 Ximena Blanco Crivelli1*
Ximena Blanco Crivelli1* María P. Bonino1
María P. Bonino1 Paula Von Wernich Castillo1
Paula Von Wernich Castillo1 Armando Navarro2
Armando Navarro2 Osvaldo Degregorio3
Osvaldo Degregorio3 Adriana Bentancor1
Adriana Bentancor1- 1Microbiología, Facultad de Ciencias Veterinarias, Universidad de Buenos Aires, Buenos Aires, Argentina
- 2Departamento de Salud Pública, Facultad de Medicina, Universidad Nacional de México, Mexico City, Mexico
- 3Salud Pública, Facultad de Ciencias Veterinarias, Universidad de Buenos Aires, Buenos Aires, Argentina
Enteropathogenic Escherichia coli (EPEC) and Shiga toxin-producing E. coli (STEC) are pathovars of E. coli that impact human health by causing childhood diseases. In this work, 118 synanthropic rodents of the genus Rattus from Buenos Aires, Argentina were evaluated as EPEC and STEC carriers. Rectal swab samples from captured animals were evaluated by conventional PCR to detect the presence of the eae, stx1, stx2, and rfbO157 genes. Twenty-one isolates were obtained (17 EPEC isolates from seven animals and four STEC isolates from the same animal). All EPEC isolates tested negative for the presence of the bfpA gene. One EPEC isolate carried the iha gene, and five EPEC isolates carried the toxB gene. STEC isolates exhibited two different virulence profiles: stx1a/stx2a/stx2c/stx2d/saa/ehxA/subA (3/4) and stx1a/stx2a/saa/ehxA/subA (1/4). EPEC isolate serotypes included O109:H46 (7), O71:H40 (4), O71:NM (2), O138:H40 (1), O108:H21 (1), O88:H25 (1), and O76:NM (1), and STEC isolates belonged to the O108:H11 (4) serotype. Antimicrobial susceptibility testing was carried out, and resistance to tetracycline was observed in one EPEC strain. Our results demonstrate that Rattus spp. may act as carriers of EPEC and STEC strains and may be involved in the epidemiology of diarrheal disease in infancy.
Introduction
Escherichia coli is a microorganism belonging to the Enterobacteriaceae family, whose habitat is the intestines of various animal species including humans. Non-pathogenic E. coli can become pathogenic following the acquisition of mobile genetic elements by horizontal transfer and interactions with bacteriophages or transposons that interface between E. coli and other bacterial species (Kaper et al., 2004; Williams et al., 2010). Non-pathogenic and pathogenic E. coli can be differentiated using molecular methods (Nataro and Kaper, 1998) and in vitro assays in cell lines (Konowalchuk et al., 1977).
Infections resulting from pathogenic E. coli may be limited to mucosal surfaces or spread throughout the body. Three clinical syndromes can result from infection with different E. coli pathovars: diarrheagenic disease, urinary tract infections, and sepsis/meningitis (Nataro and Kaper, 1998). Some E. coli pathovars that produce diarrheagenic disease cause histological lesions known as attaching and effacing (A/E) lesions in the gut mucosa of human and animal hosts (Nataro and Kaper, 1998). The formation of A/E lesions involves both plasmid and chromosomal genes that are encoded in a pathogenicity island known as the locus for enterocyte effacement (LEE) (McDaniel et al., 1995). The eae gene is present in the LEE and encodes intimin, an adhesin that mediates an intimate connection between the bacterium and the enterocyte, causing localized destruction of microvilli and the formation of an actin-rich pedestal-like structure on the apical cell membrane (Wales et al., 2005).
Enteropathogenic E. coli (EPEC) produce A/E lesions. EPEC strains have been identified as one of the causes of childhood diarrhea, which is responsible for 1000s of deaths worldwide (Ochoa et al., 2008). Cases of diarrhea caused by EPEC vary from subclinical to fatal infections (Torres et al., 2010). Children with clinical diarrhea caused by EPEC usually fail to respond to oral rehydration therapy, suffer from cow milk intolerance, and develop persistent diarrhea requiring hospitalization (Fagundes-Neto and Scaletsky, 2000). EPEC strains are classified into two groups, typical (tEPEC) and atypical (aEPEC), according to the presence or absence of the EPEC adherence factor plasmid (pEAF). This plasmid includes the bfp gene, which encodes the bundle-forming pilus necessary for localized adhesion in cell culture (Kaper et al., 2004). tEPEC strains have been isolated from humans, and aEPEC strains have been found in various animal species as well as humans (Gomes and González Pedrajo, 2010).
Shiga toxin-producing E. coli (STEC) is another pathovar that has an impact on childhood health. STEC is mainly characterized by the production of Shiga toxin (Stx), which binds to its receptor globotriaosylceramide (Gb3), disrupts protein synthesis, and kills epithelial or endothelial cells. Some STEC strains also carry the eae gene. The pathogenesis of STEC involves different presentations, from asymptomatic to mild or severe diarrhea, as well as hemolytic uremic syndrome (HUS), which may cause death. Ruminants, in particular cattle, have been identified as the main reservoir of these bacteria (Karmali, 1989). In addition, several animal species have been identified as hosts to this pathogen (Beutin et al., 1993; Leotta et al., 2006; Bentancor et al., 2007).
Furthermore, the presence of hybrid E. coli strains has been recorded in various parts of the world. In May 2011, an enteroaggregative E. coli O104:H4 transduced by a Stx2a-converting phage (EAEC/STEC) led to a food-borne outbreak in Germany with 3816 cases reported (including 54 deaths); 845 of those cases led to HUS (Frank et al., 2011). Similarly, the presence of EAEC/STEC hybrid strains associated with HUS and diarrhea has been recorded in Argentina since 2005 and has been associated with the O59:NM [H19] and ONT:H4 serogroups (Rivas, 2016). Nyholm et al. (2015) analyzed the genomes of three enterotoxigenic/STEC E. coli hybrid strains and concluded that virulence genes can co-exist in strains originating from different phylogenetic lineages. Considering the emergence of hybrid strains or heteropathogenic E. coli, the current genetic classification of strains into classical pathotypes is limited. Detection of hybrid pathovars is important because they may have a significant impact on public health.
