
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 11 October 2018
Sec. Aquatic Microbiology
Volume 9 - 2018 | https://doi.org/10.3389/fmicb.2018.02443
 Alain Rincé1*
Alain Rincé1* Charlotte Balière2
Charlotte Balière2 Dominique Hervio-Heath2
Dominique Hervio-Heath2 Joëlle Cozien2Solen Lozach2Sylvain Parnaudeau2Françoise S. Le Guyader2Simon Le Hello3
Joëlle Cozien2Solen Lozach2Sylvain Parnaudeau2Françoise S. Le Guyader2Simon Le Hello3 Jean-Christophe Giard1Nicolas Sauvageot1
Jean-Christophe Giard1Nicolas Sauvageot1 Abdellah Benachour1
Abdellah Benachour1 Sofia Strubbia3
Sofia Strubbia3 Michèle Gourmelon2
Michèle Gourmelon2During a 2-year study, the presence of human pathogenic bacteria and noroviruses was investigated in shellfish, seawater and/or surface sediments collected from three French coastal shellfish-harvesting areas as well as in freshwaters from the corresponding upstream catchments. Bacteria isolated from these samples were further analyzed. Escherichia coli isolates classified into the phylogenetic groups B2, or D and enterococci from Enterococcus faecalis and E. faecium species were tested for the presence of virulence genes and for antimicrobial susceptibility. Salmonella members were serotyped and the most abundant serovars (Typhimurium and its monophasic variants and Mbandaka) were genetically characterized by high discriminative subtyping methods. Campylobacter and Vibrio were identified at the species level, and haemolysin-producing Vibrio parahaemolyticus were searched by tdh- and trh- gene detection. Main results showed a low prevalence of Salmonella in shellfish samples where only members of S. Mbandaka were found. Campylobacter were more frequently isolated than Salmonella and a different distribution of Campylobacter species was observed in shellfish compared to rivers, strongly suggesting possible additional inputs of bacteria. Statistical associations between enteric bacteria, human noroviruses (HuNoVs) and concentration of fecal indicator bacteria revealed that the presence of Salmonella was correlated with that of Campylobacter jejuni and/or C. coli as well as to E. coli concentration. A positive correlation was also found between the presence of C. lari and the detection of HuNoVs. This study highlights the importance of simultaneous detection and characterization of enteric and marine pathogenic bacteria and human noroviruses not only in shellfish but also in catchment waters for a hazard assessment associated with microbial contamination of shellfish.
Shellfish and coastal waters contaminated by human pathogens could be sources of shellfish-borne or water-borne outbreaks (Potasman et al., 2002; Yoder et al., 2008). In fact, shellfish can accumulate and concentrate microbial pathogens present in waters by their filter-feeding activities. Because shellfish are often consumed raw or lightly cooked, they present a health risk for exposure to bacterial and viral pathogens. Indeed, crustaceans, shellfish, mollusks and products thereof constitute food vehicles in 7.3% of the strong-evidence outbreaks in the European Union (EU) in 2013 (European Food Safety Authority [EFSA] and European Centre for Disease Prevention Control [ECDC], 2015). The microbiological quality of shellfish-harvesting areas and bathing waters is assessed by enumeration of the fecal indicator bacteria (FIBs) Escherichia coli and enterococci. Their presence can lead to closures or downgrading of shellfish-harvesting areas and bathing areas. These FIBs inform of the possible presence of fecal microorganisms potentially pathogenic for humans such as members of E. coli species or Enterococcus genus, zoonotic bacteria such as Salmonella spp. and Campylobacter spp. which could originate from urban and agricultural catchments upstream coastal bathing waters and shellfish-harvesting areas. However, these indicators do not take into account the risk associated with the presence of marine bacteria such as Vibrio or HuNoVs which are also responsible for outbreaks from shellfish origin (DePaola et al., 2010; Amagliani et al., 2012; Yu et al., 2015).
Numerous bacterial human pathogens are present in coastal waters and shellfish (Escobedo-Hinojosa and Pardo-López, 2017; Leight et al., 2018). They could be classified as allochthonous bacteria, coming from upstream catchments or as autochthonous such as Vibrio. Among allochthonous bacteria, some are of fecal origin such as members of Enterobacteriaceae (e.g., pathogenic E. coli, Salmonella), pathogenic enterococci, Campylobacter and others are from aquatic environment and soil such as Aeromonas, Arcobacter, and Pseudomonas.
A selection of these pathogens was investigated in this study, starting with pathogenic bacteria among the FIB. In fact, even commensals of the intestinal microflora of humans and animals, some strains of E. coli such that producing Shiga-toxin (STEC) or enteropathogenic E. coli (EPEC) are able to cause pathologies that could potentially lead to hemolytic uremic syndrome. Members of E. coli are also responsible for extra-intestinal pathologies including urinary tract infections, meningitis or septicemia (Kaper et al., 2004). E. coli can easily be divided into four main phylogenetic groups (A, B1, B2, and D) (Clermont et al., 2000). Extraintestinal pathogenic E. coli (ExPEC) mainly belong to groups B2 and D whereas most commensal strains belong to groups A and B1 (Picard et al., 1999; Johnson et al., 2001). Since the last decades, enterococci have become nosocomial pathogens of global importance where Enterococcus faecium and E. faecalis are clinically the most feared species (Guzman Prieto et al., 2016). Moreover, E. coli and enterococci may contribute to the dissemination of antibiotic resistance genes (Boehm and Sassoubre, 2014; Andrade et al., 2015).
Among fecal pathogens, we also focus on bacteria mainly coming from livestock animals upstream coastal areas such as non-typhoidal Salmonella and Campylobacter spp. These bacteria are leading causes of bacterial gastroenteritis in many countries. With norovirus, they accounted for more than 70% of food-borne pathogens-associated illnesses and hospitalizations in France (Van Cauteren et al., 2017). Salmonellosis can be due to numerous serovars of Salmonella enterica subspecies enterica but few of them (such as Typhimurium and its monophasic variants or Enteritidis) are cause of most human infections (European Food Safety Authority [EFSA] and European Centre for Disease Prevention Control [ECDC], 2016). Salmonella spp. are present in poultry, cattle, pigs, and wild birds and also isolated from fresh and marine waters (Gorski et al., 2011; Walters et al., 2013; Flockhart et al., 2017). While S. Typhimurium is ubiquitous, S. Dublin and S. Enteritidis are more commonly found in cattle and in poultry, respectively (David et al., 2013; European Food Safety Authority [EFSA] and European Centre for Disease Prevention Control [ECDC], 2016). With over 229,000 human cases a year, campylobacteriosis is the most frequently reported food-borne illness in EU mainly due to Campylobacter jejuni followed, by far by C. coli, whereas C. lari is implicated in a lesser extend (European Food Safety Authority [EFSA] and European Centre for Disease Prevention Control [ECDC], 2016). Animals are the main reservoirs of Campylobacter. More specifically, C. jejuni, C. coli, and C. lari are mainly found in poultry and cattle, in poultry and swine, and in gulls and shorebirds, respectively (Denis et al., 2011a; Miller et al., 2014; Pitkanen and Hanninen, 2017). Campylobacter has been frequently detected in waters at the level of catchments and in coastal areas (Martinez-Urtaza et al., 2004; Van Dyke et al., 2010; Wilkes et al., 2011). Few shellfish outbreaks due to Campylobacter have been reported (Abeyta et al., 1993).
Among the allochthonous bacterial pathogens, Vibrio spp. occurring naturally in brackish and marine waters, Vibrio parahaemolyticus, V. vulnificus, and V. cholerae were selected for this study as they are the main species involved in seafood- and seawater-borne illness worldwide1. These Vibrio species are commonly isolated in coastal waters from France and Europe and the risk of further Vibrio infections might increase in the coming decades due to climate change (Hervio-Heath et al., 2002; Martinez-Urtaza et al., 2012; Esteves et al., 2015; Semenza et al., 2017).
Finally, in addition to human bacterial pathogens, enteric viruses such as HuNoVs, were also selected as they are the main cause of gastroenteritis worldwide. HuNoVs are a group of highly diverse viruses that belong to the Caliciviridae family. They are non-enveloped icosahedral viruses with a single stranded RNA genome. HuNoVs cause gastroenteritis characterized by vomiting, abdominal cramps, fever, watery diarrhea, headaches, chills and myalgia, and illness normally lasts 2–3 days. They are currently classified in seven genogroups, of which three infect humans (GI, GII, and GIV) (de Graaf et al., 2016). HuNoV is one of the most infectious pathogens, as demonstrated through human volunteer studies, or analysis of data from shellfish-related outbreaks (Thebault et al., 2013; Atmar et al., 2014). GI and GII HuNoVs are excreted at very high levels in the feces (up to 1011 copies/g) of both symptomatic and asymptomatic people during long periods (at least 27 days) (Atmar et al., 2014; Miura et al., 2017).
The aim of the present study was to detect and characterize microorganisms from shellfish-harvesting areas and their catchments in order to assess the prevalence and diversity of selected pathogenic bacteria (Salmonella, Campylobacter, and Vibrio), FIBs and HuNoVs. For this purpose, samples of shellfish batches, freshwater, seawater and surface sediments collected monthly over a 2-year period in three selected shellfish-harvesting areas and their catchments were analyzed. This study presents the interest of focusing on the simultaneous detection and/or isolation of pathogenic enteric and marine bacteria, and HuNoVs in coastal environments (shellfish and waters from the catchment).
A total of 505 samples including 237 shellfish batches, 40 surface sediments, 12 seawaters and 216 waters from nine sampling points upstream of shellfish-harvesting areas described in Balière et al. (2015) were analyzed. These samples came from three shellfish-harvesting sites located on the English Channel coast and in their catchments. On the catchment of the Brittany site (site 1, La Fresnaye), four rivers flow with their outlet in the bay: Frémur (1A, 50 km long, sub-catchment: 77.4 km2), Le Rat (1B, 10.5 km, sub-catchment: 19.2 km2), Le Clos (1C, 7 km, sub-catchment: 13.3 km2) and Kermiton (1D, 2.3 km, sub-catchment: 6.3 km2) (Figure 1). Cattle, swine and poultry are intensively bred on this catchment (Supplementary Data S1). The Normandy site 2 (Regnéville sur mer) is characterized by the intensive breeding of cattle and sheep and by the presence of poultry and swine, while its neighboring site 3 (La Vanlée) rather hosts cattle, sheep, and swine (Supplementary Data S1). The main rivers on these sites are La Sienne (2A, 93 km, sub-catchment: 794 km2) and La Soules (2B, 53 km, sub-catchment: 150 km2) on site 2, and La Vanlée (3A, 8.5 km, sub-catchment: 35 km2) and Les Hardes (3B, 6 km, sub-catchment: 15 km2) on site 3.
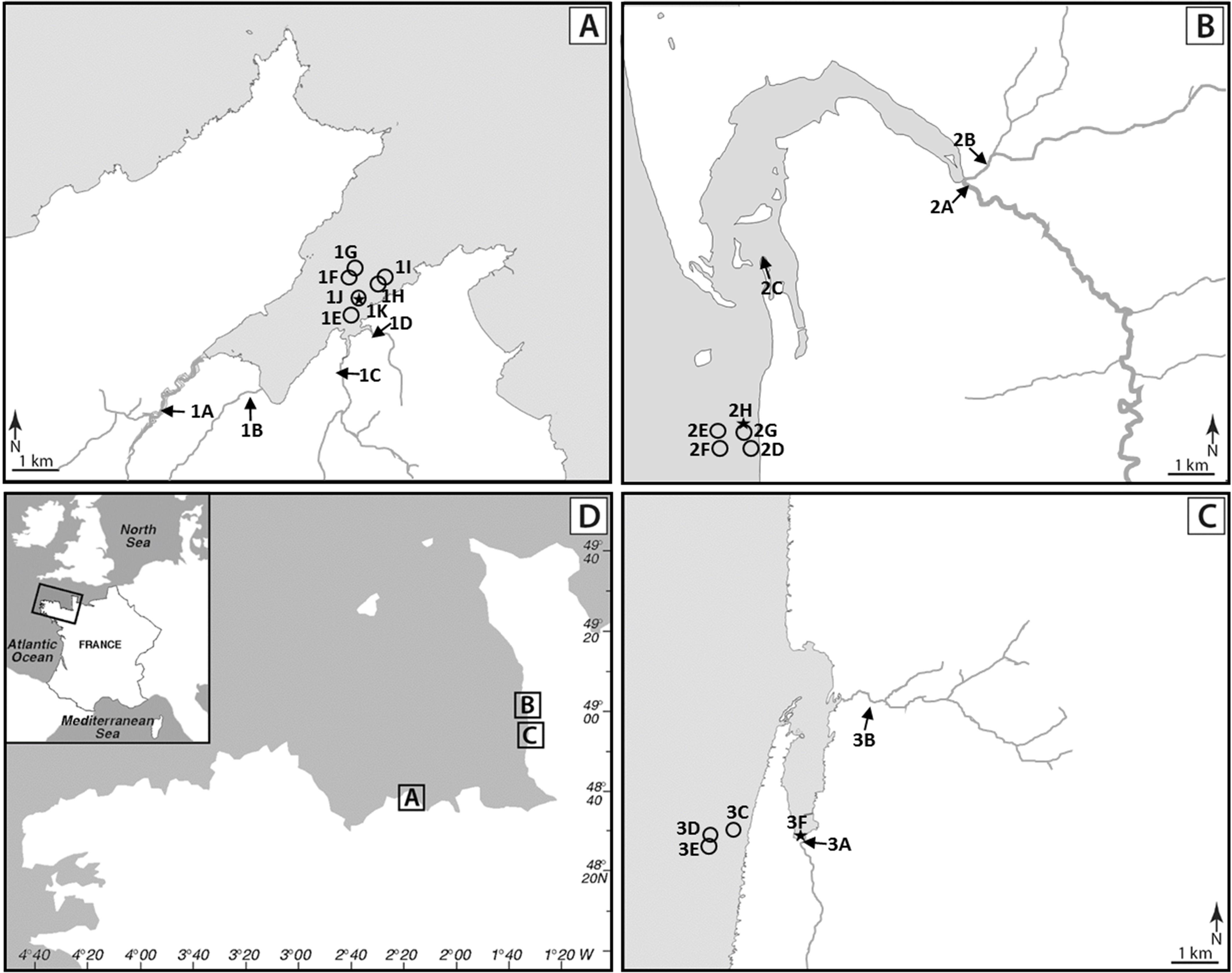
FIGURE 1. Location of sampling sites. (A) La Fresnaye (Brittany) site 1. (B) Regnéville sur mer (Normandy) site 2. (C) La Vanlée (Normandy) site 3. (D) Location of studied sites on a map of France. Sampling sites are represented by arrows (water from river and brackish water), circles (shellfish and seawater) and stars (sediment).
