 Fillipe L. R. do Carmo1,2,3
Fillipe L. R. do Carmo1,2,3 Wanderson M. Silva4,5
Wanderson M. Silva4,5 Guilherme C. Tavares6
Guilherme C. Tavares6 Izabela C. Ibraim1
Izabela C. Ibraim1 Barbara F. Cordeiro1
Barbara F. Cordeiro1 Emiliano R. Oliveira1
Emiliano R. Oliveira1 Houem Rabah2,3
Houem Rabah2,3 Chantal Cauty2
Chantal Cauty2 Sara H. da Silva1
Sara H. da Silva1 Marcus V. Canário Viana1
Marcus V. Canário Viana1 Ana C. B. Caetano1
Ana C. B. Caetano1 Roselane G. dos Santos1
Roselane G. dos Santos1 Rodrigo D. de Oliveira Carvalho7
Rodrigo D. de Oliveira Carvalho7 Julien Jardin2
Julien Jardin2 Felipe L. Pereira6
Felipe L. Pereira6 Edson L. Folador8
Edson L. Folador8 Yves Le Loir2,3
Yves Le Loir2,3 Henrique C. P. Figueiredo6
Henrique C. P. Figueiredo6 Gwénaël Jan2,3*†
Gwénaël Jan2,3*† Vasco Azevedo1†
Vasco Azevedo1†- 1Departamento de Biologia Geral, Instituto de Ciências Biológicas, Universidade Federal de Minas Gerais, Belo Horizonte, Brazil
- 2Institut National de la Recherche Agronomique, UMR1253 Science & Technologie du Lait & de l'Oeuf, Rennes, France
- 3Agrocampus Ouest, UMR1253 Science & Technologie du Lait & de l'Oeuf, Rennes, France
- 4Instituto de Biotecnología, CICVyA - Instituto Nacional de Tecnología Agropecuaria, Hurlingham, Argentina
- 5Consejo Nacional de Investigaciones Científicas y Técnicas, Buenos Aires, Argentina
- 6AQUACEN, Escola de Veterinária, Universidade Federal de Minas Gerais, Belo Horizonte, Brazil
- 7Departamento de Biointeração do Instituto de Ciências da Saúde, Universidade Federal da Bahia, Salvador, Brazil
- 8Centro de Biotecnologia, Universidade Federal da Paraíba, João Pessoa, Brazil
Propionibacterium freudenreichii is a beneficial Gram-positive bacterium, traditionally used as a cheese-ripening starter, and currently considered as an emerging probiotic. As an example, the P. freudenreichii CIRM-BIA 129 strain recently revealed promising immunomodulatory properties. Its consumption accordingly exerts healing effects in different animal models of colitis, suggesting a potent role in the context of inflammatory bowel diseases. This anti-inflammatory effect depends on surface layer proteins (SLPs). SLPs may be involved in key functions in probiotics, such as persistence within the gut, adhesion to host cells and mucus, or immunomodulation. Several SLPs coexist in P. freudenreichii CIRM-BIA 129 and mediate immunomodulation and adhesion. A mutant P. freudenreichii CIRM-BIA 129ΔslpB (CB129ΔslpB) strain was shown to exhibit decreased adhesion to intestinal epithelial cells. In the present study, we thoroughly analyzed the impact of this mutation on cellular properties. Firstly, we investigated alterations of surface properties in CB129ΔslpB. Surface extractable proteins, surface charges (ζ-potential) and surface hydrophobicity were affected by the mutation. Whole-cell proteomics, using high definition mass spectrometry, identified 1,288 quantifiable proteins in the wild-type strain, i.e., 53% of the theoretical proteome predicted according to P. freudenreichii CIRM-BIA 129 genome sequence. In the mutant strain, we detected 1,252 proteins, including 1,227 proteins in common with the wild-type strain. Comparative quantitative analysis revealed 97 proteins with significant differences between wild-type and mutant strains. These proteins are involved in various cellular process like signaling, metabolism, and DNA repair and replication. Finally, in silico analysis predicted that slpB gene is not part of an operon, thus not affecting the downstream genes after gene knockout. This study, in accordance with the various roles attributed in the literature to SLPs, revealed a pleiotropic effect of a single slpB mutation, in the probiotic P. freudenreichii. This suggests that SlpB may be at a central node of cellular processes and confirms that both nature and amount of SLPs, which are highly variable within the P. freudenreichii species, determine the probiotic abilities of strains.
Introduction
Probiotic bacteria are defined as “living microorganisms which when administered in adequate amounts confer a health benefit on the host” (Food and Agriculture Organization of the United Nations and World Health Organization, 2002). This term was further used by International Scientific Association for Probiotics and Prebiotics (ISAP) (Hill et al., 2014). Clinical proofs of efficiency were indeed obtained, in the context of antibiotic- and Clostridium difficile-associated diarrhea (Rondanelli et al., 2017), lactose intolerance (Oak and Jha, 2018), irritable bowel syndrome (IBS) (Ford et al., 2014), and ulcerative colitis, one of the disorders that constitute Inflammatory bowel disease (IBD) (Plaza-Díaz et al., 2017). The mechanisms underpinning these effects mainly belong to three categories: (i) metabolic effects, (ii) modulation of the gut microbiota, and (iii) probiotic/host molecular interactions. Although lactobacilli and bifidobacteria were mainly considered for probiotic usage, promising effects were also reported for dairy propionibacteria (Rabah et al., 2017).
The probiotic properties of dairy propionibacteria are strain-dependent and include microbiota modulation, apoptosis modulation in colonic cells and immunomodulation. Some of these probiotic abilities were validated at the clinical level. Microbiota modulation by dairy propionibacteria result in a bifidogenic effect (Roland et al., 1998; Seki et al., 2004; Suzuki et al., 2006). The corresponding molecular mechanisms were elucidated, and two molecules are shown to be involved in bifidogenic effect: 1,4-dihydroxy-2-naphtoic acid (DHNA) and 2-amino-3-carboxy-1,4-naphthoquinone (ACNQ) (Isawa et al., 2002; Furuichi et al., 2006). The pro-apoptotic effect of dairy propionibacteria was evidenced using in vitro cellular models (Jan et al., 2002) and animals models (Lan et al., 2008). This effect is mainly due to the production of the short chain fatty acids (SCFA) acetate and propionate by dairy propionibacteria (Lan et al., 2007; Cousin et al., 2016). The anti-inflammatory effect was suggested in IBD patients (Mitsuyama et al., 2007) and confirmed in animal colitis models (Foligné et al., 2010; Plé et al., 2015, 2016). Immunomodulatory properties are due to several metabolites as SCFAs and to cells wall component (Rabah et al., 2017). Indeed, surface proteins considered as microorganism-associated molecular patterns (MAMP) play a pivotal role in interaction with host's immune system (Deutsch et al., 2012; Le Maréchal et al., 2015). This includes SlpB and SlpE, surface proteins anchored to the cell wall via surface-layer homology (SLH) domains (Deutsch et al., 2017; do Carmo et al., 2018). Indeed, mutation of slpB and slpE genes clearly affected the immunomodulatory properties of P. freudenreichii (Deutsch et al., 2017). We have recently shown that SlpB is involved both in immunomodulation and in adhesion to cultured human intestinal epithelial cells (do Carmo et al., 2017).
In probiotic bacteria, extractable surface proteins play several role in bacterium/host interaction, protection against environmental stresses, inhibition of pathogens, survival within the host digestive tract, and determination or maintenance of cell shape (Hynönen and Palva, 2013; do Carmo et al., 2018). In this study, we investigated the impact of slpB gene mutation on the physiology of P. freudenreichii CIRM-BIA 129 using a proteomic approach. In this purpose, we investigated alterations in extractable surface proteins and in the whole-cell proteome. We compared wild-type CIRM-BIA 129 with mutant CB129ΔslpB. We report pleiotropic effects of this single mutation on physicochemical properties of this propionibacteria.
Materials and Methods
Bacterial Strains and Culture Conditions
The wild-type P. freudenreichii CIRM-BIA 129 (WT) strain and genetically modified P. freudenreichii CIRM-BIA 129ΔslpB strain (CB129ΔslpB) (do Carmo et al., 2017) were grown at 30°C in Yeast Extract Lactate (YEL) broth (Malik et al., 1968). For the CB129ΔslpB, YEL culture media were supplemented with chloramphenicol (10 μg.mL−1). The growth of P. freudenreichii strains was monitored spectrophotometrically by measuring the optical density at 650 nm (OD650 nm), as well as by counting colony-forming units (CFUs) in YEL medium containing 1.5% agar. P. freudenreichii strains were harvested in a stationary phase (76 h, 2 × 109 CFU.mL−1, determined by plate counts) by centrifugation (8,000 × g, 10 min, 4°C).
Inventory of Extractable Surface Proteins Using Guanidine Hydrochloride and MS/MS
Proteins were guanidine-extracted, trypsinolysed and subjected to mass spectrometry (Le Maréchal et al., 2015). Peptides were separated by Nano-LC-MS/MS using a Dionex U3000-RSLC nano-LC system fitted to a Qexactive mass spetrometer (Thermo Scientific, San Jose, CA, USA) equipped with a nano-electrospray ion source (ESI) (Proxeon Biosystems A/S, Odense, Denmark). Peptides were identified from MS/MS spectra using the X!Tandem pipeline 3.4.3 software (Langella et al., 2017) for search into two concatenated databases: (i) a homemade database containing all the predicted proteins of the P. freudenreichii CIRM-BIA 129 used in this study and (ii) a portion of the UniProtKB database corresponding to taxonomy 754252: P. freudenreichii subsp. shermanii (strain ATCC 9614/CIP 103027/CIRM-BIA1).
Zeta Potential Analysis
Electrophoretic mobility (zeta potential) was determined according to the well-described protocol of Schär-Zammaretti and Ubbink (2003). Bacteria were harvested from a 5 mL stationary phase culture by centrifugation (8.000 × g, 10 min, room temperature) and washed twice with a PBS buffer pH 7.0. Cell count of the final suspensions was approximately 108 CFU/ml. The pellet was resuspended in a 10 mM KH2PO4 solution (pH 7.0). The electrophoretic mobility was measured by using a ZetaSizer nanoZS (Malvern Instruments, Malvern, United Kingdom) and a glass capillary Zetasizer Nanoseries DTS 1061 (Malvern Instruments, Malvern, United Kingdom) as the electrophoretic cell. Electrophoretic mobilities were converted to the ζ-potential using the Helmholtz-Smoluchowski equation (Schär-Zammaretti and Ubbink, 2003). All experiments were done in biological and technical triplicates.
