 Inwoo Baek
Inwoo Baek Mincheol Kim
Mincheol Kim Imchang Lee
Imchang Lee Seong-In Na
Seong-In Na Michael Goodfellow5
Michael Goodfellow5 Jongsik Chun
Jongsik Chun
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 30 April 2018
Sec. Evolutionary and Genomic Microbiology
Volume 9 - 2018 | https://doi.org/10.3389/fmicb.2018.00834
The genus Turicella was proposed to harbor clinical strains isolated from middle-ear fluids of patients with otitis media. 16S rRNA phylogeny showed that it belonged to the mycolic acid-containing actinobacteria, currently classified in the order Corynebacteriales, and was closely related to the genus Corynebacterium. A new genus was proposed for the organisms as unlike corynebacteria they lacked mycolic acids and had different menaquinones. Here, we carried out large-scale comparative genomics on representative strains of the genera Corynebacterium and Turicella to check if this chemotaxonomic classification is justified. Three genes that are known to play an essential role in mycolic acid biosynthesis were absent in Turicella and two other mycolate-less Corynebacterium spp., explaining the lack of mycolic acids resulted from the deletion of genes and does not confer any phylogenetic context. Polyphasic phylogenetic analyses using 16S rRNA, bacterial core genes and genes responsible for synthesizing menaquinones unequivocally indicate that Turicella is a true member of the genus Corynebacterium. Here, we demonstrate that menaquinone and mycolic acid that have been used as critical taxonomic markers should be interpreted carefully, particularly when genome-based taxonomy is readily available. Based on the phylogenetic analysis, we propose to reclassify Turicella otitidis as Corynebacterium otitidis comb. nov.
The order Corynebacteriales encompasses actinobacterial strains that are important in clinical, environmental and industrial microbiology (Lehmann and Neumann, 1896; Goodfellow and Jones, 2015). The taxon is well defined by the presence of mycolic acids, a special type of long chain fatty acid only found in members of this order. Mycolic acids are known to act as a cell wall permeability barrier that confers resistance to antibiotics and phagocytosis (Gebhardt et al., 2007). The length and number of double bonds in mycolic acids are used as important chemotaxonomic markers for distinguishing between genera classified in the Corynebacteriales (Bernard et al., 2010; Marrakchi et al., 2014) though some species in the order lack mycolic acids (Funke et al., 1994; Collins et al., 1998; Wiertz et al., 2013). Historically, the classification of mycolic acid-containing taxa was guided by a combination of chemotaxonomic and 16S rRNA gene sequence data; several new genera, including Hoyosella (Jurado et al., 2009) and Lawsonella (Bell et al., 2016) were recently described accordingly. However, in some cases, a lack of congruence between these sets of data required the use of addition taxonomic evidence to clarify the situation.
The classification of the genus Turicella is a case in point. The type and only species of this genus, Turicella otitidis, was initially proposed to harbor bacterial strains isolated from the ear of a patient with otitis media (Funke et al., 1994). In this study, the type strain of T. otitidis was recovered as a sister taxon to members of the genus Corynebacterium in the 16S rRNA gene tree while showing chemotaxonomic properties that differed from those of most Corynebacterium strains, notably the absence of mycolic acids and the presence of fully unsaturated menaquinones (MK-10 and MK-11), as opposed to dihydrogenated menaquinones [MK-8(H2) and MK-9(H2)]. However, later studies using more 16S rRNA gene sequences showed that Turicella formed a phyletic lineage within the Corynebacterium clade (Goyache et al., 2003; Hall et al., 2003).
Major fatty acid biosynthetic pathways, namely the FAS-I (Fatty acid synthesis-I) and FAS-II (Fatty acid synthesis-II) cycles, have been well documented for actinobacteria (Marrakchi et al., 2014). One multifunctional gene, 3-oxoacyl-ACP synthase (fas), participates in all reactions in the FAS-I cycle (Bloch and Vance, 1977). In contrast, four essential genes are involved in the FAS-II pathway, namely beta-ketoacyl-ACP synthase (kasA) (Bhatt et al., 2005), beta-ketoacyl-ACP reductase (mabA) (Parish et al., 2007), (3R)-hydroxy acyl-ACP dehydratase subunit B (hadB) (Brown et al., 2007; Sacco et al., 2007) and (NADH)-dependent trans-2-enoyl-ACP reductase (inhA) (Vilcheze et al., 2000). Mycobacterium spp. are known to have both the FAS-I and FAS-II pathways whereas most Corynebacterium spp. contain only the FAS-I pathway. It is reported that Corynebacterium jeikeium and Corynebacterium urealyticum have neither the FAS-I nor FAS-II cycle and obtain fatty acids from the exogenous environment (Tauch et al., 2005, 2008).
Mycolic acids are synthesized by a coupling of carboxylated fatty acids (α - branch) and meromycolic acids. Two kinds of carboxylases are involved; acyl-CoA carboxylase and acetyl-CoA carboxylase (Gande et al., 2007). The meromycolic acid is formed by the modification and activation of long-chain fatty acids. In Mycobacterium tuberculosis, several genes are known to participate in the modification of fatty acids, including desaturation (NADPH-dependent stearoyl-CoA 9-desaturase; desA3) (Cole et al., 1998), then, long-chain fatty acid AMP ligase (fadD32) leads to the synthesis of the meromycolic acid (Portevin et al., 2005). Finally, mycolic acids are formed by condensing two fatty acids by polyketide synthase 13 (pks13) (Portevin et al., 2004). It is known that three of these genes (fadD32-pks13-accD4) are located in a single operon which is essential for mycolic acid synthesis (Portevin et al., 2005); two beta common subunits of carboxylases (accD4 and accD5) are also essential for mycolic acid synthesis (Gande et al., 2004).
Menaquinones are one of the major isoprenoid quinones required in bacterial electron transport systems (Nowicka and Kruk, 2010). The length and degree of saturation in isoprenoid chains of menaquinones are considered to be key taxonomic markers in the classification of Actinobacteria (Collins and Jones, 1981). Two major types of menaquinone biosynthetic pathways have been reported: the isochorismate and futalosine pathways (Seto et al., 2008). Mycobacterium tuberculosis, the most studied organism in the order Corynebacteriales, contains the former pathway; the component genes of this pathway were found in the genome of this organism (Dhiman et al., 2009). Upadhyay et al. (2015) found an enzyme, encoded by the menaquinone reductase (menJ) gene, confers the saturation of menaquinones of M. tuberculosis; the deletion of the menJ gene leads to the production of MK-9 instead of MK-9(H2).
