
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 18 April 2018
Sec. Antimicrobials, Resistance and Chemotherapy
Volume 9 - 2018 | https://doi.org/10.3389/fmicb.2018.00687
This article is part of the Research Topic New Frontiers in the Search of Antimicrobials Agents From Natural Products, Volume II View all 67 articles
 Meeta Masand1‡
Meeta Masand1‡ Kunjukrishnan Kamalakshi Sivakala2‡
Kunjukrishnan Kamalakshi Sivakala2‡ Ekta Menghani3
Ekta Menghani3 Thangathurai Thinesh4
Thangathurai Thinesh4 Rangasamy Anandham5
Rangasamy Anandham5 Gaurav Sharma1
Gaurav Sharma1 Natesan Sivakumar2
Natesan Sivakumar2 Solomon R. D. Jebakumar2§
Solomon R. D. Jebakumar2§ Polpass Arul Jose2*†
Polpass Arul Jose2*†Acquisition of Actinobacteria, especially Streptomyces from previously underexplored habitats and the exploration of their biosynthetic potential have gained much attention in the rejuvenated antibiotics search programs. Herein, we isolated some Streptomyces strains, from an arid region of the Great Indian Thar Desert, which possess an ability to produce novel bioactive compounds. Twenty-one morphologically distinctive strains differing in their aerial and substrate mycelium were isolated by employing a stamping method. Among them, 12 strains were identified by a two-level antimicrobial screening method, exerting antimicrobial effects against a panel of indicator strains including methicillin-resistant Staphylococcus aureus and vancomycin-resistant Enterococcus species. Based on their potent antimicrobial activity, four isolates were further explored by 16S rRNA gene-based identification, genetic screening, and metabolomic analysis; and it was found that these strains belong to the genus Streptomyces. The selected strains were found to have polyketide synthase and non-ribosomal peptide synthetase systems. In addition, extracellular metabolomic screening revealed that the isolates produced analogs of doxorubicinol, pyrromycin, erythromycin, and 6-13 other putative novel metabolites. These results demonstrate the significance of Streptomyces inhabiting the arid region of Thar Desert, suggesting that similar arid environments can be considered as the reservoirs of novel Streptomyces strains that could have biotechnological significance.
The escalating levels of antibiotic resistance in pathogenic microorganisms clearly dictate the need for identifying new bioactive compounds for novel drug discovery. Microorganisms, especially Actinobacteria, have been isolated from diverse environments and have most frequently been explored as a novel source for the production of bioactive compounds (Peraud et al., 2009; Poulsen et al., 2011; Subramani and Aalbersberg, 2012; Jose et al., 2013; Jose and Jebakumar, 2014; Sun et al., 2015; Singh et al., 2016). In the past 75 years, the Actinobacteria have exerted a major impact on the discovery of antibiotics and several other drugs (Baltz, 2007; Procópio et al., 2012), and continue as one of the most important sources of chemical diversity. Consequently, it makes good sense to isolate and systematically identify the potent candidates affiliated to Actinobacteria in the search for new bioactive compounds.
Recently, the search for bioactive metabolites has intensified worldwide, aided by the advancements in next generation sequencing technologies (Gomez-Escribano et al., 2016), spectroscopic methods (Forner et al., 2013; Derewacz et al., 2015; Floros et al., 2016), and bulk data processing platforms (Floros et al., 2016). There is notable rejuvenation in secondary metabolites search from Streptomyces (Gomez-Escribano et al., 2016), attested by increasing number of genome announcements and guided discoveries (Bachmann et al., 2014; Ju et al., 2015; Gomez-Escribano et al., 2016). Moreover, there are a number of reports on novel isolates with biosynthetic potential from novel sources explored by metabolomic and genomic screening methods (Guo et al., 2015; Sengupta et al., 2015).
The Actinobacteria associated with unexplored and underexplored habitats are considered as a novel source of biologically active metabolites (Guo et al., 2015; Sengupta et al., 2015). In this context, deserts are underexplored environments with limited reports on their microbiota endowed with the potential to produce novel bio-active metabolites (Takahashi et al., 1996; Hozzein et al., 2008; Okoro et al., 2009). Moreover, the desert environments (Mohammadipanah and Wink, 2016) are relatively less explored as source of novel Actinobacteria when compared to endosphere of plants (Masand et al., 2015), marine sediments (Jose and Jha, 2017), and marine animals (Mahmoud and Kalendar, 2016; Dhakal et al., 2017). A high diversity and suggested endemicity of culturable Actinobacteria have recently been found in an extremely oligotrophic desert oasis (Arocha-Garza et al., 2017).
The Great Indian Thar Desert is the ninth largest subtropical desert, which comprises of a range of arid to subhumid climatic conditions. Reports on the exploration and/or exploitation of Actinobacteria in this region are very limited, with the exception of a recent finding which claimed the recovery of 13 bioactive Actinobacteria from the Jaisalmer and Jodhpur regions of Thar Desert (Tiwari et al., 2015). This study was carried out in Bikaner, which is located right in the middle of the Thar Desert. While there is no previous report, our current focus is based on the modern metabolomics based assessment for novel Streptomyces with potential for producing bioactive metabolites. This has led to the isolation of potentially active strains that produce metabolites against pathogenic bacterial strains. We describe the isolation, molecular identification, antimicrobial potential, and secondary metabolites from these isolates.
Arid soil samples were collected from Bikaner, located in the Great Indian Thar Desert (Figure 1). Bikaner is situated between 28°01′00′′N and 73°18′43′′E, and has a hot arid climate. A total of eight soil samples were collected within a 128 m2 plot at a depth of 10-15 cm from the upper surface of the topsoil and stored at 4°C in labeled sterile bags before transporting to our laboratory for further processing.
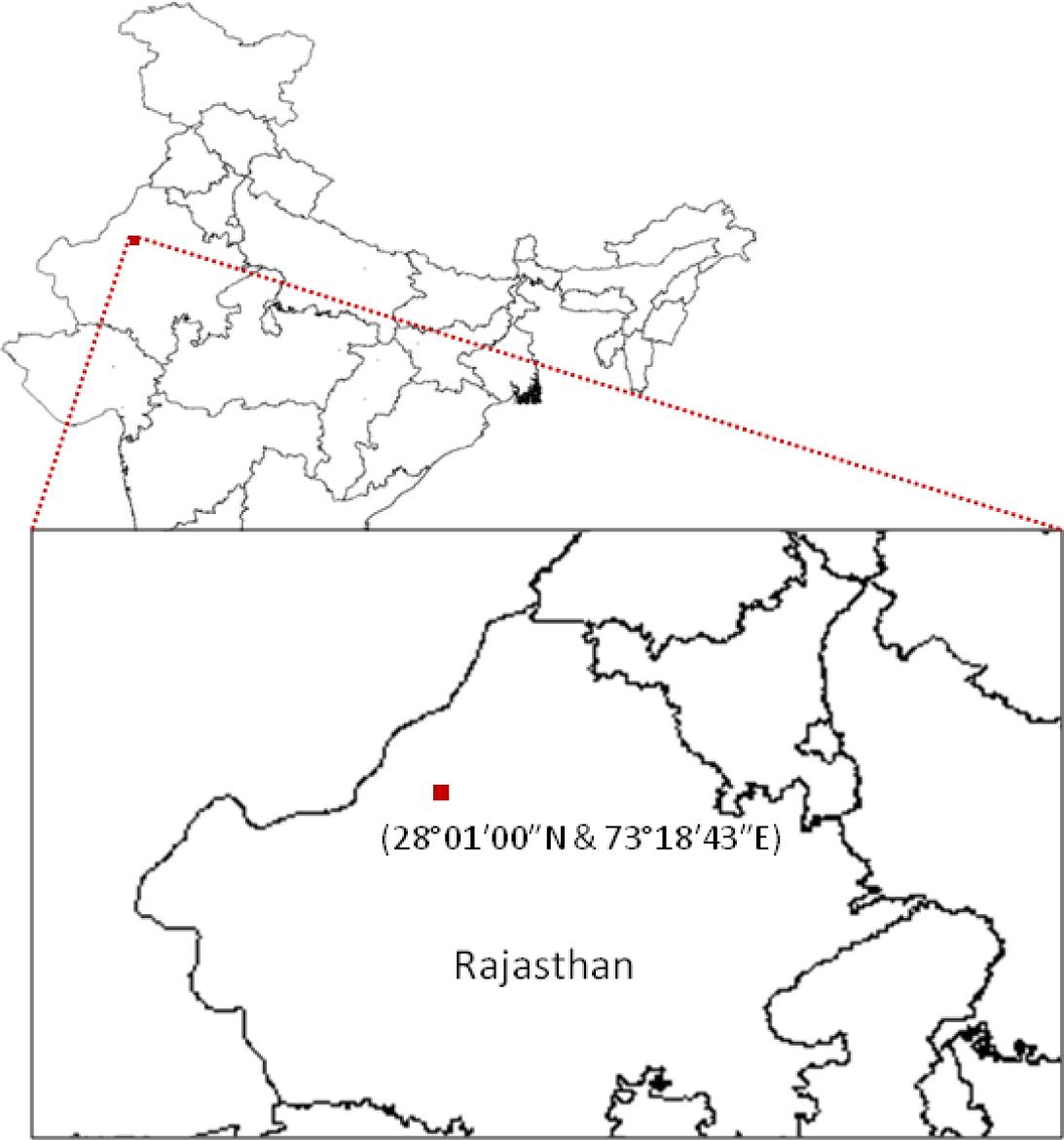
FIGURE 1. Sampling site. The map shows the sample collection site in the Thar Desert, Rajasthan (India).