The presence of EPEC and STEC strains in synanthropic animals, especially those which belong to the genus Rattus, is important because of the direct or indirect contact they may have with humans and household animals. The aim of this work was to evaluate synanthropic rodents of the Rattus genus from Buenos Aires, Argentina as carriers of EPEC and STEC strains and to assess the virulence profiles of these strains. Given the impact of diarrhea on children in Argentina, this study is of great importance to the epidemiology of transmissible diseases, as it provides health intervention criteria.
Materials and Methods
Samples
We conducted a cross-sectional epidemiological study to determine the presence of EPEC and STEC strains in rodents of the Rattus genus in Buenos Aires. Convenience non-probability sampling was carried out at accessible points in Buenos Aires, including three case animals that were temporally and spatially related to a HUS outbreak according to the National Health Surveillance System (restricted up to 50 m of HUS address within the week of the outbreak). The target population corresponded to Rattus rodents obtained in Buenos Aires; thus, all rodents belonging to the Rattus genus that were captured in urbanized areas of the city of Buenos Aires were included, while all non-Rattus rodents and non-rodent animals captured were excluded.
Rats were captured using live traps (2620 cage traps) set in different areas of Buenos Aires (Table 1). The traps remained active for four consecutive nights, and they were checked each morning, resulting in a total of 118 rats captured (Figure 1).
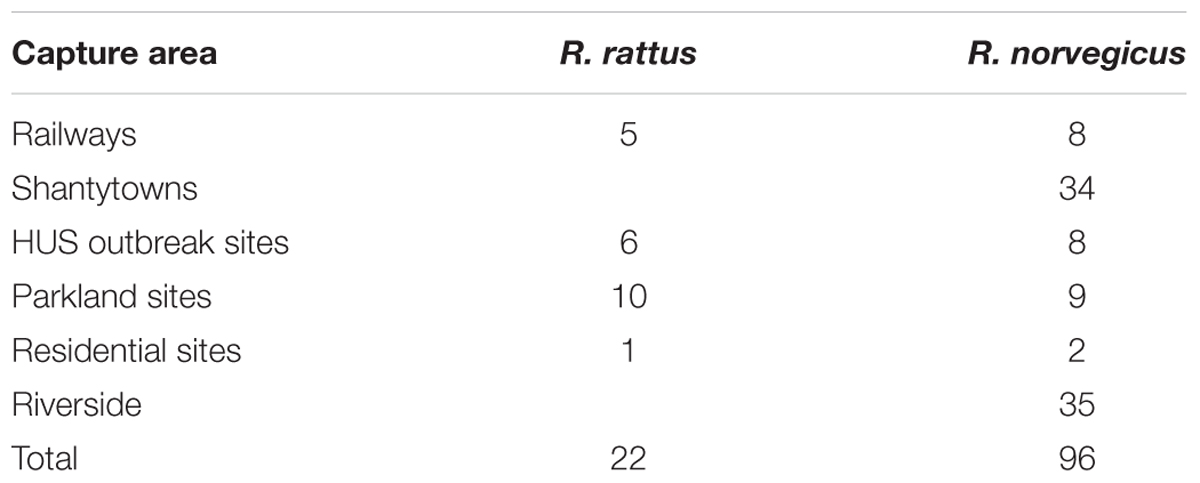
TABLE 1. Locations and species of animals captured.
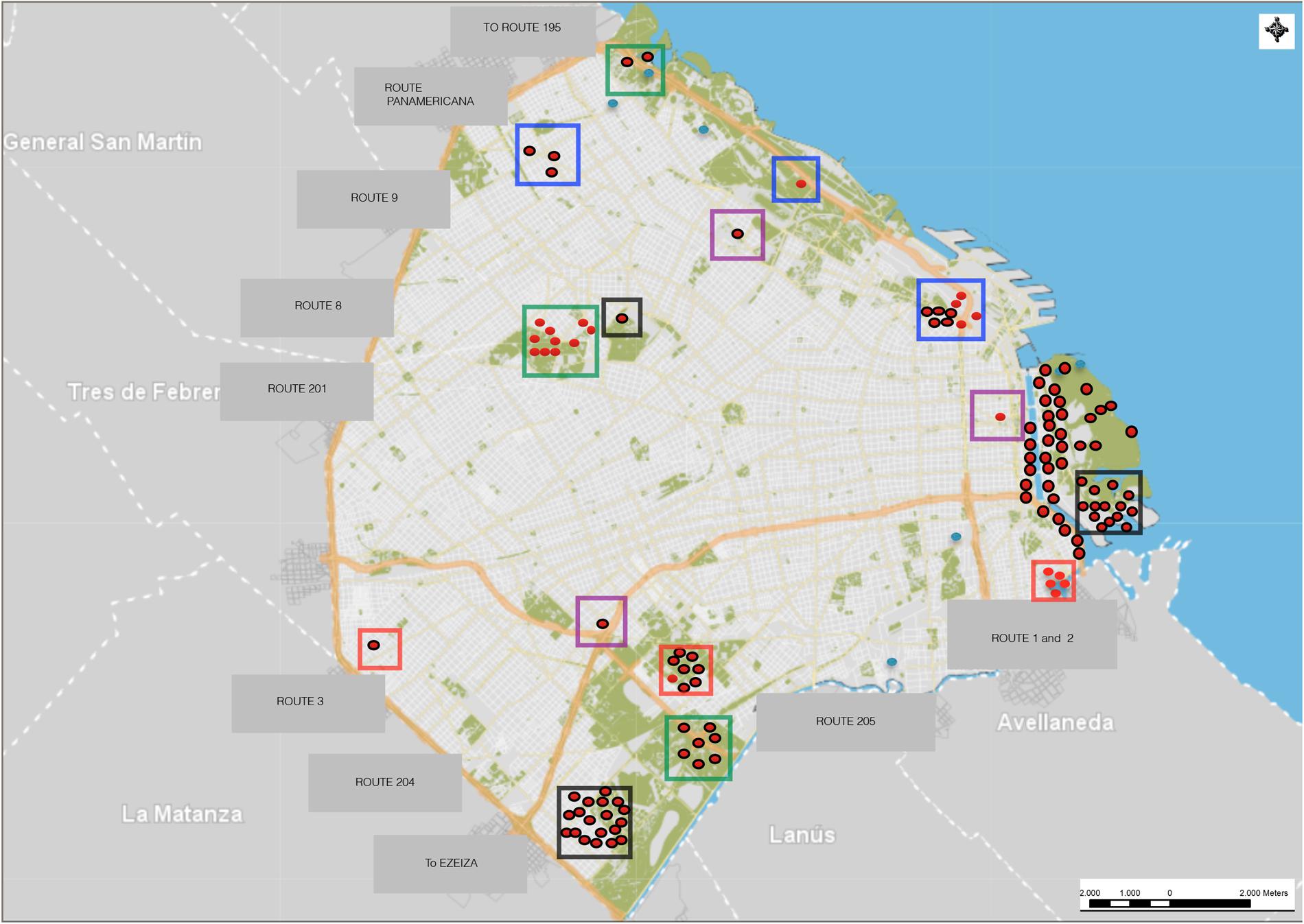
FIGURE 1. Sites of convenience sampling and trapping of Rattus spp. in Buenos Aires. Red dots: Rattus rattus captured; red dots with black outlines: Rattus norvegicus captured; light blue dots: sites with traps without capture; red squares: HUS outbreak sites; purple squares: residential sites; black squares: shantytowns; green squares: parkland sites; blue squares: railways; areas without squares: riverside.