In this study, we re-examine the confused taxonomic status of the genus Turicella using whole genome-based phylogeny and comparative genomics of genes responsible for synthesizing chemotaxonomic markers, namely fatty acids, mycolic acids, and menaquinones. On the basis of the evidence derived from the phylogenomic and comparative genomic analyses, it is proposed that Turicella otitidis be classified in the genus Corynebacterium as Corynebacterium otitidis comb. nov.
A set of 93 genome sequences includes type strains of 77 Corynebacterium spp., one T. otitidis (Brinkrolf et al., 2012), and 14 representative genera classified in the order Corynebacteriales (Dietzia alimentaria, Gordonia bronchialis, Hoyosella altamirensis, Lawsonella clevelandensis, Millisia brevis, Mycobacterium tuberculosis, Nocardia asteroides, Rhodococcus rhodochrous, Segniliparus rotundus, Skermania piniformis, Smaragdicoccus niigatensis, Tomitella biformata, Tsukamurella paurometabola, and Williamsia muralis). The type strain of Pseudonocardia thermophila was included as an outgroup. The whole-genome assemblies and their predicted CDS sequences were downloaded from the EzBioCloud database1 (Yoon et al., 2017). Genes related to the chemotaxonomic markers of the species were identified by a BLASTX search using BLAST+ v. 2.2.29 program (E-value cutoff = 1e-5, bit score cutoff = 100, identity cutoff = 30%; Rost, 1999) when compared against the reference gene sequences in the UniProt database2 (Apweiler et al., 2004). Genes of M. tuberculosis and Corynebacterium glutamicum were used as references for detecting mycolic acid biosynthetic genes (Ikeda and Nakagawa, 2003; Gande et al., 2007; Takeno et al., 2013; Marrakchi et al., 2014). Menaquinone biosynthesis genes in SwissProt, mainly M. tuberculosis and Streptomyces coelicolor (Seto et al., 2008), were used as references for the menaquinone biosynthesis pathway (Boutet et al., 2016). Description, abbreviation, and UniProt ID of all reference proteins are listed in Supplementary Table 1.
The bacterial core gene-based phylogenetic analysis was carried out using the UBCG pipeline3 (Na et al., 2018). From the concatenated gene sequences extracted by UBCG, a maximum-likelihood phylogenetic tree was inferred using RAxML version 8.2.8 (Stamatakis, 2014) with the GTRGAMMA model and 100 bootstrap replications (Felsenstein, 1985). Another core-gene based genome-wide phylogenetic analysis was performed using PhyloPhlAn (Segata et al., 2013) based on CDS sequences of each genome downloaded in the previous step. The FastTree2 software implemented in this software was used to infer an approximated maximum-likelihood tree (Price et al., 2010; Liu et al., 2011). The 16S rRNA gene sequences of the same species were downloaded from the EzBioCloud database and aligned manually using secondary structure information by the EzEditor2 program (Jeon et al., 2014) and a maximum-likelihood phylogenetic tree reconstructed using RAxML with the GTRGAMMA model and 1,000 bootstrap replications (Felsenstein, 1985).
To infer the evolutionary history of genes involved in the biosynthesis of chemotaxonomic markers, a concatenated sequence alignment was generated from seven genes responsible for menaquinone biosynthesis (menA ∼menG), then used to elucidate the phylogenetic relationship using RAxML. Prior to concatenation, each gene was aligned separately using MAFFT software (v. 7.310) (Katoh and Standley, 2014). The Evolview web server4 was used to visualize the phylogenomic trees with the information on the presence of genes (He et al., 2016).
The type strain of T. otitidis and those representing the Corynebacterium spp. formed a monophyletic clade in the phylogenomic trees inferred using both the UBCG (Figure 1) and PhyloPhlAn (Supplementary Figure 1) phylogenomic pipelines. The topology and phylogenetic relationships shown in the trees are in line with those from previous studies (Wu and Eisen, 2008; Wu et al., 2013), as well as from our analysis based on 16S rRNA phylogeny (Figure 2). It is clear that, unlike in the original study (Funke et al., 1994), T. otitidis does not form a sister taxon to the Corynebacterium clade but a phyletic lineage within the clade, indicating that it is a bona fide member of this genus.

FIGURE 1. Genome-based phylogenetic tree reconstructed by the UBCG phylogenomics pipeline (Na et al., 2018) for Turicella otitidis and neighboring species. Bar in the below signifies substitution rate per site. Blocks in the column plots on the right side indicate the presence of each gene (or gene sets). Colored blocks indicate that the gene is present in the genome, and white blocks (with Δ sign) signify its absence. Numbers at the nodes indicate the bootstrap value of RAxML. Features and their corresponding genes: MA, mycolic acid biosynthetic genes; FAS1, fatty acid synthesis cycle 1 (fas); FAS2, fatty acid synthesis cycle 2 (mabA, inhA, kasA, and hadB); DesA3, NADPH-dependent stearoyl-CoA 9-desaturase; MenJ, menaquinone reductase.

FIGURE 2. Phylogenetic tree reconstructed with 16S rRNA genes for T. otitidis and neighboring species. Bar indicates substitution rate per site. Numbers at the nodes are bootstrap values for 1,000 replicated subsamples expressed as percentage values.
The genomes of the type strain of T. otitidis and those of 61 out of the 77 Corynebacterium spp. contain the fas gene which expresses for the FAS-I pathway. Essential genes in the FAS-II pathway (mabA, inhA, kasA, and hadB) are absent from the genomes of T. otitidis and the Corynebacterium strains. The genomes of most Corynebacteriales species contain the four essential genes for the FAS-II cycle, the exceptions are Dietzia alimentaria and Lawsonella clevelandensis (Supplementary Tables 2, 3). Dietzia alimentaria lacks the kasA and hadB genes, and L. clevelandensis all of the FAS-I and FAS-II related genes. Comparative genomics and analysis of the synteny of genes responsible for mycolic acid biosynthesis suggest that T. otitidis and three mycolate-less Corynebacterium species (Corynebacterium amycolatum, Corynebacterium kroppenstedtii, and Corynebacterium lactis) lost the fadD32-pks13-accD4 operon, located between the genes coding for protein PS1 and UPF0104 membrane protein in other species (Figure 3). They also lack cmrA, whereas all other Corynebacterium species have the necessary genes. In contrast, other carboxylase subunits that are known to be essential for mycolic acid biosynthesis (accD2 and accD3) were found in the genomes of species that lack mycolic acids. Among the analyzed genomes, genes that are similar to those associated with mycolic acid modification (desaturation, cyclization, and methyl transfer) in M. tuberculosis were detected, though the sequence identity and the number of genes were not sufficient for further analysis. Exceptionally, genes homologous to desA3, a gene associated with the desaturation mycolic acids, were found in 23 of the Corynebacterium species; their sequence identity was around 50% compared to the reference (Supplementary Table 2). In contrast, two other kinds of putative desaturases (desA1 and desA2) involved in the desaturation of mycolic acids were not detected in the Corynebacterium genomes.