Three different isolation media were employed: starch casein agar (10.0 g soluble starch, 0.3 g casein, 2.0 g KNO3, 2.0 g NaCl, 2.0 g K2HPO4, 0.05 g MgSO4.7H2O, 0.02 g CaCO3, 0.01 g FeSO4.7H2O, 20.0 g agar in 1,000 mL distilled water; pH 7.0), Actinobacteria isolation agar (2.0 g sodium caseinate, 0.1 g asparagine, 4.0 g sodium propionate, 0.5 g K2HPO4, 0.1 g MgSO4.7H2O, 0.001 g FeSO4.7H2O and 20.0 g agar in 1,000 mL distilled water; pH 7.0) and modified MM agar (1.0 g glucose, 0.5 g yeast extract, 1.0 g K2HPO4, 0.5 g MgSO4⋅7H2O, and 0.5 g NaCl, and 20 g agar). All the three media were supplemented with antibiotics, cycloheximide (100 μg mL-1), and nalidixic acid (25 μg mL-1) to reduce fungal growth and contamination by fast growing bacteria.
All the soil samples were dried in a laminar flow hood for 24 h. The samples were then processed by using the heat-shock-and-dilution and plate stamping methods (Mincer et al., 2002; Jose and Jebakumar, 2013) with a minor modification, and inoculated onto the agar media. In heat-shock-and-dilution method, sterile water was used in the place of seawater. The plates were incubated for 2–10 weeks at 28–32°C and observed for the appearance of Actinobacteria-like colonies. Well-separated Actinobacteria-like colonies were removed from the original isolation plates and sub-cultured on modified ISP 4 medium (Jose and Jebakumar, 2013) to recover pure cultures with uniform colony morphology. Pure cultures of the isolates were again streaked on modified ISP4 medium and incubated for 10 days at 30°C. Aerial mycelium, substrate mycelium, pigmentation and sporulation were observed after the incubation period, and unique isolates were selected for further studies.
Isolates collected from the semi-arid soil samples were screened for antibacterial activity by cross streak (Lemos et al., 1985) and agar-plug methods (Jose and Jebakumar, 2013) against four bacteria and a yeast strain. In the cross streak method, the isolates were streaked on starch casein agar and modified ISP4 agar medium, and incubated at 29 ± 2°C for 7 days to obtain a copious ribbon-like growth. Then, overnight cultures of three clinical pathogens [methicillin-resistant Staphylococcus aureus (MRSA), vancomycin-resistant Enterococcus (VRE), and Candida albicans], and two type strains (Pseudomonas aeruginosa ATCC 10145, Escherichia coli ATCC 3739) were cross streaked at a 90° angle to actinobacterial cultures. Subsequently, plates were incubated at 29 ± 2°C for 24 h, and the zone of inhibition was observed.
In the agar plug method, the isolates were initially inoculated in the agar medium described above and incubated until sufficient growth was achieved. Then, agar plugs of 6 mm in diameter were cut from the 10-day-old agar plates and plugged into wells, bored using a sterile cork borer (diameter of 6 mm), in the Mueller–Hinton agar plates, which were seeded with bacteria and yeast. The agar plugged plates were incubated at 37°C for 24 h and observed for the zone of inhibition around the inserted agar plugs. Clinical pathogens used in this study were obtained from Kovai Medical Centre and Hospital (KMCH, Coimbatore, India).
Four potential isolates were selected for 16S rRNA gene sequencing and phylogenetic analysis. The strains were cultured in 50 mL of trypticase soy broth, shaken at 180 rpm at 29 ± 2°C for 7 days, and the resulting mycelia biomass was pelleted by centrifugation. Genomic DNA was extracted using HiPurA Streptomyces genomic DNA purification kit (HiMedia, India), according to the manufacturer’s protocol. The 16S rRNA gene was amplified using universal 27F (5′-AGA GTTTGATCCTGGCTCA-3′) and 1492R (5′-ACGGCTACCTTGTTACGACT-3′) primers with the following thermal cycling conditions: initial denaturation at 95°C for 5 min followed by 32 cycles at 95°C for 30 s, 55°C for 90 s and 72°C for 120 s, followed by a final extension at 72°C for 5 min. The reactions were performed in MyCycler (Bio-Rad, United States) and the amplified products were examined by 1% agarose gel electrophoresis. The PCR products were purified and sequenced by Applied Biosystems 3730XL DNA analyzer. The forward and reverse 16S rRNA gene sequences, obtained from each strain, were assembled and analyzed using Basic Local Alignment Search Tool (Altschul et al., 1990). To identify the isolates, the sequences were uploaded to EzTaxon server (Kim et al., 2012) and compared with 16S rRNA gene sequence of the type strains. The Ribosomal Database Project online tool (Cole et al., 2014) was used to align the sequences with the related 16S rRNA gene sequences from GenBank and downloaded in the FASTA format. The sequence file was imported into MEGA5 (Tamura et al., 2011) and a neighbor-joining phylogenetic tree was constructed with 1,000 bootstrap replicates.
Selected antagonistic actinobacterial strains SAS02, SAS09, SAS13, and SAS15 were inoculated from frozen stocks into 50 mL of modified ISP4 medium and shaken at 180 rpm and 29 ± 2°C for 3 days. The cultures were then transferred to 1 L of the same medium and incubated at the same condition for 7 days. After incubation, the cultures were filtered and the cell-free filtrate was extracted with an equal volume of ethyl acetate which is often employed for secondary metabolites extraction (Becerril-Espinosa et al., 2013; Cheng et al., 2015; Wu et al., 2016). The organic layer was separated, concentrated under vacuum, and dissolved in methanol until HR-LCMS analysis.
Ketosynthase (KS) and adenylation domain fragments of polyketide synthase (PKS) and non-ribosomal peptide synthetase (NRPS) systems, respectively, were PCR amplified using 20–50 ng of genomic DNA. The degenerate primers used in the study are given in Table 1. PCR conditions were as follows: an initial denaturation at 95°C for 5 min, followed by 31 cycles of 30 s at 95°C, 1 min at 58°C (KS) or 61°C (NRPS), and 1 min at 72°C, and a final extension at 72°C for 7 min. The tubes without template DNA were used as negative control and the tubes with DNA of previously characterized strain Streptomyces sp. JAJ06 (Jose et al., 2011) bearing PKS and NRPS systems were considered as positive controls. PCR products were checked by agarose gel (1%, w/v) electrophoresis.

TABLE 1. Primers used in PCR-based screening of isolates for PKS and NRPS biosynthetic systems.