Each rat was euthanized in accordance with the recommendations of the American Veterinary Medical Association guidelines for the euthanasia of animals (Underwood et al., 2013). We used a protocol that was approved by the Institutional Committee on Animal Care and Use of Experimental Animals (CICUAL; No. 2010726) of the Universidad de Buenos Aires, Facultad de Ciencias Veterinarias. Then, each rat was weighed, measured, sexed, and its reproductive status was recorded. Subsequently, the rats were classified according to species. All animals were sampled via two rectal swabs.
Detection and Isolation of EPEC and STEC Strains
Samples were used to inoculate 5 ml of tryptone soy broth (TSB) for pre-enrichment and incubated for 18 h at 37°C to detect EPEC and non-O157 STEC strains. The cultures were streaked onto MacConkey agar (MAC) and incubated overnight. Screening for the eae gene was carried out by PCR of cells from the confluence zone using primers reported by Blanco et al. (2004) with a previously described modification (Bentancor et al., 2007). The presence of eae-positive strains was verified by screening up to 50 CFU of positive samples from the confluence zone. For the detection of non-O157 STEC strains, screening of the confluence area for stx1 and stx2 genes was carried out by PCR using primers reported by Leotta et al. (2005). Suspected samples were evaluated for the presence of stx1-positive or stx2-positive strains by screening up to 50 CFU.
Furthermore, for the detection of O157 STEC strains, samples were inoculated in 5 ml of tellurite cefixime TSB for enrichment and incubated for 6 h at 37°C. Inmunomagnetic separation (IMS) with Dynabeads® anti-E. coli O157 was performed. Each Dynabeads–bacterial complex was streaked onto sorbitol MacConkey agar (SMAC) and onto E. coli O157:H7 chromogenic medium (Chrom ID) and incubated for 18 h at 37°C. Screening for stx1/stx2 and rfbO157 from the confluence area was performed by multiplex PCR (Leotta et al., 2005).
Detection Limit of IMS in Rodent Stool Samples
In order to establish the detection limit of the IMS in rodent stool, E. coli EDL 933 (positive control strain) was suspended in saline solution at a McFarland turbidity of 0.5. Viable cell count was determined using 10-fold dilutions and culture in brain heart agar (BHA). In addition, a pool of STEC-negative (NS) rodent stool pre-cultures that had been recovered previously was assessed. A total of 100 μl of each EDL 933 dilution was added to 900 μl of NS culture. IMS was performed for each tube according to the recommendations of the supplier by adding 20 μl of immunomagnetic beads and subsequently seeding in SMAC. Finally, PCR was performed to detect stx1, stx2, and rfbO157 from confluent growth zone. This test was repeated three times.
Biochemical Characterization and Identification of Additional Virulence Factors
Isolates were further characterized using biochemical tests (MacFaddin, 2003). Additional virulence factors such as saa, ehxA (Paton and Paton, 1998, 2002), the putative adhesion factor toxB (Toma et al., 2004), the iron regulatory gene iha (Schmidt et al., 2001), the cytolethal distending toxin cdtIII (Clark et al., 2002), cdtVA, cdtVB (Janka et al., 2003), and subtilase (subA) (Paton and Paton, 2005) were also detected by PCR according to methods reported in the above previous studies.
Identification of E. coli Diarrheagenic Hybrid Pathovars
Other markers in E. coli diarrheagenic pathovars were evaluated by conventional PCR, including the aaiC (Boisen et al., 2012) and aagR (Wieler et al., 2011) genes for enteroaggregative E. coli (EAEC), elt and estA (Itoh et al., 1992) for enterotoxigenic E. coli (ETEC), daaE (Vidal et al., 2005) for diffusely adherent E. coli (DAEC), and invE (Itoh et al., 1992) for enteroinvasive E. coli (EIEC).
Detection of bfpA-Positive EPEC Strains
Strains positive for eae and negative for stx were further analyzed to detect the bfpA gene. The primers used and the PCR conditions have been previously described (Gunzburg et al., 1995).
Subtyping of stx-Positive STEC Strains
Strains positive for stx were further analyzed to detect subtypes of stx1 and stx2 by PCR. The primers used and the PCR conditions have been previously described (Scheutz et al., 2012).
Production of Shiga Toxin by STEC Strains
The production of Stx1 and Stx2 was detected with an immunochromatographic test (Duopath Verotoxin test; Merck) (Park et al., 2003). The detection limit of this test is 25 ng/ml for Stx1 and 62.5 ng/ml for Stx2. Each isolate was inoculated in blood broth and simultaneously in 2 ml of Caye broth with Caye supplement, then incubated for 24 h at 37°C and 6 h at 37°C, respectively, following the recommendations of the supplier. Subsequently, cultures were treated with polymyxin B sulfate in a ratio of 9:1 (360 μl broth + 40 μl antibiotic) and incubated in a dry bath (Accu Block, Labnet) for 10 min at 37°C. An aliquot of the inoculum was transferred to the sample port, the test device was incubated at room temperature, and the result was interpreted after 10 min.
