
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 06 August 2018
Sec. Food Microbiology
Volume 9 - 2018 | https://doi.org/10.3389/fmicb.2018.01685
 Mohamed Zommiti1
Mohamed Zommiti1 Mélyssa Cambronel2Olivier Maillot2
Mélyssa Cambronel2Olivier Maillot2 Magalie Barreau2Khaled Sebei1
Magalie Barreau2Khaled Sebei1 Marc Feuilloley2
Marc Feuilloley2 Mounir Ferchichi1,3
Mounir Ferchichi1,3 Nathalie Connil2*
Nathalie Connil2*Enterococcus faecium strains were isolated from an original biotope, artisanal dried Tunisian meat “Dried Ossban,” and evaluated for safety and capacity as probiotics. Gram-positive, catalase negative, and bacteriocin-producing bacteria were screened using selective microbiological media. All isolates were identified by phenotypic and molecular tools. Five E. faecium strains (MZF1, MZF2, MZF3, MZF4, and MZF5) were selected and further assessed for their probiotic properties. They were found to be resistant to the physiological concentrations of bile salts, and the harsh conditions of the gastrointestinal tract, and showed autoaggregation and adhesion ability. All these isolates possess at least one enterocin and could efficiently inhibit the growth of Listeria innocua HPB13. The analysis of their safety profile revealed for almost all the strains the absence of cytotoxicity and virulence determinants, and susceptibility to clinically important antibiotics such as vancomycin. These data suggest that these bacteria, isolated from “Dried Ossban,” do not present a risk to human health, and may be considered as interesting candidates for future use as probiotics and bioprotective cultures for application in the food and/or feed industries.
Enterococci are ubiquitous microorganisms, widespread in the environment. In fact, they represent the common members of the commensal microbiota in the intestine of humans, mammals, and other animals encompassing reptiles, birds, and insects, but they are also found in other complex ecosystems such as soil, plants, water, waste, food, and feed (Foulquié Moreno et al., 2006; Byappanahalli et al., 2012). Moreover, Enterococcus species are widely present in traditional fermented foods (Giraffa, 2003) due to their amazing capacity to withstand extreme temperature (Murray, 1990), high salinity, and pH levels (Franz et al., 2011). They can grow in the presence of 40% (w/v) bile salts (Fisher and Phillips, 2009). These special characteristics of adaptability to various matrices and conditions may also allow these bacteria to cause spoilage of a wide range of food products, particularly, meat and its derivatives (Björkroth et al., 2005).
Enterococci are increasingly investigated as potential probiotic candidates. Currently, two strains belonging to the genus Enterococcus are identified as probiotics and are available on the market, also known as Enterococcus faecium SF68® (NCIMB 10415, Cerbios-Pharma SA, Barbengo, Switzerland) and Enterococcus faecalis Symbioflor 1 (SymbioPharm, Herborn, Germany). Bybee et al. (2011) have demonstrated the probiotic potential of E. faecium SF68® to prevent and treat diarrhea in pets, and E. faecalis Symbioflor 1 has been proved efficient for patients suffering from chronic sinusitis and/or bronchitis (Franz et al., 2011). The assessment of the effectiveness of probiotic strains such as E. faecium SF68® (NCIMB 10415) and E. faecalis Symbioflor 1 in humans for the treatment of diarrhea, irritable bowel syndrome, in lowering serum cholesterol and for immune regulation has been investigated in very few studies (Franz et al., 2011; Christoffersen et al., 2012; Yamaguchi et al., 2013). As a result, there are not enough data to firmly conclude on the efficiency of enterococci as probiotics for humans and their use in foods must be based on case-by-case investigations (Ogier and Serror, 2008).
Producing antimicrobial substances preventing pathogens can be a reliable criterion when selecting probiotic strains. Among lactic acid bacteria (LAB), enterococci are known to secrete various antimicrobial compounds, so these bacteria seem potentially useful to prevent diseases of bacterial foodborne origins (Franz et al., 2011). Numerous enterococci produce at least one bacteriocin, a bioactive substance of proteinaceous nature, which is ribosomally synthesized and active against a broad collection of spoiling and foodborne microbes encompassing Listeria spp. (Favaro et al., 2014). In the last decade, numerous studies have been published on bacteriocin-producing enterococci in association with food ecosystems (Todorov et al., 2010; Hadji-Sfaxi et al., 2011) and their isolation from various ecological biotopes, encompassing meat (Cintas et al., 1995), vegetables (Bennik et al., 1998), soil (Yanagida et al., 2005), crops (Todorov et al., 2005), fish (Satish Kumar et al., 2011), dairy products (Chanos and Williams, 2011), and humans (Gálvez et al., 1986).
However, Enterococci have also gained notoriety and sturdiness over the past few years as the leading nosocomial pathogens (Rosenthal et al., 2008). According to Schaberg et al. (1991), enterococcal species are well recognized as the second-most common causal agent of urinary tract infections and the third-most for nosocomial bacteraemia. Clinically, vancomycin-resistant Enterococci (VRE) represent a major problem in nosocomial infections (Arias and Murray, 2012). These bacteria often possess multiple antibiotic resistances and virulence factors such as hemolysin, adhesins, and invasins (Ogier and Serror, 2008; Franz et al., 2011). For all the reasons previously mentioned, enterococci species are generally considered to have a doubtful status for food safety (Ladero et al., 2012).
In this work, we report for the first time the isolation and characterization of five enterococci from “Dried Ossban,” a traditional Tunisian fermented meat product. Their safety, probiotic potential, and antimicrobial properties have been investigated.
Lactic acid bacteria strains were isolated from “Dried Ossban,” a Tunisian traditional dry fermented meat typically prepared from sheep intestine and meat mixed with salt and spices and dried through exposure for several days to sunlight. The samples were obtained from homemade production of fermented meat, from various governorates covering almost all the Tunisian territory. Samples were collected into sterile bottles and then transported on ice to the laboratory for analyses. For all samples, 10 g was added to 90 ml of sterile peptone saline water and homogenized for 10 min through vortexing at maximum speed. Appropriate decimal dilutions were plated onto de Man Rogosa Sharpe (MRS) agar and incubated at 37°C for 24–48 h. Colonies were randomly selected from MRS agar, and only Gram-positive and catalase-negative isolates were retained, routinely propagated, and stored at -80°C in MRS broth containing 20% glycerol. For experimental assays, working cultures were sub-cultured (1% inoculum, 24 h, 37°C) prior to use.
The Gram-positive and catalase negative LAB isolated from “Dried Ossban” were identified by analysis of the total proteome using an Autoflex III matrix-assisted laser desorption/ionization-time-of-flight mass spectrometer (MALDI-TOF MS; Bruker, Marcy-l’Etoile, France) coupled to the MALDI-Biotyper 3.1 algorithmic system for microbial identification. Briefly, each bacterium was cultured on MRS agar for 24 h and the colonies were analyzed by direct spot on the MALDI target plate or using the 70% formic acid protein extraction method, as previously described (Hillion et al., 2013).
The MALDI target plate was introduced into the MALDI-TOF MS for automated measurement and data interpretation. The instrument was calibrated using a Bruker bacterial test standard and spectra were acquired in the linear mode over a mass range from 2000 to 20,000 Da. The spectrum of each unknown bacterium was electronically transformed into the peak list and compared to the 6903 reference organisms in the database and a log(score) value between 0.00 and 3.00 was generated which represents the probability that the match is correct. A score of ≥1.7 indicates genus identification, and a score of ≥2.0 is the set threshold for determination at the species level (Sogawa et al., 2011).
For the acid tolerance assays, MRS broth was used to simulate the acidity of the gastrointestinal tract after adjusting to pH 3.0 with 1 N HCl. Fresh overnight cultures of E. faecium strains isolated from “Dried Ossban” and previously identified by MALDI-TOF MS were centrifuged (8000 rpm, 10 min), washed with phosphate buffer saline (PBS), and resuspended to approximately 107 bacteria/ml in the MRS broth pH 3.0. Samples (0.1 ml) were taken at 0 h and after 1, 2, and 3 h incubation at 37°C, simulating the time spent in the human stomach. The number of bacterial cells was then enumerated by the pour plate method of all samples using decimal serial dilutions prepared in PBS, and after 24 h incubation at 37°C. Survival bacteria were expressed as log values of colony-forming units per ml (CFU/ml).
The bile salt tolerance was tested as previously described by Anandharaj et al. (2015) with slight modifications. Bacteria from overnight cultures were inoculated (1% v/v) into MRS broth containing 0.3% (w/v) Ox-bile (Sigma–Aldrich). Total viable counts were determined after 0, 1, 2, and 3 h of exposure to bile salts, reflecting the time that food spends in the small intestine. Aliquots were diluted, plated on MRS agar, and incubated at 37°C for 24 h. Samples without addition of bile salts served as controls. Survival bacteria were expressed as described above for acid tolerance.
Autoaggregation of the E. faecium strains isolated from “Dried Ossban” was assessed according to Collado et al. (2008). Overnight cultures were centrifuged (8000 rpm, 10 min), washed twice with PBS, and resuspended in the same buffer to approximately 108 bacteria/ml. Each suspension (4 ml) was vortexed for 20 s and incubated at room temperature. The absorbance at 600 nm (A600) of the upper part of each suspension was measured at time 0 h and 24 h after incubation without vortexing. The percentage of autoaggregation of each strain was then calculated according to the following equation:
Autoaggregation (%) = [1 - (ATime/A0) × 100], where ATime refers to the absorbance of the suspension at 24 h and A0 refers to the absorbance at time 0.
The human colon adenocarcinoma cell line Caco-2/TC7 (passages 40–60) were routinely grown in Dulbecco’s Modified Eagle’s Medium (DMEM, Invitrogen, France), supplemented with 15% heat-inactivated fetal calf serum (FCS), and 100 U/ml each of penicillin and streptomycin. The cells were cultivated at 37°C in 5% CO2–95% air atmosphere, and the medium was regularly changed. For adhesion and cytotoxicity assays, the cells were seeded in 24-well tissue culture plates and incubated to confluence, and for transepithelial electrical resistance (TEER) measurements, they were grown on inserts (3 μm pore size) until full differentiation (21 days).
E. faecium strains, grown overnight in MRS broth, were harvested by centrifugation (8 000 rpm, 10 min), washed twice in PBS solution, and resuspended in cell culture medium, without serum and antibiotic, at a concentration of 108 bacteria/ml, and then applied on confluent Caco-2/TC7 monolayers as previously described for other bacteria (Messaoudi et al., 2012). Briefly, after 3 h of incubation at 37°C, in 5% CO2–95% air atmosphere, monolayers were gently washed three times with sterile pre-warmed PBS, to remove non-adherent bacteria, and disrupted by incubation for 15 min with 0.1% Triton X100. The lysates were then diluted and plated onto MRS agar to determine the number of adherent bacteria.
The cytotoxicity was determined using an enzymatic assay (Cytotox 96 Promega, France) which measures lactate dehydrogenase (LDH) released from the cytosol of damaged Caco-2/TC7 cells into the supernatant. LDH is a stable cytosolic enzyme present in many different cell types, particularly in eukaryotic cells, playing the role of an indicator of necrotic cell death when released. After overnight incubation with the E. faecium strains (108 bacteria/ml), the supernatants from confluent Caco-2/TC7 monolayers grown on 24-well tissue culture plates were collected and the concentration of the LDH was quantified. Two controls were included for calculation of cytotoxicity percentage. Caco-2/TC7 cells exposed to Triton X100 (0.9%) were used as a positive control of maximal LDH release (100% lysis) as specified by the manufacturer’s recommendations. The background level (0% LDH release) was determined with serum-free culture medium.
The TEER of differentiated Caco-2/TC7 cells was monitored during 24 h using the Millicell Electrical Resistance System (Millipore Corp., Bedford, MA, United States). To study the potential effect of E. faecium strains on the epithelial barrier integrity, these bacteria were incubated at 108 bacteria/ml on the Caco-2/TC7 cell monolayers. Control monolayers were not exposed to the potential probiotics. TEER values were expressed as percentages of the initial level measured in the insert.