Shellfish [oyster, mussel, and common cockle batches (site 1, n = 120; site 2, n = 71 and site 3, n = 46)] and waters from sampling points upstream of shellfish-harvesting areas (site 1, n = 96; site 2, n = 72; site 3, n = 48) were collected monthly from February 2013 to January 2015, whereas surface sediment samples (site 1, n = 14; site 2, n = 13; site 3, n = 13) were collected from February 2013 to January 2014 and seawater samples (site 1, n = 12) from February 2014 to January 2015 (Balière et al., 2015). If all the samples taken from the rivers Frémur, Le Rat, Le Clos, La Soules, and Les Hardes had a salinity of less than 1 g/L, one from the mouth of the river La Sienne and several samples from the estuary of Regnéville sur mer and Kermiton presented a higher salinity.
Water samples were analyzed by membrane filtration and shellfish were opened and dissected aseptically for analyses. After opening, 25 g of homogenized blended total shellfish flesh (SF; including shellfish flesh and intravalvular liquid) were introduced into 225 ml of the appropriate enrichment buffer according to the targeted bacteria. For sediment, 10 g were introduced into the appropriate buffer, while for water samples 1 L was filtered using the appropriate cellulose membranes (0.45 μm for Salmonella and E. coli and 0.2 μm for Campylobacter and Vibrio) and the filters were placed in the appropriate enrichment buffer.
Escherichia coli and enterococci from water samples were enumerated by ISO-9308-3:1998 (1998) and ISO-7899-1:1998 (1998) methods, respectively, and E. coli from shellfish batches by impedance method according to AFNOR-NF-V-08-106:2010 (2010). In addition, E. coli and enterococci were isolated as described in Supplementary Data S2. DNAs were extracted using Instagene matrix (Bio-Rad) after mechanical lysis of the bacteria with glass micro-beads.
Escherichia coli isolates were classified into the four main phylogenetic groups A, B1, B2, or D using a method based on the detection of two genes, chuA and yjaA and of a DNA fragment designated TSPE4.C2 (Clermont et al., 2000). Detection of these elements was performed by quantitative PCR (qPCR) using SYBER Green precision melt supermix (Bio-Rad) and primers described in Supplementary Data S3. Detection of stx and eae genes, encoding Shiga-toxin and intimin, respectively, and isolation of strains belonging to the STEC and EPEC pathotypes which are frequently observed as being responsible for diarrhea were the subject of a previous study (Balière et al., 2015). Thus, we focused here to detection of virulence genes characteristic of ExPEC. The presence of genes which encode fimbriae (papC and sfa), adhesins (papGII and papGIII), hemolysin (hlyC), cytotoxic necrotizing factor (cnf1), siderophore synthesis (iucC) and siderophore receptors (fyuA and iroN) was investigated by PCR using primers and protocol previously described by Bonacorsi et al. (2006) and GoTaq Flexi DNA polymerase (Promega).
Presence of virulence genes in enterococci [i.e., esp, gelE, and agg encoding the extracellular protein Esp (Shankar et al., 1999), an extracellular zinc metallo-endopeptidase (Su et al., 1991), and an aggregation substance (Kreft et al., 1992), respectively] was determined by multiplex qPCR using SYBER Green precision melt supermix and primers described in Supplementary Data S3.
Antimicrobial susceptibility testing of a selection of 556 E. coli and 446 enterococci (213 E. faecalis and 233 E. faecium) isolates was based on the disk diffusion method on Muller-Hinton medium (AES-chemunex, Bruz, France). Plates were incubated at 37°C for 24 h.
Salmonella spp. were detected and isolated as described in the Supplementary Data S2.
Serotyping of Salmonella strains was performed by agglutination tests with antisera (Bio-Rad, Marnes-la-Coquette, France) according to the White–Kauffmann–Le Minor scheme (Grimont and Weill, 2007).
Depending on the serotypes, different subtyping methods were performed in order to get suitable and discriminative information on the main Salmonella populations that were circulating. For all Salmonella Typhimurium isolates (as well as their monophasic or non-motile variants), the CRISPR (Clustered Regularly Interspaced Short Palindromic Repeats) spacer content was recognized by a high-throughput method (named CRISPOL), as previously described (Fabre et al., 2012). DNA macrorestriction, i.e., Pulsenet standard XbaI-PFGE protocol (Ribot et al., 2006), and/or whole genome sequencing (WGS) were performed for Salmonella Mbandaka isolates. Sequencing and phylogenetic analyzes were done as previously described by Fonteneau et al. (2017). Furthermore, various genetic analyses, like Multi-Locus Sequence Typing (MLST) and acquired antibiotic resistance gene content determination, were determined on assembled sequences with web-based tools2 and by an in-house Perl script. For each isolate, the paired-end reads were aligned against the S. enterica serotype Mbandaka str. 9367/03 genome3.
Campylobacter spp. were detected and isolated as described in Supplementary Data S2.
Genomic DNA was extracted from colonies suspended in water using Nuclisens protocol (BioMérieux). Isolates were confirmed as Campylobacter spp. by detecting 16S rRNA gene by qPCR (Leblanc-Maridor et al., 2011). Species were identified by mass spectrometry using a MALDI-TOF Bruker Microflex apparatus (MALDI-TOF Bruker Microflex, Billerica, MA, United States) and following the protocol described by the supplier.
Detection and isolation of V. parahaemolyticus, V. cholerae, and V. vulnificus in a selection of samples including shellfish and seawater were performed as described in Supplementary Data S2.
DNA was heat-released from APW-enriched cultures and presence of total and haemolysin-producing V. parahaemolyticus and of V. cholerae and V. vulnificus was detected by qPCR and PCR. Presumptive V. parahaemolyticus, V. vulnificus and V. cholerae colonies were selected from PCR-positive APW-enriched cultures streaked (previously) onto chromogenic (CHROMagarTM Vibrio, Humeau) and selective (Thiosulfate Citrate Bile Salts Sucrose, TCBS, Difco) media plates and incubated 24 h at 37°C. These colonies were analyzed by qPCR and PCR using the primer sets described in Supplementary Data S2.
For detection of HuNoVs, shellfish were shucked, and the digestive tissues (DT) were recovered and frozen under aliquots of 2 g. For analysis, Mengovirus (MgV) (2.106 RNA copies) was added to each sample and incubated with 2 ml of proteinase K solution (30 U/mg, Sigma-Aldrich, France) at 37°C under shaking for 15 min and then at 60°C for 15 min. After centrifugation at 3,000 g for 5 min, the supernatant was recovered. Nucleic acids (NAs) were extracted using the NucliSens extraction kit (BioMérieux) with increasing the lysis buffer volume (Le Mennec et al., 2017). After checking the extraction efficiency by amplification of MgV, HuNoVs GI and GII were detected as previously described using 5 μl of undiluted and 10-fold dilutions of the NA extract on a Mx3000P qPCR System (Agilent Technologies, France). Only sample with a MgV extraction efficiency >1% were considered for quantification. The number of RNA copies present in each positive evaluable sample was estimated by comparing the sample Cq value to standard curves. Calculated concentrations for GI and GII were added to express the final result as HuNoV RNAc/g of DT.
Rainfall data (2-days cumulative rainfall before sampling date) were provided by the meteorological stations from Meteo France at Pleurtuit (site 1) and at Coutances (sites 2 and 3). On-site water temperature, and salinity were measured using a multi sensor probe 3430 (WTW) equipped with a FDO 925 electrode and a pH-electrode Sentix 940 (site 1) or an EcoSence EC300 apparatus (VWR) (sites 2 and 3). Data on temperature and precipitation were categorized into three groups whose boundaries were defined to have a number of samples compatible with a reliable statistical analysis in each category. Comparisons of prevalence and microorganism’s characteristics between samples type, site, season, temperature and precipitation were analyzed by the chi-square test using the CHISQ.TEST function of the Excel software (Microsoft). A P-value of <0.05 was considered statistically significant. In case sample was positive for multiple pathogens of interest, it was considered positive for each of the pathogens in question.
The Shannon diversity index was calculated using the Biodiversity Calculator tool4.
Odds ratios (OR) and relative risk (RR) were calculated using MEDCALC5 and considered statistically significant when P-values were less <0.05. OR is the odds of a positive sample for a given parameter among positive samples for a second parameter while RR is the probability that a sample is positive for a given parameter among samples positive for a second parameter relative to the probability that it is positive for the given parameter among samples negative for the second parameter.
For all sites, concentrations of E. coli were variable and generally high in river water (geometric mean (GM) of 3.2 Log per 100 ml) and cockle samples (GM of 3.3 Log per 100 g) while oysters, mussels and seawaters were less contaminated (GM of 2.6, 2.9 and 1.9 Log, respectively). Mussels were found more contaminated than oysters (Figure 2A). Concentrations of enterococci in water were also variable but lower than those of E. coli with GM from 2.1 to 3.6 Log of enterococci per 100 ml in river waters and from 1.4 to 1.7 Log in seawaters (Figure 2B).
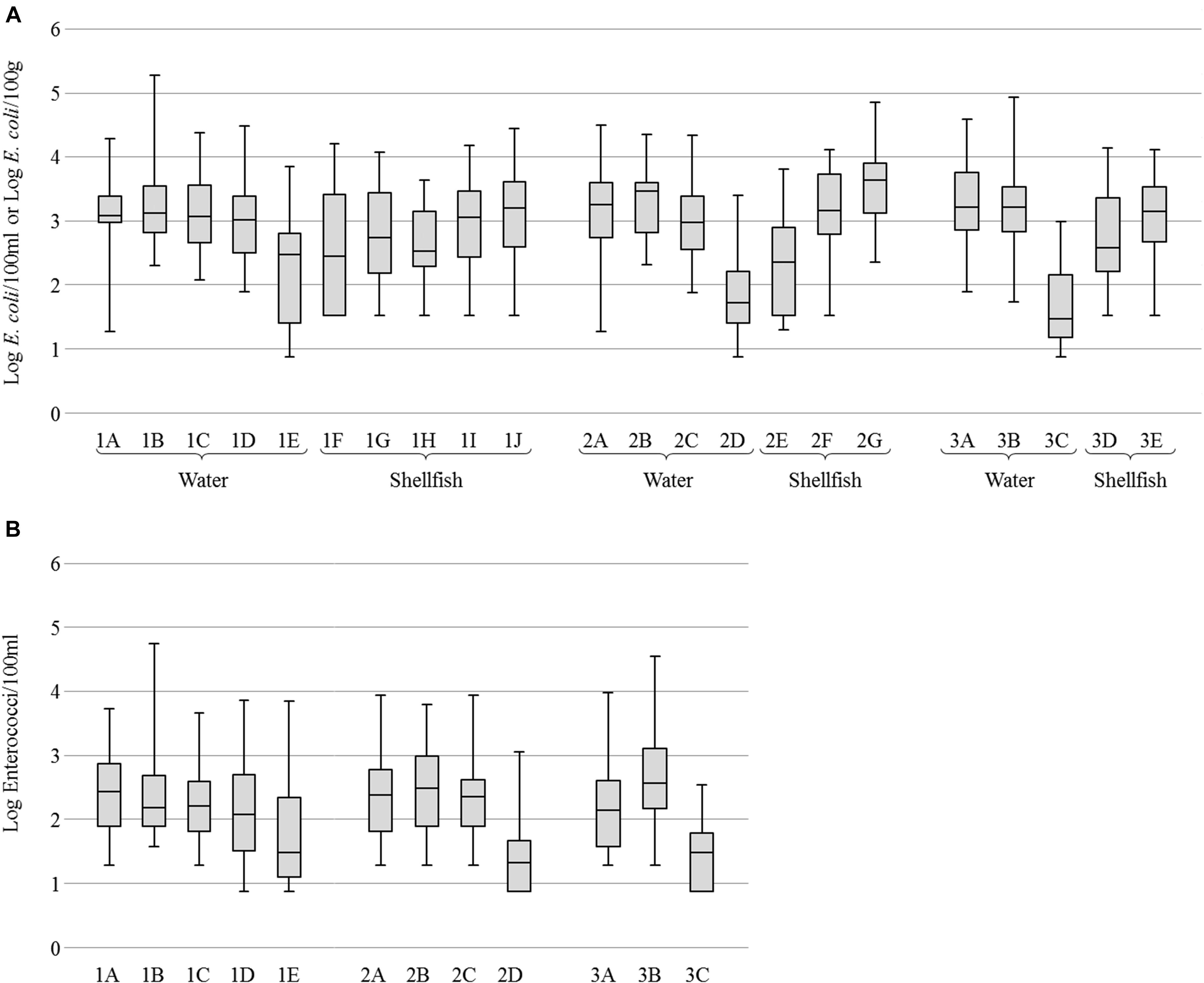
FIGURE 2. Enumeration of fecal indicator bacteria. (A) Boxplots of Escherichia coli concentration in water and shellfish. (B) Boxplots of Enterococci concentration in water. Boxplots show the minimum, the 25th percentile, the median, the 75th percentile and the maximum concentration. For water samples, 1E, 2D, and 3C correspond to seawater and others to river or brackish water. For shellfish samples, 1F, 1H, 2E, and 3D correspond to oysters, 1G, 1I, 2F, and 3E to mussels, and 1J and 2G to cockles.
