
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 13 February 2018
Sec. Infectious Agents and Disease
Volume 9 - 2018 | https://doi.org/10.3389/fmicb.2018.00212
This article is part of the Research Topic Biofilms: Current Challenges and Control Alternatives View all 29 articles
 Lvqin He1†
Lvqin He1† Ke Dai1†
Ke Dai1† Xintian Wen1Lingqiang Ding1Sanjie Cao1,2Xiaobo Huang1
Xintian Wen1Lingqiang Ding1Sanjie Cao1,2Xiaobo Huang1 Rui Wu1Qin Zhao1Yong Huang1Qigui Yan1Xiaoping Ma1Xinfeng Han1Yiping Wen1*
Rui Wu1Qin Zhao1Yong Huang1Qigui Yan1Xiaoping Ma1Xinfeng Han1Yiping Wen1*Haemophilus parasuis is known as a commensal organism discovered in the upper respiratory tract of swine where the pathogenic bacteria survive in various adverse environmental stress. QseC, a histidine protein kinase of the two-component regulatory systems CheY/QseC, is involved in the environmental adaptation in bacteria. To investigate the role of QseC in coping with the adverse environment stresses and survive in the host, we constructed a qseC mutant of H. parasuis serovar 13 strain (ΔqseC), MY1902. In this study, we found that QseC was involved in stress tolerance of H. parasuis, by the ΔqseC exhibited a decreased resistance to osmotic pressure, oxidative stress, and heat shock. Moreover, the ΔqseC weakened the ability to take up iron and biofilm formation. We also found that the QseC participate in sensing the epinephrine in environment to regulate the density of H. parasuis.
Haemophilus parasuis is a causative agent of Glässer's disease what characterized by fibrinous polyserositis, polyarthritis, and meningitis (Cai et al., 2005). H. parasuis, one of the major causes of nursery mortality in swine herds, giving rise to great economic loss in pig farms (Oliveira and Pijoan, 2004). After invading the respiratory tract and lung tissue, H. parasuis exposes to stress conditions, such as osmotic pressure, oxidative stress, and temperature. These stresses may affect bacterial survival, and result in protein denaturation and misfolding (Frees et al., 2004). H. parasuis regulates a variety of gene expression to survive these stress conditions (Huang et al., 2016). However, bacterial survival and establishment of infection require to sense and accurately to environmental cues. Two component regulatory systems are important for bacterial to adapt in the environment (Stock et al., 2000).
Two-component signal transduction system (TCSTS) consists of a sensor histidine kinase (HK) and a response regulator(RR) (Wuichet et al., 2010). QseBC, a two-component-based quorum sensing (QS) system that responds to hormone's signals AI-3, epinephrine, and norepinephrine in Escherichia coli and Salmonella (Sperandio et al., 2003; Clarke et al., 2006; Walters and Sperandio, 2006; Kalia, 2013). Enterohemorrhagic E. coli through QseC to senses AI-3, epinephrine and norepinephrine to activate flagella and motility, AE lesion formation and Shiga toxin expression (Hughes et al., 2009).
QseC sensory kinase is a bacterial adrenergic receptor that is crucial for interkingdom signaling in E. coli (Clarke et al., 2006). QseC, as a transmembrane protein with histidine protein kinase, is activated in response to host and bacterial signals, and phosphorylates the QseB response regulator, a transcription factor that regulates relevant virulence gene expression (Walters and Sperandio, 2006; Hughes et al., 2009)1. QseC can control QseB activation via a mechanism that is independent of reverse phosphotransfer. QseC-mediated dephosphorylation is required for maintaining proper QseB-PmrB-PmrA interactions in Uropathogenic E. coli (Breland et al., 2017). QseC controls biofilm formation in non-typeable Haemophilus influenza (Unal et al., 2012). Moreover, the ΔqseC diminished motility and colonization of the gastrointestinal tract compared to the wild-type parent strain in Salmonella enterica serovar Typhimurium (S. Typhimurium) (Bearson and Bearson, 2008). QseC also involved in flagellar motility, fimbrial hemagglutination, and intracellular virulence in fish pathogen Edwardsiella tarda (Wang et al., 2011).
TCSTS of bacteria are considered to form an intricate signal network to detect changes in environmental and respond by adjusting various cellular functions (Li et al., 2002; Eguchi et al., 2007)2,3. However, whether the QseC plays a role in the H. parasuis adapt to environmental changes in pigs are still unknown.
In the present study, we investigated the responses of ΔqseC to the stress conditions and biological characteristics which constructed by the natural transformation system (Bigas et al., 2005). In addition to affect biofilm formation as reported in E. coli and Salmonella, we found that QseC function in bacterial response to a variety of stimuli such as osmotic pressure, oxidative stress, and heat shock.
The bacterial strains and plasmids used in this study were listed in Table 1. E. coli DH5a and E. coli BL21 were cultured in Luria–Bertani medium at 37°C. H. parasuis serovar 13 strain, MY1902 was grown in tryptic soy broth (TSB) medium or cultivated on tryptic soy agar (TSA) (Difco, Detroit, USA) supplemented with 0.001% (w/v) nicotinamide adenine dinucleotide (NAD) (Sigma Aldrich, Missouri, USA) and 5% (v/v) inactivated bovine serum at 37°C. When necessary, the media were supplemented with 50 μL kanamycin (100 mg/mL) or 100 μL ampicillin (100 mg/mL).