Bacterial cells from overnight cultures in MRS broth of the five E. faecium strains identified from “Dried Ossban” were harvested by centrifugation at 8000 rpm for 10 min. Total genomic DNA was extracted and highly purified via GeneJET Genomic DNA Purification Kit (Thermo Scientific, France) according to the manufacturer’s recommendations. DNA quality and concentration were determined by measuring optical density (OD) at 260 and 280 nm with a spectrophotometer.
The virulence factors of the E. faecium strains were investigated by PCR amplification, revealing the presence of genes encoding for aggregation substance (agg), gelatinase (gelE), enterococcal surface protein (esp), and collagen adhesin (ace), (Eaton and Gasson, 2001; Reviriego et al., 2005). Primer sequences and fragment sizes are presented in Table 1. PCR for virulence determinant genes was performed using PCR protocols of (Mannu et al., 2003; Reviriego et al., 2005). The amplified products were separated by electrophoresis on 1% (w/v) agarose gels in 1× Tris-acetate-EDTA (TAE) buffer. Gels were stained with SYBR Safe DNA gel stain and visualized under UV light.
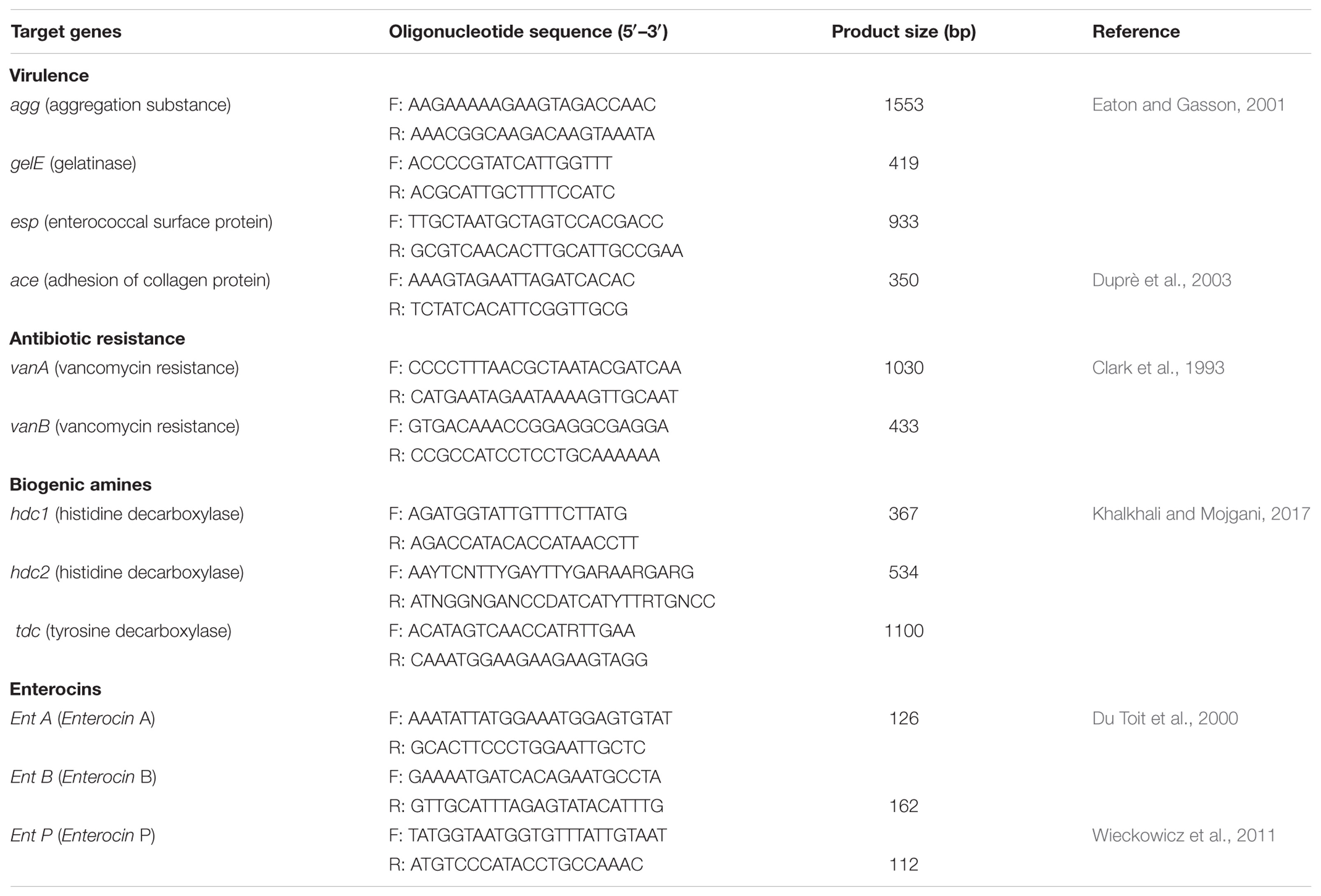
TABLE 1. PCR primers used for detection of virulence determinants, antibiotic resistance, biogenic amines, and enterocin genes.
The strains were also evaluated for vanA and vanB genes (both related to vancomycin resistance) by PCR reaction according to the protocol of Clark et al. (1993). The primer sequences are described in Table 1.
The potential of the E. faecium strains to produce histamine and tyramine was assessed via molecular tools. Genes related to the production of these biogenic amines are part of the genetic heritage of Enterococcus spp. (Connil et al., 2002). Analysis of the histidine and tyrosine decarboxylase encoding genes (hdc and tdc, respectively) in MZF1–MZF5 strains was carried out by PCR amplification with the primers previously used by Khalkhali and Mojgani (2017). PCR primers and sizes of the resulted amplicons are listed in Table 1. PCR reactions were performed using the following cycling parameters: initial denaturation for 5 min at 95°C followed by 35 cycles of 1 min at 95°C, annealing at 54°C for 30 s, and 1 min at 72°C and a final extension step of 5 min at 72°C.
The presence of the main enterocin genes was screened by PCR amplification. Total DNA from the five E. faecium strains was submitted to PCR reactions to detect genes responsible for codification of the following bacteriocins: enterocin A, enterocin B, and enterocin P (Table 1). The PCR amplification conditions were as already described (Du Toit et al., 2000; Wieckowicz et al., 2011).
For detection of antibacterial activity, a well agar diffusion test of the E. faecium isolates was carried out as previously (Jacobsen et al., 1999) using smooth brain heart infusion (BHI) agar, containing the indicator strains (Listeria innocua HPB13, E. faecalis ATCC 29212, Staphylococcus aureus ATCC 25923, or Escherichia coli DH5α).
The antibiotic susceptibility of the E. faecium strains was checked by the broth microdilutions method. The antibiotics tested were ampicillin, vancomycin, gentamycin, erythromycin, ofloxacin, tetracycline, and chloramphenicol (Table 2). In brief, 100 μl of E. faecium strains (1.105 bacteria/well final concentration) were inoculated into the wells of a 96-well plate containing 100 μl of each antibiotic in serial twofold dilutions from 512 to 0.125 μg/ml, and then were incubated for 24 h. The results were compared with growth control (enterococci alone), and the minimum inhibitory concentration (MIC) breakpoint is the concentration where no growth is observed. The level of susceptibility to antibiotics was reported as sensitive, intermediate or resistant according to the observed breaking points recommended for enterococci by the Clinical and Laboratory Standards Institute (CLSI) antimicrobial susceptibility testing standards (CLSI, 2015).

TABLE 2. List of antibiotics used in the study.
Data are expressed as a mean ± standard error (SE) calculated over three independent experiments performed in triplicate. Analysis of statistical significance was performed by ANOVA. Student’s t-test was used, when necessary, to discriminate differences between means. Differences with P-value < 0.05 were considered statistically significant.
Thirty-seven samples of artisanal dried Tunisian meat “Dried Ossban” were collected from homemade production in different governorates of the Tunisian territory to isolate potential probiotics from the lactic acid flora. The strains were first presumptively identified by phenotypic analysis (Gram staining, cellular morphology, catalase, and oxidase tests). Five presumptive Enterococcus strains (Gram+ coccus, catalase, and oxidase negative) were selected for further study and called MZF1, MZF2, MZF3, MZF4, and MZF5. Their growth in various conditions of temperature, salinity, and pH was examined. All these strains were able to grow at 45°C, in 6.5% NaCl, and at pH 9.6 but not pH 4. These characteristics are well known for Enterococcus as this genus has an extraordinary capacity to grow under hostile conditions. MZF1, MZF2, MZF3, MZF4, and MZF5 were then analyzed by Maldi Biotyper for confirmation of the genus Enterococcus and determination of the species. The five strains were identified as E. faecium with score > 2 and accordingly to the library of Maldi Biotyper, a dendrogram was generated from the minimal spanning tree (MSP) data set (Elbehiry et al., 2017). For this, the main spectra of the Maldi Biotyper taxonomy were compared with the spectra resulting in a matrix of cross-wise identification scores. This matrix was applied to estimate the distance level for each pair of main spectra. A dendrogram was created according to the distance level between the E. faecium strains isolated from Tunisian “Dried Ossban” (Figure 1A) and with the E. faecium reference strains available in the Biotyper library (Figure 1B). This phyloproteomic analysis shows the hierarchical relationship between the five isolates and suggests that E. faecium MZF5 is more distant from the four other “Dried Ossban” strains.
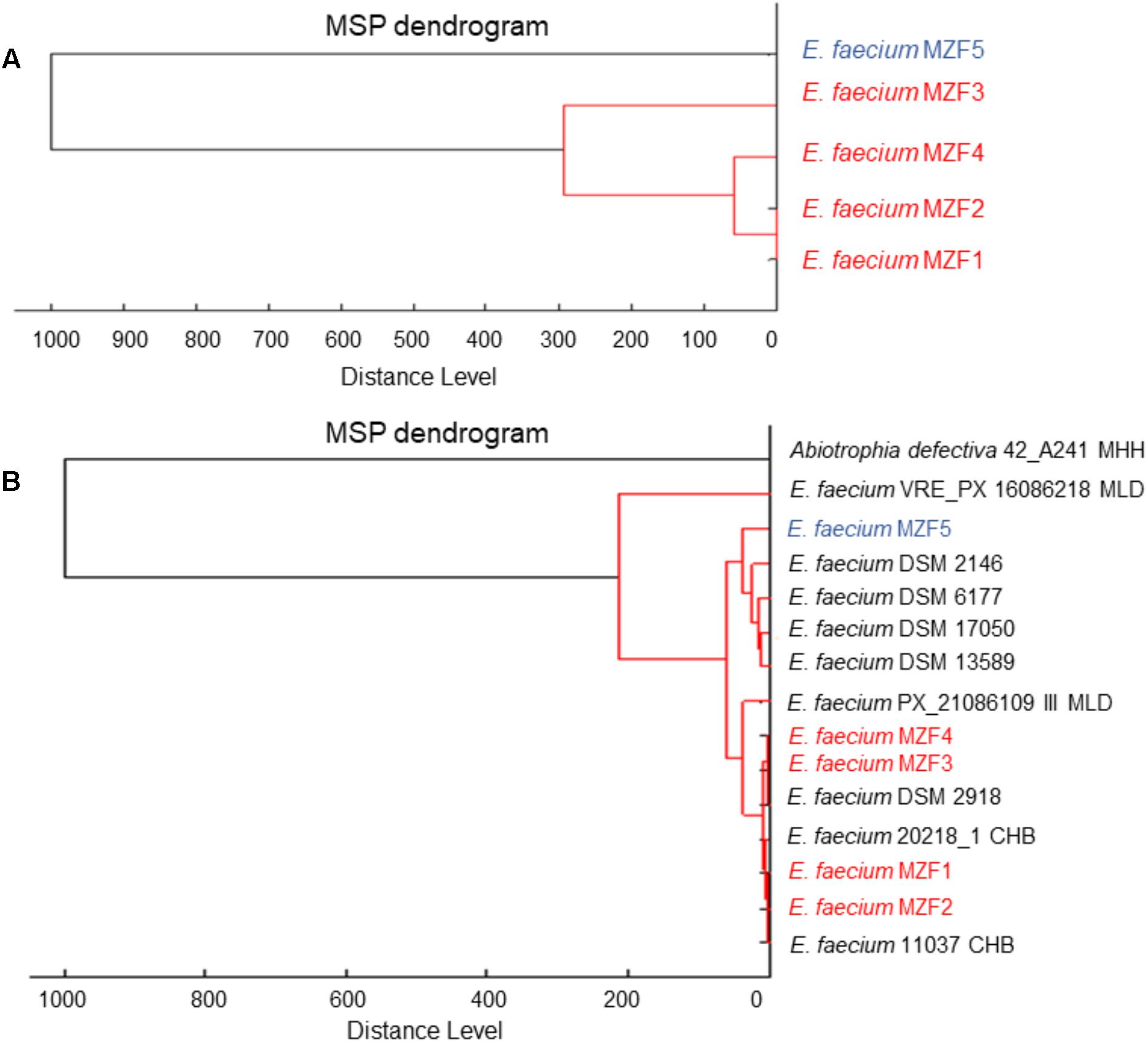
FIGURE 1. MALDI-TOF MS based phylogenetic tree. (A) Distance level between the E. faecium strains isolated from Tunisian “Dried Ossban” and (B) distance level of the isolates compared to the E. faecium reference strains available in the Biotyper library.