7,452 E. coli strains were isolated and classified in four phylogenetic groups, 40.5, 31.5, 12.1, and 15.9% of strains belong to phylogroups A, B1, B2, and D, respectively (Figure 3 and Supplementary Data S4). Two thousand and twenty four strains from phylogroups B2 and D, (1,146 from D and 876 from B2) were analyzed for the presence of nine virulence genes (fyuA, hlyC, sfa, papC, iucC, papGIII, cnf1, papGII, and iroN). This analysis (the complete results are presented in Supplementary Data S4) revealed a higher percentage of strains from site 1 carrying the fyuA, sfa, or papGIII genes compared to strains from the other sites. It also revealed that the mean virulence score was higher for strains belonging to B2 phylogroup (3.9) than for those of phylogroup D (3.0).
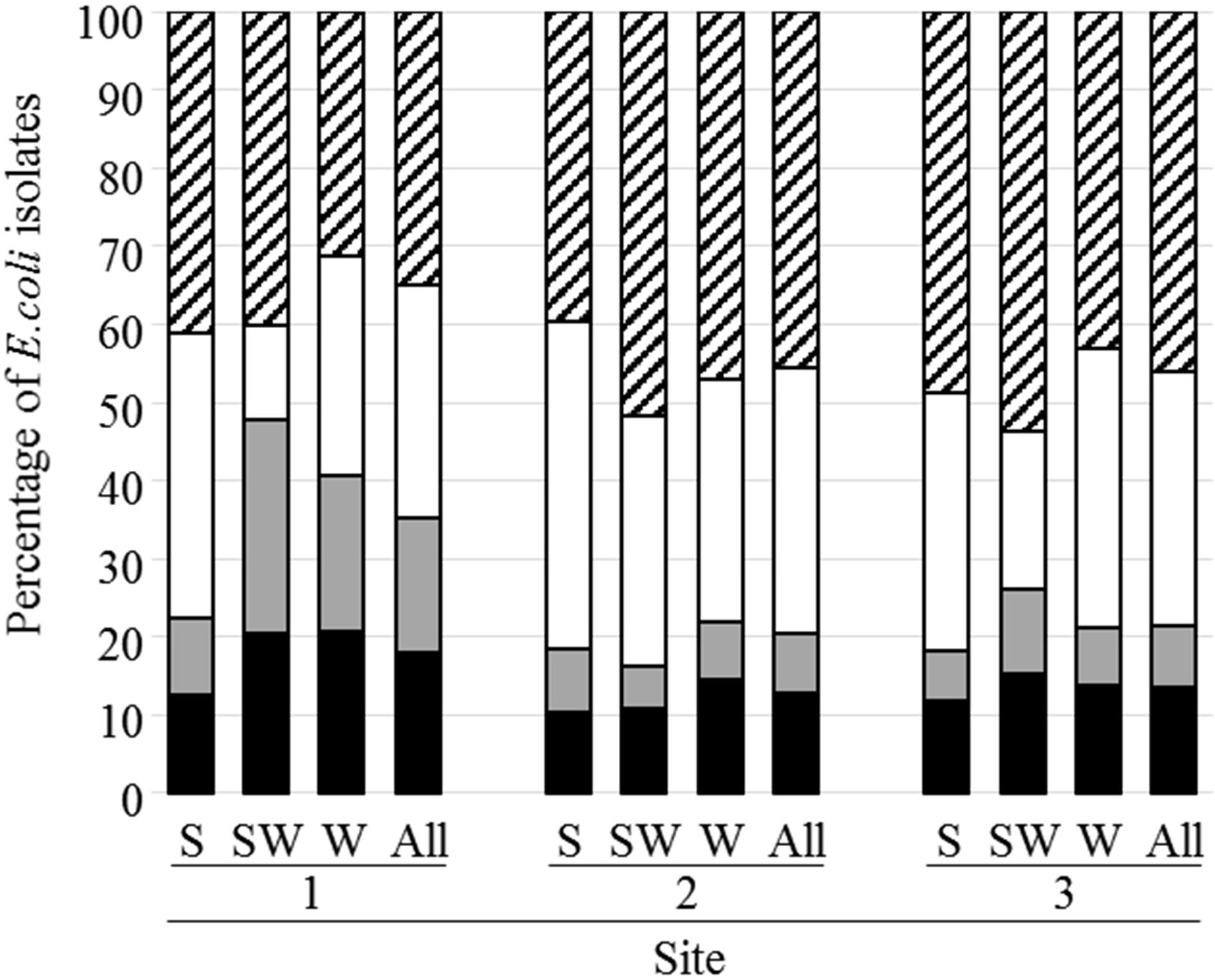
FIGURE 3. Distribution of E. coli isolates according to their phylogenetic group. The phylogenetic groups A, B1, B2, and D are represented by hatched, white, gray and black bars, respectively. S, shellfish; SW, seawater; W, river or brackish water.
Four hundred and forty nine E. coli strains were analyzed to evaluate their level of resistance against a panel of 16 antibiotics or combinations of antibiotics (Supplementary Data S4). The results showed that a higher proportion of E. coli from Les Hardes river (sub-catchment of site 3) were resistant to cefoxitin, amoxicillin + clavulanic acid, and amoxicillin when compared to isolates from the other rivers. On the other hand, isolates from Le Frémur river (sub-catchment of site 1) were statistically more frequently resistant to doxycycline.
A total of 4,344 enterococcal strains were isolated, of which 3,887 were identified by mass spectrometry. These strains belong to 26 different species where E. faecalis (32.9%), E. faecium (23.2%), E. hirae (13.7%), E. casseliflavus (8.5%) E. mundtii, (7.4%), and E. durans (6.5%) represent the predominant species (Figure 4). The H′ biodiversity index of Shannon was similar for the three sites (2.5, 2.9, and 2.7, respectively, for sites 1, 2, and 3). These species were also the most abundant in water samples (E. faecalis: 29.1%, E. faecium: 23.6%, and E. hirae: 14.0%) and in shellfish (E. faecalis: 39.0%, E. faecium: 22.6%, and E. hirae: 12.9%).
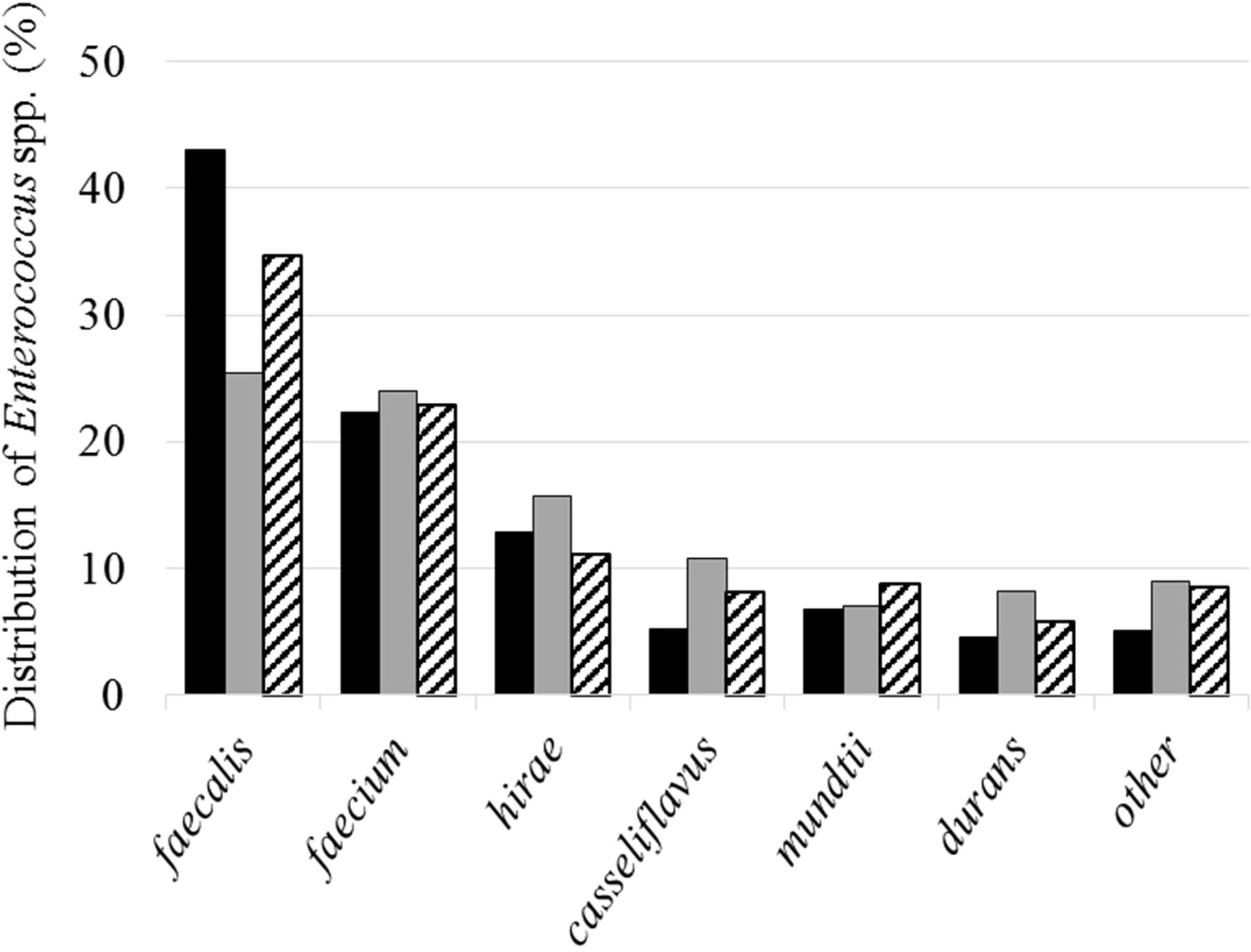
FIGURE 4. Distribution of the main isolated species. Black, gray and hatched bars represent results from sites 1, 2, and 3, respectively.
Two thousand and sixty-nine strains (1,200 E. faecalis and 869 E. faecium) were analyzed for the presence of three virulence genes (esp, gelE, and agg) (Supplementary Data S4). These genes were more frequently detected within E. faecalis than E. faecium species. Their occurrence was comparable from one site to another, except for gelE, which was more frequently detected in E. faecalis from site 1 (56.3%) than from sites 2 and 3 (45.8%).
Susceptibility of 213 E. faecalis and 233 E. faecium strains to 16 antibiotics was also determined (Supplementary Data S4). Interestingly, the percentage of E. faecium resistant to levofloxacin, ampicillin, and streptomycin was higher for strains isolated from water samples than from shellfish. Analysis of isolates from rivers, revealed that 64.3% of E. faecium strains from La Sienne river (sub-catchment of site 2) and 55.5% from La Soules (sub-catchment of site 2) were resistant to ampicillin, whereas all strains from the other rivers were sensitive to this antibiotic. Furthermore, E. faecium strains isolated from La Sienne were also statistically more frequently resistant to levofloxacin.
Salmonella spp. (468 colonies in total) were isolated from 69 samples (13.7% of the 505 samples tested). These samples were mainly from river and brackish waters (30.1% of sample with isolated Salmonella) and rarely from shellfish (3/237; 1.3%) or sediment (2.5%) while none of the 12 seawater samples analyzed allowed isolation of Salmonella (Figure 5A). The site 2 presented a higher percentage of samples with Salmonella (24.4%) and it was when the height of rainfall exceeded 10 mm per 48 h that the highest percentage of positive samples was observed (Figure 5A). 51.4% of the river and brackish water samples from site 2 and 19.8 and 18.7% of the corresponding samples from sites 1 and 3, respectively, were positive for Salmonella spp. Positive samples were found at each of the four seasons (Figure 5A).
