 Katarzyna Siegel-Hertz1
Katarzyna Siegel-Hertz1 Véronique Edel-Hermann1Emilie Chapelle2
Véronique Edel-Hermann1Emilie Chapelle2 Sébastien Terrat1
Sébastien Terrat1 Jos M. Raaijmakers2,3
Jos M. Raaijmakers2,3 Christian Steinberg1*
Christian Steinberg1*- 1Agroécologie, AgroSup Dijon, Institut National de la Recherche Agronomique, Université Bourgogne Franche-Comté, Dijon, France
- 2Laboratory of Phytopathology, Wageningen University, Wageningen, Netherlands
- 3Department of Microbial Ecology, Netherlands Institute of Ecology (NIOO-KNAW), Wageningen, Netherlands
Disease-suppressive soils are soils in which specific soil-borne plant pathogens cause only limited disease although the pathogen and susceptible host plants are both present. Suppressiveness is in most cases of microbial origin. We conducted a comparative metabarcoding analysis of the taxonomic diversity of fungal and bacterial communities from suppressive and non-suppressive (conducive) soils as regards Fusarium wilts sampled from the Châteaurenard region (France). Bioassays based on Fusarium wilt of flax confirmed that disease incidence was significantly lower in the suppressive soil than in the conducive soil. Furthermore, we succeeded in partly transferring Fusarium wilt-suppressiveness to the conducive soil by mixing 10% (w/w) of the suppressive soil into the conducive soil. Fungal diversity differed significantly between the suppressive and conducive soils. Among dominant fungal operational taxonomic units (OTUs) affiliated to known genera, 17 OTUs were detected exclusively in the suppressive soil. These OTUs were assigned to the Acremonium, Chaetomium, Cladosporium, Clonostachys, Fusarium, Ceratobasidium, Mortierella, Penicillium, Scytalidium, and Verticillium genera. Additionally, the relative abundance of specific members of the bacterial community was significantly higher in the suppressive and mixed soils than in the conducive soil. OTUs found more abundant in Fusarium wilt-suppressive soils were affiliated to the bacterial genera Adhaeribacter, Massilia, Microvirga, Rhizobium, Rhizobacter, Arthrobacter, Amycolatopsis, Rubrobacter, Paenibacillus, Stenotrophomonas, and Geobacter. Several of the fungal and bacterial genera detected exclusively or more abundantly in the Fusarium wilt-suppressive soil included genera known for their activity against F. oxysporum. Overall, this study supports the potential role of known fungal and bacterial genera in Fusarium wilt suppressive soils from Châteaurenard and pinpoints new bacterial and fungal genera for their putative role in Fusarium wilt suppressiveness.
Introduction
Plant diseases caused by soil-borne microorganisms, including fungi, oomycetes, bacteria, nematodes as well as subterranean insects, regularly result in extensive losses in agricultural production every year. Because of its role in many enzymatic activities governing the functioning of the soil, particularly in biogeochemical cycles, reorganization, and mineralization of organic matter, and feeding of plants, soil microbial diversity is a determining component of soil health (Garbeva et al., 2004; Janvier et al., 2007; Chaparro et al., 2012; Larkin, 2015; van Bruggen et al., 2015). Hence, growth of plant pathogens in soils and subsequent plant infection can be reduced through competitive interactions with the soil microbial community which results in defining disease suppressive soils (Weller et al., 2002; Raaijmakers et al., 2009; Lundgren and Fergen, 2011; Kyselková and Moënne-Loccoz, 2012; Penton et al., 2014). Indeed, soils in which disease incidence or severity commonly remain low in spite of the presence of the pathogen, a susceptible host plant and climatic conditions that would allow disease development are called disease suppressive soils (Baker and Cook, 1974). General disease suppressiveness of soils is based on multitrophic interactions and can be modulated by soil management practices that affect total microbial activity (Mazzola and Gu, 2002; Stirling et al., 2012). In addition to general suppressiveness, some soils exhibit an additional level of suppressiveness targeted to a specific soil-borne plant pathogen. Specific suppressiveness is attributed to the converging activities of specific members of the soil microbial community that interfere with the disease cycle of the pathogen. This is the case, for example, of bacteria in the Pseudomonas group that produce metabolites such as pyoverdins, iron-chelating siderophores, and make iron difficult to access for the pathogenic fungus F. oxysporum, this mechanism being added to the competition for carbon to which this same pathogen is confronted (Lemanceau et al., 1993; Alabouvette, 1999). Specific suppressiveness can be eliminated by soil sterilization, steam pasteurization, or gamma-irradiation and can be transferred by mixing a small amount of natural suppressive soil into both previously disinfected suppressive soil and natural conducive soil (Alabouvette, 1986; Weller et al., 2002; Garbeva et al., 2004; Mendes et al., 2011). Although the role of abiotic components cannot be ruled out (Amir and Alabouvette, 1993; Almario et al., 2013a), all of these demonstrations clearly indicate that a focus must be placed on the biotic component (Alabouvette, 1999).
This intriguing phenomenon of specific disease suppressiveness has been described for a number of soil-borne pathogens, including bacteria, fungi, oomycetes, and nematodes. Among fungi, special attention has been paid to soils suppressive to take-all disease of wheat (Weller et al., 2002), damping-off diseases caused by Rhizoctonia solani (Mendes et al., 2011) and wilt diseases caused by different formae speciales of Fusarium oxysporum (Toussoun, 1975; Louvet et al., 1976; Alabouvette, 1986; Alabouvette et al., 2009). For decades, many vegetables were and are still produced in the market gardening region of Châteaurenard, in France and yet very few symptoms of Fusarium wilt are observed despite the presence of F. oxysporum in the soil which led to consider this soil as suppressive to this disease (Louvet et al., 1976; Alabouvette, 1986). Because the situation was unique, we conducted various studies, over the past 40 years, to determine the microorganisms and mechanisms involved in Fusarium wilt suppressiveness of this soil from the Châteaurenard region. Alabouvette (1986) postulated that the suppressive nature of this soil relied on both (i) a general mechanism of competition for nutrients caused by the whole soil microflora and (ii) a specific competition between pathogenic and non-pathogenic Fusarium strains. Subsequent studies further revealed that bacteria of the Pseudomonas genus contributed, in combination with non-pathogen F. oxysporum, to suppressiveness via siderophore-mediated competition for iron and via the production of antifungal phenazines (Lemanceau and Alabouvette, 1993; Duijff et al., 1999; Mazurier et al., 2009). We used next-generation sequencing to conduct a comparative metabarcoding analysis of the microbiomes of both the Fusarium wilt suppressive soil of Châteaurenard and a non-suppressive soil from a nearby field. The analysis was based on 454-pyrosequencing of the fungal internal transcribed spacer (ITS) region and the bacterial 16S rRNA gene. The aim was to detect and identify fungal and bacterial genera associated with Fusarium-wilt-suppressive soils not yet identified and get new insights into potentially novel microbial genera and mechanisms involved in Fusarium wilt suppressiveness.
Materials and Methods
Soil Sampling
The Fusarium-wilt-suppressive soil was harvested from a field that had remained a fallow for several years in the Châteaurenard region, France (43°53′15″N, 4°51′36″E). The conducive soil was sampled nearby from a temporarily uncultivated field following muskmelon cropping and before lettuce cropping (43°51′53″N, 4°50′04″E). The two soils were geographically very close (3 km apart). Both were collected in October 2011, air-dried and sieved to 4 mm. Both were loamy soils. Physicochemical profiles were respectively 17.1% clay, 47.4% silt, 35.5% sand, 1.98% organic matter (OM), 0.9 N (g/kg), and pH 8.5 for the suppressive soil, and 20.6% clay, 43.2% silt, 36.2% sand, 4.6% OM, 1.5 N (g/kg), pH 8.06 for the conducive soil. Differences in organic matter content, nitrogen content and pH were related to the current uses of the soils. To confirm the biological nature of the soil suppressiveness, the suppressive soil was autoclaved three times on three consecutive days for 20 min at 120°C and stored at room temperature for 1 week before use.
Assessment of Soil Disease Suppressiveness
Soil disease suppressiveness was tested under greenhouse conditions with flax as the host plant and the flax pathogen F. oxysporum f. sp. lini MIAE00347 (Collection of Microorganisms of Interest for Agriculture and Environment, INRA Dijon, France). The plants were grown in different types of soil: suppressive soil (S), conducive soil (C), conducive soil amended with 10% (w/w) of suppressive soil (referred to as mixed soil or M), and heat-treated suppressive soil. Soils were inoculated with 103 conidia of the pathogen per mL of soil, and flax (Linum usitatissimum L. variety Opaline) seeds were sown on the same day. For each modality, three replicates were performed with 20 plants/replicate. Non-inoculated soils were used as controls. Plants were grown in a growth chamber with 70% relative humidity, a 16/8 h daylight/dark photoperiod at 17°C (day) and 15°C (night) the first week, and 25°C (day) and 22°C (night) afterwards. Diseased plants were recorded 25 days after inoculation. Data were subjected to one-way analysis of variance (ANOVA). Pairwise comparisons were performed using Fisher's test (P < 0.05).
Rhizospheric Soil Sampling
Flax seeds were sown in 9/9/9.5 cm pots containing 300 g of soil. Twenty plants per pot were grown under the same conditions of light and humidity as above. Four modalities were prepared: soil S, soil C, soil M, and suppressive soil inoculated with 103 conidia of F. oxysporum f. sp. lini MIAE00347 per mL of soil (referred to as pathogen-inoculated suppressive soil or IS soil). The inoculum dose used was relatively low (103 conidia per mL) to ensure that plants do not die too quickly in the conducive soil and prevent to collect the active rhizospheric soil needed to conduct microbial communities comparative analyses. Three replicates per modality were performed. After 25 days, the rhizospheric soil was collected from each replicate. The root systems were isolated and shaken to remove free soil particles. The soil around the roots influenced by root development and plant exudates was considered as rhizospheric soil. It was sieved to 2 mm to remove fine roots and organic debris. Large pieces of roots were removed manually with tweezers. Two grams of soil samples from each modality were placed in 2-ml cryotubes and frozen at −20°C for further DNA extractions.
DNA Extraction, PCR Amplification, and Construction of Pyrosequencing Libraries
The protocol used for DNA extractions was described by Plassart et al. (2012). Extraction was performed by mechanical lysis using FastPrep®-24 (MP-Biomedicals, NY, USA) with a lysis buffer and two purification steps with PVPP (polyvinyl polypyrrolydone) minicolumns (BIORAD, Marne-la-Coquette, France) and Geneclean Turbo kit (Q-Biogene, Illkirch, France). Total DNA was extracted from each replicate of the four modalities. Each purified DNA sample was quantified by a fluorometric assay using the PicoGreen dsDNA Assay Kit (Invitrogen Life Technologies) and the StepOnePlus™System (Applied Biosystems®) according to the manufacturer's instructions. DNA extracted from each soil sample served as a template in PCR reactions to amplify a fungal barcode and a bacterial barcode. Three replicates were used for each of the four soil modalities. PCR reactions were performed in quadruplicate for each replicate, with 20 ng of soil DNA.