Simultaneously, a PCR amplification with the E. faecium specific primers (EfmF1 5′-ACGGAGATCGTGGATTCAAA-3′ and EfmR1 5′-CGTACGGGAAGTGATTCGAC-3′) from Park et al. (2017) has been done and gave positive results. Indeed, as expected, a PCR product of 1032 bp was obtained, corresponding to the specific amplification of the M protein trans-acting positive regulator target gene of E. faecium (data not shown). Finally, 16S rRNA genes of the five E. faecium isolates (MZF1, MZF2, MZF3, MZF4 and MZF5) have been sequenced and the partial nucleotide sequences have been deposited in the NCBI Genbank under the accession numbers MH569603, MH569604, MH588167, MH591462, and MH591463, respectively.
The efficiency of a bacterium as a potential probiotic depends firmly on its aptitude to survive the passageway through the upper digestive tract to the intestine, where its beneficial effect is expected, and this represents a crucial requirement (Marteau et al., 1997; Tuomola et al., 2001). According to Maragkoudakis et al. (2006), the pH inside the human stomach ranges from 1 (during fasting) to 4.5 (after a meal), and food ingestion can take up to 3 h. Our findings indicated that the five isolates exhibited high tolerance to acidity after exposure to media pH 3 (Table 3) and were also tolerant to bile salts, since all of them survived successfully in MRS broth medium supplemented with 30 g/l bile salt, reflecting the physiological concentration of human bile (Table 4). Our results are in total accordance with data published by other authors for Enterococcus spp. isolated from food niches suggesting that these “Dried Ossban” strains could have the potential to reach the intestinal lumen and thus stay alive in that environment (Ahmadova et al., 2013; Gupta and Tiwari, 2015; Aspri et al., 2016).
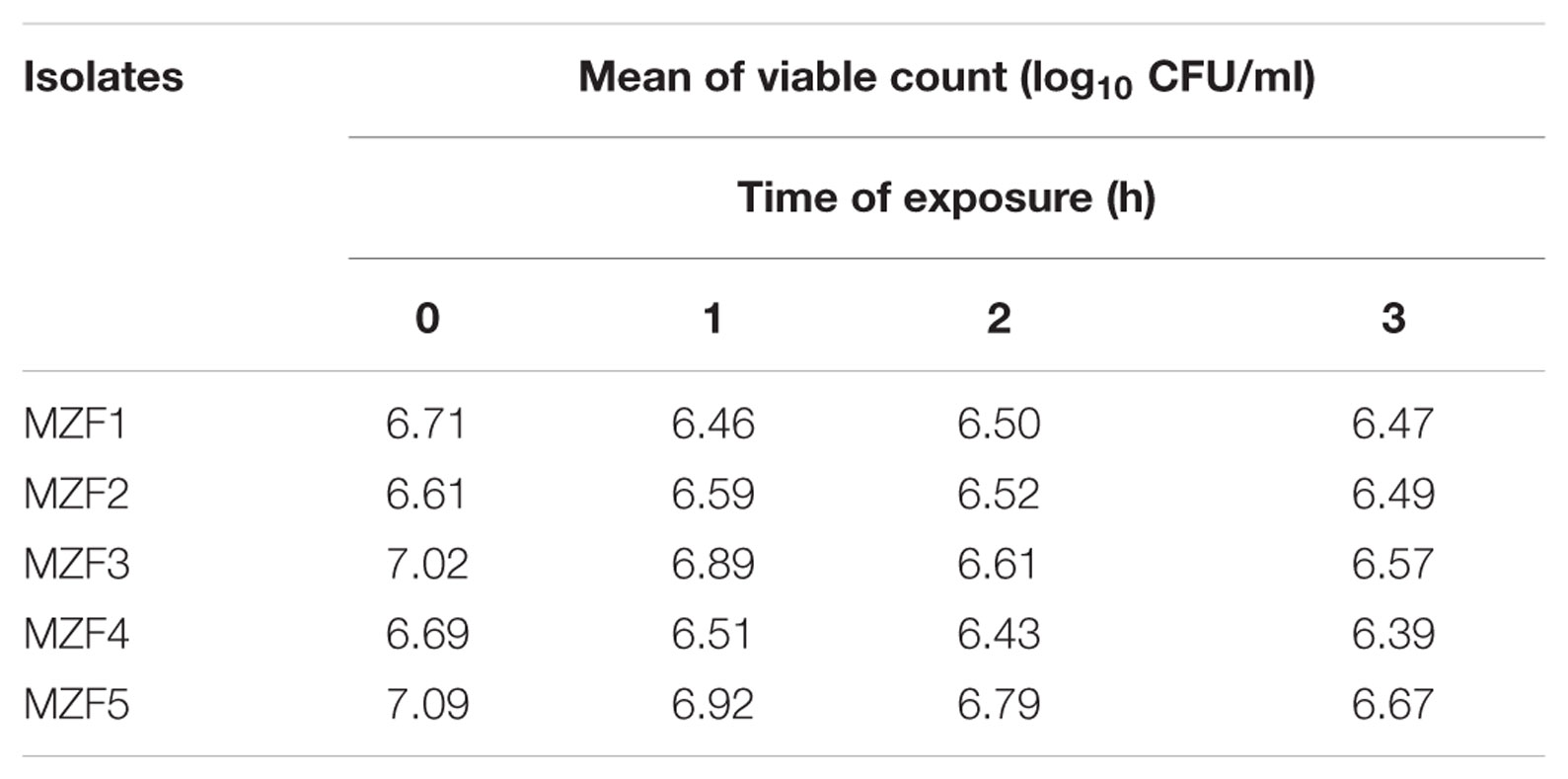
TABLE 3. Low pH and acidic conditions tolerance.
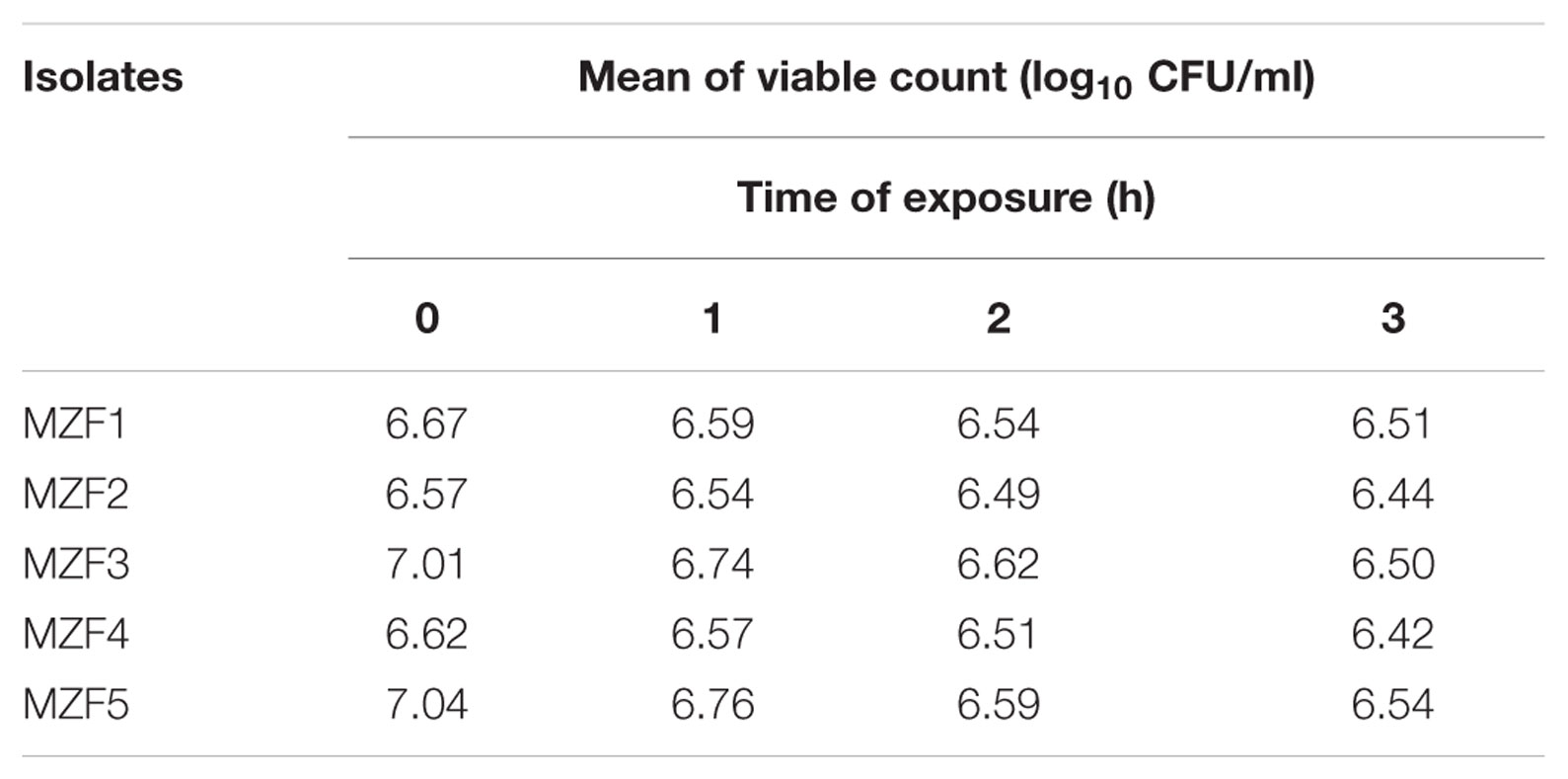
TABLE 4. Bile salts tolerance.
According to the investigations of Collado et al. (2007), cell aggregation properties represent one of the most important phenotypic characteristics of a potential probiotic strain. In this study, the ability of the enterococcal strains, isolated from “Dried Ossban,” to aggregate was evaluated. The results showed autoaggregation values ranged from 50 to 96% (Figure 2). MZF4 showed the lower level, 50%, whereas MZF2 and MZF1 showed the highest levels with 94 and 96%, respectively. These findings are supported by previous data demonstrating high values of autoaggregation for E. faecium strains (Dos Santos et al., 2015; Veljović et al., 2017). Autoaggregation proved to be strain-specific and may vary inside the same taxonomic group (Dos Santos et al., 2015). This strain specificity in autoaggregation has been reported in a study led by Todorov et al. (2008) for Lactobacillus pentosus ST712BZ and Lactobacillus paracasei ST284BZ. Bacterial autoaggregation is considered an important phenomenon in several ecological niches (Del Re et al., 2000). Among LAB, enterococcal species represent beneficial members of the microbiota residing the human and animal gastrointestinal and urogenital tracts (Bhardwaj et al., 2009). One beneficial feature of bacteria with autoaggregative potential is the ability to prevent the colonization of pathogenic bacteria via the formation of a barrier through autoaggregation to intestinal mucosa (Prince et al., 2012).
