 Frédérique Pasquali1*
Frédérique Pasquali1* Federica Palma1
Federica Palma1 Laurent Guillier2
Laurent Guillier2 Alex Lucchi1
Alex Lucchi1 Alessandra De Cesare1
Alessandra De Cesare1 Gerardo Manfreda1
Gerardo Manfreda1- 1Dipartimento di Scienze e Tecnologie Agro-Alimentari, Alma Mater Studiorum – Università di Bologna, Bologna, Italy
- 2Laboratoire de Sécurité des Aliments, Agence Nationale de Sécurité Sanitaire de l’Alimentation, de l’Environnement et du Travail, Maisons-Alfort, France
Listeria monocytogenes is a foodborne pathogen adapted to survive and persist in multiple environments. Following two previous studies on prevalence and virulence of L. monocytogenes ST121 and ST14 repeatedly collected in a the same rabbit-meat processing plant, the research questions of the present study were to: (1) assess persistence of L. monocytogenes isolates from the rabbit-plant; (2) select genes associated to physiological adaptation to the food-processing environment; (3) compare presence/absence/truncation of these genes in newly sequenced and publicly available ST121 and ST14 genomes. A total of 273 draft genomes including ST121 and ST14 newly sequenced and publicly available draft genomes were analyzed. Whole-genome Single Nucleotide Polymorfism (wgSNP) analysis was performed separately on the assemblies of ST121 and ST14 draft genomes. SNPs alignments were used to infer phylogeny. A dataset of L. monocytogenes ecophysiology genes was built based on a comprehensive literature review. The 94 selected genes were screened on the assemblies of all ST121 and ST14 draft genomes. Significant gene enrichments were evaluated by statistical analyses. A persistent ST14 clone, including 23 out of 27 newly sequenced genomes, was circulating in the rabbit-meat plant along with two not persistent clones. A significant enrichment was observed in ST121 genomes concerning stress survival islet 2 (SSI-2) (alkaline and oxidative stress), qacH gene (resistance to benzalkonium chloride), cadA1C gene cassette (resistance to 70 mg/l of cadmium chloride) and a truncated version of actA gene (biofilm formation). Conversely, ST14 draft genomes were enriched with a full-length version of actA gene along with the Listeria Genomic Island 2 (LGI 2) including the ars operon (arsenic resistance) and the cadA4C gene cassette (resistance to 35 mg/l of cadmium chloride). Phenotypic tests confirmed ST121 as a weak biofilm producer in comparison to ST14. In conclusion, ST121 carried the qacH gene and was phenotypically resistant to quaternary ammonium compounds. This property might contribute to the high prevalence of ST121 in food processing plants. ST14 showed greater ability to form biofilms, which might contribute to the occasional colonization and persistence on harborage sites where sanitizing procedures are difficult to display.
Introduction
Listeria monocytogenes is a foodborne pathogen adapted to survive in a variety of environmental locations including soil, groundwater, decaying vegetation (Gray et al., 2006). In food-processing plants, L. monocytogenes has been repeatedly isolated from both food and the environment. Based on different molecular typing methods, isolates sharing the same profile have been collected over months or years in fish, meat, dairy and vegetable processing plants (Leong et al., 2014; Stasiewicz et al., 2015; Véghová et al., 2017). Strains repeatedly isolated over time in the same plant are considered as persistent. Unfortunately, there is not yet an agreement on specific issues related to the definition of persistence. In particular, the number of times of re-isolation, the sources as well as the period of isolation are not yet uniquely defined (Ferreira et al., 2014).
Persistent strains of high virulence are of major concern since they commonly colonize harborage sites difficult to clean or to reach by sanitizing procedures. These strains typically contaminate different lots of food during several months of production and have been described as responsible of outbreaks including few to hundreds cases spread in time and geographical areas (Tompkin, 2002).
The reasons why L. monocytogenes persists in food processing plants is still on debate. One strain can colonize harborage sites by chance (Carpentier and Cerf, 2011). In this view, the persistence of the strain was supposed as more related to characteristics of the environment rather than of the strain itself (Carpentier and Cerf, 2011). Authors failed to identify associations among persistence and particular genes/features of the strain (Ferreira et al., 2011; Stasiewicz et al., 2015). Nevertheless, a differential distribution of specific subtypes in food environments and clinical samples was observed, suggesting that L. monocytogenes strains might harbor unique genotypic and phenotypic features facilitating survival and growth and ultimately spread to humans. In foods and food processing environments, L. monocytogenes of lineage II and serotype 1/2a has been more frequently collected than lineage I (Orsi et al., 2011). Within serotype 1/2a, Clonal Complex (CC) 121 was the most prevalent clone (17%) (Maury et al., 2016). In particular within CC 121, food source was overrepresented in comparison to clinical one (92.9% vs. 7.0%) (Maury et al., 2016). Human strains belonging to CC121 showed prevalence of 9.5 and 2.3% among 116 strains of the Institute Pasteur L. monocytogenes database (Moura et al., 2016) and 262 sporadic cases collected throughout Europe, respectively (Nielsen et al., 2017). Low frequency of CC121 in clinical samples was associated to an attenuated virulence of this subtype, which often carries Premature Stop Codon Mutations in the virulence gene marker inlA. (Olier et al., 2002, 2003; Van Stelten et al., 2010; Beier and Bertilsson, 2013; Palma et al., 2017).
The CC14 is another clonal complex of serotype 1/2a. Compared to CC121, CC14 was seldom isolated in foods in France (1.4%) (Maury et al., 2016). Similarly, low detection values were reported in 19 meat processing plants located in Northern Italy (5.7% over 69 tested isolates) in comparison to CC121 (23%) (Morganti et al., 2016). Besides a low prevalence in foods, CC14 was described to gather a high percentage of isolates of clinical source (29.5%) (Maury et al., 2016). Within CC14, hypervirulent strains were described. In particular, a Multi Locus Sequence Type (ST) 14 strain was associated to a case of invasive listeriosis. Further molecular analyses revealed that this strain belongs to epidemic clone (EC) III (corresponding to Multi-Virulence Locus Type (VT) 1) previously linked to a sporadic case as well as a multi-state outbreak occurred in United States in 1988 and 2000, respectively (Kathariou, 2002; Mammina et al., 2013).
Physiological adaptation (ecophysiology) to environmental stresses including resistance to antimicrobials, heavy metals and quaternary ammonium compounds (QAC) as well as adaptation to cold, salt, acid, oxidative stresses, desiccation and ability of biofilm formation are often described in L. monocytogenes isolated from food processing plants (Lebrun et al., 1994; Kathariou, 2002; Srinivasan et al., 2005; Mullapudi et al., 2008; Ryan et al., 2010; Hein et al., 2011; Ratani et al., 2012; Müller et al., 2013; Ortiz et al., 2015; Kovacevic et al., 2016; Xu et al., 2016; Harter et al., 2017; Hingston et al., 2017). Studies on ST121 strains have been recently performed aiming at identifying genes associated to persistence and/or adaptation to food-processing environmental stresses. In particular, deletions of lmo02774-lmo2776 were described along with no significant association of genes (Holch et al., 2013; Knudsen et al., 2017). In one study, ST121 isolates were found to harbor the qacH carrying transposon Tn1688 (QAC resistance) (Ortiz et al., 2015). No similar studies are available on ST14.
Analyses based on Whole Genome Sequencing (WGS) have recently revealed an unprecedented potential for multiple investigations as a one-serve-all approach. WGS-based typing showed valuable potential in pathogen fingerprinting. In particular, a high discrimination power has been achieved by single nucleotide polymorphisms (SNPs) analysis. This is particularly relevant to discriminate strains showing high genetic similarity as determined by same Pulse Field Gel Electrophoresis (PFGE) or 7-loci MLST, as it is the case during a persistent event (Ferreira et al., 2014).
Based on WGS-data, studies on presence/absence/truncation of a wide number of genes can be performed at once. Genes described as strongly associated to a specific phenotype can be selected through a literature search, and traced on sequenced genomes for prediction of that specific phenotype. At present, freely available tools unable a quick screening of genomes for thousands of genes selected for their association to virulence or antimicrobial resistance (McArthur et al., 2013; Zankari et al., 2012). Genes, useful to predict phenotypic traits associated to L. monocytogenes physiological adaptation to environmental conditions, have not been collected so far in a unique dataset.
In a previous study on prevalence of L. monocytogenes in four Italian rabbit meat processing plants, isolates indistinguishable by 7-loci MLST, Multi Locus Variable number tandem repeats Analysis (MLVA) type and ApaI-PFGE were repeatedly collected over time. Isolates were sampled from carcasses, meat cuts, meat products and the meat-processing environment (De Cesare et al., 2017). The dataset of this study was used to investigate further the potential virulence of specific subtypes. For this purpose, in a second study a specific focus was addressed on two subtypes gathering isolates of 7-loci MLST ST121 and ST14 collected over 1 year in the same processing plant (Palma et al., 2017). In this latter study, ST14 isolates showed higher virulence potential than ST121 as suggested by in silico virulotyping (Palma et al., 2017). In particular, all ST14 isolates belonged to VT107, which differed from ECIII (VT1) for only four nucleotides (Murugesan et al., 2015). Moreover, all ST121 and no ST14 genomes carried a truncated version of the actA gene and an inlA gene with a premature stop codon of type 6, both described as associated to attenuated virulence (Smith and Portnoy, 1997; Olier et al., 2002, 2003; Van Stelten et al., 2010).