For fungal identification, the variable internal transcribed spacer 1 (ITS1) region was amplified using specific fungal primers ITS1F (5′-CTTGGTCATTTAGAGGAAGTAA-3′) and ITS2 (5′-GCTGCGTTCTTCATCGATGC-3′; White et al., 1990; Gardes and Bruns, 1993; Buée et al., 2009). The primers were tagged with four-base-pair multiplex identifiers (MIDs) at the 5′ and 3′ positions to specifically identify each sample, as recommended by the manufacturer (Beckman Coulter Genomics). PCR conditions were as follows: 95°C for 5 min, 35 cycles of 30 s at 95°C, 40 s at 53°C, and 45 s at 72°C, followed by 7 min at 72°C.
For bacterial identification, a 16S rRNA gene fragment (partial V3, V4 and partial V5) was amplified using primers 530F (5′-ACTCCTACGGGAGGCAGCAG-3′; Acosta-Martínez et al., 2008) and 803R (5′-CTACCNGGGTATCTAAT-3′; Zancarini et al., 2013). Ten-base-pair MIDs at the 5′ and 3′ positions were added to the primers to specifically identify each sample, as recommended by the manufacturer (Beckman Coulter Genomics). PCR conditions were as follows: 95°C for 10 min, 30 cycles of 30 s at 95°C, 30 s at 62°C, and 60 s at 72°C, followed by 10 min at 72°C. All amplifications were performed in a Mastercycler (Eppendorf, Hamburg, Germany). For each sample, amplicons of the four replicated PCRs were pooled and purified using a MinElute PCR Purification Kit (Qiagen, Courtaboeuf, France) following the manufacturer's protocol. Amplicon concentrations were then estimated by fluorometric assay (PicoGreen dsDNA Assay Kit). For each of the two amplified barcodes, an equimolar pooling of all samples was prepared (total DNA amount: ~3.2 μg per library). Adapter sequences were added by ligation as recommended by the manufacturer, and 454-pyrosequencing was carried out by Beckman Coulter Genomics (Danvers, USA) on a Genome Sequencer FLX 454 (Life Sciences/Roche Applied Biosystems).
Bioinformatics Analysis
Concerning the sequenced ITS1 region, sequences were sorted into different files according to their MID using the sfffile program of Roche 454 main software with default parameters. Mismatches were not allowed for MIDs and were removed. Sequences were then cleaned by trimming and denoising using trim.flows in Mothur version 1.20.1 with the default parameters (Schloss et al., 2009), and reverse-complemented if needed. ITS1 was extracted using Fungal ITS Extractor, version 2 (Nilsson et al., 2010) and sequences were filtered based on a minimal length of 100 bp. Operational taxonomic units (OTUs) were generated after two successive clustering steps using Uclust version 3.0 [Usearch version 5.2.32, (Edgar, 2010)] at 97% similarity. The first clustering included all sequences, and the second clustering was conducted with the batch of consensus sequences from the previous clustering.
Concerning the sequenced 16S region, sequences were analyzed using the GnS-PIPE pipeline initially developed by the GenoSol platform (INRA, Dijon, France) and recently optimized (Terrat et al., 2012, 2015). First, all the raw 16S reads were sorted according to their multiplex identifier sequences. Then, a preprocessing step was realized to filter and delete low-quality reads based on (i) their length (<350 bases), (ii) their number of ambiguities (deletion of reads with one N or more), and (iii) their primer sequence(s) (the distal and proximal primer sequences must be complete and without errors). A PERL program was then applied for rigorous dereplication (i.e., clustering of strictly identical sequences with same length). The dereplicated reads were aligned using INFERNAL alignment (Cole et al., 2009; v1.0.2 with chosen parameters: –hbanded, –sub, –dna) to obtain a global alignment. Then, aligned sequences were clustered into OTUs at 93.8% of similarity using the CrunchClust V43 program (Hartmann et al., 2012) that groups rare reads with abundant ones and does not count differences in homopolymer lengths (default parameters were selected). Here, an OTU is defined by the most abundant read, known as the centroid, and every read in the OTU must have similarity above the given similarity threshold with the centroid). The chosen level of similarity was defined by an in silico approach (data not shown). We first extracted all known and reliable microbial sequences (bacteria and archaea) from the SILVA database (release v111) (Quast et al., 2013). We kept from these sequences only the amplified regions of sequences using our primer set, and deleted those with too many mismatches with our primer set (more than 3 mismatches with one primer). Then, all these “artificial reads” with a reliable taxonomy were clustered using our specific program of clustering at various threshold levels (100% to 90%, with a 0.1% step). Regarding the results and the amplified regions, the 93.8% threshold was the best suited to efficiently define the genus level, as it was the closest one to the genus level defined by the SILVA taxonomy of analyzed sequences.
For both the sequenced ITS and 16S regions, a filtering step was carried out to check all singletons (OTUs supported by only one sequence likely to be artifacts, such as PCR chimeras) and potentially to delete them from downstream analysis based on the quality of their taxonomic assignments. Then, to efficiently compare datasets and avoid biased community comparisons, high-quality reads were normalized by random selection: each dataset with a large number of reads was randomly cut down to the same number of reads as the dataset with the lowest number of reads (3,451 and 2,505 reads for fungal and bacterial communities, respectively) using a homemade Perl program (similar to the single_rarefaction.py with default parameters from QIIME). The retained high-quality reads were then used for taxonomy-independent analyses, and several diversity and richness indices were determined using the defined OTU composition. More precisely, we used the number of bacterial and fungal OTUs from each sample to determine Shannon and Evenness indices as indicators of soil microbial diversity and structure, respectively. Taxonomy-based analysis was also performed using similarity approaches against: (i) the UNITE database (fungal rDNA ITS sequence database; Kõljalg et al., 2013) using the Basic Local Alignment Search Tool algorithm BLASTn version 2.2.23 (Altschul et al., 1997) with the filter turned off for fungal sequences; (ii) a dedicated reference database from SILVA (Quast et al., 2013), using the USEARCH tool (Edgar, 2010) for bacterial sequences. Finally, global analysis of the soil samples was computed by merging all homogenized high-quality reads from each sample into one global file and defining OTUs as previously described before subsequent analyses.
All raw sequences collected in this study have been deposited in the European Bioinformatics Institute nucleotide sequence database system under the accession number PRJEB24081.
Statistical Analysis and Heat Map Computation
The OTUs defined for fungal and bacterial datasets were used to perform rarefaction analysis using Analytic Rarefaction version 1.3 (Hunt Mountain Software, Department of Geology, University of Georgia, Athens, GA, USA). The differences in fungal and bacterial community compositions (number of genera, OTUs, Shannon and Evenness indices) were assessed by Kruskal–Wallis tests under XLSTAT software (Addinsoft®). The OTU tables obtained from ITS1 and 16S datasets were used to perform an ordination analysis by means of Principal Component Analysis (PCA), using ADE-4 version 2001 (Biometry and Evolutionary Biology Laboratory, University Claude Bernard Lyon 1). For each microbial community, the numbers of unique OTUs and shared OTUs among the different soils were compared in Venn diagrams using VENNY (Oliveros, 2007).
Heat maps were built by applying the heatmap.2 function implemented in “gplots” R package based on the relative abundance values of the dominant fungal OTUs (for which the relative abundance level in one soil at least was higher than 1%) and dominant bacterial OTUs (for which the relative abundance level in one soil at least was higher than 1.5%) across all samples using the global matrices. Datasets were analyzed together, so the mean values of Z-scores were from the total dataset. The relative abundance levels of each row (corresponding to each OTU) were expressed as median-centered Z-scores between all samples, and the colors scaled to standard deviations of the corresponding Z-scores. All these analyses were performed with R free software (version 3.2.3). Moreover, to minimize the probability of incorrect assignments of the dominant OTUs, taxonomic assignments of representative sequences were refined by checking taxonomic assignments of dominant fungal OTUs against the GenBank database. The default algorithm parameters were used, and environmental sequences were excluded. Only sequences associated with a publication and a description of the corresponding strain were considered. Assignments were taken into account at the genus level. Some synonyms were corrected using the online Species Fungorum database (Kirk, 2017). For the few OTUs that had the same percentage of similarity with several fungal genera or families, assignment was made at the family or order level, respectively. Concerning the dominant bacterial OTUs, their taxonomic assignments were checked using SINA online (Pruesse et al., 2012), the SILVA database (version 123), and default parameters. SINA uses the search result to derive a classification with the LCA (lowest common ancestor) method. Each query sequence is assigned the shared part of the classifications of the search results. OTU abundance levels in the different soils were compared based on Kruskal-Wallis tests using XLSTAT software (Addinsoft®).
Results
Disease Suppressiveness of the Soils
The plants grown in the non-inoculated soils showed no wilt disease symptoms. Typical symptoms of Fusarium wilt were observed in pathogen-inoculated soils. The percentage of healthy plants was significantly higher (P < 0.05) in soil S (46.6 ± 10.6) and soil M (43.5 ± 19.4) than in soil C (15.3 ± 5.7). The percentage of healthy plants was significantly lower (P < 0.05) in heat-treated suppressive soil (9.2 ± 0.3) than in soils S, M and C, further confirming the biological nature of the soil suppressiveness.
Microbiome Data Acquisition and Statistics
Fungal and bacterial targeted regions were successfully amplified by PCR and sequenced for all soils. The raw sequence libraries were filtered to remove reads originating from sequencing errors or putative chimeric sequences. For the fungal and bacterial datasets, 81,902 reads (88% of raw sequences), and 67,675 reads (89% of raw sequences) passed all quality controls, respectively. The number of high-quality reads ranged from 3,451 to 10,821 for fungal datasets, and from 2,505 to 8,842 for bacterial datasets. After homogenization, a total of 41,412 high-quality fungal ITS sequences were clustered into 1,798 OTUs for all soil samples. Among them, 570 OTUs were considered as singletons, and represented 31.7% of the detected richness. Regarding bacterial communities, 2,280 distinct OTUs, including 1,162 singletons (50.9% of the richness), were observed for 30,060 high-quality bacterial 16S rDNA sequences. Rarefaction curves were drawn for fungal and bacterial datasets based on OTUs (Supplementary Figure 1). Fungal curves showed that: (i) the number of OTUs at 97% similarity increased with the number of sequences, and saturation was not reached for all soils; (ii) based on soil replicates, however, the number of sequences was sufficient to obtain a representative coverage of the major fungal groups; and (iii) differences among soils were recorded in the slope and level of the curves (Supplementary Figure 1A). As regards bacterial richness, rarefaction curves also revealed that the number of OTUs increased with the number of reads, without reaching a plateau, and reads were in sufficient numbers to allow for an accurate description of the major bacterial genera in each of the soil samples (Supplementary Figure 1B). Interestingly, no significant differences were detected in the slope and level of bacterial curves in any of the soils, showing less heterogeneity of bacterial richness among the four modalities.