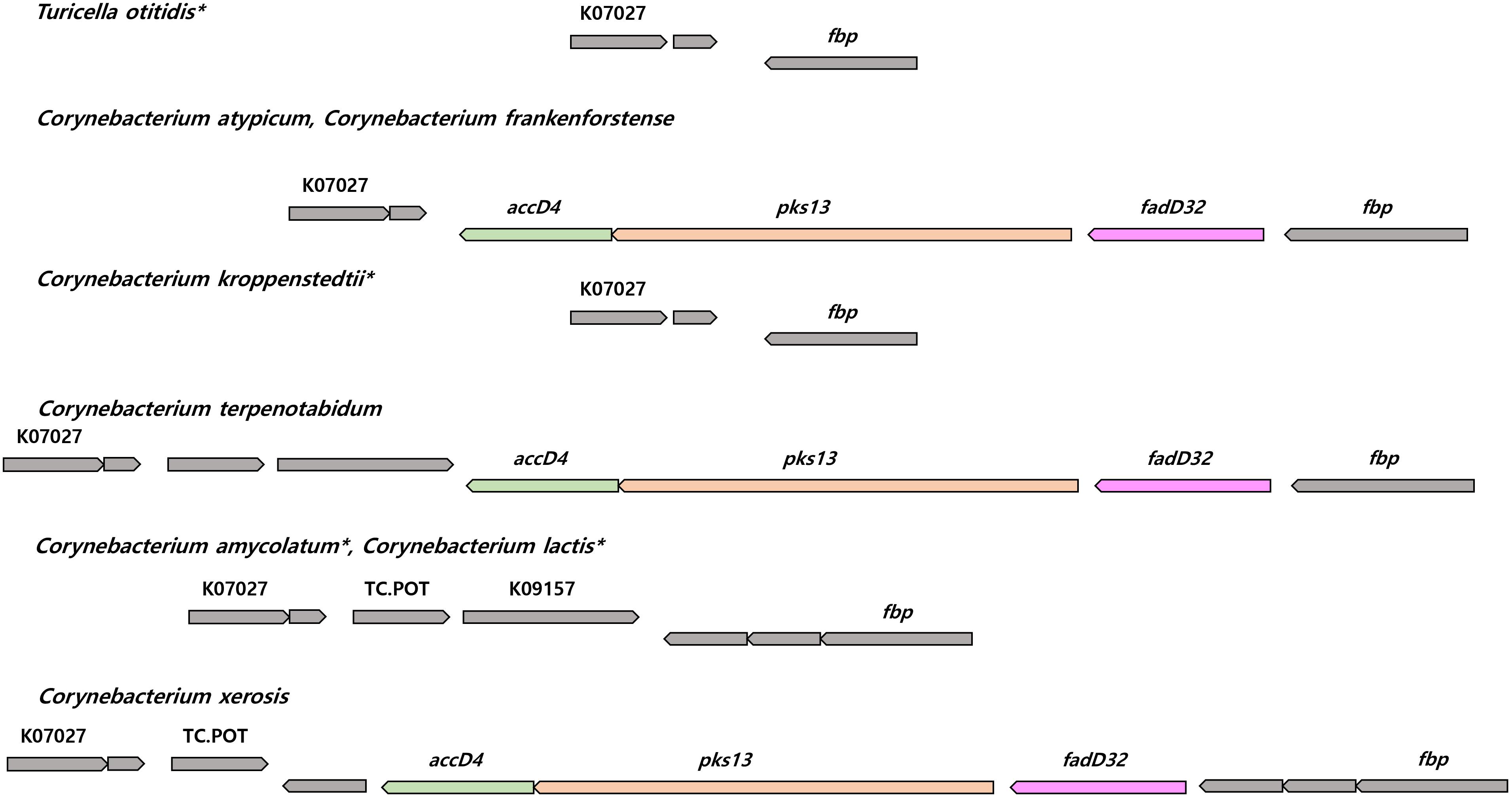
FIGURE 3. The synteny plot of gene families near fadD32-pks13-accD4 operon site of species whose mycolic acid is absent and their phylogenetically neighboring species. Mycolic acid lacked species are marked with an asterisk. accD4, acyl-CoA carboxylase subunit beta 1; pks13, polyketide synthase; fadD32, long-chain-fatty-acid-AMP ligase; K07027, UPF0104 membrane protein; fbp, protein PS1; TC.POT, di/tripeptide transporter; K09157, UPF0210 protein.
The presence of genes responsible for menaquinone biosynthesis, including those participating in the elongation of isoprenoid side chains, was checked. Seven menaquinone biosynthetic genes (menA ∼menG), and a menaquinone side chain elongation gene (uppS and hepST) were common amongst the genomes of Corynebacteriales species including T. otitidis. On the other hand, a menJ gene implicated in menaquinone side chain saturation was not found in either T. otitidis or C. kroppenstedtii, though it was present in all of the other Corynebacterium genomes. This gene was also detected in the other Corynebacteriales species analyzed in this study, apart from those of Hoyosella altamirensis, Lawsonella clevelandensis, and Tsukamurella paurometabola which are reported to have fully unsaturated menaquinones (Table 1). The menaquinone composition of C. kroppenstedtii was not reported in the initial description (Collins et al., 1998). Comparative analysis of the gene synteny showed that the lack of menJ gene located between hepST and menG genes is apparent in both T. otitidis and C. kroppenstedtii, suggesting C. kroppenstedtii likely has fully unsaturated menaquinones (Figure 4).
