 Gwenn M. M. Hennon
Gwenn M. M. Hennon María D. Hernández Limón
María D. Hernández Limón Sheean T. Haley
Sheean T. Haley Andrew R. Juhl
Andrew R. Juhl Sonya T. Dyhrman
Sonya T. Dyhrman- 1Lamont-Doherty Earth Observatory, Biology and Paleo Environment, Palisades, NY, United States
- 2Department of Earth and Environmental Sciences, Columbia University, New York, NY, United States
With rising atmospheric CO2, phytoplankton face shifts in ocean chemistry including increased dissolved CO2 and acidification that will likely influence the relative competitive fitness of different phytoplankton taxa. Here we compared the physiological and gene expression responses of six species of phytoplankton including a diatom, a raphidophyte, two haptophytes, and two dinoflagellates to ambient (~400 ppm) and elevated (~800 ppm) CO2. Dinoflagellates had significantly slower growth rates and higher, yet variable, chlorophyll a per cell under elevated CO2. The other phytoplankton tended to have increased growth rates and/or decreased chlorophyll a per cell. Carbon and nitrogen partitioning of cells shifted under elevated CO2 in some species, indicating potential changes in energy fluxes due to changes in carbon concentrating mechanisms (CCM) or photorespiration. Consistent with these phenotypic changes, gene set enrichment analyses revealed shifts in energy, carbon and nitrogen metabolic pathways, though with limited overlap between species in the genes and pathways involved. Similarly, gene expression responses across species revealed few conserved CO2-responsive genes within CCM and photorespiration categories, and a survey of available transcriptomes found high diversity in biophysical CCM and photorespiration expressed gene complements between and within the four phyla represented by these species. The few genes that displayed similar responses to CO2 across phyla were from understudied gene families, making them targets for further research to uncover the mechanisms of phytoplankton acclimation to elevated CO2. These results underscore that eukaryotic phytoplankton have diverse gene complements and gene expression responses to CO2 perturbations and highlight the value of cross-phyla comparisons for identifying gene families that respond to environmental change.
Introduction
Anthropogenic CO2 emissions have increased the concentration of CO2 in the atmosphere from ~280 ppm at the start of the Industrial Revolution to ~400 ppm at present (Le Quéré et al., 2012), with levels predicted to rise to ~800 ppm by 2100 (Ciais et al., 2013). CO2 in the atmosphere acts as a greenhouse gas, warming the planet, and it dissolves into the oceans decreasing the concentration of carbonate ions and acidifying the water (Feely et al., 2004; Sabine et al., 2004). These changes in carbonate chemistry, called ocean acidification, can increase dissolution of carbonates, favor photosynthesis, and alter other cellular processes (Doney et al., 2009). For example, the differential impact of ocean acidification on phytoplankton growth rates is predicted to have a greater impact on phytoplankton community composition and global primary production than ocean warming and decreases in nutrient fluxes from stratification (Dutkiewicz et al., 2015).
Eukaryotic phytoplankton, in particular, play important roles in aquatic biogeochemical cycles and marine ecosystems. For example, diatoms alone are responsible for ~20% of global primary production (Nelson et al., 1995). Coastal and upwelling ecosystems, where larger eukaryotic phytoplankton thrive, host some of the largest fisheries through efficient trophic transfer (Ryther, 1969). Diatoms and coccolithophores also create biominerals that serve to transport silica (Nelson et al., 1995) and calcium carbonate (Milliman, 1993) with organic carbon from the surface to the deep ocean (Ducklow et al., 2001), sequestering this carbon for potentially thousands of years. In addition, some species of phytoplankton can cause harmful algal blooms (HABs) that disrupt ecosystems and human activities by growing to high concentrations or producing toxins (Trainer and Yoshida, 2014). Dissolved CO2 in the oceans is highly variable on regional and seasonal scales (Takahashi et al., 2014) and can be particularly high in coastal upwelling regions (Feely et al., 2008) where eukaryotic phytoplankton are abundant, making it likely that coastal eukaryotic phytoplankton regularly experience large fluctuations in CO2. Phytoplankton can change their carbon affinity based on acclimation to different concentrations of CO2 (Rost et al., 2003), but this acclimation effect varies by species. Predicting how the future structure of marine ecosystems will change with rising CO2 requires an understanding of how shifting carbonate chemistry will impact phytoplankton from diverse eukaryotic lineages (Hallegraeff, 2010; Dutkiewicz et al., 2015).