Enterohemolysin Production
For ehxA-positive strains, we investigated the production of enterohemolysin on tryptose blood agar base supplemented with 10 mM CaCl2 and 5% defibrinated sheep blood (Beutin et al., 1989).
Serotyping of Isolated Strains
Somatic antigen (O) and flagellar antigen (H) were examined according to a previously reported methodology (Orskov and Orskov, 1984). We used 187 rabbit sera against somatic antigens (O) and 53 rabbit sera against flagellar antigens (H) from the E. coli antigenic scheme. The rabbit sera were previously obtained using reference strains of E. coli according to a proposed method by Ewing (1986).
Antimicrobial Susceptibility Testing
Isolated strains were examined for resistance to amoxicillin/clavulanic acid (30 μg), imipenem (10 μg), aztreonam (30 μg), gentamicin (10 μg), amikacin (30 μg), nalidixic acid (30 μg), ciprofloxacin (5 μg), nitrofurantoin (300 μg), and tetracycline (30 μg) using commercial monodisks (Oxoid) and tablets (Rosco) by the disk diffusion method in Müeller Hinton agar as recommended by the Clinical Laboratory and Standards Institute (CLSI). Strains were characterized as being susceptible, having reduced susceptibility, or being resistant (Clinical Laboratory and Standards Institute [CLSI], 2014).
Statistical Analysis
The data obtained were analyzed using Fisher’s exact test (InfoStat 2016e).
Results
Among the Rattus spp. captured, 96 were R. norvegicus (81%) and 22 were R. rattus (19%). The proportion of EPEC (eae-positive) carriers among R. norvegicus specimens was 3.12% (3/96). Most of these belonged to serotype O109:H46, followed by O108:H21 and O138:H21 (Table 2). Interestingly, multiple isolates belonging to serotype O109:H46 recovered from the same R. norvegicus specimen exhibited distinct characteristics: while one isolate was eae+/iha+, the others were eae+/toxB+. All R. norvegicus EPEC carriers were captured from shantytown sites (Table 2).
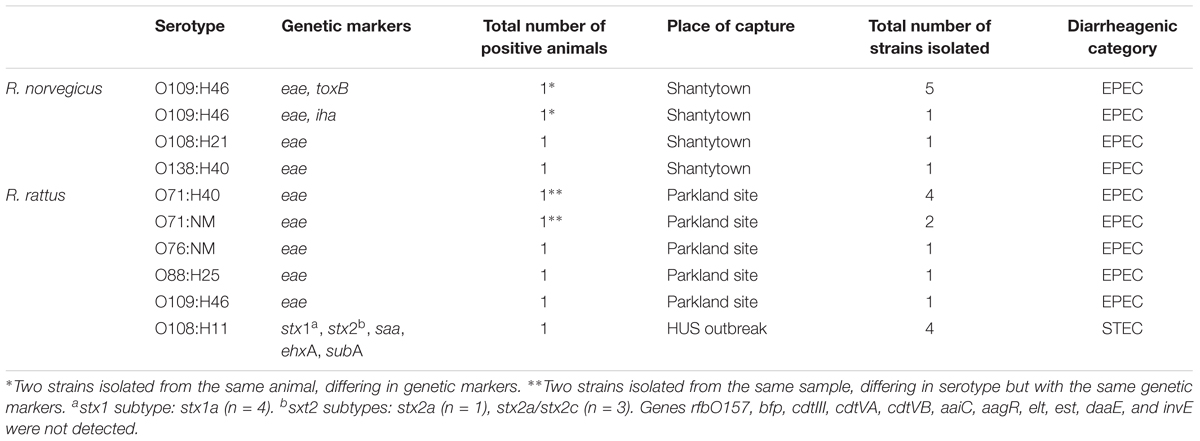
TABLE 2. Enteropathogenic and Shiga toxin-producing Escherichia coli isolated from Rattus spp.
The proportion of EPEC carriers among R. rattus specimens was 18.18% (4/22). Two EPEC strains isolated from the same sample and belonging to the O71 serogroup belonged to different serotypes (O71:H40/NM) according to serological tests. All R. rattus EPEC carriers were captured from parkland sites (Table 2).
Based on these results, there was a statistically significant difference in the rate of EPEC strains carried by the two Rattus species (p = 0.04).
A total of 17 isolates were obtained from the seven EPEC-positive Rattus specimens (Table 2). All eae-positive isolates were negative for stx1, stx2, saa, ehxA, cdtIII, cdtVA, cdtVB, and subA. In addition, all eae-positive isolates were identified as E. coli and were classified as aEPEC (bfpA-negative). One EPEC isolate was iha-positive, and five EPEC isolates were iha-negative/toxB-positive. Resistance to tetracycline was observed in one EPEC strain.
Only one sample obtained from an R. rattus specimen captured during a HUS outbreak was found to be stx1/stx2-positive/rfbO157-negative/eae-negative. Four isolates were obtained from this animal (Table 2), all of which were identified as E. coli and classified as STEC. These strains exhibited the genetic profiles stx1a/stx2a/stx2c/saa/ehxA/subA (3/4) and stx1a/stx2a/saa/ehxA/subA (1/4). All STEC isolates produced Stx1 and Stx2 and exhibited an enterohemolytic phenotype.
The serotypes of the strains isolated in this study are shown in Table 2. Other markers for E. coli diarrheagenic pathovars were not detected in EPEC or STEC isolates, and there were no hybrids. The detection limit for IMS in rodent stool samples was established to be 12.6 × 102 CFU/ml.
Discussion
In this study, we evaluated the presence of EPEC and STEC strains in rodents of the genus Rattus. The sampling methodology (rectal swab) differed from that used by other researchers working with synanthropic rodents. While Nielsen et al. (2004) took fecal samples from the environment, Rahn et al. (1997) collected feces present in live catch traps for rodents, and Hancock et al. (1998) and Cižek et al. (1999) used feces from the gastrointestinal tracts of captured animals. However, in this study, rectal swabs were used for sampling, as Greenquist et al. (2005) used swabs of the rectoanal junction mucosa to identify the presence of STEC O157 and determine the epidemiological role of cattle as a reservoir. Moreover, the presence of STEC O157 in stool samples can be related to the transient microbiota in these samples.