Fungal and bacterial communities were also evaluated using richness and diversity indices (Table 1). Similar numbers of genera were detected in the different soils. However, significant differences among soils were recorded (P < 0.05) for the number of fungal OTUs, and for Shannon and Evenness indices. Thus, soil M harbored the highest fungal richness (462.0 ± 26 OTUs on average), and soils C and IS the lowest. However, soils C and M had the highest Shannon indices, and soil IS the lowest. Finally, evenness was significantly higher in soil C (P < 0.05) than in soils S and IS. Contrary to fungal communities, bacterial communities did not significantly differ in the number of genera, the number of OTUs, and the Shannon and Evenness indices between the different soils (Table 1).
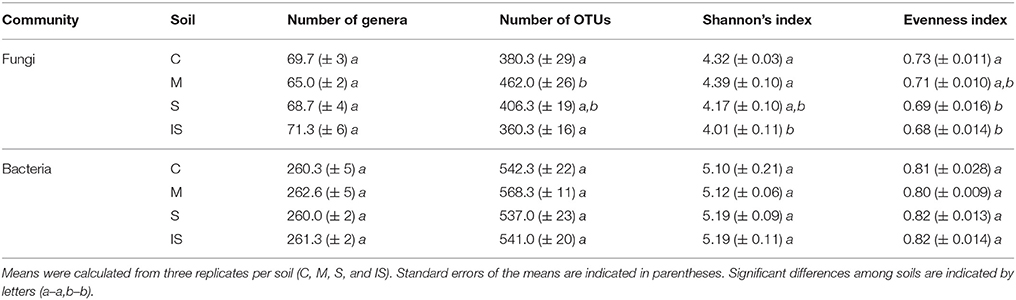
Table 1. Fungal and bacterial richness and diversity indices of the conducive soil (C), mixed soil (M), suppressive soil (S), and pathogen-inoculated suppressive soil (IS).
Fungal and Bacterial Community Composition
Fungal and bacterial sequences were assigned at different taxonomic levels (phylum, class, order, family, and genus; Supplementary Table 1). Concerning fungal communities, most assigned sequences belonged to the Ascomycota phylum (90.9%). Basidiomycota and Zygomycota accounted for 2.6 and 1.5% of the sequences, respectively. The proportion of Chytridiomycota and Glomeromycota was <1%. Finally, 4.4% of the sequences were not assigned and were considered as undefined. At the class level, Sordariomycetes were dominant within Ascomycota, followed by Eurotiomycetes, Dothideomycetes, and Pezizomycetes (Figure 1A).
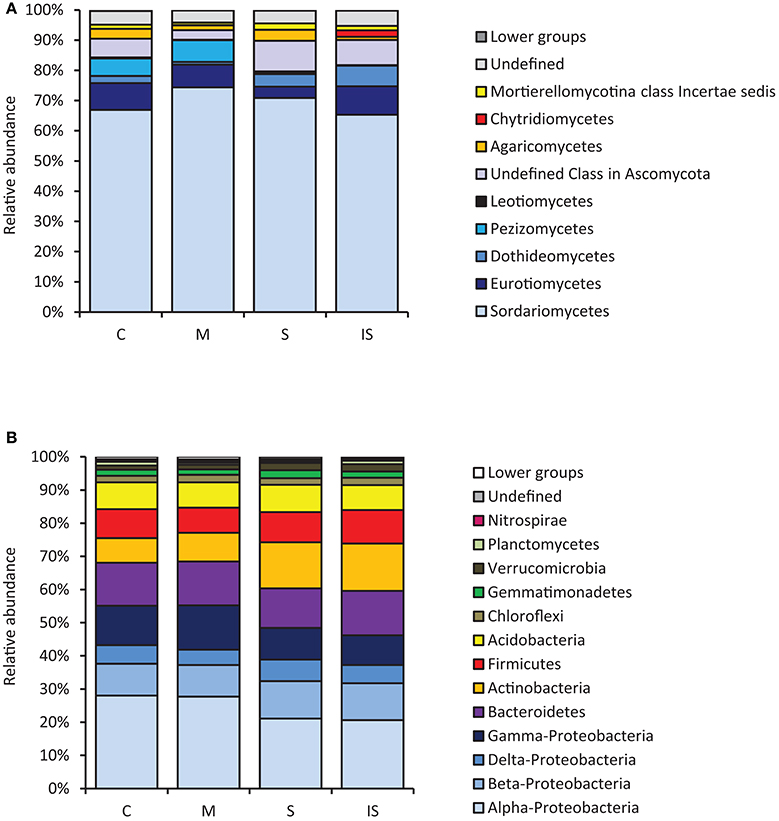
Figure 1. Relative abundance levels of fungal classes (A) and bacterial phyla (B), expressed as percentages of all sequences, detected in the conducive soil (C), mixed soil (M), suppressive soil (S), and pathogen-inoculated suppressive soil (IS).
Concerning bacterial communities, the soils had the same overall bacterial composition, with Proteobacteria, Bacteroidetes, Actinobacteria, Firmicutes, and Acidobacteria as major phyla (Figure 1B). Proteobacteria was the dominant bacterial phylum and represented 46–55% of all bacterial DNA sequences. Several minor phyla (Chloroflexi, Gemmatimonadetes, Verrucomicrobia, Planctomycetes, and Nitrospirae) were also identified; they accounted for ca. 7% of bacterial communities. However, a few differences were found among soils, with more Gamma-Proteobacteria in soil M than in soils S and IS (P < 0.05). In contrast, Actinobacteria were more represented in soils S and IS than in soil C (P < 0.05).
Microbial Community Composition Associated With Suppressive and Conducive Soils
The structure of the fungal and bacterial communities was analyzed by PCA to obtain an ordination on a factorial map and to compare the soils based on global OTU composition (Figure 2). A clear discrimination of fungal communities based on soil types was observed (Figure 2A). In particular, two groups of soils were separated along the first axis: soils C and M on the one hand, and soils S and IS on the other hand. Moreover, soil C grouped apart from soil M, and soil S apart from soil IS along the second axis. Concerning bacterial communities, sample discrimination was not so obvious (Figure 2B): soils C and M were separated from soils S and IS along the first axis, but no distinction of these two groups was visible along the second axis.
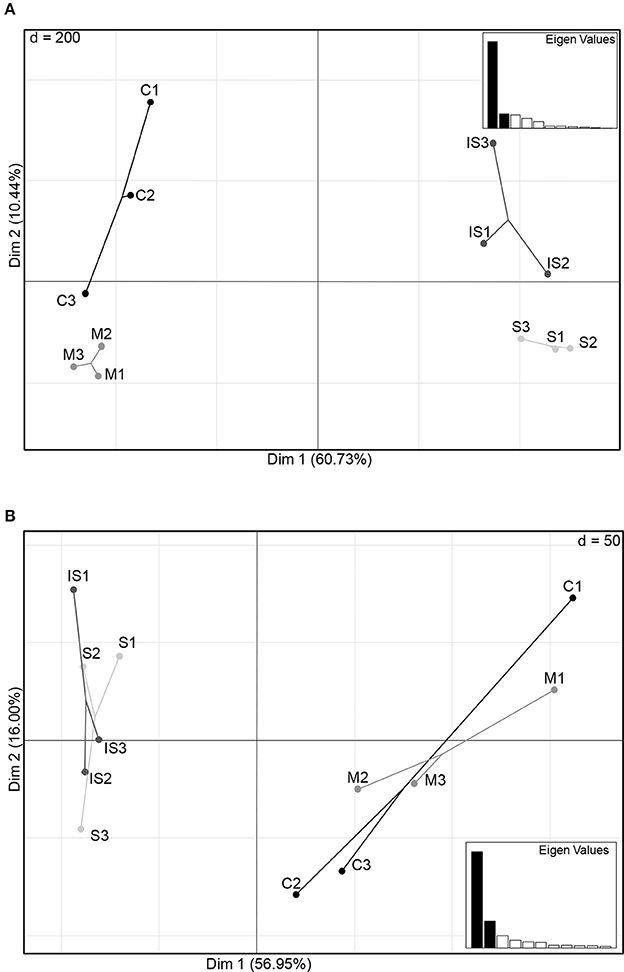
Figure 2. Principal component plots of the conducive soil (C), mixed soil (M), suppressive soil (S), and pathogen-inoculated suppressive soil (IS) generated from fungal (A) and bacterial (B) OTU matrices of ITS1 and 16S relative abundance levels.
Regarding OTU composition, 189 fungal OTUs and 414 bacterial OTUs were shared between soils C and S (Figure 3). In the fungal community, 205 OTUs were unique in soil C, and 271 in soil S. In the bacterial community, 355 OTUs were unique in soil C, and 293 in soil S. To further focus on the microbial genera associated with disease suppressiveness, we compared the fungal and bacterial community compositions between soils S and M on the one hand, and between soils S and IS on the other hand. 122 fungal OTUs and 100 bacterial OTUs were shared between soils S and M but absent from soil C. In addition, 214 fungal OTUs and 145 bacterial OTUs shared between soils S and IS were potentially promoted by the presence of the pathogen.
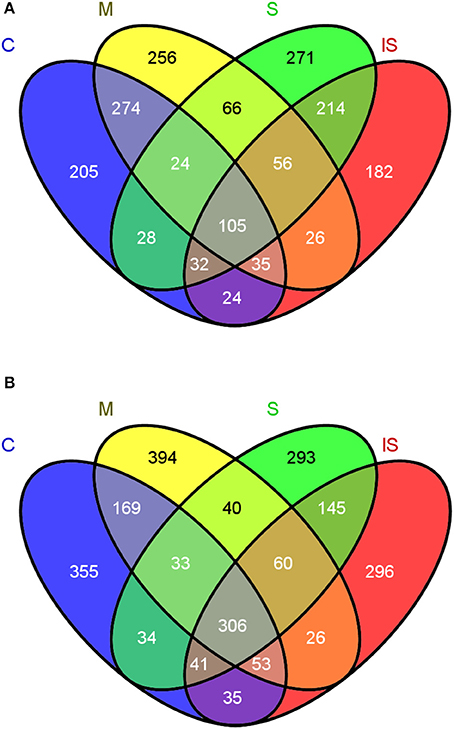
Figure 3. Venn diagrams indicating the numbers of shared and unique OTUs between the conducive soil (C), mixed soil (M), suppressive soil (S), and pathogen-inoculated suppressive soil (IS) in fungal (A) and bacterial communities (B).
To combine taxonomic assignment and soil global OTU composition, we compared microbial community compositions based on the relative abundance of the detected microbial genera using a heat map. Due to the large size of the dataset, only the most dominant fungal OTUs (relative abundance level > 1%) and bacterial OTUs (relative abundance level > 1.5%) were mapped (Figures 4, 5). The 143 dominant fungal OTUs represented in the heat map accounted for 83% of total reads on average, and the 122 dominant bacterial OTUs kept in the heat map accounted for 72% of total reads on average (Figures 4, 5). Firstly, these heat maps showed that biological replicates from a same soil were more similar to each other than replicates from other soils, whatever the soil. Moreover, evidence of clear differences between soils C and M on the one hand, and soils S and IS on the other hand was illustrated by both bacterial and fungal community compositions. Concerning fungal communities, 24 OTUs affiliated to 15 known genera were relatively more abundant in soils S and IS than in soil C (P < 0.05; Figure 4, underlined OTUs). Among them, 17 OTUs were not detected at all in soil C. They were assigned to the Acremonium, Chaetomium, Cladosporium, Clonostachys, Fusarium, Ceratobasidium, Mortierela, Penicillium, Scytalidium, and Verticillium genera. Interestingly, these OTUs were also detected in soil M. All 10 genera mentioned above, exclusively detected in soils S, IS and M, have been described for their antagonistic activity against diverse plant pathogens (Table 2). Moreover, the Acremonium, Chaetomium, Clonostachys, Fusarium, and Penicillium genera include strains used in the biological control of pathogenic F. oxysporum (Table 2).