All phytoplankton share the rate-limiting enzyme of carbon fixation, ribulose-1,5-bisphosphate carboxylase/oxygenase (Rubisco), though the form differs in different groups, e.g., form II in most dinoflagellates (Morse et al., 1995) and form 1D for other red algal lineages (Tabita et al., 2008). Rubisco originated under much higher ambient CO2 and low oxygen levels and evolved in eukaryotic phytoplankton lineages as oxygen increased and CO2 levels decreased (Tortell, 2000; Young et al., 2012). Rubisco remains vulnerable to fixing oxygen in the place of CO2, resulting in the loss of fixed carbon—a metabolic pathway known as photorespiration (Bauwe et al., 2010). Although photorespiration is typically considered to be detrimental to photoautotrophs (Bauwe et al., 2010), it can also serve as a mechanism for phytoplankton to eliminate excess energy in times of high light or variable nutrient environments (Parker and Armbrust, 2005; Bagby and Chisholm, 2015). Rubisco form 1D and II differ in structural (Tabita et al., 2008) and kinetic properties (Badger et al., 1998); form II Rubisco has fewer subunits and a relatively higher rate of oxygen fixation compared with form 1D Rubisco.
Nearly all marine phytoplankton have carbon concentrating mechanisms (CCM) to increase the concentration of carbon around the active site of Rubisco by ~3 to 80-fold in order to favor carbon fixation and prevent oxygen fixation (Badger et al., 1998). The genes involved in CCMs remain largely uncharacterized for most species. Evidence for fixation of carbon into a four-carbon compound (C4 metabolism) has been described for a few species of diatoms (Reinfelder et al., 2000; McGinn and Morel, 2007) and for a coccolithophore (Tsuji et al., 2012). Yet the evidence remains mixed as to whether C4 compounds can be used as a CCM in phytoplankton (Kroth et al., 2008) and whether C4 enzymes are regulated by CO2 (Roberts et al., 2007; Kustka et al., 2014). Biophysical CCMs consisting of, at minimum, a bicarbonate pump and a carbonic anhydrase are thought to be potentially more efficient than C4 CCMs (Hopkinson et al., 2011), yet the genes that could be used for biophysical CCMs are multi-copy and diverse even within a single species (Tachibana et al., 2011), making a systematic study across phytoplankton lineages challenging. Large uncertainties persist in the range of potential CCM genes in phytoplankton and what trade-offs might result from regulating CCMs in response to rising CO2. Decreased expression of biophysical (Hennon et al., 2015) and C4 CCM gene expression (Kustka et al., 2014) has been observed in the diatom, Thalassiosira pseudonana, in response to elevated CO2. Concurrent rearrangements in metabolic pathways and physiology have also been observed in T. pseudonana (Hennon et al., 2014, 2015), likely as a result of metabolic rearrangement with a decreased need for the energy-consuming CCM. Similar responses for a few well-studied coccolithophores (Rokitta et al., 2012) have been reported. It is an open question how many other ecologically-important species of phytoplankton will alter their CCM and photorespiration genes in response to CO2 and what impact this will have on core metabolic processes and growth rate responses.
To examine how CCM and photorespiration genes are modulated in response to CO2 and what impact this has on core metabolic processes and growth rate responses, six eukaryotic phytoplankton species from four phyla were cultured under elevated (~800 ppm) and ambient (~400 ppm) CO2. These phytoplankton included: a cosmopolitan diatom (Chaetoceros affinis), a raphidophyte (Heterosigma akashiwo), a calcifying haptophyte (Gephyrocapsa oceanica), a non-calcifying haptophyte (Chrysochromulina polylepis), and two dinoflagellates (Alexandrium monilatum and Prorocentrum minimum). All species are ecologically important marine phytoplankton, with four documented to cause HABs: A. monilatum (Juhl, 2005; Anderson et al., 2012), P. minimum (Heil et al., 2005), H. akashiwo (Honjo, 1992), and C. polylepis (Dahl et al., 1989). Changes in physiology and gene expression in response to elevated CO2 were evaluated for all six phytoplankton to determine the impact of future ocean chemistry on these phyla. Additionally, a comprehensive analysis of all consensus sequences from the same four phyla within the marine microbial eukaryote transcriptome project (MMETSP) database was conducted to assess the potential for these phytoplankton lineages to acclimate to rising CO2 based on the expressed gene complement of CCM and photorespiration pathways.