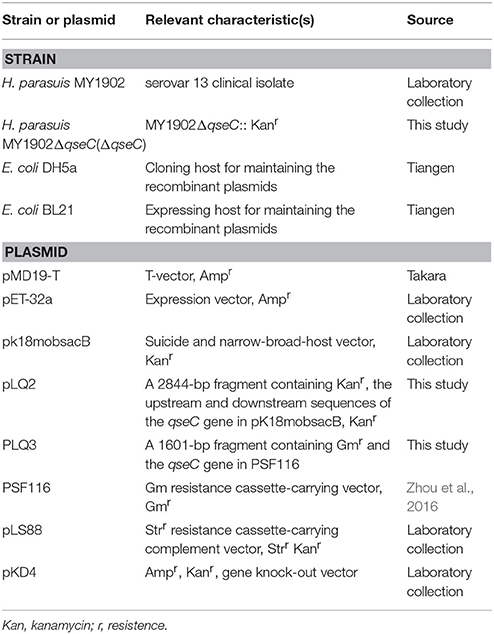
Table 1. Bacterial strains and plasmids used in this study.
The primers used to construct the qseC mutant are listed in Table 2. The 964-bp upstream and 945-bp downstream fragments of qseC were amplified from the genome of MY1902 using primers qseC-Up-F/R and qseC-Down-F/R, A kanamycin resistant (kanR) cassette(935 bp) was amplified from pKD4 using primers Kan-F/R. These three PCR fragments were combined by overlap PCR with primers qseC-up-F and qseC–down-R. Then the overlapped product was cloned into pK18mobsacB at BamHI and HindIII to construct the recombinant plasmid pLQ2. The recombinant plasmid pLQ2 was mobilized into H. parasuis strain MY1902 by natural transformations (Zhang et al., 2012b). To construct the complementing Plasmid pLQ3, the qseC gene was amplified from MY1902 using primers qseC-Comp-F/R and cloned into KpnI and BamHI digested pSF116 (Zhou et al., 2016). Both two DNA fragments (upstream homologous arm and down homologous arm) contained a 9-bp core DNA uptake signal sequence (USS) of 5′- ACCGCTTGT−3′ (Zhang et al., 2012a).
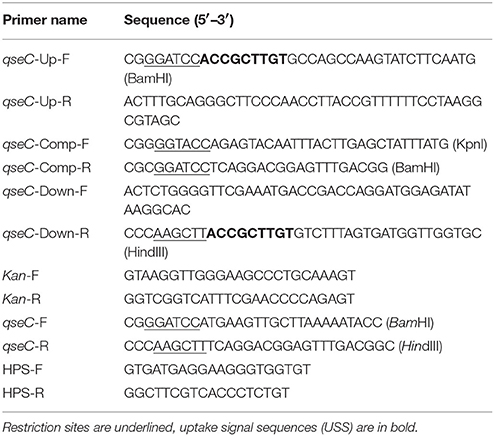
Table 2. Primers used in this study.
The RNAs of MY1902, ΔqseC, and C-ΔqseC were extracted using the Bacterial RNA Kit (5) according to the instructions(OMEGA R6950-00, America). RT-PCR using a PrimeScript™ RT reagent Kit (Perfect Real Time) according to the instructions (TaKaRa, Japan). The cDNA synthesis of both wild strain MY1902 and ΔqseC was detected with primers HPS-F/R, Kan-F/R, and qseC-F/R (Table 2). Western blotting assay was performed as described previously (Wang et al., 2013). 1 ml overnight bacterial cultures of the wild strain MY1902 and the ΔqseC were harvested by centrifugation for 1 min at 12000 rpm/min. Then resuspended with 40 μL ultrapure water, followed by adding 10 μL five-fold protein loading buffer, then boil for 10 min and ice bath for 2 min. The samples (10 ml) were electrophoresed on 12% SDS-PAGE gel and transferred onto Nitrocellulose(NC) membranes. The proteins on membrane were detected with Clarity™ Western ECL Substrate kit according to the instructions (BIO-RAD, America).
Stress resistance assays were performed as the previously described methods (Wong et al., 2007; Allen and Schmitt, 2009; Liu et al., 2013; Xie et al., 2013, 2016; Nasrallah et al., 2014; Huang et al., 2016). Fifty microliters of overnight cultures of MY1902, ΔqseC, and C-ΔqseC were subcultured at a dilution of 1:100 into 5 mL fresh TSB with 5% inactivated bovine serum and 0.01% NAD and the cells were grown at 37°C with 220 rpm. For the osmotic tolerance assay, the cells were cultivated on 40, 60, 80, and 100 mM NaCl TSA respectively. For the oxidative stress tolerance assay, the bacterial suspension was treated with 0.5, 1, 2, 4, 8, 16 mM H2O2 for 30 min respectively. For the heat-shock assay, bacterial cultures were placed in a 39, 42, and 45°C water bath for 30 min respectively and plated on TSA plates. Stress resistance was calculated as [(stressed sample CFU mL−1)/(control sample CFU mL−1)] × 100. The experiments were carried out independently three times.
Growth curve of the low-iron environment assay was performed as the previously described (Deslandes et al., 2007; Xie et al., 2013)4. Briefly, 1 mL overnight cultures of MY1902, ΔqseC, and C-ΔqseC were subcultured at a dilution of 1:100 into 100 mL fresh TSB with 100 μM, 200 μM EDDHA (ethylenediamine di(o-hydroxyphenylacetic) acid), a concentration sufficient to cause iron restriction (Beddek et al., 2004; Deslandes et al., 2007), or 33.33 μM FeSO4, 66.66 μM FeSO4 respectively, OD600 values were measured every hour. The experiments were carried out in triplicate independently.