In this work, we isolated EPEC strains, and our results were similar to those obtained by other authors (Heidemanns et al., 2009; Firth et al., 2014). Ten animals were suspected to be carriers of the pathogen, and we isolated EPEC strains from seven animals. A higher proportion of EPEC strains were found among R. rattus specimens; this result is similar to a previous finding by our group (Rumi et al., 2012). However, we also found EPEC strains in R. norvegicus, a finding which is similar to those obtained by Firth et al. (2014). All EPEC isolates were classified as aEPEC because they tested negative for the bfpA gene. Certain features may favor the prolonged intestinal colonization of aEPEC compared to other intestinal pathogens, allowing aEPEC strains to persist for a longer period of time in the intestine (Ochoa et al., 2008). Currently, it has been reported that the proportion of aEPEC strains has increased and is associated with childhood diarrhea, while the proportion of tEPEC strains is declining (Ochoa et al., 2008; Hu and Torres, 2015); this could also be a possible explanation for the high proportion of aEPEC strains among the isolates.
The differences in the carriage of EPEC between species in this study should not be considered due to geographic clustering because we carried out a convenience non-probability sampling. Thus, this finding should be interpreted cautiously and further studies should be performed in order to examine if the difference in carriage between the species is real.
EPEC O71:H40, which was isolated from R. rattus (n = 4), has previously been isolated from ruminants (Horcajo et al., 2012) and from patients with diarrhea or other gastrointestinal disturbances (Hien et al., 2008; Horcajo et al., 2012). Moreover, an EPEC isolate from an R. rattus specimen presented the serotype O88:H25, which has a high incidence in Brazil without being associated with cases of diarrhea in humans (Pedroso et al., 1993). This has also been identified as a new serotype in Mexico, where it was isolated from children under 5 years of age with acute diarrhea and bloody diarrhea (Navarro and Estrada-García, 2010).
To our knowledge, there are no previous records of serotypes O138:H40, O109:H46, O108:H21, and O76:NM among EPEC strains, indicating that this study is the first to identify aEPEC isolates of these serotypes.
Inmunomagnetic separation appears to be an efficient method for the detection of STEC O157, as the detection limit in rodent stool samples was 12.6 × 102 CFU/ml. This value is lower than that obtained by Bentancor et al. (2007) for dog swabs (6.6 × 103 CFU/ml). However, the possibility of competition by the resident microbiota of the gastrointestinal tracts of rodents cannot be ruled out. It has been established that cross-reactions with antigenically similar microorganisms such as Escherichia hermannii, Salmonella spp., and Proteus spp. may occur in IMS. Non-specific binding with microorganisms such as Pseudomonas spp. and Serratia liquefaciens may also occur (Dynal Biotech A S.A.). Although the microbiotas of synanthropic Rattus spp. were not analyzed, they could include a high proportion of one of these microorganisms.
Screening for STEC O157 was negative, even though a selective enrichment protocol was used. The absence of STEC O157 in rat samples is in agreement with results obtained by other authors (Hancock et al., 1998; Rahn et al., 1997; Nielsen et al., 2004). However, Cižek et al. (1999) found STEC O157 in R. norvegicus. The low rate of detection of this pathogen in Rattus spp. may be due to the fact that it can be excreted quickly or sporadically by these animals, and we should not rule out the possibility of the existence of carriers with levels of the pathogen below the detection limit of the diagnostic test.
The screening for non-O157 STEC strains identified four isolates from a R. rattus specimen. The presence of non-O157 STEC in Rattus spp. coincides with the results obtained by Nielsen et al. (2004), who isolated STEC O136:H12 (stx1-positive) in R. norvegicus. However, there is no history of STEC isolates in R. rattus, except for in our previous study involving a HUS outbreak (Bentancor et al., 2009; Blanco Crivelli et al., 2012).
Shiga toxin-producing E. coli isolates carried subA, a gene that codes for subunit A of SubAB. The presence of this cytotoxin in non-O157 STEC strains was previously recorded (Paton et al., 2004). This toxin is 10–100 times more potent than Stx in Vero cells (Noda et al., 1987; Fujii et al., 2003) and may play a role in the pathogenesis of HUS (Irino et al., 2010).
The enterohemolytic phenotype was observed in all STEC isolates. This is important because enterohemolysis is associated with diarrheal disease in humans (Beutin et al., 1989). Moreover, all the STEC isolates obtained in this work corresponded to the serotype O108:H11 and the profile stx1/stx2/saa/ehxA/subA. Toxin production was verified by immunochromatography, which has a sensitivity of 100% and results in no false positives when testing individual fecal cultures (Park et al., 2003).
The STEC-positive Rattus specimen that was captured was related to a HUS outbreak according to the National Health Surveillance System. In this case, no STEC strains were isolated from affected children, so it is not possible to establish a connection with our isolated strains.
Only one EPEC isolate showed resistance to tetracycline. This is expected among wild-type strains obtained from synanthropic animals, suggesting that the isolated strains originated among the microbiota of Rattus spp. and are not derived from other sources, such as human waste, which may could have been ingested by these animals.
The typical intestinal inhabitants of rodents are not well-known, so it is possible that the intestines of these animals constitute a niche that favors variability among the strains (Heidemanns et al., 2009). Our results demonstrated that Rattus spp. can be carriers of EPEC and STEC strains. Moreover, we found a significant association between the Rattus species and the presence of EPEC. In Argentina, studies of the presence of EPEC in urban, domestic, and synanthropic animals are limited. There are few epidemiological studies of EPEC in dogs and cats, and to our knowledge, this is the first report describing synanthropic carriers of EPEC in urban areas of Argentina. Synanthropic rodents from the Rattus genus could play a role in the epidemiology of EPEC. It is therefore imperative to perform continuous monitoring and identification of carriers and risk factors for EPEC, STEC, and other E. coli pathovars of significance for public health.
Author Contributions
Conceived and designed the experiments: AB and XB. Performed the experiments: XB, MB, PVWC, AN, and AB. Analyzed the data and wrote the paper: XB, OD, and AB.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The research in this study was performed at the Universidad de Buenos Aires, Facultad de Ciencias Veterinarias and was funded by the Agencia Nacional de Promoción Científica y Tecnológica, Ministerio de Ciencia y Tecnología e Innovación Productiva of Argentina, PICT 2010-1355, UBACyT 20020150100159BA and UBACyT 20020130100463BA. We thank DGAPA-PAPIIT for their support of this project IN216417.