Methods
Culturing Conditions
Experiments were performed on six species of phytoplankton: A. monilatum (CCMP3105), C. affinis (CCMP159), C. polylepis (CCMP1757), G. oceanica (RCC1303), H. akashiwo (CCMP2393), and P. minimum (CCMP1329), available from the National Center for Marine Algae and Microbiota (NCMA, formerly CCMP) and Roscoff Culture Collections (RCC). All cultures were uni-algal and uni-eukaryotic, but not axenic. Strains were cultured in L1 media (with silica for the diatom C. affinis), except G. oceanica which is native to open ocean habitats and was grown in a reduced nutrient medium (L1/25 vitamins and trace metals with L1/10 nitrate and L1/15 phosphate), to ensure calcification. The natural seawater for the media base for all species was collected from Vineyard Sound, MA (salinity 32). Cultures were grown at 18°C, except for species with different temperature optima: A. monilatum (24°C) and C. polylepis (15°C). Light levels in all experimental treatments were ~100 μmol photon m−2 s−1 with a 14:10 light:dark cycle. Culture conditions are summarized in Supplementary Table 1.
Strains were acclimated to experimental conditions in 100-mL culture flasks for 3 days (or ~1–3 generations). Triplicate experimental batch cultures were inoculated with 25 mL of acclimated culture. Experimental batch cultures were grown in 1-L volumes in 2.5-L polycarbonate bottles under gentle rotation (75 rpm). To control the carbonate chemistry of the water, the headspace of each bottle was purged continuously with either ambient outside air of 400 ppm (±15 ppm) checked with a LI-7200RS CO2 Analyzer (LI-COR, Lincoln NB, USA) or a custom gas mix (TechAir, NY, USA) of 800 ppm (± 12 ppm) carbon dioxide with 21% oxygen and balance nitrogen. The gas streams were pre-filtered through 0.2-μm HEPA filters and directed through a sterile glass pipette to break the boundary layer of the media without contacting the surface. The bottles were not bubbled to avoid the adverse effects of turbulence on phytoplankton growth (Thomas and Gibson, 1990; Juhl and Latz, 2002). Cell concentration and carbonate chemistry were monitored over the course of the experiments, which were typically 4–5 days. At this point, experimental batch cultures were harvested for physiological and gene expression measurements. This approach both constrained the biomass to ecologically-relevant levels and permitted the calculation of growth rates with confidence.
Carbonate Chemistry
Differences in media carbonate chemistry were confirmed by measuring pH using an electrode (300728.1, Denver Instruments) calibrated with NIST buffers (pH 4–10). All cultures were grown in media from the same base seawater from Vineyard Sound, Massachusetts with salinity (S) of 32. The total alkalinity (TA) was estimated with an empirical formula derived from an offshore transect from Woods Hole, MA to the mid-Atlantic Bight (TA = 73.4 × S −188.7) (Cai et al., 2010). Calculations of carbonate chemistry parameters were performed by CO2SYS (Lewis and Wallace, 1998) and summarized in Supplementary Table 1. Although all species were grown at ecologically-relevant cell concentrations with headspace continuously purged with elevated (~800 ppm) or ambient (~400 ppm) CO2 (Supplementary Table 1), aqueous CO2 was drawn down by the phytoplankton as they fixed carbon, resulting in lower mean dissolved CO2 concentrations compared to the headspace for some species. Thus, the changes in dissolved CO2 concentrations varied between species (Supplementary Table 1). Despite this variability, dissolved CO2 concentrations exceeded 400 ppm in all flasks purged with elevated CO2 and the dissolved CO2 levels in ambient and elevated treatments for each species were offset from each other, with the difference ranging 0.2–0.5 pH units and equating to a difference of ~200–600 ppm dissolved CO2 (Supplementary Table 1). For clarity of the manuscript, the treatments are referred to as elevated (~800 ppm) or ambient (~400 ppm) CO2 throughout the text.