The crude extracts were subjected to HR-LCMS analysis in an Infinity Nano HPLC-Chip cube system coupled with iFunnel MS-Q-TOFs (Agilent Technologies, United States). The sample injection volume was 5 μL. The compound separation was performed at a flow rate of 0.3 mL/min under a gradient program. Mobile phase A was HPLC grade water containing 0.1% (v/v) formic acid and mobile phase B was HPLC grade acetonitrile supplemented with 10% (v/v) water and 0.1% (v/v) formic acid. The separation was performed in gradient mode, using 5–95% of mobile phase B in mobile phase A over 20 min. For MS, dual Agilent Jet Stream ESI source was used and data were acquired in both positive and negative modes. Spectral data were analyzed using Agilent Mass Hunter Qualitative Analysis software (version B.05.00; Agilent Technologies) and compared with known compounds based on similar molecular features. HR-LCMS analysis was performed at the Sophisticated Analytical Instrumentation Facility (SAIF), Indian Institute of Technology, Mumbai, India.
A total of 21 morphologically unique Actinobacteria were isolated from the eight soil samples collected from the Thar Desert. The pure culture of the isolates was obtained by a series of repeated streaking on an agar medium. The isolates that showed distinct colony morphology (Figure 2), indicative of the order Actinomycetales, were taken for further analysis. Among the two different pre-treatment techniques employed, plate stamping resulted in the isolation of 16 (76%) strains. Heat-shock-dilution method resulted in isolation of relatively less number of (5, 24%) strains. Morphological characteristics of the 21 isolates are summarized in Table 2.

FIGURE 2. Morphological characterization of actinobacterial isolates. Colony morphology of different actinobacterial isolates derived from semi-arid soil samples was photographed after growing on modified ISP4 media for 10 days.
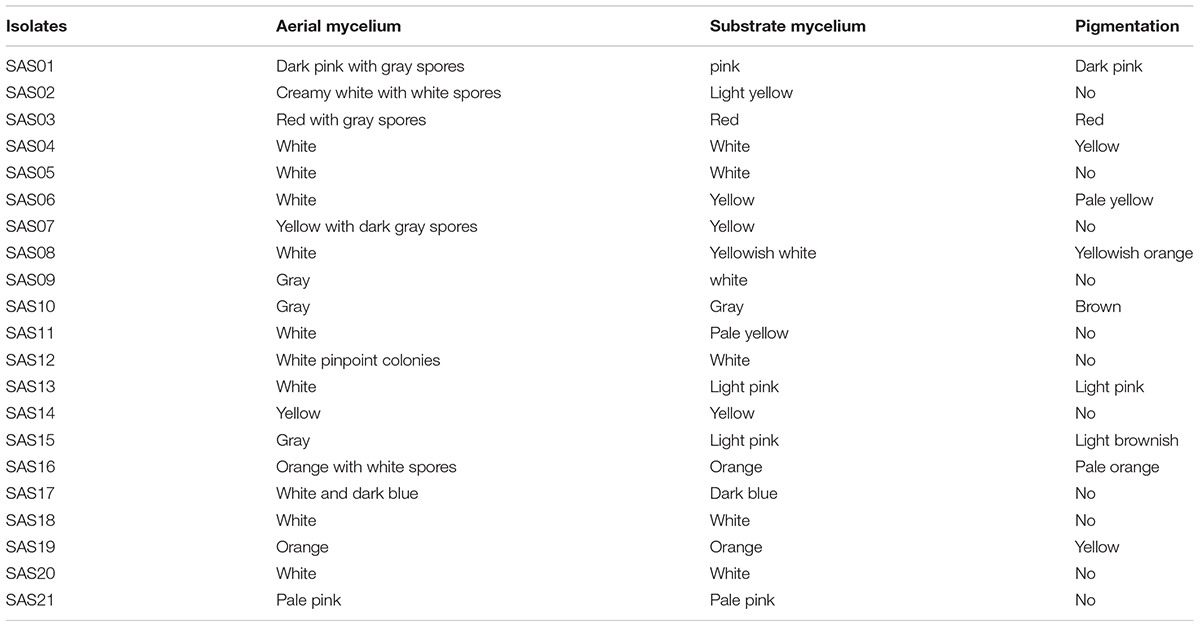
TABLE 2. Morphological characteristics of actinobacterial isolates acquired from the desert soils.
We screened the different isolates at two levels with two different production media against both the clinical pathogens (clinical isolates) and ATCC type strains. Notably, in the cross-streak method, where the antimicrobial activity of the isolates was tested against one yeast and five different bacterial strains, a total of 12 (57%) actinobacterial isolates were found to produce bioactive metabolites, in either or both SCA and M-ISP4, against at least one of the test strains. The antimicrobial metabolite producers included the following strains: SAS01–SAS06, SAS08, SAS09, SAS11, SAS13, SAS15, and SAS19.
Results of secondary screening for antimicrobial activity were presented as the diameter of inhibition zone against the test strains (Table 3). Out of the 12 antimicrobial isolates identified, nine (75%) exhibited considerable inhibitory activity against both Gram-negative and Gram-positive bacteria. The remaining three (25%) isolates exhibited antibacterial activity against only Gram-positive bacteria. In the case of antifungal activity against Candida albicans, except for SAS08 and SA11, all the isolates showed considerable growth inhibition. Based on the broad-spectrum activity exhibited, strains SAS02, SAS09, SAS13, and SAS15 (Figure 3) were selected for further analysis, including molecular identification and secondary metabolites analysis.
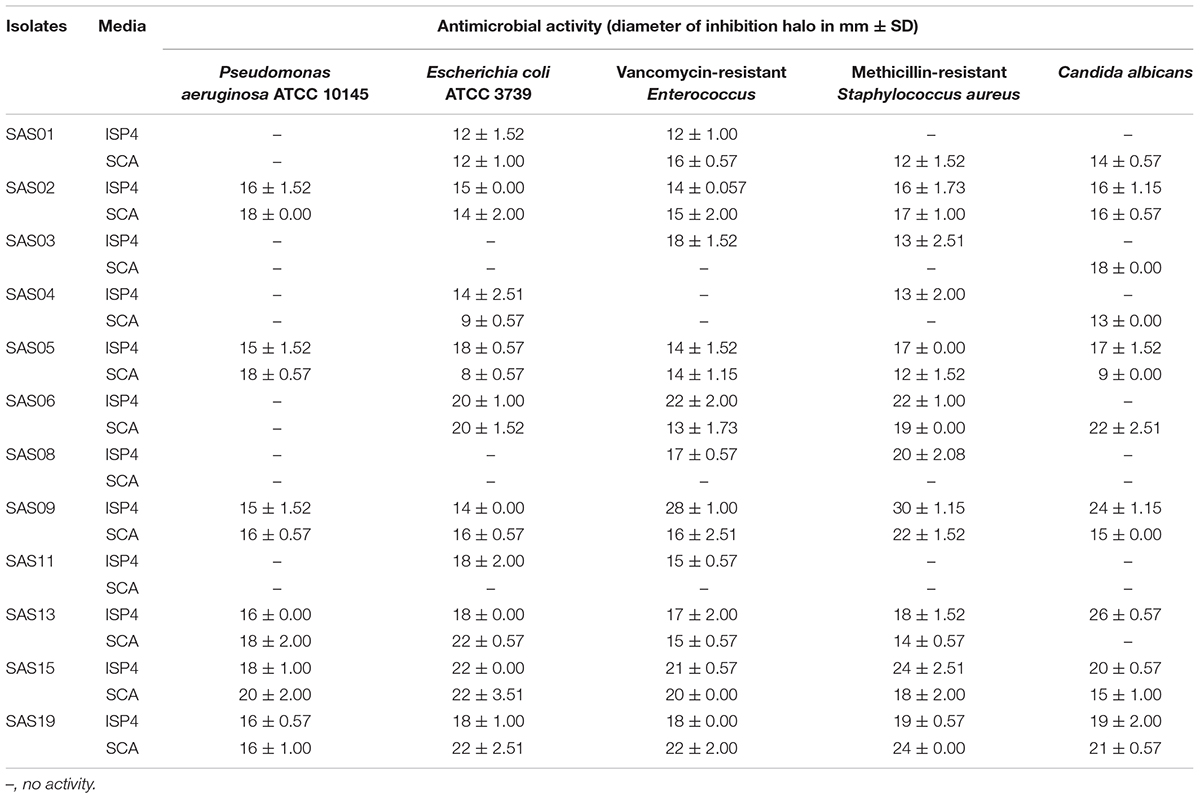
TABLE 3. Antimicrobial activity of active isolates against both clinical and ATCC strains.