Biofilm formation ability was measured as described previously with some modifies (Kaplan and Mulks, 2005; Tremblay et al., 2013; Xie et al., 2016). Twenty microliters overnight cultures of MY1902, ΔqseC, and C-ΔqseC were subcultured at a dilution of 1:100 into fresh TSB in 6-well tissue culture plate for 24, 48, 72, 96 h at 37°C respectively. Biofilms were washed with water and stained with 1 ml of 0.1% crystal violet for30 min. Excess staining was rinsed off under water, drying, and 100 μL of 33% (v/v) acetic acid was added to each well, then transferred to 96-well polystyrenemicrotiter plates and measured at wavelength of 595 nm. All tests were repeated independently times.
Confocal laser scanning microscopy (CLSM) assay was performed as previously described (Tremblay et al., 2013). Thirty microliters overnight cultures of MY1902, ΔqseC, and C-ΔqseC were subcultured were diluted 1:100 in the following media: fresh TSB, TSB with 50 μM epinephrine both supplemented with 5% inactivated bovine serum and 0.01% NAD in the six well microtiter plate with 20 × 20 mm cell climbing tablets for 18 h. The cell climbing tablets were removed and washed three times with phosphate buffer saline and stained with (LIVE/DEAD@ BacLight ™ Bacterial Viability kits, Invitrogen; live bacteria stain fluorescent green, whereas dead bacteria stain fluorescent red). The plates were incubated at room temperature in the dark for 20 min and washed three times with phosphate buffer saline. SYTO 9 was excitation (with an Ar laser) at 488 nm, and propidium iodide was excitation (with a HeNe laser) at 559 nm using Nikon AIR confocal scanning laser microscope (CLSM). The images were analyzed with the NIS-Elements AR software.
QseC is a chromosomally encoded polypeptide with 466 amino acids. In order to determine the similarity between QseC, we aligned the protein sequences of QseC from H. parasuis (SH0165) with Actinobacillus pleuropneumoniae (L20), H. influenzae (Rd KW20), S. enterica subsp (enterica serovar Typhimurium str. LT2), and E. coli (str. K-12 substr. MG1655). The multiple-sequence alignment revealed 60.71% identity (Figure 1).

Figure 1. Sequence alignment of H. parasuis QseC (HpQseC) against A. pleuropneumoniae QseC(ApQseC), H. influenzae QseC (HiQseC), Salmonella QseC (SaQseC), and E. coli QseC (EcQseC).
In this study, we constructed the qseC mutant (ΔqseC) and complementary strain (C-ΔqseC) of H. parasuis serovar 13 strain, MY1902 (Figure 2 in Supplementary Material). And the result of reverse transcription PCR(RT-PCR) verificated the H. parasuis ΔqseC was successfully constructed (Figure 3 in Supplementary Material). Additionally, the result of western blot showed that the expression of a protein of ~69.08 kDa was absent in the ΔqseC compared with the wild-type strain MY1902 (Figure 4 in Supplementary Material). These results indicated that the qseC gene have been knocked out from the genome of the wild-type strain MY1902.
Compared with the wild strain MY1902, we found that the ΔqseC didn't exhibit obvious growth defects compared with the wild strain (Figure 2).
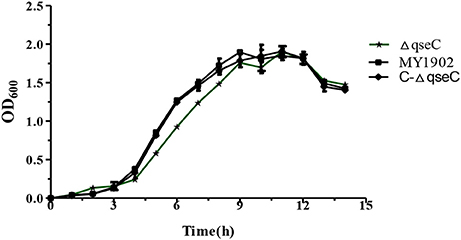
Figure 2. Growth of the wild strain MY1902, ΔqseC and C-ΔqseC in TSB supplemented with 5% inactivated bovine serum and 0.01% NAD. Error bars represent the standard deviations of three independent experiments.
We investigated various stress conditions included osmotic pressure, oxidative stress, and heat shock of the wild strain, ΔqseC and C-ΔqseC. When bacterial were treated with 40, 60, 80, and 100 mM NaCl TSA, the survival rate of ΔqseC was 32, 27.07, 1.88%, 0, which much lower than the wild strain with 85.13, 87.22, 60.88, 42.1% survival (Figure 3A). Similar results were observed in the oxidative stress assay, when bacterial exposed to 0.5, 1, 2, 4, 8, 16 mM H2O2 for 30 min, the survival rate of wild strain were 85.04, 81.64, 77.24, 82.07, 88.24, 73.8%, while only 75.13, 61.73, 46.6, 51.13, 61.38, 55.01% of the ΔqseC cells survived (Figure 3B). When incubated in 39, 42 and 45°C water bath for 30 min, the wild strain exhibited survival rates of 68.25, 71.30, 34.6%. However, the resistance of ΔqseC to heat-shock was significantly decreased, with survival rate of 61.74, 20.59, and 17.30% (Figure 3C). These findings indicated that QseC played an important role in stress tolerance in H. parasuis.