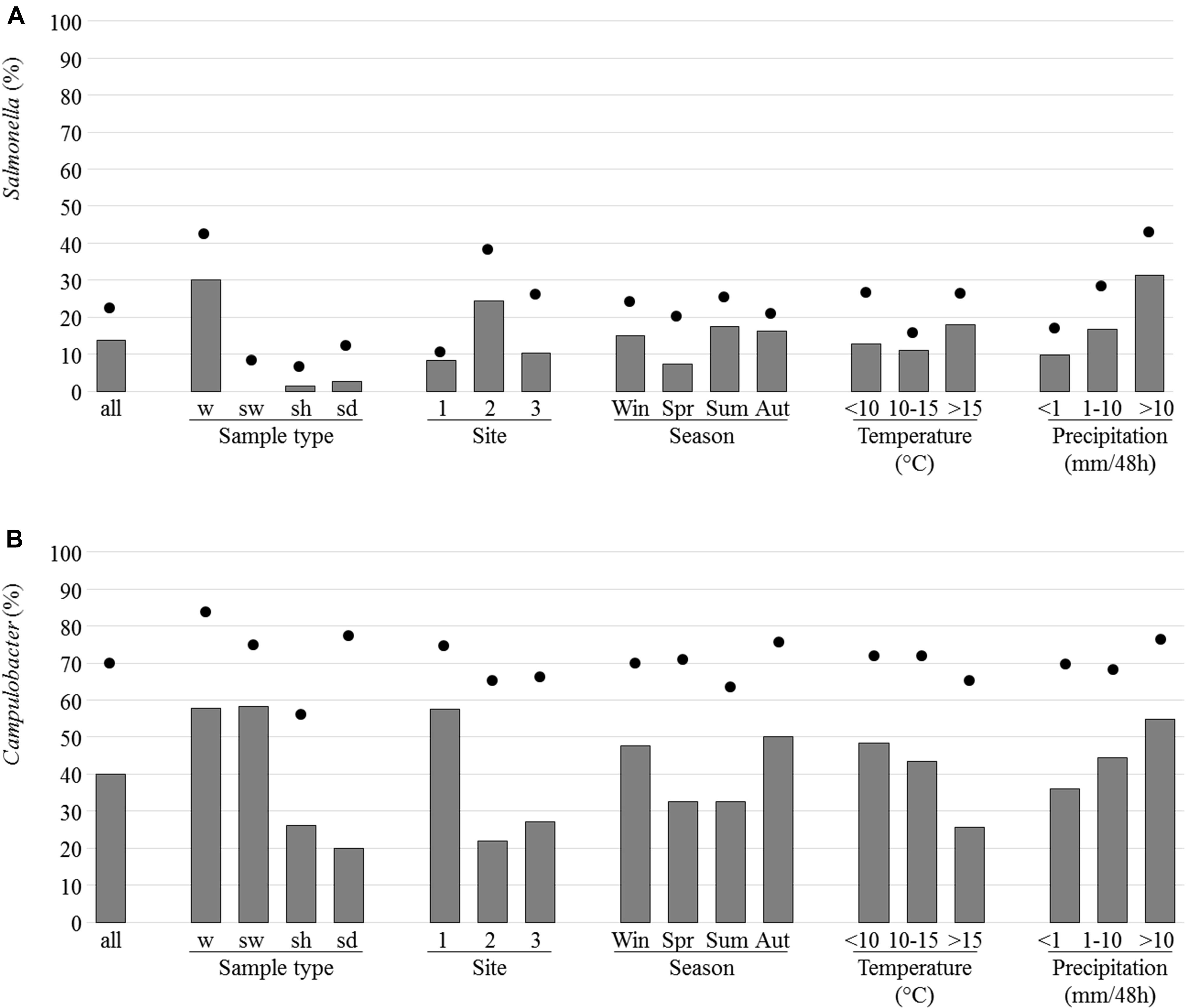
FIGURE 5. Prevalence and isolation of Salmonella (A) and Campylobacter (B). The prevalence which corresponds to the presence of the invA or ttrBCA genes in selective enrichments for Salmonella or 16S RNA genes in Bolton broth for Campylobacter (dots) and the percentage of samples for which at least one strain was isolated (bars) are indicated as regard to sample type [water from river and brackish water (w), seawater (sw), shellfish (sh), and sediment (sd), site, season, temperature, and precipitation].
Of the 468 colonies, 75 Salmonella isolates were retained for further analyses. The remaining 393 isolates were considered replicates for the purpose of the present study (same serovar from the same sample or isolates from the same sample not serotyped). Seventy-one originated from water, three from shellfish (one from each type of shellfish and from each site) and one from sediment. These strains were assigned to Salmonella enterica subsp. enterica for 73 isolates and to Salmonella enterica subsp. diarizonae for two isolates belonging to the same 21:z10:z53 antigenic formulae. Among S. enterica subsp. enterica, the isolates were divided into 13 serovars (Table 1). Mbandaka was the most frequent serovar (36.0%) and was the only one found in the three positive shellfish batches. Eight strains from Typhimurium serovar were isolated (10.7%) and 10 others carrying O antigens O:4,5,12 were non-motile (two) or monophasic (eight) with antigen H:i, and one carrying O:4,12 antigen was monophasic with antigen H:i. CRISPOL typing confirmed that Salmonella Typhimurium and monophasic or non-motile variants belonged to the Typhimurium population of Salmonella and displayed diverse CRISPOL types (CT) 1, 18, 21, 30, 76, or 255. The CRISPOL types the most frequently found were CT 1 and 18 and mainly came from the Brittany site (three different rivers).
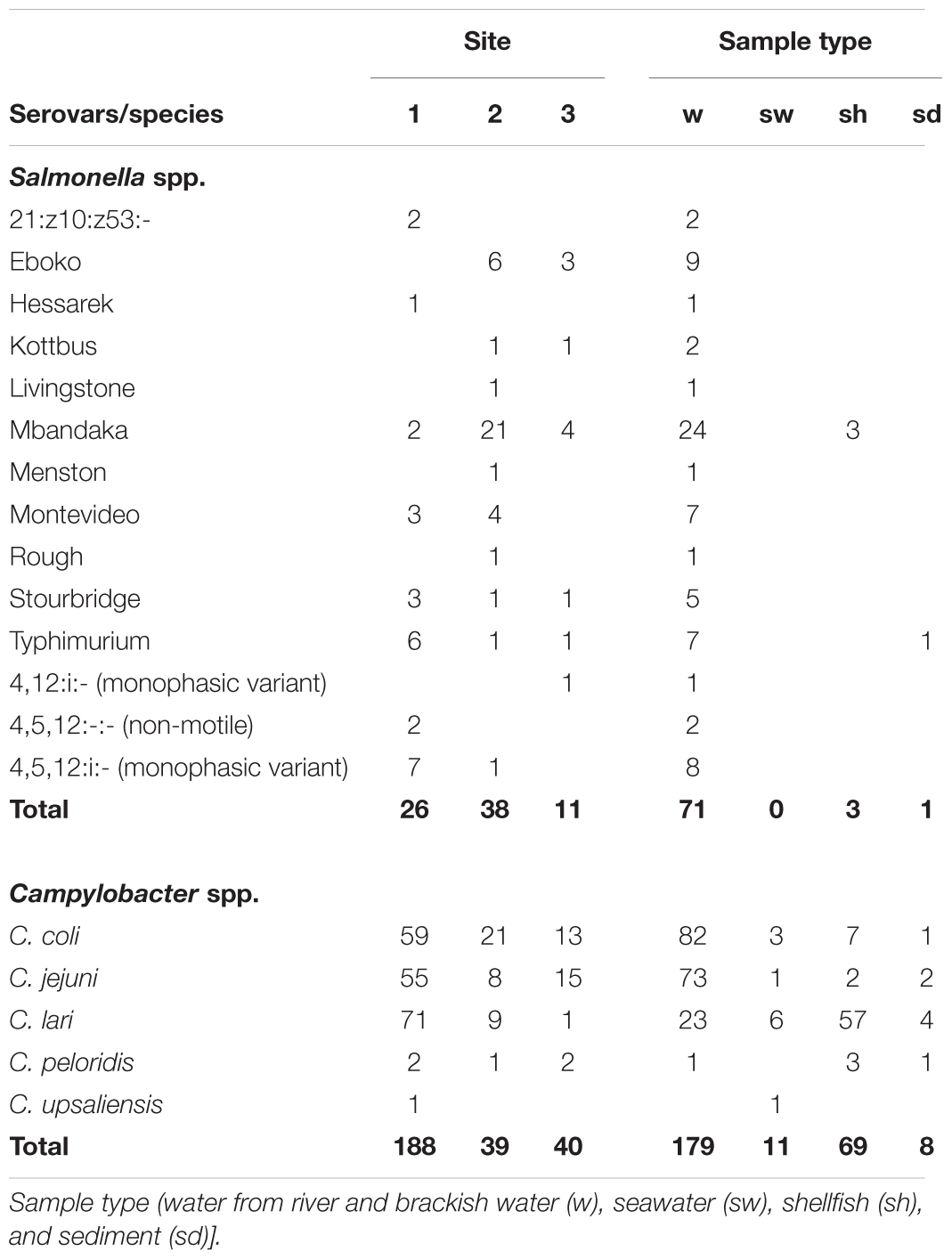
TABLE 1. Presence of Salmonella serovars and Campylobacter species in the three sites and in different types of samples (only isolates from different samples and different species from the same sample were retained).
Several Salmonella serovars were isolated from each site and their distribution was different from one site to another. On the site 1, mainly S. Typhimurium and its variants were isolated whereas S. Mbandaka and, in a lesser extent, S. Eboko were mainly isolated on the sites 2 and 3 (Table 1). If S. Typhimurium, monophasic variants, S. Mbandaka, and S. Stourbridge were isolated at least once from all sites, the serovars Eboko or Kottbus and Montevideo were only isolated from sites 2 and 3, and sites 1 and 2, respectively. The other serovars were isolated once or twice from only one site. On the site 1, the S. Typhimurium and monophasic variants were mainly isolated from Le Rat sampling site.
Among 14 S. Mbandaka typed by PFGE, we found six different pulsotypes. WGS analysis performed on 21 S. Mbandaka indicated that all belonged to the MLST type ST413 and none antibiotic gene was found. On all of them we counted 1,296 SNPs and phylogenic analysis did not allowed us to detect any clusters under 50 SNP divergence, suggesting high diversity population and so absence of a unique source.
Campylobacter (1,400 colonies in total) were isolated from 204 of the 505 samples corresponding to 58.3% water, 26.6% shellfish and 20% sediment samples analyzed (Figure 5B). It was on site 1 and when rainfall height exceeded 10 mm per 48 h that the highest percentage of positive samples was observed (Figure 5B).
Campylobacter jejuni and C. coli were the most frequently found species in river and brackish water samples with 33.8 and 38% of samples positive for these species, respectively, versus 10.6 and 0.5% of samples positive for C. lari and for C. peloridis, respectively. C. lari was the most frequently isolated species in shellfish with 26.4% of samples positive for this species versus 0.8% for C. jejuni, 2.9% for C. coli, and 1.3% for C. peloridis (Table 1). More positive samples were observed in autumn and winter with temperature mainly under 15°C (Figure 5B). C. jejuni and C. coli were more frequently isolated in autumn (36.3% of positive samples) and C. lari in winter (34.6%). One species was isolated in 72.1% of the samples positive for Campylobacter spp. (147 of 204) whereas two species were isolated in 25% of the samples and only six samples (2.9%) allowed the isolation of three species.
Of the 1,400 clones, 267 Campylobacter isolates were retained for further analyses (isolates from different samples and different species from the same sample). One hundred and seventy nine originated from river and brackish waters, 11 from seawater, 69 from shellfish and eight from sediment. One hundred and eighty eight were isolated from site 1, 39 from site 2, and 40 from site 3.
Campylobacter spp. were more frequently isolated than Salmonella spp on these sites. In fact, 169 of the 505 samples (33.5%) were positive for Campylobacter spp. only versus 35 (6.9%) positive for Salmonella only and 34 (6.7%) positive for both pathogens.
Among the 188 analyzed samples (43 seawaters and 145 shellfish batches), 56.9, 25.5, and 30.9% were positive for V. parahaemolyticus, V. cholerae, and V. vulnificus, respectively. For these Vibrio species, the highest prevalence was observed during the summer months and when water temperatures were above 15°C and/or when the height of precipitation exceeded 10 mm per 48 h (Figure 6). None of them was detected when water temperature was below 10°C.
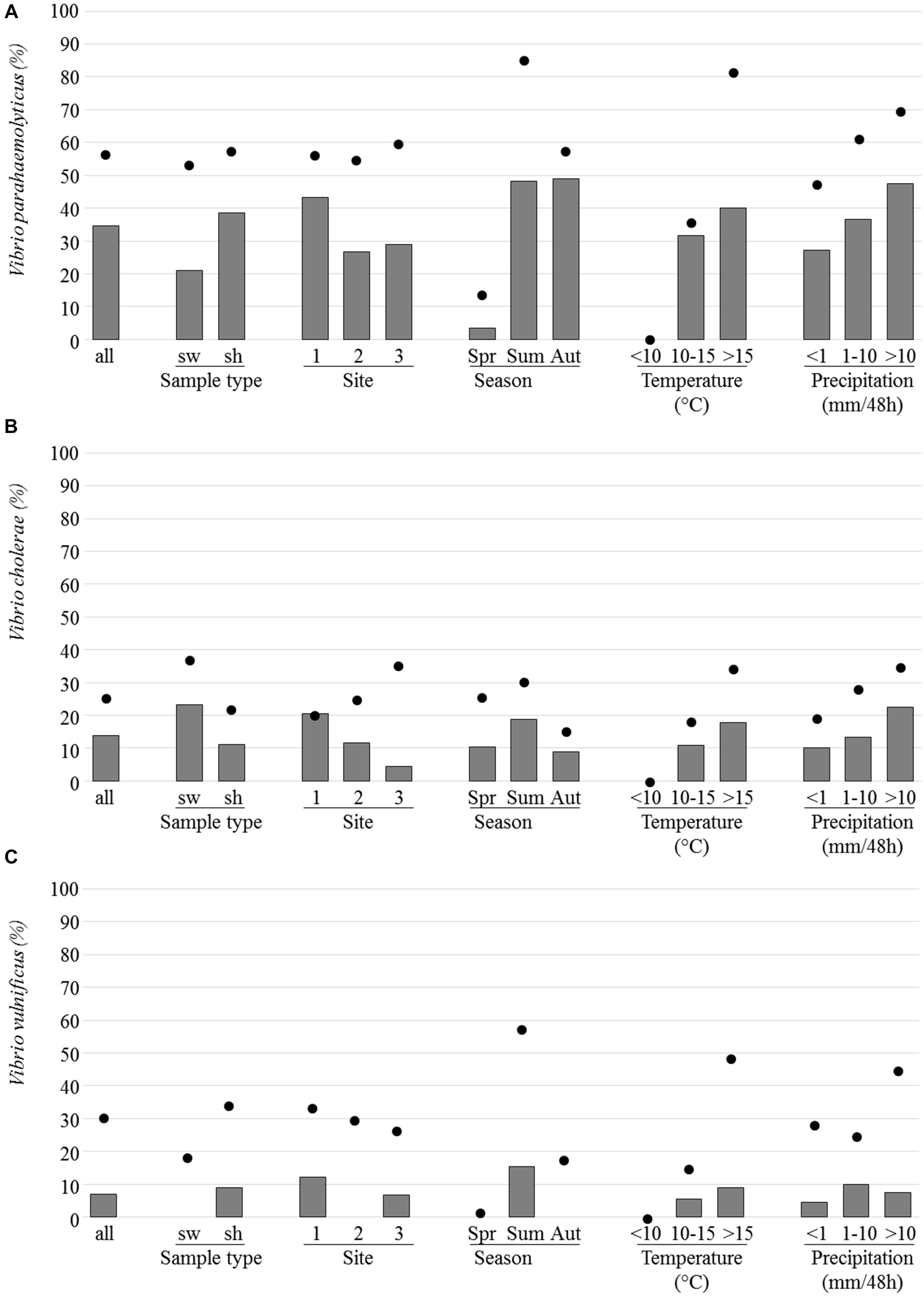
FIGURE 6. Prevalence and isolation of Vibrio parahaemolyticus, Vp (A), Vibrio cholerae, Vc (B), and Vibrio vulnificus, Vv (C). The prevalence which corresponds to the presence of the toxR gene, the IGS region and the vvhA gene in selective enrichments for Vp, Vc, and Vv, respectively (dots) and the percentage of samples for which at least one strain was isolated (bars) are indicated as regard to sample type [seawater (sw), and shellfish (sh), site, season, temperature, and precipitation].
