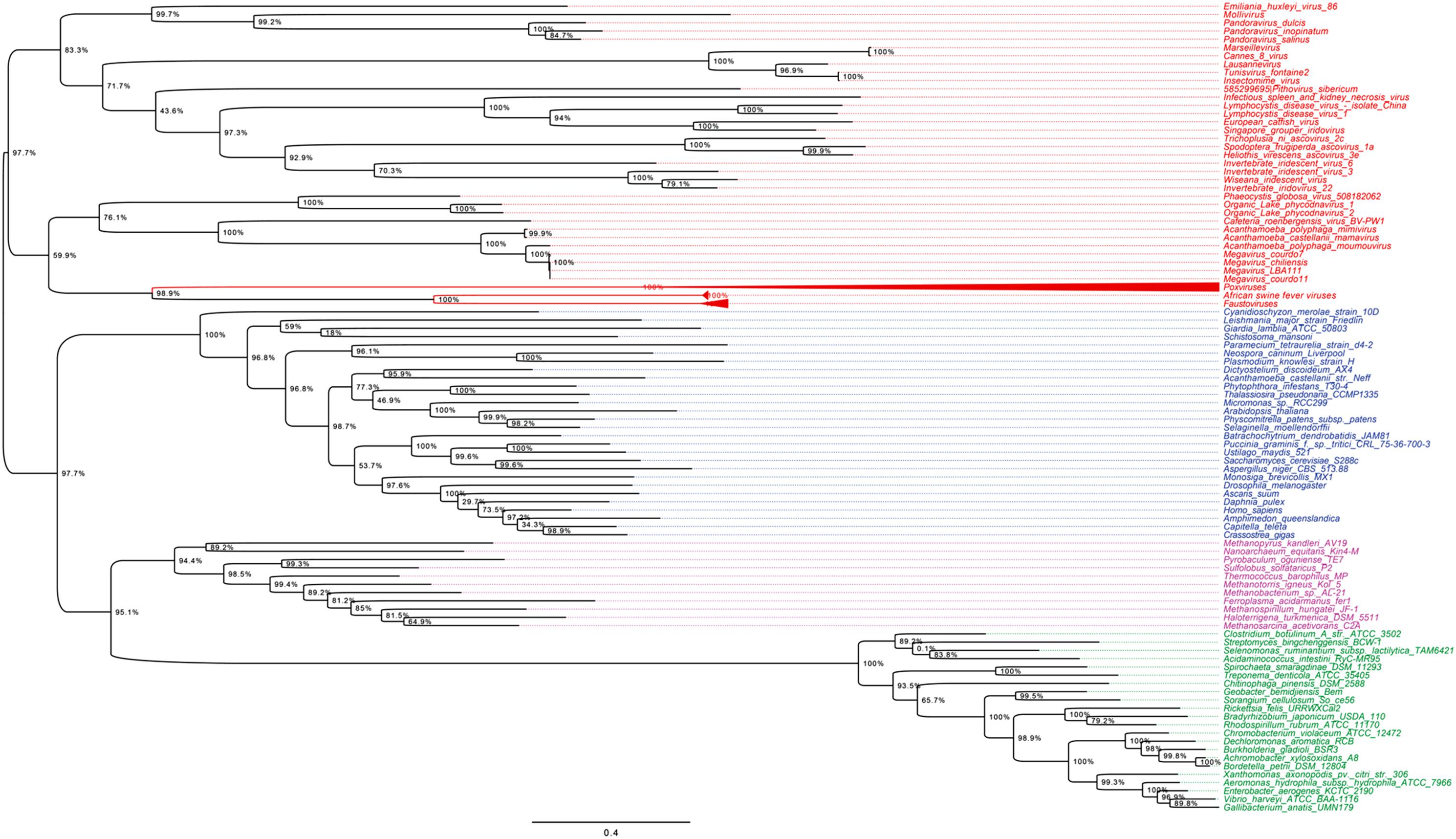
FIGURE 5. RNAP1 phylogenetic tree. The RNAP1 tree was built by using aligned protein sequences from Megavirales (red), Bacteria (green), Archaea (pink), and Eukarya (blue). Confidence values were calculated by the Shimodaira-Hasegawa (SH) test using the FastTree program (Price et al., 2010). Average length of sequences was 1,336 amino acids. The scale bar represents the number of estimated changes per position.
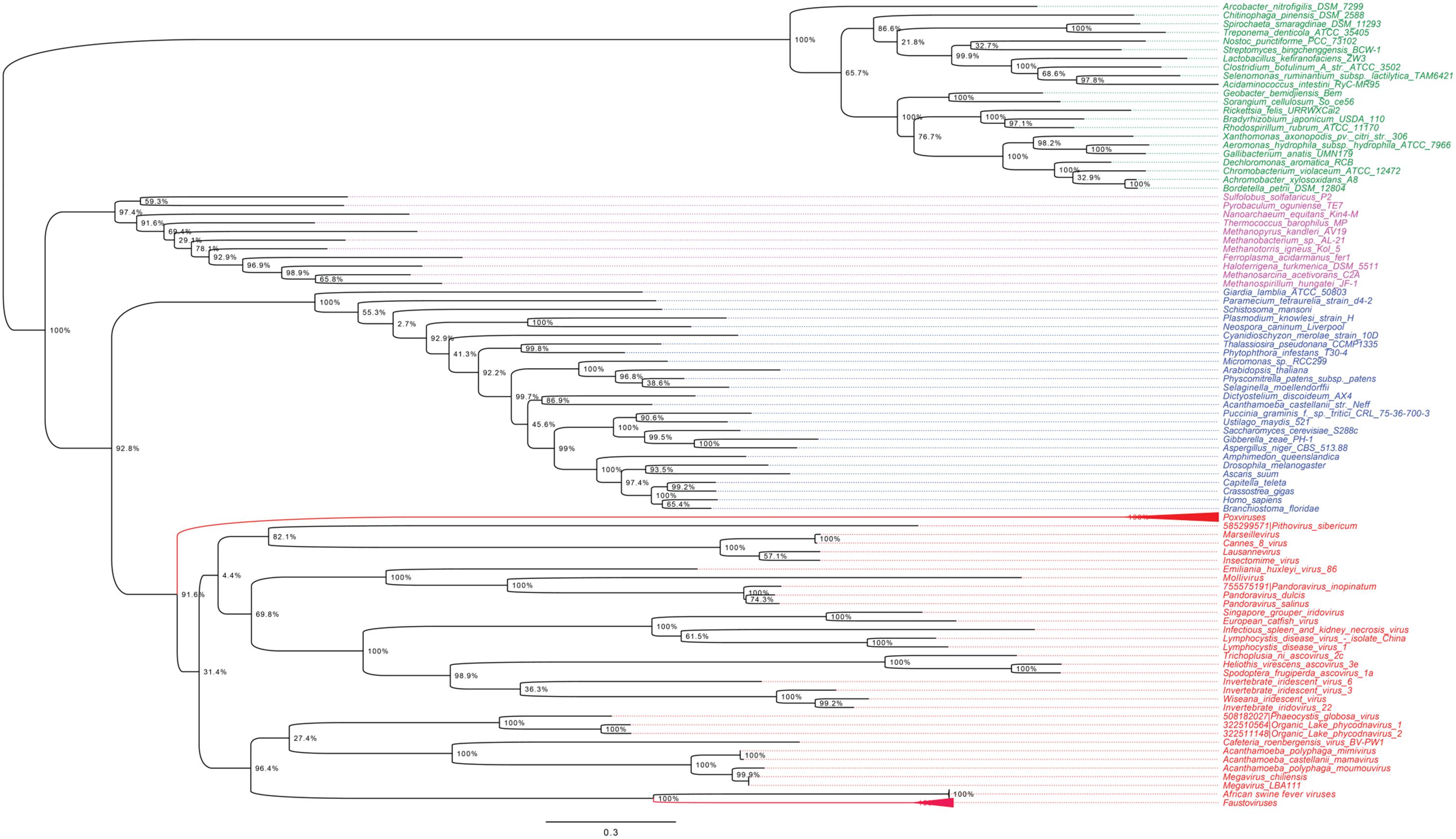
FIGURE 6. RNAP2 phylogenetic tree. The RNAP2 tree was built by using aligned protein sequences from Megavirales (red), Bacteria (green), Archaea (pink), and Eukarya (blue). Confidence values were calculated by the SH test using the FastTree program (Price et al., 2010). Average length of sequences was 1,188 amino acids. The scale bar represents the number of estimated changes per position.

FIGURE 7. DNA polymerase phylogenetic tree. The DNA polymerase tree was built by using aligned protein sequences from Megavirales (red), Bacteria (green), Archaea (pink), and Eukarya (blue). Confidence values were calculated by the SH support using the FastTree program (Price et al., 2010). Average length of sequences was 1,134 amino acids. The scale bar represents the number of estimated changes per position.
FIGURE 8. Hierarchical clustering by phyletic pattern based on the presence/absence of informational Clusters of Orthologous Groups (COGs) of proteins. The Megavirales members are represented in red, Bacteria members in green, Archaea members in pink, and Eukarya members in blue.