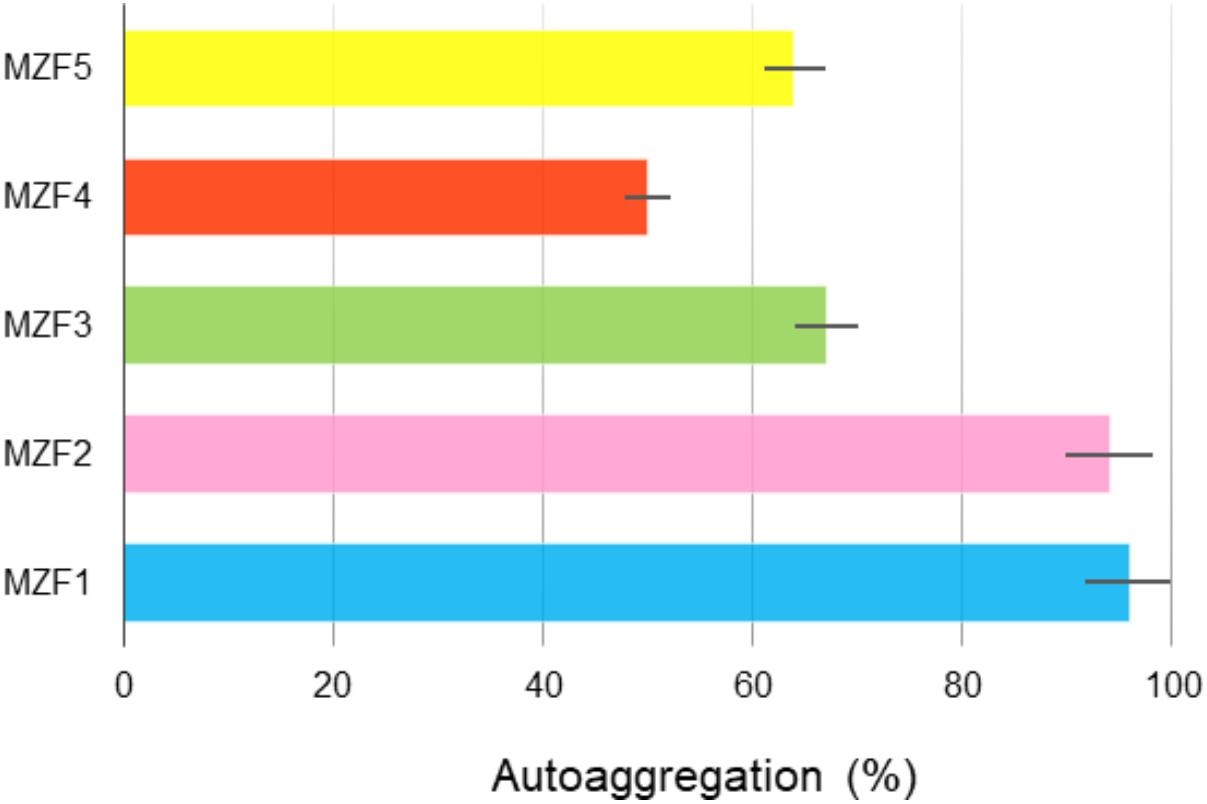
FIGURE 2. Autoaggregation of the E. faecium strains isolated from “Dried Ossban.” Data are expressed as mean ± SE.
Saarela et al. (2000) demonstrated the importance of the adhesion step. Once the probiotic bacteria have reached the gut, the adhesion to the intestinal mucosa and epithelial cells represents a requirement step for colonization. Probiotics must adhere to the mucus layer to prevent being removed from the colon by peristalsis. To complete the study of the E. faecium isolated from “Dried Ossban,” in vitro assessment of their adhesive potential to human colon adenocarcinoma epithelial cells (Caco-2/TC7 cell line) was conducted. Figure 3 shows three isolates with very low adhesion percentages, 0.7, 0.9, and 1.4% for MZF1, MZF2, and MZF4, respectively. A moderate value is noticed with MZF3 isolates, 7.8% of adhesion, and the highest value is accorded to the MZF5 isolate with more than 21% of adhesive potential to Caco-2/TC7 cells. This agrees with other studies demonstrating strong adhesiveness for E. faecium BGGO9-28 (Veljović et al., 2017) and other LAB strains than Enterococcal species (Juntunen et al., 2001). Our findings showed a variability of results among the isolates. This is in line with Turpin et al. (2012) who reported that 30 LAB strains displayed adhesion capacities from 0.6 to 30.0% on the HT-29 cells. For the MZF5 isolate with 21.8% of adhesion capacity, this value was greater than that reported for the E. faecalis UGRA10 (Cebrián et al., 2012). It is significant to highlight that in vitro experiments have many restrictions, since adhesion properties and mucus production are strictly cell line-dependent in every single study (Turpin et al., 2012). In vitro test results using Caco-2 cells for the evaluation of adhesion ability to intestinal epithelium cells have been reported to have a good correlation with in vivo results (Crociani et al., 1995) and this characteristic is often strain-specific (Duary et al., 2011).
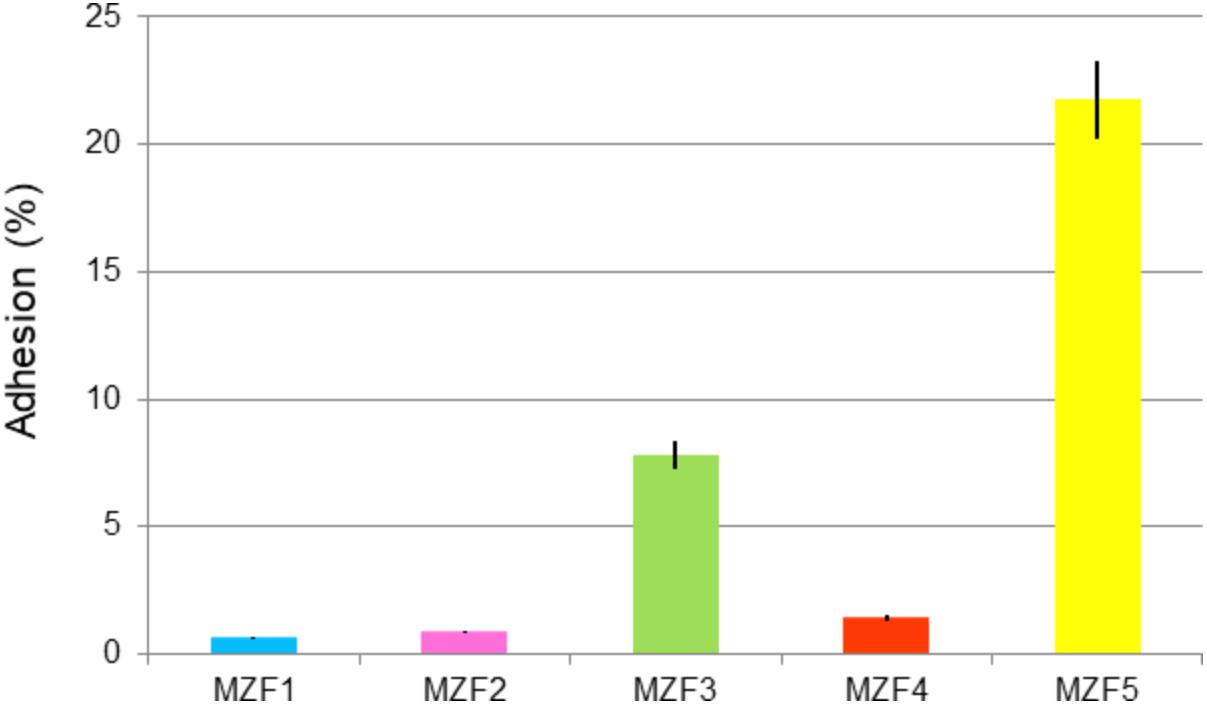
FIGURE 3. Adhesion of the E. faecium isolates to the intestinal Caco-2/TC7 cells. Data are expressed as mean ± SE.
As E. faecium contains concurrently harmless and pathogenic strains, a cytotoxicity assay was needed to ensure the safety of MZF1, MZF2, MZF3, MZF4, and MZF5. The five strains were applied overnight on Caco-2/TC7 monolayers and the cytotoxicity was estimated by measurement of LDH release and microscopic observation. Our findings showed that the weakest level of cytotoxicity is attributed to the MZF3 isolate with 4% of mortality, reflecting a high rate of viability, about 96%. This isolate exhibited no significant effect on the viability of Caco-2 cells compared to cells alone. MZF1 and MZF2 showed about 8 and 6% of cytotoxicity rate; 15 and 13% of the mortality was obtained with strains MZF4 and MZF5, respectively (Figure 4). These results are in harmony with previous studies showing that various probiotic bacteria had no cytotoxic effect on Caco-2 cells. For instance, Er et al. (2015) studied the cytotoxicity of three LAB (Pediococcus pentosaceus, Lactobacillus plantarum, and Weissella confusa) on Caco-2 cells and showed dose-dependent cytotoxic effects of the cell-free filtrates and cell-free lyophilized filtrates of the three LAB. Similarly, a recent work performed by Castro et al. (2016) demonstrated that E. faecalis CECT7121 strain is not cytotoxic for Caco-2 cells.
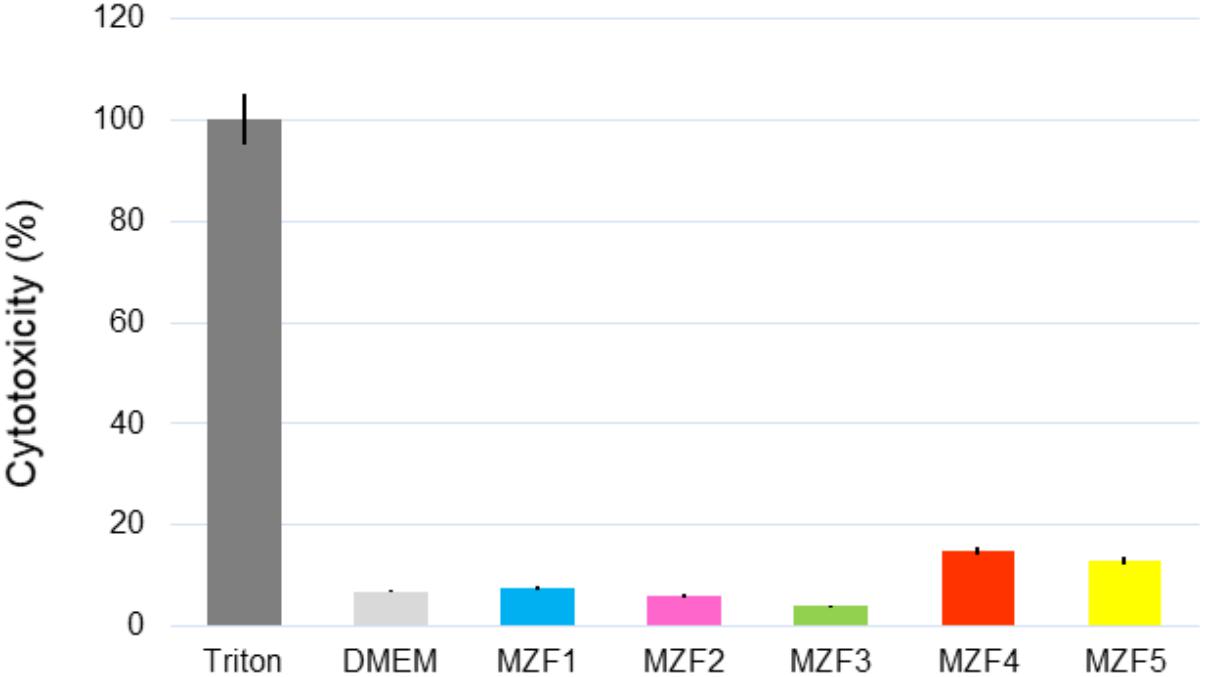
FIGURE 4. Cytotoxicity of the E. faecium isolates measured by LDH release after overnight incubation of Caco-2/TC7 cells with 108 bacteria/ml. Data are expressed as mean ± SE.