Physiological Measurements
Daily, 1 mL of each triplicate culture was preserved in 2% Lugols solution (final concentration) for microscopic cell counts. Growth rate within each flask was calculated as the slope of the natural log of biomass (cell abundance) vs. time. There was no evidence of a lag phase in any of the six species, suggesting cultures were appropriately acclimated (Berge et al., 2010), but growth rate was conservatively calculated from only the last 3–5 days of the experiment to most closely represent growth rate at harvest. For harvest, volumes of 10, 25, and 50 mL of each culture were gently filtered on 25-mm, pre-combusted glass fiber filters (Whatman, GF/F) and frozen at −20°C until processing for chlorophyll a, total particulate carbohydrates and elemental analyses respectively. Chlorophyll a concentration was determined according to Strickland and Parsons (1972). Briefly, the samples were extracted in 10 mL of 90% acetone, vortexed for 15 s, and stored in the dark at −20°C for 12 h. The fluorescence in the supernatant was then measured on a Turner Designs Aquafluor fluorometer before and after acidification with 0.1 N hydrochloric acid. The fluorometer was calibrated with chlorophyll a from Anacystis nidulans (Sigma). Total particulate carbohydrates (pCHO) were measured according to a modified phenol-sulfuric acid method (DuBois et al., 1956). Filters were extracted in 5% phenol and concentrated sulfuric acid in 15-mL polypropylene tubes at 30°C for 12 h. After removal of the filter, light absorption of the sample at 490 nm was measured using a Hach DR2700 spectrophotometer and compared to a glucose calibration curve. Elemental analysis of particulate organic carbon and nitrogen (POC:PON) was performed by the Nutrient Analytical Services Laboratory at the Chesapeake Bay Laboratory (University of Maryland, Solomon, MD) with a CE-440 Elemental Analyzer following the methods of USEPA (1997). All physiological measurements were normalized to cell concentration from their respective flask.
RNA Extraction and Sequencing
At the time of harvest, triplicate 150-mL subsamples of each replicate in each treatment were filtered onto polycarbonate filters (47 mm, 0.2–3 μm depending on species). Filters were immediately flash frozen and stored in liquid nitrogen until extraction. RNA extractions were performed with RNeasy Mini Kit (Qiagen, Valencia, CA), with a slight modification to the lysis procedure. Lysis was performed by adding 1.4 mL of Buffer RLT and ~250 μL zirconium/silica beads (0.5 mm) and vortexing for 1 min at 250 rpm. DNA was removed using TurboDNase (Ambion, Austin, TX). Total RNA extracts of the triplicate cultures for each treatment were pooled for sequencing, quantified using a Qubit fluorometer (Invitrogen, Carlsbad, CA) and verified to be high quality using a 2100 Bioanalyzer (Agilent, Santa Clara, CA). The sequencing libraries were prepared using the Illumina TruSeq RNA Sample Preparation Kit to select mRNA with polyA tails. Library preparation and sequencing of ~50-bp, paired-end reads from each library was performed on an Illumina HiSeq 2000 at the National Center for Genome Resources (Sante Fe, NM).
Sequence Assembly and Alignment
Combined assemblies and alignments for sequences from each phytoplankton strain were generated as part of the Marine Microbial Eukaryote Transcriptome Project (MMETSP) described in detail in Keeling et al. (2014). Briefly, sequence reads were pre-processed using SGA (Simpson and Durbin, 2012). After trimming, reads less than 25 bp in length were discarded. Quality-controlled sequences were assembled into contigs with ABySS (Simpson et al., 2009) for each sample. The resulting contigs from all samples from a single strain were combined into one consensus assembly using a combination of assembly tools: CD-HIT-EST (Li and Godzik, 2006), ABySS, CAP3 (Huang and Madan, 1999), GapCloser (SOAP, Li et al., 2008), with redundant contigs removed at a threshold of 98% identity. Reads were aligned to combined assemblies with BWA (Li and Durbin, 2009). Combined assemblies and alignment counts are available from iMicrobe (ftp://ftp.imicrobe.us/camera/combined_assemblies/) or raw reads from BioProject (PRJNA248394) for IDs: A. monilatum (control, MMETSP0093; elevated CO2 MMETSP0097), C. affinis (control, MMETSP0088; elevated CO2, MMETSP0092), C. polylepis (control, MMETSP0143; elevated CO2, MMETSP0147), G. oceanica (control, MMETSP1363; elevated CO2, MMETSP1364), H. akashiwo (control, MMETSP0292; elevated CO2, MMETSP0296), P. minimum (control, MMETSP0053; elevated CO2, MMETSP0057).