Pangenome and Core Genome for One Member of Each of the Three Cellular Domains of Life and of a Giant Virus
A pangenome and core genome was determined for one representative of each of the four TRUCs of microbes: namely R. bellii (bacteria, 1,430 genes), M. luminyensis (archaea, 2,533 genes), E. intestinalis (eukaryota, 1,910 genes), and Tupanvirus soda lake (giant virus, 1,269 genes). The pangenome describes the full complement of genes in a group of organisms, in our case the four microbes, and is comprised by the core genome that contains genes present in all 4 microbes and by the dispensable genome composed of genes that are unique to each microbe and genes absent from one or more microbes. The pangenome of these four microbes was composed of 6,531 genes, and their core genome (shared by all four organisms) was composed of 33 genes that represented between 1.3 and 2.6% of their gene contents. This core genome included notably genes encoding a DNA-directed RNA polymerase, a ribonucleoside-diphosphate reductase, a translation elongation factor 2, and several aminoacyl-tRNA synthetases. A majority of these genes therefore consisted of translation components. In addition, 23 (1.6%), 68 (5.4%), 13 (0.7%), and 68 (5.4%) genes from R. bellii, M. luminyensis, E. intestinalis, and Tupanvirus, respectively, had homologs in the genomes of two other microbes. Finally, 261 genes in R. bellii (18.3%), 362 in M. luminyensis (14.3%), 298 in E. intestinalis (15.6%), and 132 in Tupanvirus (10.4%) had homologs in at least one of the three other microbes. These results show that beyond the fact that the number of genes for Tupanvirus is in the same order of magnitude than for the three cellular microorganisms, a substantial proportion of the genes of this giant virus overlaps with those of the bacteria, the archaeon and the eukaryote.
Rhizomes of Genomes and Genes as Appropriate Representations of the Origin and Evolution of Members From the Four TRUCs of Microbes
A substantial genome mosaicism, consisting of genomes composed by genes with sequences suggesting different evolutionary origins and histories, was observed for representatives of the four TRUCs, including R. bellii, M. luminyensis, E. intestinalis, and Tupanvirus (Figure 9). This mosaicism was particularly predominant in the Tupanvirus genome as described previously (Abrahao et al., 2018), with 51, 11, 8, and 0.2% of its genes best matching with viruses, eukaryota, bacteria, and archaea, respectively, but it was a shared feature of the three non-eukaryotic microorganisms. This illustrates that a rhizome is the most appropriate representation of the evolutionary history at a genome scale, as individual genes can have distinct and distant origins (Raoult, 2010). Such representation notably takes into account introgressive descent as a result of lateral sequence transfers. Moreover, it appears that genes themselves may be subject to lateral sequence transfer rearrangements (through gene conversion), as shown here for the case of the methionyl-tRNA synthetase encoding gene of the four microorganisms (Figure 10). Indeed, 40 amino acid-long fragments of these genes alternately found as best hits, apart from relatives from the same family or genus, sequences from archaea, bacteria, eukaryota, or viruses. Such a gene sequence mosaicism was particularly broad for Tupanvirus and M. luminyensis. For the case of Tupanvirus soda lake, 15, 3, 2, and 1 methionyl-tRNA synthetase gene fragments found as best hits an eukaryote, a virus, a bacterium and an archaeon, respectively. This was also remarkably exemplified with the case of the glutaminyl-tRNA synthetase of Klosneuvirus, a mimivirus relative (Schulz et al., 2017). Indeed, fragments of this glutaminyl-tRNA synthetase gene showed a mixture of sequences from eukaryotes, bacteria and of unknown sources, or of sequences retrieved from metagenomes, in particular those of Antarctic dry valleys (Abrahao et al., 2018). These findings make the notion of gene lateral transfer obsolete, as sequences, rather than genes, are transferred (Merhej et al., 2011). Thus, the source of a gene may be better defined by a rhizome than by a tree, as previously proposed for organisms (Raoult, 2010) (Figure 11). Examples of chimeric genes have been previously described. Thus, ORF13 of the Sputnik virophage encodes a primase-helicase whose N-terminal region is of archaea-eukaryotic source and C-terminal portion was inferred to originate from giant viruses (La Scola et al., 2008). In the fern Adiantum capillus-veneris, a chimeric photoreceptor was identified that may have been critical in the divergence and rise of some fern species under low luminosity environments (Kawai et al., 2003). More broadly, it has been described that the creation of novel chimeric genes, referred as chimeric nuclear symbiogenetic genes (S-genes), occurred during eukaryogenesis through the fusion of bacterial and archaeal genes; this gave rise in early eukaryotes to novel chimeric proteins with central functions (Meheust et al., 2018). These data confirm and expand to genes the concept that no single tree can define the chimeric nature of genomes, as genes themselves are mosaics (Dagan and Martin, 2006; Merhej et al., 2011). As a consequence, trees made with homologous sequences make no sense if not all fragments of these sequences have a common source. Phylogeny reconstructions based on concatenated genes are still worse when the trees built based on the separate genes do not have the same topology, because they consist in mixing sequences from different, and eventually very distant, origins.
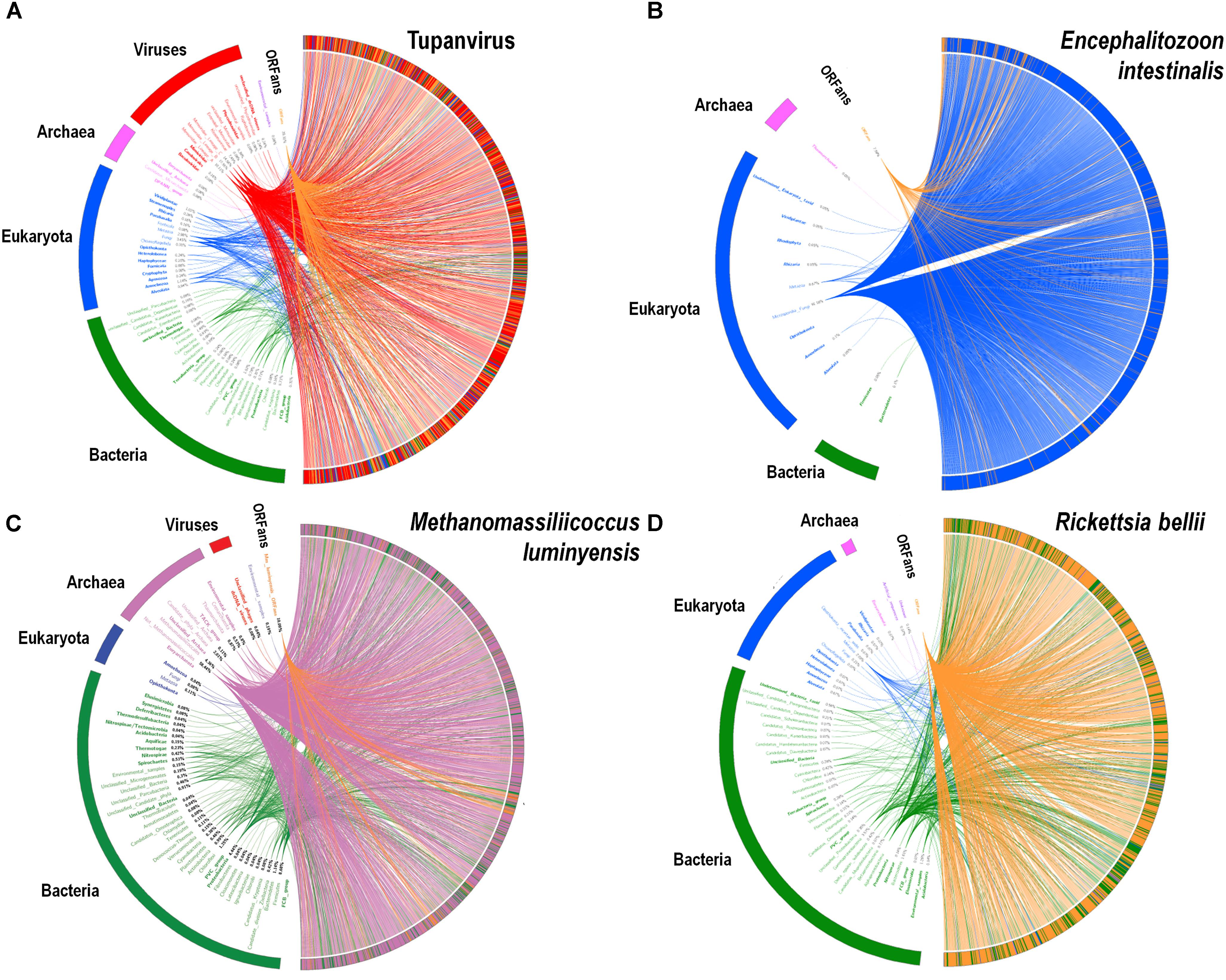
FIGURE 9. Rhizomes of genomes illustrative of the mosaicism of the genomes of representatives of the four TRUCs of microbes including Tupanvirus soda lake (a mimivirus) (A); Encephalitozoon intestinalis (a microbial eukaryote) (B); Methanomassiliicoccus luminyensis (an archaeon) (C); and Rickettsia bellii (a bacterium) (D). The genes of these four microorganisms were linked to their most similar sequences in the NCBI GenBank protein sequence database according to the BLAST program (https://blast.ncbi.nlm.nih.gov/Blast.cgi), classified according to their belonging to viruses, eukaryotes, bacteria or archaea, and integrated in a circular gene data visualization. The figures were performed using the CIRCOS online tool (http://mkweb.bcgsc.ca/tableviewer/visualize/). Circular representations in A and C are the same than those produced for figures from articles Abrahao et al. (2018) and Levasseur et al. (2017), respectively, as they originate from the same data. These representations are licensed under CC BY 4.0 (https://creativecommons.org/licenses/by/4.0/) and CC-BY-NC (https://creativecommons.org/licenses/by-nc/4.0/), respectively.