Cell Surface Hydrophobicity Analysis
The Microbial Adhesion To Hydrocarbons (MATH) assay was performed as described by Kos et al. (2003). The optical density of the stationary phase bacteria was adjusted to an OD650 nm = 1. The samples were centrifuged for 5 min, 10,000 × g at room temperature and the pellets washed twice with the same volume of PBS pH 7.0 prior to resuspension in 15 mL of 0.1M KNO3, pH 6.2. An aliquot of each bacterial suspension (4 ml) was mixed with 1 mL of the solvent (Xylene, chloroform and ethyl acetate), incubated for 5 min at room temperature and mixed by vortex during 120 s. Subsequently, samples were incubated during 60 min to allow phases separation, the aqueous phase was carefully removed and absorbance (OD600 nm) was determined as above. Cell surface hydrophobicity in terms of per cent (H %) was calculated using the following formula: H % = (1–A1/A0) × 100. All experiments were done in biological and technical triplicates.
Transmission Electron Microscopy Assay
Cultures were grown on YEL medium to an OD650 nm of 1. Transmission electron microscopy was executed after bacteria were washed with PBS and fixed overnight at 4°C in 0.1 M sodium cacodylate buffer (pH 7.2) containing 2% glutaraldehyde. Fixed bacteria were rinsed and stored at 4°C in cacodylate buffer containing 0.2 M sucrose. They were then postfixed with 1% osmium tetroxide containing 1.5% potassium cyanoferrate and 2% uranyl acetate in water before gradual dehydration in ethanol (30% to 100%) and embedding in Epon. Thin sections (70 nm) were collected on 200-mesh cooper grids and counterstained with lead citrate before examination. The thickness of the cell wall was determined using the imageJ software in both strains analyzed by Transmission Electron Microscopy (TEM) as described (Foligné et al., 2010; Deutsch et al., 2012).
Stress Conditions Challenge
P. freudenreichii strains in stationary phase were subjected to lethal doses of different stresses. The acid challenge was carried out at pH 2.0 for 1 h as described (Jan et al., 2000). The bile salts stress was induced by adding 1.0 g/l of bile salts for 60 s as described (Leverrier et al., 2003). For the thermic stress, bacteria were heated for 30 min at 63°C. Viable cells were determined by serial dilutions of samples made up in peptone water (0.1% bacteriological peptone, Kasvi, Brazil), adjusted to pH 7.0 and containing 0.9% NaCl, into YEL medium containing 1.5% agar. CFU were counted after 6 days of anoxic incubation at 30°C (Anaerocult® A - Merck Millipore). All experiments were done in biological and technical triplicates.
Whole-Cell Protein Extraction and Preparation of Total Bacterial Lysates
The optical density of the stationary phase bacteria was adjusted to an OD650 nm = 1. The cultures were centrifuged for 5 min, 10,000 × g at room temperature and the bacterial pellets from biological triplicates were resuspended in 1 mL of lysis buffer containing 42% urea, 15% thiourea, 4% SDC (sodium deoxycholate), 12.5 mM Tris-HCl pH 7.5 and 1.5% dithiothreitol (DTT) with 10 μL of protease inhibitor (GE HealthCare, Pittsburgh, USA). Next, whole-cell proteins were extracted as described (Silva et al., 2014) and quantified by Qubit 2.0 fluorometer (Invitrogen, Carlsbad, USA). 100 μg of each protein extract were denatured with 0.2% of RapiGest SF solution (Waters, Milford, USA) at 80°C for 15 min, reduced with 100 mM DTT at 60°C for 30 min, and alkylated with 300 mM iodoacetamide at room temperature in a dark room for 30 min (Leibowitz et al., 2017). Subsequently, proteins were enzymatically digested with 10 μl of trypsin at 0.5 μg.μL−1 (Promega, Madison, USA), and the digestion stopped with the addition of 10 μL of 5% trifluoroacetic acid (TFA) (Sigma Aldrich, Saint Louis, USA) (Silva et al., 2017). Tryptic peptides were subjected to SDC removal (Lin et al., 2010), desalted using C18 MacroSpin Columns (Harvard Apparatus, Holliston, USA), according to the manufacturer's instructions, and dried under vacuum in the Eppendorf™ Vacufuge™ Concentrator (Eppendorf, Hamburg, Germany) (Wong et al., 2013). Prior to injection, the peptides were resuspended in 20 mM ammonium formate (Sigma Aldrich) and transferred to Waters Total Recovery vials (Waters).
LC-HDMSE Analysis and Data Processing
Quantitative proteomics analyses were conducted with Bidimensional Nano Ultra-Performance Liquid Chromatography (nanoUPLC) tandem Nano Electrospray High Definition Mass Spectrometry (nanoESI-HDMSE) both using a 1-h reverse-phase (RP) gradient from 7 to 40% (v/v) acetonitrile (0.1% v/v formic acid) and a 500 nL.min−1 nanoACQUITY UPLC 2D Technology system (Waters) (Gilar et al., 2005). A nanoACQUITY UPLC High Strength Silica (HSS) T3 1.8 μm, 75 μm × 150 mm column (pH 3) was used in conjunction with a RP Acquity UPLC Nano Ease XBridge BEH130 C18 5 μm, 300 μm × 50 mm nanoflow column (pH 10) (Silva et al., 2017). Typical on-column sample loads were 500 ng of total protein digests for each sample of the 5 fractions (500 ng per fraction/load).
The measurements for all samples by mass spectrometer was operated in resolution mode with a typical m/z resolving power of at least 25,000 Full Width at Half Maximum (FWHM) and an ion mobility cell that was filled with helium gas and a cross-section resolving power at least 40 Ω/Δ Ω. The effective resolution with the conjoined ion mobility was 25,000 FWHM. Analyses were performed using nano-electrospray ionization in positive ion mode nanoESI (+) and a NanoLock-Spray (Waters) ionization source. The lock mass channel was sampled every 30 s. The mass spectrometer was calibrated with an MS/MS spectrum of [Glu1]-Fibrinopeptide B human (Glu-Fib) solution (100 fmol.μL−1) that was delivered through the reference sprayer of the NanoLock-Spray source. The double-charged ion ([M + 2H]2+ = 785.8426) was used for initial single-point calibration, and MS/MS fragment ions of Glu-Fib were used to obtain the final instrument calibration.
The multiplexed data-independent acquisition (DIA) scanning with added specificity and selectivity of a non-linear “T-wave” ion mobility (HDMSE) device was performed with a Synapt G2-Si HDMS mass spectrometer (Waters) (Giles et al., 2011). Synapt G2-Si HDMS was automatically planned to switch between standard MS (3 eV) and elevated collision energies HDMSE (19–45 eV) applied to the transfer “T-wave” collision-induced dissociation cell with nitrogen gas. The trap collision cell was adjusted to 1 eV, using a millisecond scan time that was previously adjusted based on the linear velocity of the chromatographic peak that was delivered through nanoACQUITY UPLC (Waters). A minimum of 20 scan points was generated for each single peak, both in low-energy and high-energy transmission at an orthogonal acceleration time-of-flight (oa-TOF) and a mass range from m/z 50 to 2,000.
Mass spectrometric analysis of tryptic peptides was performed using a mass spectrometer equipped with a T-Wave-IMS device (Waters) in MSE mode following the method previously described (Distler et al., 2014). Stoichiometric measurements based on scouting runs of the integrated total ion account prior to analysis were performed to ensure standardized molar values across all samples. Therefore, the tryptic peptides of each strain were injected with the same amount on the column. The radio frequency (RF) offset (MS profile) was adjusted such that the nanoESI-HDMSE data were effectively acquired from m/z 400 to 2000, which ensured that any masses less than m/z 400 that were observed in the high energy spectra with arose from dissociations in the collision cell (Silva et al., 2017).
The mass spectrometry proteomics data have been deposited to the ProteomeXchange Consortium via the PRIDE (Vizcaíno et al., 2016) partner repository with the dataset identifier PXD009804.
Proteins Identification and Quantification
HDMSE raw data were processed using Progenesis QI for Proteomics (QIP) v.2.0 (Nonlinear Dynamics, Newcastle, UK) as described by Kuharev et al. (2015). For proteins identification, the peptides were searching against a P. freudenreichii strain CIRM-BIA 129 database as described above. The reversed sequences were joined together to the original sequences using ProteinLynx Global Server (PLGS) v 3.0.2 (Waters) database management tool. The reversed sequences were used to calculate the false positive rate during identification process. Next, the following parameters were used for peptide identification: digest reagent = trypsin; maximum missed cleavage = one; maximum protein mass = 600 kDa; modifications: carbamidomethyl of cysteine (fixed), acetyl N-terminal (variable), phosphoryl (variable), oxidation of methionine (variable); search tolerance parameters: peptide tolerance = 10 ppm, fragment tolerance = 20 ppm, maximum false discovery rate (FDR) = 1%.
The protein-level quantitation was performed with Relative Quantitation using Hi-N algorithm. Proteins identified with at least two peptides and presents in at least two of the three biological replicates were considered (Silva et al., 2014). The proteins list was exported by the function “export protein measurements” and was used to subsequent bioinformatics analysis. Proteins were considered to be differentially expressed between mutant and wild type if there were a significant (p < 0,05, ANOVA) change in expression ≥ 2-fold (log2 ratio ≥ 1.0). A volcano plot was generated to visualize the differentially expressed proteins across these strains.
Extraction of Genomic DNA of the CB129ΔslpB Strain
Genomic DNA was extracted from CB129ΔslpB culture grown in YEL medium supplemented with chloramphenicol (10 μg ml−1), during the phase (76 h at 30°C). Samples was centrifuged at 4°C and 8,000 × g for 10 min. Bacterial pellets were resuspended in 1 ml Tris/EDTA/RNase [10 mM Tris/HCl (pH 7.0), 10 mM EDTA (pH 8.0), 300 mM NaCl, 50 μg RNase A ml−1] with 50 mg of Glass beads VK01 and cell lysis occurred in Precellys®24 by 2 cylces of 15 s at 6,500 rpm. DNA was purified using phenol/chloroform/isoamyl alcohol and precipitated with ethanol according with Sambrook and Russell (2001). DNA concentrations were determined spectrophotometrically in Thermo Scientific NanoDrop 1000.
Genome Sequencing, Assembly and Annotation of the CB129ΔslpB Strain
CB129ΔslpB strain sequencing libraries were constructed using 100 ng of genomic DNA. The gDNA was sheared with the Ion Shear™ Plus Reagents Kit and barcoded using the Ion Xpress Fragment Library kit and Ion Xpress™ Barcode Adapters (Life Technologies, USA), according to the manufacturer's protocol. Size selection of ~400 bp was performed with 2% E-Gel® SizeSelect™ Agarose Gels (Invitrogen, USA) and quantified with the Ion Library Quantitation Kit. The libraries were amplified with the OneTouch Template 400 kit on the Ion One Touch™ 2 (Life Technologies) and enriched on the Ion OneTouch™ ES (Life Technologies). Genomic libraries were enriched using Ion PI™ Hi-Q™ Sequencing Polymerase in the Ion 318™ v2 Chip, according to the manufacturer's protocols, and they were sequenced using Ion Torrent Personal Genome Machine (PGM). The amplification processes were performed using Ion PGM™ Hi-Q™ Sequencing 400 Polymerase with required 1,100 flows. Finally, signal processing was performed using Torrent Suite 4.2.1 to conclude the sequencing process.