In the present study, the ST121 and ST14 L. monocytogenes isolates, collected over 1 year on the same rabbit meat processing plant, were studied with a specific focus on persistence and physiological adaptation to food-processing environmental stresses. In particular, the research questions were: (1) assess persistence of L. monocytogenes isolates from the rabbit-plant; (2) select genes associated to physiological adaptation to the food-processing environment; (3) compare presence/absence/truncation of selected genes in newly sequenced and publicly available ST121 and ST14 genomes. A particular focus has been placed on the evaluation of putative gene enrichment in the two subtypes as well as identification of putative markers of ecophysiology associated to survival and growth of rare subtype ST14 of L. monocytogenes.
Materials and Methods
Listeria monocytogenes Isolates From Italian Meat Processing Plants, Sequencing and de Novo Assembly
Listeria monocytogenes isolates included in the present study belong to a subset of isolates collected from November 2005 to November 2006 within a previous study on prevalence of L. monocytogenes in rabbit meat-processing plants (De Cesare et al., 2017). This subset included isolates belonging to two subtypes and collected from plant A. Isolates were considered as potentially persistent when they belonged to the same subtype and they were collected more than six times over a period of more than 6 months from different sources (rabbit carcasses, rabbit meat cuts, rabbit meat products, and the food processing environment). Each subtype gathered isolates indistinguishable by serotype, ApaI-PFGE, 7-loci MLST, MLVA and automated ribotype (De Cesare et al., 2017). The genomes of this subset of L. monocytogenes isolates were sequenced and de novo assembled in a previous study (Palma et al., 2017). Briefly, the genomes of 33 L. monocytogenes isolates belonging to ST121 (6) and ST14 (27) were previously sequenced by MiSeq (Illumina) platform. Paired-end reads were de novo assembled using the INNUca pipeline1, which consists of several modules (e.g., Trimmomatic, SPAdes, Pilon) and QA/QC steps. Information on quality parameters of de novo assemblies were included in the previous study (Palma et al., 2017).
Publicly Available L. monocytogenes Genomes Included in the Present Study
A selection of draft genomes from ENA was carried out in order to: (1) explore the genetic distances of newly sequenced L. monocytogenes genomes in comparison to publicly available genomes; (2) study ST-related genetic markers associated to ecophysiology. In particular, 196 ST121 and 44 ST14 publicly available draft genomes were included. These genomes were from strains isolated from both food processing environments and humans. Moreover, they were widely distributed over time and geographical locations (Supplementary Table S2).
Whole-Genome SNP Analysis
Single nucleotide polymorphisms (SNPs) calling was performed using the Snippy v2.6 pipeline2. De novo assemblies of ST121 and ST14 genomes were analyzed separately. A newly generated draft genome for each L. monocytogenes ST was chosen as reference genome: LSALM51 for ST121 and LSALM1 for ST14. After removal of Illumina Nextera adapters and low-quality sequences (Phred scores of <10), the de novo assemblies were mapped to the reference genome with the Burrows-Wheeler Aligner (BWA) v0.7.12 using default parameters (Li and Durbin, 2009). After mapping, average depths were determined with SAMtools v1.3 (Li et al., 2009). Variants were called using Freebayes v0.9.20 (Garrison and Marth, 2012) with the following parameters: minimum base quality of 20, minimum read coverage of 10X, and 90% read concordance at a locus. Snippy was used to pool all SNP positions called in at least one isolate and investigate all isolates. Alignments of all SNPs were produced in Snippy and used to infer a high-resolution phylogeny. A maximum likelihood (ML) tree was constructed using the PhyML v2.4.4 program to analyze the SNP differences between isolates and FigTree v1.4.3 software3 was used to visualize the tree rooting at midpoint for each ST. Draft genomes were tentatively considered as belonging to the same persistent clone if the following criteria were fulfilled: (1) the difference between draft genomes and the reference genome was equal or lower than 25 SNPs; (2) draft genomes belonged to isolates collected from different origins (food and the processing environment) for at least six times during a time frame of at least 6 months. The cut-off of 25 SNPs was used as previously suggested (Nielsen et al., 2017).
Dataset of Putative Gene Markers of Ecophysiology in L. monocytogenes
An extensive literature review was conducted on PubMed4 using following keywords: “antimicrobials,” “QAC,” “heavy metals,” “bacteriocins,” “cold,” “high salt concentration,” “low pH,” “desiccation,” “blue-light,” “biofilm,” “L. Monocytogenes,” and “genes.” At the time of analysis (October 2017), the system retrieved overall 664 peer-reviewed published papers. The abstract of each of these papers was read with the purpose to identify genes associated to the response of L. monocytogenes to each stress. Around 100 papers were selected based on the abstract. These papers were thoroughly read in order to identify and specifically select genes with a strong association to the related phenotype. For this purpose, only genes confirmed by insertional mutagenesis or mutant selection experiments were included (Supplementary Table S1). All GenBank accession numbers and related sequences reported in published papers were checked by nucleotide BLAST5 and CLUSTAL Omega6 for alignment. For each gene, the following informations were reported: name of the gene, locus tag, annotation, main function, localization, reference paper, GenBank accession number and direct link to the web page of the sequence in NCBI7 (Supplementary Table S1).
Screening of Genes of Ecophysiology
A multifasta file was generated with the sequences of all selected genes (Supplementary Table S1). Nucleotide BLAST was run locally using ABRricate pipeline8 on 202 ST121 as well as 71 ST14 draft genomes (Supplementary Table S2) with the mutifasta file as gene database. Based on the output matrix of gene presence/absence, a heatmap was built using morpheus software9. Morever, results were analyzed for statistically significant differences at 95% confidence by t-test.
Antimicrobial Susceptibility
For phenotypic confirmation, newly sequenced strains carrying antimicrobial resistant genes (ampC, tetA) were tested for susceptibility against ampicillin and tetracycline by disk diffusion method (Clinical and Laboratory Standards Institute [CLSI], 2010; Jamali et al., 2015). The assay was performed on Mueller Hinton Agar plates supplemented with 5% defibrinated sheep blood (Thermo Scientific, Milan, Italy). Ampicillin (10 μg) and tetracycline (30 μg), were applied as antibiotic agents following Clinical and Laboratory Standards Institute [CLSI] recommendations for fastidious organisms (document M45-A2).
Cadmium Chloride and Benzalkonium Chloride Susceptibility
Determination of cadmium chloride and benzalkonium chloride resistance was performed as previously described (Mullapudi et al., 2008). Briefly, a single colony from a blood agar plate culture was suspended in 100 μl of tryptic soy broth (Thermo Fisher Scientific, Milan, Italy). Three microliters of the suspension were spotted in duplicate onto: (1) Mueller Hinton Agar Cation adjusted (MHBII) containing 2% defibrinated sheep blood (Thermo Fisher Scientific) (control); (2) MHBII (Thermo Fisher Scientific) containing 2% defibrinated sheep blood (Thermo Fisher Scientific) and 70 mg/l of cadmium chloride anhydrous (Sigma, Milan, Italy); (3) MHBII (Thermo Fisher Scientific) containing 2% defibrinated sheep blood (Thermo Fisher Scientific) and 35 mg/l of cadmium chloride anhydrous (Sigma, Milan, Italy); (4) Mueller Hinton Agar (MHB) (Thermo Fisher Scientific) containing 2% defibrinated sheep blood (Thermo Fisher Scientific) and 10 mg/l benzalkonium chloride (Sigma). Positive and negative control strains were included. All plates were incubated at 37°C for 48 h.
Crystal Violet Staining Assay
In order to test the ability of biofilm formation, representative isolates of identified clones of ST121 and ST14 isolates of L. monocytogenes were submitted to the crystal violet staining assay as previously described (Stepanović et al., 2004). Briefly, 20 μl of an overnight bacterial culture were added to 230 μl of Brain Heart Infusion broth (BHI, Thermo Scientific, Milan, Italy) into each well of a sterile 96-well not tissue treated polystyrene microplate (Sarstedt, Milan Italy). Plates were incubated at 35°C for 24 h. The content of each plate was discarded and 300 μl of sterile distilled water were added to each well. This washing step was repeated three times. Adherent bacteria were fixed with 250 μl of methanol per well. After 15 min, methanol was discarded and plates air-dried overnight. Biofilms were stained with 250 μl per well of Crystal violet (Gram-color staining set for microscopy; Merck) for 5 min. Excess stain was rinsed off and microplates air-dried. Then, attached bacteria were solubilized with 250 μl of 33% (v/v) glacial acetic acid per well. Finally, the optical density (OD) of each well was measured at 570 nm using Infinite® F50 Absorbance Microplate Reader (Tecan Group Ltd, Männedorf, Switzerland). According to Stepanović et al. (2004), based on the OD produced by bacterial films, strains were classified as no, weak, moderate or strong biofilm producers. All isolates were tested in triplicate.