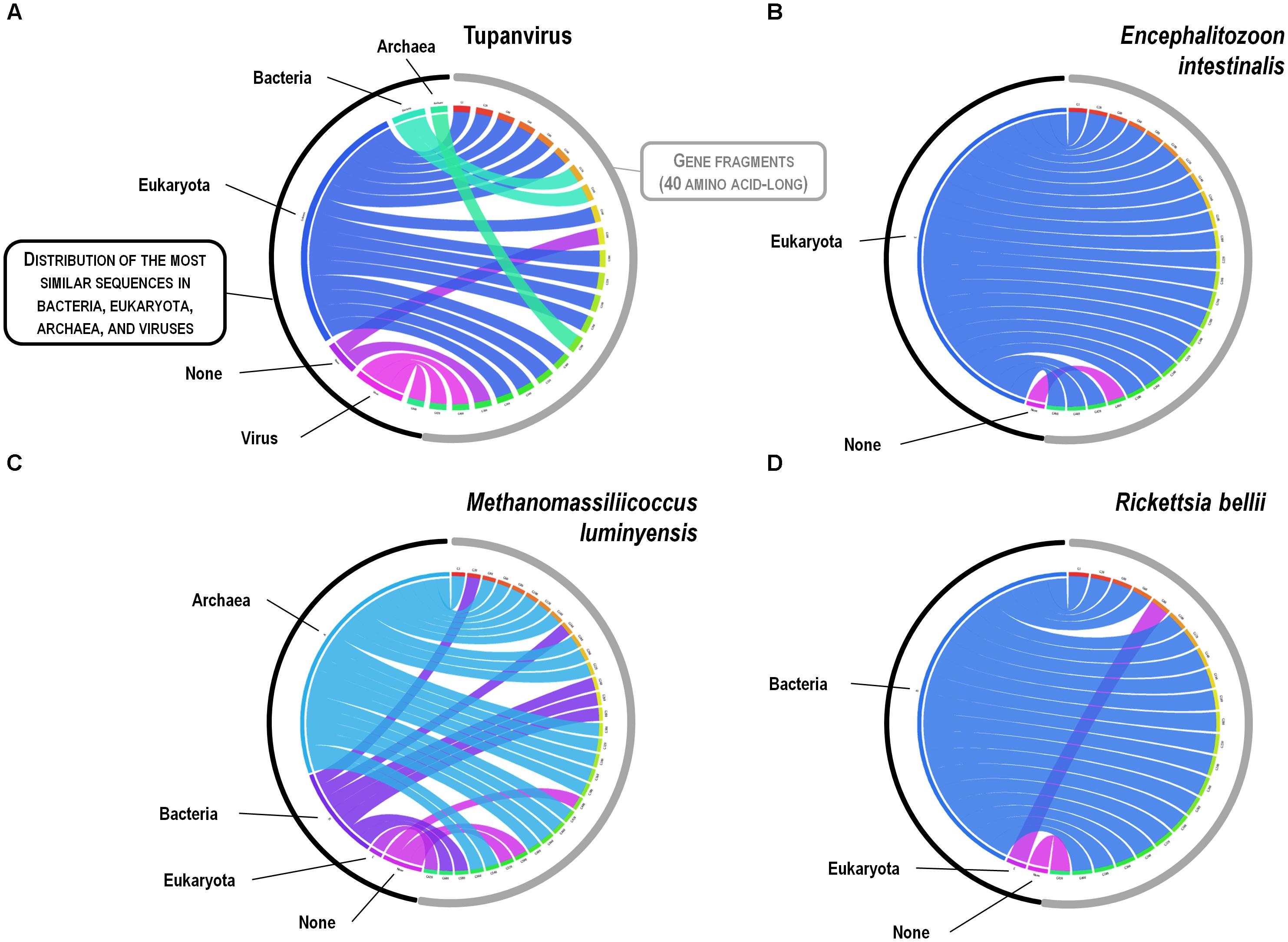
FIGURE 10. Rhizomes of methionyl-tRNA synthetase gene fragments illustrative of the mosaicism of the genes of representatives of the four TRUCs of microbes including Tupanvirus soda lake (a mimivirus) (A); Encephalitozoon intestinalis (a microbial eukaryote) (B); Methanomassiliicoccus luminyensis (an archaeon) (C); and Rickettsia bellii (a bacterium) (D). Forty amino acid-long fragments of the methionyl-tRNA synthetase encoding genes of the four microorganisms were linked to their most similar sequences in the NCBI GenBank protein sequence database according to the BLAST program (https://blast.ncbi.nlm.nih.gov/Blast.cgi), classified according to their belonging to viruses, eukaryotes, bacteria or archaea, and integrated in a circular gene data visualization. The figures were performed using the CIRCOS online tool (http://mkweb.bcgsc.ca/tableviewer/visualize/).
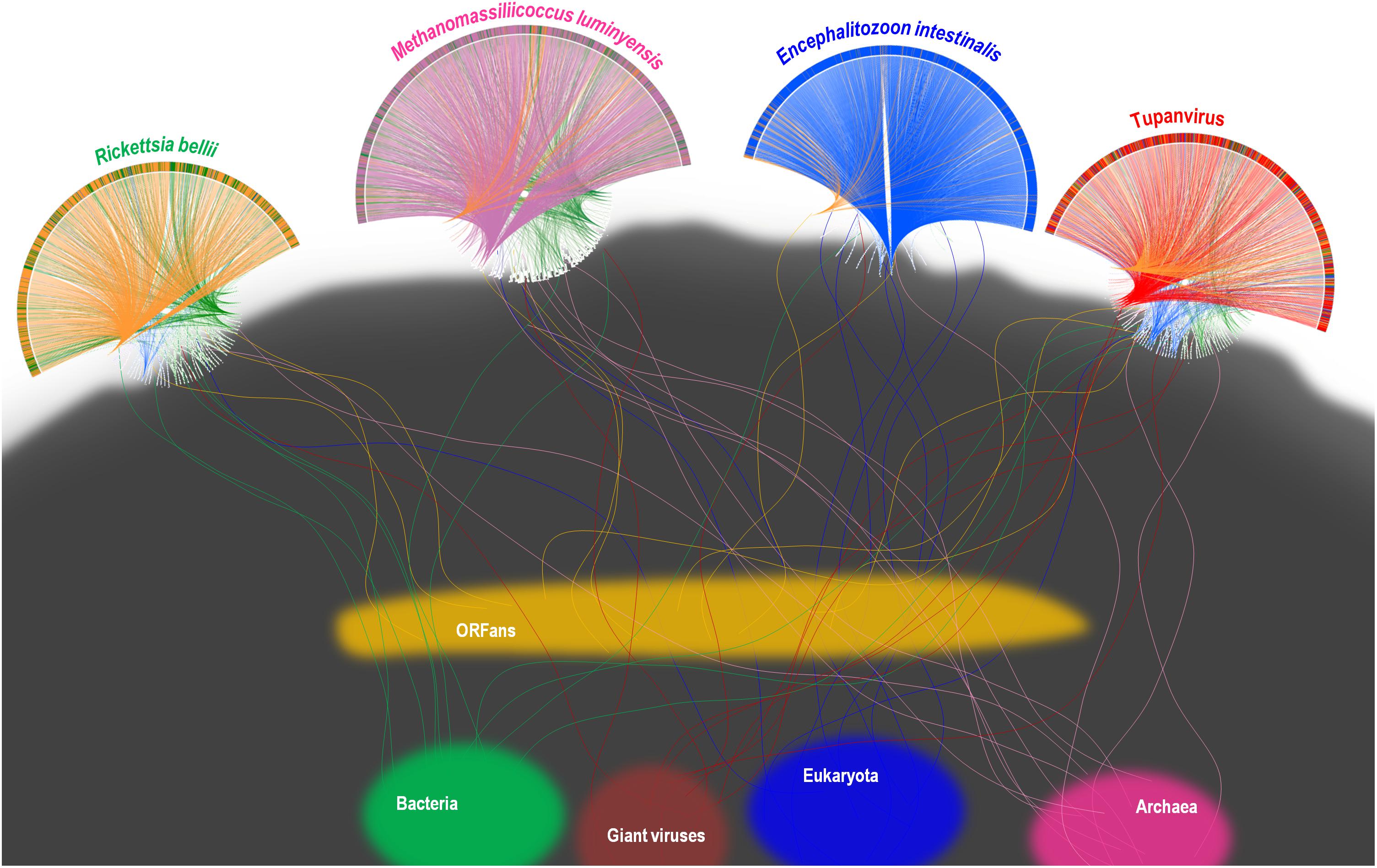
FIGURE 11. Representation as a rhizome of the genetic evolution for four current intracellular parasites of the four TRUCs of microbes with a comparable genome size, including Rickettsia bellii (a bacterium), Methanomassiliicoccus luminyensis (an archaeon), Encephalitozoon intestinalis (a microbial eukaryote), and Tupanvirus soda lake (a mimivirus). Rhizomes consist in a representation of genome evolution and mosaicism that takes into account that genes and intragenic sequences do not have the same evolutionary history, being proposed as better paradigm of genetic evolution than phylogenetic trees. The genomes of each of the four represented current microorganisms harbor mixtures of sequences of different origins. Sequences corresponding to current bacteria, Archaea, eukaryota, giant viruses, and to ORFans are colored in green, purple, blue, red, and orange, respectively. Rhizomes of the genomes of Tupanvirus and Methanomassiliicoccus luminyensis were adapted from same representations than representations from Levasseur et al. (2017) and Abrahao et al. (2018), respectively, licensed under CC BY 4.0 (https://creativecommons.org/licenses/by/4.0/) and CC-BY-NC (https://creativecommons.org/licenses/by-nc/4.0/), respectively (see legend to Figure 9).