De novo assembly was conducted using the software Newbler v 2.9 (Roche 454, USA). The assembled contigs were oriented to generate a scaffold using CONTIGuator v 2.7 (Galardini et al., 2011) and the strains P. freudenreichii CIRM-BIA 1 (FN806773.1) and P. freudenreichii JS17 (LT618789) as reference. The P. freudenreichii CIRM-BIA 1 strain (without the slpB gene) was used for comparative analysis as it is a reference from INRA strain collection strain and P. freudenreichii JS17 strain was used due to the presence of the s-layer gene slpB. CLC Genomics Workbench 7.0 (Qiagen, USA) was used to map the raw reads against the reference genome and to generate the consensus sequence used to the gap filling. The plasmid that integrated within and disrupted the slpB gene was not found in the scaffold, but its sequence was found within the contigs that were excluded during the scaffold generation. It was manually inserted to the scaffold by mapping its ends on the slpB gene and using the overlap sequences as coordinates for the insertion. The insertion was validated by mapping the reads on the assembly and checking for mismatches on the regions flanking the plasmid. The genome of CB129ΔslpB strain was annotated automatically using RAST pipeline (Aziz et al., 2008; Brettin et al., 2015).
Bioinformatics Analyses
The predicted proteins of CB129ΔslpB and WT strain were analyzed using the SurfG+ v1.0 tool (Barinov et al., 2009) to predict sub-cellular localization. It enabled the classification of proteins within the following categories: cytoplasmic (CYT), membrane (MEM), potentially surface-exposed (PSE) and secreted (SEC). The prediction of orthologous groups by functional category the sequences was performed using Cluster of Orthologous Genes (COG) database version 2014db (Galperin et al., 2015). The COG database search was performed using an in-house script (available at https://github.com/aquacen/blast_cog). The number of predicted proteins in relation to subcellular localization and functional category were visualized in plots generated using TIBCO SpotFire software 7.0 (TIBCO, Boston, USA) from the protein list exported of QIP. The Interactivenn web-based tool (Heberle et al., 2015) was used to evaluate the shared proteins among strains through Venn diagram.
Protein-protein interaction (PPI) network was constructed using interolog mapping methodology and metrics according to Folador et al. (2014). To generate a preview of the interaction network was generated using Cytoscape version 2.8.3 (Shannon et al., 2003) with a spring-embedded layout. To indicate the reliability of our predicted PPIs in the database STRING, the network was selected using the score 500 (0.5). In the PPI network, the interactions with score close to 500 are with red or yellow lines and, above 700 in dark green lines. The score indicating how much the pair of proteins in the interaction is similar (homologous) to the interaction according to the database. In the PPI, they interact with at least 65% identity with at least 65% coverage.
A circular map comparing the chromosome of CB129ΔslpB with P. freudenreichii CIRM-BIA 1 and JS17 strains was generated using BLAST Ring Image Generator(BRIG) software v0.95 (Alikhan et al., 2011). Operon prediction in CB129ΔslpB strain was performed using FGENESB (http://www.softberry.com).
Statistical Analyses
Growth curve, MATH assay, Zeta potential measure, and stress challenges were performed with three technical replicates and three biological replicates. The results were expressed as means ± standard deviations. Statistical analyses were performed in GraphPad Prism Software version 7 (GraphPad Software) using Student's t-test, one-way or two-way ANOVA with SIDAK's or Tukey post-hoc analyses for multiple comparisons. Asterisks represent statistically significant differences and were indicated as follows: *p < 0.05; **p < 0.01; ***p < 0.001.
Results
Impact of slpB Mutation on P. freudenreichii Extractable Surface Proteins
SLPs play a key role in probiotic/host interactions and we have shown that such interactions are impaired in an slpB mutant. Electrophoretic analysis of guanidine extracts confirmed the disappearance of the corresponding SlpB protein (do Carmo et al., 2017). In the present study, we further investigated these extractable fractions in order to decipher the impact of such a single mutation on the inventory of SLPs, and more widely, of extractable surface proteins, including surface layer associated proteins (SLAPs). Using nanoLC-MS/MS, we identified 40 surface extractable proteins in CB129ΔslpB strain, yet 33 in the parental wild-type CIRM BIA 129 one (Table 1). The core of extractable proteins, non-covalently bound to the cell wall, common to mutant and parental strains, was composed of 23 proteins, including solute-binding protein of the ABC transport system (BopA), internalin A (InlA), surface protein with SLH Domain E (SlpE), and surface-Layer Protein A (SlpA). Moreover, it comprised a series of cytoplasmic proteins involved in different biological processes like Heat shock 70 kDa protein 1 (HSP70 1), Clp chaperone, GroL1 and GroL2, Elongation factor Tu, and subunits of Methylmalonyl-CoA mutase and subunits of Methylmalonyl-CoA carboxytransferase. Among extractable proteins specific of the CB129ΔslpB, we identified proteins involved in metabolic processes like Coenzyme A transferase involved in acetyl-CoA metabolic process and Pyruvate phosphate dikinase Pyruvate synthase involved in pyruvate metabolic process. Furthermore, this specific subset also comprised another protein involved in stress response (HSP70 2). As expected, the SlpB protein was found only in the parental wild type CIRM BIA 129, yet not in the CB129ΔslpB mutant.
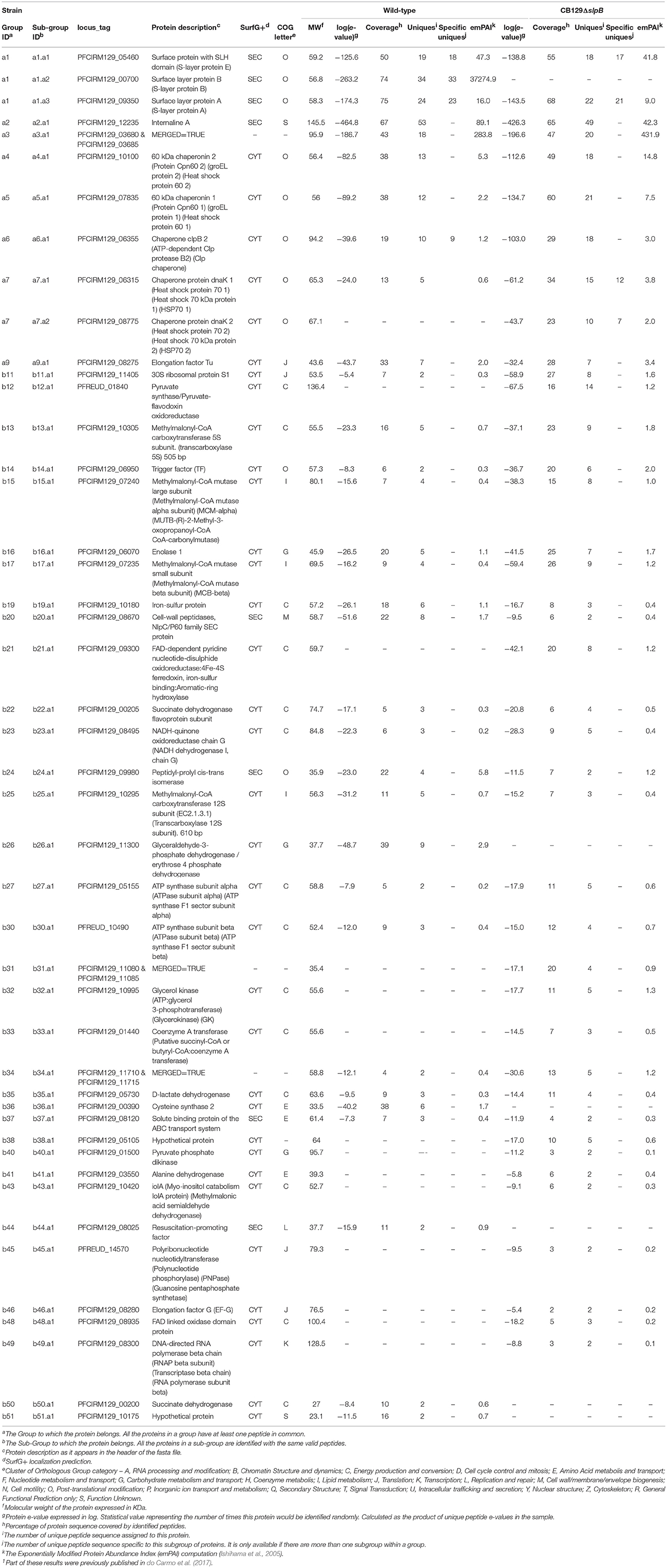
Table 1. Proteins identified in the extraction of surface proteins non-covalently bound to the cell wall using guanidine hydrochloride of CB 129 wild-type and CB129ΔslpB strains1.
Impact of slpB Mutation on P. freudenreichii ζ-Potential and Cell Surface Hydrophobicity
Propionibacterial SLPs, with a low isoelectric point, confer negative charges to the cell surface. In order to identify if the net surface charge was altered in the mutant strain, we conducted ζ-potential and cell surface hydrophobicity assays in both strains. As shown in the Figure 1A, the WT strain exhibited a zeta potential of −21.73 ± 1.63 mV, reflecting a negative net charge, in accordance with the low isoelectric point of P. freudenreichii SlpB protein. By contrast, mutation of slpB gene significantly affected the zeta potential of the CB129ΔslpB strain, which was −6.75 ± 0.55 mV, showing a reduced electronegativity, in accordance with a disorganization of the S-layer at the bacterial cell surface. As shown in Figure 1B, the wild type strain also showed a high affinity to the hydrocarbons tested, whereas the CB129ΔslpB mutant showed a decreased adhesion, whatever the hydrocarbon used in the assay. Adhesion, respectively to mutant and WT strains, were as follow: to Xylol, 0.33 ± 0.52% and 43.67 ± 6.31%, to Chloroform 16.5 ± 10.7% and 75 ± 5.88, and to Ethyl Acetate 5.33 ± 7.17% and 43.83 ± 5.74%. Cell surface properties being drastically affected, we then sought morphological changes caused by the mutation (Figure 2). Both strains exhibited a similar cell wall thickness, 24.33 ± 0.4154 nm and 24.90 ± 0.4154 nm, respectively. No significant difference in term of bacteria morphology, cell wall thickness and shape was observed between the two strains using transmission electron microscopy.