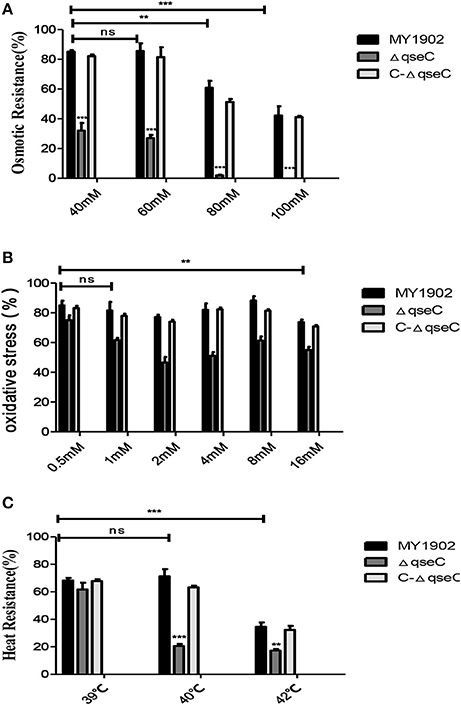
Figure 3. Analysis of the stress tolerance of wild strain MY1902, ΔqseC and C-ΔqseC. (A) The bacterial were treated with 40, 60, 80, and 100 mM NaCl TSA, (B) The bacterial exposed to 0.5, 1, 2, 4, 8, 16 mM H2O2 for 30 min, (C) The bacterial were incubated in 39, 42, and 45°C water bath for 30 min. Data indicate the mean of three independent experiments performed in duplicates and error bars show SDs. Asterisks indicate statistical significance using two-way ANOVA (**P < 0.01; ***P < 0.001).
The ability of utilize iron in the wild strain MY1902 and the qseC mutant strain were studied by using of iron restricted medium with 100, 200 μM EDDHA or 33.33 μM FeSO4, 66.66 μM FeSO4 respectively, containing 5% inactivated bovine serum and 0.01%NAD. As shown in Figure 3, when exposed to 100 /200 μM EDDHA, the ΔqseC decreased the growth compared with the wild strain in H. parasuis. Whereas supplement of FeSO4, the ΔqseC restored the growth rate (Figure 4).
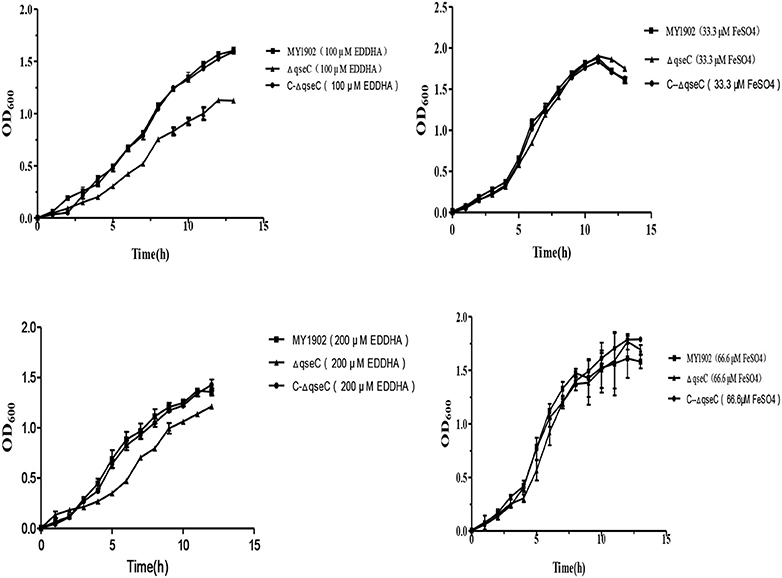
Figure 4. The capability of utilize iron of the wild strain MY1902, ΔqseC, and C-ΔqseC. Growth of the wild strain MY1902, ΔqseC and C-ΔqseC in iron restricted medium and iron supplementation medium. Error bars represent the standard deviations of three independent experiments.
The biofilm formation assay results show that QseC was involved in biofilm formation in H. parasuis (Figure 5). Biofilm productions was measured at wavelength of 595 nm.
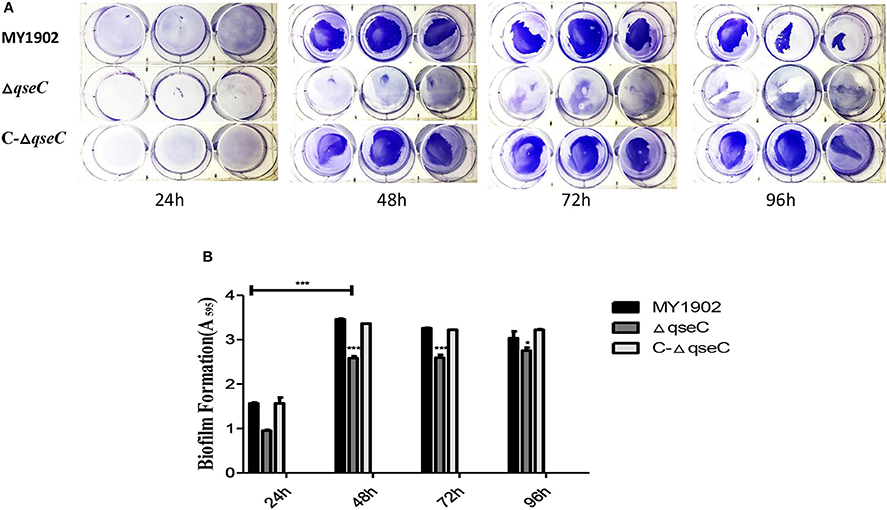
Figure 5. (A) Biofilms were stained with crystal violet. (B) Quantification of biofilm productions. Error bars represent the standard deviations of three independent experiments. Asterisks indicate statistical significance using two-way ANOVA (*P < 0.05; ***P < 0.001).