Definition Criteria for Giant Viruses or Megavirales
As shown in Table 1, giant viruses exhibit unique phenotypic and genotypic features that differentiate them from ‘classical’ viruses, indicate their much greater complexity, and bring them close to small micro-organisms. These characteristics can be classified as follows: (i) Giant sizes of the virions and their genomes. (ii) Complexity, with presence in virions of dozens of proteins, and of messenger RNA. (iii) Presence of translation components unique among viruses; in this view, the recent characterization of klosneuviruses (Schulz et al., 2017) and tupanviruses (Abrahao et al., 2018) has led to a considerable expansion of the set of such translation components. Notably, the tupanvirus isolates encode for 67–70 tRNA, 20 aminoacyl tRNA-synthetases, and 11 translation factors. (iv) Presence of a specific mobilome in mimiviruses that includes virophages, transpovirons, introns, and endonucleases (Desnues et al., 2012), as well as MIMIVIRE, a defense system against virophages (Levasseur et al., 2016b; Dou et al., 2018). (v) Based on phylogenetic, phyletic, and protein fold superfamilies analyses, delineation of a fourth group of micro-organisms comprised by giant amoebal viruses alongside bacterial, archeal and eukaryotic microbes, and evidence of an archaic origin (Boyer et al., 2010; Sharma et al., 2014; Nasir and Caetano-Anollés, 2015). Moreover, the recent comparison of the genomes of a fossil and a modern pithovirus highlighted that giant viruses evolve with a mutation rate estimated to be lower than that of RNA viruses and comparable to those determined for bacteria and archaea, and by classical mechanisms of evolution, including through long-term fixation of genes that are acquired by horizontal gene transfer (Levasseur et al., 2016a).
Giant viruses of amoebae certainly exhibit several criteria that are hallmarks and definition criteria of viruses. These include the occurrence of an eclipse phase during their replicative cycle, an obligatory replication into host cells, and the presence of a capsid (Lwoff, 1957; La Scola et al., 2003; Raoult and Forterre, 2008). Nevertheless, regarding the capsid, pandoraviruses, pithoviruses, mollivirus, and cedratviruses have virions surrounded by a tegument-like structure and no known capsid morphology (Philippe et al., 2013; Yutin and Koonin, 2013; Legendre et al., 2014, 2015). Pandoraviruses do not have a recognizable capsid-encoding gene, pithoviruses have a barely identifiable capsid-encoding gene, while capsid proteins are detected in Mollivirus virions but they are not part of the virion structure. Other giant virions with an ovoid or spherical shape such as cedratviruses and Orpheovirus are also devoid of a morphology resembling those provided by known capsids. An atypical capsid structure was previously described for Megavirales representatives. Thus, most poxviruses have brick-shaped virions, the capsid precursors being assembled following icosahedral symmetry and the final shape being reached after proteolytical cleavages (Condit et al., 2006), and ascoviruses harbor allantoid capsids (Federici et al., 1990).
Moreover, although giant viruses of amoebae share phenotypic and genotypic features with cellular microorganisms, they were described to lack key cellular hallmarks. A first one consists in proteins involved in the production of energy. This might not be strictly true as tupanviruses harbor genes encoding a putative citrate synthase (Abrahao et al., 2018), and the genome of a distant mimivirus relative (Tetraselmis virus 1) that infects a green alga was shown to harbor key fermentation genes (a pyruvate formate-lyase and a pyruvate formate-lyase activating enzyme) that might ensure energy requirements (Schvarcz and Steward, 2018). A second one consists in ribosomal DNA and proteins, which are absent from giant viruses. Nevertheless, two distinct copies of an 18S rRNA intronic region were recently described in tupanviruses (Abrahao et al., 2018). These sequences were found to be highly expressed, and led to detect similar 18S rRNA intronic region in the majority of other mimivirus genomes. A third cellular hallmark that lacks in giant viruses of amoebae is binary fission as multiplication mechanism.
Conversely, it must be also considered that some bacteria display viral specific features and also lack hallmark features of cellular microorganisms. Numerous bacteria are indeed obligatory intracellular parasites. Moreover, some small cellular microorganisms such as Carsonella ruddii lack a comprehensive ATP generation machinery and, in addition, have a not comprehensive set of ribosomal proteins and aminoacyl-tRNA synthetases (Nakabachi et al., 2006; Tamames et al., 2007). Other cellular microorganisms, such as Chlamydia spp. (Abdelrahman et al., 2016; Bou Khalil et al., 2016), Ehrlichia spp. (Zhang et al., 2007), and Babela sp. (Pagnier et al., 2015) have no bona fide binary fission step during their multiplication. These data highlight that both classical viruses and cellular microorganisms can lack one or several pillar defining features. Finally, while a few viruses, including pandoraviruses, are devoid of capsid (Philippe et al., 2013; Koonin and Dolja, 2014), two classes of icosahedral compartments exist in bacteria and archaea that resemble to viral capsids: they include encapsulin nanocompartments structurally similar to and possibly derived from major capsid proteins of tailed bacterial and archaeal caudaviruses, and microcompartments present in bacteria (including cyanobacteria and many chemotropic bacteria) that encapsulate enzymes involved in metabolic pathways (Tanaka et al., 2008; Krupovic and Koonin, 2017).
Conclusion And Perspectives
Viruses have long been considered as parasitic entities invisible by light microscopy and with a limited repertoire of genes (Raoult and Forterre, 2008). The fact that they are devoid of ribosomal genes has confined them outside of the “tree of life.” Giant viruses of amoebae have undermined this paradigm due to their characteristics that are, at the scale of classical viruses, outstanding (Raoult et al., 2007; Sharma et al., 2016). Phylogenies that were constructed here based on three ancient genes, including RNAP1/2 and DNA polymerase, delineate a fourth TRUC of microbes, as previously reported (Boyer et al., 2010; Sharma et al., 2014, 2015b). Hierarchical clustering performed using a set of informational COGs also shows a fourth independent branch alongside the three cellular branches. Because the tree of proteomes provides a more global and conserved phylogenomic view of protein domain composition in proteomes, their topologies can differ from single-gene based phylogenies that can independently indicate different evolutionary histories. However, here, the four branch topology was maintained in both sequence and structure based trees.