Measuring the TEER is a method for evaluating the intestinal barrier integrity in vitro (Schneeberger and Lynch, 2004). Results found for the effect of “Dried Ossban” isolates on Caco-2/TC7 monolayers are presented in Figure 5. After 24 h of incubation, MZF1 and MZF3 showed no significant impact on TEER, whereas MZF2 and MZF4 led to a slight 20% increase of TEER, and MZF5 a 30% increase. These findings are in agreement with the results obtained for other well-known and characterized probiotics [Bifidobacterium infantis, Lactobacillus acidophilus, Lactobacillus rhamnosus, E. coli Nissle 1917, Streptococcus thermophilus, the probiotic complex VSL#3] on different cell lines (T84, HT29/cl.19A, and Caco-2) (Resta-Lenert and Barrett, 2003; Sherman et al., 2005; Ewaschuk et al., 2008). Indeed, it has been demonstrated that the application of probiotics on intestinal epithelial cells can give rise to two kinds of response. The first one was a significant enhancement of TEER values (Resta-Lenert and Barrett, 2003; Ewaschuk et al., 2008; Klingspor et al., 2015), while the other type of response was no effect on TEER measurements (Sherman et al., 2005). Furthermore, the effects were frequently showed to be dose- and time-dependent (Klingberg et al., 2005).
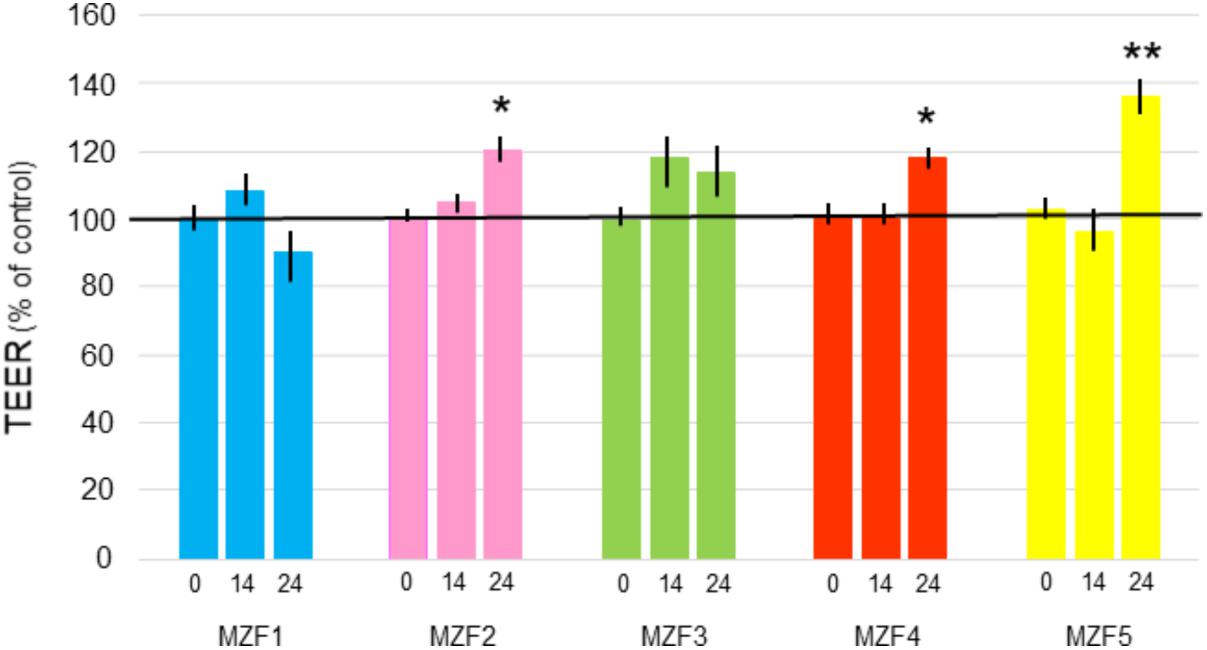
FIGURE 5. TEER at time 0, 14, and 24 h of Caco-2/TC7 cells exposed to the E. faecium isolates (108 bacteria/ml). Data are expressed as percentages of the initial level measured in the insert and as a mean ± SE. ∗P < 0.05 and ∗∗P < 0.01 compared to Caco-2/TC7 control.
Virulence determinants greatly contribute to enhancing infection because of the risk of genetic transfer. Since these genes are usually located in conjugative plasmids (Eaton and Gasson, 2001), potential virulence genes of E. faecium, isolated from food sources, need to be assessed. Therefore, it seemed needful to test the five E. faecium isolates for their virulence factors profile (Table 5). Results showed that only the isolate MZF1 carried the agg gene, whereas gelE gene was detected in MZF1 and MZF4. The MZF2, MZF3, and MZF5 isolates do not harbor any of the tested virulence genes. Our findings are consistent with the data reported by Zheng et al. (2015), proving the absence of these virulence factors in food-isolated E. faecium strains, and with the study conducted by Eaton and Gasson (2001) who showed that the presence or absence of virulence factors is strain-specific. Indeed, Eaton and Gasson (2001) and Ahmadova et al. (2013) reported that a high frequency of the esp gene is only found in medical E. faecium strains. Besides, the presence of some virulence determinants such as agg and gelE genes in MZF1 and MZF4 cannot be considered as a negative trend since some commercial enterococci starter cultures with a long history of safe use are also known to possess several virulence genes such as ace, asa1, gelE, and esp. Moreover, Cebrián et al. (2012) and Popović et al. (2018) suggested that some virulence factors as the agg substance and the esp surface protein may play a beneficial role for probiotic bacteria. In fact, this may help the bacterium colonizing the gastrointestinal tract, and proliferating inside it, revealing its probiotic properties.
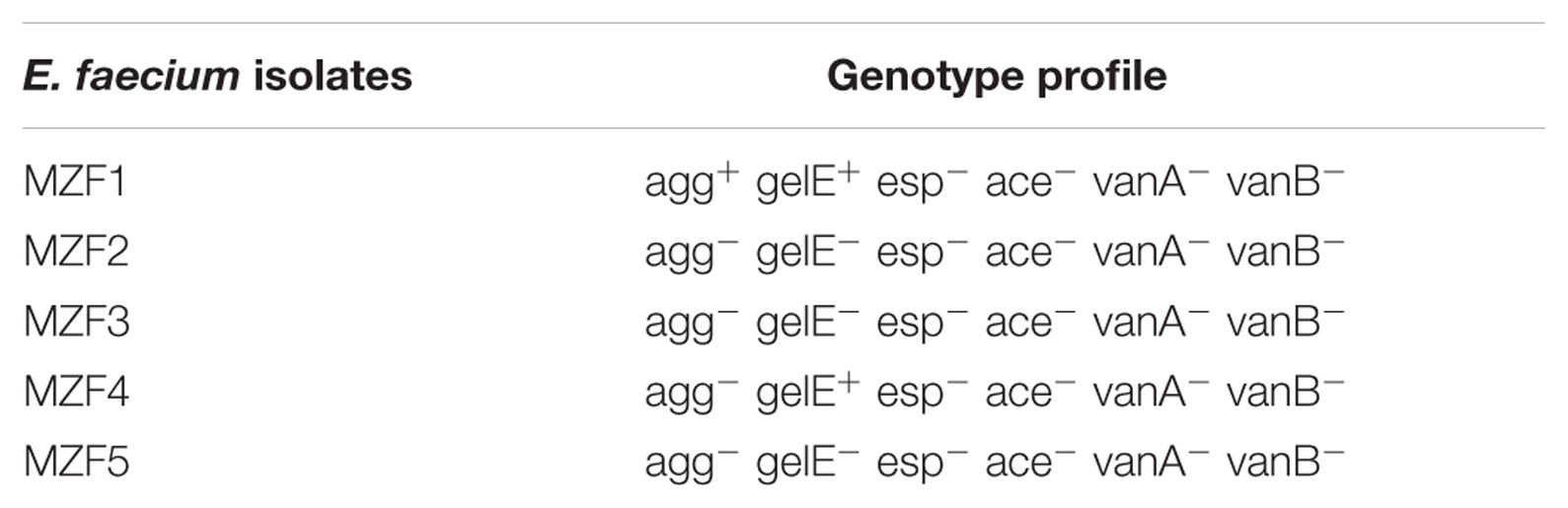
TABLE 5. PCR results of virulence and antibiotic resistance genes.
The isolated DNA of the E. faecium strains was subjected to PCR amplification to detect the presence of histidine and tyrosine decarboxylase encoding genes (hdc and tdc, respectively). All the five strains showed the expected amplicon of 1100 bp corresponding to the tdc gene (Figure 6) but none harbored the histidine decarboxylase encoding gene, hdc. Our findings are in line with studies conducted by Inoǧlu and Tuncer (2013), Ruiz et al. (2016), and Avci and Özden Tuncer (2017) where the authors found that all the analyzed E. faecium strains did not harbor the hdc gene but had the tdc gene. Histamine is the more toxic biogenic amine but the level of tyramine in food must also be controlled. A physiological assay showed that the five “Dried Ossban” strains were not, or weak producers of tyramine (data not shown). This will have to be confirmed later by a direct inoculation test of these bacteria in meat product.
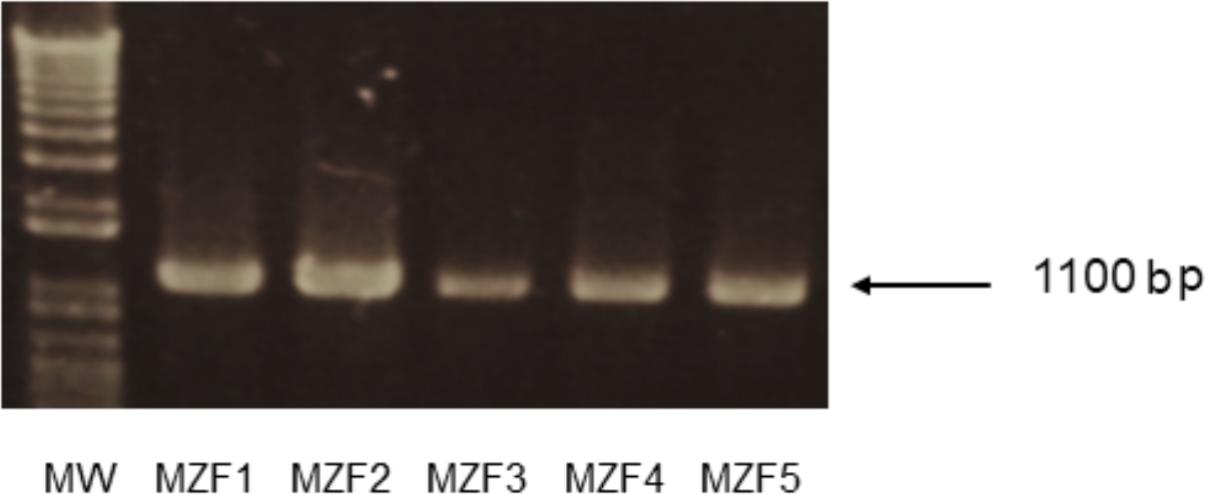
FIGURE 6. PCR amplification of the tyrosine decarboxylase (tdc) gene in the five E. faecium strains isolated from “Dried Ossban” Molecular Weight marker (MW) 1 kB.
Antibiotic resistance represents an essential issue in the safety evaluation of Enterococcus spp. Based on the broth microdilution procedure, all enterococci isolates showed sensitivity toward ampicillin and vancomycin (Table 6) which represent the commonly used antibiotics to treat enterococcal infections (Favaro et al., 2014). Vancomycin susceptibility was expected as PCR detection of the antibiotic resistance genes vanA and vanB gave negative results (Table 5). The five “Dried Ossban” strains were also sensitive to chloramphenicol, gentamicin, and tetracyclin, whereas they showed resistance to erythromycin (Macrolides) and ofloxacin (Quinolones).