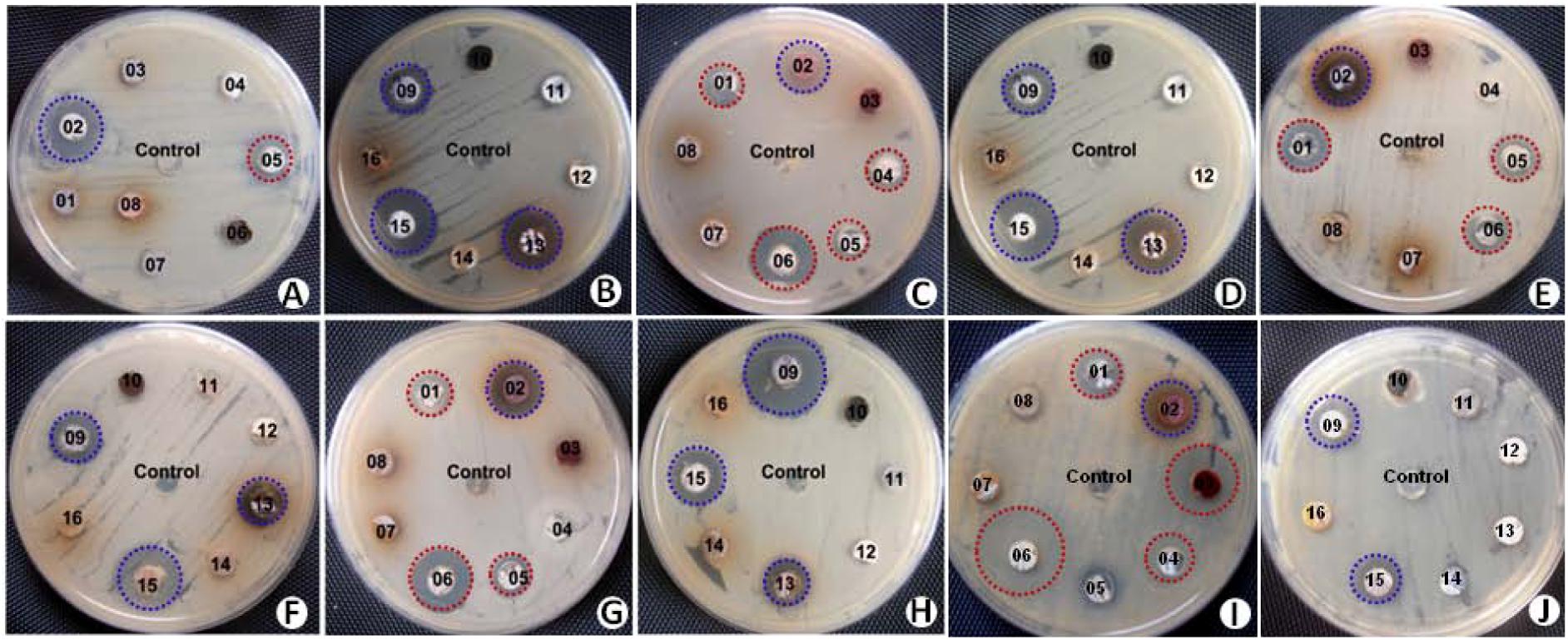
FIGURE 3. Screening of actinobacterial isolates for antimicrobial activity against series of test strains by agar-plug method. Antimicrobial assay plates showing antimicrobial activity of selected strains (shaded with blue color) against Pseudomonas aeruginosa ATCC 10145 (A,B), Escherichia coli ATCC 3739 (C,D), vancomycin-resistant Enterococcus (E,F), methicillin-resistant Staphylococcus aureus (G,H), and Candida albicans (I,J).
The phylogeny of the selected strains, which showed broad-spectrum antimicrobial activity, was evaluated on the basis of their partial 16S rRNA gene sequences. PCR amplification, sequencing, and processing of 16S rRNA gene fragment yielded final contigs of the size ranging from 1,375 bp to 1,391 bp. The identification of 16S rRNA gene sequence using EzTaxon server revealed that all the four isolates belong to a single genus, Streptomyces. Table 4 summarizes the 16S rRNA gene sequence coverage, accession numbers, and similarity with known strains.
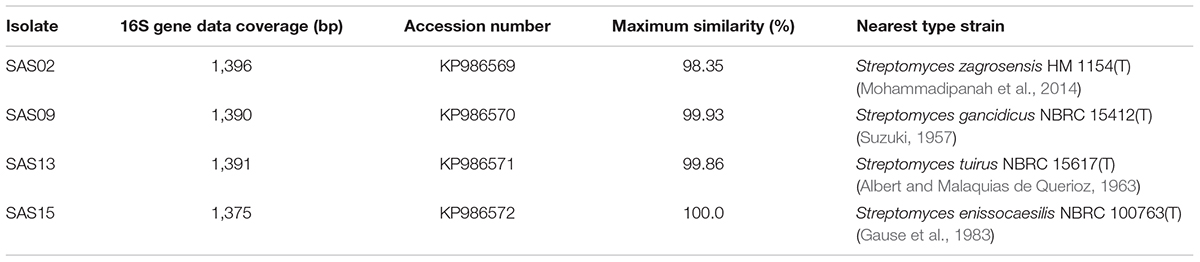
TABLE 4. 16S rRNA gene sequence data in comparison with nearest valid members of Streptomyces genus.
Strain SAS02 showed 93.84–98.35% sequence similarity with the members of genus Streptomyces, with the maximum (98.35 %) similarity to Streptomyces zagrosensis HM 1154T (Mohammadipanah et al., 2014). Strain SAS09 shared 93.44–98.93% sequence similarity to the members of Streptomyces, with the highest similarity to Streptomyces gancidicus NBRC 15412T (Suzuki, 1957). Strain SAS13 showed 93.38–99.86% sequence similarity, showing maximum similarity to Streptomyces tuirus NBRC 15617T (Albert and Malaquias de Querioz, 1963). Strain SAS15 showed 92.94–100% similarity to the members of Streptomyces, with the highest similarity to Streptomyces enissocaesilis NRRL B–16365T (Gause et al., 1983). Consequently, SAS02, SAS09, SAS13, and SAS15 were designated as Streptomyces sp. SAS02, Streptomyces sp. SAS09, Streptomyces sp. SAS13, and Streptomyces enissocaesilis strain SAS15, respectively.
The neighbor-joining phylogenetic tree analysis segregated SAS02, SAS09, SAS13, and SAS15 into three different clusters (Figure 4) representing the genus Streptomyces. Strains SAS13 and SAS15 were assigned to the same cluster, while SAS02 and SAS09 were grouped into two separate clusters. Interestingly, strain SAS02 clearly separated from its nearest strain type, suggesting that it may be a novel strain.