Nucleotide Sequence Accession Numbers
De novo assembled genomes of the 33 L. monocytogenes isolates included in this study were submitted to GenBank under BioProject no. PRJNA396103 with individual BioSample identification (ID) numbers SAMN07420940 to SAMN07420973.
Results
Whole-Genome SNP Analysis
In order to increase the resolution of whole-genome SNP analysis, assemblies of ST121 and ST14 sequenced isolates were mapped separately against de novo assemblies of the reference genomes (i.e., LSALM51, LSALM1 strains). The resulting ML trees are reported in Figures 1, 2. Within ST14, 23 out of 27 isolates collected from the rabbit meat-processing environment as well as from rabbit meat carcasses, meat cuts and products from November 2005 to November 2006, shared SNPs counts ranging from 0 to 25 SNPs in comparison to the reference genome LSALM1 and were considered as belonging to the same persistent clone. Three isolates (LSALM8, LSALM9, LSALM10), collected from rabbit carcasses from June to August 2006, showed 29–33 SNPs whereas one isolate (LSALM22), collected in November 2006 showed 359 SNPs (Figure 1).
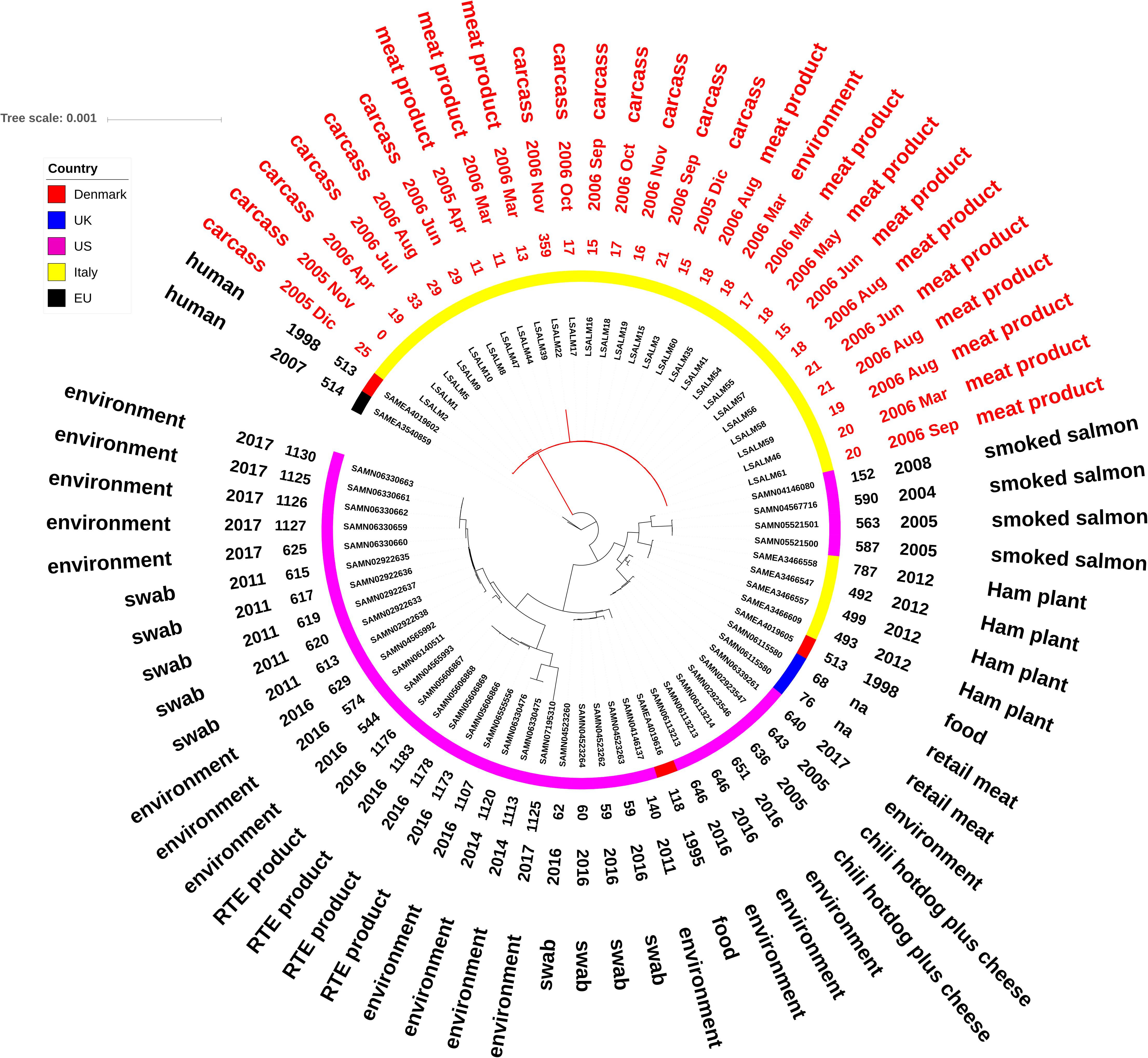
FIGURE 1. Maximum likelihood tree showing genetic distances between ST14 L. monocytogenes draft genomes based on whole-genome core SNPs. The tree tips list study ID of the isolate, date of isolation, origin (rabbit carcass, rabbit meat cut, rabbit meat product, the food processing environment) and the SNP count of the ID genome in comparison to the reference (LSALM1). The scale bar refers to the branch length representing the number of nucleotide substitutions per site.
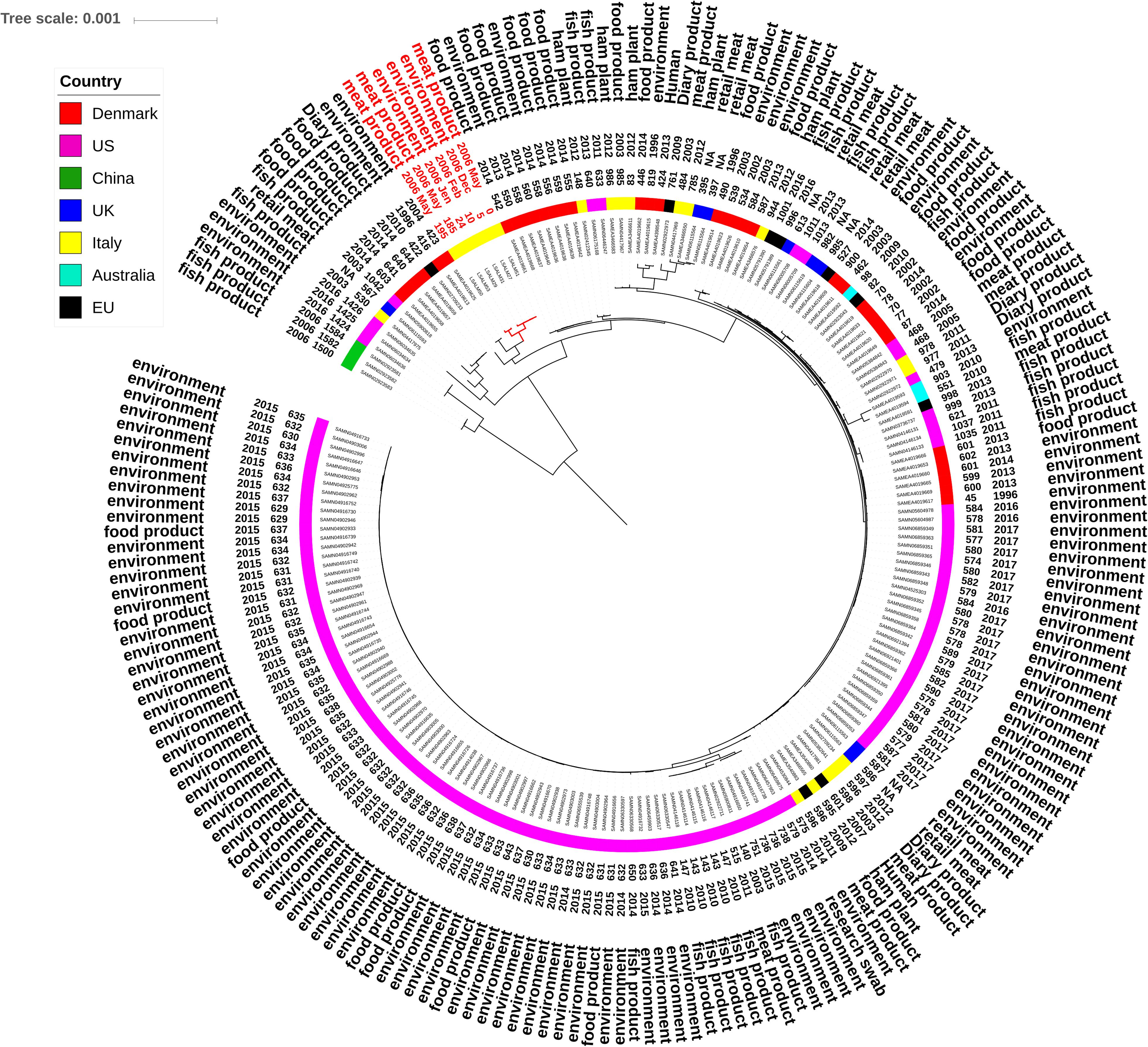
FIGURE 2. Maximum likelihood tree showing genetic distances between ST121 L. monocytogenes draft genomes based on whole-genome core SNPs. The tree tips list study ID of the isolate, date of isolation, origin (rabbit meat cut, the food processing environment) and the SNP count of the ID genome in comparison to the reference (LSALM51). The scale bar refers to the branch length representing the number of nucleotide substitutions per site.