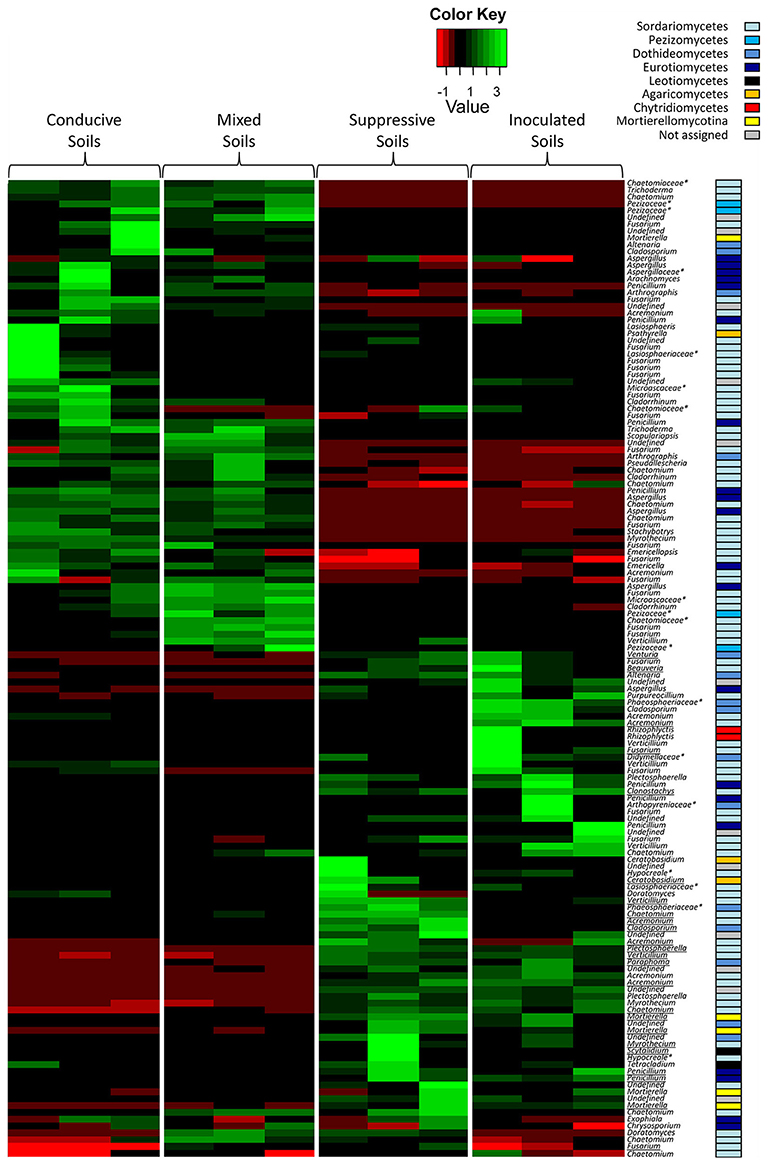
Figure 4. Heat map comparison of the dominant fungal genera detected in the soils according to each modality. The four different modalities (conducive soil, mixed soil, suppressive soil, and pathogen-inoculated suppressive soil) were organized based on the UPGMA dendrogram of UNIFRAC weighted and normalized distances between corresponding soil samples. The legend shows the Z-scores (relative abundance levels are expressed as median-centered Z-scores between all samples, and colors are scaled to standard deviations). OTUs with a star (*) indicate groups that were not assigned to a precise fungal genus, but to a higher taxonomical group. Underlined OTUs indicate significant differences in the relative abundance levels of particular fungal genera in the four modalities.
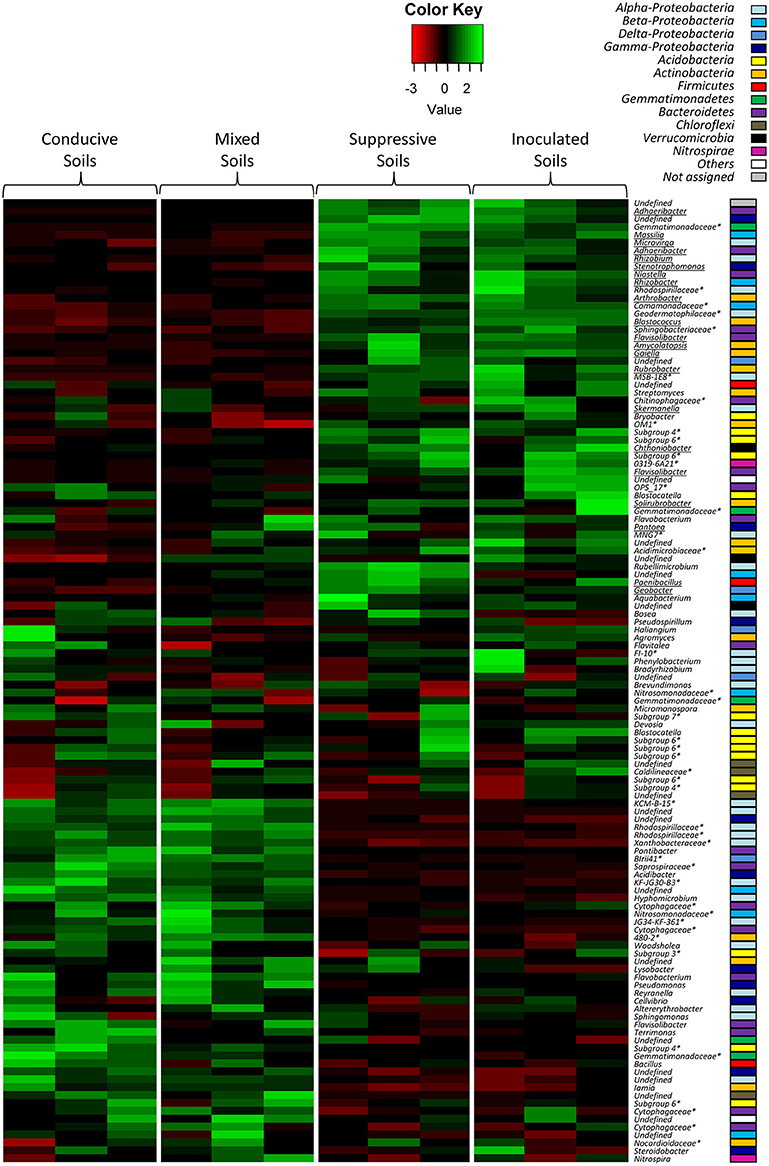
Figure 5. Heat map comparison of the dominant bacterial genera detected in the soils according to each modality. The four different modalities (conducive soil, mixed soil, suppressive soil, and pathogen-inoculated suppressive soil) were organized based on the UPGMA dendrogram of UNIFRAC weighted and normalized distances between corresponding soil samples. The legend shows the Z-scores (relative abundance levels are expressed as median-centered Z-scores between all samples, and colors are scaled to standard deviations). OTUs with a star (*) indicate groups that were not assigned to a precise bacterial genus, but to a higher taxonomical group. Underlined OTUs indicate significant differences in the relative abundance levels of particular bacterial genera in the four modalities.
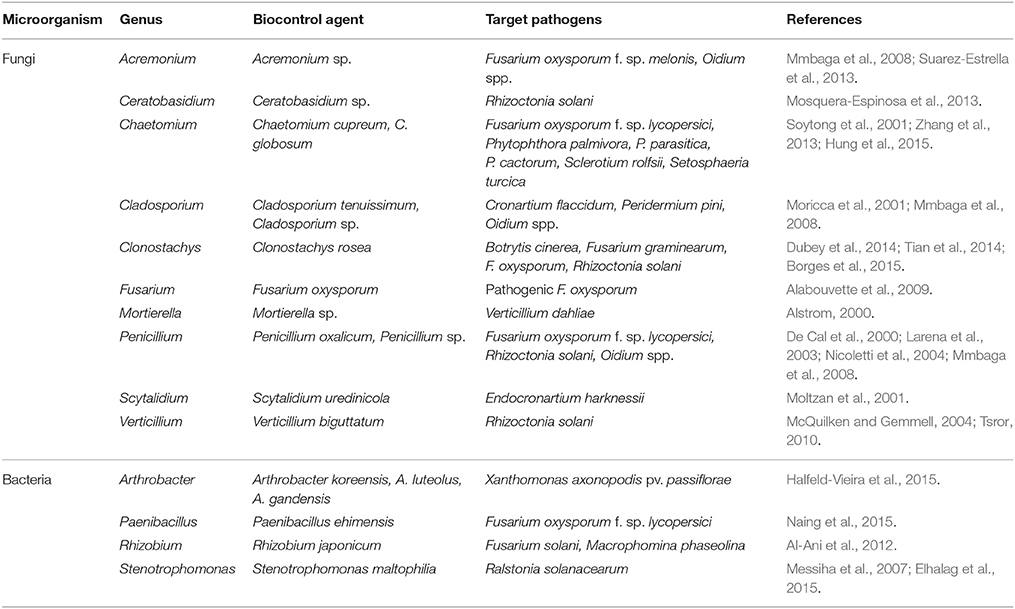
Table 2. Microbial genera including OTUs associated with suppressive soil (S), pathogen-inoculated suppressive soil (IS), and mixed soil (M) and known to include biological control agents against fungal diseases.
Concerning bacterial communities, the OTUs detected in soils C and M also differed from those of soils S and IS, with dominant Alpha-Proteobacteria and Gamma-Proteobacteria in soils C and M, and dominant Actinobacteria in soils S and IS (Figure 5). More precisely, several OTUs affiliated to known bacterial genera were more abundant in soils S than in soil C (P < 0.05), e.g., Adhaeribacter, Amycolatopsis, Arthrobacter, Geobacter, Massilia, Microvirga, Paenibacillus, Rhizobacter, Rhizobium, Rubrobacter, and Stenotrophomonas (Figure 5, underlined OTUs). Interestingly, some of these OTUs (i.e., Adhaeribacter, Amycolatopsis, Arthrobacter, Paenibacillus, Rhizobacter, Rubrobacter, and Stenotrophomonas) were also more abundant in soil IS than in soil C (P < 0.05). Finally, some of these OTUs (i.e., Adhaeribacter, Amycolatopsis, Paenibacillus, Rhizobacter, and Rubrobacter) were also more abundant in soil M than in soil C (P < 0.05). Interestingly, four genera mentioned above (Arthrobacter, Paenibacillus, Rhizobium, and Stenotrophomonas) are known to have pathogen suppression potential (Table 2), and two of them (Paenibacillus and Rhizobium) include strains used in the biological control of pathogenic Fusarium (Table 2).