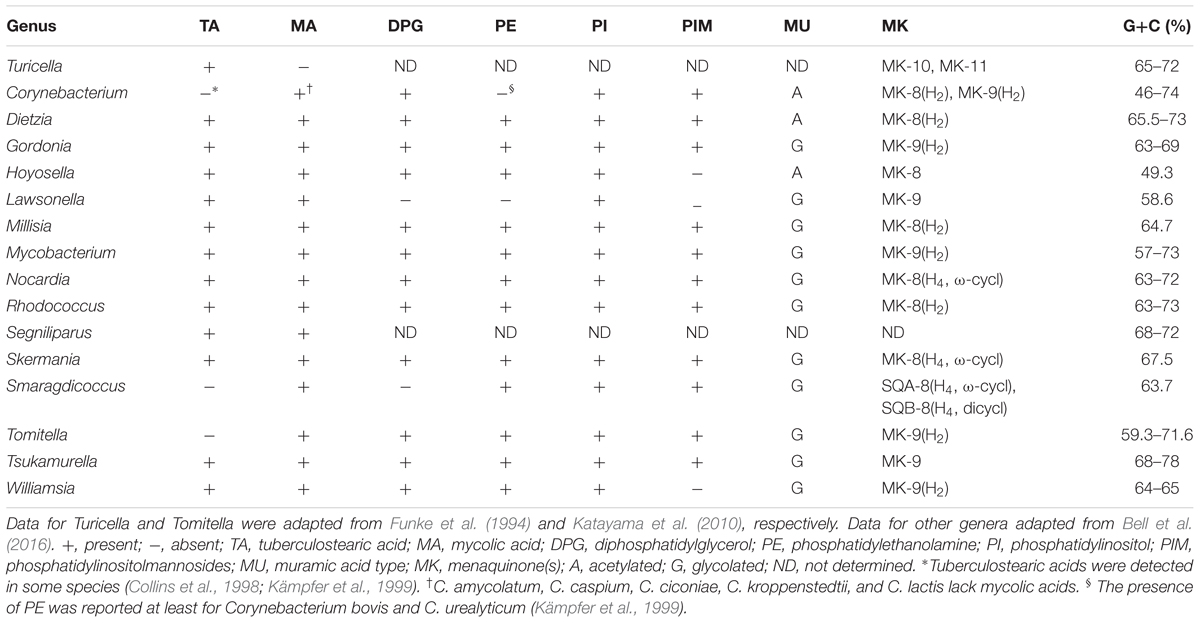
TABLE 1. Differential chemotaxonomic characteristics of genera in the order Corynebacteriales.
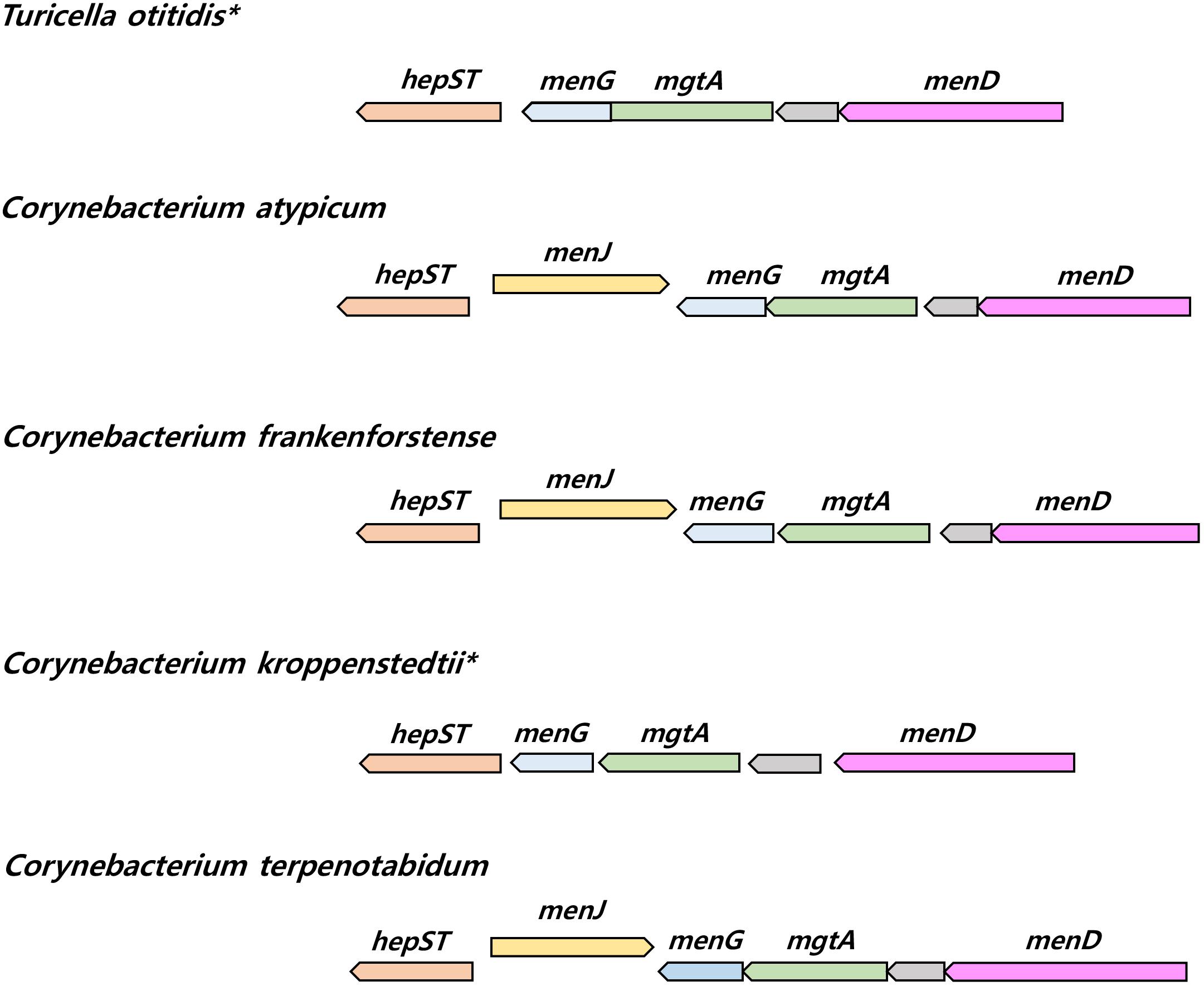
FIGURE 4. The synteny plot of gene families near menJ gene site of T. otitidis and related species. Species who lacked menJ gene are marked with an asterisk. hepST, geranyl pyrophosphate synthase; menG, demethylmenaquinone methyltransferase; mgtA, GDP-mannose-dependent alpha-mannosyltransferase; menD, 2-succinyl-5-enolpyruvyl-6-hydroxy-3-cyclohexene-1-carboxylate synthase.
Gene-concatenated phylogenetic trees based on the seven genes (menA ∼menG) responsible for menaquinone biosynthesis were generated (Figure 5). As with the phylogenetic trees based on bacterial core and 16S rRNA genes, both nucleotide and amino acid gene concatenated trees support the inclusion of T. otitidis with the Corynebacterium species, even though the detail topologies within the evolutionary radiation occupied by the Corynebacterium-Turicella clade differ between trees.

FIGURE 5. (A) Phylogenetic tree reconstructed with amino acid sequences of seven menaquinone biosynthetic genes for T. otitidis and its close species. (B) Phylogenetic tree reconstructed with nucleotide sequences of seven menaquinone biosynthetic gene families for T. otitidis and neighboring species. Bar indicates substitution rate per site. Numbers at the nodes are bootstrap values for 1,000 replicated subsamples expressed as a percentage value.