Previously study proved that qseBC is activated by AI-3. AI-3, and Epi are recognized by the same receptor, and ΔqseC is unable to respond to both AI-3 and Epi in Enterohemorrhagic E. coli (Sperandio et al., 2003). We found that when exposed to 50 μM exogenous epinephrine, the ΔqseC weakened the ability of feeling the signal AI-3 (Figure 6). The specific mechanisms of interkingdom communication still unkown, and further research are needed in this specific mechanisms, which is important in understanding bacterial pathogenesis.
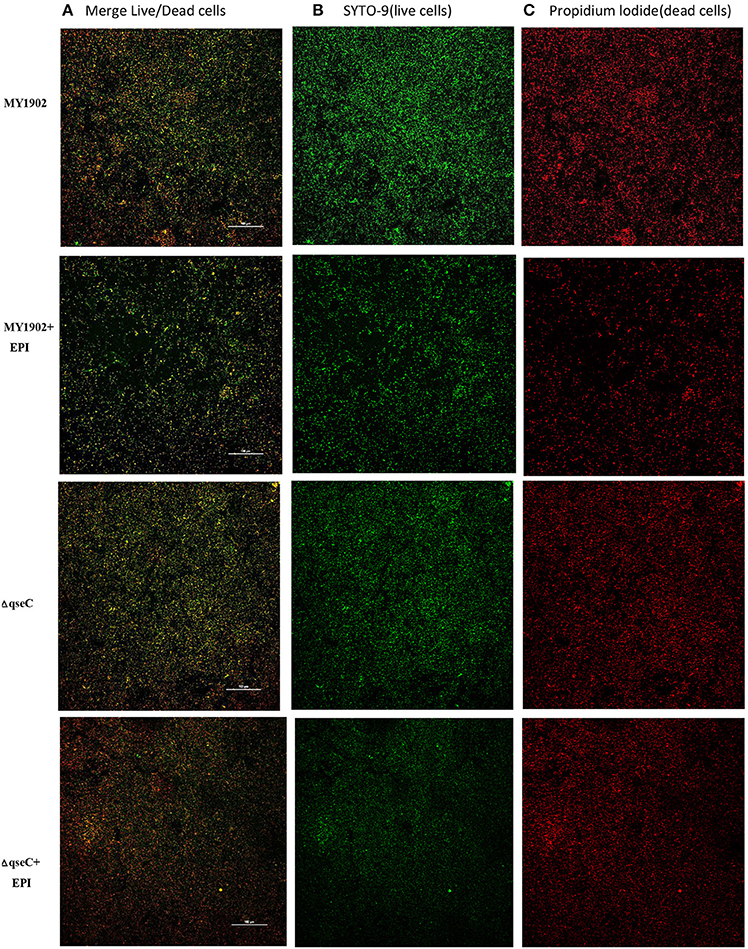
Figure 6. CLSM images of MY1902 and ΔqseC. H. parasuis were cultured in six-well microtiter plates were stained with the SYTO-9 and propidium iodide to label live versus dead cells.
Bacteria have multiple mechanisms for sensing the environment and regulate gene expressions in response to different niches of their host organisms, which is frequently mediated by TCSTS (Labandeira-Rey et al., 2010; Xu et al., 2014). CheY/QseC is one of the TCSTS in H. parasuis, and involved in virulence gene expression in a number of pathogenic bacterias (Kostakioti et al., 2009). QseC controls the phosphorylation of QseB in order to optimize expression patterns (metabolic and virulence genes) in E. coli (Hadjifrangiskou et al., 2011). QseBC plays an important role in flagellar motility, fimbrial hemagglutination, and intracellular virulence in fish pathogen Edwardsiella tarda (Wang et al., 2011). QseBC of E. coli shares the homology with H. parasuis regulator Y and C (CheY/QseC). However, the functions of QseC in the H. parasuis adapts to the environment are unknown. It is necessary to explore the function of the QseC in H. parasuis.
In this study, we constructed a qseC deletion mutant of H. parasuis serovar 13 strain MY1902(ΔqseC), and studied the survival rate under a variety of stress conditions as well as relevant biological characteristics. Results showed that the QseC played an important role in stress tolerance and biofilm formation of H. parasuis strain. Whereas we found that the ΔqseC didn't exhibit obvious growth defects compared with the wild strain. In this study, we observed that the ΔqseC was more sensitive to osmotic pressure, oxidative stress, and heat shock. Most notably the ΔqseC significantly osmotic pressure tolerance, in which the survival rate of wild strain was 42.1% whereas the ΔqseC didn't grow when exposed to 100 mM NaCl. These data suggested that the QseBC two-component system participated in H. parasuis responded to signal in the environment and survived in the stress conditions.
Iron is essential for bacterial growth, and it's an environmental signal that regulates the expression of virulence factors (Jacques, 2004). Iron contributes a lot to the growth of H. parasuis, and low iron availability in the host is a primary pressure for the pathogenic bacterium and considered a signal that leads to significant changes in cell processes (Deslandes et al., 2007). In this study we found that QseC might regulate the expression of some of the genes involved in iron uptake, and further studies will be necessary to evaluate the impact of the two-component regulatory systems CheY/QseC during the course of iron acquisition in H. parasuis.
Furthermore, we observed that the biofilm formation ability of the ΔqseC was weaker than the wild-type strain. Bacterial biofilm formation is a complex multifactor process, which is involved in adherence, competence, quorum sensing, cell wall synthesis, metabolism, and the stress response (Hasona et al., 2007). Previous studies have demonstrated that qseC controls biofilm formation of non-typeable H. influenza (Unal et al., 2012). Similarly, protein CheY was proved to influence biofilm formation in H. parasuis (He et al., 2016), which might closely related with QseC. In this study, we demonstrated that QseC was involved in biofilms formation in H. parasuis.