Discussion
Châteaurenard soil has been so far a unique model for understanding the microbial nature of soils naturally suppressive to Fusarium wilts (Alabouvette, 1986, 1999). The uniqueness of this soil lies in the native and long-lasting character of disease suppressiveness, whereas suppressiveness of other soils and other diseases is acquired and transient (Weller et al., 2002; Mazzola, 2007; Mendes et al., 2011). In most cases, attention was mainly paid to decipher the role of bacterial communities (Kyselková et al., 2009; Mendes et al., 2011; Gómez Expósito et al., 2017) while in the present study, both bacterial and fungal communities are taken into consideration. Our suppressive and conducive soils subjected to globally equivalent intensive cropping systems (market gardening) under the same pedoclimatic conditions harbored similar specific bacterial richness. Moreover, diversity indices did not at all highlight specific traits likely to explain the different behaviors with respect to disease. Only fungal communities slightly varied, based on OTU numbers and Shannon and Evenness indices. The highest number of OTUs recorded in the conducive soil amended with suppressive soil may have resulted from the accumulation of specific microorganisms from both soils. Analyses of assigned OTUs showed that bacterial community and even more markedly fungal community membership and structure differed in this Châteaurenard suppressive soil as compared to the nearby conducive soil. Some of these differing OTUs belonged to known genera that harbored strains with biocontrol activity against plant pathogenic fungi including F. oxysporum, like Acremonium, Chaetomium, Clonostachys, Fusarium, and Penicillium. Analyzing the diversity of microbial communities remains descriptive, nevertheless, the presence of these taxa in the suppressive soil of Châteaurenard cannot leave indifferent as for their possible contribution to the control of F. oxysporum. For example, the presence and abundance of the Acremonium and Chaetomium genera were associated with reduced disease and reduced F. oxysporum abundance during field observations following specific agricultural practices (Zhao et al., 2014; Huang et al., 2015; Ma et al., 2015). These two genera, along with the Penicillium genus, had a similar effect on cereal diseases caused by the Fusarium Head Blight species complex in which F. oxysporum is however little involved (Vujanovic et al., 2012; Kohl et al., 2015). Further evidence of the involvement of these genera in F. oxysporum control was highlighted under controlled conditions. For example, an A. chrysogenum strain controlled F. oxysporum f. sp. melonis development and reduced disease severity in dual conditions, in vermiculite (Suarez-Estrella et al., 2013). These were analytical studies that revealed a huge diversity of mechanisms within genera, such as mentioned above, that confer an important bioprotective potential against numerous phytopathogenic targets including F. oxysporum (Daguerre et al., 2016). These mechanisms are mainly based on the production of secondary metabolites. In the Chaetomium genus, and more specifically in the C. globosum species, strains produce polyketides such as chaetoviridin A or chaetoglobosin X, which inhibit F. oxysporum development (Park et al., 2005; Wang et al., 2012). Penicillium chrysogenum strains produce small cysteine-rich fungicidal proteins that target several fungal species including F. oxysporum (Kaiserer et al., 2003). Clonostachys catenulatum and C. rosea are more known to inhibit F. graminearum development by producing lactonohydrolases or hydrophobins (Dubey et al., 2014; Popiel et al., 2014), but their ability to control F. oxysporum development by producing β-1,3-glucanases besides mycoparasitism has also been demonstrated (Chatterton and Punja, 2009; Tian et al., 2014). Finally, inside the F. oxysporum species itself, in addition to the already mentioned trophic competition mechanisms (Alabouvette et al., 2009), strains produce secondary metabolites such as terpenes. These specifically impacted formae speciales such as F. oxysporum f. sp. lactucae, whose growth was inhibited by α-humulene from strain F. oxysporum MSA35 (Minerdi et al., 2009). On the other hand, F. oxysporum has developed mechanisms of resistance to mycotoxins produced by other species from the Fusarium genus (Dawidziuk et al., 2016). Therefore, these other species probably contribute only weakly to soil suppressiveness of Fusarium wilt, or do so via other modes of action which remain to be identified.
None of the few examples of antagonistic activity mentioned above is highly specific or completely biocidal and inhibitory of F. oxysporum infectious activity, or permanently active. On the other hand, the combination of these multiple mechanisms most probably exerts a permanent pressure that only allows the pathogen to survive; F. oxysporum is still present in Châteaurenard soil, it has not been eradicated. However, due to the multi-factorial nature of this pressure linked to the diversity of mechanisms affecting its metabolism, F. oxysporum has not as yet developed means to circumvent this pressure, despite the remarkable adaptability that it is otherwise able to express in various abiotic conditions (Steinberg et al., 2016). This could explain why its pathogenic activity remains controlled in the suppressive soil of Châteaurenard.
Taxa were also found in studies aimed at identifying bacteria associated with soil suppressiveness to Fusarium wilt (Shen et al., 2014, 2015) or with agricultural practices reducing the infectious activity of pathogenic F. oxysporum (Klein et al., 2013; Naing et al., 2015). This was particularly the case of Paenibacillus, Arthrobacter, and Rhizobium. The co-occurrence of these bacteria with disease suppression does not prove that they are necessarily involved in the control of pathogenic F. oxysporum. However, hypotheses can be advanced about potential roles they may have since bacteria of the genera highlighted in our analysis have been described for their ability to produce metabolites that can directly or indirectly affect the metabolism of F. oxysporum. Thus, bacteria of the Paenibacillus genus produce secondary metabolites such as fusaricidins A-D which are specifically toxic to certain fungi including F. oxysporum (Mousa and Raizada, 2015). Actinobacteria are the most prolific production source of bioactive metabolites, among which various polyketides– antifungal compounds–thanks to many secondary metabolite gene clusters in their genomes (Hamedi and Mohammadipanah, 2015). By the way, they are more abundant in a soil suppressive to Rhizoctonia damping-off of sugarbeet than in conducive soils (Mendes et al., 2011). Streptomyces spp. strains produce molecules that inhibit F. oxysporum development (Cha et al., 2016). The co-occurrence of Rhizobium with disease suppression was a priori more difficult to explain. However, Rhizobium isolates inhibited F. oxysporum f. sp. ciceris growth by producing volatiles, mainly cyanides. Although they did not totally control the pathogen in the field, they contributed to decreasing disease incidence (Arfaoui et al., 2006). It may seem strange that OTUs referring to the Pseudomonas genus should not be included in the list of bacterial taxa specifically present in the suppressive soil of Châteaurenard, while Lemanceau and Alabouvette (1993) and Mazurier et al. (2009) isolated fluorescent Pseudomonas and showed that these strains indirectly reinforced trophic competition between pathogenic and non-pathogenic F. oxysporum by reducing Fe availability in the rhizosphere or producing antifungal phenazines. However, the Pseudomonas genus is ubiquitous and was indeed detected in all soils, but given their discrimination level due to the length of obtained reads, OTUs did not distinguish the Pseudomonas strains directly involved.
The suppressive character of Châteaurenard soil is not only due to the presence of non-pathogenic F. oxysporum and Pseudomonas fluorescens producing siderophores even though the role of these microorganisms was previously highlighted a lot (Alabouvette, 1986; Lemanceau et al., 1993). Since this suppressive character has been observed for years, that it is stable, it results more likely from a combination of complementary mechanisms of fungal and bacterial origin, besides the role of the abiotic components (pH, clays type) which, although it has not been approached here, also participate in the microbiological functioning of the soil (Höper et al., 1995; Almario et al., 2013a,b). Thus, the sum of the mechanisms expressed concurrently or successively would constrain F. oxysporum development and explain the suppressive nature of Châteaurenard soil. F. oxysporum survived but did not express its infectious activity. This also explains why Châteaurenard soil is suppressive to Fusarium wilts and not to diseases caused by pathogens with different ecological requirements. These mechanisms are driven by consortiums that include microorganisms directly involved in F. oxysporum control; but they are also very likely driven by microorganisms rather believed to act on compatibility and communication between these biocontrol agents, which structure the community assemblage and give the consortium a synergistic value. Analyzing co-occurrence networks within suppressive and conducive soils microbial communities would test such hypothesis (Faust et al., 2015; Karimi et al., 2017).
The taxa revealed by the OTUs are generally presented as genera, but they more often correspond to a sum of individuals whose biological functionality or role may not be detectable at the genus level, but rather at the strain level (Mendes et al., 2011). It is therefore difficult to assign a direct or indirect antagonistic activity to these OTUs against F. oxysporum. Nevertheless, we found them associated with suppressive soil, and they were associated with Fusarium disease decline in other studies. Consequently, we may believe that certain strains from these taxa are actually involved in the mechanisms of soil suppressiveness. To confirm this hypothesis, it would be necessary to isolate these microorganisms by conventional methods, insofar as Pasteurian methods allow for it, which is not always the case. However, intrageneric or even intraspecific diversity is such within these taxa that the number of isolated microorganisms should be exhaustive to detect the bioactive strain(s) actually involved in the control of pathogenic F. oxysporum. Additionally, there is no evidence that any given strain taken separately can exert its antagonistic activity in the same way as when it is within the microbial community. On the contrary, a better efficiency of the biological control agents is obtained when microbial consortia and not single strains are used to control pathogenic fungi in soils and substrates (Danon et al., 2010; Jain et al., 2012; Pertot et al., 2017). Thus, it is becoming clearer and clearer that soil functionality depends on the community pattern and that the activity of the microorganisms directly involved in the targeted function is only possible in the presence of the different components of this community, without these components being directly involved in the function of interest (Tyc et al., 2014; Williams et al., 2014; Chao et al., 2016). Therefore, an alternative strategy to identify the members of the microbial consortium responsible for soil suppressiveness would be to erode diversity by performing serial dilutions of the suppressive soil until it loses its suppressiveness. The dilution level preceding the loss of suppressiveness would contain the minimum consortium necessary for pathogenic F. oxysporum control. This erosion of diversity and/or destructuring of the community pattern may well be responsible for the loss of the suppressive character of soil plots in the Châteaurenard region. Actually, the soil is suppressive to Fusarium wilts but not to other soil-borne diseases. In market gardening, producers use disinfection methods (fumigation or/and steam) to control other soil-borne diseases such as damping-off during rotations (Navarrete et al., 2006). Consequently, the repeated use of these practices strongly and more or less permanently disrupted the microbial balance to reach a similar situation to the one in our comparative study. Other farming practices such as intensive monoculture also led to the accumulation of pathogens and reduced the number and/or activity of beneficial bacteria, with a loss of soil suppressiveness to cotton Fusarium wilt (Li et al., 2015). Conversely, modifying the structure of microbial soil communities through agricultural practices can make soils temporarily suppressive to one disease or another (Westphal and Becker, 2001; Mazzola, 2007; Klein et al., 2013; Raaijmakers and Mazzola, 2016; Vida et al., 2016). Unfortunately the sustainability of this suppressiveness seems difficult to acquire as durably as in Châteaurenard soil. It is therefore likely that the balance reached by Châteaurenard soil microbial communities results from a long natural evolutionary process; the suppressiveness of this soil is called “native,” this is the reason why it is so stable. In the cases of “acquired” soil suppressiveness, the agricultural practices used to manage microbial community patterns are still too recent to fix their assemblage (structure) in a sustainable way, but all the results reported in the literature show that this is an alternative to be favored.