References
Bentancor, A., Calviño, F., Manfredi, F., Miccio, L., Ameal, L. A., Aguirre, S. M., et al. (2009). “Isolation of Shiga toxin-producing Escherichia coli from household pets and Rattus rattus related to outbreaks of hemolytic uremic syndrome,” in Procedings of the VTEC 2009 - 7th International Symposium on Shiga Toxin (Verocytotoxin) – Producing Escherichia coli Infections, Buenos Aires.
Bentancor, A., Rumi, M. V., Gentilini, M. V., Sardoy, C., Irino, K., Agostini, A., et al. (2007). Shiga toxin-producing and attaching and effacing Escherichia coli in cats and dogs in a high hemolytic uremic syndrome incidence region in Argentina. FEMS Microbiol. Lett. 267, 251–256. doi: 10.1111/j.1574-6968.2006.00569.x
Beutin, L., Geier, D., Steinruck, H., Zimmermann, S., and Scheutz, F. (1993). Prevalence and some properties of verotoxin (Shiga-like toxin)-producing Escherichia coli in seven different species of healthy domestic animals. J. Clin. Microbiol. 31, 2483–2488.
Beutin, L., Montenegro, M. A., Orskov, I., Orskov, F., Prada, J., Zimmermann, S., et al. (1989). Close association of verotoxin (Shiga-like toxin) production with enterohemolysin production in strains of Escherichia coli. J. Clin. Microbiol. 27, 2559–2564.
Blanco, M., Blanco, J. E., Mora, A., Dahbi, G., Alonso, M. P., González, E. A., et al. (2004). Serotypes, virulence genes, and intimin types of Shiga Toxin (Verotoxin)-producing Escherichia coli isolates from cattle in Spain and identification of a new intimin variant gene (eae-ξ). J. Clin. Microbiol. 42, 645–651. doi: 10.1128/JCM.42.2.645-651.2004
Blanco Crivelli, X., Rumi, M. V., Carfagnini, J. C., Degregorio, O., and Bentancor, A. B. (2012). Synanthropic rodents as possible reservoirs of shigatoxigenic Escherichia coli strains. Front. Cell. Infect. Microbiol. 2:134. doi: 10.3389/fcimb.2012.00134
Boisen, N., Scheutz, F., Rasko, D. A., Redman, J. C., Persson, S., Simon, J., et al. (2012). Genomic characterization of enteroaggregative Escherichia coli from children in Mali. J. Infect. Dis. 205, 431–444. doi: 10.1093/infdis/jir757
Cižek, A., Alexa, P., Literák, I., Hamřik, J., Novák, P., and Smola, J. (1999). Shiga toxin-producing Escherichia coli O157 in feedlot cattle and Norwegian rats from a large-scale farm. Lett. Appl. Microbiol. 28, 435–439. doi: 10.1046/j.1365-2672.1999.00549.x
Clark, C. G., Johnson, S. T., Easy, R. H., Campbell, J. L., and Rodgers, F. G. (2002). PCR for detection of cdt-III and the relative frequencies of cytolethal distending toxin variant-producing Escherichia coli isolates from humans and cattle. J. Clin. Microbiol. 40, 2671–2674. doi: 10.1128/JCM.40.7.2671-2674.2002
Clinical Laboratory and Standards Institute [CLSI] (2014). Performance Standards for Antimicrobial Susceptibility Testing. Twenty Four International Supplement. 2014; M100-S24. Wayne, PA: CLSI.
Ewing, W. H. (1986). Edwards and Ewing’s Identification of Enterobacteriaceae. New York, NY: Elsevier Science Publishing Inc.
Fagundes-Neto, U., and Scaletsky, I. C. (2000). The gut at war: the consequences of enteropathogenic Escherichia coli infection strain that causes diarrhea and malnutrition. Sao Paulo Med. J. 118, 21–29. doi: 10.1590/S1516-31802000000100006
Firth, C., Bhat, M., Firth, M. A., Williams, S. H., Frye, M. J., Simmonds, P., et al. (2014). Detection of zoonotic pathogens and characterization of novel viruses carried by commensal Rattus norvegicus in New York City. mBio 5:e01933-14. doi: 10.1128/mBio.01933-14
Frank, C., Werber, D., Cramer, J. P., Askar, M., Faber, M., An der Heiden, M., et al. (2011). Epidemic profile of Shiga-toxin-producing Escherichia coli O104:H4 outbreak in Germany. N. Engl. J. Med. 365, 1771–1780. doi: 10.1056/NEJMoa1106483
Fujii, J., Matsui, T., Heatherly, D. P., Schlegel, K. H., Lobo, P. I., Yutsudo, T., et al. (2003). Rapid apoptosis induced by Shiga toxin in Hela cells. Infect. Immun. 71, 2724–2735. doi: 10.1128/IAI.71.5.2724-2735.2003
Gomes, T. A. T., and González Pedrajo, B. (2010). “Enteropathogenic Escherichia coli (EPEC),” in Pathogenic Escherichia coli, Latin America. ed. A. G. Torres (Sharjah: Bentham Science Publishers).
Greenquist, M. A., Drouillard, J. S., Sargeant, J. M., Depenbusch, B. E., Shi, X., Lechtenberg, K. F., et al. (2005). Comparison of rectoanal mucosal swab cultures and fecal cultures for determining prevalence of Escherichia coli O157:H7 in feedlot cattle. Appl. Environ. Microbiol. 71, 6131–6133. doi: 10.1128/AEM.71.10.6431-6433.2005
Gunzburg, S. T., Tornieporth, N. G., and Riley, L. W. (1995). Identification of Enteropathogenic Escherichia coli by PCR-based detection of the bundle-forming pilus gene. J. Clin. Microbiol. Lett. 33, 1375–1377.