ST121 isolates were clearly differentiated in two distinct clusters (Figure 2). One cluster included isolates LSALM50 and LSALM53, collected from rabbit meat cuts in May 2006 and sharing 204 and 214 SNPs in comparison to the reference genome LSALM51. The second cluster included the reference along with LSALM27, LSALM29, LSALM31. These isolates, sharing from 0 to 17 SNPs, were collected from the rabbit meat-processing environment as well as from meat cuts from December 2005 to May 2006 (Figure 2).
Public ST121 draft genomes showed genetic distances ranging from 45 SNPs (processing environment, Danmark) to 1584 SNPs (fish product, China) in comparison to the internal reference sequence LSALM51 (Figure 1). Public ST14 draft genomes displayed SNPs differences ranging from 60 SNPs (environmental swab, United States) to 1183 SNPs (RTE product, United States). These results confirm the high clonality of L. monocytogenes especially within the same ST.
Dataset of Putative Gene Markers of Ecophysiology in L. monocytogenes
Overall 94 genes, described in 41 published papers, were included in the dataset (Supplementary Table S1). All selected genes were identified as strongly associated to specific phenotypes related to physiological adaptation of L. monocytogenes to environmental stresses encountered in food processing plants. In particular the genes were associated to: resistance to antimicrobials, QAC, heavy metals and bacteriocins; adaptation to cold, high salt concentration, low pH, desiccation and biofilm formation (Supplementary Table S1).
Fourteen genes were included related to resistance to different antimicrobial classes such as tetracycline, ampicillin, vancomycin, streptomycin, chloramphenicol/florfenicol, sulphonamides, erythromycin, and fluoroquinolones (Poyart-Salmeron et al., 1992; Charpentier et al., 1993; Roberts et al., 1996; Godreuil et al., 2003; Srinivasan et al., 2005; Lungu et al., 2011; Jamali et al., 2015).
Seven genes associated to resistance to benzalkonium chloride were included (qacH, qacA, qacC, bcrA, bcrB, bcrC, and emrE). These genes are associated to the active efflux pump of QAC (Elhanafi et al., 2010; Müller et al., 2013; Xu et al., 2014; Kovacevic et al., 2016; Nielsen et al., 2017).
Sixteen genes associated to resistance to cadmium and arsenic were included. In particular, three gene cassettes, cadA1C, cadA2C, and cadA3C were associated to resistance to 70 mg/l of cadmium chloride, whereas cadA4C was associated to resistance to 35 mg/l (Mullapudi et al., 2010; Parsons et al., 2017). Regarding resistance to arsenic, ars genes were associated to resistance to 500 mg/l of sodium (meta) arsenite (Lee et al., 2013).
Regarding bacteriocins resistance, 5 genes associated to the response of L. monocytogenes to cell-envelope stress were included: virR, virS, mprF, liaR, anrB (Thedieck et al., 2006; Collins et al., 2010; Bergholz et al., 2013; Kang et al., 2015).
As far as stress adaptation is considered, different mutant selection experiments demonstrated the important role of the sigB gene (sigma factor B) in the regulation of expression of several genes associated to adaptation to environmental stresses. In particular the knockout of sigB was directly associated to adaptation to desiccation in L. monocytogenes (Huang et al., 2015).
As far as blue-light is considered, the gene lmo0799, coding for a blue-light receptor, was strongly associated to the adaptation of L. monocytogenes to this particular stress (Ondrusch and Kreft, 2011; O’Donoghue et al., 2016).
Listeria monocytogenes can adapt to cold following different pathways. One pathway includes cold shock proteins cspB and cspD (Schmid et al., 2009). Another pathway includes the glycine/betaine transporter system which mediates the uptake of osmolytes, such as glycine, betaine and carnitine, important for adaptation to both cold and high salt concentration (Angelidis and Smith, 2003). Overall, 13 genes related to adaptation of L. monocytogenes to cold and/or high salt concentration were included in the dataset (Angelidis and Smith, 2003; Schmid et al., 2009; Markkula et al., 2012; Pöntinen et al., 2015).
The Stress Survival Islet 1, corresponding to a cassette of five genes (lmo0444-lmo0448), was associated to the survival and growth of L. monocytogenes under suboptimal conditions. In particular, the knockout of the entire SSI-1 was associated to an impaired ability of this food-borne pathogen to grow at low pH and high salt concentrations (Ryan et al., 2010). Within this gene cassette, gadD1 (lmo0447) and gadT1 (lmo0448) encode for a glutamate decarboxylase and an amino acid transporter both described as specifically involved in the adaptation to low pH (Feehily et al., 2014).
Along the GAD System, the ADI system might be activated in response to low pH. The arc gene is involved in transformation of arginine into ornithine with ammonia as by product, which increases the pH. The ADI system has been described in response to mild acid pH (Feehily et al., 2014). Overall, 16 genes associated to low pH adaptation were included in the dataset (Abram et al., 2008; Ryan et al., 2009; Ryan et al., 2010; Feehily et al., 2014).
The Stress Survival Islet 2 (SSI-2) was firstly described as a cassette of two genes of Listeria innocua often present in place of SSI-1 in L. monocytogenes ST121 (Hein et al., 2011). SSI-2 was more recently associated to alkaline and oxidative stress in L. monocytogenes (Harter et al., 2017).
Regarding adaptation to desiccation, seven genes related to motility of L. monocytogenes, were recently associated to this specific phenotype (Hingston et al., 2015). In particular, these genes were found to be downregulated in desiccation tolerant L. monocytogenes.
As far as the ability of biofilm is concerned, 8 genes associated to biofilm formation were included in the dataset. Biofilm formation is essential for survival of L. monocytogenes and further contributes to bacterial persistence in the food processing environment (Popowska et al., 2017).
Screening of Putative Gene Markers of Ecophysiology and Related Phenotypic Tests
The 94 putative gene markers of ecophysiology were screened on 202 and 71 genomes of L. monocytogenes ST121 and ST14, respectively (Figure 3). Twenty-one genes were not found in any of the tested draft genomes and were not included into the heatmap (Figure 3).
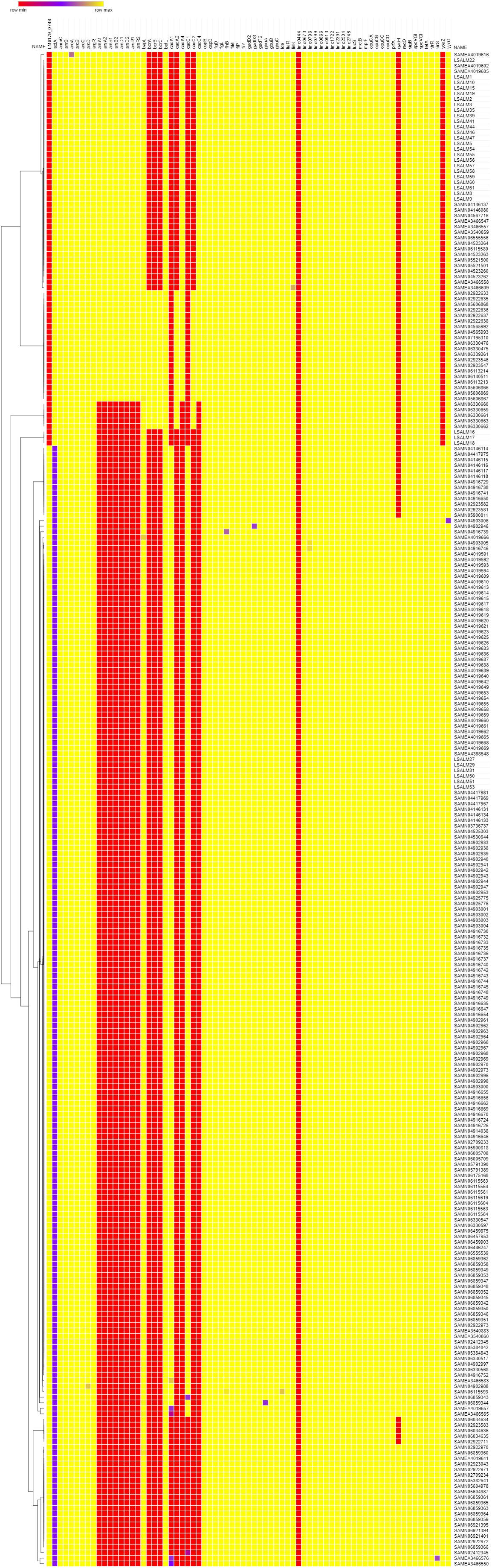
FIGURE 3. Heatmap of the 202 ST121 and 71 ST 14 genomes function to the ecophysiology gene dataset. The rows represent the genomes included while columns represent each gene of the dataset. The colors represent the percentage of identity and length of genes. Red represents lowest percentage while yellow highest percentage. Genes not found in any of the tested genomes were not included.