Author Contributions
The project was supervised by CS. KS-H, VE-H, and EC: carried out the experiments and acquired the data; VE-H and ST: carried out bioinformatics analyzes; VE-H, ST, JR, and CS: animated scientific discussions over the duration of the project and contributed equally to the writing of the article.
Funding
This work was supported by a grant to KS-H from the Conseil Régional de Bourgogne (CRB 2011-9201AAO048S05563). It was also supported by the European Commission within the framework of the EcoFINDERS project (FP7-264465) and the Plant Health and Environment department (INRA http://www.spe.inra.fr/en). The funders had no role in study design, data collection and analysis, decision to publish, or preparation of the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors wish to thank the colleagues of the Genosol platform (https://www2.dijon.inra.fr/plateforme_genosol/en/homepage) for technical discussions and relevant advice regarding the metabarcoding approach. Thanks also to Mrs. Buchwalter for language edition of this manuscript.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.00568/full#supplementary-material
References
Acosta-Martínez, V., Acosta-Mercado, D., Sotomayor-Ramírez, D., and Cruz-Rodríguez, L. (2008). Microbial communities and enzymatic activities under different management in semiarid soils. Appl. Soil Ecol. 38, 249–260. doi: 10.1016/j.apsoil.2007.10.012
Alabouvette, C. (1986). Fusarium-wilt suppressive soils from the Châteaurenard region: review of a 10-year study. Agronomie 6, 273–284. doi: 10.1051/agro:19860307
Alabouvette, C. (1999). Fusarium wilt suppressive soils: an example of disease-suppressive soils. Australas. Plant Pathol. 28, 57–64. doi: 10.1071/AP99008
Alabouvette, C., Olivain, C., Migheli, Q., and Steinberg, C. (2009). Microbiological control of soil-borne phytopathogenic fungi with special emphasis on wilt-inducing Fusarium oxysporum. New Phytol. 184, 529–544. doi: 10.1111/j.1469-8137.2009.03014.x
Al-Ani, R. A., Adhab, M. A., Mahdi, M. H., and Abood, H. M. (2012). Rhizobium japonicum as a biocontrol agent of soybean root rot disease caused by Fusarium solani and Macrophomina phaseolina. Plant Protect. Sci. 48, 149–155. doi: 10.17221/16/2012-PPS
Almario, J., Kyselková, M., Kopecky, J., Sagova-Mareckova, M., Muller, D., Grundmann, G. L., et al. (2013a). Assessment of the relationship between geologic origin of soil, rhizobacterial community composition and soil receptivity to tobacco black root rot in Savoie region (France). Plant Soil 371, 397–408. doi: 10.1007/s11104-013-1677-1
Almario, J., Prigent-Combaret, C., Muller, D., and Moënne-Loccoz, Y. (2013b). Effect of clay mineralogy on iron bioavailability and rhizosphere transcription of 2,4-diacetylphloroglucinol biosynthetic genes in biocontrol Pseudomonas protegens. Mol. Plant. Microbe Interact. 26, 566–574. doi: 10.1094/MPMI-11-12-0274-R
Alstrom, S. (2000). Root-colonizing fungi from oilseed rape and their inhibition of Verticillium dahliae. J. Phytopathol. 148, 417–423. doi: 10.1046/j.1439-0434.2000.00511.x
Altschul, S. F., Madden, T. L., Schäffer, A. A., Zhang, J., Zhang, Z., Miller, W., et al. (1997). Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25, 3389–3402. doi: 10.1093/nar/25.17.3389
Amir, H., and Alabouvette, C. (1993). Involvement of soil abiotic factors in the mechanisms of soil suppressiveness to fusarium wilts. Soil Biol. Biochem. 25, 157–164. doi: 10.1016/0038-0717(93)90022-4
Arfaoui, A., Sifi, B., Boudabous, A., El Hadrami, I., and Cherif, M. (2006). Identification of Rhizobium isolates possessing antagonistic activity against Fusarium oxysporum f.sp ciceris, the causal agent of Fusarium wilt of chickpea. J. Plant Pathol. 88, 67–75. doi: 10.4454/jpp.v88i1.832
Baker, K. F., and Cook, R. J. (1974). Biological Control of Plant Pathogens. St. Paul, MN: The American Phytopathological Society. 433.
Borges, A. V., Saraiva, R. M., and Maffia, L. A. (2015). Biocontrol of gray mold in tomato plants by Clonostachys rosea. Trop. Plant Pathol. 40, 71–76. doi: 10.1007/s40858-015-0010-3
Buée, M., Reich, M., Murat, C., Morin, E., Nilsson, R. H., Uroz, S., et al. (2009). 454 Pyrosequencing analyses of forest soils reveal an unexpectedly high fungal diversity. New Phytol. 184, 449–456. doi: 10.1111/j.1469-8137.2009.03003.x
Cha, J. Y., Han, S., Hong, H. J., Cho, H., Kim, D., Kwon, Y., et al. (2016). Microbial and biochemical basis of a Fusarium wilt-suppressive soil. ISME J. 10, 119–129. doi: 10.1038/ismej.2015.95
Chao, Y. Q., Liu, W. S., Chen, Y. M., Chen, W. H., Zhao, L. H., Ding, Q. B., et al. (2016). Structure, variation, and co-occurrence of soil microbial communities in abandoned sites of a rare earth elements mine. Environ. Sci. Technol. 50, 11481–11490. doi: 10.1021/acs.est.6b02284
Chaparro, J. M., Sheflin, A. M., Manter, D. K., and Vivanco, J. M. (2012). Manipulating the soil microbiome to increase soil health and plant fertility. Biol. Fertil. Soils 48, 489–499. doi: 10.1007/s00374-012-0691-4
Chatterton, S., and Punja, Z. K. (2009). Chitinase and beta-1,3-glucanase enzyme production by the mycoparasite Clonostachys rosea f. catenulata against fungal plant pathogens. Can. J. Microbiol. 55, 356–367. doi: 10.1139/W08-156
Cole, J. R., Wang, Q., Cardenas, E., Fish, J., Chai, B., Farris, R. J., et al. (2009). The ribosomal database Project: improved alignments and new tools for rRNA analysis. Nucleic Acids Res. 37, D141–D145. doi: 10.1093/nar/gkn879
Daguerre, Y., Edel-Hermann, V., and Steinberg, C. (2016). “Fungal genes and metabolites associated with the biocontrol of soil-borne plant pathogenic fungi,” in Fungal Metabolites, eds J.-M. Mérillon, and K. G. Ramawat (Basel: Springer International Publishing), 1–72.
Danon, M., Chen, Y., and Hadar, Y. (2010). Ascomycete communities associated with suppression of Sclerotium rolfsii in compost. Fungal Ecol. 3, 20–30. doi: 10.1016/j.funeco.2009.05.003
Dawidziuk, A., Koczyk, G., and Popiel, D. (2016). Adaptation and response to mycotoxin presence in pathogen-pathogen interactions within the Fusarium genus. World Mycotoxin J. 9, 565–575. doi: 10.3920/WMJ2015.2010
De Cal, A., Garcia-Lepe, R., and Melgarejo, P. (2000). Induced resistance by Penicillium oxalicum against Fusarium oxysporum f. sp lycopersici: histological studies of infected and induced tomato stems. Phytopathology 90, 260–268. doi: 10.1094/PHYTO.2000.90.3.260
Dubey, M. K., Jensen, D. F., and Karlsson, M. (2014). Hydrophobins are required for conidial hydrophobicity and plant root colonization in the fungal biocontrol agent Clonostachys rosea. BMC Microbiol. 14:14. doi: 10.1186/1471-2180-14-18
Duijff, B. J., Recorbet, G., Bakker, P., Loper, J. E., and Lemanceau, P. (1999). Microbial antagonism at the root level is involved in the suppression of Fusarium wilt by the combination of nonpathogenic Fusarium oxysporum Fo47 and Pseudomonas putida WCS358. Phytopathology 89, 1073–1079. doi: 10.1094/PHYTO.1999.89.11.1073
Edgar, R. C. (2010). Search and clustering orders of magnitude faster than BLAST. Bioinformatics 26, 2460–2461. doi: 10.1093/bioinformatics/btq461
Elhalag, K. M., Emara, H. M., Messiha, N. A. S., Elhadad, S. A., and Abdallah, S. A. (2015). The relation of different crop roots exudates to the survival and suppressive effect of Stenotrophomonas maltophilia (PD4560), biocontrol agent of bacterial xilt of potato. J. Phytopathol. 163, 829–840. doi: 10.1111/jph.12381
Faust, K., Lima-Mendez, G., Lerat, J. S., Sathirapongsasuti, J. F., Knight, R., Huttenhower, C., et al. (2015). Cross-biome comparison of microbial association networks. Front. Microbiol. 6:1200. doi: 10.3389/fmicb.2015.01200
Garbeva, P., van Veen, J. A., and van Elsas, J. D. (2004). Microbial diversity in soil: selection of microbial populations by plant and soil type and implications for disease suppressiveness. Annu. Rev. Phytopathol. 42, 243–270. doi: 10.1146/annurev.phyto.42.012604.135455
Gardes, M., and Bruns, T. D. (1993). ITS primers with enhanced specificity for basidiomycetes - application to the identification of mycorrhizae and rusts. Mol. Ecol. 2, 113–118. doi: 10.1111/j.1365-294X.1993.tb00005.x
Gómez Expósito, R., de Bruijn, I., Postma, J., and Raaijmakers, J. M. (2017). Current insights into the role of rhizosphere bacteria in disease suppressive soils. Front. Microbiol. 8:2529. doi: 10.3389/fmicb.2017.02529
Halfeld-Vieira, B. D., da Silva, W. L. M., Schurt, D. A., Ishida, A. K. N., de Souza, G. R., and Nechet, K. D. (2015). Understanding the mechanism of biological control of passionfruit bacterial blight promoted by autochthonous phylloplane bacteria. Biol. Control 80, 40–49. doi: 10.1016/j.biocontrol.2014.09.011
Hamedi, J., and Mohammadipanah, F. (2015). Biotechnological application and taxonomical distribution of plant growth promoting actinobacteria. J. Ind. Microbiol. Biotechnol. 42, 157–171. doi: 10.1007/s10295-014-1537-x
Hartmann, M., Howes, C. G., Vaninsberghe, D., Yu, H., Bachar, D., Christen, R., et al. (2012). Significant and persistent impact of timber harvesting on soil microbial communities in Northern coniferous forests. ISME J. 6, 2199–2218. doi: 10.1038/ismej.2012.84
Höper, H., Steinberg, C., and Alabouvette, C. (1995). Involvement of clay type and pH in the mechanisms of soil suppressiveness to fusarium wilt of flax. Soil Biol. Biochem. 27, 955–967. doi: 10.1016/0038-0717(94)00238-V
Huang, X. Q., Liu, L. L., Wen, T., Zhu, R., Zhang, J. B., and Cai, Z. C. (2015). Illumina MiSeq investigations on the changes of microbial community in the Fusarium oxysporum f.sp cubense infected soil during and after reductive soil disinfestation. Microbiol. Res. 181, 33–42. doi: 10.1016/j.micres.2015.08.004
Hung, P. M., Wattanachai, P., Kasem, S., and Poaim, S. (2015). Biological control of Phytophthora palmivora causing root rot of pomelo using Chaetomium spp. Mycobiology 43, 63–70. doi: 10.5941/MYCO.2015.43.1.63
Jain, A., Singh, S., Sarma, B. K., and Singh, H. B. (2012). Microbial consortium-mediated reprogramming of defence network in pea to enhance tolerance against Sclerotinia sclerotiorum. J. Appl. Microbiol. 112, 537–550. doi: 10.1111/j.1365-2672.2011.05220.x
Janvier, C., Villeneuve, F., Alabouvette, C., Edel-Hermann, V., Mateille, T., and Steinberg, C. (2007). Soil health through soil disease suppression: which strategy from descriptors to indicators? Soil Biol. Biochem. 39, 1–23. doi: 10.1016/j.soilbio.2006.07.001
Kaiserer, L., Oberparleiter, C., Weiler-Görz, R., Burgstaller, W., Leiter, E., and Marx, F. (2003). Characterization of the Penicillium chrysogenum antifungal protein PAF. Arch. Microbiol. 180, 204–210. doi: 10.1007/s00203-003-0578-8
Karimi, B., Maron, P. A., Boure, N. C. P., Bernard, N., Gilbert, D., and Ranjard, L. (2017). Microbial diversity and ecological networks as indicators of environmental quality. Environ. Chem. Lett. 15, 265–281. doi: 10.1007/s10311-017-0614-6
Kirk, P. M. (2017). “Species Fungorum (version Jan 2016),” in Species 2000 & ITIS Catalogue of Life, eds Y. Roskov, L. Abucay, T. Orrell, D. Nicolson, N. Bailly, P. M. Kirk, T. Bourgoin, R. E. DeWalt, W. Decock, A. De Wever, E. Nieukerken, J. van Zarucchi, and L. Penev (Leiden: Species 2000: Naturalis). Available online at: www.catalogueoflife.org/col (Accessed December 20, 2017).