Hancock, D. D., Besser, T. E., Rice, D. H., Ebel, E. D., Herriott, D. E., and Carpenter, L. V. (1998). Multiple sources of Escherichia coli O157 in feedlots and dairy farms in the Northwestern USA. Prev. Vet. Med. 35, 11–19. doi: 10.1016/S0167-5877(98)00050-6
Heidemanns, M., Nordhoff, M., Semmler, T., Tietze, E., Fruth, A., Karch, A., et al. (2009). “Atypical EPEC in rodents as STEC-ancestor?” in Proceedings of the VTEC 2009 - 7th International Symposium on Shiga Toxin (Verocytotoxin) – Producing Escherichia coli Infections, Buenos Aires.
Hien, B. T. T., Scheutz, F., Cam, P. D., Serichantalergs, O., Huong, T. T., Thu, T. M., et al. (2008). Diarrheagenic Escherichia coli and Shigella strains isolated from children in a hospital case-control study in Hanoi Vietnam. J. Clin. Microbiol. 46, 996–1004. doi: 10.1128/JCM.01219-07
Horcajo, P., Domínguez-Bernal, G., de la Fuente, R., Ruiz-Santa-Quiteria, J. A., Blanco, J. E., Blanco, M., et al. (2012). Comparison of ruminant and human attaching and effacing Escherichia coli (AEEC) strains. Vet. Microbiol. 155, 341–348. doi: 10.1016/j.vetmic.2011.08.034
Hu, J., and Torres, A. G. (2015). Enteropathogenic Escherichia coli: foe or innocent bystander? Clin. Microbiol. Infect. 21, 729–734. doi: 10.1016/j.cmi.2015.01.015
Irino, K., Midolli Vieira, M. A., Tardelli Gomes, T. A., Cabilio Guth, B. E., Furtado Naves, Z. V., Gomes Oliveira, M., et al. (2010). Subtilase cytotoxin-encoding subAB operon found exclusively among Shiga toxin-producing Escherichia coli strains. J. Clin. Microbiol. 48, 988–990. doi: 10.1128/JCM.00010-10
Itoh, F., Ogino, T., Itoh, F., and Watanabe, H. (1992). Differentiation and detection of pathogenic determinants among diarrheagenic Escherichia coli by polymerase chain reaction using mixed primers. Jap. J. Clin. Med. 50, 343–347.
Janka, A., Bielaszewska, M., Dobrindt, U., Greune, L., Schmidt, M. A., and Karch, H. (2003). Cytolethal distending toxin gene cluster in enterohemorrhagic Escherichia coli O157:NM and O157:H7 characterization and evolutionary considerations. Infect. Immun. 71, 3634–3638. doi: 10.1128/IAI.71.6.3634-3638.2003
Kaper, J. B., Nataro, J. P., and Mobley, H. L. (2004). Pathogenic Escherichia coli. Nat. Rev. Microbiol. 2, 123–140. doi: 10.1038/nrmicro818
Karmali, M. A. (1989). Infection by verocytotoxin-producing Escherichia coli. Clin. Microbiol. Rev. 2, 15–38. doi: 10.1128/CMR.2.1.15
Konowalchuk, J., Speirs, J. I., and Stavric, S. (1977). Vero response to a cytotoxin of Escherichia coli. Infect. Immun. 18, 775–779.
Leotta, G., Chinen, I., Epszteyn, S., Miliwebsky, E., Melamed, I. C., Motter, M., et al. (2005). Validación de una técnica de PCR múltiple para la detección de Escherichia coli productor de toxina Shiga. Rev. Arg. Microbiol. 37, 1–10.
Leotta, G. A., Deza, N., Origlia, J., Toma, C., Chinen, I., Miliwebsky, E., et al. (2006). Detection and characterization of Shiga toxin-producing Escherichia coli in captive non-domestic mammals. Vet. Microbiol. 118, 151–157. doi: 10.1016/j.vetmic.2006.07.006
MacFaddin, J. F. (2003). Pruebas Bioquímicas Para la Identificación de Bacterias de Importancia Clínica. Barcelona: Ed Médica Panamericana.
McDaniel, T. K., Jarvis, K. G., Donnenberg, M. S., and Kaper, J. B. (1995). A genetic locus of enterocyte effacement conserved among diverse enterobacterial pathogens. Proc. Natl. Acad. Sci. U.S.A. 92, 1664–1668. doi: 10.1073/pnas.92.5.1664
Nataro, J. P., and Kaper, J. B. (1998). Diarrheagenic Escherichia coli. Clin. Microbiol. Rev. 11, 142–201.
Navarro, A., and Estrada-García, T. (2010). “Epidemiology of diarrheagenic Escherichia coli pathotypes in Mexico, past and present,” in Pathogenic Escherichia coli, Latin America. ed. A. G. En Torres (Sharjah: Bentham Science Publishers), 191–208.
Nielsen, E. M., Skov, M. N., Madsen, J. J., Lodal, J., Jespersen, J. B., and Baggesen, D. L. (2004). Verocytotoxin-producing Escherichia coli in wild birds and rodents in close proximity to farms. Appl. Environ. Microbiol. 70, 6944–6947. doi: 10.1128/AEM.70.11.6944-6947.2004
Noda, M., Yutsudo, T., Nakabayashi, N., Hirayama, T., and Takeda, Y. (1987). Purification and some properties of Shiga-like toxin of Escherichia coli O157:H7 that is immunologically identical to Shiga toxin. Microb. Pathog. 2, 339–349. doi: 10.1016/0882-4010(87)90076-3
Nyholm, O., Halkilahti, J., Wiklund, G., Okeke, U., Paulin, L., Auvinen, P., et al. (2015). Comparative genomics and characterization of hybrid shigatoxigenic and enterotoxigenic Escherichia coli (STEC/ETEC) strains. PLOS ONE 10:e0135936. doi: 10.1371/journal.pone.0135936
Ochoa, T. J., Barletta, F., Contreras, C., and Mercado, E. (2008). New insights into epidemiology of enteropathogenic Escherichia coli infection. Natl. Inst. Health 102, 852–856. doi: 10.1016/j.trstmh.2008.03.017
Orskov, F., and Orskov, I. (1984). “Serotyping of Escherichia coli,” in En Methods in Microbiology, ed. T. Bergan (London: Academic Press Ltd.), 43–112. doi: 10.1016/S0580-9517(08)70447-1
Park, C. H., Kim, H. J., Hixon, D. L., and Bubert, A. (2003). Evaluation of the Duopath verotoxin test for detection of Shiga toxins in cultures of human stools. J. Clin. Microbiol. 41, 2650–2653. doi: 10.1128/JCM.41.6.2650-2653.2003
Paton, A. W., and Paton, J. C. (2002). Direct detection and characterization of shiga toxigenic Escherichia coli by multiplex PCR for stx1,stx2, eae, ehxA, and saa. J. Clin. Microbiol. 40, 271–274. doi: 10.1128/JCM.40.1.271-274.2002
Paton, A. W., and Paton, J. C. (2005). Multiplex PCR for direct detection of Shiga toxigenic Escherichia coli strains producing the novel subtilase cytotoxin. J. Clin. Microbiol. 43, 2944–2947. doi: 10.1128/JCM.43.6.2944-2947.2005
Paton, A. W., Srimanote, P., Talbot, U. M., Wang, H., and Paton, J. C. (2004). A new family of a potent AB5citotoxines produced by Shiga toxigenic Escherichia coli. J. Exp. Med. 200, 35–46. doi: 10.1084/jem.20040392
Paton, J. C., and Paton, A. W. (1998). Pathogenesis and diagnosis of Shiga toxin-producing Escherichia coli infections. Clin. Microbiol. Rev. 11, 450–479.