As far as antimicrobial resistance gene markers are concerned, all genomes were positive for ampC and tetA associated to penicillin and tetracycline resistance, respectively. However, phenotypic tests did not confirm genetic results. Although disk diffusion breakpoints are not available for Listeria, all isolates showed zone diameters equal or higher than 30 mm to tetracycline and ampicillin, suggesting susceptibility (data not shown). Further analysis should be performed in order to assess the reason behind these discordant results.
Regarding resistance to QAC, 25 ST14 genomes (35.2%) and 184 (91.2%) ST121 genomes were positive for the bcrABC locus and the qacH gene, respectively. All ST14 genomes but 8 (88.7%) and none of the ST121 genomes were positive for the ars operon (arsenic resistance) and for the cadA4C gene cassette (cadmium resistance). Further analyses on the localization of these two gene clusters within the genome, revealed that both the ars operon and the cadA4C gene cassette are located in a genomic region already identified as Listeria genomic Island 2 (LGI-2) (Lee et al., 2013; Parsons et al., 2017) (data not shown). In ST14 isolates included in the present study, LGI-2 was found in 24 out of 27 isolates and it was inserted within the gtfA2 gene (data not shown). This gene codes for a sucrose phosphorylase involved in O-glycosylation of proteins. Glycosylation of flagellins is essential for bacterial flagellar assembly, motility, virulence, and host specificity (Merino and Tomás, 2014). The three isolates lacking LGI-2 (LSALM16, LSALM17, LSALM18) belonged to the persistent clone and were collected in September and November 2006 at the end of the sampling period. Further analyses should be performed in order to confirm whether these three isolates are natural mutants, which lost the accessory genome sequence of SGI-2.
Twenty cadA4C positive ST14 genomes additionally carried cadA2C gene cassette. The cadA1C gene cassette was found only in ST121 and in particular in 177 out of the 202 tested genomes (87.6%). Phenotypic tests conducted on the 33 L. monocytogenes strains from the rabbit meat-plant, confirmed that all qacH positive ST121 isolates were resistant to benzalkonium chloride and that all cadA1C positive ST121 isolates were resistant to 70 mg/l of cadmium chloride, whereas all cadA4C positive ST14 isolates were resistant to 35 mg/l of cadmium chloride.
As for environmental stress adaptation, no genomes were positive for the Stress Survival Islet 1 (SSI-1) and all ST121 but no ST14 were positive for the Stress Survival Islet 2 (SSI-2). All tested genomes carried a full-length version of genes associated to: cell-envelope stress response linked to bacteriocins resistance, adaptation to cold and/or high salt concentration, low pH, blue-light and desiccation.
Regarding biofilm formation, the actA gene was truncated in all ST121 and in none of the ST14 genomes. Phenotypic tests confirmed the differential biofilm forming ability of ST121 in comparison to ST14. In particular, tested ST121 isolates were classified as weak biofilm producers with ODC (OD Control) median value of 0.15 and OD values ranging from 0.17 to 0.24 OD, whereas all ST14 isolates but three (LSALM8, LSALM9, LSALM10) were categorized as moderate biofilm producers, with OD values ranging from 0.30 to 0.52 OD at 570 nm. OD values of ST121 isolates were statistically significant different in comparison to ST14 ones (P = 0,00238).
Discussion
In the present study, the persistence of L. monocytogenes ST121 and ST14 repeatedly isolated within 1 year of sampling in a rabbit meat processing plant was studied by a genomic approach. Moreover, 94 putative gene markers of L. monocytogenes physiological adaptation to the food processing environment were investigated in newly sequenced as well as publicly available genomes of ST121 and ST14 strains. The aim was to evaluate the significant enrichment of these genes in the two subtypes, with a particular focus on those genes associated to ecophysiology in ST14, a subtype rarely isolated in food processing plants.
Based on PFGE-Typing, persistence of L. monocytogenes in dairy, meat, fish and vegetable sectors was extensively observed (Leong et al., 2014; Ortiz et al., 2015; Véghová et al., 2017). However, more recently, whole genome SNPs analysis revealed a superior discriminatory power. This is particularly relevant in studies in which highly similar strains have to be differentiated in order to distinguish true persistent from sporadic strains (Ferreira et al., 2014). SNPs calling revealed that the majority but not all of the ST14 isolates sharing the same PFGE-typing, belonged to the same clone. This clone, gathering 23 out of 27 isolates, was re-isolated more than six times within 1 year from rabbit carcasses, meat cuts, meat products and the processing environment in the same rabbit meat processing plant. Although an agreement has not been achieved on the definition of persistence, for the purpose of the present study, the ST14 clone was considered as persistent (Ferreira et al., 2014). This persistent clone included ST14 genomes sharing a maximum of 25 SNPs differences. A difference of 25 SNPs has been already proposed as cut off for definition of genetically related strains belonging to a single persistent clone of L. monocytogenes (Nielsen et al., 2017). The persistence of a L. monocytogenes strain for long periods confirms a higher risk of food contamination and human exposure to specific pathogen strains (Tompkin, 2002). The concern is even higher when the persistent clone belongs to subtype ST 14, which was described as including hypervirulent strains (e.g., ST14) (Mammina et al., 2013; Voronina et al., 2015; Maury et al., 2016; Palma et al., 2017).
Single nucleotide polymorphisms (SNPs) analysis gathered ST121 isolates in two clusters of 2 and 4 isolates each. SNPs analyses showed a higher discriminatory power in comparison to other molecular typing methods (ApaI-PFGE, 7-loci MLST, MLVA, ribotyping), all of which identified all ST121 isolates as belonging to the same clone (De Cesare et al., 2017). The analyzed dataset suggests that one ST121 clone gathering four isolates, survived over 5 months in the same rabbit meat processing plant. However, since those strains were repeatedly isolated in a relative short time frame, this clone cannot be considered as persistent.
In order to gain more insights on the genomic bases behind the differential frequency of ST121 and ST14 in food processing plants, a comprehensive literature review was performed in order to identify genes associated to physiological adaptation of L. monocytogenes to specific food-processing environmental stresses. In the literature, different dataset of genes associated to particular phenotypes, such as antimicrobial resistance and virulence of L. monocytogenes, have been described (Zankari et al., 2012; McArthur et al., 2013). However, to the best of author’s knowledges, no dataset on genes associated to ecophysiology of L. monocytogenes is available.
In the present study, 94 genes associated to ecophysiology were included in a dataset available for the public. Genes were included in the dataset only if their association to the specific phenotype was confirmed by insertional mutagenesis or deletion mutants experiments (Supplementary Table S2). In particular the dataset includes genes associated to resistance to antimicrobials, QAC, heavy metals and bacteriocins as well as associated to adaptation to cold, high salt concentration, low pH, desiccation and biofilm formation.
The results on 202 ST121 and 71 ST14 genomes outlined interesting findings with particular reference to genes significantly enriched in one or the other subtype (Figure 3). In particular genes qacH, cadA1C, Stress Survival Islet -2 and a truncated version of the actA gene were significantly enriched within the ST121 genomes (P < 0,000001), whereas the ars operon and cadA4C gene cassettes included in the Listeria Genomic Islands 2, the bcrABC locus and a full-length version of the actA gene were significantly enriched in ST14 genomes (P < 0,000001). The presence of a QAC associated genetic determinant (either the qacH gene or the bcrABC locus) was significantly enriched in ST121 genomes in comparison to ST14 (P < 0,000001). This observation underlines that subtypes ST121 and ST14 have different patterns of genes associated to ecophysiology. Further studies should confirm whether other subtypes with high and low frequency show the same pattern of genes as identified in ST121 and ST14 subtypes, respectively. Comparing the two patterns it appears that ST121 showed high adaptation to sanitizing procedures (resistance to QAC and adaptation to alkaline stress) (qacH; SSI-2) along with adaptation to high cadmium concentration (cadA1C) (Mullapudi et al., 2010; Müller et al., 2013; Harter et al., 2017). Moreover, ST121 harbored a truncated version of the actA gene whereas ST14 harbored a full-length version of the same gene. The actA gene is involved in the polymerization of actin which is important for motility of L. monocytogenes within the host cell as well as in the first steps of biofilm formation and in particular in cell-to-cell aggregation (Smith and Portnoy, 1997; Travier et al., 2013). Deletion mutants ΔactA showed attenuated virulence and inability to form biofilms (Travier et al., 2013). The higher biofilm forming ability of ST14 in comparison to ST121, was phenotypically confirmed.