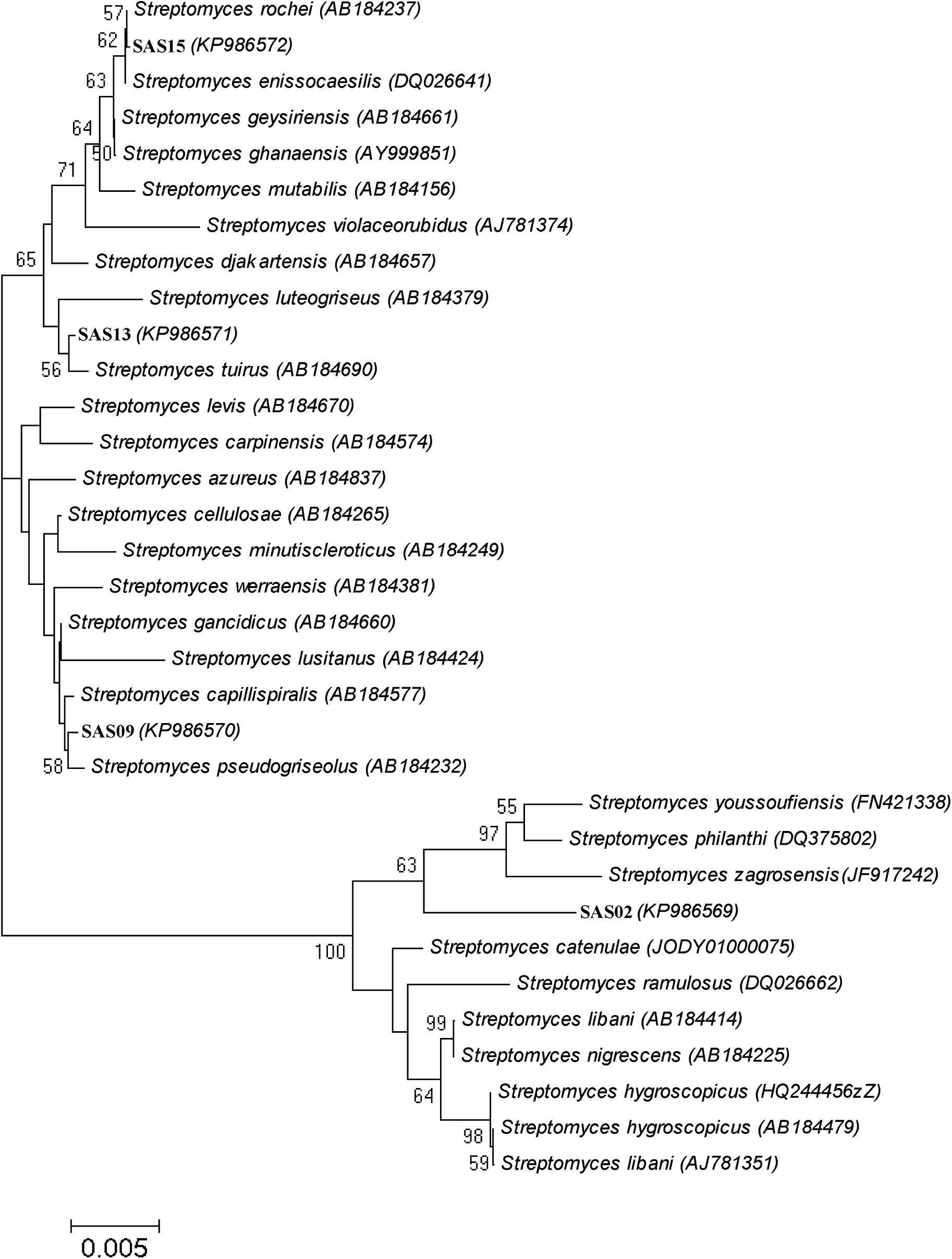
FIGURE 4. Phylogenetic analysis of selected actinobacterial strains. Neighbor-joining phylogenetic tree showing evolutionary relationships of selected isolates based on 16S rRNA sequence alignments. Organisms represented are the isolates from this study and their closest valid GenBank matches. Numbers at nodes indicate percentages of 1,000 bootstrap resamplings; only values above 50% are given. Bar indicates 0.005 substitutions per nucleotide position.
The biotechnological significance of the isolates was examined using PCR-based screening for type II PKS and NRPS systems (Figure 5). Strains SAS02, SAS09, and SAS13 were positive for both type II PKS and NRPS systems, whereas SAS15 was positive only for NRPS system. Presence of type II PKS and NRPS systems proved biosynthetic potential of the selected isolates that showed antimicrobial activity.
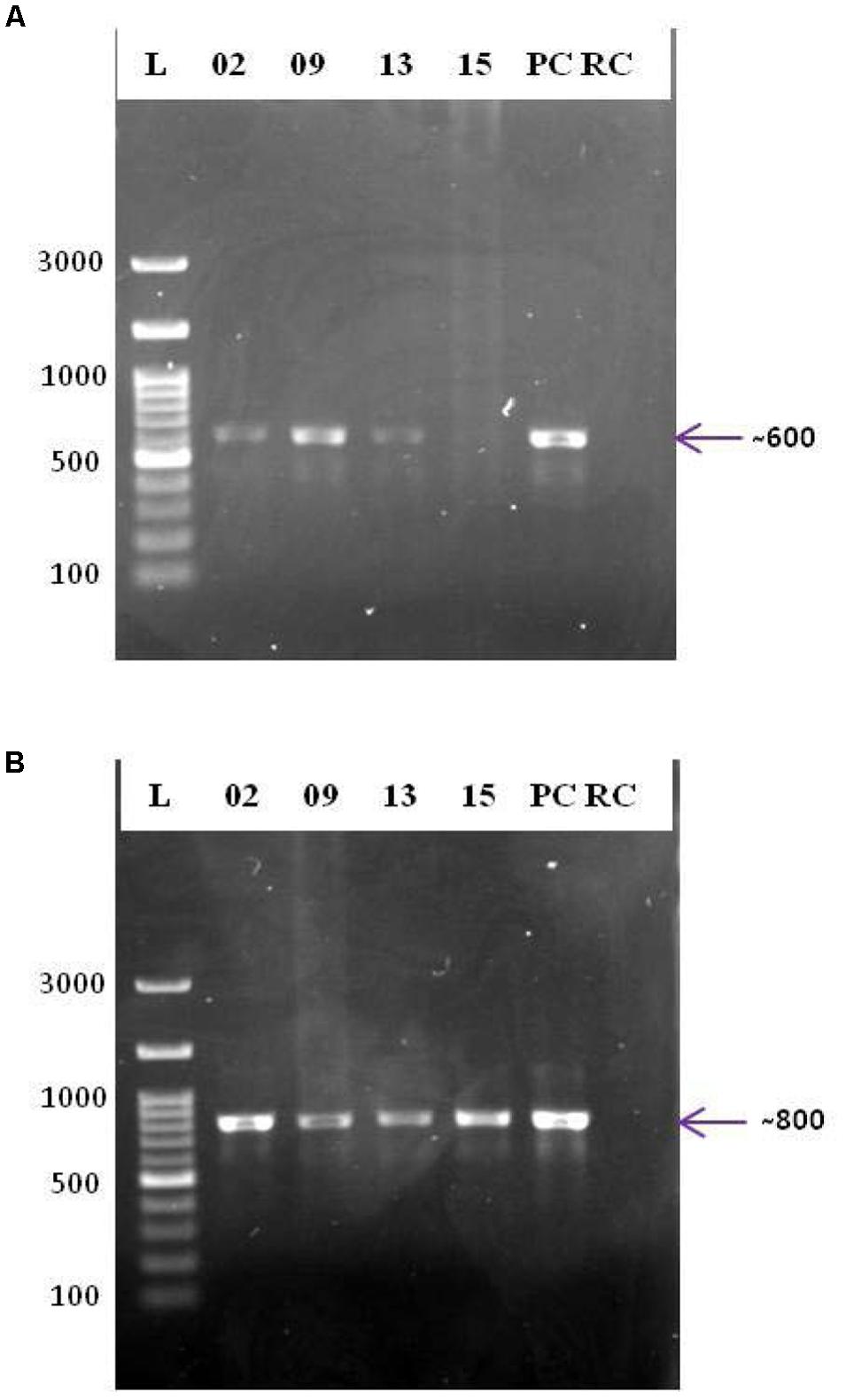
FIGURE 5. PCR based screening for PKS and NRPS systems. Agarose gels (1%, w/v) showing the amplicons of PCR targeted the ketosynthase domain of type II PKS (A) and the adenylation domain of NRPS (B). L: 100 bp DNA ladder; 02, 09, 13, and 15: PCR amplicons of strains SAS02, SAS09, SAS13, and SAS15, respectively; PC, positive control; RC: reaction control.
Secondary metabolites, present in the crude extracts fractionated with an equal volume of ethyl acetate from the fermentation broths of the selected strains were subjected to high resolution LC-MS-Q-TOF analysis (Chromatograms can be found in Supplementary Figure S1) and database search. The compounds produced by the different strains are summarized in Table 5. Notably, the strain SAS02 produced a known microbial bioactive metabolite, related to doxorubicinol, and several putative novel metabolites. Strains SAS09 and SAS13 produced compounds that were related to known compounds like pyrromycin and erythromycin. Similarly, strain SAS15 produced erythromycin derivatives. Further, about 5-12 putatively novel metabolites having molecular features unlike of any other known compounds were also identified. Among them, SAS02 produced 12 putatively novel compounds, followed by SAS15, SAS13, and SAS09 with 9, 6, and 5 novel compounds, respectively (Table 4).