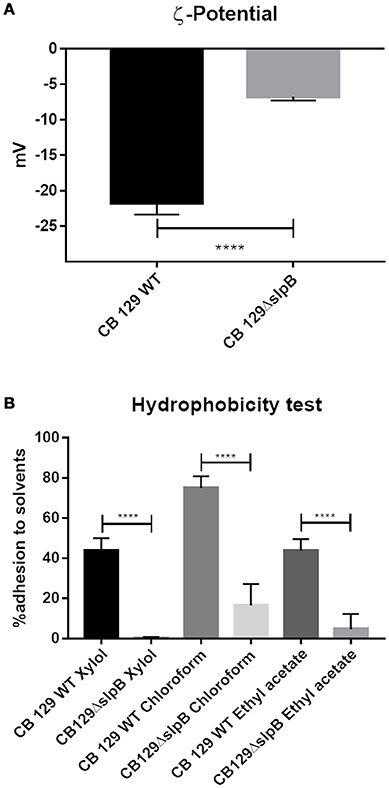
Figure 1. Mutation of slpB gene drastically affects surface proprieties in P. freudenreichii CIRM-BIA 129. (A) The surface net charge was determined by measuring the ζ-potential. (B) The surface hydrophobicity was determined by quantifying adhesion to solvents (Xylol, Chloroform, and Ethyl Acetate). Wild-type (WT) and mutant CB129ΔslpB strains were compared. Bar represents the mean SD of three biological replicates and three technical replicates. The asterisks (****) denotes the statistical significance of the represented value between CB 129 WT and CB 129ΔslpB (p > 0.0001).

Figure 2. Mutation of slpB gene does not affect envelope thickness in P. freudenreichii CIRM-BIA 129. Wild-type (WT) and mutant CB129ΔslpB strains were analyzed by transmission electron microscopy (TEM). No difference in morphology and cell wall tickeness was found.
Impact of slpB Mutation on P. freudenreichii Growth and Stress Tolerance
A single mutation, inactivating a key gene, may affect bacterial fitness and thus probiotic efficacy. We therefore monitored P. freudenreichii growth and tolerance toward acid, bile salts and heat challenges, in the wild type and in the mutant. The growth curves showed a similar pattern for both strains (Figure 3A). The bacterial count at the stationary phase end was also equivalent for both strains, with a viable population count of 1.63 × 109 CFU.mL−1 and 1.75 × 109 CFU.mL−1 for the wild type and the mutant strains, respectively. Tolerance toward stress challenges is reported in Figure 3B. In the case of acid stress, we observed a significant decrease in viability for the CB129ΔslpB strain 0.71 ± 0.13% (7.3 × 106 CFU.mL−1) compared to the WT strain 5.76 ± 1.48% (5.76 × 107 CFU.mL−1). During the bile salts stress, we observed the same trend in the tolerance. Indeed, the survival rate for the CB129ΔslpB strain was significantly decreased 0.37 ± 0.24% (3.71 × 106 CFU.mL−1), compared to the WT strain 2.19 ± 1.01% (2.19 × 107 CFU.mL−1). The same stands for heat challenge, with a reduced survival in CB129ΔslpB 0.71 ± 0.16% (9.01 × 106 CFU.mL−1) compared to WT strain 5.76 ± 1.35% (5.86 × 107 CFU.mL−1).
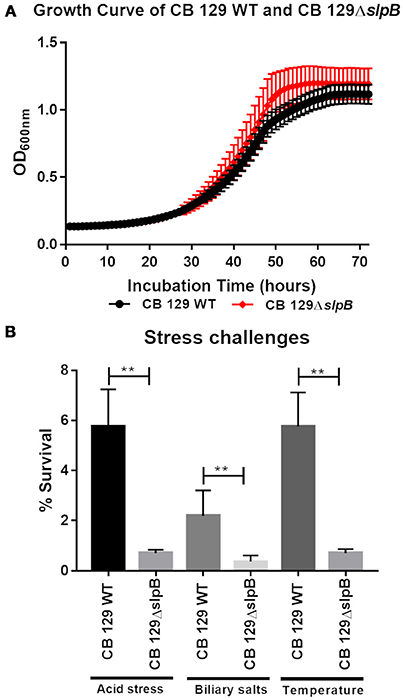
Figure 3. Mutation of SlpB drastically affects stress tolerance in P. freudenreichii CIRM-BIA 129. (A) The growth curve of Wild-type (WT) and mutant CB129ΔslpB strains was determined at 30°C in YEL broth until stationary phase (72 h). Growth was monitored by OD650 nm as a function of time. No statistically significant difference was found in growth curve between strains. (B) Wild-type (WT) and mutant 129ΔslpB strains were subjected to acid, bile salts and thermal challenges. Viable propionibacteria were enumerated by plate counting before and after each challenge. Asterix represent statistically significant differences between strains and were indicated as follows: **p < 0.01.
Impact of slpB Mutation on P. freudenreichii Qualitative and Quantitative Proteome
Considering the major alterations in surface extractable proteins, bacteria cell surface physicochemical properties, and stress tolerance, a qualitative and quantitative analysis of the total proteome was performed to elucidate the impact of the slpB gene knockout in the mutant strain. A total of 1,288 quantifiable proteins (53.26% of predicted proteome) wherein 1,253 proteins (reported in Figure 4A) were identified (Table S1). In the WT strain 1,227 proteins were found, whereas in the CB129ΔslpB strain, we detected 1,252 proteins. Comparative analysis revealed a core-proteome, composed by 1,226 proteins, shared by both strains (Figure 4A). Differences in protein abundance were observed by proteomic quantitative analysis (Figure 4B). A total of 97 proteins (4.2% of the predicted proteome) of these common proteins showed differences in the level of expression among strains, including 36 up-regulated and 61 down-regulated proteins in CB129ΔslpB in comparison with the WT strain (Table 2).

Figure 4. Label-free quantification of proteins from P. freudenreichii CIRM-BIA 129 and CB 129ΔslpB strain. (A) Distribution of the proteins identified in the proteome of WT and CB129ΔslpB strains, represented by a Venn diagram. (B) Volcano Plot showing Log(2) Fold Change of the differentially expressed proteins in CB129ΔslpB strain in relation to WT strain. Green (up-regulated proteins) and red circles (down-regulated proteins) represent proteins statistically different (p ≤ 0.05, ANOVA) in abundance between strains by 2-fold or more. (C) Prediction of the subcellular localization of the proteins identified by LC/MS and organized as cytoplasmic (CYT), membrane (MEM), potentially surface-exposed (PSE) or secreted proteins (SEC).
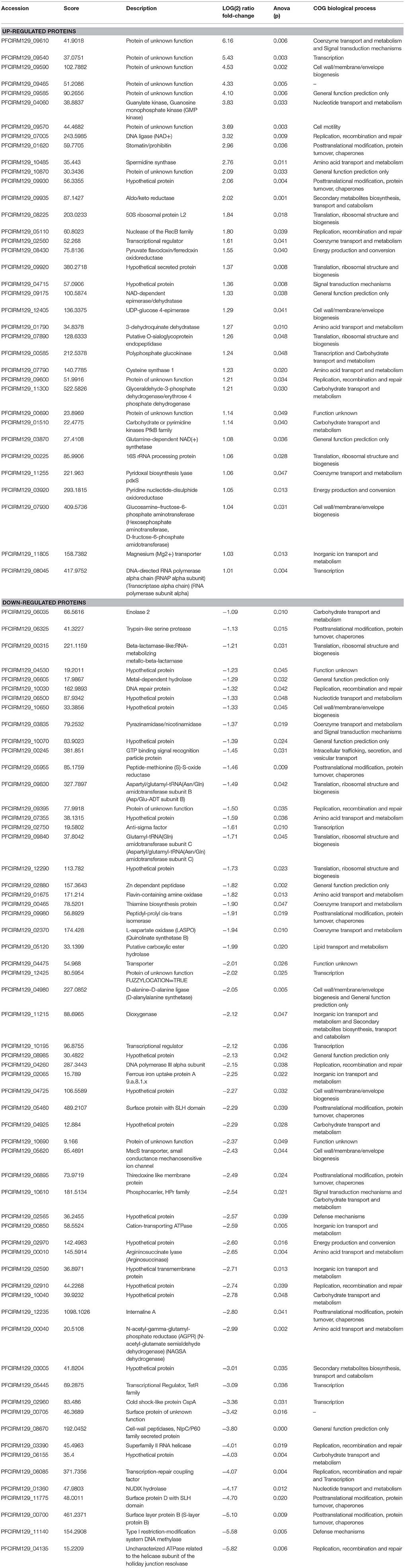
Table 2. Differentially regulated proteins at CB129ΔslpB in relation to CB 129 wild-type.
According to the predicted subcellular localization of the 1,253 proteins identified, 1,081 proteins are CYT (61% of predicted proteome), 71 are MEM (18% of predicted proteome), 77 are PSE (41% of predicted proteome) and 24 are SEC (38% of predicted proteome). In the analysis of non-differentially expressed proteins, we classified 1,001 as CYT proteins, 67 as MEM proteins, 70 as PSE proteins and 22 as SEC proteins (Figure 4C). Meanwhile, between the P. freudenreichii WT and the CB129ΔslpB strains, from 97 proteins differentially expressed, the subcellular localization were predicted as follow: 81 CYT, 2 MEM, 7 PSE, and 7 SEC proteins (Figure 4C).
According to COG functional classifications, the differentially expressed proteins were classified into 20 biological processes (Figure 5A). A general category of differentially regulated proteins in CB129ΔslpB strain core proteome showed 27 proteins involved in information storage and processing, 25 associated to metabolism and, 18 proteins related to cellular processes and signaling (Figure 5A). Proteins that mediate different biological process were dysregulated in the mutant strain. As seen in Figure 5B, 11 proteins were classified as having general functions, 10 proteins related to process of replication, recombination and repair, other 10 proteins linked to posttranslational modification, chaperones, protein turnover, and 9 proteins involved in the transcription process. The differentially expressed proteins between wild-type and mutant strains detected in each functional category are shown in Table 2. In addition, we detected proteins exclusive to the proteome of each strain. WT strain exhibits a unique exclusive protein, the Putative carboxylic ester hydrolase, which is involved in metabolism, especially in hydrolase activity. Interestingly, 27 proteins were found exclusively in the mutant strain, they are involved in several processes like metabolism and replication, recombination and repair (Table S1).