All tested genomes carried a full-length version of genes associated to: cell-envelope stress response linked to bacteriocins resistance; adaptation to cold and/or high salt concentration, low pH and desiccation. These genes common to all L. monocytogenes genomes were not informative for the differentiation of the two ST121 and ST14 subtypes. For those genes, mostly located in the core genome, specific allele types can be identified. Genome wide association studies can be performed to associate specific allele types to specific phenotypes. Allele types with a strong association to the phenotype could be then included in the dataset. The usefulness of this approach was already demonstrated. For example a genome-wide association study (GWAS) on Campylobacter jejuni, revealed that, based on overrepresented genetic elements, different C. jejuni subtypes show distinct genotypes associated with survival from farm to fork (Yahara et al., 2017).
Conclusion
The different gene enrichment found in ST121 and ST14 supported by phenotypic confirmations, suggest that ST121 mainly include strains resistant to sanitizers. This feature might in part explain the high frequency of detection of this subtype in food-processing plants. ST14 include biofilm producer strains. This feature suggest that ST14 might occasionally contaminate harborage sites where sanitizing procedures are difficult to be performed. If confirmed, this route of contamination might be of concern since ST14 could potentially spread from harborage sites to different food lots scattered in months or years. The concern is even higher if a hypervirulent strain is the driver of this repeated contamination.
Author Contributions
FrP designed the study, performed the literature review of genes related to ecophysiology, and draft the manuscript. FeP performed data analysis and contributed in manuscript writing. LG contributed to the setup of the dataset. AL performed the phenotypic tests. ADC contributed to the literature review. GM collected the samples, revised the manuscript, and coordinated the study. All authors have contributed to data interpretation, have critically reviewed the manuscript, and approved the final version as submitted.
Funding
This study was funded by European Union Horizon 2020 Research and Innovation Program: COMPARE (collaborative management platform for detection and analyses of [re-]emerging and foodborne outbreaks in Europe: grant number 643476).
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors gratefully acknowledge Prof. Mirko Rossi of University of Helsinki for supportive, critical and helpful discussions. Moreover, the authors wish to thank Dr. Jani Halkilahti for support in collecting public Listeria genomes.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.00596/full#supplementary-material
TABLE S1 | Dataset of putative gene markers of ecophysiology in L. monocytogenes.
TABLE S2 | Public genomes of L. monocytogenes ST121 and ST14 included in the present study.
Footnotes
- ^https://github.com/B-UMMI/INNUca
- ^https://github.com/tseemann/snippy
- ^https://github.com/rambaut/figtree
- ^https://www.ncbi.nlm.nih.gov/pubmed/
- ^https://blast.ncbi.nlm.nih.gov/Blast.cgi
- ^https://www.ebi.ac.uk/Tools/msa/clustalo/
- ^https://www.ncbi.nlm.nih.gov/nuccore/
- ^https://github.com/tseemann/abricate
- ^https://software.broadinstitute.org/morpheus/
References
Abram, F., Starr, E., Karatzas, K. A., Matlawska-Wasowska, K., Boyd, A., Wiedmann, M., et al. (2008). Identification of components of the sigma B regulon in Listeria monocytogenes that contribute to acid and salt tolerance. Appl. Environ. Microbiol. 74, 6848–6858. doi: 10.1128/AEM.00442-08
Angelidis, A. S., and Smith, G. M. (2003). Three transporters mediate uptake of glycine betaine and carnitine by Listeria monocytogenes in response to hyperosmotic stress. Appl. Environ. Microbiol. 69, 1013–1022. doi: 10.1128/AEM.69.2.1013-1022.2003
Beier, S., and Bertilsson, S. (2013). Bacterial chitin degradation-mechanisms and ecophysiological strategies. Front. Microbiol. 4:149. doi: 10.3389/fmicb.2013.00149
Bergholz, T. M., Tang, S., Wiedmann, M., and Boor, K. J. (2013). Nisin resistance of Listeria monocytogenes is increased by exposure to salt stress and is mediated via LiaR. Appl. Environ. Microbiol. 79, 5682–5688. doi: 10.1128/AEM.01797-13
Carpentier, B., and Cerf, O. (2011). Review — Persistence of Listeria monocytogenes in food industry equipment and premises. Int. J. Food Microbiol. 145, 1–8. doi: 10.1016/j.ijfoodmicro.2011.01.005
Charpentier, E., Gerbaud, G., and Courvalin, P. (1993). Characterization of a new class of tetracycline-resistance gene tet(S) in Listeria monocytogenes BM4210. Gene 131, 27–34. doi: 10.1016/0378-1119(93)90665-P
Clinical and Laboratory Standards Institute [CLSI] (2010). Methods for Antimicrobial Dilution and Disk Susceptibility Testing of Infrequently Isolated or Fastidious Bacteria, 2nd Edn. Wayne, PA: CLSI.
Collins, B., Curtis, N., Cotter, P. D., Hill, C., and Ross, R. P. (2010). The ABC transporter AnrAB contributes to the innate resistance of Listeria monocytogenes to nisin, bacitracin, and various beta-lactam antibiotics. Antimicrob. Agents Chemother. 54, 4416–4423. doi: 10.1128/AAC.00503-10
De Cesare, A., Parisi, A., Mioni, R., Comin, D., Lucchi, A., and Manfreda, G. (2017). Listeria monocytogenes circulating in rabbit meat products and slaughterhouses in Italy: prevalence data and comparison among typing results. Foodborne Pathog. Dis. 14, 167–176. doi: 10.1089/fpd.2016.2211
Elhanafi, D., Dutta, V., and Kathariou, S. (2010). Genetic characterization of plasmid-associated benzalkonium chloride resistance determinants in a Listeria monocytogenes strain from the 1998-1999 outbreak. Appl. Environ. Microbiol. 76, 8231–8238. doi: 10.1128/AEM.02056-10
Feehily, C., Finnerty, A., Casey, P. G., Hill, C., Gahan, C. G., O’Byrne, C. P., et al. (2014). Divergent evolution of the activity and regulation of the glutamate decarboxylase systems in Listeria monocytogenes EGD-e and 10403S: roles in virulence and acid tolerance. PLoS One 9:e112649. doi: 10.1371/journal.pone.0112649
Ferreira, V., Barbosa, J., Stasiewicz, M., Vongkamjan, K., Moreno Switt, A., Hogg, T., et al. (2011). Diverse geno- and phenotypes of persistent Listeria monocytogenes isolates from fermented meat sausage production facilities in Portugal. Appl. Environ. Microbiol. 77, 2701–2715. doi: 10.1128/AEM.02553-10
Ferreira, V., Wiedmann, M., Teixeira, P., and Stasiewicz, M. J. (2014). Listeria monocytogenes persistence in food-associated environments: epidemiology, strain characteristics, and implications for public health. J. Food Prot. 77, 150–170. doi: 10.4315/0362-028X.JFP-13-150
Garrison, E., and Marth, G. (2012). Haplotype-Based Variant Detection from Short-Read Sequencing. Available at: http://arxiv.org/abs/1207.3907 [accessed August 6, 2017].