Klein, E., Ofek, M., Katan, J., Minz, D., and Gamliel, A. (2013). Soil suppressiveness to fusarium disease: shifts in root microbiome associated with reduction of pathogen root colonization. Phytopathology 103, 23–33. doi: 10.1094/PHYTO-12-11-0349
Kohl, J., Lombaers, C., Moretti, A., Bandyopadhyay, R., Somma, S., and Kastelein, P. (2015). Analysis of microbial taxonomical groups present in maize stalks suppressive to colonization by toxigenic Fusarium spp.: a strategy for the identification of potential antagonists. Biol. Control 83, 20–28. doi: 10.1016/j.biocontrol.2014.12.007
Kõljalg, U., Nilsson, R. H., Abarenkov, K., Tedersoo, L., Taylor, A. F. S., Bahram, M., et al. (2013). Towards a Unified Paradigm for Sequence-Based Identification of Fungi. Mol. Ecol. 22, 5271–5277. doi: 10.1111/mec.12481
Kyselková, M., and Moënne-Loccoz, Y. (2012). “Pseudomonas and other microbes in disease-suppressive soils,” in Organic Fertilisation, Soil Quality and Human Health, ed E. Lichtfouse (Dordrecht: Springer Science+Business Media), 93–140.
Kyselková, M., Kopecky, J., Frapolli, M., Defago, G., Sagova-Mareckova, M., Grundmann, G. L., et al. (2009). Comparison of rhizobacterial community composition in soil suppressive or conducive to tobacco black root rot disease. ISME J. 3, 1127–1138. doi: 10.1038/ismej.2009.61
Larena, I., Sabuquillo, P., Melgarejo, P., and De Cal, A. (2003). Biocontrol of Fusarium and Verticillium wilt of tomato by Penicillium oxalicum under greenhouse and field conditions. J. Phytopathol. 151, 507–512. doi: 10.1046/j.1439-0434.2003.00762.x
Larkin, R. P. (2015). Soil health paradigms and implications for disease management. Annu. Rev. Phytopathol. 53, 199–221. doi: 10.1146/annurev-phyto-080614-120357
Lemanceau, P., and Alabouvette, C. (1993). Suppression of Fusarium wilts by fluorescent pseudomonads: mechanisms and applications. Biocontrol Sci. Technol. 3, 219–234. doi: 10.1080/09583159309355278
Lemanceau, P., Bakker, P., Jan de Kogel, W., Alabouvette, C., and Schippers, B. (1993). Antagonistic effect of nonpathogenic Fusarium oxysporum Fo47 and pseudobactin 358 upon pathogenic Fusarium oxysporum f. sp. dianthi. Appl. Environ. Microbiol. 59, 74–82.
Li, X. G., Zhang, Y. N., Ding, C. F., Jia, Z. J., He, Z. L., Zhang, T. L., et al. (2015). Declined soil suppressiveness to Fusarium oxysporum by rhizosphere microflora of cotton in soil sickness. Biol. Fertil. Soils 51, 935–946. doi: 10.1007/s00374-015-1038-8
Louvet, J., Rouxel, F., and Alabouvette, C. (1976). Recherches sur la résistance des sols aux maladies. I. Mise en évidence de la nature microbiologique de la résistance d'un sol au développement de la fusariose vasculaire du melon. Ann. Phytopathol. 8, 425–436.
Lundgren, J. G., and Fergen, J. K. (2011). Enhancing predation of a subterranean insect pest: a conservation benefit of winter vegetation in agroecosystems. Appl. Soil Ecol. 51, 9–16. doi: 10.1016/j.apsoil.2011.08.005
Ma, Y., Gentry, T., Hu, P., Pierson, E., Gu, M. M., and Yin, S. X. (2015). Impact of brassicaceous seed meals on the composition of the soil fungal community and the incidence of Fusarium wilt on chili pepper. Appl. Soil Ecol. 90, 41–48. doi: 10.1016/j.apsoil.2015.01.016
Mazurier, S., Corberand, T., Lemanceau, P., and Raaijmakers, J. M. (2009). Phenazine antibiotics produced by fluorescent pseudomonads contribute to natural soil suppressiveness to Fusarium wilt. ISME J. 3, 977–991. doi: 10.1038/ismej.2009.33
Mazzola, M. (2007). Manipulation of rhizosphere bacterial communities to induce suppressive soil. J. Nematol. 39, 213–220.
Mazzola, M., and Gu, Y. H. (2002). Wheat genotype-specific induction of soil microbial communities suppressive to disease incited by Rhizoctonia solani anastomosis group AG-5 and AG-8. Phytopathology 92, 1300–1307. doi: 10.1094/PHYTO.2002.92.12.1300
McQuilken, M. P., and Gemmell, J. (2004). Enzyme production by the mycoparasite Verticillium biguttatum against Rhizoctonia solani. Mycopathologia 157, 201–205. doi: 10.1023/B:MYCO.0000020590.20040.4a
Mendes, R., Kruijt, M., de Bruijn, I., Dekkers, E., van der Voort, M., Schneider, J. H. M., et al. (2011). Deciphering the rhizosphere microbiome for disease-suppressive bacteria. Science 332, 1097–1100. doi: 10.1126/science.1203980
Messiha, N. A. S., van Diepeningen, A. D., Farag, N. S., Abdallah, S. A., Janse, J. D., and van Bruggen, A. H. C. (2007). Stenotrophomonas maltophilia: a new potential biocontrol agent of Ralstonia solanacearum, causal agent of potato brown rot. Eur. J. Plant Pathol. 118, 211–225. doi: 10.1007/s10658-007-9136-6
Minerdi, D., Bossi, S., Gullino, M. L., and Garibaldi, A. (2009). Volatile organic compounds: a potential direct long-distance mechanism for antagonistic action of Fusarium oxysporum strain MSA35. Environ. Microbiol. 11, 844–854. doi: 10.1111/j.1462-2920.2008.01805.x
Mmbaga, M. T., Sauve, R. J., and Mrema, F. A. (2008). Identification of microorganisms for biological control of powdery mildew in Cornus florida. Biol. Control 44, 67–72. doi: 10.1016/j.biocontrol.2007.10.018
Moltzan, B. D., Blenis, P. V., and Hiratsuka, Y. (2001). Temporal occurrence and impact of Scytalidium uredinicola, a mycoparasite of western gall rust. Can. J. Plant Pathol. 23, 384–390. doi: 10.1080/07060660109506959
Moricca, S., Ragazzi, A., Mitchelson, K. R., and Assante, G. (2001). Antagonism of the two-needle pine stem rust fungi Cronartium flaccidum and Peridermium pini by Cladosporium tenuissimum in vitro and in planta. Phytopathology 91, 457–468. doi: 10.1094/PHYTO.2001.91.5.457
Mosquera-Espinosa, A. T., Bayman, P., Prado, G., Gómez-Carabal,í, A., Otero, J. T., Gomez-Carabali, A., et al. (2013). The double life of Ceratobasidium: orchid mycorrhizal fungi and their potential for biocontrol of Rhizoctonia solani sheath blight of rice. Mycologia 105, 141–150. doi: 10.3852/12-079
Mousa, W. K., and Raizada, M. N. (2015). Biodiversity of genes encoding anti-microbial traits within plant associated microbes. Front. Plant Sci. 6:231. doi: 10.3389/fpls.2015.00231
Naing, K. W., Nguyen, X. H., Anees, M., Lee, Y. S., Kim, Y. C., Kim, S. J., et al. (2015). Biocontrol of Fusarium wilt disease in tomato by Paenibacillus ehimensis KWN38. World J. Microbiol. Biotechnol. 31, 165–174. doi: 10.1007/s11274-014-1771-4
Navarrete, M., Le Bail, M., Papy, F., Bressoud, F., and Tordjman, S. (2006). Combining leeway on farm and supply basin scales to promote technical innovations in lettuce production. Agron. Sustain. Dev. 26, 77–87. doi: 10.1051/agro:2005062
Nicoletti, R., De Stefano, M., De Stefano, S., Trincone, A., and Marziano, F. (2004). Antagonism against Rhizoctonia solani and fungitoxic metabolite production by some Penicillium isolates. Mycopathologia 158, 465–474. doi: 10.1007/s11046-004-3712-5
Nilsson, R. H., Veldre, V., Hartmann, M., Unterseher, M., Amend, A., Bergsten, J., et al. (2010). An open source software package for automated extraction of ITS1 and ITS2 from fungal ITS sequences for use in high-throughput community assays and molecular ecology. Fungal Ecol. 3, 284–287. doi: 10.1016/j.funeco.2010.05.002
Oliveros, J. C. (2007). Venny. An Interactive Tool for Comparing Lists with Venn's diagrams. (Madrid: CNB-Csic), http://bioinfogp.cnb.csic.es/tools/venny/index.html
Park, J. H., Choi, G. J., Jang, K. S., Lim, H. K., Kim, H. T., Cho, K. Y., et al. (2005). Antifungal activity against plant pathogenic fungi of chaetoviridins isolated from Chaetomium globosum. FEMS Microbiol. Lett. 252, 309–313. doi: 10.1016/j.femsle.2005.09.013
Penton, C. R., Gupta, V., Tiedje, J. M., Neate, S. M., Ophel-Keller, K., Gillings, M., et al. (2014). Fungal community structure in disease suppressive soils assessed by 28S LSU gene sequencing. PLoS ONE 9:4. doi: 10.1371/journal.pone.0093893
Pertot, I., Giovannini, O., Benanchi, M., Caffi, T., Rossi, V., and Mugnai, L. (2017). Combining biocontrol agents with different mechanisms of action in a strategy to control Botrytis cinerea on grapevine. Crop Protect. 97, 85–93. doi: 10.1016/j.cropro.2017.01.010
Plassart, P., Terrat, S., Thomson, B., Griffiths, R., Dequiedt, S., Lelievre, M., et al. (2012). Evaluation of the ISO Standard 11063 DNA extraction procedure for assessing soil microbial abundance and community structure. PLoS ONE 7:e44279. doi: 10.1371/journal.pone.0044279