With the recent expansion of the proposed order Megavirales, the number of genes that are shared by these viruses and cellular organisms has shrunk, making it more difficult to build a fourth branch. Nevertheless, among the genes that still show a monophyly are polymerases, which were shown to be among the most ancient protein fold superfamilies (Nasir and Caetano-Anollés, 2015). The ancestrality of conserved genes such as the RNA polymerases, which are suspected to be more ancient than the ribosome (Nasir and Caetano-Anollés, 2015), highlights that evolution can be the result of structural constraints. This concept was described by Gould and Lewontin who used San Marco Cathedral’s spandrels to illustrate that adaptation through selection cannot comprehensively explain the evolution of genomes, and that biological constraints have to be considered (Gould and Lewontin, 1979). The structural, functional and evolutionary units of proteins are the structural domains, highly compact and recurrent segments of the molecules that often combine with others to perform major molecular and cellular tasks (Caetano-Anollés et al., 2009). Domains are evolutionarily highly conserved since they are defined by three-dimensional (3D) structural folds rather than amino acid sequences (Illergard et al., 2009). A rough estimate of evolutionary change suggests that a new fold structure takes millions of years to unfold, while a stable new sequence appears on Earth at least once every microsecond (Caetano-Anollés et al., 2009). In addition, hairpin-forming palindromes, which are possible primordial functional RNAs, are widely distributed among living entities, and they were found to be represented in giant viruses and virophages (Seligmann and Raoult, 2016). Short hairpin structures exist in the genomes of Mimivirus and the Sputnik virophage that may be involved in determining the polyadenylation site of transcripts (Byrne et al., 2009; Claverie and Abergel, 2009). While viral diversification appears fundamentally tailored by reductive evolution, the enrichment of viral genomes with primordial superfamilies of structural domains provides a strong support to the development of the viral proteome core prior to the inception of the ribosome but after the appearance of synthetase-like proteins capable of specific aminoacylation of tRNA molecules (Nasir and Caetano-Anollés, 2015). This could explain the existence of remnants of the translation machinery, the number of which has recently expanded considerably through the isolation of tupanviruses (Abrahao et al., 2018) and the assembly of klosneuvirus genomes (Schulz et al., 2017). As a matter of fact, it is unlikely that there has been a gradual and random acquisition of such large numbers of translation components in giant viruses, such as in mimiviruses, without using it. Hence, this translation machinery might have been acquired in a single step, or, alternatively, might have originated with giant viruses.
The classification of microbes, including the giant viruses, is more realistically based on their genomic content, which reflects their lifestyle, rather than on the phylogenies of supposedly representative genes, which may be confusing because of their mosaicism. This mosaicism results from sequence (and not gene) exchanges occurring during billion years of interactions between emerging lineages or organisms, and is particularly frequent between sympatric microorganisms (Moliner et al., 2010; Raoult and Boyer, 2010). Indeed, microorganisms that encounter and multiply or replicate in same biological niches are particularly prone to exchange nucleic acid sequences. This is well-suggested by the case of Acanthamoeba spp. that can be infected concomitantly by several amoeba-resistant microorganisms including intracellular bacteria and giant viruses with significantly larger repertoires than other related organisms (Moliner et al., 2010; Raoult and Boyer, 2010). Genes evolve by point mutations, but also by fusion, shuffling and fission of genetic fragments, which likely produce gene sequences that are mosaics (Long et al., 1999; Meheust et al., 2018; Pathmanathan et al., 2018). Such chimeric genes have been described in several studies (Ben et al., 2008; Merhej et al., 2011; Meheust et al., 2018), and we found here hints of such gene sequence mosaicism. In addition, many of the genes studied here encode for multi-domain proteins, which makes them mosaics of domains of different ages and histories. The phylogenomic tree reconstructed from domain structures that we describe here disentangles evolutionary histories because each domain becomes a separate phylogenetic character used to build the tree of proteomes. We note however that structural domains and their complex 3D topologies are also built from smaller module-like pieces of arrangements of helix, strand and turn segments (e.g., αα-hairpins, ββ-hairpins, βαβ-motifs) that act as evolutionary building blocks. Recent studies identified combinable (Goncearenco and Berezovsky, 2015) and no-combinable (Alva et al., 2015) ‘loop’ modules of these kinds. In fact, we recently studied the evolutionary combination of loops in domains by generating networks of loops and domains and by tracing their evolution along a timeline of billions of years (Aziz et al., 2016). We uncovered remarkable patterns such as the existence of two functional ‘waves’ of innovation associated with the ‘p-loop’ and ‘winged helix’ general domain structures, the preferential recruitment of ancient loops into new domain structures, and a pervasive network tendency toward hierarchical modularity. Given this difficult ‘mosaic’ problem that affects the sequences of genes and demands phylogenetic dissection, it is interesting to observe here that the tree of proteomes and the trees reconstructed from central genes provided a same overall phylogenetic insight of four TRUCs.
In summary, we highlight here the quantum leap that exists between classical and giant viruses. Our analyses confirm previous evidence of the existence of a fourth TRUC of life that includes viruses, and highlight its ancestrality and mosaicism. Results suggest that best representations for the evolution of giant viruses and cellular microorganisms are rhizomes, and, beyond, that mosaicism has to be considered at the genome (gene content) level but, more generally, at the gene and sequence level. Giant viruses may be represented as comprised by an evolutionary core inferred from highly conserved protein fold structures and gene sequences of very central and ancient proteins, surrounded by a larger and more dynamic gene complement characterized by genome and gene sequence mosaicisms. Such an abductive path as we use, which is based on phenotypic observations, is propitious to provide novel insight on microbial evolution. The “Fourth TRUC” club should, beyond any doubt, continue to expand in the near future, which may be boosted by using new amoebae as co-culture supports and by implementing high-throughput isolation strategies (Khalil et al., 2016). These giant viruses, as new biological entities, should continue to challenge previous paradigms, and a first step is to describe extensively these parasitic microbes without ribosomes.
Author Contributions
DR, PC, PP, GC-A, BLS, and AL designed the experiments. PC, AL, GC-A, and DR wrote the manuscript. PC, AL, VS, AN, and GC-A performed the experiments. All authors analyzed the data and reviewed the manuscript.
Funding
This work was supported by a grant from the French State managed by the National Research Agency under the “Investissements d’Avenir (Investments for the Future)” program with the reference ANR-10-IAHU-03 (Méditerranée Infection) and by the région Provence Alpes Côte d’Azur and European funding FEDER PRIMI. Nisrine Chelkha was financially supported through a grant from the Infectiopole Sud Foundation. Research at Illinois was supported by the USDA National Institute of Food and Agriculture, Hatch project 1014249 and a Blue Waters allocation to GC-A.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.02668/full#supplementary-material