Godreuil, S., Galimand, M., Gerbaud, G., Jacquet, C., and Courvalin, P. (2003). Efflux pump Lde is associated with fluoroquinolone resistance in Listeria monocytogenes. Antimicrob. Agents Chemother. 47, 704–708. doi: 10.1128/AAC.47.2.704-708.2003
Gray, M. J., Freitag, N. E., and Boor, K. J. (2006). How the bacterial pathogen Listeria monocytogenes mediates the switch from environmental Dr. Jekyll to Pathogenic Mr. Hyde. Infect. Immun. 74, 2505–2512. doi: 10.1128/IAI.74.5.2505-2512.2006
Harter, E., Wagner, E. M., Zaiser, A., Halecker, S., Wagner, M., and Rychli, K. (2017). Stress survival islet 2, predominantly present in Listeria monocytogenes strains of sequence type 121, is involved in the alkaline and oxidative stress responses. Appl. Environ. Microbiol. 83:e00827-17. doi: 10.1128/AEM.00827-17
Hein, I., Klinger, S., Dooms, M., Flekna, G., Stessl, B., Leclercq, A., et al. (2011). Stress survival islet 1 (SSI-1) survey in Listeria monocytogenes reveals an insert common to Listeria innocua in sequence type 121 L. monocytogenes strains. Appl. Environ. Microbiol. 77, 2169–2173. doi: 10.1128/AEM.02159-10
Hingston, P., Chen, J., Dhillon, B. K., Laing, C., Bertelli, C., Gannon, V., et al. (2017). Genotypes associated with Listeria monocytogenes isolates displaying impaired or enhanced tolerances to cold, salt, acid, or desiccation stress. Front. Microbiol. 8:369. doi: 10.3389/fmicb.2017.00369
Hingston, P. A., Piercey, M. J., and Truelstrup Hansen, L. (2015). Genes associated with desiccation and osmotic stress in Listeria monocytogenes as revealed by insertional mutagenesis. Appl. Environ. Microbiol. 81, 5350–5362. doi: 10.1128/AEM.01134-15
Holch, A., Webb, K., Lukjancenko, O., Ussery, D., Rosenthal, B. M., and Gram, L. (2013). Genome sequencing identifies two nearly unchanged strains of persistent Listeria monocytogenes isolated at two different fish processing plants sampled 6 years apart. Appl. Environ. Microbiol. 79, 2944–2951. doi: 10.1128/AEM.03715-12
Huang, Y., Ells, T. C., and Truelstrup Hansen, L. (2015). Role of sigB and osmolytes in desiccation survival of Listeria monocytogenes in simulated food soils on the surface of food grade stainless steel. Food Microbiol. 46, 443–451. doi: 10.1016/j.fm.2014.09.007
Jamali, H., Paydar, M., Ismail, S., Looi, C. Y., Wong, W. F., Radmehr, B., et al. (2015). Prevalence, antimicrobial susceptibility and virulotyping of Listeria species and Listeria monocytogenes isolated from open-air fish markets. BMC Microbiol. 15:144. doi: 10.1186/s12866-015-0476-7
Kang, J., Wiedmann, M., Boor, K. J., and Bergholz, T. M. (2015). VirR-Mediated resistance of Listeria monocytogenes against food antimicrobials and cross-protection induced by exposure to organic acid salts. Appl. Environ. Microbiol. 81, 4553–4562. doi: 10.1128/AEM.00648-15
Kathariou, S. (2002). Listeria monocytogenes virulence and pathogenicity, a food safety perspective. J. Food Prot. 65, 1811–1829. doi: 10.4315/0362-028X-65.11.1811
Knudsen, G. M., Nielsen, J. B., Marvig, R. L., Ng, Y., Worning, P., Westh, H., et al. (2017). Genome-wide-analyses of Listeria monocytogenes from food-processing plants reveal clonal diversity and date the emergence of persisting sequence types. Environ. Microbiol. Rep. 9, 428–440. doi: 10.1111/1758-2229.12552
Kovacevic, J., Ziegler, J., Wałecka-Zacharska, E., Reimer, A., Kitts, D. D., and Gilmour, M. W. (2016). Tolerance of Listeria monocytogenes to quaternary ammonium sanitizers is mediated by a novel efflux pump encoded by emrE. Appl. Environ. Microbiol. 82, 939–953. doi: 10.1128/AEM.03741-15
Lebrun, M., Audurier, A., and Cossart, P. (1994). Plasmid-borne cadmium resistance genes in Listeria monocytogenes are similar to cadA and cadC of Staphylococcus aureus and are induced by cadmium. J. Bacteriol. 176, 3040–3048. doi: 10.1128/jb.176.10.3040-3048.1994
Lee, S., Rakic-Martinez, M., Graves, L. M., Ward, T. J., Siletzky, R. M., and Kathariou, S. (2013). Genetic determinants for cadmium and arsenic resistance among Listeria monocytogenes serotype 4b isolates from sporadic human listeriosis patients. Appl. Environ. Microbiol. 79, 2471–2476. doi: 10.1128/AEM.03551-12
Leong, D., Alvarez-Ordóñ A., and Jordan, K. (2014). Monitoring occurrence and persistence of Listeria monocytogenes in foods and food processing environments in the Republic of Ireland. Front. Microbiol. 5:436. doi: 10.3389/fmicb.2014.00436
Li, H., and Durbin, R. (2009). Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 25, 1754–1760. doi: 10.1093/bioinformatics/btp324
Li, H., Handsaker, B., Wysoker, A., Fennell, T., Ruan, J., Homer, N., et al. (2009). The Sequence Alignment/Map format and SAMtools. Bioinformatics 25, 2078–2079. doi: 10.1093/bioinformatics/btp352
Lungu, B., O’Bryan, C. A., Muthaiyan, A., Milillo, S. R., Johnson, M. G., Crandall, P. G., et al. (2011). Listeria monocytogenes: antibiotic resistance in food production. Foodborne Pathog. Dis. 8, 569–578. doi: 10.1089/fpd.2010.0718
Mammina, C., Parisi, A., Guaita, A., Aleo, A., Bonura, C., Nastasi, A., et al. (2013). Enhanced surveillance of invasive listeriosis in the Lombardy region, Italy, in the years 2006-2010 reveals major clones and an increase in serotype 1/2a. BMC Infect. Dis. 13:152. doi: 10.1186/1471-2334-13-152
Markkula, A., Mattila, M., Lindström, M., and Korkeala, H. (2012). Genes encoding putative DEAD-box RNA helicases in Listeria monocytogenes EGD-e are needed for growth and motility at 3°C. Environ. Microbiol. 14, 2223–2232. doi: 10.1111/j.1462-2920.2012.02761.x
Maury, M. M., Tsai, Y.-H., Charlier, C., Touchon, M., Chenal-Francisque, V., Leclercq, A., et al. (2016). Uncovering Listeria monocytogenes hypervirulence by harnessing its biodiversity. Nat. Genet. 48, 308–313. doi: 10.1038/ng.3501
McArthur, A. G., Waglechner, N., Nizam, F., Yan, A., Azad, M. A., Baylay, A. J., et al. (2013). The comprehensive antibiotic resistance database. Antimicrob. Agents Chemother. 57, 3348–3357. doi: 10.1128/AAC.00419-13
Merino, S., and Tomás, J. M. (2014). Gram-Negative Flagella Glycosylation. Int. J. Mol. Sci. 15, 2840–2857. doi: 10.3390/ijms15022840
Morganti, M., Scaltriti, E., Cozzolino, P., Bolzoni, L., Casadei, G., Pierantoni, M., et al. (2016). Processing-dependent and clonal contamination patterns of Listeria monocytogenes in the cured ham food chain revealed by genetic analysis. Appl. Environ. Microbiol. 82, 822–831. doi: 10.1128/AEM.03103-15
Moura, A., Criscuolo, A., Pouseele, H., Maury, M. M., Leclercq, A., Tarr, C., et al. (2016). Whole genome-based population biology and epidemiological surveillance of Listeria monocytogenes. Nat. Microbiol. 2:16185. doi: 10.1038/nmicrobiol.2016.185
Mullapudi, S., Siletzky, R. M., and Kathariou, S. (2008). Heavy-metal and benzalkonium chloride resistance of Listeria monocytogenes isolates from the environment of turkey-processing plants. Appl. Environ. Microbiol. 74, 1464–1468. doi: 10.1128/AEM.02426-07
Mullapudi, S., Siletzky, R. M., and Kathariou, S. (2010). Diverse cadmium resistance determinants in Listeria monocytogenes isolates from the turkey processing plant environment. Appl. Environ. Microbiol. 76, 627–630. doi: 10.1128/AEM.01751-09
Müller, A., Rychli, K., Muhterem-Uyar, M., Zaiser, A., Stessl, B., Guinane, C. M., et al. (2013). Tn6188 - a novel transposon in Listeria monocytogenes responsible for tolerance to benzalkonium chloride. PLoS One 8:e76835. doi: 10.1371/journal.pone.0076835
Murugesan, L., Kucerova, Z., Knabel, S. J., and LaBorde, L. F. (2015). Predominance and distribution of a persistent Listeria monocytogenes clone in a commercial fresh mushroom processing environment. J. Food Prot. 78, 1988–1998. doi: 10.4315/0362-028X.JFP-15-195
Nielsen, E. M., Björkman, J. T., Kiil, K., Grant, K., Dallman, T., Painset, A., et al. (2017). Closing Gaps for Performing a Risk Assessment on Listeria monocytogenes in Ready-to-Eat (RTE) Foods: Activity 3, the Comparison of Isolates from Different Compartments Along the Food Chain, and from Humans using Whole Genome Sequencing (WGS) Analysis. Parma: EFSA. doi: 10.2903/sp.efsa.2017.EN-1151