The application of chemotaxonomic procedures led to marked improvements in prokaryotic systematics (Goodfellow et al., 2012) though little attention was paid to the confidence that could be placed on chemical characters. Experimental protocols used to acquire chemical data can be influenced by cultivation conditions, fatty acid profiles, for instance, are known to be sensitive to changes in temperature (Russell, 1984). Again, chemical markers generally are seen to be reliable, such as the presence or absence of mycolic acids in members of the order Corynebacteriales (Goodfellow and Jones, 2015), have been shown to be prone to experimental error. Thus, early reports that Corynebacterium atypicum, Hoyosella altarmirensis, and Hoyosella subflava lacked mycolic acids (Hall et al., 2003; Jurado et al., 2009; Wang et al., 2010) were found to be misplaced in light of later studies (Laneelle et al., 2012; Tippelt et al., 2014; Hamada et al., 2016; Li et al., 2016). Similarly, the report that the type strain of Corynebacterium amycolatum contained major amounts of dihydrogenated menaquinones with eight and nine isoprenoid units (Collins et al., 1988) was overturned by the discovery that this organism was rich in fully unsaturated menaquinones with nine isoprene units (Kämpfer et al., 1999; Jurado et al., 2009). In a broader context, it has been pointed out that the interpretation of polar lipid patterns based on two-dimensional thin-layer-chromatography is inherently subjective (Sutcliffe et al., 2012; Sutcliffe et al., 2013) while evidence that menaquinone composition can be significantly influenced when biomass is harvested at different stages of the growth cycle has generally been overlooked (Saddler et al., 1986). Such problems are compounded by the loss of single or small numbers of genes coding for chemotaxonomic features, as like the case of fatty acid biosynthesis gene loss in some Corynebacterium species (Figure 1), and by the fact that differences in a set of chemotaxonomic traits may be affected by one or a few genes and hence do not reflect phylogeny. For example, only menJ gene present species have saturated site in isoprenyl chain in their menaquinone (Figure 1 and Table 1). Furthermore, there are two kinds of menaquinone biosynthetic pathways whose component genes are completely different (Seto et al., 2008), and the evolutionary history of those pathways was revealed (Zhi et al., 2014). Experimental chemotaxonomy cannot detect phylogenetic variance like this case. It is noteworthy that at present, we were not able to predict the isoprenyl chain length of menaquinones.
It is apparent from the present study that phylogenetic approaches can provide reliable ways of establishing whether Corynebacteriales strains are able to synthesis mycolic acids. It is particularly interesting that the gene set indispensable for the biosynthesis of mycolic acids (fadD32-pks13-accD4 operon and cmrA) is present in the genome of the C. atypicum and Hoyosella strains but have been lost from the genome of the T. otitidis strain. The genomes of the type strains of Corynebacterium caspium and Corynebacterium ciconiae also contain this gene set suggesting that they may synthesize mycolic acids (Figure 1 and Supplementary Tables 2, 3) even though these strains have been reported to lack these components (Collins et al., 2004; Fernandez-Garayzabal et al., 2004). It is also interesting that the genes essential for the FAS-II fatty acid pathway (hadB, inhA, kasA, and mabA) are absent from the genome of T. otitidis and Corynebacterium strains but present in the genome of Mycobacterium tuberculosis, Nocardia asteroides, and Rhodococcus erythropolis. These observations suggest that genes involved in the same pathway evolve relative to one another. Furthermore, as in these cases, consecutive gene transfer or loss in a pathway can be used for the evidence for the classification when considering the parsimony of such events.
At present, genome-based phylogenies tend to be based on single-copy ubiquitous orthologous genes (Wu and Eisen, 2008; Wu et al., 2013). This practice yields consistent results albeit ones that may be biased as many of the target genes encode ribosomal proteins. Nevertheless, genes associated with key metabolic pathways can be used to supplement genome-derived phylogenetic inference. In this study, a phylogenetic tree based on concatenated sequences of seven menaquinone biosynthetic genes was used as a basis for phylogenetic re-identification given the significance of menaquinone composition in the identification of bacteria. Again, the current genomic analyses provided information on microbial structural phenotypes (Figure 1), without recourse to wet laboratory experimental data (Table 1 and Supplementary Tables 2, 3). For example, the menaquinone profile of the type strain of C. kroppenstedtii is reported as MK-8 (Jurado et al., 2009), and the genome data can be used to infer that the major menaquinone of this organism, that is, fully unsaturated type given the absence of the menJ gene. It is noteworthy that the presence of genes or metabolic pathways derived from genomic information does not necessarily mean that it is always expressed to confer the phenotypes. However, this information can provide crucial clues about the phenotypes, including the chemotaxonomic markers.
At the time of the proposal of the genus Turicella (Funke et al., 1994), only three 16S rRNA gene sequences of the genus Corynebacterium were included in the phylogenetic analysis, leading to the conclusion of creating a new genus. The presence of fully unsaturated menaquinones in Turicella also supported this proposal. In this study, we showed that all aspects of phylogenetic evidence, including that based on menaquinone biosynthetic genes, indicate that Turicella is a genuine member of the genus Corynebacterium thereby supporting the view that phylogenomics provides more reliable data than chemotaxonomy in describing their evolutionary history and natural relationships, or that phylogenomics trumps chemotaxonomy. It can be inferred from the phenotypic inference of the genomes of T. otitidis that this organism has chemotaxonomic properties consistent with its classification in the genus Corynebacterium, notably the presence of genes associated with the FAS-II pathway. Consequently, it is proposed that T. otitidis be reclassified as Corynebacterium otitidis comb. nov.
The description is that given by Bernard et al. (2010) with the following changes. Most species produce menaquinones with partially saturated isoprenoid chains [mainly MK-8(H2) and/or MK-9(H2)], but for some species the fully unsaturated menaquinones MK-8, MK-9, MK-10, and MK-11 were found.
Corynebacterium otitidis (o.ti’ti.dis. L. gen. n. of otitis, inflammation of the ear). Basonym: Turicella otitidis Funke et al. (1994). This description is the same as that given by Funke et al. (1994). The type strain is 234/92T (=DSM 8821T = ATCC 51513T).
The idea for this study was conceived by JC and IB, who was also charged with supervising the whole project. Data collecting and reference searching, analyzing genomic data, and writing a draft of the manuscript scheme were done by IB. Phylogenomic trees were reconstructed by IB, IL, and S-IN, with applying the mechanisms and proposed the idea. The manuscript was written, curated, and confirmed by IB, MK, MG, and JC.