Vibrio parahaemolyticus was the most frequently detected Vibrio species in the three sites with 56.6, 55.0, and 60.0% of positive samples from sites 1, 2, and 3, respectively. The prevalence of total V. parahaemolyticus was quite similar in seawater (53.5%) and shellfish (57.9%) (Figure 6). It was detected alone (39.2% of the 107 positive samples) or in combination with one (V. vulnificus, 29.0%; V. cholerae, 16.8%) or the two other species (15.0%). Enteropathogenic tdh positive V. parahaemolyticus was present in only six shellfish and two seawater samples and in the three sites but no tdh positive Vp strain was isolated. The prevalence of potentially pathogenic trh positive V. parahaemolyticus samples was higher in seawater than in shellfish samples (92.3 and 88.1%, respectively). It was detected in all the V. parahaemolyticus positive samples collected in Brittany (site 1). Trh strains were isolated from 58% of the V. parahaemolyticus positive samples.
Vibrio cholerae was found in seawater and shellfish (37.2 and 22.1%, respectively) and most prevalent in site 3 whereas V. vulnificus was detected more often in shellfish (34.5%) than in seawater samples (18.5%) and most prevalent in site 1 and during the summer months. V. cholerae strains were isolated from 13.8% of analyzed samples and V. vulnificus strains from 6.9%.
A total of 150 shellfish samples were analyzed for HuNoVs (Table 2). The average extraction efficiency was 9% and varied from 2% for cockle samples to 12% for mussel samples. HuNoVs were detected in 33 (22%) of the samples analyzed, 12 being under the limit of quantification (35 RNAc/g of DT). GM concentrations calculated for each species were comparable. As expected HuNoV positive samples were detected mainly during autumn and winter (Table 3). A similar rate of positive samples for HuNoV was observed between site 1 (21.1%) and site 2 (23.6%).
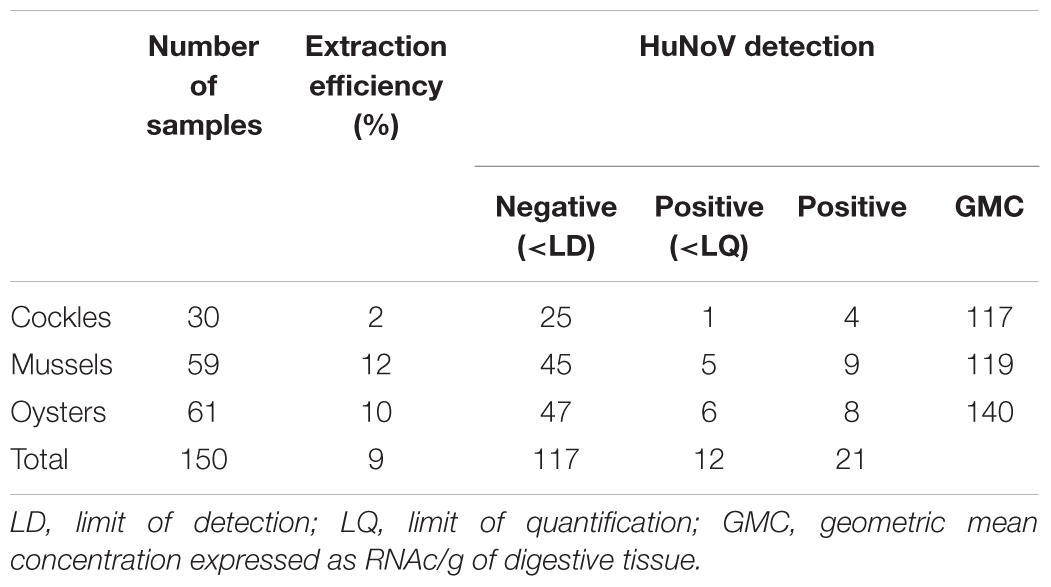
TABLE 2. HuNoV contamination in the shellfish species analyzed.
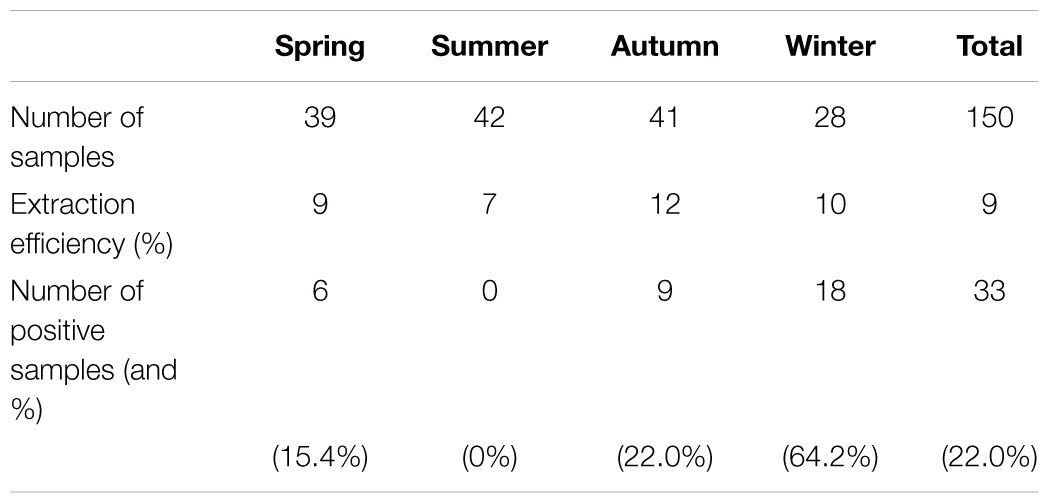
TABLE 3. Seasonal variation of HuNoV contamination.
Comparison of results from isolation of enteric bacterial pathogens or detection of HuNoV with E. coli enumeration showed a positive correlation between the presence of Salmonella and E. coli concentration (OR = 3.94; Table 4). The relative risk of isolating Salmonella in samples whose E. coli concentration exceeds the median was 3.28 times higher than in those with E. coli concentration below the median. A positive correlation was also observed between the presence of STEC and/or EPEC and samples with the highest E. coli concentration (OR = 1.74 and RR = 1.57). In addition, a positive correlation between the presence of enteric pathogens was observed for Salmonella with C. jejuni and/or C. coli (OR = 2.58, RR = 2.21), for STEC and/or EPEC with C. jejuni and/or C. coli (OR = 2.79, RR = 2.00), and for C. lari with HuNoV (OR = 2.42, RR = 1.77).
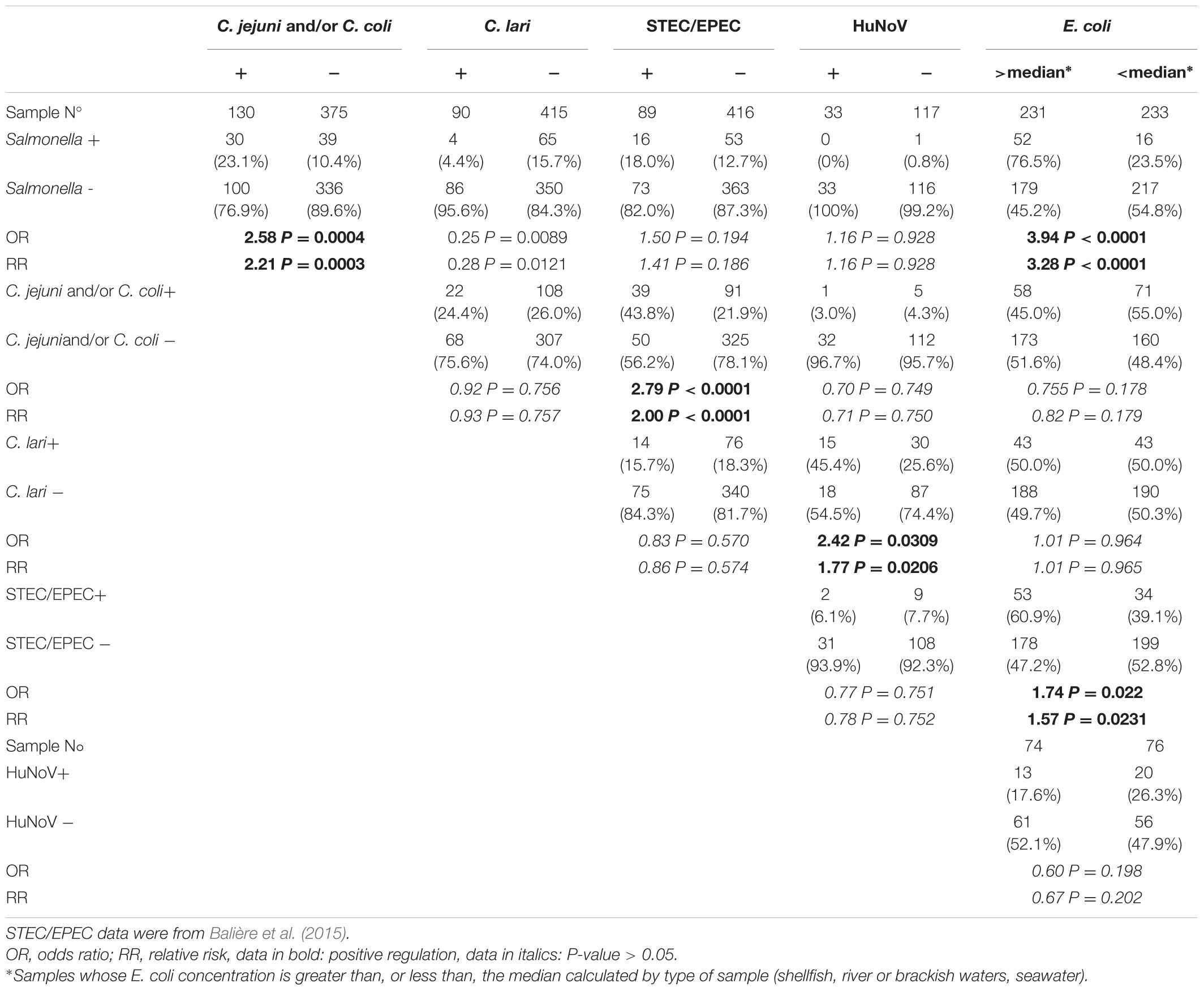
TABLE 4. Correlations between E. coli concentration, isolation of pathogenic enteric bacteria and detection of HuNoVs.
This original study aims to detect and characterize a large panel of micro-organisms from fecal contamination (Salmonella spp., Campylobacter spp., E. coli and enterococci strains and HuNoVs) as well as some marine bacteria such as Vibrio spp.. For this study samples were collected from shellfish-harvesting areas and their catchments, but also from seawater. This has rarely been performed, most of other reporting data on these pathogens specifically relate either in shellfish or in water samples.
The enumerations of the FIB showed that water from all three catchments is characterized by fecal pollution that is variable and can reach relatively high levels at certain periods.
Classification of E. coli into phylogenetic groups revealed that isolates belonging to phylogroup A or B1 were more predominant than clones from group B2 or D. This observation is in agreement with previous results of surface water samples and shellfish (Stange et al., 2016; Vignaroli et al., 2016). Previous data obtained from the same river water samples showed that 5.6 and 21.3% of these waters were contaminated with STEC and EPEC, respectively (Balière et al., 2015). This means that these pathotypes were less common than other pathogenic bacteria mentioned above. Here, we focused on characterization of E. coli strains belonging to phylogroups B2 or D, the most frequently represented within ExPEC.
Analysis of antibiotic susceptibility of 338 E. coli isolated from river and brackish water samples and which belongs to phylogroups B2 or D showed a high proportion of bacteria resistant to cefoxitin, amoxicillin + clavulanic acid and amoxicillin in Les Hardes river and of clones resistant to doxycycline in Le Frémur.
Numerous studies have clearly shown that extraintestinal pathogenic strains generally contain more virulence factors than commensal strains. Here, we found that the average virulence score was higher for strains isolated from shellfish in site 1 than in sites 2 and 3. Recently, Johnson et al. (2017) assigned the ExPEC status to E. coli isolated from surface waters and animals based on the detection of a set of virulence genes. Their analysis focused on the detection of papAH and/or papC (encoding P fimbriae), sfa/focDE (S and F1 fimbriae), afa/draDC (Dr-binding adhesins), kpsMI (group 2 capsule) and iutA (aerobactin system); a strain being considered ExPEC when positive for at least two of these genes. In the present study, papC and sfa were selected and the percentage of strains isolated from shellfish that were positive for these two genes was also higher for site 1 (12.5%) than for sites 2 and 3 (8.5 and 9.3%, respectively). This would mean that the risk associated with the presence of ExPEC would be higher at the site 1.
In water, the predominant Enterococcus species were E. faecalis, E. faecium, and E. hirae followed by E. casseliflavus, E. mundtii, and E. durans which is in agreement with the species usually cited as the most abundantly detected in surface waters (Lanthier et al., 2011; Furtula et al., 2013; Sidhu et al., 2014; Veljović et al., 2015). As previously observed by Sidhu et al. (2014) virulence genes were detected more frequently in E. faecalis than in E. faecium isolates. For the collection of environmental E. faecalis tested here, 34.9, 49.8, and 46.0% of strains were positive for genes esp, gelE, and agg, respectively. These values, although high, are lower than those of clinical isolates described by Soares et al. (2014) who observed 70.1, 78.7, and 63.4% E. faecalis strains containing esp, gelE, and agg genes. As described by Lanthier et al. (2011), the presence of enterococci with multiple virulence genes is an additional risk factor for public health. Among the E. faecalis analyzed here, it is in the site 1 that we observed the highest percentage of isolates positive for at least two virulence genes.