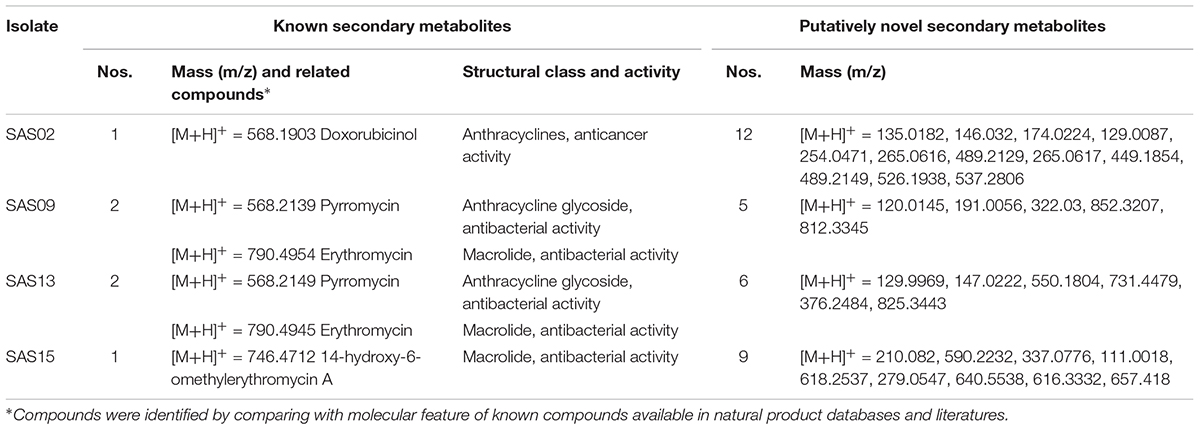
TABLE 5. Known and putatively novel secondary metabolites found from the selected isolates in chemical analysis.
Actinobacteria from poorly explored habitats are an important source of medically relevant secondary metabolites (Fenical and Jensen, 2006; Bull and Stach, 2007; Monciardini et al., 2014; Singh et al., 2016). India is a country well endowed with natural resources, like large forest area, sea coast, and deserts having diverse climatic conditions; however, they are largely unexplored. In the present study, 21 morphologically distinct Actinobacteria were isolated from arid soil samples collected from a previously unexplored location in the Thar Desert. Although a few studies have recently been resulted in isolation of relatively larger number of isolates from desert ecosystem (Arocha-Garza et al., 2017; Mohamed et al., 2017), this study represents the occurrence of diverse culturable Actinobacteria in a relatively small sampling area (128 m2).
Morphological characteristics, notably aerobic growth, chalky and heaped appearance, aerial and substrate mycelia with different colors, and an earthy odor suggested that the isolates are affiliated to genus Streptomyces. These characteristics of Streptomyces are widely considered for their preliminary identification (Taddei et al., 2006).
The Streptomyces are a well-established source of diverse bioactive compounds possessing antimicrobial activity. This agrees well with the current study, which uncovered the antimicrobial activity of 12 different actinobacterial strains. Further, we observed antibacterial activity against MRSA and VRE, implying the need for detailed characterization of the active isolates and their principal components. Streptomycetes from deserts have been previously investigated for the production of antimicrobial metabolites. For instance, a new type of ansamycin and 22 members of macrolactones with antibacterial and antitumor activity were isolated from Streptomyces species that were isolated from the Atacama Desert soils (Rateb et al., 2011). Similarly, novel Abenquines A–D, which demonstrated inhibitory activity against bacteria and dermatophytic fungi, were purified from a Streptomyces strain isolated from the Chilean Atacama Desert (Schulz et al., 2011). The current study further adds to these observations and shows that Thar Desert is a source of actinobacterial strains with a potential for the exploitation of bioactive compounds.
The strains showed potent antimicrobial activity were characterized by 16S rDNA sequence analysis to assign them a particular molecular taxonomic unit. It is interesting to note that three out of four isolates showed dissimilarity to the already reported members of genus Streptomyces. Recent studies have raised the 16S rRNA gene sequence similarity threshold to 99% for Actinobacteria (Stach et al., 2003; Guo et al., 2015). Accordingly, strain SAS02 represents a novel species of the genus Streptomyces. The taxonomic novelty of SAS02 was further supported by the formation of distinct sub-clade in the neighbor-joining phylogenetic tree.
Microbial natural products with interesting biological activities are mainly synthesized by PKS and NRPS systems (Fischbach and Walsh, 2006). The occurrence of genes that encode such biosynthetic systems in a microorganism strongly indicates their biosynthetic potential (Ayuso-Sacido and Genilloud, 2005). In this report, a PCR-based screening method was adopted for the detection of PKS and NRPS systems in streptomycetes. Interestingly, out of the four strains selected, three were found to be positive for both type II PKS and NRPS systems. The presence of PKS and NRPS systems in the selected strains provides evidence for their biosynthetic potential. Since genome-guided screening of isolates is helpful in the discovery of novel drug leads (Heine et al., 2014; Huang et al., 2016), future studies will focus on the genomes of these isolates.
The application of LC-MS in identifying novel metabolites is a well-recognized methodology (Abdelmohsen et al., 2015; Sun et al., 2015). We could obtain only a qualitative estimate of the abundance of bioactive secondary metabolites in the selected isolates. Janso and Carter (2010) employed this approach to uncover the biosynthetic potential of endophytic Actinobacteria. Similarly, an LC-MS based study by Tiwari et al. (2015) suggested that the culturable Actinobacteria of Indian desert regions are a source of bioactive Actinobacteria. This study further revealed the biosynthetic potential of Actinobacteria inhabiting arid environments.
Among the four selected isolates, known antibacterial compounds related to pyrromycin and erythromycin derivatives were detected from the ethyl acetate extract of SAS09, SAS13, and SAS15. In contrast, known antibacterial compounds were not found from SAS02, which indicates the presence of putatively novel antimicrobial compounds. Moreover, strain SAS02 is a novel strain and supports a fact that antimicrobial activity against the highest impact may come from the novel, undescribed strains present in the unexplored environment (Donadio et al., 2007; Guo et al., 2015; Parrot et al., 2015). Overall, production of 5–12 putatively novel compounds was detected from the selected strains and it is attesting their importance in secondary metabolites production. It is important to note that these compounds were identified from the spent broth of selected isolates by single-solvent (ethyl acetate) extraction, revealing the abundance of secondary metabolites and their complete chemical space may be further studied under a wide range of cultivation conditions, culture media, and secondary metabolites extraction methods. Since the comprehensive exploration of Actinobacteria is indispensable in the discovery of bioactive compounds (Chater and Chandra, 2006; Kersten et al., 2011; Harrison and Studholme, 2014), further efforts are necessary for the structural characterization of the metabolites produced by SAS02 under different laboratory conditions, and whole genome sequencing is required to explore its complete potential.
The research on the microbial natural product is expected to move ahead in a new direction, aided by the innovations in Next Generation Sequencing and spectroscopic methods. This study reports an investigation of the biosynthetic potential of Actinobacteria from the soil samples collected from the semi-arid region of India. The relatively small number of isolates screened here demonstrates their novelty, significance, and biosynthetic potential. Actinobacteria thus identified seem to be a promising source of new and interesting natural products that will be further explored for their biotechnological applications.
The work was conceived and designed by SJ, PJ, EM, and GS. Experiments were done by MM, KS, and TT. Data analysis and verification were done by RA, NS, MM, and KS. The manuscript was drafted by KS, MM, and PJ. The manuscript was approved by all the authors.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
PJ thanks Council of Scientific and Industrial Research (Government of India) for the continuous support to the research on Actinobacteria. The authors are grateful to Sophisticated Analytical Instrumentation Facility (SAIF) of IIT Bombay for HR-LCMS analysis.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.00687/full#supplementary-material
Abdelmohsen, U. R., Grkovic, T., Balasubramanian, S., Kamel, M. S., Quinn, R. J., and Hentschel, U. (2015). Elicitation of secondary metabolism in actinomycetes. Biotechnol. Adv. 33, 798–811. doi: 10.1016/j.biotechadv.2015.06.003
Albert, C. A., and Malaquias de Querioz, V. M. (1963). Streptomyces tuirus nov. sp., productor do antibiotico tuoromicina. Rev. Inst. Antibiot. 5, 43–51.