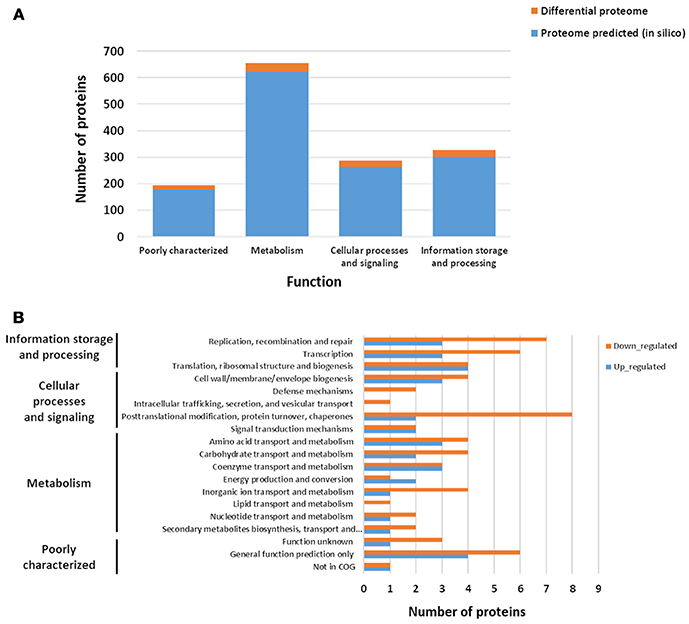
Figure 5. Repartition of differential proteins in biological processes. (A) Functional distribution of the predicted theoretical proteome and of the experimental differential proteins when comparing Wild-type (WT) and mutant CB129ΔslpB strains. (B) Repartition of differential proteins in Biological processes. Functional distribution and Biological process were predicted based on the functional classifications of COG database.
slpB Gene Mutagenesis and Whole-Genome Co-localization
Complete genome of CB129ΔslpB (BioProject - PRJNA476583, Accession - CP030279) strain was sequenced and assembled in a circular chromosome, which exhibits a length of 2.6815.18 bp, with a G+C content of 67.28%, and a total of 2,479 CDSs, 6 rRNA genes (5S, 16S, and 23S), and 45 tRNA genes. The circular map showed a high similarity when comparing CB129ΔslpB with the CIRM-BIA 1 and the JS17 reference strains (Figure 6A). Figure 6B shows the localization of the plasmid inserted within the slpB gene during its knockout and Figures S1, S2 shows the read mapping before and after the insertion. Genomic analyses of genetic context, i.e., the sequences upstream and downstream the slpB gene, confirmed that this locus is not part of an operon and thus should not affect the expression of downstream genes or upstream genes. Complete genome sequence of CB129ΔslpB strain further ruled out any homologous recombination (HR) in other genome sites.
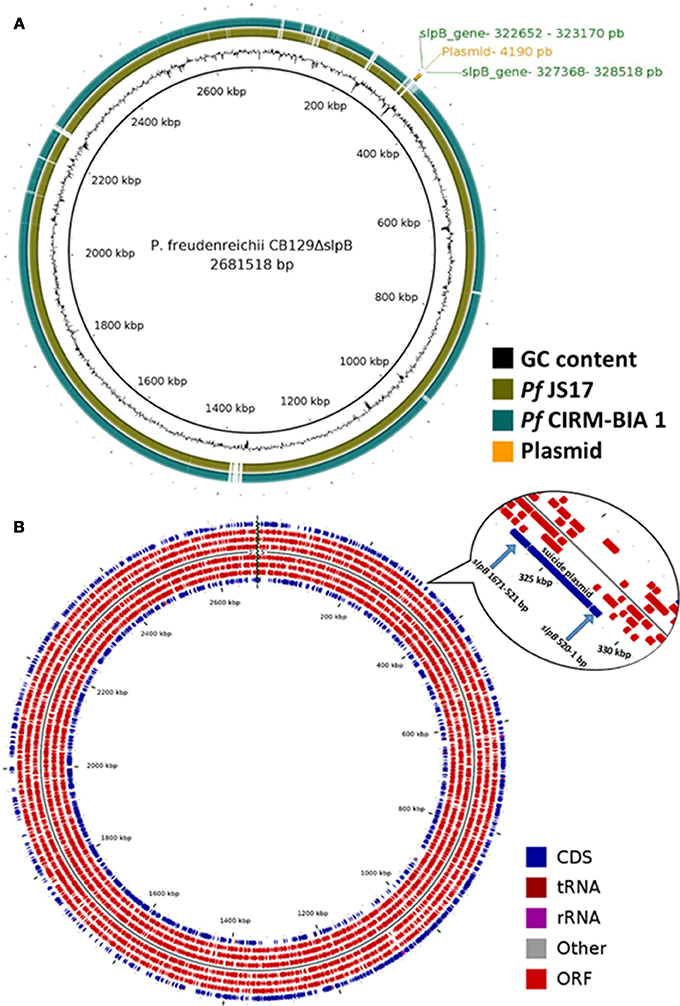
Figure 6. Comparative genomic map generated with BRIG and Map of Circular genome generated with CGview. (A) P. freudenreichii CIRM-BIA 1 and P. freudenreichii JS17 were aligned using CB129ΔslpB strain as a reference. (B) In the outermost ring the genes localization in genome, followed by CDS, tRNAs, rRNAs, other RNAs, and CDSs. The insertion site of the plasmid for the slpB gene mutation is visualized in the zoom image.
Protein-Protein Interaction (PPI)
We performed a PPI network to evaluate the interactions among the proteins differentially regulated in WT and CB129ΔslpB strains (Figure 7). The interactome analysis revealed 118 interactions between identified proteins. In PPI network, we observed that upregulated proteins, such as DNA-directed RNA polymerase alpha chain (PFCIRM129_08045), and 50S ribosomal protein L2 (PFCIRM129_08225), which exhibit high interaction, are involved in Transcription and Translation, respectively. Moreover, downregulated proteins such as GTP binding signal recognition particle protein (PFCIRM129_00245), DNA polymerase III alpha subunit (PFCIRM129_04260) and Enolase 2 (PFCIRM129_06035) showing high interaction, are involved in metabolism, DNA repair and main glycolytic pathway, respectively.
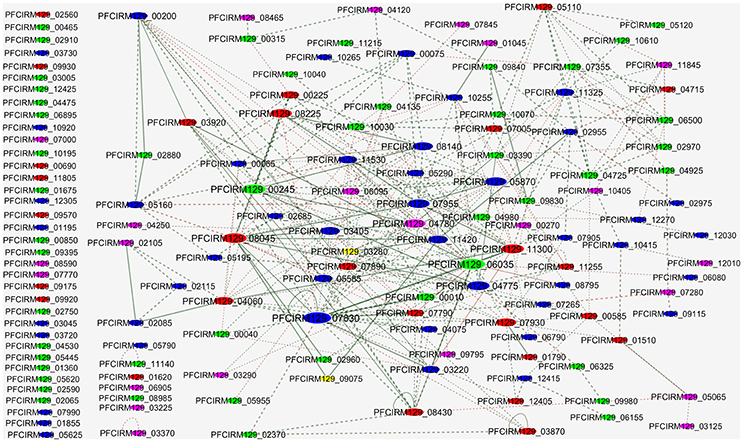
Figure 7. Protein-protein interactions of the proteins identified as differentially expressed in CB 129ΔslpB. The sizes of the nodes represent the degree of interaction for each gene/protein; the major nodes demonstrate greater interactions. Red, up-regulated; Blue, unchanged; Green, down-regulated; Yellow, Exclusive identified at WT strain; Purple, Exclusive identified at CB129ΔslpB strain.
Discussion
Propionibacterium freudenreichii CIRM-BIA 129 has emerged as a probiotic strain with a great immunomodulatory potential in the context of inflammatory bowel disease, according to promising results obtained in animal models (Plé et al., 2015, 2016). Recently, our group has studied the role of the surface SlpB protein of P. freudenreichii CIRM-BIA 129 in adhesion to the intestinal epithelial cells, a probiotic property linked to beneficial effects. Knocking-out of the slpB gene evidenced a direct involvement of this protein in the adhesion to HT-29 cells. Electrophoretic analysis of guanidine extracts confirmed the disappearance of the corresponding SlpB protein ( do Carmo et al., 2017). Surface layer proteins are associated to several functions (do Carmo et al., 2018). Therefore, in order to better understand the impact of this mutation, we performed a more thorough proteomic analysis by applying nanoLC-MS/MS to these extracts. Differences were found between the parental wild type CIRM BIA 129 and the isogenic CB129ΔslpB mutant strains of P. freudenreichii, in terms of surface extractable proteins. As shown in Table 1, proteins previously identified in CB 129 WT strain guanidine-extracted proteins (Le Maréchal et al., 2015) were detected in both strains, including in particular, surface proteins anchored in the peptidoglycan cell wall via surface layer homology (SLH) domains, such as SlpA, SlpB, SlpE, and InlA like as previously reported by Carmo and collaborators (do Carmo et al., 2017). However, this set of SLH domain-containing proteins was reduced in the mutant strain guanidine-extracted proteins, with the expected absence of SlpB protein, thus validating the directed mutagenesis. Analysis of CB129ΔslpB strain guanidine-extracted proteins, identified several proteins, including chaperones, such as ClpB, DnaK, and GroEL, and Enolase (carbohydrate metabolism) involved in stress tolerance, as previously reported for Propionibacterium ssp. strains by enzymatic shaving of the surface proteins using trypsin (Jan et al., 2000; Gagnaire et al., 2015; Huang et al., 2016). Another noticeable difference was the higher number of guanidine-extracted proteins, in the mutant strain, compared to the wild type strain. This included proteins usually described as cytoplasmic: enzymes of the central carbon metabolic pathways, such as pyruvate synthase, or the two subunits of the methylmalonyl-CoA mutase, a recognized cytoplasmic marker, previously described as an extracellular marker of autolysis (Valence et al., 2000). Interestingly, the HSP 70 cytoplasmic stress-related protein present at the surface of the mutant strain could be responsible for preventing protein denaturation. It is as such considered a factor of virulence and pathogenesis in some specific pathogens (Ghazaei, 2017), in Neisseri meingitidis (Knaust et al., 2007) and in Mycobacterium spp. (Das Gupta et al., 2008). This appeals further investigation, as it suggests a profound modification of the envelope structure and cell surface properties of the mutant strain.