Analysis of antibiotic susceptibility of Enterococcus isolates indicated that the sub-catchments of the rivers La Sienne and La Soules constitute the main source of ampicillin-resistant E. faecium and that most of the E. faecium isolates from the river La Sienne were resistant to levofloxacin. Additional analyses including the study of antibiotics used in farms or in human medicine on this particular site should make it possible to identify more precisely the corresponding source.
Salmonella was frequently detected in the three coastal catchments and Salmonella spp. was isolated in 30.1% of samples (range from 17.8% in site 1 to 51.4% in site 2). Such variable frequency of Salmonella positive samples has been already reported in the literature. Indeed, 43% of water samples from Georgia (United States) and 23% of surface water in Canada were also positive for Salmonella (Vereen et al., 2013; Flockhart et al., 2017).
A different distribution of serovars was observed according to the site. S. Typhimurium, and monophasic variants were mainly isolated on the site 1 whereas S. Mbandaka was mainly observed on site 2. This could be explained by the different agricultural activities on these catchments, with mainly swine, cattle and poultry breeding in Brittany and cattle, sheep and poultry in Normandy (Supplementary Data S1). Salmonella Mbandaka are known to be mainly present in cattle and poultry (European Food Safety Authority [EFSA] and European Centre for Disease Prevention Control [ECDC], 2015) while swine are one of the major sources of Salmonella Typhimurium, in particular its monophasic variants (Martelli et al., 2018). Furthermore, the main CRISPOL type identified in these latter strains was the CT1, a Salmonella population known to be responsible for most of human infections and frequently abundant in swine in France (Fabre et al., 2012). S. Mbandaka was described as highly persistent in the environment near farms due to its ability to produce biofilms and to persist and grow in the external environment and on animal feed (Hayward et al., 2016). Furthermore, S. Mbandaka was found to be more resistant to sunlight (solar simulator; passing wavelength, 290 nm < λ < 800 nm) than S. Typhimurium and other serovars (Boehm et al., 2012). Contrary to S. Mbandaka, S. Typhimurium and monovariants are one of the serotypes frequently implicated in food-borne outbreaks (European Food Safety Authority [EFSA] and European Centre for Disease Prevention Control [ECDC], 2016). The main presence of S. Typhimurium in water samples was already observed in coastal catchments in United States and catchments in Canada with also the presence of S. Mbandaka (Walters et al., 2013; Jokinen et al., 2015).
In shellfish, the prevalence of Salmonella spp. was low (1.3%) and similar to those observed in market oysters in United States (DePaola et al., 2010) but lower than those from other studies (8% of molluscs in Northern Ireland, Wilson and Moore, 1996; 7.4% of oysters from 36 United States bays, Brands et al., 2005; 10% of mussels in Morocco; Setti et al., 2009). Interestingly, the presence of only one serovar that is S. Mbandaka (three shellfish batches in the three sites) was different from previous studies in which several serovars such as S. Newport, S. Typhimurium, S. Agona, S. Blockey, S. Kentucky were isolated (Brands et al., 2005; Setti et al., 2009). The presence of only S. Mbandaka in shellfish could be explained by the better survival of this serovar in the environment and the presence of poultry and cattle breeding, main reservoir of S. Mbandaka, upstream the shellfish-harvesting areas. The identification of this serovar suggests a low risk of contamination by Salmonella in shellfish. However, S. Mbandaka ST413 (the main ST of this serovar and found in all isolates here) was responsible of food-borne illnesses in Poland (Hoszowski et al., 2016).
As for Salmonella, Campylobacter has been frequently isolated in the studied catchments, with an isolation in 58.3% of water samples. On the three sites, water samples were more frequently positive for C. coli and C. jejuni than C. lari. The highest prevalence of C. jejuni and C. coli was also observed by Denis et al. (2011b) and Khan et al. (2014) who found more frequently C. jejuni than C. coli and C. lari in Canada and in France (Brittany), respectively. These different species could arise from both human and animal origins (Pitkanen and Hanninen, 2017). If we consider the breeding sites and the prevalence described for C. coli, C. jejuni, and C. lari in the feces and manure of the corresponding animals, C. coli could arise mainly from swine, poultry, and sheep and C. jejuni from cattle, poultry and sheep. Furthermore, another important source of Campylobacter, including C. lari could be wild birds (Ryu et al., 2014).
In shellfish, the prevalence of C. jejuni (0.8%) and C. coli (2.9%) was very low whereas C. lari was most frequently isolated in these samples (24.1%) than in river waters. Campylobacter was the most often bacterial enteric pathogen isolated in shellfish (27.8% of samples positive for Campylobacter) which is lower than the 42% of shellfish with thermophilic Campylobacter spp. observed by Wilson and Moore (1996). Interestingly, a similar distribution among the species was observed in both studies. Indeed, C. jejuni, C. coli, and C. lari represented 3, 10.6, and 86.1% of the Campylobacter spp. isolated in the present study, respectively, and 2, 8, and 81% of the Campylobacter spp. isolated by Wilson and Moore, respectively. The fact that C. lari was more present in marine environment and especially in shellfish comparing to C. jejuni and C. coli, whereas the opposite was observed in upstream waters, suggests a better persistence of this species in marine environment and/or an input from wild seabirds, as observed previously in Morecambe Bay (United Kingdom) by Obiri-Danso et al. (2001). These two hypotheses will be investigated in the future.
Concerning the marine bacteria, the incidence of total Vibrio parahaemolyticus was similar in shellfish and seawater samples (57.9 and 53.5%) and equally distributed between the three sites (55–60%). Culturable V. parahaemolyticus was detected in summer and autumn although the highest incidence (85.9%) was found in summer. Previous studies conducted in European coastal areas showed lower occurrence of V. parahaemolyticus in seawaters (12–31.4%) (Deter et al., 2010; Rodriguez-Castro et al., 2010; Martinez-Urtaza et al., 2012). However, similar or higher prevalence of V. parahaemolyticus was recorded in shellfish samples analyzed over the same seasonal period (Collin and Rehnstam-Holm, 2011; Suffredini et al., 2014).
The first incidence of V. parahaemolyticus tdh gene in Northern French waters confirm the recent reports of the presence of V. parahaemolyticus carrying this gene in Northern European waters (Collin and Rehnstam-Holm, 2011; Powell et al., 2013). However, no tdh+ strain was isolated in any of the three costal sites. The prevalence of V. parahaemolyticus trh gene in the environment appears very high (88.8%) and contrasts with previous observations in Southern and Northern Europe (Hervio-Heath et al., 2002; Esteves et al., 2015; Passalacqua et al., 2016).
Vibrio vulnificus and V. cholerae non-O1/non-O139 were detected in 30.9 and 25.5% of the environmental samples (34.5 and 22.2% in shellfish and 18.6 and 37.2% in seawater, respectively). This occurrence is much higher than previously reported in Europe for environmental samples except for blue mussels collected in 2006 in The Sound between Denmark and Sweden (63 and 53%, respectively) (Collin and Rehnstam-Holm, 2011). Recent emergence of wound and otitis infections caused by these two Vibrio species in Northern Europe (North Sea and Baltic Sea region) coincided with warm weather anomalies during summer months (Baker-Austin et al., 2013). These observations have to be considered and suggest that the presence of V. cholerae and V. vulnificus in French coastal waters could pose a potential hazard to shellfish consumers and for susceptible people exposed to the seawater especially during the summer months.
For HuNoVs the percentage of positive samples was higher compared to other studies performed in our country confirming the fecal contamination shown by FIB counts (Schaeffer et al., 2013). Extraction efficiency was always above the quality criteria stipulated in the ISO method, and is similar to results from previous studies (Schaeffer et al., 2013, 2018; Le Mennec et al., 2017). Even if most of samples presented a low concentration, this can be an issue for marketing considering the high infectivity of these viruses (Polo et al., 2016). As expected, most of positive samples were detected during the cold season when the virus is circulating in the human population and thus excreted in sewage (de Graaf et al., 2016).
The seasonal influence on the presence of human potential pathogens in shellfish was also observed for other human potential pathogens in this study. Firstly, as discussed above, Vibrio spp. were mainly detected/isolated on summer at the opposite to HuNoVs mainly detected in winter.
Concerning bacterial enteric pathogens, human Salmonella and Campylobacter infections typically occur in summer although environmental studies often show varied seasonal peaks for these pathogens (Vereen et al., 2013). Here, Campylobacter jejuni, C. coli, and C. lari were detected all year round in the environmental samples. However, a seasonal effect was observed with more positive samples in autumn and winter. C. jejuni and C. coli were more frequently isolated in autumn (36.3% of positive samples) whereas C. lari was more frequently isolated in winter (34.6%). These results are in agreement with those previously obtained in river water (Llobregat river, Spain), freshwater bathing sites (River Lune, United Kingdom) and coastal areas (Morecambe Bay and Lune Estuary, United Kingdom) which have shown higher Campylobacter concentrations in winter than in summer (Obiri-Danso and Jones, 1999; Jones, 2001; Rodriguez and Araujo, 2012). Vereen et al. (2013) also observed lower detection frequencies of Campylobacter in water of the Satilla River Basin (United States) in summer, reflecting a reverse association with temperature. Furthermore, a lower survival rate of Campylobacter in surface water in presence of sunlight (elevated UV levels) and higher temperatures has been observed (Obiri-Danso et al., 2001; Rodriguez and Araujo, 2012). In contrast, we did not show seasonal variation for Salmonella with year-round detection in rivers and only three isolations of Salmonella in shellfish (one in March and two in August).
Salmonella, Campylobacter, and Vibrio were more often detected after rainfall events in the three sites. This was also the case for E. coli concentrations (data not shown). For the enteric pathogens, these higher frequencies could be due to a higher release of these bacteria from wastes disposal in the fields by runoff and/or an exceedance of the waste water treatment plant capacity of collective plants, individual septic systems and/or non-collective sanitation. Rainfall was found to be the most significant environmental parameter driving the transfer of fecal contamination from soil to streams in three catchments in Brittany (including the site 1). In this later site, human-, bovine- and pig-associated markers of source of fecal contamination were more detected when rainfall >10 mm (Jardé et al., 2018).
For Vibrio, an input of nutrients to coastal waters by runoff could be in favor of a higher growth of these bacteria.
A variable presence of human potential pathogens according to the site was observed here for several pathogens. Firstly, Campylobacter was more frequently isolated in Brittany (site 1) than in Normandy (sites 2 and 3). Furthermore, S. Typhimurium and monophasic variants were the most frequently isolated Salmonella on the site 1 whereas it was S. Mbandaka on the site 2. These differences could be due to the urban and agricultural activities which are different in the studied sites and to the respective size of these catchments. If the variable densities of the different types of livestock (bovine, poultry, sheep, and pig) could explain the difference from one site to another, it is less clear at the level of sub-catchments. This lack of clear relationship in sub-catchments could be due to animal waste disposal practices. For example, in Brittany (site 1), poultry and pig were landless breeded whereas cattle are both reared outdoors on grass and in farms. These different rearing methods produce different types of wastes in this catchment which could be carried out both inside and outside the sub-catchment (pig manure), could be exported (poultry manure) or directly input on the field (cowpats).
Correlations were observed between the isolation of enteric bacterial pathogens and/or E. coli concentrations and likely reflect variations in waste disposal and runoff. In the selection of shellfish samples for which noroviruses were studied (n = 150), no correlation was found between the detection levels of E. coli or enteric bacteria Salmonella, STEC/EPEC or C. jejuni and/or C. coli, and HuNoVs whose circulation, as mentioned above, is more important in winter. On the other hand, a correlation was observed between HuNoVs and C. lari and this could be due to a similar seasonal distribution of C. lari and HuNoVs. To the best of our knowledge norovirus and bacteria co-infection in humans has not been reported yet. However, the discovery that human norovirus binds to some enteric bacteria raised hypothesis on the impact for their resistance in the environment, or to heat stress (Miura et al., 2013; Li et al., 2015) or may also play a role during infection (Sullender and Baldridge, 2018).
We demonstrate a high prevalence of potential enteric bacterial pathogens in these coastal catchments but a low prevalence of Salmonella and C. jejuni and C. coli in shellfish from the downstream harvesting areas. No correlation between detection levels of E. coli and that of HuNoVs neither Campylobacter spp. was shown, at the opposite to Salmonella. However, more importantly, a positive correlation between the presence of Salmonella and C. jejuni and/or C. coli and between the presence of C. lari and HuNoVs was observed; this latter observation could be due to their similar season distributions or to other factors that need to be investigated. The different distribution of Campylobacter species in rivers and shellfish could be in favor of additional inputs of bacteria such as wild birds.
MG and AR designed the study, participated in the bacterial analysis, and wrote the manuscript. CB participated in the bacterial analysis and especially in the isolation of E. coli and enterococci. J-CG performed the characterization of E. coli and enterococci. DH-H participated in the Vibrio analysis and in manuscript writing. SL and JC isolated and analyzed Salmonella and Campylobacter, respectively. SLH characterized Salmonella (serotyping and WGS) and revised the manuscript. NS and AB contributed to the isolation and the analysis of bacteria. SP and SS performed the HuNoVs analysis. FLG analyzed the HuNoVs data and participated in the writing of the manuscript.