Altschul, S. F., Gish, W., Miller, W., Myers, E. W., and Lipman, D. J. (1990). Basic local alignment search tool. J. Mol. Biol. 215, 403–410. doi: 10.1016/S0022-2836(05)80360-2
Arocha-Garza, H. F., Castillo, R. C.-D., Eguiarte, L. E., Souza, V., and De la Torre-Zavala, S. (2017). High diversity and suggested endemicity of culturable Actinobacteria in an extremely oligotrophic desert oasis. PeerJ 5:e3247. doi: 10.7717/peerj.3247
Ayuso-Sacido, A., and Genilloud, O. (2005). New PCR primers for the screening of NRPS and PKS-I systems in actinomycetes: detection and distribution of these biosynthetic gene sequences in major taxonomic groups. Microb. Ecol. 49, 10–24. doi: 10.1007/s00248-004-0249-6
Bachmann, B. O., Van Lanen, S. G., and Baltz, R. H. (2014). Microbial genome mining for accelerated natural products discovery: is a renaissance in the making? J. Ind. Microbiol. Biotechnol. 41, 175–784. doi: 10.1007/s10295-013-1389-9
Becerril-Espinosa, A., Freel, K. C., Jensen, P. R., and Soria-Mercado, I. E. (2013). Marine Actinobacteria from the Gulf of California: diversity, abundance and secondary metabolite biosynthetic potential. Antonie Van Leeuwenhoek 103, 809–819. doi: 10.1007/s10482-012-9863-3
Bull, A. T., and Stach, J. E. M. (2007). Marine Actinobacteria: new opportunities for natural product search and discovery. Trends Microbiol. 11, 491–499. doi: 10.1016/j.tim.2007.10.004
Chater, K. F., and Chandra, G. (2006). The evolution of development in Streptomyces analysed by genome comparisons. FEMS Microbiol. Rev. 30, 651–672. doi: 10.1111/j.1574-6976.2006.00033.x
Cheng, C., MacIntyre, L., Abdelmohsen, U. R., Horn, H., Polymenakou, P. N., Edrada-Ebel, R., et al. (2015). Biodiversity, anti-trypanosomal activity screening, and metabolomic profiling of actinomycetes isolated from Mediterranean sponges. PLoS One 10:e0138528. doi: 10.1371/journal.pone.0138528
Cole, J. R., Wang, Q., Fish, J. A., Chai, B., McGarrell, D. M., Sun, Y., et al. (2014). Ribosomal database project: data and tools for high throughput rRNA analysis. Nucleic Acids Res. 42, D633–D642. doi: 10.1093/nar/gkt1244
Derewacz, D. K., Covington, B. C., McLean, J. A., and Bachmann, B. O. (2015). Mapping microbial response metabolomes for induced natural product discovery. ACS Chem. Biol. 10, 1998–2006. doi: 10.1021/acschembio.5b00001
Dhakal, D., Pokhrel, A. R., Shrestha, B., and Sohng, J. K. (2017). Marine rare Actinobacteria: isolation, characterization, and strategies for harnessing bioactive compounds. Front. Microbiol. 8:1106. doi: 10.3389/fmicb.2017.01106
Donadio, S., Brandi, L., Monciardini, P., Sosio, M., and Gualerzi, C. O. (2007). Novel assays and novel strains – promising routes to new antibiotics? Expert Opin. Drug Discov. 2, 789–798. doi: 10.1517/17460441.2.6.789
Fenical, W., and Jensen, P. R. (2006). Developing a new resource for drug discovery: marine actinomycete bacteria. Nat. Chem. Biol. 2, 666–673. doi: 10.1038/nchembio841
Fischbach, M. A., and Walsh, C. T. (2006). Assembly-line enzymology for polyketide and nonribosomal peptide antibiotics: logic, machinery, and mechanisms. Chem. Rev. 106, 3468–3496. doi: 10.1021/cr0503097
Floros, D. J., Jensen, P. R., Dorrestein, P. C., and Koyama, N. (2016). A metabolomics guided exploration of marine natural product chemical space. Metabolomics 12:145. doi: 10.1007/s11306-016-1087-5
Forner, D., Berrué, F., Correa, H., Duncan, K., and Kerr, R. G. (2013). Chemical dereplication of marine actinomycetes by liquid chromatography-high resolution mass spectrometry profiling and statistical analysis. Anal. Chim. Acta 805, 70–79. doi: 10.1016/j.aca.2013.10.029
Gause, G. F., Preobrazhenskaya, T. P., Sveshnikova, M. A., Terekhova, L. P., and Maximova, T. S. (1983). A Guide for the Determination of Actinomycetes. Genera Streptomyces, Streptoverticillium, and Chainia. Moscow: Nauka.
Gomez-Escribano, J. P., Alt, S., and Bibb, M. J. (2016). Next generation sequencing of Actinobacteria for the discovery of novel natural products. Mar. Drugs 14:E78. doi: 10.3390/md14040078
Guo, X., Liu, N., Li, X., Ding, Y., Shang, F., Gao, Y., et al. (2015). Red soils harbor diverse culturable actinomycetes that are promising sources of novel secondary metabolites. Appl. Environ. Microbiol. 81, 3086–3103. doi: 10.1128/AEM.03859-14
Harrison, J., and Studholme, D. J. (2014). Recently published Streptomyces genome sequences. Microb. Biotechnol. 7, 373–380. doi: 10.1111/1751-7915.12143
Heine, D., Martin, K., and Hertweck, C. (2014). Genomics-guided discovery of endophenazines from Kitasatospora sp. HKI 714. J. Nat. Prod. 77, 1083–1087. doi: 10.1021/np400915p
Hozzein, W. N., Ali, M. I. A., and Rabie, W. (2008). A new preferential medium for enumeration and isolation of desert actinomycetes. World J. Microbiol. Biotechnol. 24, 1547–1552. doi: 10.1007/s11274-007-9641-y
Huang, C., Leung, R. K.-K., Guo, M., Tuo, L., Guo, L., Yew, W. W., et al. (2016). Genome-guided investigation of antibiotic substances produced by Allosalinactinospora lopnorensis CA15-2T from Lop Nor region, China. Sci. Rep. 6:20667. doi: 10.1038/srep20667
Janso, J. E., and Carter, G. T. (2010). Biosynthetic potential of phylogenetically unique endophytic actinomycetes from tropical plants. Appl. Environ. Microbiol. 76, 4377–4386. doi: 10.1128/AEM.02959-09
Jose, P. A., and Jebakumar, S. R. D. (2013). Diverse actinomycetes from Indian coastal solar salterns–a resource for antimicrobial screening. J. Pure Appl. Microbiol. 7, 2569–2575.
Jose, P. A., and Jebakumar, S. R. D. (2014). Unexplored hypersaline habitats are sources of novel actinomycetes. Front. Microbiol. 5:242. doi: 10.3389/fmicb.2014.00242
Jose, P. A., and Jha, B. (2017). Intertidal marine sediment harbours Actinobacteria with promising bioactive and biosynthetic potential. Sci. Rep. 7:10041. doi: 10.1038/s41598-017-09672-6
Jose, P. A., Santhi, V. S., and Jebakumar, S. R. D. (2011). Phylogenetic-affiliation, antimicrobial potential and PKS gene sequence analysis of moderately halophilic Streptomyces sp. inhabiting an Indian saltpan. J. Basic Microbiol. 51, 348–356. doi: 10.1002/jobm.201000253
Jose, P. A., Sivakala, K. K., and Jebakumar, S. R. D. (2013). Formulation and statistical optimization of culture medium for improved production of antimicrobial compound by Streptomyces sp. JAJ06. Int. J. Microbiol. 2013:526260. doi: 10.1155/2013/526260
Ju, K. S., Gao, J., Doroghazi, J. R., Wang, K. K., Thibodeaux, C. J., Li, S., et al. (2015). Discovery of phosphonic acid natural products by mining the genomes of 10,000 actinomycetes. Proc. Natl. Acad. Sci. U.S.A. 112, 12175–12180. doi: 10.1073/pnas.1500873112
Kersten, R. D., Yang, Y.-L., Xu, Y., Cimermancic, P., Nam, S.-J., Fenical, W., et al. (2011). A mass spectrometry–guided genome mining approach for natural product peptidogenomics. Nat. Chem. Biol. 7, 794–802. doi: 10.1038/nchembio.684
Kim, O. S., Cho, Y. J., Lee, K., Yoon, S. H., Kim, M., Na, H., et al. (2012). Introducing EzTaxon-e: a prokaryotic 16S rRNA gene sequence database with phylotypes that represent uncultured species. Int. J. Syst. Evol. Microbiol. 62, 716–721. doi: 10.1099/ijs.0.038075-0
Lemos, M. L., Toranzo, A. E., and Barja, J. L. (1985). Antibiotic activity of epiphytic bacteria isolated from intertidal seaweeds. Microbiol. Ecol. 11, 149–163. doi: 10.1007/BF02010487
Mahmoud, H. M., and Kalendar, A. A. (2016). Coral-associated Actinobacteria: diversity, abundance, and biotechnological potentials. Front. Microbiol. 7:204. doi: 10.3389/fmicb.2016.00204
Masand, M., Jose, P. A., Menghani, E., and Jebakumar, S. R. D. (2015). Continuing hunt for endophytic actinomycetes as a source of novel biologically active metabolites. World J. Microbiol. Biotechnol. 31, 1863–1875. doi: 10.1007/s11274-015-1950-y
Metsa-Ketela, M., Salo, V., Halo, L., Hautala, A., and Hakala, J. (1999). An efficient approach for screening minimal PKS genes from Streptomyces. FEMS Microbiol. Lett. 180, 1–6. doi: 10.1016/S0378-1097(99)00453-X
Mincer, T. J., Jensen, P. R., Kauffman, C. A., and Fenical, W. (2002). Widespread and persistent populations of a major new marine actinomycete taxon in ocean sediments. Appl. Environ. Microbiol. 68, 5005–5011. doi: 10.1128/AEM.68.10.5005-5011.2002
Mohamed, H., Miloud, B., Zohra, F., García-Arenzana, J. M., Veloso, A., and Rodríguez-Couto, S. (2017). Isolation and characterization of Actinobacteria from Algerian Sahara soils with antimicrobial activities. Int. J. Mol. Cell. Med. 6, 109–120.