O’Donoghue, B., NicAogáin, K., Bennett, C., Conneely, A., Tiensuu, T., and Johansson, J. (2016). Blue-light inhibition of Listeria monocytogenes growth is mediated by reactive oxygen species and is influenced by σB and the blue-light sensor Lmo0799. Appl. Environ. Microbiol. 82, 4017–4027. doi: 10.1128/AEM.00685-16
Olier, M., Pierre, F., Lemaître, J.-P., Divies, C., Rousset, A., and Guzzo, J. (2002). Assessment of the pathogenic potential of two Listeria monocytogenes human faecal carriage isolates. Microbiology 148, 1855–1862. doi: 10.1099/00221287-148-6-1855
Olier, M., Pierre, F., Rousseaux, S., Lemaître, J.-P., Rousset, A., Piveteau, P., et al. (2003). Expression of truncated Internalin A is involved in impaired internalization of some Listeria monocytogenes isolates carried asymptomatically by humans. Infect. Immun. 71, 1217–1224. doi: 10.1128/IAI.71.3.1217-1224.2003
Ondrusch, N., and Kreft, J. (2011). Blue and red light modulates SigB-dependent gene transcription, swimming motility and invasiveness in Listeria monocytogenes. PLoS One 6:e16151. doi: 10.1371/journal.pone.0016151
Orsi, R. H., den Bakker, H. C., and Wiedmann, M. (2011). Listeria monocytogenes lineages: genomics, evolution, ecology, and phenotypic characteristics. Int. J. Med. Microbiol. 301, 79–96. doi: 10.1016/j.ijmm.2010.05.002
Ortiz, S., López-Alonso, V., Rodríguez, P., and Martínez-Suárez, J. V. (2015). The connection between persistent, disinfectant-resistant Listeria monocytogenes strains from two geographically separate Iberian pork processing plants: evidence from comparative genome analysis. Appl. Environ. Microbiol. 82, 308–317. doi: 10.1128/AEM.02824-15
Palma, F., Pasquali, F., Lucchi, A., Cesare, A. D., and Manfreda, G. (2017). Whole genome sequencing for typing and characterisation of Listeria monocytogenes isolated in a rabbit meat processing plant. Ital. J. Food Saf. 6:6879. doi: 10.4081/ijfs.2017.6879
Parsons, C., Lee, S., Jayeola, V., and Kathariou, S. (2017). Novel cadmium resistance determinant in Listeria monocytogenes. Appl. Environ. Microbiol. 83:e2580-e16. doi: 10.1128/AEM.02580-16
Pöntinen, A., Markkula, A., Lindström, M., and Korkeala, H. (2015). Two-component-system histidine kinases involved in growth of Listeria monocytogenes EGD-e at low temperatures. Appl. Environ. Microbiol. 81, 3994–4004. doi: 10.1128/AEM.00626-15
Popowska, M., Krawczyk-Balska, A., Ostrowski, R., and Desvaux, M. (2017). InlL from Listeria monocytogenes is involved in biofilm formation and Adhesion to Mucin. Front. Microbiol. 8:660. doi: 10.3389/fmicb.2017.00660
Poyart-Salmeron, C., Trieu-Cuot, P., Carlier, C., MacGowan, A., McLauchlin, J., and Courvalin, P. (1992). Genetic basis of tetracycline resistance in clinical isolates of Listeria monocytogenes. Antimicrob. Agents Chemother. 36, 463–466. doi: 10.1128/AAC.36.2.463
Ratani, S. S., Siletzky, R. M., Dutta, V., Yildirim, S., Osborne, J. A., Lin, W., et al. (2012). Heavy metal and disinfectant resistance of Listeria monocytogenes from foods and food processing plants. Appl. Environ. Microbiol. 78, 6938–6945. doi: 10.1128/AEM.01553-12
Roberts, M. C., Facinelli, B., Giovanetti, E., and Varaldo, P. E. (1996). Transferable erythromycin resistance in Listeria spp. isolated from food. Appl. Environ. Microbiol. 62, 269–270.
Ryan, S., Begley, M., Gahan, C. G. M., and Hill, C. (2009). Molecular characterization of the arginine deiminase system in Listeria monocytogenes: regulation and role in acid tolerance. Environ. Microbiol. 11, 432–445. doi: 10.1111/j.1462-2920.2008.01782.x
Ryan, S., Begley, M., Hill, C., and Gahan, C. G. M. (2010). A five-gene stress survival islet (SSI-1) that contributes to the growth of Listeria monocytogenes in suboptimal conditions. J. Appl. Microbiol. 109, 984–995. doi: 10.1111/j.1365-2672.2010.04726.x
Schmid, B., Klumpp, J., Raimann, E., Loessner, M. J., Stephan, R., and Tasara, T. (2009). Role of cold shock proteins in growth of Listeria monocytogenes under cold and osmotic stress conditions. Appl. Environ. Microbiol. 75, 1621–1627. doi: 10.1128/AEM.02154-08
Smith, G. A., and Portnoy, D. A. (1997). How the Listeria monocytogenes ActA protein converts actin polymerization into a motile force. Trends Microbiol. 5, 272–276. doi: 10.1016/S0966-842X(97)01048-2
Srinivasan, V., Nam, H. M., Nguyen, L. T., Tamilselvam, B., Murinda, S. E., and Oliver, S. P. (2005). Prevalence of antimicrobial resistance genes in Listeria monocytogenes isolated from dairy farms. Foodborne Pathog. Dis. 2, 201–211. doi: 10.1089/fpd.2005.2.201
Stasiewicz, M. J., Oliver, H. F., Wiedmann, M., and den Bakker, H. C. (2015). Whole-genome sequencing allows for improved identification of persistent Listeria monocytogenes in food-associated environments. Appl. Environ. Microbiol. 81, 6024–6037. doi: 10.1128/AEM.01049-15
Stepanović, S., Cirković, I., Ranin, L., and Svabić-Vlahović, M. (2004). Biofilm formation by Salmonella spp. and Listeria monocytogenes on plastic surface. Lett. Appl. Microbiol. 38, 428–432. doi: 10.1111/j.1472-765X.2004.01513.x
Thedieck, K., Hain, T., Mohamed, W., Tindall, B. J., Nimtz, M., Chakraborty, T., et al. (2006). The MprF protein is required for lysinylation of phospholipids in listerial membranes and confers resistance to cationic antimicrobial peptides (CAMPs) on Listeria monocytogenes. Mol. Microbiol. 62, 1325–1339. doi: 10.1111/j.1365-2958.2006.05452.x
Tompkin, R. B. (2002). Control of Listeria monocytogenes in the food-processing environment. J. Food Prot. 65, 709–725. doi: 10.4315/0362-028X-65.4.709
Travier, L., Guadagnini, S., Gouin, E., Dufour, A., Chenal-Francisque, V., Cossart, P., et al. (2013). ActA promotes Listeria monocytogenes aggregation, intestinal colonization and carriage. PLoS Pathog. 9:e1003131. doi: 10.1371/journal.ppat.1003131
Van Stelten, A., Simpson, J. M., Ward, T. J., and Nightingale, K. K. (2010). Revelation by single-nucleotide polymorphism genotyping that mutations leading to a premature stop codon in inlA Are common among Listeria monocytogenes isolates from ready-to-eat foods but not human listeriosis cases. Appl. Environ. Microbiol. 76, 2783–2790. doi: 10.1128/AEM.02651-09
Véghová, A., Minarovièová, J., Koreňová, J., Drahovská, H., and Kaclíková, E. (2017). Prevalence and tracing of persistent Listeria monocytogenes strains in meat processing facility production chain. J. Food Saf. 37: e12315. doi: 10.1111/jfs.12315
Voronina, O. L., Kunda, M. S., Ryzhova, N. N., Aksenova, E. I., Semenov, A. N., Kurnaeva, M. A., et al. (2015). Regularities of the ubiquitous polyhostal microorganisms selection by the example of three taxa. Mol. Biol. 49, 430–441. doi: 10.7868/S0026898415030179
Xu, D., Li, Y., Zahid, M. S. H., Yamasaki, S., Shi, L., Li, J., et al. (2014). Benzalkonium chloride and heavy-metal tolerance in Listeria monocytogenes from retail foods. Int. J. Food Microbiol. 190, 24–30. doi: 10.1016/j.ijfoodmicro.2014.08.017
Xu, D., Nie, Q., Wang, W., Shi, L., and Yan, H. (2016). Characterization of a transferable bcrABC and cadAC genes-harboring plasmid in Listeria monocytogenes strain isolated from food products of animal origin. Int. J. Food Microbiol. 217, 117–122. doi: 10.1016/j.ijfoodmicro.2015.10.021
Yahara, K., Méric, G., Taylor, A. J., de Vries, S. P. W., Murray, S., Pascoe, B., et al. (2017). Genome-wide association of functional traits linked with Campylobacter jejuni survival from farm to fork. Environ. Microbiol. 19, 361–380. doi: 10.1111/1462-2920.13628
Keywords: Listeria monocytogenes, the food processing environment, persistence, stress, adaptation
Citation: Pasquali F, Palma F, Guillier L, Lucchi A, De Cesare A and Manfreda G (2018) Listeria monocytogenes Sequence Types 121 and 14 Repeatedly Isolated Within One Year of Sampling in a Rabbit Meat Processing Plant: Persistence and Ecophysiology. Front. Microbiol. 9:596. doi: 10.3389/fmicb.2018.00596
Received: 16 December 2017; Accepted: 15 March 2018;
Published: 29 March 2018.
Edited by:
Giovanna Suzzi, Università di Teramo, ItalyReviewed by:
Beatrix Stessl, Veterinärmedizinische Universität Wien, AustriaHana Drahovska, Comenius University, Slovakia
Copyright © 2018 Pasquali, Palma, Guillier, Lucchi, De Cesare and Manfreda. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Frédérique Pasquali, ZnJlZGVyaXF1ZS5wYXNxdWFsaUB1bmliby5pdA==