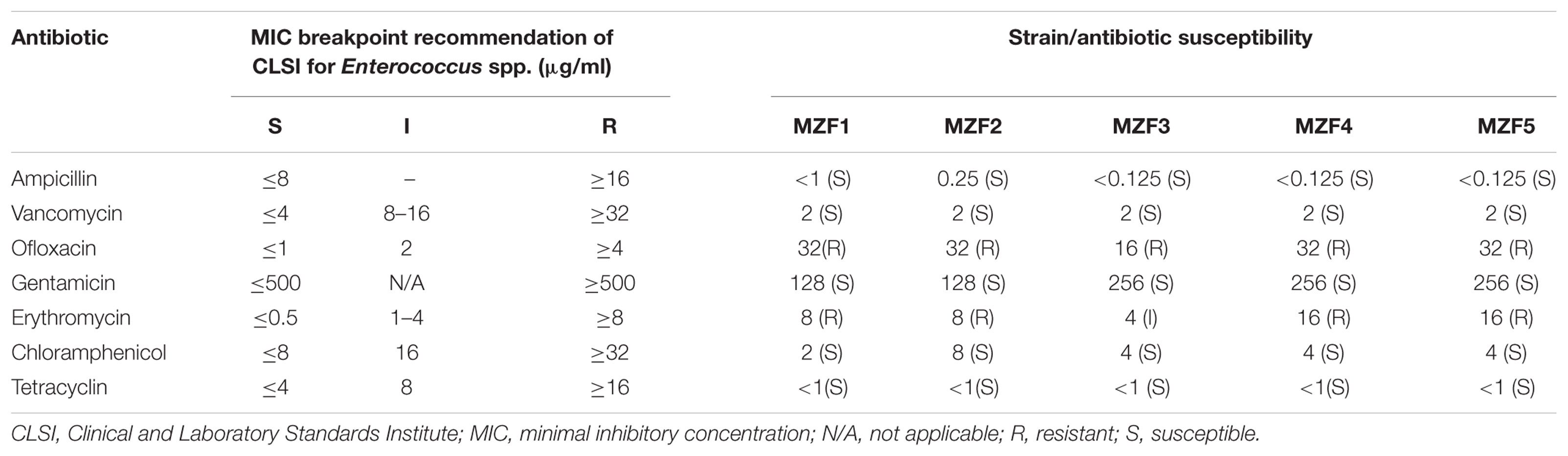
TABLE 6. Antibiotic susceptibility.
The major criterion for the safety assessment of enterococci is their susceptibility to glycopeptides such as vancomycin. Fortunately, Franz et al. (2001) reported that vanA and vanB genes are rarely detected in enterococci isolates from food sources, which is in congruence with our results. Moreover, whereas many strains of Enterococcus are known to be intrinsically resistant to β-lactam antibiotics (de Fátima Silva Lopes et al., 2005), the five E. faecium isolated from “Dried Ossban” were sensitive to ampicillin. Concerning aminoglycosides, high rates of resistance have been previously noticed in enterococcal strains from dairy products (Hammad et al., 2015; Aspri et al., 2016). On the contrary, all the “Dried Ossban” isolates were sensitive to gentamicin. High levels of tetracycline resistance have been found previously in Enterococci from animal origin, and this may be due to the wide use of this antibiotic in husbandry activities (Busani et al., 2004). It is worth noting that on the contrary, the strains isolated from “Dried Ossban” were all sensitive to tetracycline. These bacteria were also sensitive to chloramphenicol, and this is in harmony with previous studies (Zheng et al., 2015; Aspri et al., 2016). Conversely, a high rate of erythromycin resistance was detected among the “Dried Ossban” Enterococci, 80% (4/5 isolates) and the remaining 20% (1/5 isolates) were classified as intermediate. Macrolides are frequently used in animal husbandry; this could contribute to the emergence of many resistant strains (Diarra et al., 2010).
For safety reasons, a critical criterion for Enterococcus to be used in foods is the lack of transferable antibiotic resistance (Cebrián et al., 2012). All the isolates in this study showed sensitive traits to five antibiotics.
Numerous studies have reported the potential of several enterococcal species such as Enterococcus mundtii, E. faecium, and E. faecalis to produce bacteriocins, also known as enterocins, highly active against sturdy spoilage and pathogenic microorganisms in foods and food products, such as Listeria spp. (Favaro et al., 2014). Most of these enterocins have been genetically characterized; it seems that most of them are class II bacteriocins, heat stable, cationic, hydrophobic, and low molecular weight peptides. The classification of enterocins includes four main classes. Class II is subdivided into three subclasses: the pediocin family of enterocins such as enterocin A and enterocin P (IIa) with strong anti-listerial effects, bacteriocins synthesized without a leader peptide such as enterocin L50 A and B (IIb), linear enterocins that do not belong to the pediocin family (IIc) and the class (IId) including enterocin B (Aymerich et al., 1996; Casaus et al., 1997; Franz et al., 2007). Bacteriocin production is being increasingly considered as an important criterion in the selection of a probiotic strain (Dobson et al., 2012; Hegarty et al., 2016). In our study, MZF1, MZF2, MZF4, and MZF5 have been shown to be highly active against L. innocua HPB13 (Table 7). These isolates were found to harbor the EntA, EntB, and EntP genes, whereas the MZF3 isolate has been shown carrying out only the EntB gene with a weak activity against L. innocua HPB13. Similarly, Strompfová et al. (2008) found the presence of a combination of the three structural genes EntA, EntB, and EntP in various food matrices including French and Slovak meat products. Other results are also in agreement with our findings showing the presence of the combination of these three enterocin genes in E. faecium strains from food and clinical sources (De Vuyst et al., 2003; Ogaki et al., 2016).
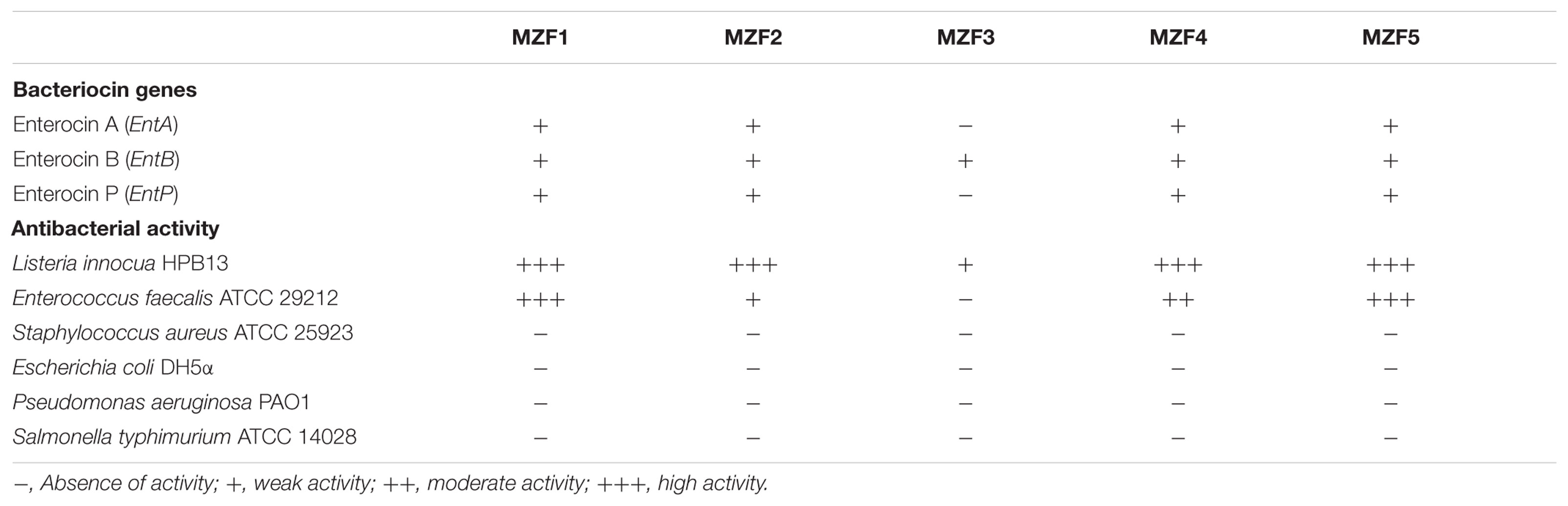
TABLE 7. Analysis for enterocins A, B, and P genes and antibacterial activity.
However, all the five E. faecium isolates displayed no inhibition of the reference strain S. aureus ATCC 25923, neither against the Gram-negative indicator bacteria used in this study. Besides, they showed inhibitory potential, strain-dependent, against E. faecalis ATCC 29212 except for MZF3 that showed no activity against this indicator strain. All the results of antibacterial activity are summarized in Table 7.
In our investigation, preliminary treatments on culture-free supernatant (CFS) have been applied in purpose to determine the nature of the antimicrobial compound. The activity of the inhibitory substance was retained in the presence of catalase indicating that the microbial inhibition is not due to H2O2, but not in the presence of proteinase K, reflecting the proteinaceous nature of the antimicrobial substance, also known as bacteriocin which is in our case an enterocin.
To our knowledge, this is the first report on the study of E. faecium isolated from a Tunisian artisanal biotope called “Dried Ossban.” The outcomes of this work claim that enterococcal isolates are resistant against the harsh gastrointestinal conditions (acidity and bile salts). The safety assessment revealed that these bacteria were susceptible to clinically relevant antibiotics such as vancomycin. They do not have the capacity to produce histamine, the more toxic biogenic amine, and most of the E. faecium strains do not harbor the virulence genes for agg, gelE, esp, and ace. Conversely, all the isolates possess at least one enterocin and could efficiently inhibit the growth of Listeria spp. These results showed that these strains, in particular MZF5, according to its high adhesion capacity, absence of cytotoxicity and virulence genes, may be considered as interesting candidates for future use as probiotics and bioprotective cultures for application in the food and/or feed industries. Further tests will be needed to evaluate their technological characteristics and behavior in meat products.
MZ carried out most of the experimental work, interpreted the data, and wrote the manuscript. MC and OM contributed to the PCR assays. MB helped with the identification of bacteria by MALDI-TOF MS. MOF and NC designed and supervised the entire project. All authors edited the manuscript, read and approved the final version of the manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
MZ is recipient of a Tunisian grant. This work was supported by Évreux Porte de Normandie, Region Normandie and European Union (FEDER), and by Tunisian Ministry of Higher Education and Scientific Research.
Ahmadova, A., Todorov, S. D., Choiset, Y., Rabesona, H., Zad, T. M., Kuliyev, A., et al. (2013). Evaluation of antimicrobial activity, probiotic properties and safety of wild strain Enterococcus faecium AQ71 isolated from Azerbaijani Motal cheese. Food Control 30, 631–641. doi: 10.1016/j.foodcont.2012.08.009
Anandharaj, M., Sivasankari, B., Santhanakaruppu, R., Manimaran, M., Rani, R. P., and Sivakumar, S. (2015). Determining the probiotic potential of cholesterol-reducing Lactobacillus and Weissella strains isolated from gherkins (fermented cucumber) and south Indian fermented koozh. Res. Microbiol. 166, 428–439. doi: 10.1016/j.resmic.2015.03.002
Arias, C. A., and Murray, B. E. (2012). The rise of the Enterococcus: beyond vancomycin resistance. Nat. Rev. Microbiol. 10, 266–278. doi: 10.1038/nrmicro2761
Aspri, M., Bozoudi, D., Tsaltas, D., Hill, C., and Papademas, P. (2016). Raw donkey milk as a source of Enterococcus diversity: assessment of their technological properties and safety characteristics. Food Control 73, 81–90. doi: 10.1016/j.foodcont.2016.05.022
Avci, M., and Özden Tuncer, B. (2017). Safety evaluation of enterocin producer Enterococcus sp. Strains isolated from traditional Turkish cheeses. Pol. J. Microbiol. 66, 223–233. doi: 10.5604/01.3001.0010.7839
Aymerich, T., Holo, H., Havarstein, L. S., Hugas, M., Garriga, M., and Nes, I. F. (1996). Biochemical and genetic characterization of enterocin A from Enterococcus faecium, a new antilisterial bacteriocin in the pediocin family of bacteriocins. Appl. Environ. Microbiol. 62, 1676–1682.