Pedroso, M. Z., Freymüller, E., Trabulsi, L. R., and Gomes, T. A. T. (1993). Attaching-Effacing lesions and intracellular penetration in Hela cells and human duodenal mucosa by two Escherichia coli strains not belonging to the classical Enteropathogenic E. coli serogroups. Infect. Immun. 61, 1152–1156.
Rahn, K., Renwick, S. A., Johnson, R. P., Wilson, J. B., Clarke, R. C., Alves, D., et al. (1997). Persistence of Escherichia coli O157:H7 in dairy cattle and the dairy farm environment. Epidemiol. Infect. 119, 251–259. doi: 10.1017/S0950268897007929
Rivas, M. (2016). Available at: http://www.sap.org.ar/uploads/archivos/files_dra-rivas-epidemiologia-del-sindrome-uremicohemolitico-en-argentina-situacion-actual-e-innovaciones-diagnosticas_1494446234.pdf
Rumi, M. V., Blanco Crivelli, X., Calviño, M. F., Regalía, A., Cueto, G., Degregorio, O., et al. (2012). Escherichia coli shigatoxigénica en animales relacionados con casos de diarreas sanguinolentas o síndrome urémico hemolítico y prevalencia de cepas STEC en roedores de la ciudad de Buenos Aires. Rev. Arg. Salud Pública 3, 23–29.
Scheutz, F., Teel, L. D., Beutin, L., Pierard, D., Buvens, G., Karch, H., et al. (2012). Multicenter evaluation of a sequence-based protocol for subtyping Shiga toxins and standardizing Stx nomenclature. J. Clin. Microbiol. 50, 2951–2963. doi: 10.1128/JCM.00860-12
Schmidt, H., Zhang, W. L., Hemmrich, U., Jelacic, S., Brunder, W., Tarr, P. I., et al. (2001). Identification and characterization of a novel genomic island integrated at selC in locus of enterocyte effacement-negative, Shiga toxin-producing Escherichia coli. Infect. Inmun. 69, 6863–6873. doi: 10.1128/IAI.69.11.6863-6873.2001
Toma, C., Espinosa, E. M., Song, T., Miliwebsky, E., Chinen, I., Iyoda, S., et al. (2004). Distribution of putative adhesins in different seropathotypes of Shiga toxin-producing Escherichia coli. J. Clin. Microbiol. 42, 4937–4946. doi: 10.1128/JCM.42.11.4937-4946.2004
Torres, A. G., Arenas-Hernández, M., and Martinez-Laguna, Y. (2010). “Overview of Escherichia coli,” in Pathogenic Escherichia coli, Latin America. ed. A. G. En Torres (Sharjah: Bentham Science Publishers), 1–7.
Underwood, W., Anthony, R., Gwaltney-Brant, S., Poison, A. S. P. C. A., and Meyer, R. (2013). AVMA Guidelines for the Euthanasia of Animals: 2013 Edition. Schaumburg, IL: American Veterinary Medical Association.
Vidal, M., Kruger, C., Duran, R., Lagos, R., Levine, M., Prado, V., et al. (2005). Single PCR assay to identify simultaneously the six categories of diarrheagenic Escherichia coli associated with enteric infections. J. Clin. Microbiol. 43, 5362–5365. doi: 10.1128/JCM.43.10.5362-5365.2005
Wales, D. A., Woodward, M. J., and Person, G. R. (2005). Attaching –effacing bacteria in animals. J. Comp. Pathol. 132, 1–26. doi: 10.1016/j.jcpa.2004.09.005
Wieler, L. H., Semmler, T., Eichhorn, I., Antao, E. M., Kinnemann, B., Geue, L., et al. (2011). No evidence of the Shiga toxin-producing E. coli O104:H4 outbreak strain o enteroaggregative E. coli (EAEC) found in cattle faeces in northern Germany, the hostpot of the 2011 HUS outbreak area. Gut Pathog. 3:17. doi: 10.1186/1757-4749-3-17
Keywords: Rattus, synanthropic, EPEC, STEC, childhood diarrhea
Citation: Blanco Crivelli X, Bonino MP, Von Wernich Castillo P, Navarro A, Degregorio O and Bentancor A (2018) Detection and Characterization of Enteropathogenic and Shiga Toxin-Producing Escherichia coli Strains in Rattus spp. from Buenos Aires. Front. Microbiol. 9:199. doi: 10.3389/fmicb.2018.00199
Received: 10 August 2017; Accepted: 29 January 2018;
Published: 14 February 2018.
Edited by:
Jacob Moran-Gilad, Ben-Gurion University of the Negev, IsraelReviewed by:
Eyal Klement, Koret School of Veterinary Medicine, IsraelTânia Aparecida Tardelli Gomes, Federal University of São Paulo, Brazil
Copyright © 2018 Blanco Crivelli, Bonino, Von Wernich Castillo, Navarro, Degregorio and Bentancor. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ximena Blanco Crivelli, eGJsYW5jb2NyaXZlbGxpQGZ2ZXQudWJhLmFy