SLAPs are known to determine key parameters of the surface layer of bacteria, in terms of charge and hydrophobicity (Wilson et al., 2001). Not only amino acid residues, but also covalent modification may endow the S-layer lattice with a strong negative charge. Thus, we determined the surface charge in both P. freudenreichii WT and CB129ΔslpB strains by measuring the zeta potential, which reflects the mobility rate of cells within an electric field. A lower negative value is reportedly linked with higher hydrophobicity, consequently improving adhesion (de Wouters et al., 2015). Likewise, considering the presence of surface proteins and their role in zeta potential, van der Mei et al. have shown that some wild type strains, like the L. acidophilus ATCC4356, with SLPs, are more negatively charged at pH 7 than strains without SLPs, such as L. johnsonii LMG9436 and L. gasseri LMG9203 (van der Mei et al., 2003). We thus further investigated the hydrophobicity of the cell surface, a parameter thought to be correlated with in vitro adhesion of bacteria to mucin, collagen, fibronectin, and to human epithelial cells (Duary et al., 2011). The cell surface hydrophobic and hydrophilic properties have been studied in lactic acid bacteria (Sandes et al., 2017) and can be correlated to the adhesion process to intestinal epithelial cells of apolar surface proteins (Guo et al., 2010). Using the MATH assay, we showed that the CB129ΔslpB strain has a strongly decreased ability to adhere to xylol, as well as to chloroform and to ethyl acetate solvents, indicating a change in the global properties of the cell surface, affecting adhesion to surfaces. These results corroborate with the previous study showing a decreased adhesion to HT-29 human intestinal epithelial cells (do Carmo et al., 2017). Hydrophobicity and ζ-potential are factors correlated with bacterial adhesion to the epithelial cells, which are guided by charge and hydrophobicity of the bacterial surface.
The presence of surface layers being reportedly linked to tolerance toward stresses (do Carmo et al., 2018), we decided to investigate the impact of such a mutation on the CB129ΔslpB strain tolerance toward stress challenges that are relevant for the selection of new probiotics. The ability to survive acid stress in the stomach and bile salts stress in the duodenum during the passage through the digestive tract, is important for probiotic interaction with the host (Rabah et al., 2017). Accordingly, in vitro assays can be used to simulate digestive stresses, mimicking the exposure to acidic conditions (pH 2.0) or to biliary salts (1 g.L−1) (Jan et al., 2000). For P. freudenreichii, commonly used as a cheese starter, the heat stress tolerance constitutes a relevant technological ability of this strain (Rosa do Carmo et al., 2017). Overall, we observed a large decrease in tolerance to the environmental stresses, confirming a role of SlpB in toughness. In the guanidine-extracted proteins of the mutant strain, the chaperones and heat shock proteins, DnaK1, DnaK2, ClpB 2, GroE1, and GroE2 were found. Inside the cell, they are responsible for protein folding and are correlated to acid and bile adaptation (Leverrier et al., 2005; Gagnaire et al., 2015). Here, they were found at the surface of the CB129ΔslpB mutant, which was more susceptible to extreme acid stress and temperature, compared to wild type strain. Previous work showed that L. acidophilus ATCC 4356 adapts to harsh environments by increasing the expression of the s-layer SlpA protein upon bile, acidic pH and heat stress exposition (Khaleghi et al., 2010; Khaleghi and Kasra, 2012). Moreover, changes in the cell surface properties could alter the transmembrane protein complex responsible for the extrusion of protons from the cytoplasm, which are responsible for surviving environmental stresses (Ruiz et al., 2013; Rosa do Carmo et al., 2017).
Profound modifications of P. freudenreichii physiology and surface properties suggested that modifications, wider than the disappearance of a single protein, occurred as a result of slpB gene inactivation. To understand this impact of the mutation, a comparative proteomic analysis was performed to identify significant alterations in the whole proteome profile of the mutant strain, using label-free quantitative proteomic analysis. Prediction of sub cellular localization using the SurfG+ tool (Barinov et al., 2009) evidenced changes in all the categories (CYT, MEM, PSE and SEC) in the differential proteome of CB129ΔslpB. In addition, differential proteome was functionally classified using COG, showing a functional implication of differential proteins in cellular processes such as signaling, information storage, processing, and metabolism. Specifically, this study showed that the moonlighting enolase and NlpC/P60 are both exported (Frohnmeyer et al., 2018), as it was recently observed in the cutaneous Propionibacterium acnes strain (Jeon et al., 2017). These moonlighting proteins were downregulated in CB129ΔslpB. Interestingly, in the Bifidobacterium and Lactobacillus genera, moonlighting proteins, such as enolase, also play a role in immunomodulation and adhesion (Sánchez et al., 2010; Kainulainen and Korhonen, 2014; Vastano et al., 2016). Furthermore, in the PPI network we observed high interactions between the downregulated Enolase (PFCIRM129_06035), reportedly involved in human gut colonization and stress adaptation (Ruiz et al., 2009), with other proteins involved in several other processes, including metabolism and DNA repair. Moreover, all surface layer-associated proteins SlpA, SlpD, SlpE, and InlA were downregulated in CB129ΔslpB. These proteins form a protective layer on the surface of the bacteria, and have been associated with environmental stress tolerance (Fagan and Fairweather, 2014). As seen previously, a decreased amount of these proteins could be directly associated with stress susceptibility and with altered hydrophobicity. SLAPs can directly influence these properties (Pum et al., 2013), and consequently alter adhesion to epithelial cells (do Carmo et al., 2017).
We performed the complete genome DNA sequencing of the CB129ΔslpB, which, in turn, allowed us to evaluate whether the slpB gene disruption had major consequences on the mutant strain genome. The slpB gene is not part of an operon, which suggests that homologous recombination using the suicide plasmid pUC:ΔslpB:CmR (do Carmo et al., 2017) did not affect the expression of upstream and downstream genes. Analysis of the genetic context, upstream and downstream, revealed that the homologous recombination process was site-specific, and not affecting other genes in the genome of the mutant strain CB129ΔslpB. However, we were unable to evaluate possible rearrangements in the genome of CB129ΔslpB, which could have affected the transcription of other genes. Therefore, more studies are necessary to explore whether any probiotic potential was lost after the single mutation of the slpB gene in Propionibacterium freudenreichii CIRM-BIA 129 strain.
Conclusion
This study evidenced the pleiotropic impact of the surface layer protein slpB mutation in the probiotic strain Propionibacterium freudenreichii CIRM-BIA 129 in relation to its physicochemical proprieties, stress challenges, surfaceome and whole cell quantitative proteome. It confirmed the key role of SLPs and strongly suggests that expression of specific ones, such as P. freudenreichii SlpB, should be used as criteria for selecting strains with probiotic potential.
Author Contributions
FC performed in vitro assays, microscopy, proteomic assays and data interpretation. WS, FP, GT, and ROC performed proteomic assays, data interpretation and bioinformatics analyses. BC, EO, and SS performed in vitro assays. II and HR data interpretation. EF performed PPI network. CC performed microscopy. MC, AC, and RS performed genomics and data interpretation. VA, GJ, HF, and YL contributed to the supervision, analysis, and interpretation of data and were major contributors to revising the manuscript. All authors contributed in writing the manuscript.
Funding
This work was supported by Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq), Fundação de Amparo à Pesquisa do Estado de Minas Gerais (FAPEMIG), and Coordenação de Aperfeiçoamento de Pessoal de Nível Superior (CAPES).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer MAS declared a shared affiliation, with no collaboration, with one of the authors, WS, to the handling editor at time of review.
Acknowledgments
The authors thank Prof. Dra. Mônica Cristina de Oliveira UFMG, Prof. Dr. Leonardo Borges Acúrcio UNIFOR-MG, Dra. Cristiana Perdigão Rezende AQUACEN, Dra. Fernanda Dorella AQUACEN, Miss. Gabriella Borba de Assis AQUACEN for expert technical assistance and useful discussions and advices. GJ also thanks A.B.I. Timadeuc for sharing experiences.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.01807/full#supplementary-material
Figure S1. Verification of assembly error by read mapping. The plasmid pUC:ΔslpB:CmR was not inserted in the slpB gene during de novo genome assembly. The read mapping on the slpB gene shows misalignments upstream and downstream insertion site, confirming the assembly error. The read mapping was performed using in CLC Genomics Workbench 7.0.
Figure S2. Assembly curation and validation by read mapping. The manual insertion of plasmid pUC:ΔslpB:CmR in the slpB gene was validated by read mapping. The correct read alignments upstream (A) and downstream (B) the plasmid validate the manual insertion. The read mapping was performed using in CLC Genomics Workbench 7.0.
Table S1. Total list of proteins identified in the core-proteome of CB 129 wild-type and CB129ΔslpB.
References
Alikhan, N.-F., Petty, N. K., Ben Zakour, N. L., and Beatson, S. A. (2011). BLAST ring image generator (BRIG): simple prokaryote genome comparisons. BMC Genomics 12:402. doi: 10.1186/1471-2164-12-402
Aziz, R. K., Bartels, D., Best, A. A., DeJongh, M., Disz, T., Edwards, R. A., et al. (2008). The RAST server: rapid annotations using subsystems technology. BMC Genomics 9:75. doi: 10.1186/1471-2164-9-75
Barinov, A., Loux, V., Hammani, A., Nicolas, P., Langella, P., Ehrlich, D., et al. (2009). Prediction of surface exposed proteins in Streptococcus pyogenes, with a potential application to other Gram-positive bacteria. Proteomics 9, 61–73. doi: 10.1002/pmic.200800195
Brettin, T., Davis, J. J., Disz, T., Edwards, R. A., Gerdes, S., Olsen, G. J., et al. (2015). RASTtk: a modular and extensible implementation of the RAST algorithm for building custom annotation pipelines and annotating batches of genomes. Sci. Rep. 5:8365. doi: 10.1038/srep08365
Cousin, F. J., Jouan-Lanhouet, S., Théret, N., Brenner, C., Jouan, E., Le Moigne-Muller, G., et al. (2016). The probiotic Propionibacterium freudenreichii as a new adjuvant for TRAIL-based therapy in colorectal cancer. Oncotarget 7, 7161–7178. doi: 10.18632/oncotarget.6881
Das Gupta, T., Bandyopadhyay, B., and Das Gupta, S. K. (2008). Modulation of DNA-binding activity of Mycobacterium tuberculosis HspR by chaperones. Microbiology 154, 484–490. doi: 10.1099/mic.0.2007/012294-0
de Wouters, T., Jans, C., Niederberger, T., Fischer, P., and Rühs, P. A. (2015). Adhesion potential of intestinal microbes predicted by physico-chemical characterization methods. PLoS ONE 10:e0136437. doi: 10.1371/journal.pone.0136437
Deutsch, S.-M., Mariadassou, M., Nicolas, P., Parayre, S., Le Guellec, R., Chuat, V., et al. (2017). Identification of proteins involved in the anti-inflammatory properties of Propionibacterium freudenreichii by means of a multi-strain study. Sci. Rep. 7:46409. doi: 10.1038/srep46409
Deutsch, S. M., Parayre, S., Bouchoux, A., Guyomarc'h, F., Dewulf, J., Dols-Lafargue, M., et al. (2012). Contribution of surface β-glucan polysaccharide to physicochemical and immunomodulatory properties of Propionibacterium freudenreichii. Appl. Environ. Microbiol. 78, 1765–1775. doi: 10.1128/AEM.07027-11
Distler, U., Kuharev, J., Navarro, P., Levin, Y., Schild, H., and Tenzer, S. (2014). Drift time-specific collision energies enable deep-coverage data-independent acquisition proteomics. Nat. Methods 11, 167–170. doi: 10.1038/nmeth.2767
do Carmo, F. L. R., Rabah, H., De Oliveira Carvalho, R. D., Gaucher, F., Cordeiro, B. F., da Silva, S. H., et al. (2018). Extractable bacterial surface proteins in probiotic–host interaction. Front. Microbiol. 9:645. doi: 10.3389/fmicb.2018.00645
do Carmo, F. L. R., Rabah, H., Huang, S., Gaucher, F., Deplanche, M., Dutertre, S., et al. (2017). Propionibacterium freudenreichii surface protein SlpB is involved in adhesion to intestinal HT-29 cells. Front. Microbiol. 8:1033. doi: 10.3389/fmicb.2017.01033
Duary, R. K., Rajput, Y. S., Batish, V. K., and Grover, S. (2011). Assessing the adhesion of putative indigenous probiotic lactobacilli to human colonic epithelial cells. Ind. J. Med. Res. 134, 664–671. doi: 10.4103/0971-5916.90992
Fagan, R. P., and Fairweather, N. F. (2014). Biogenesis and functions of bacterial S-layers. Nat. Rev. Microbiol. 12, 211–222. doi: 10.1038/nrmicro3213
Folador, E. L., Hassan, S. S., Lemke, N., Barh, D., Silva, A., Ferreira, R. S., et al. (2014). An improved interolog mapping-based computational prediction of protein–protein interactions with increased network coverage. Integr. Biol. 6, 1080–1087. doi: 10.1039/C4IB00136B
Foligné, B., Deutsch, S. M., Breton, J., Cousin, F. J., Dewulf, J., Samson, M., et al. (2010). Promising immunomodulatory effects of selected strains of dairy propionibacteria as evidenced in vitro and in vivo. Appl. Environ. Microbiol. 76, 8259–8264. doi: 10.1128/AEM.01976-10
Food and Agriculture Organization of the United Nations and World Health Organization (eds.). (2002). Probiotics in Food: Health and Nutritional Properties and Guidelines for Evaluation. Rome: Food and Agriculture Organization of the United Nations : World Health Organization.