Read Count Normalization and Annotation
Contig counts resulting from sequence assembly and alignment were normalized using the trimmed mean of M-values (function tmm, NOISeq package, R) and then normalized to read counts from ambient CO2 controls. Translated contigs (peptide sequences) were annotated with GhostKOALA (Kanehisa et al., 2016) using the KEGG database of genus-level prokaryotic and family-level eukaryotic sequences.
Orthologous Group Clustering and Comparison
Consensus contigs from the six eukaryotic phytoplankton species were clustered into orthologous groups by OrthoFinder (Emms and Kelly, 2015), and orthologous groups represented in at least two species were kept for further analysis. Orthologous groups that had a greater than 2-fold change (|log2FC| > 1) under elevated compared to ambient CO2 and total of 10 reads or greater across both treatments (>−16 log2 counts per million) were then compared across the six species. Overlaps in CO2-responsive orthologous groups were visualized with UpSet (http://caleydo.org/tools/upset/).
CCM and Photorespiration Gene Identification
Putative CCM genes, biophysical and C4, and photorespiration pathway genes were compiled from the literature. Hidden Markov Model (HMM) seed files were created for each gene using peptide sequences from complete genomes of at least four species belonging to at least three phyla. Seed file genes, applicable citations (Branson et al., 2004; Jitrapakdee et al., 2008; Kroth et al., 2008; Milenkovic et al., 2008; Hackenberg et al., 2011; Tachibana et al., 2011; Tsuji et al., 2012; Nakajima et al., 2013; Romero et al., 2013; Chi et al., 2014; Hennon et al., 2015), and their accession IDs from GenBank are detailed in Supplementary Table 2. Multiple alignments of HMM seed files were performed by MAFFT (Katoh and Standley, 2013) (on setting auto) and checked with Jalview (Waterhouse et al., 2009) (version 2) visualization software. HMM searches for each seed file were performed on translated contigs using HMMER (Eddy, 2011) (version 3.1b2, HMMbuild, HMMsearch) with an E-value cut-off of 10−4, the results are summarized in Supplementary Table 3 and contig IDs for each HMM hit are listed in Supplementary Table 4. Putative CCM and photorespiration genes were identified from the six eukaryotic phytoplankton species as well as all available consensus contigs from the same four phyla in the MMETSP database of combined assemblies (ftp://ftp.imicrobe.us/camera/combined_assemblies/).
Statistical Analysis
Differences in physiological measurements between ambient and elevated CO2 for each species were tested with student's t-test (t-test, stats package, R). Due to the pooling of biological replicates in each of the six eukaryotic phytoplankton transcriptomes the significance of differential expression for individual genes was not calculated. Rather, a cut-off of greater than 2-fold change (|log2FC| > 1), thought to represent a substantial biological response (Mock et al., 2008; Shrestha et al., 2012), and a minimum of 10 reads or greater across both treatments (>−16 log2 counts per million) was used to define contigs with increased and decreased expression under elevated CO2. Rather than comparing expression for individual genes, analyses focused on whether changes in sets of genes were statistically significant between treatments. Gene set enrichment, the fraction of pathway genes present in increased or decreased contigs compared to total contigs, was tested for 10 sets defined by KEGG pathways involved in energy, carbon and nitrogen fluxes: photosynthesis (00195), photosynthesis antenna proteins (00196), porphyrin and chlorophyll metabolism (00860), carbon fixation in photosynthetic eukaryotes (00710), tricarboxylic acid cycle (00020), glycolysis and gluconeogenesis (00010), nitrogen metabolism (00910), fatty acid elongation (00062), fatty acid degradation (00071), and biosynthesis of unsaturated fatty acids (01040). The hypergeometric test (phyper, stats, R) was used to test for statistical significance. To test whether the CCM and photorespiration gene sets were significantly differently distributed compared to all genes, two-sample Kolmogorov-Smirnov tests (ks_2samp, SciPy library, Python) were used. Custom python scripts are available on github (https://github.com/MDHDZ91).