Footnotes
- ^ ftp://ftp.ncbi.nih.gov/genomes/Viruses/
- ^ http://tree.bio.ed.ac.uk/software/figtree/
- ^ http://circos.ca/
References
Abdelrahman, Y., Ouellette, S. P., Belland, R. J., and Cox, J. V. (2016). Polarized cell division of Chlamydia trachomatis. PLoS Pathog. 12:e1005822. doi: 10.1371/journal.ppat.1005822
Abergel, C., Legendre, M., and Claverie, J. M. (2015). The rapidly expanding universe of giant viruses: mimivirus, Pandoravirus, Pithovirus and Mollivirus. FEMS Microbiol. Rev. 39, 779–796. doi: 10.1093/femsre/fuv037
Abrahao, J., Silva, L., Santos Silva, L., Bou Khalil, J. Y., Rodrigues, R., Arantes, T., et al. (2018). Tupanvirus, a tailed giant virus and distant relative of Mimiviridae, possesses the most complete translational apparatus of the virosphere. Nat. Commun. 9:749. doi: 10.1038/s41467-018-03168-1
Altschul, S. F., Gish, W., Miller, W., Myers, E. W., and Lipman, D. J. (1990). Basic local alignment search tool. J. Mol. Biol. 215, 403–410. doi: 10.1016/S0022-2836(05)80360-2
Alva, V., Söding, J., and Lupas, A. N. (2015). A vocabulary of ancient peptides at the origin of folded proteins. eLife 4:e09410. doi: 10.7554/eLife.09410
Andreani, J., Aherfi, S., Bou Khalil, J. Y., Di Pinto, F., Bitam, I., Raoult, D., et al. (2016). Cedratvirus, a double-cork structured giant virus, is a distant relative of pithoviruses. Viruses 8:E300. doi: 10.3390/v8110300
Andreani, J., Khalil, J. Y. B., Baptiste, E., Hasni, I., Michelle, C., Raoult, D., et al. (2018). Orpheovirus IHUMI-LCC2: a new virus among the giant viruses. Front. Microbiol. 8:2643. doi: 10.3389/fmicb.2017.02643
Andreani, J., Khalil, J. Y. B., Sevvana, M., Benamar, S., Di Pinto, F., Bitam, I., et al. (2017). Pacmanvirus, a new giant icosahedral virus at the crossroads between Asfarviridae and faustoviruses. J. Virol. 91:e00212-17. doi: 10.1128/JVI.00212-17
Andreeva, A., Howorth, D., Chandonia, J. M., Brenner, S. E., Hubbard, T. J., Chothia, C. et al. (2008). Data growth and its impact on the SCOP database: new developments. Nucleic Acids Res. 36, D419–D425. doi: 10.1093/nar/gkm993
Aziz, M. F., Caetano-Anollés, K., and Caetano-Anollés, G. (2016). The early history and emergence of molecular functions and modular scale-free network behavior. Sci. Rep. 6:25058. doi: 10.1038/srep25058
Baird, N. L., York, J., and Nunberg, J. H. (2012). Arenavirus infection induces discrete cytosolic structures for RNA replication. J. Virol. 86, 11301–11310. doi: 10.1128/JVI.01635-12
Bajrai, L. H., Benamar, S., Azhar, E. I., Robert, C., Levasseur, A., Raoult, D., et al. (2016). Kaumoebavirus, a new virus that clusters with faustoviruses and Asfarviridae. Viruses 8:E278. doi: 10.3390/v8110278
Ben, S. I., Adekambi, T., Raoult, D., and Drancourt, M. (2008). RpoB sequence-based identification of Mycobacterium avium complex species. Microbiology 154, 3715–3723. doi: 10.1099/mic.0.2008/020164-0
Benamar, S., Reteno, D. G., Bandaly, V., Labas, N., Raoult, D., and La Scola, B. (2016). Faustoviruses: comparative genomics of new Megavirales family members. Front. Microbiol. 7:3. doi: 10.3389/fmicb.2016.00003
Bou Khalil, J. Y., Benamar, S., Baudoin, J. P., Croce, O., Blanc-Tailleur, C., Pagnier, I., et al. (2016). Developmental cycle and genome analysis of “Rubidus massiliensis,” a New Vermamoeba vermiformis Pathogen. Front. Cell. Infect. Microbiol. 6:31. doi: 10.3389/fcimb.2016.00031
Boyer, M., Madoui, M. A., Gimenez, G., La Scola, B., and Raoult, D. (2010). Phylogenetic and phyletic studies of informational genes in genomes highlight existence of a 4 domain of life including giant viruses. PLoS One 5:e15530. doi: 10.1371/journal.pone.0015530
Boyer, M., Yutin, N., Pagnier, I., Barrassi, L., Fournous, G., Espinosa, L., et al. (2009). Giant Marseillevirus highlights the role of amoebae as a melting pot in emergence of chimeric microorganisms. Proc. Natl. Acad. Sci. U.S.A. 106, 21848–21853. doi: 10.1073/pnas.0911354106
Byrne, D., Grzela, R., Lartigue, A., Audic, S., Chenivesse, S., Encinas, S., et al. (2009). The polyadenylation site of Mimivirus transcripts obeys a stringent ‘hairpin rule’. Genome Res. 19, 1233–1242. doi: 10.1101/gr.091561.109
Caetano-Anollés, G., and Caetano-Anollés, D. (2003). An evolutionarily structured universe of protein architecture. Genome Res. 13, 1563–1571. doi: 10.1101/gr.1161903
Caetano-Anollés, G., Kim, K. M., and Caetano-Anollés, D. (2012). The phylogenomic roots of modern biochemistry: origins of proteins, cofactors and protein biosynthesis. J. Mol. Evol. 74, 1–34. doi: 10.1007/s00239-011-9480-1
Caetano-Anollés, G., Wang, M., and Caetano-Anollés, D. (2013). Structural phylogenomics retrodicts the origin of the genetic code and uncovers the evolutionary impact of protein flexibility. PLoS One 8:e72225. doi: 10.1371/journal.pone.0072225
Caetano-Anollés, G., Wang, M., Caetano-Anollés, D., and Mittenthal, J. E. (2009). The origin, evolution and structure of the protein world. Biochem. J. 417, 621–637. doi: 10.1042/BJ20082063
Caetano-Anollés, K., Caetano-Anollés, D., Nasir, A., Kim, K. M., and Caetano-Anollés, G. (2018). Order and polarity in character state transformation models that root the tree of life. Biochimie 149, 135–136. doi: 10.1016/j.biochi.2018.04.001
Claverie, J. M., and Abergel, C. (2009). Mimivirus and its virophage. Annu. Rev. Genet. 43, 49–66. doi: 10.1146/annurev-genet-102108-134255
Colson, P., de Lamballerie, X., Yutin, N., Asgari, S., Bigot, Y., Bideshi, D. K., et al. (2013a). “Megavirales”, a proposed new order for eukaryotic nucleocytoplasmic large DNA viruses. Arch. Virol. 158, 2517–2521. doi: 10.1007/s00705-013-1768-6
Colson, P., Pagnier, I., Yoosuf, N., Fournous, G., La Scola, B., and Raoult, D. (2013b). “Marseilleviridae”, a new family of giant viruses infecting amoebae. Arch. Virol. 158, 915–920. doi: 10.1007/s00705-012-1537-y
Colson, P., La Scola, B., Levasseur, A., Caetano-Anollés, G., and Raoult, D. (2017a). Mimivirus: leading the way in the discovery of giant viruses of amoebae. Nat. Rev. Microbiol. 15, 243–254. doi: 10.1038/nrmicro.2016.197
Colson, P., La Scola, B., and Raoult, D. (2017b). Giant viruses of amoebae: a journey through innovative research and paradigm changes. Annu. Rev. Virol. 4, 61–85. doi: 10.1146/annurev-virology-101416-041816
Condit, R. C., Moussatche, N., and Traktman, P. (2006). In a nutshell: structure and assembly of the vaccinia virion. Adv. Virus Res. 66, 31–124. doi: 10.1016/S0065-3527(06)66002-8
Corradi, N., Pombert, J. F., Farinelli, L., Didier, E. S., and Keeling, P. J. (2010). The complete sequence of the smallest known nuclear genome from the microsporidian Encephalitozoon intestinalis. Nat. Commun. 1:77. doi: 10.1038/ncomms1082
Dagan, T., and Martin, W. (2006). The tree of one percent. Genome Biol. 7:118. doi: 10.1186/gb-2006-7-10-118
Desnues, C., La Scola, B., Yutin, N., Fournous, G., Robert, C., Azza, S., et al. (2012). Provirophages and transpovirons as the diverse mobilome of giant viruses. Proc. Natl. Acad. Sci. U.S.A 109, 18078–18083. doi: 10.1073/pnas.1208835109
Dou, C., Yu, M., Gu, Y., Wang, J., Yin, K., Nie, C., et al. (2018). Structural and mechanistic analyses reveal a unique Cas4-like protein in the mimivirus virophage resistance element system. iScience 3, 1–10. doi: 10.1016/j.isci.2018.04.001
Edgar, R. C. (2004). MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 32, 1792–1797. doi: 10.1093/nar/gkh340
Fabre, E., Jeudy, S., Santini, S., Legendre, M., Trauchessec, M., Coute, Y., et al. (2017). Noumeavirus replication relies on a transient remote control of the host nucleus. Nat. Commun. 8:15087. doi: 10.1038/ncomms15087
Farris, J. S. (1983). “The logical basis of phylogenetic analysis,” in Advances in Cladistics Proceedings of the Second Meeting of the Willi Hennig Society, Vol. 2, eds N. I. Platnick and V. A. Funk (New York, NY: Columbia University Press), 7–36.