Bennik, M. H., Vanloo, B., Brasseur, R., Gorris, L. G., and Smid, E. J. (1998). A novel bacteriocin with a YGNGV motif from vegetable-associated Enterococcus mundtii: full characterization and interaction with target organisms. Biochim. Biophys. Acta 1373, 47–58. doi: 10.1016/S0005-2736(98)00086-8
Bhardwaj, A., Kapila, S., Mani, J., and Malik, R. K. (2009). Comparison of susceptibility to opsonic killing by in vitro human immune response of Enterococcus strains isolated from dairy products, clinical samples and probiotic preparation. Int. J. Food Microbiol. 128, 513–515. doi: 10.1016/j.ijfoodmicro.2008.10.010
Björkroth, J., Ristiniemi, M., Vandamme, P., and Korkeala, H. (2005). Enterococcus species dominating in fresh modified-atmosphere-packaged, marinated broiler legs are overgrown by Carnobacterium and Lactobacillus species during storage at 6°C. Int. J. Food Microbiol. 97, 267–276. doi: 10.1016/j.ijfoodmicro.2004.04.011
Busani, L., Del Grosso, M., Paladini, C., Graziani, C., Pantosti, A., Biavasco, F., et al. (2004). Antimicrobial susceptibility of vancomycin-susceptible and resistant enterococci isolated in Italy from raw meat products, farm animals, and human infections. Int. J. Food Microbiol. 97, 17–22. doi: 10.1016/j.ijfoodmicro.2004.04.008
Byappanahalli, M. N., Nevers, M. B., Korajkic, A., Staley, Z. R., and Harwood, V. J. (2012). Enterococci in the environment. Microbiol. Mol. Biol. Rev. 76, 685–706. doi: 10.1128/MMBR.00023-12
Bybee, S. N., Scorza, A. V., and Lappin, M. R. (2011). Effect of the probiotic Enterococcus faecium SF68 on presence of diarrhea in cats and dogs housed in an animal shelter. J. Vet. Intern. Med. 25, 856–860. doi: 10.1111/j.1939-1676.2011.0738.x
Casaus, P., Nilsen, T., Cintas, L. M., Nes, I. F., Hernández, P. E., and Holo, H. (1997). Enterocin B, a new bacteriocin from Enterococcus faecium T136 which can act synergistically with enterocin A. Microbiology 143, 2287–2294. doi: 10.1099/00221287-143-7-2287
Castro, M. S., Molina, M. A., Azpiroz, M. B., Díaz, A. M., Ponzio, R., Sparo, M. D., et al. (2016). Probiotic activity of Enterococcus faecalis CECT7121: effects on mucosal immunity and intestinal epithelial cells. J. Appl. Microbiol. 121, 1117–1129. doi: 10.1111/jam.13226
Cebrián, R., Baños, A., Valdivia, E., Pérez-Pulido, R., Martínez-Bueno, M., and Maqueda, M. (2012). Characterization of functional, safety, and probiotic properties of Enterococcus faecalis UGRA 10, a new AS-48-producer strain. Food Microbiol. 30, 59–67. doi: 10.1016/j.fm.2011.12.002
Chanos, P., and Williams, D. R. (2011). Anti-Listeria bacteriocin-producing bacteria from raw ewe’s milk in northern Greece. J. Appl. Microbiol. 110, 757–768. doi: 10.1111/j.1365-2672.2010.04932.x
Christoffersen, T. E., Jensen, H., Kleiveland, C. R., Dorum, G., Jacobsen, M., and Lea, T. (2012). In vitro comparison of commensal, probiotic and pathogenic strains of Enterococcus faecalis. Br. J. Nutr. 108, 2043–2053. doi: 10.1017/S0007114512000220
Cintas, L. M., Rodríguez, J. M., Fernández, M. F., Sletten, K., Nes, I. F., Hernández, P. E., et al. (1995). Isolation and characterization of pediocin L 50, a new bacteriocin from Pediococcus acidilactici with a broad inhibitory spectrum. Appl. Environ. Microbiol. 61, 2643–2648.
Clark, N. C., Cooksey, R. C., Hill, B. C., Swenson, J. M., and Tenover, F. C. (1993). Characterization of glycopeptide-resistant enterococci from US hospitals. Antimicrob. Agents Chemother. 37, 2311–2317. doi: 10.1128/AAC.37.11.2311
CLSI (2015). M100-S25 Performance Standards for Antimicrobial Susceptibility Testing; Twenty-Fifth Informational Supplement. Wayne, PA: CLSI.
Collado, M. C., Meriluoto, J., and Salminen, S. (2007). Measurement of aggregation properties between probiotics and pathogens: in vitro evaluation of different methods. J. Microbiol. Methods 71, 71–74. doi: 10.1016/j.mimet.2007.07.005
Collado, M. C., Meriluoto, J., and Salminen, S. (2008). Adhesion and aggregation properties of probiotic and pathogen strains. Eur. Food Res. Technol. 226, 1065–1073. doi: 10.1007/s00217-007-0632-x
Connil, N., Le Breton, Y., Dousset, X., Auffray, Y., Rincé, A., and Prévost, H. (2002). Identification of the Enterococcus faecalis tyrosine decarboxylase operon involved in tyramine production. Appl. Environ. Microbiol. 68, 3537–3544. doi: 10.1128/AEM.68.7.3537-3544.2002
Crociani, J., Grill, J. P., Huppert, M., and Ballongue, J. (1995). Adhesion of different bifidobacteria strains to human enterocyte-like Caco-2 cells and comparison with in vivo study. Lett. Appl. Microbiol. 21, 146–148. doi: 10.1111/j.1472-765X.1995.tb01027.x
de Fátima Silva Lopes, M., Ribeiro, T., Abrantes, M., Figueiredo Marques, J. J., Tenreiro, R., and Crespo, M. T. B. (2005). Antimicrobial resistance profiles of dairy and clinical isolates and type strains of enterococci. Int. J. Food Microbiol. 103, 191–198. doi: 10.1016/j.ijfoodmicro.2004.12.025
De Vuyst, L., Foulquié Moreno, M. R., and Revets, H. (2003). Screening for enterocins and detection of hemolysin and vancomycin resistance in enterococci of different origins. Int. J. Food Microbiol. 84, 299–318. doi: 10.1016/S0168-1605(02)00425-7
Del Re, B., Sgorbati, B., Miglioli, M., and Palenzona, D. (2000). Adhesion, autoaggregation and hydrophobicity of 13 strains of Bifidobacterium longum. Lett. Appl. Microbiol. 31, 438–442. doi: 10.1046/j.1365-2672.2000.00845.x
Diarra, M. S., Rempel, H., Champagne, J., Masson, L., Pritchard, J., and Topp, E. (2010). Distribution of antimicrobial resistance and virulence genes in Enterococcus spp. and characterization of isolates from broiler chickens. Appl. Environ. Microbiol. 76, 8033–8043. doi: 10.1128/AEM.01545-10
Dobson, A., Cotter, P. D., Ross, R. P., and Hill, C. (2012). Bacteriocin production: a probiotic trait? Appl. Environ. Microbiol. 78, 1–6. doi: 10.1128/AEM.05576-11
Dos Santos, K. M., Vieira, A. D., Salles, H. O., Oliveira Jda, S., Rocha, C. R., Borges Mde, F., et al. (2015). Safety, beneficial and technological properties of Enterococcus faecium isolated from Brazilian cheeses. Braz. J. Microbiol. 46, 237–249. doi: 10.1590/S1517-838246120131245
Du Toit, M., Franz, C. M., Dicks, L. M., and Holzapfel, W. H. (2000). Preliminary characterization of bacteriocins produced by Enterococcus faecium and Enterococcus faecalis isolated from pig faeces. J. Appl. Microbiol. 88, 482–494. doi: 10.1046/j.1365-2672.2000.00986.x
Duary, R. K., Rajput, Y. S., Batish, V. K., and Grover, S. (2011). Assessing the adhesion of putative indigenous probiotic lactobacilli to human colonic epithelial cells. Ind. J. Med. Res. 134, 664–671. doi: 10.4103/0971-5916.90992
Duprè, I., Zanetti, S., Schito, A. M., Fadda, G., and Sechi, L. A. (2003). Incidence of virulence determinants in clinical Enterococcus faecium and Enterococcus faecalis isolates collected in Sardinia (Italy). J. Med. Microbiol. 52, 491–498. doi: 10.1099/jmm.0.05038-0
Eaton, T. J., and Gasson, M. J. (2001). Molecular screening of Enterococcus virulence determinants and potential for genetic exchange between food and medical isolates. Appl. Environ. Microbiol. 67, 1628–1635. doi: 10.1128/AEM.67.4.1628-1635.2001
Elbehiry, A., Marzouk, E., Hamada, M., Al-Dubaib, M., Alyamani, E., Moussa, I. M., et al. (2017). Application of MALDI-TOF MS fingerprinting as a quick tool for identification and clustering of foodborne pathogens isolated from food products. New Microbiol. 40, 269–278.
Er, S., Koparal, A. T., and Kivanc, M. (2015). Cytotoxic effects of various lactic acid bacteria on Caco-2 cells. Turk. J. Biol. 39, 23–30. doi: 10.3906/biy-1402-62
Ewaschuk, J. B., Diaz, H., Meddings, L., Diederichs, B., Dmytrash, A., Backer, J., et al. (2008). Secreted bioactive factors from Bifidobacterium infantis enhance epithelial cell barrier function. Am. J. Physiol. Gastrointest. Liver Physiol. 295, G1025–G1034. doi: 10.1152/ajpgi.90227.2008
Favaro, L., Basaglia, M., Casella, S., Hue, I., Dousset, X., Gombossy de Melo Franco, B. D., et al. (2014). Bacteriocinogenic potential and safety evaluation of non-starter Enterococcus faecium strains isolated from home made white brine cheese. Food Microbiol. 38, 228–239. doi: 10.1016/j.fm.2013.09.008
Fisher, K., and Phillips, P. (2009). The ecology, epidemiology and virulence of Enterococcus. Microbiology 155, 1749–1757. doi: 10.1099/mic.0.026385-0
Foulquié Moreno, M. R., Sarantinopoulos, P., Tsakalidou, E., and De Vuyst, L. (2006). The role and application of enterococci in food and health. Int. J. Food Microbiol. 106, 1–24. doi: 10.1016/j.ijfoodmicro.2005.06.026
Franz, C. M., Huch, M., Abriouel, H., Holzapfel, W., and Gálvez, A. (2011). Enterococci as probiotics and their implications in food safety. Int. J. Food Microbiol. 151, 125–140. doi: 10.1016/j.ijfoodmicro.2011.08.014
Franz, C. M., Muscholl-Silberhorn, A. B., Yousif, N. M., Vancanneyt, M., Swings, J., and Holzapfel, W. H. (2001). Incidence of virulence factors and antibiotic resistance among Enterococci isolated from food. Appl. Environ. Microbiol. 67, 4385–4389. doi: 10.1128/AEM.67.9.4385-4389.2001
Franz, C. M., van Belkum, M. J., Holzapfel, W. H., Abriouel, H., and Gálvez, A. (2007). Diversity of enterococcal bacteriocins and their grouping in a new classification scheme. FEMS Microbiol. Rev. 31, 293–310. doi: 10.1111/j.1574-6976.2007.00064.x
Gálvez, A., Maqueda, M., Valdivia, E., Quesada, A., and Montoya, E. (1986). Characterization and partial purification of a broad spectrum antibiotic AS-48 produced by Streptococcus faecalis. Can. J. Microbiol. 32, 765–771. doi: 10.1139/m86-141
Giraffa, G. (2003). Functionality of enterococci in dairy products. Int. J. Food Microbiol. 88, 215–222. doi: 10.1016/S0168-1605(03)00183-1
Gupta, A., and Tiwari, S. K. (2015). Probiotic potential of bacteriocin-producing Enterococcus hirae strain LD3 isolated from dosa batter. Ann. Microbiol. 65, 2333–2342. doi: 10.1007/s13213-015-1075-4
Hadji-Sfaxi, I., El-Ghaish, S., Ahmadova, A., Batdorj, B., Le Blay, G., Barbier, G., et al. (2011). Antimicrobial activity and safety of use of Enterococcus faecium PC4.1 isolated from Mongol yogurt. Food Control 22, 2020–2027. doi: 10.1016/j.foodcont.2011.05.023
Hammad, A. M., Hassan, H. A., and Shimamoto, T. (2015). Prevalence, antibiotic resistance and virulence of Enterococcus spp. in Egyptian fresh raw milk cheese. Food Control 50, 815–820. doi: 10.1016/j.foodcont.2014.10.020
Hegarty, J. W., Guinane, C. M., Ross, R. P., Hill, C., and Cotter, P. D. (2016). Bacteriocin production: a relatively unharnessed probiotic trait? F1000Res. 5:2587. doi: 10.12688/f1000research.9615.1
Hillion, M., Mijouin, L., Jaouen, T., Barreau, M., Meunier, P., Lefeuvre, L., et al. (2013). Comparative study of normal and sensitive skin aerobic bacterial populations. Microbiologyopen 2, 953–961. doi: 10.1002/mbo3.138
Inoǧlu, Z. N., and Tuncer, Y. (2013). Safety assessment of Enterococcus faecium and Enterococcus faecalis strains isolated from Turkish Tulum Cheese. J. Food Saf. 33, 369–377. doi: 10.1111/jfs.12061
Jacobsen, C. N., Nielsen, V., Hayford, A. E., Møller, P. L., Michaelsen, K. F., et al. (1999). Screening of probiotic activities of forty-seven strains of Lactobacillus spp. by in vitro techniques and evaluation of the colonization ability of five selected strains in humans. Appl. Environ. Microbiol. 65, 4949–4956.