Results and Discussion
Section 1: CO2 Physiology of Six Eukaryotic Phytoplankton Species
Six eukaryotic phytoplankton species were grown in nutrient replete media under headspaces purged with elevated (~800 ppm) or ambient (~400 ppm) CO2 (Supplementary Table 1). Changes in growth rate and chlorophyll a per cell showed high variability between species (Table 1). The dinoflagellates, A. monilatum, and P. minimum, both grew significantly slower in elevated CO2 (Table 1), averaging 0.67 ± 0.07 (mean ± 1SE) of the growth rate at ambient CO2 (Figure 1). The chlorophyll a content trended higher under elevated CO2 in both dinoflagellates, although it was highly variable within biological replicates and not statistically significantly different between CO2 treatments (Figure 1, Table 1). In contrast, C. polylepis and H. akashiwo had statistically significant increases in growth rate (Table 1) consistent with previous studies of large diatoms (Wu et al., 2014). Only one species, H. akashiwo, had significantly less chlorophyll a per cell (Table 1). The differences in the impact of CO2 on growth and chlorophyll content in these species may be driven by differences in cell size, Rubisco form or gene complement, disentangling the mechanisms driving physiological differences will be important to accurately predict the future competitive fitness of different groups of eukaryotic phytoplankton.
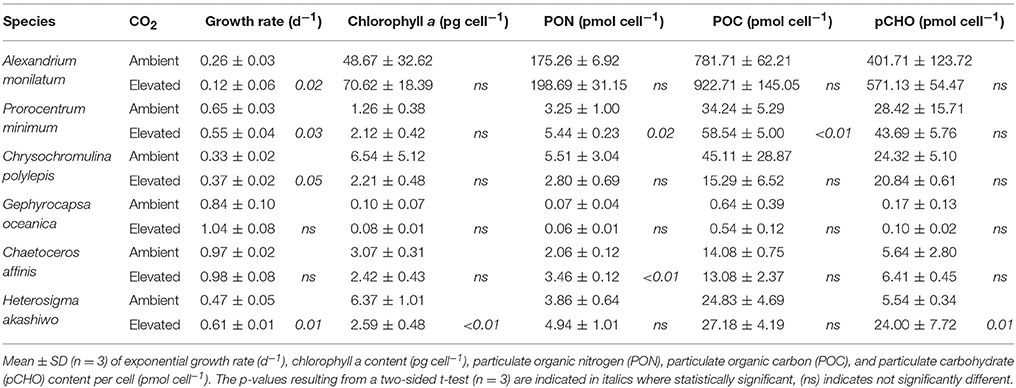
Table 1. Physiology of six eukaryotic phytoplankton grown under ambient and elevated CO2.
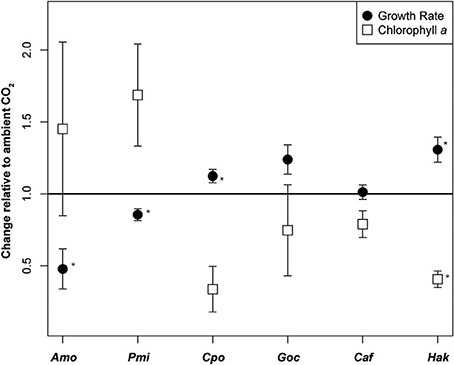
Figure 1. Growth rate response and change in chlorophyll a content under elevated CO2. Change in growth rate (d−1) and chlorophyll a (pg cell−1) under elevated relative to ambient CO2 in six species of phytoplankton. Black circles indicate growth rate response and squares indicate the change in chlorophyll a content per cell (mean ± SE). An asterisk represents statistically significant differences in elevated vs. ambient CO2 (p < 0.05, two-sided t-test, n = 3). Amo, Alexandrium monilatum; Pmi, Prorocentrum minimum; Cpo, Chrysochromulina polylepis; Goc, Gephyrocapsa oceanica; Caf, Chaetoceros affinis; Hak, Heterosigma akashiwo.