This work was supported by a National Science Foundation grant (NRF-2014M3C9A3063541/NRF-2015R1A2A2A01008404) through the National Research Foundation of Korea (NRF) funded by the Ministry of Science and ICT, South Korea.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.00834/full#supplementary-material
Apweiler, R., Bairoch, A., Wu, C. H., Barker, W. C., Boeckmann, B., Ferro, S., et al. (2004). UniProt: the Universal Protein knowledgebase. Nucleic Acids Res. 32, D115–D119. doi: 10.1093/nar/gkh131
Bell, M. E., Bernard, K. A., Harrington, S. M., Patel, N. B., Tucker, T. A., Metcalfe, M. G., et al. (2016). Lawsonella clevelandensis gen. nov., sp. nov., a new member of the suborder Corynebacterineae isolated from human abscesses. Int. J. Syst. Evol. Microbiol. 66, 2929–2935. doi: 10.1099/ijsem.0.001122
Bernard, K. A., Wiebe, D., Burdz, T., Reimer, A., Ng, B., Singh, C., et al. (2010). Assignment of Brevibacterium stationis (ZoBell and Upham 1944) Breed 1953 to the genus Corynebacterium, as Corynebacterium stationis comb. nov., and emended description of the genus Corynebacterium to include isolates that can alkalinize citrate. Int. J. Syst. Evol. Microbiol. 60(Pt 4), 874–879. doi: 10.1099/ijs.0.012641-0
Bhatt, A., Kremer, L., Dai, A. Z., Sacchettini, J. C., and Jacobs, W. R. Jr. (2005). Conditional depletion of KasA, a key enzyme of mycolic acid biosynthesis, leads to mycobacterial cell lysis. J. Bacteriol. 187, 7596–7606. doi: 10.1128/JB.187.22.7596-7606.2005
Bloch, K., and Vance, D. (1977). Control mechanisms in the synthesis of saturated fatty acids. Annu. Rev. Biochem. 46, 263–298. doi: 10.1146/annurev.bi.46.070177.001403
Boutet, E., Lieberherr, D., Tognolli, M., Schneider, M., Bansal, P., Bridge, A. J., et al. (2016). UniProtKB/Swiss-Prot, the manually annotated section of the UniProt KnowledgeBase: how to use the entry view. Methods Mol. Biol. 1374, 23–54. doi: 10.1007/978-1-4939-3167-5_2
Brinkrolf, K., Schneider, J., Knecht, M., Ruckert, C., and Tauch, A. (2012). Draft genome sequence of Turicella otitidis ATCC 51513, isolated from middle ear fluid from a child with otitis media. J. Bacteriol. 194, 5968–5969. doi: 10.1128/JB.01412-12
Brown, A. K., Bhatt, A., Singh, A., Saparia, E., Evans, A. F., and Besra, G. S. (2007). Identification of the dehydratase component of the mycobacterial mycolic acid-synthesizing fatty acid synthase-II complex. Microbiology 153(Pt 12), 4166–4173. doi: 10.1099/mic.0.2007/012419-0
Cole, S. T., Brosch, R., Parkhill, J., Garnier, T., Churcher, C., Harris, D., et al. (1998). Deciphering the biology of Mycobacterium tuberculosis from the complete genome sequence. Nature 393, 537–544. doi: 10.1038/31159
Collins, M. D., Burton, R. A., and Jones, D. (1988). Corynebacterium amycolatum sp. nov. a new mycolic acid-less Corynebacterium species from human skin. FEMS Microbiol. Lett. 49, 349–352. doi: 10.1111/j.1574-6968.1988.tb02755.x
Collins, M. D., Falsen, E., Akervall, E., Sjoden, B., and Alvarez, A. (1998). Corynebacterium kroppenstedtii sp. nov., a novel Corynebacterium that does not contain mycolic acids. Int. J. Syst. Bacteriol. 48(Pt 4), 1449–1454. doi: 10.1099/00207713-48-4-1449
Collins, M. D., Hoyles, L., Foster, G., and Falsen, E. (2004). Corynebacterium caspium sp. nov., from a Caspian seal (Phoca caspica). Int. J. Syst. Evol. Microbiol. 54(Pt 3), 925–928. doi: 10.1099/ijs.0.02950-0
Collins, M. D., and Jones, D. (1981). Distribution of isoprenoid quinone structural types in bacteria and their taxonomic implications. Microbiol. Rev. 45, 316–354.
Dhiman, R. K., Mahapatra, S., Slayden, R. A., Boyne, M. E., Lenaerts, A., Hinshaw, J. C., et al. (2009). Menaquinone synthesis is critical for maintaining mycobacterial viability during exponential growth and recovery from non-replicating persistence. Mol. Microbiol. 72, 85–97. doi: 10.1111/j.1365-2958.2009.06625.x
Felsenstein, J. (1985). Confidence Limits on phylogenies: an approach using the bootstrap. Evolution 39, 783–791. doi: 10.1111/j.1558-5646.1985.tb00420.x
Fernandez-Garayzabal, J. F., Vela, A. I., Egido, R., Hutson, R. A., Lanzarot, M. P., Fernandez-Garcia, M., et al. (2004). Corynebacterium ciconiae sp. nov., isolated from the trachea of black storks (Ciconia nigra). Int. J. Syst. Evol. Microbiol. 54(Pt 6), 2191–2195. doi: 10.1099/ijs.0.63165-0
Funke, G., Stubbs, S., Altwegg, M., Carlotti, A., and Collins, M. D. (1994). Turicella otitidis gen. nov., sp. nov., a coryneform bacterium isolated from patients with otitis media. Int. J. Syst. Bacteriol. 44, 270–273. doi: 10.1099/00207713-44-2-270
Gande, R., Dover, L. G., Krumbach, K., Besra, G. S., Sahm, H., Oikawa, T., et al. (2007). The two carboxylases of Corynebacterium glutamicum essential for fatty acid and mycolic acid synthesis. J. Bacteriol. 189, 5257–5264. doi: 10.1128/JB.00254-07
Gande, R., Gibson, K. J., Brown, A. K., Krumbach, K., Dover, L. G., Sahm, H., et al. (2004). Acyl-CoA carboxylases (accD2 and accD3), together with a unique polyketide synthase (Cg-pks), are key to mycolic acid biosynthesis in Corynebacterianeae such as Corynebacterium glutamicum and Mycobacterium tuberculosis. J. Biol. Chem. 279, 44847–44857. doi: 10.1074/jbc.M408648200
Gebhardt, H., Meniche, X., Tropis, M., Kramer, R., Daffe, M., and Morbach, S. (2007). The key role of the mycolic acid content in the functionality of the cell wall permeability barrier in Corynebacterineae. Microbiology 153(Pt 5), 1424–1434. doi: 10.1099/mic.0.2006/003541-0
Goodfellow, M., and Jones, A. L. (2015). Bergey’s Manual of Systematics of Archaea and Bacteria. New York, NY: Springer.