Ford, A. C., Quigley, E. M. M., Lacy, B. E., Lembo, A. J., Saito, Y. A., Schiller, L. R., et al. (2014). Efficacy of prebiotics, probiotics, and synbiotics in irritable bowel syndrome and chronic idiopathic constipation: systematic review and meta-analysis. Am. J. Gastroenterol. 109, 1547–1561; quiz 1546, 1562. doi: 10.1038/ajg.2014.202
Frohnmeyer, E., Deptula, P., Nyman, T. A., Laine, P. K. S., Vihinen, H., Paulin, L., et al. (2018). Secretome profiling of Propionibacterium freudenreichii reveals highly variable responses even among the closely related strains. Microb. Biotechnol. 11, 510–526. doi: 10.1111/1751-7915.13254
Furuichi, K., Hojo, K., Katakura, Y., Ninomiya, K., and Shioya, S. (2006). Aerobic culture of Propionibacterium freudenreichii ET-3 can increase production ratio of 1,4-dihydroxy-2-naphthoic acid to menaquinone. J. Biosci. Bioeng. 101, 464–470. doi: 10.1263/jbb.101.464
Gagnaire, V., Jardin, J., Rabah, H., Briard-Bion, V., and Jan, G. (2015). Emmental cheese environment enhances Propionibacterium freudenreichii stress tolerance. PLoS ONE 10:e0135780. doi: 10.1371/journal.pone.0135780
Galardini, M., Biondi, E. G., Bazzicalupo, M., and Mengoni, A. (2011). CONTIGuator: a bacterial genomes finishing tool for structural insights on draft genomes. Source Code Biol. Med. 6:11. doi: 10.1186/1751-0473-6-11
Galperin, M. Y., Makarova, K. S., Wolf, Y. I., and Koonin, E. V. (2015). Expanded microbial genome coverage and improved protein family annotation in the COG database. Nucleic Acids Res. 43, D261–D269. doi: 10.1093/nar/gku1223
Ghazaei, C. (2017). Role and mechanism of the Hsp70 molecular chaperone machines in bacterial pathogens. J. Med. Microbiol. 66, 259–265. doi: 10.1099/jmm.0.000429
Gilar, M., Olivova, P., Daly, A. E., and Gebler, J. C. (2005). Two-dimensional separation of peptides using RP-RP-HPLC system with different pH in first and second separation dimensions. J. Sep. Sci. 28, 1694–1703. doi: 10.1002/jssc.200500116
Giles, K., Williams, J. P., and Campuzano, I. (2011). Enhancements in travelling wave ion mobility resolution. Rapid Commun. Mass Spectrom. RCM 25, 1559–1566. doi: 10.1002/rcm.5013
Guo, X.-H., Kim, J.-M., Nam, H.-M., Park, S.-Y., and Kim, J.-M. (2010). Screening lactic acid bacteria from swine origins for multistrain probiotics based on in vitro functional properties. Anaerobe 16, 321–326. doi: 10.1016/j.anaerobe.2010.03.006
Heberle, H., Meirelles, G. V., da Silva, F. R., Telles, G. P., and Minghim, R. (2015). InteractiVenn: a web-based tool for the analysis of sets through Venn diagrams. BMC Bioinformatics 16:169. doi: 10.1186/s12859-015-0611-3
Hill, C., Guarner, F., Reid, G., Gibson, G. R., Merenstein, D. J., Pot, B., et al. (2014). Expert consensus document: the international scientific association for probiotics and prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 11, 506–514. doi: 10.1038/nrgastro.2014.66
Huang, S., Rabah, H., Jardin, J., Briard-Bion, V., Parayre, S., Maillard, M.-B., et al. (2016). Hyperconcentrated sweet whey, a new culture medium that enhances Propionibacterium freudenreichii stress tolerance. Appl. Environ. Microbiol. 82, 4641–4651. doi: 10.1128/AEM.00748-16
Hynönen, U., and Palva, A. (2013). Lactobacillus surface layer proteins: structure, function and applications. Appl. Microbiol. Biotechnol. 97, 5225–5243. doi: 10.1007/s00253-013-4962-2
Isawa, K., Hojo, K., Yoda, N., Kamiyama, T., Makino, S., Saito, M., et al. (2002). Isolation and identification of a new bifidogenic growth stimulator produced by Propionibacterium freudenreichii ET-3. Biosci. Biotechnol. Biochem. 66, 679–681. doi: 10.1271/bbb.66.679
Ishihama, Y., Oda, Y., Tabata, T., Sato, T., Nagasu, T., Rappsilber, J., et al. (2005). Exponentially modified protein abundance index (emPAI) for estimation of absolute protein amount in proteomics by the number of sequenced peptides per protein. Mol. Cell. Proteomics MCP 4, 1265–1272. doi: 10.1074/mcp.M500061-MCP200
Jan, G., Belzacq, A.-S., Haouzi, D., Rouault, A., Métivier, D., Kroemer, G., et al. (2002). Propionibacteria induce apoptosis of colorectal carcinoma cells via short-chain fatty acids acting on mitochondria. Cell Death Differ. 9, 179–188. doi: 10.1038/sj.cdd.4400935
Jan, G., Rouault, A., and Maubois, J.-L. (2000). Acid stress susceptibility and acid adaptation of Propionibacterium freudenreichii subsp. shermanii. Le Lait 80, 325–336. doi: 10.1051/lait:2000128
Jeon, J., Mok, H. J., Choi, Y., Park, S. C., Jo, H., Her, J., et al. (2017). Proteomic analysis of extracellular vesicles derived from Propionibacterium acnes. PROTEOMICS - Clin. Appl. 11:1600040. doi: 10.1002/prca.201600040
Kainulainen, V., and Korhonen, T. K. (2014). Dancing to another tune-adhesive moonlighting proteins in bacteria. Biology 3, 178–204. doi: 10.3390/biology3010178
Khaleghi, M., and Kasra, R. (2012). “Effect of environmental stresses on S-layer production in Lactobacillus acidophilus ATCC 4356,” in Advances in Applied Biotechnology, ed. M. Petre (London: InTech), 209–224.
Khaleghi, M., Kermanshahi, R. K., Yaghoobi, M. M., Zarkesh-Esfahani, S. H., and Baghizadeh, A. (2010). Assessment of bile salt effects on s-layer production, slp gene expression and some physicochemical properties of Lactobacillus acidophilus ATCC 4356. J. Microbiol. Biotechnol. 20, 749–756. doi: 10.4014/jmb.0906.06050
Knaust, A., Weber, M. V. R., Hammerschmidt, S., Bergmann, S., Frosch, M., and Kurzai, O. (2007). Cytosolic proteins contribute to surface plasminogen recruitment of Neisseria meningitidis. J. Bacteriol. 189, 3246–3255. doi: 10.1128/JB.01966-06
Kos, B., Susković, J., Vuković, S., Simpraga, M., Frece, J., and Matosić, S. (2003). Adhesion and aggregation ability of probiotic strain Lactobacillus acidophilus M92. J. Appl. Microbiol. 94, 981–987. doi: 10.1046/j.1365-2672.2003.01915.x
Kuharev, J., Navarro, P., Distler, U., Jahn, O., and Tenzer, S. (2015). In-depth evaluation of software tools for data-independent acquisition based label-free quantification. Proteomics 15, 3140–3151. doi: 10.1002/pmic.201400396
Lan, A., Bruneau, A., Bensaada, M., Philippe, C., Bellaud, P., Rabot, S., et al. (2008). Increased induction of apoptosis by Propionibacterium freudenreichii TL133 in colonic mucosal crypts of human microbiota-associated rats treated with 1,2-dimethylhydrazine. Br. J. Nutr. 100, 1251–1259. doi: 10.1017/S0007114508978284
Lan, A., Lagadic-Gossmann, D., Lemaire, C., Brenner, C., and Jan, G. (2007). Acidic extracellular pH shifts colorectal cancer cell death from apoptosis to necrosis upon exposure to propionate and acetate, major end-products of the human probiotic propionibacteria. Apoptosis Int. J. Program. Cell Death 12, 573–591. doi: 10.1007/s10495-006-0010-3
Langella, O., Valot, B., Balliau, T., Blein-Nicolas, M., Bonhomme, L., and Zivy, M. (2017). X!TandemPipeline: a tool to manage sequence redundancy for protein inference and phosphosite identification. J. Proteome Res. 16, 494–503. doi: 10.1021/acs.jproteome.6b00632
Le Maréchal, C., Peton, V., Plé, C., Vroland, C., Jardin, J., Briard-Bion, V., et al. (2015). Surface proteins of Propionibacterium freudenreichii are involved in its anti-inflammatory properties. J. Proteomics 113, 447–461. doi: 10.1016/j.jprot.2014.07.018
Leibowitz, M. P., Tavares, G. C., Pereira, F. L., Perdigão, C., Azevedo, V., and Figueiredo, H. C. P. (2017). Shotgun label-free proteomic analyses of the oyster parasite Perkinsus marinus. J. Proteomics Genomics Res. 2:13. doi: 10.14302/issn.2326-0793.JPGR-17-1571
Leverrier, P., Dimova, D., Pichereau, V., Auffray, Y., Boyaval, P., and Jan, G. (2003). Susceptibility and adaptive response to bile salts in Propionibacterium freudenreichii: physiological and proteomic analysis. Appl. Environ. Microbiol. 69, 3809–3818.