QseC, a histidine sensor kinase that can sense epinephrine (EPI)/norepinephrine (NE) was the quorum-sensing regulator of E. coli and Eschericha coli and S. enterica (Yang et al., 2014; Weigel et al., 2015). Weigel et al. demonstrated that iron and catecholamines may be signals that activate the QseC sensor, and detection of catecholamines and iron by the QseBC two-component system may essential for the adaptation of A. actinomycetemcomitans to the host cell environment (Weigel et al., 2015). Privious study demonstrated that epinephrine (EPI) and norepinephrine (NE) can promote the growth of a variety of bacteria, including Pseudomonas aeruginosa, Yersinia enterocolitica, E. coli (Green et al., 2003). But some scholars noted that the addition of 50 μM EPI or NE did not accelerate E. coli growth, which was in contrast to prior reports describing that NE increased the growth of E. coli and other bacteria (Yang et al., 2014). Interestingly, we found that the addition of 50 μM epinephrine reduced the cell density of MY1902 in H. parasuis, which might had an relationship with the QseC quorum-sensing sensor kinase. The ΔqseC, which lacks the ability to sense the hormones, showed little difference when exposure to epinephrine in the environment. Detection of AI-3 by the QseBC two-component system may play an important role in the growth of H. parasuis.
In conclusion, we successfully constructed the ΔqseC, C-ΔqseC and investigated the functions of QseC in the H. parasuis on stress response, iron utilization, biofilm formation and sense epinephrine. The ΔqseC obviously weakened the ability of stress tolerance such as osmotic pressure, oxidative stress, heat shock. In addition, the ΔqseC decreased the ability of iron acquisition and biofilm formation compared with the wild-type strain MY1902 in H. parasuis, which suggested that QseBC two-component system played an important role in sensing the external stimuli and adapt to environmental pressures. Further studies are needed to determine the regulatory mechanism of transmembrane protein QseC interacted with the response regulator CheY in H. parasuis.
XW, SC, XHu, RW, YH, and QZ: designed this experiment; LH, KD, LD, and YW: implement the experimental program the experimental program; QY, XM, and XHa: modify the articles; LH: organize data and write articles.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
This work, including the efforts of XW, was funded by Ministry of Agriculture of the People's Republic of China (MOA) (201303034). The plasmid PSF116 was provided by the Key Laboratory of Veterinary Vaccine Innovation of the Ministry of Agriculture, College of Veterinary Medicine, South China Agricultural University, Guangzhou, China.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2018.00212/full#supplementary-material
1. ^Quorum sensing Escherichia coli regulators B and C (QseBC)_ a novel two-component regulatory systeminvolved in the regulation of flagella and motility byquorum sensing in E. coli.pdf.
2. ^Signaling by two-component system non-cognate partners promotes intrinsic tolerance to polymyxin B in uropathogenic Escherichia coli.pdf.
3. ^Strong cross-system interactions drive the activation of the Qse B response regulator in the absence of its cognate sensor.pdf.
4. ^Molecular cloning and characterization of the ferric hydroxamate uptake (fhu) operon in Actinobacillus pleuropneumoniae.pdf.
Allen, C. E., and Schmitt, M. P. (2009). HtaA is an iron-regulated hemin binding protein involved in the utilization of heme iron in Corynebacterium diphtheriae. J. Bacteriol. 191, 2638–2648. doi: 10.1128/JB.01784-08
Bearson, B. L., and Bearson, S. M. (2008). The role of the QseC quorum-sensing sensor kinase in colonization and norepinephrine-enhanced motility of Salmonella enterica serovar Typhimurium. Microb. Pathog. 44, 271–278. doi: 10.1016/j.micpath.2007.10.001
Beddek, A. J., Sheehan, B. J., Bossé, J. T., Rycroft, A. N., Kroll, J. S., and Langford, P. R. (2004). Two TonB systems in Actinobacillus pleuropneumoniae: their roles in iron acquisition and virulence. Infect. Immun. 72, 701–708. doi: 10.1128/IAI.72.2.701-708.2004
Bigas, A., Garrido, M. E., de Rozas, A. M., Badiola, I., Barbé, J., and Llagostera, M. (2005). Development of a genetic manipulation system for Haemophilus parasuis. Vet. Microbiol. 105, 223–228. doi: 10.1016/j.vetmic.2004.10.015
Breland, E. J., Zhang, E. W., Bermudez, T., Martinez, C. R. III., and Hadjifrangiskou, M. (2017). The histidine residue of QseC is required for canonical signaling between QseB and PmrB in uropathogenic Escherichia coli. J. Bacteriol. 199:e00060–17. doi: 10.1128/JB.00060-17