Popiel, D., Koczyk, G., Dawidziuk, A., Gromadzka, K., Blaszczyk, L., and Chelkowski, J. (2014). Zearalenone lactonohydrolase activity in Hypocreales and its evolutionary relationships within the epoxide hydrolase subset of a/b-hydrolases. BMC Microbiol. 14:82. doi: 10.1186/1471-2180-14-82
Pruesse, E., Peplies, J., and Glöckner, F. O. (2012). SINA: accurate high-throughput multiple sequence alignment of ribosomal RNA genes. Bioinformatics 28, 1823–1829. doi: 10.1093/bioinformatics/bts252
Quast, C., Pruesse, E., Yilmaz, P., Gerken, J., Schweer, T., Yarza, P., et al. (2013). The SILVA ribosomal RNA gene database project: improved data processing and web-based tools. Nucleic Acids Res. 41, D590–D596. doi: 10.1093/nar/gks1219
Raaijmakers, J. M., and Mazzola, M. (2016). Soil immune responses. Science 352, 1392–1393. doi: 10.1126/science.aaf3252
Raaijmakers, J. M., Paulitz, T. C., Alabouvette, C., Steinberg, C., and Moënne-Loccoz, Y. (2009). The rhizosphere: a playground and battlefield for soilborne pathogens and beneficial microorganisms. Plant Soil 321, 341–361. doi: 10.1007/s11104-008-9568-6
Schloss, P. D., Westcott, S. L., Ryabin, T., Hall, J. R., Hartmann, M., Hollister, E. B., et al. (2009). Introducing mothur: open-source, platform-independent community-supported software for describing and comparing microbial communities. Appl. Environ. Microbiol. 75, 7537–7541. doi: 10.1128/AEM.01541-09
Shen, Z. Z., Ruan, Y. Z., Xue, C., Zhong, S. T., Li, R., and Shen, Q. R. (2015). Soils naturally suppressive to banana Fusarium wilt disease harbor unique bacterial communities. Plant Soil 393, 21–33. doi: 10.1007/s11104-015-2474-9
Shen, Z. Z., Wang, D. S., Ruan, Y. Z., Xue, C., Zhang, J., Li, R., et al. (2014). Deep 16S rRNA pyrosequencing reveals a bacterial community associated with banana Fusarium wilt disease suppression induced by Bio-organic fertilizer application. PLoS ONE 9:e98420. doi: 10.1371/journal.pone.0098420
Soytong, K., Kanokmedhakul, S., Kukongviriyapa, V., and Isobe, M. (2001). Application of Chaetomium species (Ketomium®) as a new broad spectrum biological fungicide for plant disease control: a review article. Fungal Divers. 7, 1–15.
Steinberg, C., Lecomte, C., Alabouvette, C., and Edel-Hermann, V. (2016). “Root interactions with non-pathogenic Fusarium. Hey Fusarium oxysporum, what do you do in life when you do not infect a plant?,” in Belowground Defence Strategies in Plants, eds C. M. Vos and K. Kazan (Basel: Springer International Publishing Switzerland), 281–299.
Stirling, G. R., Smith, M. K., Smith, J. P., Stirling, A. M., and Hamill, S. D. (2012). Organic inputs, tillage and rotation practices influence soil health and suppressiveness to soilborne pests and pathogens of ginger. Australas. Plant Pathol. 41, 99–112. doi: 10.1007/s13313-011-0096-0
Suarez-Estrella, F., Arcos-Nievas, M. A., Lopez, M. J., Vargas-Garcia, M. C., and Moreno, J. (2013). Biological control of plant pathogens by microorganisms isolated from agro-industrial composts. Biol. Control 67, 509–515. doi: 10.1016/j.biocontrol.2013.10.008
Terrat, S., Christen, R., Dequiedt, S., Lelièvre, M., Nowak, V., Regnier, T., et al. (2012). Molecular biomass and metataxogenomic assessment of soil microbial communities as influenced by soil DNA extraction procedure. Microb. Biotechnol. 5, 135–141. doi: 10.1111/j.1751-7915.2011.00307.x
Terrat, S., Plassart, P., Bourgeois, E., Ferreira, S., Dequiedt, S., Adele-Dit-De-Renseville, N., et al. (2015). Meta-barcoded evaluation of the ISO standard 11063 DNA extraction procedure to characterize soil bacterial and fungal community diversity and composition. Microb. Biotechnol. 8, 131–142. doi: 10.1111/1751-7915.12162
Tian, T., Li, S. D., and Sun, M. H. (2014). Synergistic effect of dazomet soil fumigation and Clonostachys rosea against cucumber Fusarium wilt. Phytopathology 104, 1314–1321. doi: 10.1094/PHYTO-11-13-0297-R
Toussoun, T. A. (1975). “Fusarium-suppressive soils,” in Biology and control of soil-borne plant pathogens, ed G. W. Bruehl (St. Paul, MN: American Phytopathological Society), 145–151.
Tsror, L. (2010). Biology, epidemiology and management of Rhizoctonia solani on potato. J. Phytopathol. 158, 649–658. doi: 10.1111/j.1439-0434.2010.01671.x
Tyc, O., van den Berg, M., Gerards, S., van Veen, J. A., Raaijmakers, J. M., de Boer, W., et al. (2014). Impact of interspecific interactions on antimicrobial activity among soil bacteria. Front. Microbiol. 5:567. doi: 10.3389/fmicb.2014.00567
van Bruggen, A. H. C., Sharma, K., Kaku, E., Karfopoulos, S., Zelenev, V. V., and Blok, W. J. (2015). Soil health indicators and Fusarium wilt suppression in organically and conventionally managed greenhouse soils. Appl. Soil Ecol. 86(Suppl. C), 192–201. doi: 10.1016/j.apsoil.2014.10.014
Vida, C., Bonilla, N., Vicente, A. D., and Cazorla, F. M. (2016). Microbial profiling of a suppressiveness-induced agricultural soil amended with composted almond shells. Front. Microbiol. 7:4. doi: 10.3389/fmicb.2016.00004
Vujanovic, V., Mavragani, D., and Hamel, C. (2012). Fungal communities associated with durum wheat production system: a characterization by growth stage, plant organ and preceding crop. Crop Prot. 37, 26–34. doi: 10.1016/j.cropro.2012.02.006
Wang, Y., Xu, L., Ren, W., Zhao, D., Zhu, Y., and Wu, X. (2012). Bioactive metabolites from Chaetomium globosum L18, an endophytic fungus in the medicinal plant Curcuma wenyujin. Phytomedicine 19, 364–368. doi: 10.1016/j.phymed.2011.10.011
Weller, D. M., Raaijmakers, J. M., Gardener, B. B. M., and Thomashow, L. S. (2002). Microbial populations responsible for specific soil suppressiveness to plant pathogens. Annu. Rev. Phytopathol. 40, 309–348. doi: 10.1146/annurev.phyto.40.030402.110010
Westphal, A., and Becker, J. O. (2001). Soil suppressiveness to Heterodera schachtii under different cropping sequences. Nematology 3, 551–558. doi: 10.1163/156854101753389167
White, T. J., Bruns, T., Lee, S., and Taylor, J. (1990). “Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics,” in PCR Protocols. A Guide to Methods and Applications, eds M. A. Innis, D. H. Gelfand, J. J. Sninsky, and T. J. White (San Diego, CA: Academic Press), 315–322.
Williams, R. J., Howe, A., and Hofmockel, K. S. (2014). Demonstrating microbial co-occurrence pattern analyses within and between ecosystems. Front. Microbiol. 5:358. doi: 10.3389/fmicb.2014.00358
Zancarini, A., Mougel, C., Terrat, S., Salon, C., and Munier-Jolain, N. (2013). Combining ecophysiological and microbial ecological approaches to study the relationship between Medicago truncatula genotypes and their associated rhizosphere bacterial communities. Plant Soil 365, 183–199. doi: 10.1007/s11104-012-1364-7
Zhang, G. Z., Wang, F. T., Qin, J. C., Wang, D., Zhang, J. Y., Zhang, Y. H., et al. (2013). Efficacy assessment of antifungal metabolites from Chaetomium globosum No.05, a new biocontrol agent, against Setosphaeria turcica. Biol. Control 64, 90–98. doi: 10.1016/j.biocontrol.2012.10.005
Zhao, S., Liu, D. Y., Ling, N., Chen, F. D., Fang, W. M., and Shen, Q. R. (2014). Bio-organic fertilizer application significantly reduces the Fusarium oxysporum population and alters the composition of fungi communities of watermelon Fusarium wilt rhizosphere soil. Biol. Fertil. Soils 50, 765–774. doi: 10.1007/s00374-014-0898-7
Keywords: bacterial community, fungal community, diversity, metabarcoding, 454 pyrosequencing, biocontrol agent
Citation: Siegel-Hertz K, Edel-Hermann V, Chapelle E, Terrat S, Raaijmakers JM and Steinberg C (2018) Comparative Microbiome Analysis of a Fusarium Wilt Suppressive Soil and a Fusarium Wilt Conducive Soil From the Châteaurenard Region. Front. Microbiol. 9:568. doi: 10.3389/fmicb.2018.00568
Received: 11 October 2017; Accepted: 13 March 2018;
Published: 04 April 2018.
Edited by:
Choong-Min Ryu, Korea Research Institute of Bioscience and Biotechnology (KRIBB), South KoreaReviewed by:
Jordan Vacheron, Université de Lausanne, SwitzerlandYoun-Sig Kwak, Gyeongsang National University, South Korea
Copyright © 2018 Siegel-Hertz, Edel-Hermann, Chapelle, Terrat, Raaijmakers and Steinberg. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Christian Steinberg, Y2hyaXN0aWFuLnN0ZWluYmVyZ0BpbnJhLmZy