Leverrier, P., Fremont, Y., Rouault, A., Boyaval, P., and Jan, G. (2005). In vitro tolerance to digestive stresses of propionibacteria: influence of food matrices. Food Microbiol. 22, 11–18. doi: 10.1016/j.fm.2004.05.003
Lin, Y., Liu, Y., Li, J., Zhao, Y., He, Q., Han, W., et al. (2010). Evaluation and optimization of removal of an acid-insoluble surfactant for shotgun analysis of membrane proteome. Electrophoresis 31, 2705–2713. doi: 10.1002/elps.201000161
Malik, A. C., Reinbold, G. W., and Vedamuthu, E. R. (1968). An evaluation of the taxonomy of Propionibacterium. Can. J. Microbiol. 14, 1185–1191.
Mitsuyama, K., Masuda, J., Yamasaki, H., Kuwaki, K., Kitazaki, S., Koga, H., et al. (2007). Treatment of ulcerative colitis with milk whey culture with Propionibacterium freudenreichii. J. Intest. Microbiol. 21, 143–147. doi: 10.11209/jim.21.143
Oak, S. J., and Jha, R. (2018). The effects of probiotics in lactose intolerance: a systematic review. Crit. Rev. Food Sci. Nutr. 9, 1–9. doi: 10.1080/10408398.2018.1425977
Plaza-Díaz, J., Ruiz-Ojeda, F. J., Vilchez-Padial, L. M., and Gil, A. (2017). Evidence of the anti-inflammatory effects of probiotics and synbiotics in intestinal chronic diseases. Nutrients 9:E555. doi: 10.3390/nu9060555
Plé, C., Breton, J., Richoux, R., Nurdin, M., Deutsch, S.-M., Falentin, H., et al. (2016). Combining selected immunomodulatory Propionibacterium freudenreichii and Lactobacillus delbrueckii strains: reverse engineering development of an anti-inflammatory cheese. Mol. Nutr. Food Res. 60, 935–948. doi: 10.1002/mnfr.201500580
Plé, C., Richoux, R., Jardin, J., Nurdin, M., Briard-Bion, V., Parayre, S., et al. (2015). Single-strain starter experimental cheese reveals anti-inflammatory effect of Propionibacterium freudenreichii CIRM BIA 129 in TNBS-colitis model. J. Funct. Foods 18, 575–585. doi: 10.1016/j.jff.2015.08.015
Pum, D., Toca-Herrera, J. L., and Sleytr, U. B. (2013). S-layer protein self-assembly. Int. J. Mol. Sci. 14, 2484–2501. doi: 10.3390/ijms14022484
Rabah, H., Rosa do Carmo, F. L., and Jan, G. (2017). Dairy Propionibacteria: versatile probiotics. Microorganisms 5:E24. doi: 10.3390/microorganisms5020024
Roland, N., Bougle, D., Lebeurrier, F., Arhan, P., and Maubois, J. L. (1998). Propionibacterium freudenreichii stimulates the growth of Bifidobacterium bifidum in vitro and increases fecal bifidobacteria in healthy human volunteers. Int. Dairy J. 8, 587–588.
Rondanelli, M., Faliva, M. A., Perna, S., Giacosa, A., Peroni, G., and Castellazzi, A. M. (2017). Using probiotics in clinical practice: where are we now? A review of existing meta-analyses. Gut Microbes 8, 521–543. doi: 10.1080/19490976.2017.1345414
Rosa do Carmo, F. L., Rabah, H., Fernandes Cordeiro, B., Da Silva, S. H., Jan, G., Azevedo, V. A., et al. (2017). “Applications of Probiotic Bacteria and Dairy Foods in Health,” in Current Research in Microbiology (Wilmington, DE). Available online at: https://hal.archives-ouvertes.fr/hal-01583804
Ruiz, L., Couté, Y., Sánchez, B., de los Reyes-Gavilán, C. G., Sanchez, J.-C., and Margolles, A. (2009). The cell-envelope proteome of Bifidobacterium longum in an in vitro bile environment. Microbiology 155, 957–967. doi: 10.1099/mic.0.024273-0
Ruiz, L., Margolles, A., and Sánchez, B. (2013). Bile resistance mechanisms in Lactobacillus and Bifidobacterium. Front. Microbiol. 4:396. doi: 10.3389/fmicb.2013.00396
Sambrook, J., and Russell, D. W. (2001). Molecular Cloning: a Laboratory Manual. 3rd edn. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press.
Sánchez, B., Urdaci, M. C., and Margolles, A. (2010). Extracellular proteins secreted by probiotic bacteria as mediators of effects that promote mucosa-bacteria interactions. Microbiol. Read. Engl. 156, 3232–3242. doi: 10.1099/mic.0.044057-0
Sandes, S., Alvim, L., Silva, B., Acurcio, L., Santos, C., Campos, M., et al. (2017). Selection of new lactic acid bacteria strains bearing probiotic features from mucosal microbiota of healthy calves: looking for immunobiotics through in vitro and in vivo approaches for immunoprophylaxis applications. Microbiol. Res. 200, 1–13. doi: 10.1016/j.micres.2017.03.008
Schär-Zammaretti, P., and Ubbink, J. (2003). The cell wall of lactic acid bacteria: surface constituents and macromolecular conformations. Biophys. J. 85, 4076–4092. doi: 10.1016/S0006-3495(03)74820-6
Seki, K., Nakao, H., Umino, H., Isshiki, H., Yoda, N., Tachihara, R., et al. (2004). Effects of fermented milk whey containing novel bifidogenic growth stimulator produced by Propionibacterium on fecal bacteria, putrefactive metabolite, defecation frequency and fecal properties in senile volunteers needed serious nursing-care taking enteral nutrition by tube feeding. J. Intest. Microbiol. 18, 107–115. doi: 10.11209/jim.18.107
Shannon, P., Markiel, A., Ozier, O., Baliga, N. S., Wang, J. T., Ramage, D., et al. (2003). Cytoscape: a software environment for integrated models of biomolecular interaction networks. Genome Res. 13, 2498–2504. doi: 10.1101/gr.1239303
Silva, W. M., Carvalho, R. D., Soares, S. C., Bastos, I. F., Folador, E. L., Souza, G. H., et al. (2014). Label-free proteomic analysis to confirm the predicted proteome of Corynebacterium pseudotuberculosis under nitrosative stress mediated by nitric oxide. BMC Genomics 15:1065. doi: 10.1186/1471-2164-15-1065
Silva, W. M., Folador, E. L., Soares, S. C., Souza, G. H. M. F., Santos, A. V., Sousa, C. S., et al. (2017). Label-free quantitative proteomics of Corynebacterium pseudotuberculosis isolates reveals differences between Biovars ovis and equi strains. BMC Genomics 18:451. doi: 10.1186/s12864-017-3835-y
Suzuki, A., Mitsuyama, K., Koga, H., Tomiyasu, N., Masuda, J., Takaki, K., et al. (2006). Bifidogenic growth stimulator for the treatment of active ulcerative colitis: a pilot study. Nutrition 22, 76–81.doi: 10.1016/j.nut.2005.04.013
Valence, F., Deutsch, S. M., Richoux, R., Gagnaire, V., and Lortal, S. (2000). Autolysis and related proteolysis in Swiss cheese for two Lactobacillus helveticus strains. J. Dairy Res. 67, 261–271. doi: 10.1017/S0022029900004118
van der Mei, H., van de Belt-Gritter, B., Pouwels, P., Martinez, B., and Busscher, H. (2003). Cell surface hydrophobicity is conveyed by S-layer proteins—a study in recombinant lactobacilli. Colloids Surf. B Biointerfaces 28, 127–134. doi: 10.1016/S0927-7765(02)00144-3
Vastano, V., Pagano, A., Fusco, A., Merola, G., Sacco, M., and Donnarumma, G. (2016). The Lactobacillus plantarum Eno A1 enolase is involved in immunostimulation of Caco-2 cells and in biofilm development. Adv. Exp. Med. Biol. 897, 33–44. doi: 10.1007/5584_2015_5009
Vizcaíno, J. A., Csordas, A., del-Toro, N., Dianes, J. A., Griss, J., Lavidas, I., et al. (2016). 2016 update of the PRIDE database and its related tools. Nucleic Acids Res. 44, D447–D456. doi: 10.1093/nar/gkv1145
Wilson, W. W., Wade, M. M., Holman, S. C., and Champlin, F. R. (2001). Status of methods for assessing bacterial cell surface charge properties based on zeta potential measurements. J. Microbiol. Methods 43, 153–164. doi: 10.1016/S0167-7012(00)00224-4
Keywords: bacteria genomic, bacteria proteomic, surface layer protein, HDMSE, shotgun proteomic
Citation: do Carmo FLR, Silva WM, Tavares GC, Ibraim IC, Cordeiro BF, Oliveira ER, Rabah H, Cauty C, da Silva SH, Canário Viana MV, Caetano ACB, dos Santos RG, de Oliveira Carvalho RD, Jardin J, Pereira FL, Folador EL, Le Loir Y, Figueiredo HCP, Jan G and Azevedo V (2018) Mutation of the Surface Layer Protein SlpB Has Pleiotropic Effects in the Probiotic Propionibacterium freudenreichii CIRM-BIA 129. Front. Microbiol. 9:1807. doi: 10.3389/fmicb.2018.01807
Received: 18 May 2018; Accepted: 18 July 2018;
Published: 17 August 2018.
Edited by:
Aleš Berlec, Jožef Stefan Institute (IJS), SloveniaReviewed by:
Paulina Deptula, University of Helsinki, FinlandMaria de los Angeles Serradell, Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET), Argentina
Copyright © 2018 do Carmo, Silva, Tavares, Ibraim, Cordeiro, Oliveira, Rabah, Cauty, da Silva, Canário Viana, Caetano, dos Santos, de Oliveira Carvalho, Jardin, Pereira, Folador, Le Loir, Figueiredo, Jan and Azevedo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Gwénaël Jan, Z3dlbmFlbC5qYW5AaW5yYS5mcg==
†These authors shared senior authorship.