Cai, X., Chen, H., Blackall, P. J., Yin, Z., Wang, L., Liu, Z., et al. (2005). Serological characterization of Haemophilus parasuis isolates from China. Vet. Microbiol. 111, 231–236. doi: 10.1016/j.vetmic.2005.07.007
Clarke, M. B., Hughes, D. T., Zhu, C., Boedeker, E. C., and Sperandio, V. (2006). The QseC sensor kinase: a bacterial adrenergic receptor. Proc. Natl. Acad. Sci. U.S.A. 103, 10420–10425. doi: 10.1073/pnas.0604343103
Deslandes, V., Nash, J. H., Harel, J., Coulton, J. W., and Jacques, M. (2007). Transcriptional profiling of Actinobacillus pleuropneumoniae under iron-restricted conditions. BMC Genomics 8:72. doi: 10.1186/1471-2164-8-72
Eguchi, Y., Itou, J., Yamane, M., Demizu, R., Yamato, F., Okada, A., et al. (2007). B1500, a small membrane protein, connects the two-component systems EvgS/EvgA and PhoQ/PhoP in Escherichia coli. Proc. Natl. Acad. Sci. U.S.A. 104, 18712–18717. doi: 10.1073/pnas.0705768104
Frees, D., Chastanet, A., Qazi, S., Sørensen, K., Hill, P., Msadek, T., et al. (2004). Clp ATPases are required for stress tolerance, intracellular replication and biofilm formation in Staphylococcus aureus. Mol. Microbiol. 54, 1445–1462. doi: 10.1111/j.1365-2958.2004.04368.x
Green, B. T., Lyte, M., Kulkarni-Narla, A., and Brown, D. R. (2003). Neuromodulation of enteropathogen internalization in Peyer's patches from porcine jejunum. J. Neuroimmunol. 141, 74–82. doi: 10.1016/S0165-5728(03)00225-X
Hadjifrangiskou, M., Kostakioti, M., Chen, S. L., Henderson, J. P., Greene, S. E., and Hultgren, S. J. (2011). A central metabolic circuit controlled by QseC in pathogenic Escherichia coli. Mol. Microbiol. 80, 1516–1529. doi: 10.1111/j.1365-2958.2011.07660.x
Hasona, A., Zuobi-Hasona, K., Crowley, P. J., Abranches, J., Ruelf, M. A., Bleiweis, A. S., et al. (2007). Membrane composition changes and physiological adaptation by Streptococcus mutans signal recognition particle pathway mutants. J. Bacteriol. 189, 1219–1230. doi: 10.1128/JB.01146-06
He, L., Wen, X., Yan, X., Ding, L., Cao, S., Huang, X., et al. (2016). Effect of cheY deletion on growth and colonization in a Haemophilus parasuis serovar 13 clinical strain EP3. Gene 577, 96–100. doi: 10.1016/j.gene.2015.11.046
Huang, J., Wang, X., Cao, Q., Feng, F., Xu, X., and Cai, X. (2016). ClpP participates in stress tolerance and negatively regulates biofilm formation in Haemophilus parasuis. Vet. Microbiol. 182, 141–149. doi: 10.1016/j.vetmic.2015.11.020
Hughes, D. T., Clarke, M. B., Yamamoto, K., Rasko, D. A., and Sperandio, V. (2009). The QseC adrenergic signaling cascade in Enterohemorrhagic E. coli (EHEC). PLoS Pathog. 5:e1000553. doi: 10.1371/journal.ppat.1000553
Jacques, M. (2004). Surface polysaccharides and iron-uptake systems of Actinobacillus pleuropneumoniae. Can. J. Vet. Res. 68, 81–85.
Kalia, V. C. (2013). Quorum sensing inhibitors: an overview. Biotechnol. Adv. 31, 224–245. doi: 10.1016/j.biotechadv.2012.10.004
Kaplan, J. B., and Mulks, M. H. (2005). Biofilm formation is prevalent among field isolates of Actinobacillus pleuropneumoniae. Vet. Microbiol. 108, 89–94. doi: 10.1016/j.vetmic.2005.02.011
Kostakioti, M., Hadjifrangiskou, M., Pinkner, J. S., and Hultgren, S. J. (2009). QseC-mediated dephosphorylation of QseB is required for expression of genes associated with virulence in uropathogenic Escherichia coli. Mol. Microbiol. 73, 1020–1031. doi: 10.1111/j.1365-2958.2009.06826.x
Labandeira-Rey, M., Brautigam, C. A., and Hansen, E. J. (2010). Characterization of the CpxRA regulon in Haemophilus ducreyi. Infect. Immun. 78, 4779–4791. doi: 10.1128/IAI.00678-10
Li, Y. H. P., Lau, C. Y., Tang, N., Svensäter, G., Ellen, R. P., and Cvitkovitch, D. G. (2002). Novel two-component regulatory system involved in biofilm formation and acid resistance in Streptococcus mutans. J. Bacteriol. 184, 6333–6342. doi: 10.1128/JB.184.22.6333-6342.2002
Liu, M., Bouhsira, E., Boulouis, H. J., and Biville, F. (2013). The Bartonella henselae SitABCD transporter is required for confronting oxidative stress during cell and flea invasion. Res. Microbiol. 164, 827–837. doi: 10.1016/j.resmic.2013.06.009
Nasrallah, G. K., Abdelhady, H., Tompkins, N. P., Carson, K. R., and Garduño, R. A. (2014). Deletion of potD, encoding a putative spermidine-binding protein, results in a complex phenotype in Legionella pneumophila. Int. J. Med. Microbiol. 304, 703–716. doi: 10.1016/j.ijmm.2014.05.004
Oliveira, S., and Pijoan, C. (2004). Haemophilus parasuis: new trends on diagnosis, epidemiology and control. Vet. Microbiol. 99, 1–12. doi: 10.1016/j.vetmic.2003.12.001