Mohammadipanah, F., Hamedi, J., Spröer, C., Rohde, M., Montero-Calasanz, M. D., and Klenk, H. P. (2014). Streptomyces zagrosensis sp. nov., isolated from soil. Int. J. Syst. Evol. Microbiol. 64, 3434–3440. doi: 10.1099/ijs.0.064527-0
Mohammadipanah, F., and Wink, J. (2016). Actinobacteria from arid and desert habitats: diversity and biological activity. Front. Microbiol. 6:1541. doi: 10.3389/fmicb.2015.01541
Monciardini, P., Iorio, M., Maffioli, S., Sosio, M., and Donadio, S. (2014). Discovering new bioactive molecules from microbial sources. Microb. Biotechnol. 7, 209–220. doi: 10.1111/1751-7915.12123
Okoro, C. K., Brown, R., Jones, A. L., Andrews, B. A., Asenjo, J. A., Goodfellow, M., et al. (2009). Diversity of culturable actinomycetes in hyper-arid soils of the Atacama Desert, Chile. Antonie Van Leeuwenhoek 95, 121–133. doi: 10.1007/s10482-008-9295-2
Parrot, D., Antony-Babu, S., Intertaglia, L., Grube, M., Tomasi, S., and Suzuki, M. T. (2015). Littoral lichens as a novel source of potentially bioactive Actinobacteria. Sci. Rep. 5:15839. doi: 10.1038/srep15839
Peraud, O., Biggs, J. S., Hughen, R. W., Light, A. R., Concepcion, G. P., and Concepcion, G. P. (2009). Microhabitats within venomous cone snails contain diverse Actinobacteria. Appl. Environ. Microbiol. 75, 6820–6826. doi: 10.1128/AEM.01238-09
Poulsen, M., Oh, D. C., Clardy, J., and Currie, C. R. (2011). Chemical analyses of wasp-associated Streptomyces bacteria reveal a prolific potential for natural products discovery. PLoS One 6:e16763. doi: 10.1371/journal.pone.0016763
Procópio, R. E., Silva, I. R., Martins, M. K., Azevedo, J. L., and Araújo, J. M. (2012). Antibiotics produced by Streptomyces. Braz. J. Infect. Dis. 16, 466–471. doi: 10.1016/j.bjid.2012.08.014
Rateb, M. E., Houssen, W. E., Harrison, W. T. A., Deng, H., Okoro, C. K., Asenjo, J. A., et al. (2011). Diverse metabolic profiles of a Streptomyces strain isolated from a hyper-arid environment. J. Nat. Prod. 74, 1965–1971. doi: 10.1021/np200470u
Schulz, D., Beese, P., Ohlendorf, B., Erhard, A., Zinecker, H., Dorador, C., et al. (2011). Abenquines A–D: aminoquinone derivatives produced by Streptomyces sp. strain DB634. J. Antibiot. 64, 763–768. doi: 10.1038/ja.2011.87
Sengupta, S., Pramanik, A., Ghosh, A., and Bhattacharyya, M. (2015). Antimicrobial activities of actinomycetes isolated from unexplored regions of Sundarbans mangrove ecosystem. BMC Microbiol. 15:170. doi: 10.1186/s12866-015-0495-4
Singh, V., Haque, S., Singh, H., Verma, J., Vibha, K., Singh, R., et al. (2016). Isolation, screening, and identification of novel isolates of actinomycetes from India for antimicrobial applications. Front. Microbiol. 7:1921. doi: 10.3389/fmicb.2016.01921
Stach, J. E., Maldonado, L. A., Masson, D. G., Ward, A. C., Goodfellow, M., and Bull, A. T. (2003). Statistical approaches for estimating actinobacterial diversity in marine sediments. Appl. Environ. Microbiol. 69, 6189–6200. doi: 10.1128/AEM.69.10.6189-6200.2003
Subramani, R., and Aalbersberg, W. (2012). Marine actinomycetes: an ongoing source of novel bioactive metabolites. Microbiol. Res. 167, 571–580. doi: 10.1016/j.micres.2012.06.005
Sun, W., Zhang, F., He, L., Karthik, L., and Li, Z. (2015). Actinomycetes from the South China Sea sponges: isolation, diversity, and potential for aromatic polyketides discovery. Front. Microbiol. 6:1048. doi: 10.3389/fmicb.2015.01048
Suzuki, M. (1957). Studies on an antitumor substance, gancidin. Mycological study on the strain AAK-84 and production, purification of active fractions. J. Chiba Med. Soc. 33, 535–542.
Taddei, A., Rodríguez, M. J., Márquez-Vilchez, E., and Castelli, C. (2006). Isolation and identification of Streptomyces spp. from Venezuelan soils: morphological and biochemical studies. I. Microbiol. Res. 161, 222–231. doi: 10.1016/j.micres.2005.08.004
Takahashi, Y., Matsumoto, A., Seino, A., Iwai, Y., and Omura, S. (1996). Rare actinomycetes isolated from desert soils. Actinomycetologica 10, 91–97. doi: 10.3209/saj.10_91
Tamura, K., Peterson, D., Peterson, N., Stecher, G., Nei, M., and Kumar, S. (2011). MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 28, 2731–2739. doi: 10.1093/molbev/msr121
Tiwari, K., Upadhyay, D. J., Mösker, E., Süssmuth, R., and Gupta, R. K. (2015). Culturable bioactive actinomycetes from the Great Indian Thar Desert. Ann. Microbiol. 65, 1901–1914. doi: 10.1007/s13213-014-1028-3
Keywords: arid, desert, Streptomyces, metabolomics, secondary metabolome, antimicrobial activity
Citation: Masand M, Sivakala KK, Menghani E, Thinesh T, Anandham R, Sharma G, Sivakumar N, Jebakumar SRD and Jose PA (2018) Biosynthetic Potential of Bioactive Streptomycetes Isolated From Arid Region of the Thar Desert, Rajasthan (India). Front. Microbiol. 9:687. doi: 10.3389/fmicb.2018.00687
Received: 19 May 2017; Accepted: 23 March 2018;
Published: 18 April 2018.
Edited by:
Maria Tereza dos Santos Correia, Federal Rural University of Pernambuco, BrazilReviewed by:
Osmar Nascimento Silva, Universidade Católica Dom Bosco, BrazilCopyright © 2018 Masand, Sivakala, Menghani, Thinesh, Anandham, Sharma, Sivakumar, Jebakumar and Jose. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Polpass Arul Jose, YXJ1bG1rdUBnbWFpbC5jb20=
†Present address: Polpass Arul Jose, Central Salt and Marine Chemicals Research Institute, Bhavnagar, India
‡These authors have contributed equally to this work.
§Deceased
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.