Juntunen, M., Kirjavainen, P. V., Ouwehand, A. C., Salminen, S. J., and Isolauri, E. (2001). Adherence of probiotic bacteria to human intestinal mucus in healthy infants and during rotavirus infection. Clin. Diagn. Lab. Immunol. 2, 293–296. doi: 10.1128/CDLI.8.2.293-296.2001
Khalkhali, S., and Mojgani, N. (2017). Bacteriocinogenic potential and virulence traits of Enterococcus faecium and E. faecalis isolated from human milk. Iran. J. Microbiol. 9, 224–233.
Klingberg, T. D., Pedersen, M. H., Cencic, A., and Budde, B. B. (2005). Application of measurements of transepithelial electrical resistance of intestinal epithelial cell monolayers to evaluate probiotic activity. Appl. Environ. Microbiol. 71, 7528–7530. doi: 10.1128/AEM.71.11.7528-7530.2005
Klingspor, S., Bondzio, A., Martens, H., Aschenbach, J. R., Bratz, K., Tedin, K., et al. (2015). Enterococcus faecium NCIMB 10415 modulates epithelial integrity, heat shock protein, and proinflammatory cytokine response in intestinal cells. Mediators Inflamm. 2015:304149. doi: 10.1155/2015/304149
Ladero, V., Fernández, M., Calles-Enríquez, M., Sánchez-Llana, E., Cañedo, E., Martín, M. C., et al. (2012). Is the production of the biogenic amines tyramine and putrescine a species-level trait in enterococci? Food Microbiol. 30, 132–138. doi: 10.1016/j.fm.2011.12.016
Mannu, L., Paba, A., Daga, E., Comunian, R., Zanetti, S., Duprè, I., et al. (2003). Comparison of the incidence of virulence determinants and antibiotic resistance between Enterococcus faecium strains of dairy, animal and clinical origin. Int. J. Food Microbiol. 88, 291–304. doi: 10.1016/S0168-1605(03)00191-0
Maragkoudakis, P. A., Zoumpopoulou, G., Miaris, C., Kalantzopoulos, G., Pot, B., and Tsakalidou, E. (2006). Probiotic potential of Lactobacillus strains isolated from dairy products. Int. Dairy J. 16, 189–199. doi: 10.1016/j.idairyj.2005.02.009
Marteau, P., Minekus, M., Havenaar, R., and Huis in’t Veld, J. H. J. (1997). Survival of lactic acid bacteria in a dynamic model of the stomach and small intestine: validation and the effects of bile. J. Dairy Sci. 80, 1031–1037. doi: 10.3168/jds.S0022-0302(97)76027-2
Messaoudi, S., Madi, A., Prévost, H., Feuilloley, M., Manai, M., Dousset, X., et al. (2012). In vitro evaluation of the probiotic potential of Lactobacillus salivarius SMXD51. Anaerobe 18, 584–589. doi: 10.1016/j.anaerobe.2012.10.004
Murray, B. E. (1990). The life and times of the Enterococcus. Clin. Microbiol. Rev. 3, 46–65. doi: 10.1128/CMR.3.1.46
Ogaki, M. B., Rocha, K. R., Terra, M. R., Furlaneto, M. C., and Maia, L. F. (2016). Screening of the enterocin-encoding genes and antimicrobial activity in Enterococcus species. J. Microbiol. Biotechnol. 26:10261034. doi: 10.4014/jmb.1509.09020
Ogier, J. C., and Serror, P. (2008). Safety assessment of dairy microorganisms: the Enterococcus genus. Int. J. Food Microbiol. 126, 291–301. doi: 10.1016/j.ijfoodmicro.2007.08.017
Park, J., Jin, G. D., Pak, J. I., Won, J., and Kim, E. B. (2017). Development of a rapid identification method for the differentiation of Enterococcus species using a species-specific multiplex PCR based on comparative genomics. Curr. Microbiol. 74, 476–483. doi: 10.1007/s00284-017-1210-5
Popović, N., Dinić, M., Tolinaèki, M., Mihajlović, S., Terzić-Vidojević, A., Bojić, S., et al. (2018). New insight into biofilm formation ability, the presence of virulence genes and probiotic potential of Enterococcus sp. dairy isolates. Front. Microbiol. 9:78. doi: 10.3389/fmicb.2018.00078
Prince, T., McBain, A. J., and O’Neill, C. A. (2012). Lactobacillus reuteri protects epidermal keratinocytes from Staphylococcus aureus-induced cell death by competitive exclusion. Appl. Environ. Microbiol. 78, 5119–5126. doi: 10.1128/AEM.00595-12
Resta-Lenert, S., and Barrett, K. E. (2003). Live probiotics protect intestinal epithelial cells from the effects of infection with enteroinvasive Escherichia coli (EIEC)”. Gut 52, 988–997. doi: 10.1136/gut.52.7.988
Reviriego, C., Eaton, T., Martin, R., Jimenez, E., Fernandez, L., Gasson, M. J., et al. (2005). Screening of virulence determinants in Enterococcus faecium strains isolated from breast milk. J. Hum. Lact. 21, 131–138. doi: 10.1177/0890334405275394
Rosenthal, V. D., Maki, D. G., Mehta, A., Alvarez-Moreno, C., Leblebicioglu, H., Higuera, F., et al. (2008). International nosocomial infection control consortium members 2002–2007. Am. J. Infect. Control 36, 627–637. doi: 10.1016/j.ajic.2008.03.003
Ruiz, P., Pérez-Martín, F., Seseña, S., and Llanos Palop, M. (2016). Seasonal diversity and safety evaluation of enterococci population from goat milk in a farm. Dairy Sci. Technol. 96, 359–375. doi: 10.1007/s13594-015-0273-y
Saarela, M., Mogensen, G., Fondén, R., Mättö, J., and Mattila-Sandholm, T. (2000). Probiotic bacteria: safety, functional and technological properties. J. Biotechnol. 84, 197–215. doi: 10.1016/S0168-1656(00)00375-8
Satish Kumar, R., Kanmani, P., Yuvaraj, N., Paari, K. A., Pattukumar, V., and Arul, V. (2011). Purification and characterization of enterocin MC13 produced by a potential aquaculture probiont Enterococcus faecium MC13 isolated from the gut of Mugil cephalus. Can. J. Microbiol. 57, 993–1001. doi: 10.1139/w11-092
Schaberg, D. R., Culver, D. H., and Gaynes, R. P. (1991). Major trends in the microbial etiology of nosocomial infection. Am. J. Med. 91, 72S–75S. doi: 10.1016/0002-9343(91)90346-Y
Schneeberger, E. E., and Lynch, R. D. (2004). The tight junction: a multifunctional complex. Am. J. Physiol. Cell Physiol. 286, C1213–C1228. doi: 10.1152/ajpcell.00558.2003
Sherman, P. M., Johnson-Henry, K. C., Yeung, H. P., Ngo, P. S., Goulet, J., and Tompkins, T. A. (2005). Probiotics reduce enterohemorrhagic Escherichia coli O157:H7- and enteropathogenic E. coli O127:H6-induced changes in polarized T84 epithelial cell monolayers by reducing bacterial adhesion and cytoskeletal rearrangements. Infect. Immun. 73, 5183–5188. doi: 10.1128/IAI.73.8.5183-5188.2005
Sogawa, K., Watanabe, M., Sato, K., Segawa, S., Ishii, C., Miyabe, A., et al. (2011). Use of the MALDI BioTyper system with MALDI-TOF mass spectrometry for rapid identification of microorganisms. Anal. Bioanal. Chem. 400, 1905–1911. doi: 10.1007/s00216-011-4877-7
Strompfová, V., Lauková, A., Simonová, M., and Marcináková, M. (2008). Occurrence of the structural enterocin A, P, B, L50B genes in enterococci of different origin. Vet. Microbiol. 132, 293–301. doi: 10.1016/j.vetmic.2008.05.001
Todorov, S. D., Botes, M., Guigas, C., Schillinger, U., Wiid, I., Wachsman, M. B., et al. (2008). Boza, a natural source of probiotic lactic acid bacteria. J. Appl. Microbiol. 104, 465–477. doi: 10.1111/j.1365-2672.2007.03558.x
Todorov, S. D., Wachsman, M., Tomé, E., Dousset, X., Destro, M. T., Milner, L., et al. (2010). Characterisation of an antiviral pediocin-like bacteriocin produced by Enterococcus faecium. Food Microbiol. 27, 869–879. doi: 10.1016/j.fm.2010.05.001
Todorov, S. D., Wachsman, M. B., Knoetze, H., Meincken, M., and Dicks, L. M. T. (2005). An antibacterial and antiviral peptide produced by Enterococcus mundtii ST4 V isolated from soya beans. Int. J. Antimicrob. Agents 25, 508–513. doi: 10.1016/j.ijantimicag.2005.02.005
Tuomola, E., Crittenden, R., Playne, M., Isolauri, E., and Salminen, S. (2001). Quality assurance criteria for probiotic bacteria. Am. J. Clin. Nutr. 73, 393s–398s. doi: 10.1093/ajcn/73.2.393s
Turpin, W., Humbolt, C., Noordine, M. L., Thomas, M., and Guyot, J. P. (2012). Lactobacillaceae and cell adhesion: genomic and functional screening. PLoS One 7:e38034. doi: 10.1371/journal.pone.0038034
Veljović, K., Popović, N., Miljković, M., Tolinaèki, M., Terzić-Vidojević, A., and Kojić, M. (2017). Novel aggregation promoting factor AggE contributes to the probiotic properties of Enterococcus faecium BGGO9-28. Front. Microbiol. 8:1843. doi: 10.3389/fmicb.2017.01843
Wieckowicz, M., Schmidt, M., Sip, A., and Graje, W. (2011). Development of a PCR-based assay for rapid detection of class IIa bacteriocin genes. Lett. Appl. Microbiol. 52, 281–289. doi: 10.1111/j.1472-765X.2010.02999.x
Yamaguchi, T., Miura, Y., and Matsumoto, T. (2013). Antimicrobial susceptibility of Enterococcus strains used in clinical practice as probiotics. J. Infect. Chemother. 19, 1109–1115. doi: 10.1007/s10156-013-0633-6
Yanagida, F., Chen, Y., Onda, T., and Shinohara, T. (2005). Durancin L28-1A, a new bacteriocin from Enterococcus durans L28- 1, isolated from soil. Lett. Appl. Microbiol. 40, 430–435. doi: 10.1111/j.1472-765X.2005.01693.x
Keywords: Enterococcus faecium, probiotics, Dried Ossban, safety, antibiotic resistance, virulence determinants
Citation: Zommiti M, Cambronel M, Maillot O, Barreau M, Sebei K, Feuilloley M, Ferchichi M and Connil N (2018) Evaluation of Probiotic Properties and Safety of Enterococcus faecium Isolated From Artisanal Tunisian Meat “Dried Ossban”. Front. Microbiol. 9:1685. doi: 10.3389/fmicb.2018.01685
Received: 06 May 2018; Accepted: 06 July 2018;
Published: 06 August 2018.
Edited by:
Fatih Ozogul, Çukurova University, TurkeyReviewed by:
Giuseppe Spano, University of Foggia, ItalyCopyright © 2018 Zommiti, Cambronel, Maillot, Barreau, Sebei, Feuilloley, Ferchichi and Connil. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nathalie Connil, bmF0aGFsaWUuY29ubmlsQHVuaXYtcm91ZW4uZnI=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.