Goodfellow, M., Kampfer, P., Busse, H.-J., Trujillo, M. E., Suzuki, K.-I., Ludwig, W., et al. (2012). Bergey’s Manual of Systematics Bacteriology. New York, NY: Springer. doi: 10.1007/978-0-387-68233-4
Goyache, J., Ballesteros, C., Vela, A. I., Collins, M. D., Briones, V., Hutson, R. A., et al. (2003). Corynebacterium sphenisci sp. nov., isolated from wild penguins. Int. J. Syst. Evol. Microbiol. 53(Pt 4), 1009–1012. doi: 10.1099/ijs.0.02502-0
Hall, V., Collins, M. D., Hutson, R. A., Lawson, P. A., Falsen, E., and Duerden, B. I. (2003). Corynebacterium atypicum sp. nov., from a human clinical source, does not contain corynomycolic acids. Int. J. Syst. Evol. Microbiol. 53(Pt 4), 1065–1068. doi: 10.1099/ijs.0.02442-0
Hamada, M., Shibata, C., Sakurai, K., Hosoyama, A., Oji, S., Teramoto, K., et al. (2016). Reclassification of Amycolicicoccus subflavus as Hoyosella subflava comb. nov. and emended descriptions of the genus Hoyosella and Hoyosella altamirensis. Int. J. Syst. Evol. Microbiol. 66, 4711–4715. doi: 10.1099/ijsem.0.001415
He, Z., Zhang, H., Gao, S., Lercher, M. J., Chen, W. H., and Hu, S. (2016). Evolview v2: an online visualization and management tool for customized and annotated phylogenetic trees. Nucleic Acids Res. 44, W236–W241. doi: 10.1093/nar/gkw370
Ikeda, M., and Nakagawa, S. (2003). The Corynebacterium glutamicum genome: features and impacts on biotechnological processes. Appl. Microbiol. Biotechnol. 62, 99–109. doi: 10.1007/s00253-003-1328-1
Jeon, Y. S., Lee, K., Park, S. C., Kim, B. S., Cho, Y. J., Ha, S. M., et al. (2014). EzEditor: a versatile sequence alignment editor for both rRNA- and protein-coding genes. Int. J. Syst. Evol. Microbiol. 64(Pt 2), 689–691. doi: 10.1099/ijs.0.059360-0
Jurado, V., Kroppenstedt, R. M., Saiz-Jimenez, C., Klenk, H. P., Mouniee, D., Laiz, L., et al. (2009). Hoyosella altamirensis gen. nov., sp. nov., a new member of the order Actinomycetales isolated from a cave biofilm. Int. J. Syst. Evol. Microbiol. 59(Pt 12), 3105–3110. doi: 10.1099/ijs.0.008664-0
Kämpfer, P., Andersson, M. A., Rainey, F. A., Kroppenstedt, R. M., and Salkinoja-Salonen, M. (1999). Williamsia muralis gen. nov., sp. nov., isolated from the indoor environment of a children’s day care centre. Int. J. Syst. Bacteriol. 49(Pt 2), 681–687. doi: 10.1099/00207713-49-2-681
Katayama, T., Kato, T., Tanaka, M., Douglas, T. A., Brouchkov, A., Abe, A., et al. (2010). Tomitella biformata gen. nov., sp. nov., a new member of the suborder corynebacterineae isolated from a permafrost ice wedge. Int. J. Syst. Evol. Microbiol. 60, 2803–2807. doi: 10.1099/ijs.0.017962-0
Katoh, K., and Standley, D. M. (2014). MAFFT: iterative refinement and additional methods. Methods Mol. Biol. 1079, 131–146. doi: 10.1007/978-1-62703-646-7_8
Laneelle, M. A., Launay, A., Spina, L., Marrakchi, H., Laval, F., Eynard, N., et al. (2012). A novel mycolic acid species defines two novel genera of the Actinobacteria. Hoyosella and Amycolicicoccus. Microbiology 158(Pt 3),843–855. doi: 10.1099/mic.0.055509-0
Lehmann, K. B., and Neumann, R. (1896). Atlas und Grundriss der Bakteriologie und Lehrbuch der speciellen bakteriologischen Diagnostik. Am. Nat. 364, 312–314.
Li, X. J., Liu, S. W., Jiang, Z. K., Wu, G., and Sun, C. H. (2016). Hoyosella rhizosphaerae sp. nov., a halotolerant actinobacterium isolated from rhizosphere soil of Suaeda salsa, and emended description of the genus Hoyosella. Int. J. Syst. Evol. Microbiol. 66, 4716–4722. doi: 10.1099/ijsem.0.001416
Liu, K., Linder, C. R., and Warnow, T. (2011). RAxML and FastTree: comparing two methods for large-scale maximum likelihood phylogeny estimation. PLoS One 6:e27731. doi: 10.1371/journal.pone.0027731
Marrakchi, H., Laneelle, M. A., and Daffe, M. (2014). Mycolic acids: structures, biosynthesis, and beyond. Chem. Biol. 21, 67–85. doi: 10.1016/j.chembiol.2013.11.011
Na, S.-I., Kim, Y. O., Yoon, S.-H., Ha, S.-M., Baek, I., and Chun, J. (2018). UBCG: Up-to-date bacterial core gene set and pipeline for phylogenomic tree reconstruction. J. Microbiol. 56, 281–285. doi: 10.1007/s12275-018-8014-6
Nowicka, B., and Kruk, J. (2010). Occurrence, biosynthesis and function of isoprenoid quinones. Biochim. Biophys. Acta 1797, 1587–1605. doi: 10.1016/j.bbabio.2010.06.007
Parish, T., Roberts, G., Laval, F., Schaeffer, M., Daffe, M., and Duncan, K. (2007). Functional complementation of the essential gene fabG1 of Mycobacterium tuberculosis by Mycobacterium smegmatis fabG but not Escherichia coli fabG. J. Bacteriol. 189, 3721–3728. doi: 10.1128/JB.01740-06
Portevin, D., De Sousa-D’Auria, C., Houssin, C., Grimaldi, C., Chami, M., Daffe, M., et al. (2004). A polyketide synthase catalyzes the last condensation step of mycolic acid biosynthesis in mycobacteria and related organisms. Proc. Natl. Acad. Sci. U.S.A. 101, 314–319. doi: 10.1073/pnas.0305439101
Portevin, D., de Sousa-D’Auria, C., Montrozier, H., Houssin, C., Stella, A., Laneelle, M. A., et al. (2005). The acyl-AMP ligase FadD32 and AccD4-containing acyl-CoA carboxylase are required for the synthesis of mycolic acids and essential for mycobacterial growth: identification of the carboxylation product and determination of the acyl-CoA carboxylase components. J. Biol. Chem. 280, 8862–8874. doi: 10.1074/jbc.M408578200
Price, M. N., Dehal, P. S., and Arkin, A. P. (2010). FastTree 2–approximately maximum-likelihood trees for large alignments. PLoS One 5:e9490. doi: 10.1371/journal.pone.0009490
Rost, B. (1999). Twilight zone of protein sequence alignments. Protein Eng. 12, 85–94. doi: 10.1093/protein/12.2.85
Russell, N. J. (1984). Mechanisms of thermal adaptation in bacteria - blueprints for survival. Trends Biochem. Sci. 9, 108–112. doi: 10.1016/0968-0004(84)90106-3
Sacco, E., Covarrubias, A. S., O’Hare, H. M., Carroll, P., Eynard, N., Jones, T. A., et al. (2007). The missing piece of the type II fatty acid synthase system from Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. U.S.A. 104, 14628–14633. doi: 10.1073/pnas.0704132104
Saddler, G. S., Goodfellow, M., Minnikin, D. E., and O’Donnell, A. G. (1986). Influence of the growth cycle on the fatty acid and menaquinone composition of Streptomyces cyaneus NCIB 9616. J. Appl. Microbiol. 60, 51–56.