This work was funded by the European Regional Development Fund Interreg IVA Programme, as part of the collaborative project RiskManche. The thesis of CB was supported by a grant from Ifremer and the Agence de l’Eau Loire-Bretagne.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank Marie-Paule Caprais, Jean-Claude Le Saux, Emmanuelle Quenot, Céline Hubert, Pascal Garry, Antoine Véron from Ifremer, Clémence Balière, Isabelle Rincé, Quentin Bruet from the University of Caen-Normandy, and Emilie Jardé, Loïc Harrault and Laurent Jeanneau from Geosciences Rennes for their participation in the sampling and/or technical assistance. We are especially grateful for the assistance of Julien Chevé from Ifremer (Dinard) for the analysis of catchment data on the Brittany site, and for the permission of the local stakeholders (Dinan agglomeration; Elvis Desnieuls and Michel Eldejam) to take samples and to share catchment characteristic data.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.02443/full#supplementary-material
Abeyta, C., Deeter, F. G., Kaysner, C. A., Stott, R. F., and Wekell, M. M. (1993). Campylobacter jejuni in a Washington state shellfish growing bed associated with illness. J. Food Prot. 56, 323–325. doi: 10.4315/0362-028X-56.4.323
AFNOR-NF-V-08-106:2010 (2010). Microbiologie des aliments: Dnombrement des E. coli présumés dans les coquillages vivants -Technique indirecte par impédancemétrie directe.
Amagliani, G., Brandi, G., and Schiavano, G. F. (2012). Incidence and role of Salmonella in seafood safety. Food Res. Int. 45, 780–788. doi: 10.1016/j.foodres.2011.06.022
Andrade, V. C., Zampieri, B. B., Ballesteros, E. R., Pinto, A. B., and de Oliveira, A. J. (2015). Densities and antimicrobial resistance of Escherichia coli isolated from marine waters and beach sands. Environ. Monit. Assess. 187:342. doi: 10.1007/s10661-015-4573-8
Atmar, R. L., Opekun, A. R., Gilger, M. A., Estes, M. K., Crawford, S. E., Neill, F. H., et al. (2014). Determination of the 50% human infectious dose for Norwalk virus. J. Infect. Dis. 209, 1016–1022. doi: 10.1093/infdis/jit620
Baker-Austin, C., Trinanes, J. A., Taylor, N. G. H., Hartnell, R., Siitonen, A., and Martinez-Urtaza, J. (2013). Emerging Vibrio risk at high latitudes in response to ocean warming. Nature Climate Change 3, 73–77. doi: 10.1038/nclimate1628
Balière, C., Rincé, A., Blanco, J., Dahbi, G., Harel, J., Vogeleer, P., et al. (2015). Prevalence and characterization of shiga toxin-producing and Enteropathogenic Escherichia coli in shellfish-harvesting areas and their watersheds. Front. Microbiol. 6:1356. doi: 10.3389/fmicb.2015.01356
Boehm, A. B., and Sassoubre, L. M. (2014). “Enterococci as indicators of environmental fecal contamination,” in Enterococci: From Commensals to Leading Causes of Drug Resistant Infection, eds M. S. Gilmore, D. B. Clewell, Y. Ike, and N. Shankar (Boston, MA: Massachusetts Eye and Ear Infirmary).
Boehm, A. B., Soetjipto, C., and Wang, D. (2012). Solar inactivation of four Salmonella serovars in fresh and marine waters. J. Water Health 10, 504–510. doi: 10.2166/wh.2012.084
Bonacorsi, S., Houdouin, V., Mariani-Kurkdjian, P., Mahjoub-Messai, F., and Bingen, E. (2006). Comparative prevalence of virulence factors in Escherichia coli causing urinary tract infection in male infants with and without bacteremia. J. Clin. Microbiol. 44, 1156–1158. doi: 10.1128/JCM.44.3.1156-1158.2006
Brands, D. A., Inman, A. E., Gerba, C. P., Maré, C. J., Billington, S. J., Saif, L. A., et al. (2005). Prevalence of Salmonella spp. in oysters in the United States. Appl. Environ. Microbiol. 71, 893–897. doi: 10.1128/AEM.71.2.893-897.2005
Clermont, O., Bonacorsi, S., and Bingen, E. (2000). Rapid and simple determination of the Escherichia coli phylogenetic group. Appl. Environ. Microbiol. 66, 4555–4558. doi: 10.1128/AEM.66.10.4555-4558.2000
Collin, B., and Rehnstam-Holm, A.-S. (2011). Occurrence and potential pathogenesis of Vibrio cholerae, Vibrio parahaemolyticus and Vibrio vulnificus on the South coast of Sweden. FEMS Microbiol. Ecol. 78, 306–313. doi: 10.1111/j.1574-6941.2011.01157.x
David, J. M., Sanders, P., Bemrah, N., Granier, S. A., Denis, M., Weill, F.-X., et al. (2013). Attribution of the French human Salmonellosis cases to the main food-sources according to the type of surveillance data. Prev. Vet. Med. 110, 12–27. doi: 10.1016/j.prevetmed.2013.02.002
de Graaf, M., van Beek, J., and Koopmans, M. P. G. (2016). Human norovirus transmission and evolution in a changing world. Nat. Rev. Microbiol. 14, 421–433. doi: 10.1038/nrmicro.2016.48
Denis, M., Henrique, E., Chidaine, B., Tircot, A., Bougeard, S., and Fravalo, P. (2011a). Campylobacter from sows in farrow-to-finish pig farms: risk indicators and genetic diversity. Vet. Microbiol. 154, 163–170. doi: 10.1016/j.vetmic.2011.07.001
Denis, M., Tanguy, M., Chidaine, B., Laisney, M.-J., Mégraud, F., and Fravalo, P. (2011b). Description and sources of contamination by Campylobacter spp. of river water destined for human consumption in Brittany, France. Pathologie Biologie 59, 256–263. doi: 10.1016/j.patbio.2009.10.007
DePaola, A., Jones, J. L., Woods, J., Burkhardt, W., Calci, K. R., Krantz, J. A., et al. (2010). Bacterial and viral pathogens in live oysters: 2007 United States market survey. Appl. Environ. Microbiol. 76, 2754–2768. doi: 10.1128/AEM.02590-09
Deter, J., Lozach, S., Derrien, A., Véron, A., Chollet, J., and Hervio-Heath, D. (2010). Chlorophyll a might structure a community of potentially pathogenic culturable Vibrionaceae. Insights from a one-year study of water and mussels surveyed on the French Atlantic coast. Environ. Microbiol. Rep. 2, 185–191. doi: 10.1111/j.1758-2229.2010.00133.x
Escobedo-Hinojosa, W., and Pardo-López, L. (2017). Analysis of bacterial metagenomes from the Southwestern Gulf of Mexico for pathogens detection. Pathog. Dis. 75:ftx058. doi: 10.1093/femspd/ftx058
Esteves, K., Hervio-Heath, D., Mosser, T., Rodier, C., Tournoud, M.-G., Jumas-Bilak, E., et al. (2015). Rapid proliferation of Vibrio parahaemolyticus, Vibrio vulnificus, and Vibrio cholerae during freshwater flash floods in French Mediterranean coastal lagoons. Appl. Environ. Microbiol. 81, 7600–7609. doi: 10.1128/AEM.01848-15
European Food Safety Authority [EFSA] and European Centre for Disease Prevention Control [ECDC] (2015). The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2014. EFSA J. 13, 1–190. doi: 10.2903/j.efsa.2015.4329
European Food Safety Authority [EFSA] and European Centre for Disease Prevention Control [ECDC] (2016). The European Union summary report on trends and sources of zoonoses, zoonotic agents and food-borne outbreaks in 2015. EFSA J. 14, 1–231. doi: 10.2903/j.efsa.2016.4634
Fabre, L., Zhang, J., Guigon, G., Le Hello, S., Guibert, V., Accou-Demartin, M., et al. (2012). CRISPR typing and subtyping for improved laboratory surveillance of Salmonella infections. PLoS One 7:e36995. doi: 10.1371/journal.pone.0036995
Flockhart, L., Pintar, K., Cook, A., McEwen, S., Friendship, R., Kelton, D., et al. (2017). Distribution of Salmonella in humans, production animal operations and a watershed in a FoodNet Canada Sentinel Site. Zoonoses Public Health 64, 41–52. doi: 10.1111/zph.12281
Fonteneau, L., Jourdan Da Silva, N., Fabre, L., Ashton, P., Torpdahl, M., Müller, L., et al. (2017). Multinational outbreak of travel-related Salmonella Chester infections in Europe, summers 2014 and 2015. Euro Surveill. 22:30463. doi: 10.2807/1560-7917.ES.2017.22.7.30463
Furtula, V., Jackson, C. R., Farrell, E. G., Barrett, J. B., Hiott, L. M., and Chambers, P. A. (2013). Antimicrobial resistance in Enterococcus spp. isolated from environmental samples in an area of intensive poultry production. Int. J. Environ. Res. Public Health 10, 1020–1036. doi: 10.3390/ijerph10031020
Gorski, L., Parker, C. T., Liang, A., Cooley, M. B., Jay-Russell, M. T., Gordus, A. G., et al. (2011). Prevalence, distribution, and diversity of Salmonella enterica in a major produce region of California. Appl. Environ. Microbiol. 77, 2734–2748. doi: 10.1128/AEM.02321-10
Grimont, P. A. D., and Weill, F. X. (2007). Antigenic Formulae of the Salmonella serovars, 9th Edn. Paris: WHO Collaborating Center for Reference and Research on Salmonella.
Guzman Prieto, A. M., van Schaik, W., Rogers, M. R. C., Coque, T. M., Baquero, F., Corander, J., et al. (2016). Global emergence and dissemination of Enterococci as nosocomial pathogens: attack of the Clones? Front. Microbiol. 7:788. doi: 10.3389/fmicb.2016.00788
Hayward, M. R., Petrovska, L., Jansen, V. A. A., and Woodward, M. J. (2016). Population structure and associated phenotypes of Salmonella enterica serovars Derby and Mbandaka overlap with host range. BMC Microbiol. 16:15. doi: 10.1186/s12866-016-0628-4
Hervio-Heath, D., Colwell, R. R., Derrien, A., Robert-Pillot, A., Fournier, J. M., and Pommepuy, M. (2002). Occurrence of pathogenic vibrios in coastal areas of France. J. Appl. Microbiol. 92, 1123–1135. doi: 10.1046/j.1365-2672.2002.01663.x
Hoszowski, A., Zająac, M., Lalak, A., Przemyk, P., and Wasyl, D. (2016). Fifteen years of successful spread of Salmonella enterica serovar Mbandaka clone ST413 in Poland and its public health consequences. Ann. Agric. Environ. Med. 23, 237–241. doi: 10.5604/12321966.1203883
ISO-7899-1:1998 (1998). Qualité de l’eau: Recherche et dénombrement des entérocoques intestinaux -Partie 1: Méthode miniaturisée (nombre le plus probable) pour les eaux de surface et résiduaires. Available at: https://www.iso.org/fr/standard/14852.html
ISO-9308-3:1998 (1998). Qualité de l’eau: Recherche et dénombrement des Escherichia coli et des bactéries coliformes -Partie 3: Méthode miniaturisée (nombre le plus probable) pour la recherche et le dénombrement des E. coli dans les eaux de surface et résiduaires. Available at: https://www.iso.org/fr/standard/20878.html
Jardé, E., Jeanneau, L., Harrault, L., Quenot, E., Solecki, O., Petitjean, P., et al. (2018). Application of a microbial source tracking based on bacterial and chemical markers in headwater and coastal catchments. Sci. Total Environ. 61, 55–63. doi: 10.1016/j.scitotenv.2017.07.235
Johnson, J. R., Delavari, P., Kuskowski, M., and Stell, A. L. (2001). Phylogenetic distribution of extraintestinal virulence-associated traits in Escherichia coli. J. Infect. Dis. 183, 78–88. doi: 10.1086/317656
Johnson, J. R., Porter, S. B., Johnston, B., Thuras, P., Clock, S., Crupain, M., et al. (2017). Extraintestinal pathogenic and antimicrobial-resistant Escherichia coli, including sequence type 131 (ST131), from retail chicken breasts in the United States in 2013. Appl. Environ. Microbiol. 83:e02956-16. doi: 10.1128/AEM.02956-16
Jokinen, C. C., Koot, J., Cole, L., Desruisseau, A., Edge, T. A., Khan, I. U. H., et al. (2015). The distribution of Salmonella enterica serovars and subtypes in surface water from five agricultural regions across Canada. Water Res. 76, 120–131. doi: 10.1016/j.watres.2015.02.038
Jones, K. (2001). Campylobacters in water, sewage and the environment. Symp. Ser. Soc. Appl. Microbiol. 30, 68S–79S. doi: 10.1046/j.1365-2672.2001.01355.x
Kaper, J. B., Nataro, J. P., and Mobley, H. L. (2004). Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2, 123–140. doi: 10.1038/nrmicro818
Khan, I. U., Gannon, V., Jokinen, C. C., Kent, R., Koning, W., Lapen, D. R., et al. (2014). A national investigation of the prevalence and diversity of thermophilic Campylobacter species in agricultural watersheds in Canada. Water Res. 61, 243–252. doi: 10.1016/j.watres.2014.05.027
Kreft, B., Marre, R., Schramm, U., and Wirth, R. (1992). Aggregation substance of Enterococcus faecalis mediates adhesion to cultured renal tubular cells. Infect. Immun. 60, 25–30.