Federici, B. A., Vlak, J. M., and Hamm, J. J. (1990). Comparative study of virion structure, protein composition and genomic DNA of three ascovirus isolates. J. Gen. Virol. 71, 1661–1668. doi: 10.1099/0022-1317-71-8-1661
Fischer, M. G., and Suttle, C. A. (2011). A virophage at the origin of large DNA transposons. Science 332, 231–234. doi: 10.1126/science.1199412
Forterre, P., and Gaia, M. (2016). Giant viruses and the origin of modern eukaryotes. Curr. Opin. Microbiol. 31, 44–49. doi: 10.1016/j.mib.2016.02.001
Glass, J. I., Assad-Garcia, N., Alperovich, N., Yooseph, S., Lewis, M. R., Maruf, M., et al. (2006). Essential genes of a minimal bacterium. Proc. Natl. Acad. Sci. U.S.A. 103, 425–430. doi: 10.1073/pnas.0510013103
Goncearenco, A., and Berezovsky, I. N. (2015). Protein function from its emergence to diversity in contemporary proteins. Phys. Biol. 12:045002. doi: 10.1088/1478-3975/12/4/045002
Gorlas, A., Robert, C., Gimenez, G., Drancourt, M., and Raoult, D. (2012). Complete genome sequence of Methanomassiliicoccus luminyensis, the largest genome of a human-associated Archaea species. J. Bacteriol. 194:4745. doi: 10.1128/JB.00956-12
Gough, J., and Chothia, C. (2002). SUPERFAMILY: HMMs representing all proteins of known structure. SCOP sequence searches, alignments and genome assignments. Nucleic Acids Res. 30, 268–272. doi: 10.1093/nar/30.1.268
Gough, J., Karplus, K., Hughey, R., and Chothia, C. (2001). Assignment of homology to genome sequences using a library of hidden Markov models that represent all proteins of known structure. J. Mol. Biol. 313, 903–919. doi: 10.1006/jmbi.2001.5080
Gould, S. J., and Lewontin, R. C. (1979). The spandrels of San Marco and the Panglossian paradigm: a critique of the adaptationist programme. Proc. R. Soc. Lond. B Biol. Sci. 205, 581–598. doi: 10.1098/rspb.1979.0086
Harish, A., and Caetano-Anollés, G. (2012). Ribosomal history reveals origins of modern protein synthesis. PLoS One 7:e32776. doi: 10.1371/journal.pone.0032776
Hedges, S. B., Dudley, J., and Kumar, S. (2006). TimeTree: a public knowledge-base of divergence times among organisms. Bioinformatics 22, 2971–2972. doi: 10.1093/bioinformatics/btl505
Illergard, K., Ardell, D. H., and Elofsson, A. (2009). Structure is three to ten times more conserved than sequence–a study of structural response in protein cores. Proteins 77, 499–508. doi: 10.1002/prot.22458
Iyer, L. M., Aravind, L., and Koonin, E. V. (2001). Common origin of four diverse families of large eukaryotic DNA viruses. J. Virol. 75, 11720–11734. doi: 10.1128/JVI.75.23.11720-11734.2001
Iyer, L. M., Balaji, S., Koonin, E. V., and Aravind, L. (2006). Evolutionary genomics of nucleo-cytoplasmic large DNA viruses. Virus Res. 117, 156–184. doi: 10.1016/j.virusres.2006.01.009
Kawai, H., Kanegae, T., Christensen, S., Kiyosue, T., Sato, Y., Imaizumi, T., et al. (2003). Responses of ferns to red light are mediated by an unconventional photoreceptor. Nature 421, 287–290. doi: 10.1038/nature01310
Khalil, J. Y., Robert, S., Reteno, D. G., Andreani, J., Raoult, D., and La Scola, B. (2016). High-throughput isolation of giant viruses in liquid medium using automated flow cytometry and fluorescence staining. Front. Microbiol. 7:26. doi: 10.3389/fmicb.2016.00026
Koonin, E. V., and Dolja, V. V. (2014). Virus world as an evolutionary network of viruses and capsidless selfish elements. Microbiol. Mol. Biol. Rev. 78, 278–303. doi: 10.1128/MMBR.00049-13
Koonin, E. V., Senkevich, T. G., and Dolja, V. V. (2006). The ancient virus world and evolution of cells. Biol. Direct 1:29. doi: 10.1186/1745-6150-1-29
Koonin, E. V., and Yutin, N. (2010). Origin and evolution of eukaryotic large nucleo-cytoplasmic DNA viruses. Intervirology 53, 284–292. doi: 10.1159/000312913
Krupovic, M., and Koonin, E. V. (2017). Cellular origin of the viral capsid-like bacterial microcompartments. Biol. Direct 12:25. doi: 10.1186/s13062-017-0197-y
La Scola, B., Audic, S., Robert, C., Jungang, L., de Lamballerie, X., Drancourt, M., et al. (2003). A giant virus in amoebae. Science 299:2033. doi: 10.1126/science.1081867
La Scola, B., de Lamballerie, X. N., Claverie, J. M., Drancourt, M., and Raoult, D. (2005). “Genus Mimivirus,” in Virus Taxonomy, eds M. Fauquet, M. A. Mayo, J. Maniloff, U. Desselberger, and L. A. Ball (San Diego: Elsevier Academic Press), 275–276.
La Scola, B., Desnues, C., Pagnier, I., Robert, C., Barrassi, L., Fournous, G., et al. (2008). The virophage as a unique parasite of the giant mimivirus. Nature 455, 100–104. doi: 10.1038/nature07218
Lartillot, N., Brinkmann, H., and Philippe, H. (2007). Suppression of long-branch attraction artefacts in the animal phylogeny using a site-heterogeneous model. BMC Evol. Biol. 7(Suppl. 1):S4. doi: 10.1186/1471-2148-7-S1-S
Lechner, M., Findeiss, S., Steiner, L., Marz, M., Stadler, P. F., and Prohaska, S. J. (2011). Proteinortho: detection of (co-)orthologs in large-scale analysis. BMC Bioinformatics 12:124. doi: 10.1186/1471-2105-12-124
Legendre, M., Arslan, D., Abergel, C., and Claverie, J. M. (2012). Genomics of Megavirus and the elusive fourth domain of Life. Commun. Integr. Biol. 5, 102–106. doi: 10.4161/cib.18624
Legendre, M., Bartoli, J., Shmakova, L., Jeudy, S., Labadie, K., Adrait, A., et al. (2014). Thirty-thousand-year-old distant relative of giant icosahedral DNA viruses with a pandoravirus morphology. Proc. Natl. Acad. Sci. U.S.A. 111, 4274–4279. doi: 10.1073/pnas.1320670111
Legendre, M., Lartigue, A., Bertaux, L., Jeudy, S., Bartoli, J., Lescot, M., et al. (2015). In-depth study of Mollivirus sibericum, a new 30,000-y-old giant virus infecting Acanthamoeba. Proc. Natl. Acad. Sci. U.S.A. 112, E5327–E5335. doi: 10.1073/pnas.1510795112
Levasseur, A., Andreani, J., Delerce, J., Bou Khalil, J., Catherine, R., La Scola, B., et al. (2016a). Comparison of a modern and fossil pithovirus reveals its genetic conservation and evolution. Genome Biol. Evol. 8, 2333–2339. doi: 10.1093/gbe/evw153
Levasseur, A., Bekliz, M., Chabriere, E., Pontarotti, P., La Scola, B., and Raoult, D. (2016b). MIMIVIRE is a defence system in mimivirus that confers resistance to virophage. Nature 531, 249–252. doi: 10.1038/nature17146
Levasseur, A., Merhej, V., Baptiste, E., Sharma, V., Pontarotti, P., and Raoult, D. (2017). The Rhizome of Lokiarchaeota illustrates the mosaicity of archaeal genomes. Genome Biol. Evol. 9, 2635–2639. doi: 10.1093/gbe/evx208
Li, L., Stoeckert, C. J. Jr., and Roos, D. S. (2003). OrthoMCL: identification of ortholog groups for eukaryotic genomes. Genome Res. 13, 2178–2189. doi: 10.1101/gr.1224503
Long, M., Wang, W., and Zhang, J. (1999). Origin of new genes and source for N-terminal domain of the chimerical gene, jingwei, in Drosophila. Gene 238, 135–141. doi: 10.1016/S0378-1119(99)00229-2
Lundberg, J. G. (1972). Wagner networks and ancestors. Syst. Zool. 21, 398–413. doi: 10.1093/sysbio/21.4.398
Lwoff, A. (1957). The concept of virus. J. Gen. Microbiol. 17, 239–253. doi: 10.1099/00221287-17-2-239
Meheust, R., Bhattacharya, D., Pathmanathan, J. S., McInerney, J. O., Lopez, P., and Bapteste, E. (2018). Formation of chimeric genes with essential functions at the origin of eukaryotes. BMC Biol. 16:30. doi: 10.1186/s12915-018-0500-0
Merhej, V., Notredame, C., Royer-Carenzi, M., Pontarotti, P., and Raoult, D. (2011). The rhizome of life: the sympatric Rickettsia felis paradigm demonstrates the random transfer of DNA sequences. Mol. Biol. Evol. 28, 3213–3223. doi: 10.1093/molbev/msr239
Moliner, C., Fournier, P. E., and Raoult, D. (2010). Genome analysis of microorganisms living in amoebae reveals a melting pot of evolution. FEMS Microbiol. Rev. 34, 281–294. doi: 10.1111/j.1574-6976.2010.00209.x
Moreira, D., and Lopez-Garcia, P. (2009). Ten reasons to exclude viruses from the tree of life. Nat. Rev. Microbiol. 7, 306–311. doi: 10.1038/nrmicro2108
Moreira, D., and Lopez-Garcia, P. (2015). Evolution of viruses and cells: do we need a fourth domain of life to explain the origin of eukaryotes? Philos. Trans. R. Soc. Lond. B Biol. Sci. 370:20140327. doi: 10.1098/rstb.2014.0327
Nakabachi, A., Yamashita, A., Toh, H., Ishikawa, H., Dunbar, H. E., Moran, N. A., et al. (2006). The 160-kilobase genome of the bacterial endosymbiont Carsonella. Science 314:267. doi: 10.1126/science.1134196
Nasir, A., and Caetano-Anollés, G. (2015). A phylogenomic data-driven exploration of viral origins and evolution. Sci. Adv. 1:e1500527. doi: 10.1126/sciadv.1500527