Segata, N., Bornigen, D., Morgan, X. C., and Huttenhower, C. (2013). PhyloPhlAn is a new method for improved phylogenetic and taxonomic placement of microbes. Nat. Commun. 4:2304. doi: 10.1038/ncomms3304
Seto, H., Jinnai, Y., Hiratsuka, T., Fukawa, M., Furihata, K., Itoh, N., et al. (2008). Studies on a new biosynthetic pathway for menaquinone. J. Am. Chem. Soc. 130, 5614–5615. doi: 10.1021/ja710207s
Stamatakis, A. (2014). RAxML version 8: a tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 30, 1312–1313. doi: 10.1093/bioinformatics/btu033
Sutcliffe, I. C., Trujillo, M. E., and Goodfellow, M. (2012). A call to arms for systematists: revitalising the purpose and practises underpinning the description of novel microbial taxa. Antonie Van Leeuwenhoek 101, 13–20. doi: 10.1007/s10482-011-9664-0
Sutcliffe, I. C., Trujillo, M. E., Whitman, W. B., and Goodfellow, M. (2013). A call to action for the international committee on systematics of prokaryotes. Trends Microbiol. 21, 51–52. doi: 10.1016/j.tim.2012.11.004
Takeno, S., Takasaki, M., Urabayashi, A., Mimura, A., Muramatsu, T., Mitsuhashi, S., et al. (2013). Development of fatty acid-producing Corynebacterium glutamicum strains. Appl. Environ. Microbiol. 79, 6776–6783. doi: 10.1128/AEM.02003-13
Tauch, A., Kaiser, O., Hain, T., Goesmann, A., Weisshaar, B., Albersmeier, A., et al. (2005). Complete genome sequence and analysis of the multiresistant nosocomial pathogen Corynebacterium jeikeium K411, a lipid-requiring bacterium of the human skin flora. J. Bacteriol. 187, 4671–4682. doi: 10.1128/JB.187.13.4671-4682.2005
Tauch, A., Trost, E., Tilker, A., Ludewig, U., Schneiker, S., Goesmann, A., et al. (2008). The lifestyle of Corynebacterium urealyticum derived from its complete genome sequence established by pyrosequencing. J. Biotechnol. 136, 11–21. doi: 10.1016/j.jbiotec.2008.02.009
Tippelt, A., Mollmann, S., Albersmeier, A., Jaenicke, S., Ruckert, C., and Tauch, A. (2014). Mycolic acid biosynthesis genes in the genome sequence of Corynebacterium atypicum DSM 44849. Genome Announc. 2:e00845-14. doi: 10.1128/genomeA.00845-14
Upadhyay, A., Fontes, F. L., Gonzalez-Juarrero, M., McNeil, M. R., Crans, D. C., Jackson, M., et al. (2015). Partial saturation of menaquinone in Mycobacterium tuberculosis: function and essentiality of a novel reductase. ACS Cent. Sci. 1, 292–302. doi: 10.1021/acscentsci.5b00212
Vilcheze, C., Morbidoni, H. R., Weisbrod, T. R., Iwamoto, H., Kuo, M., Sacchettini, J. C., et al. (2000). Inactivation of the inhA-encoded fatty acid synthase II (FASII) enoyl-acyl carrier protein reductase induces accumulation of the FASI end products and cell lysis of Mycobacterium smegmatis. J. Bacteriol. 182, 4059–4067. doi: 10.1128/JB.182.14.4059-4067.2000
Wang, Y. N., Chi, C. Q., Cai, M., Lou, Z. Y., Tang, Y. Q., Zhi, X. Y., et al. (2010). Amycolicicoccus subflavus gen. nov., sp. nov., an actinomycete isolated from a saline soil contaminated by crude oil. Int. J. Syst. Evol. Microbiol. 60(Pt 3), 638–643. doi: 10.1099/ijs.0.010546-0
Wiertz, R., Schulz, S. C., Muller, U., Kampfer, P., and Lipski, A. (2013). Corynebacterium frankenforstense sp. nov. and Corynebacterium lactis sp. nov., isolated from raw cow milk. Int. J. Syst. Evol. Microbiol. 63(Pt 12), 4495–4501. doi: 10.1099/ijs.0.050757-0
Wu, D., Jospin, G., and Eisen, J. A. (2013). Systematic identification of gene families for use as “markers” for phylogenetic and phylogeny-driven ecological studies of bacteria and archaea and their major subgroups. PLoS One 8:e77033. doi: 10.1371/journal.pone.0077033
Wu, M., and Eisen, J. A. (2008). A simple, fast, and accurate method of phylogenomic inference. Genome Biol. 9:R151. doi: 10.1186/gb-2008-9-10-r151
Yoon, S. H., Ha, S. M., Kwon, S., Lim, J., Kim, Y., Seo, H., et al. (2017). Introducing EzBioCloud: a taxonomically united database of 16S rRNA gene sequences and whole-genome assemblies. Int. J. Syst. Evol. Microbiol. 67, 1613–1617. doi: 10.1099/ijsem.0.001755
Keywords: Turicella otitidis, Corynebacterium, phylogenomic analysis, chemotaxonomy, mycolic acid, menaquinone
Citation: Baek I, Kim M, Lee I, Na S-I, Goodfellow M and Chun J (2018) Phylogeny Trumps Chemotaxonomy: A Case Study Involving Turicella otitidis. Front. Microbiol. 9:834. doi: 10.3389/fmicb.2018.00834
Received: 28 February 2018; Accepted: 12 April 2018;
Published: 30 April 2018.
Edited by:
Svetlana N. Dedysh, Winogradsky Institute of Microbiology (RAS), RussiaReviewed by:
André Lipski, Universität Bonn, GermanyCopyright © 2018 Baek, Kim, Lee, Na, Goodfellow and Chun. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jongsik Chun, amNodW5Ac251LmFjLmty
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.