Sperandio, V., Torres, A. G., Jarvis, B., Nataro, J. P., and Kaper, J. B. (2003). Bacteria-host communication: the language of hormones. Proc. Natl. Acad. Sci. U.S.A. 100, 8951–8956. doi: 10.1073/pnas.1537100100
Stock, A. M., Robinson, V. L., and Goudreau, P. N. (2000). Two-component signal transduction. Annu. Rev. Biochem. 69, 183–215. doi: 10.1146/annurev.biochem.69.1.183
Tremblay, Y. D., Deslandes, V., and Jacques, M. (2013). Actinobacillus pleuropneumoniae genes expression in biofilms cultured under static conditions and in a drip-flow apparatus. BMC Genomics 14:364. doi: 10.1186/1471-2164-14-364
Unal, C. M., Singh, B., Fleury, C., Singh, K. L., Chávez de Paz Svensáter, G., et al. (2012). QseC controls biofilm formation of non-typeable Haemophilus influenzae in addition to an AI-2-dependent mechanism. Int. J. Med. Microbiol. 302, 261–269. doi: 10.1016/j.ijmm.2012.07.013
Walters, M., and Sperandio, V. (2006). Quorum sensing in Escherichia coli and Salmonella. Int. J. Med. Microbiol. 296, 125–131. doi: 10.1016/j.ijmm.2006.01.041
Wang, X., Wang, Q., Yang, M., Xiao, J., Liu, Q., Wu, H., et al. (2011). QseBC controls flagellar motility, fimbrial hemagglutination and intracellular virulence in fish pathogen Edwardsiella tarda. Fish Shellfish Immunol. 30, 944–953. doi: 10.1016/j.fsi.2011.01.019
Wang, X., Xu, X., Wu, Y., Li, L., Cao, R., Cai, X., et al. (2013). Polysaccharide biosynthesis protein CapD is a novel pathogenicity-associated determinant of Haemophilus parasuis involved in serum-resistance ability. Vet. Microbiol. 164, 184–189. doi: 10.1016/j.vetmic.2013.01.037
Weigel, W. A., Demuth, D. R., Torres-Escobar, A., and Juárez-Rodríguez, M. D. (2015). Aggregatibacter actinomycetemcomitans QseBC is activated by catecholamines and iron and regulates genes encoding proteins associated with anaerobic respiration and metabolism. Mol. Oral Microbiol. 30, 384–398. doi: 10.1111/omi.12101
Wong, S. M., Alugupalli, K. R., Ram, S., and Akerley, B. J. (2007). The ArcA regulon and oxidative stress resistance in Haemophilus influenzae. Mol. Microbiol. 64, 1375–1390. doi: 10.1111/j.1365-2958.2007.05747.x
Wuichet, K., Cantwell, B. J., and Zhulin, I. B. (2010). Evolution and phyletic distribution of two-component signal transduction systems. Curr. Opin. Microbiol. 13, 219–225. doi: 10.1016/j.mib.2009.12.011
Xie, F., Li, G., Zhang, Y., Zhou, L., Liu, S., Liu, S., et al. (2016). The Lon protease homologue LonA, not LonC, contributes to the stress tolerance and biofilm formation of Actinobacillus pleuropneumoniae. Microb. Pathog. 93, 38–43. doi: 10.1016/j.micpath.2016.01.009
Xie, F., Zhang, Y., Li, G., Zhou, L., Liu, S., and Wang, C. (2013). The ClpP protease is required for the stress tolerance and biofilm formation in Actinobacillus pleuropneumoniae. PLoS ONE 8:e53600. doi: 10.1371/journal.pone.0053600
Xu, J., Fu, S., Liu, M., Xu, Q., Bei, W., Chen, H., et al. (2014). The two-component system NisK/NisR contributes to the virulence of Streptococcus suis serotype 2. Microbiol. Res. 169, 541–546. doi: 10.1016/j.micres.2013.11.002
Yang, K., Meng, J., Huang, Y. C., Ye, L. H., Li, G. J., Huang, J., et al. (2014). The role of the QseC quorum-sensing sensor kinase in epinephrine-enhanced motility and biofilm formation by Escherichia coli. Cell Biochem. Biophys. 70, 391–398. doi: 10.1007/s12013-014-9924-5
Zhang, B., Feng, S., Xu, C., Zhou, S., He, Y., Zhang, L., et al. (2012a). Serum resistance in Haemophilus parasuis SC096 strain requires outer membrane protein P2 expression. FEMS Microbiol. Lett. 326, 109–115. doi: 10.1111/j.1574-6968.2011.02433.x
Zhang, B., He, Y., Xu, C., Xu, L., Feng, S., Liao, M., et al. (2012b). Cytolethal distending toxin (CDT) of the Haemophilus parasuis SC096 strain contributes to serum resistance and adherence to and invasion of PK-15 and PUVEC cells. Vet. Microbiol. 157, 237–242. doi: 10.1016/j.vetmic.2011.12.002
Keywords: Haemophilus parasuis, QseC, stress tolerance, biofilm formation, iron utilization
Citation: He L, Dai K, Wen X, Ding L, Cao S, Huang X, Wu R, Zhao Q, Huang Y, Yan Q, Ma X, Han X and Wen Y (2018) QseC Mediates Osmotic Stress Resistance and Biofilm Formation in Haemophilus parasuis. Front. Microbiol. 9:212. doi: 10.3389/fmicb.2018.00212
Received: 15 August 2017; Accepted: 30 January 2018;
Published: 13 February 2018.
Edited by:
Felipe Lombó, Universidad de Oviedo Mieres, SpainReviewed by:
Stephen Peter Kidd, University of Adelaide, AustraliaCopyright © 2018 He, Dai, Wen, Ding, Cao, Huang, Wu, Zhao, Huang, Yan, Ma, Han and Wen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yiping Wen, eXVlbGlhbmc1MTg5QDE2My5jb20=
†These authors have contributed equally to this work.
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.