Nasir, A., Kim, K. M., and Caetano-Anollés, G. (2012). Giant viruses coexisted with the cellular ancestors and represent a distinct supergroup along with superkingdoms Archaea, Bacteria and Eukarya. BMC Evol. Biol. 12:156. doi: 10.1186/1471-2148-12-156
Nasir, A., Kim, K. M., and Caetano-Anollés, G. (2017). Phylogenetic tracings of proteome size support the gradual accretion of protein structural domains and the early origin of viruses from primordial cells. Front. Microbiol. 8:1178. doi: 10.3389/fmicb.2017.01178
Nasir, A., Sun, F. J., Kim, K. M., and Caetano-Anollés, G. (2015). Untangling the origin of viruses and their impact on cellular evolution. Ann. N. Y. Acad. Sci. 1341, 61–74. doi: 10.1111/nyas.12735
Netherton, C. L., and Wileman, T. (2011). Virus factories, double membrane vesicles and viroplasm generated in animal cells. Curr. Opin. Virol. 1, 381–387. doi: 10.1016/j.coviro.2011.09.008
Nissimov, J. I., Pagarete, A., Ma, F., Cody, S., Dunigan, D. D., Kimmance, S. A., et al. (2017). Coccolithoviruses: a review of cross-kingdom genomic thievery and metabolic thuggery. Viruses 9:E52. doi: 10.3390/v9030052
Ogata, H., La Scola, B., Audic, S., Renesto, P., Blanc, G., Robert, C., et al. (2006). Genome sequence of Rickettsia bellii illuminates the role of amoebae in gene exchanges between intracellular pathogens. PLoS Genet. 2:e76. doi: 10.1371/journal.pgen.0020076
Pagnier, I., Yutin, N., Croce, O., Makarova, K. S., Wolf, Y. I., Benamar, S., et al. (2015). Babela massiliensis, a representative of a widespread bacterial phylum with unusual adaptations to parasitism in amoebae. Biol. Direct 10:13. doi: 10.1186/s13062-015-0043-z
Pathmanathan, J. S., Lopez, P., Lapointe, F. J., and Bapteste, E. (2018). Composite Search: a generalized network approach for composite gene families detection. Mol. Biol. Evol. 35, 252–255. doi: 10.1093/molbev/msx283
Philippe, N., Legendre, M., Doutre, G., Coute, Y., Poirot, O., Lescot, M., et al. (2013). Pandoraviruses: amoeba viruses with genomes up to 2.5 Mb reaching that of parasitic eukaryotes. Science 341, 281–286. doi: 10.1126/science.1239181
Price, M. N., Dehal, P. S., and Arkin, A. P. (2010). FastTree 2–approximately maximum-likelihood trees for large alignments. PLoS One 5:e9490. doi: 10.1371/journal.pone.0009490
Raoult, D. (2010). The post-Darwinist rhizome of life. Lancet 375, 104–105. doi: 10.1016/S0140-6736(09)61958-9
Raoult, D. (2013). TRUC or the need for a new microbial classification. Intervirology 56, 349–353. doi: 10.1159/000354269
Raoult, D. (2014). How the virophage compels the need to readdress the classification of microbes. Virology 477, 119–124. doi: 10.1016/j.virol.2014.11.014
Raoult, D., Audic, S., Robert, C., Abergel, C., Renesto, P., Ogata, H., et al. (2004). The 1.2-megabase genome sequence of Mimivirus. Science 306, 1344–1350. doi: 10.1126/science.1101485
Raoult, D., and Boyer, M. (2010). Amoebae as genitors and reservoirs of giant viruses. Intervirology 53, 321–329. doi: 10.1159/000312917
Raoult, D., and Forterre, P. (2008). Redefining viruses: lessons from Mimivirus. Nat. Rev. Microbiol. 6, 315–319. doi: 10.1038/nrmicro1858
Raoult, D., La Scola, B., and Birtles, R. (2007). The discovery and characterization of Mimivirus, the largest known virus and putative pneumonia agent. Clin. Infect. Dis. 45, 95–102. doi: 10.1086/518608
Reteno, D. G., Benamar, S., Khalil, J. B., Andreani, J., Armstrong, N., Klose, T., et al. (2015). Faustovirus, an asfarvirus-related new lineage of giant viruses infecting amoebae. J. Virol. 89, 6585–6594. doi: 10.1128/JVI.00115-15
Schulz, F., Yutin, N., Ivanova, N. N., Ortega, D. R., Lee, T. K., Vierheilig, J., et al. (2017). Giant viruses with an expanded complement of translation system components. Science 356, 82–85. doi: 10.1126/science.aal4657
Schvarcz, C. R., and Steward, G. F. (2018). A giant virus infecting green algae encodes key fermentation genes. Virology 518, 423–433. doi: 10.1016/j.virol.2018.03.010
Seligmann, H., and Raoult, D. (2016). Unifying view of stem-loop hairpin RNA as origin of current and ancient parasitic and non-parasitic RNAs, including in giant viruses. Curr. Opin. Microbiol. 31, 1–8. doi: 10.1016/j.mib.2015.11.004
Sharma, V., Colson, P., Chabrol, O., Pontarotti, P., and Raoult, D. (2015a). Pithovirus sibericum, a new bona fide member of the “Fourth TRUC” club. Front. Microbiol. 6:722. doi: 10.3389/fmicb.2015.00722
Sharma, V., Colson, P., Chabrol, O., Scheid, P., Pontarotti, P., and Raoult, D. (2015b). Welcome to pandoraviruses at the ‘Fourth TRUC’ club. Front. Microbiol. 6:423. doi: 10.3389/fmicb.2015.00423
Sharma, V., Colson, P., Giorgi, R., Pontarotti, P., and Raoult, D. (2014). DNA-dependent RNA polymerase detects hidden giant viruses in published databanks. Genome Biol. Evol. 6, 1603–1610. doi: 10.1093/gbe/evu128
Sharma, V., Colson, P., Pontarotti, P., and Raoult, D. (2016). Mimivirus inaugurated in the 21st century the beginning of a reclassification of viruses. Curr. Opin. Microbiol. 31, 16–24. doi: 10.1016/j.mib.2015.12.010
Tamames, J., Gil, R., Latorre, A., Pereto, J., Silva, F. J., and Moya, A. (2007). The frontier between cell and organelle: genome analysis of Candidatus Carsonella ruddii. BMC Evol. Biol. 7:181. doi: 10.1186/1471-2148-7-181
Tanaka, S., Kerfeld, C. A., Sawaya, M. R., Cai, F., Heinhorst, S., Cannon, G. C., et al. (2008). Atomic-level models of the bacterial carboxysome shell. Science 319, 1083–1086. doi: 10.1126/science.1151458
Terhune, S. S., Schroer, J., and Shenk, T. (2004). RNAs are packaged into human cytomegalovirus virions in proportion to their intracellular concentration. J. Virol. 78, 10390–10398. doi: 10.1128/JVI.78.19.10390-10398.2004
Wang, M., Jiang, Y. Y., Kim, K. M., Qu, G., Ji, H. F., Mittenthal, J. E., et al. (2011). A universal molecular clock of protein folds and its power in tracing the early history of aerobic metabolism and planet oxygenation. Mol. Biol. Evol. 28, 567–582. doi: 10.1093/molbev/msq232
Williams, T. A., Embley, T. M., and Heinz, E. (2011). Informational gene phylogenies do not support a fourth domain of life for nucleocytoplasmic large DNA viruses. PLoS One 6:e21080. doi: 10.1371/journal.pone.0021080
Wilson, W. H., Van Etten, J. L., and Allen, M. J. (2009). The Phycodnaviridae: the story of how tiny giants rule the world. Curr. Top. Microbiol. Immunol. 328, 1–42. doi: 10.1007/978-3-540-68618-7_1
Wu, D., Wu, M., Halpern, A., Rusch, D. B., Yooseph, S., Frazier, M., et al. (2011). Stalking the fourth domain in metagenomic data: searching for, discovering, and interpreting novel, deep branches in marker gene phylogenetic trees. PLoS One 6:e18011. doi: 10.1371/journal.pone.0018011
Yutin, N., and Koonin, E. V. (2012). Hidden evolutionary complexity of nucleo-cytoplasmic large DNA viruses of eukaryotes. Virol. J. 9:161. doi: 10.1186/1743-422X-9-161
Yutin, N., and Koonin, E. V. (2013). Pandoraviruses are highly derived phycodnaviruses. Biol. Direct 8, 25–28. doi: 10.1186/1745-6150-8-25
Yutin, N., Wolf, Y. I., and Koonin, E. V. (2014). Origin of giant viruses from smaller DNA viruses not from a fourth domain of cellular life. Virology 46, 38–52. doi: 10.1016/j.virol.2014.06.032
Yutin, N., Wolf, Y. I., Raoult, D., and Koonin, E. V. (2009). Eukaryotic large nucleo-cytoplasmic DNA viruses: clusters of orthologous genes and reconstruction of viral genome evolution. Virol. J. 17:223. doi: 10.1186/1743-422X-6-223
Keywords: giant virus, TRUC, megavirales, mimivirus, informational genes, protein structural domains
Citation: Colson P, Levasseur A, La Scola B, Sharma V, Nasir A, Pontarotti P, Caetano-Anollés G and Raoult D (2018) Ancestrality and Mosaicism of Giant Viruses Supporting the Definition of the Fourth TRUC of Microbes. Front. Microbiol. 9:2668. doi: 10.3389/fmicb.2018.02668
Received: 22 May 2018; Accepted: 18 October 2018;
Published: 27 November 2018.
Edited by:
Steven M. Short, University of Toronto Mississauga, CanadaReviewed by:
Jessica Labonté, Texas A&M University at Galveston, United StatesJozef I. Nissimov, Rutgers, The State University of New Jersey, United States
David Robert Wessner, Davidson College, United States
Copyright © 2018 Colson, Levasseur, La Scola, Sharma, Nasir, Pontarotti, Caetano-Anollés and Raoult. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Didier Raoult, ZGlkaWVyLnJhb3VsdEBnbWFpbC5jb20=