Lanthier, M., Scott, A., Zhang, Y., Cloutier, M., Durie, D., Henderson, V. C., et al. (2011). Distribution of selected virulence genes and antibiotic resistance in Enterococcus species isolated from the South Nation River drainage basin, Ontario, Canada. J. Appl. Microbiol. 110, 407–421. doi: 10.1111/j.1365-2672.2010.04893.x
Le Mennec, C., Parnaudeau, S., Rumebe, M., Le Saux, J.-C., Piquet, J.-C., and Le Guyader, F. S. (2017). Follow-up of norovirus contamination in an oyster production area linked to repeated outbreaks. Food Environ. Virol. 9, 54–61. doi: 10.1007/s12560-016-9260-6
Leblanc-Maridor, M., Beaudeau, F., Seegers, H., Denis, M., and Belloc, C. (2011). Rapid identification and quantification of Campylobacter coli and Campylobacter jejuni by real-time PCR in pure cultures and in complex samples. BMC Microbiol. 11:113. doi: 10.1186/1471-2180-11-113
Leight, A. K., Crump, B. C., and Hood, R. R. (2018). Assessment of fecal indicator bacteria and potential pathogen co-occurrence at a shellfish growing area. Front. Microbiol. 9:384. doi: 10.3389/fmicb.2018.00384
Li, D., Breiman, A., le Pendu, J., and Uyttendaele, M. (2015). Binding to histo-blood group antigen-expressing bacteria protects human norovirus from acute heat stress. Front. Microbiol. 6:659. doi: 10.3389/fmicb.2015.00659
Martelli, F., Andres, V. M., Davies, R., and Smith, R. P. (2018). Observations on the introduction and dissemination of Salmonella in three previously low prevalence status pig farms in the United Kingdom. Food Microbiol. 71, 129–134. doi: 10.1016/j.fm.2017.05.004
Martinez-Urtaza, J., Blanco-Abad, V., Rodriguez-Castro, A., Ansede-Bermejo, J., Miranda, A., and Rodriguez-Alvarez, M. X. (2012). Ecological determinants of the occurrence and dynamics of Vibrio parahaemolyticus in offshore areas. ISME J. 6, 994–1006. doi: 10.1038/ismej.2011.156
Martinez-Urtaza, J., Saco, M., de Novoa, J., Perez-Piñeiro, P., Peiteado, J., Lozano-Leon, A., et al. (2004). Influence of environmental factors and human activity on the presence of Salmonella serovars in a marine environment. Appl. Environ. Microbiol. 70, 2089–2097. doi: 10.1128/AEM.70.4.2089-2097.2004
Miller, W. G., Yee, E., Chapman, M. H., Smith, T. P. L., Bono, J. L., Huynh, S., et al. (2014). Comparative genomics of the Campylobacter lari group. Genome Biol. Evol. 6, 3252–3266. doi: 10.1093/gbe/evu249
Miura, T., Sano, D., Suenaga, A., Yoshimura, T., Fuzawa, M., Nakagomi, T., et al. (2013). Histo-blood group antigen-like substances of human enteric bacteria as specific adsorbents for human noroviruses. J. Virol. 87, 9441–9451. doi: 10.1128/JVI.01060-13
Miura, T., Schaeffer, J., Le Saux, J.-C., Le Mehaute, P., and Le Guyader, F. S. (2017). Virus type-specific removal in a full-scale membrane bioreactor treatment process. Food Environ. Virol. 10, 176–186. doi: 10.1007/s12560-017-9330-4
Obiri-Danso, K., and Jones, K. (1999). Distribution and seasonality of microbial indicators and thermophilic campylobacters in two freshwater bathing sites on the River Lune in northwest England. J. Appl. Microbiol. 87, 822–832. doi: 10.1046/j.1365-2672.2001.01239.x
Obiri-Danso, K., Paul, N., and Jones, K. (2001). The effects of UVB and temperature on the survival of natural populations and pure cultures of Campylobacter jejuni, Camp. coli, Camp. lari and urease-positive thermophilic campylobacters (UPTC) in surface waters. J. Appl. Microbiol. 90, 256–267. doi: 10.4081/ijfs.2016.5709
Passalacqua, P. L., Zavatta, E., Bignami, G., Serraino, A., and Serratore, P. (2016). Occurrence of Vibrio parahaemolyticus, Vibrio cholerae and Vibrio vulnificus in the clam Ruditapes Philippinarum (Adams & Reeve, 1850) from Emilia Romagna and Sardinia, Italy. Ital. J. Food Saf. 5:5709. doi: 10.4081/ijfs.2016.5709
Picard, B., Garcia, J. S., Gouriou, S., Duriez, P., Brahimi, N., Bingen, E., et al. (1999). The link between phylogeny and virulence in Escherichia coli extraintestinal infection. Infect. Immun. 67, 546–553.
Pitkanen, T., and Hanninen, M. L. (2017). Members of the family Campylobacteraceae: Campylobacter jejuni, Campylobacter coli. Global Water Pathogen Project. Available at: http://www.waterpathogens.org/book/campylobacter doi: 10.3201/eid2212.160841
Polo, D., Schaeffer, J., Fournet, N., Le Saux, J.-C., Parnaudeau, S., McLeod, C., et al. (2016). Digital PCR for quantifying norovirus in oysters implicated in outbreaks, France. Emerg. Infect. Dis. 22, 2189–2191. doi: 10.3201/eid2212.160841
Potasman, I., Paz, A., and Odeh, M. (2002). Infectious outbreaks associated with bivalve shellfish consumption: a worldwide perspective. Clin. Infect. Dis. 35, 921–928. doi: 10.1086/342330
Powell, A., Baker-Austin, C., Wagley, S., Bayley, A., and Hartnell, R. (2013). Isolation of pandemic Vibrio parahaemolyticus from UK water and shellfish produce. Microb. Ecol. 65, 924–927. doi: 10.1007/s00248-013-0201-8
Ribot, E. M., Fair, M. A., Gautom, R., Cameron, D. N., Hunter, S. B., Swaminathan, B., et al. (2006). Standardization of pulsed-field gel electrophoresis protocols for the subtyping of Escherichia coli O157:H7, Salmonella, and Shigella for PulseNet. Foodborne Pathog. Dis. 3, 59–67. doi: 10.1089/fpd.2006.3.59
Rodriguez, S., and Araujo, R. (2012). Effect of environmental parameters on the inactivation of the waterborne pathogen Campylobacter in a Mediterranean river. J. Water Health 10, 100–107. doi: 10.2166/wh.2011.044
Rodriguez-Castro, A., Ansede-Bermejo, J., Blanco-Abad, V., Varela-Pet, J., Garcia-Martin, O., and Martinez-Urtaza, J. (2010). Prevalence and genetic diversity of pathogenic populations of Vibrio parahaemolyticus in coastal waters of Galicia. Spain Environ. Microbiol. Rep. 2, 58–66. doi: 10.1111/j.1758-2229.2009.00064.x
Ryu, H., Grond, K., Verheijen, B., Elk, M., Buehler, D. M., and Santo Domingo, J. W. (2014). Intestinal microbiota and species diversity of Campylobacter and Helicobacter spp. in migrating shorebirds in Delaware Bay. Appl. Environ. Microbiol. 80, 1838–1847. doi: 10.1128/AEM.03793-13
Schaeffer, J., Le Saux, J.-C., Lora, M., Atmar, R. L., and Le Guyader, F. S. (2013). Norovirus contamination on French marketed oysters. Int. J. Food Microbiol. 166, 244–248. doi: 10.1016/j.ijfoodmicro.2013.07.022
Schaeffer, J., Treguier, C., Piquet, J.-C., Gachelin, S., Cochennec-Laureau, N., Le Saux, J.-C., et al. (2018). Improving the efficacy of sewage treatment decreases norovirus contamination in oysters. Int. J. Food Microbiol. 286, 1–5. doi: 10.1016/j.ijfoodmicro.2018.07.016
Semenza, J. C., Trinanes, J., Lohr, W., Sudre, B., Löfdahl, M., Martinez-Urtaza, J., et al. (2017). Environmental suitability of Vibrio Infections in a warming climate: an Early Warning System. Environ. Health Perspect. 125:107004. doi: 10.1289/EHP2198
Setti, I., Rodriguez-Castro, A., Pata, M. P., Cadarso-Suarez, C., Yacoubi, B., Bensmael, L., et al. (2009). Characteristics and dynamics of Salmonella contamination along the coast of Agadir, Morocco. Appl. Environ. Microbiol. 75, 7700–7709. doi: 10.1128/AEM.01852-09
Shankar, V., Baghdayan, A. S., Huycke, M. M., Lindahl, G., and Gilmore, M. S. (1999). Infection-derived Enterococcus faecalis strains are enriched in esp, a gene encoding a novel surface protein. Infect. Immun. 67, 193–200. doi: 10.1021/es4050083
Sidhu, J., Skelly, E., Hodgers, L., Ahmed, W., Li, Y., and Toze, S. (2014). Prevalence of enterococcus species and their virulence genes in fresh water prior to and after storm events. Environ. Sci. Technol. 48, 2979–2988. doi: 10.1021/es4050083
Soares, R. O., Fedi, A. C., Reiter, K. C., Caierão, J., and d’Azevedo, P. A. (2014). Correlation between biofilm formation and gelE, esp, and agg genes in Enterococcus spp. clinical isolates. Virulence 5, 634–637. doi: 10.4161/viru.28998
Stange, C., Sidhu, J. P. S., Tiehm, A., and Toze, S. (2016). Antibiotic resistance and virulence genes in coliform water isolates. Int. J. Hyg. Environ. Health 219, 823–831. doi: 10.1016/j.ijheh.2016.07.015
Su, Y. A., Sulavik, M. C., He, P., Makinen, K. K., Makinen, P. L., Fiedler, S., et al. (1991). Nucleotide sequence of the gelatinase gene (gelE) from Enterococcus faecalis subsp. liquefaciens. Infect. Immun. 59, 415–420. doi: 10.1016/j.ijfoodmicro.2014.04.016
Suffredini, E., Mioni, R., Mazzette, R., Bordin, P., Serratore, P., Fois, F., et al. (2014). Detection and quantification of Vibrio parahaemolyticus in shellfish from Italian production areas. Int. J. Food Microbiol. 184, 14–20. doi: 10.1016/j.ijfoodmicro.2014.04.016
Sullender, M. E., and Baldridge, M. T. (2018). Norovirus interactions with the commensal microbiota. PLoS Pathog. 14:e1007183. doi: 10.1371/journal.ppat.1007183
Thebault, A., Teunis, P. F. M., Le Pendu, J., Le Guyader, F. S., and Denis, J.-B. (2013). Infectivity of GI and GII noroviruses established from oyster related outbreaks. Epidemics 5, 98–110. doi: 10.1016/j.epidem.2012.12.004
Van Cauteren, D., Le Strat, Y., Sommen, C., Bruyand, M., Tourdjman, M., Da Silva, N. J., et al. (2017). Estimated annual numbers of foodborne pathogen-associated Illnesses, hospitalizations, and deaths, France, 2008-2013. Emerg. Infect. Dis. 23, 1486–1492. doi: 10.3201/eid2309.170081
Van Dyke, M. I., Morton, V. K., McLellan, N. L., and Huck, P. M. (2010). The occurrence of Campylobacter in river water and waterfowl within a watershed in southern Ontario, Canada. J. Appl. Microbiol. 109, 1053–1066. doi: 10.1111/j.1365-2672.2010.04730.x
Veljović, K., Popović, N., Vidojević, A. T., Tolinaèki, M., Mihajlović, S., Jovèić, B., et al. (2015). Environmental waters as a source of antibiotic-resistant Enterococcus species in Belgrade, Serbia. Environ. Monit. Assess. 187:599. doi: 10.1007/s10661-015-4814-x
Vereen, E., Lowrance, R. R., Jenkins, M. B., Adams, P., Rajeev, S., and Lipp, E. K. (2013). Landscape and seasonal factors influence Salmonella and Campylobacter prevalence in a rural mixed use watershed. Water Res. 47, 6075–6085. doi: 10.1016/j.watres.2013.07.028
Vignaroli, C., Di Sante, L., Leoni, F., Chierichetti, S., Ottaviani, D., Citterio, B., et al. (2016). Multidrug-resistant and epidemic clones of Escherichia coli from natural beds of Venus clam. Food Microbiol. 59, 1–6. doi: 10.1016/j.fm.2016.05.003
Walters, S. P., González-Escalona, N., Son, I., Melka, D. C., Sassoubre, L. M., and Boehm, A. B. (2013). Salmonella enterica diversity in central Californian coastal waterways. Appl. Environ. Microbiol. 79, 4199–4209. doi: 10.1128/AEM.00930-13
Wilkes, G., Edge, T. A., Gannon, V. P. J., Jokinen, C., Lyautey, E., Neumann, N. F., et al. (2011). Associations among pathogenic bacteria, parasites, and environmental and land use factors in multiple mixed-use watersheds. Water Res. 45, 5807–5825. doi: 10.1016/j.watres.2011.06.021
Wilson, I. G., and Moore, J. E. (1996). Presence of Salmonella spp. and Campylobacter spp. in shellfish. Epidemiol. Infect. 116, 147–153.
Yoder, J. S., Hlavsa, M. C., Craun, G. F., Hill, V., Roberts, V., Yu, P. A., et al. (2008). Surveillance for waterborne disease and outbreaks associated with recreational water use and other aquatic facility-associated health events–United States, 2005-2006. MMWR Surveill. Summ. 57, 1–29. doi: 10.1128/AEM.01729-15
Keywords: Campylobacter, Salmonella, Vibrio, HuNoVs, fecal bacterial indicators, shellfish, water
Citation: Rincé A, Balière C, Hervio-Heath D, Cozien J, Lozach S, Parnaudeau S, Le Guyader FS, Le Hello S, Giard J-C, Sauvageot N, Benachour A, Strubbia S and Gourmelon M (2018) Occurrence of Bacterial Pathogens and Human Noroviruses in Shellfish-Harvesting Areas and Their Catchments in France. Front. Microbiol. 9:2443. doi: 10.3389/fmicb.2018.02443
Received: 26 April 2018; Accepted: 24 September 2018;
Published: 11 October 2018.
Edited by:
Jean-Christophe Avarre, Institut de Recherche pour le Développement (IRD), FranceReviewed by:
Christopher Karl Yost, University of Regina, CanadaCopyright © 2018 Rincé, Balière, Hervio-Heath, Cozien, Lozach, Parnaudeau, Le Guyader, Le Hello, Giard, Sauvageot, Benachour, Strubbia and Gourmelon. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Alain Rincé, YWxhaW4ucmluY2VAdW5pY2Flbi5mcg==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.