The carbon and nitrogen composition of these six phytoplankton species was also examined for changes under elevated CO2 (Figure 2, Table 1). P. minimum was the only species that showed a significant change in POC per cell, while the raphidophyte, H. akashiwo, was the only species showing significant differences in pCHO per cell and in pCHO:POC. The dinoflagellates did not significantly alter their C:N ratios, while C:N of phytoplankton from other lineages dropped with elevated CO2, although this change was only significant in the diatom, C. affinis, and the coccolithophore, G. oceanica (Figure 2). For C. affinis, the shift in C:N was driven by a significant increase in nitrogen cell quota (Table 1), suggesting an increase in nitrate assimilation rate. These results differ from previous studies of phytoplankton communities in mesocosms (Riebesell et al., 2007), yet are similar to the findings of Eberlein et al. (2016) for dinoflagellates suggesting that there may be a wide range of species-specific responses in C:N ratio to elevated CO2. It is important to understand changes in phytoplankton elemental composition for predicting future biogeochemistry and marine food web dynamics, as a decrease in C:N ratio could decrease the efficiency of carbon sequestration (Riebesell et al., 2007) while improving quality of phytoplankton as food for higher trophic levels (Elser et al., 2000).
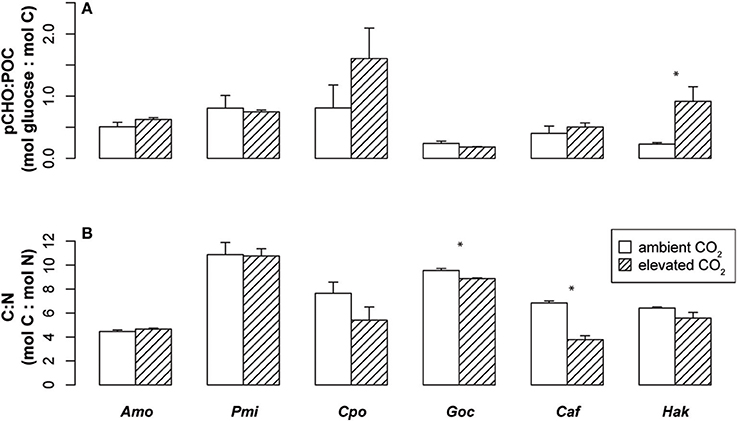
Figure 2. Carbon and nitrogen composition of phytoplankton under ambient and elevated CO2. Mean ± SE of (A) the ratio of carbohydrate (pCHO units of pmol glucose) to total particulate organic carbon (POC units of pmol carbon) and (B) carbon to nitrogen (C:N) for six species of phytoplankton. Asterisks indicate significant differences in means between ambient and elevated CO2 (p < 0.05, n = 3, two-sided t-test). Abbreviations are as in Figure 1.
Section 2: CO2-Induced Changes in Gene Expression
Transcriptomes were sequenced for each species to investigate how these phytoplankton shift metabolic pathways to acclimate to elevated CO2. The changes in core carbon and nitrogen metabolic pathways differed among the six species (Figure 3). C. polylepis and H. akashiwo were the two species with the highest number of gene sets with significantly increased expression, including many carbon and nitrogen metabolism genes, particularly biosynthesis of unsaturated fatty acids (Figure 3A). The increased expression of these biosynthesis genes in C. polylepis and H. akashiwo, which also had significantly higher growth rates under elevated CO2, likely corresponded to faster rates of lipid biosynthesis. Both C. polylepis and H. akashiwo also had significant enrichment of Glycolysis/Gluconeogenesis pathway genes without a corresponding significant enrichment of TCA pathway genes (Figure 3A), suggesting that the genetic underpinning for accumulation of carbohydrates in these two species (Figure 2A) may stem from favoring gluconeogenesis over glycolysis and respiration. The diatom, C. affinis, had significantly enriched expression of fatty acid elongation as well as degradation genes (Figure 3A) with no significant change in growth rate (Table 1), suggesting there may be futile cycling of lipid synthesis and degradation. C. affinis also displayed decreased expression of nitrogen and chlorophyll metabolism genes with significant gene set enrichment (Figure 3B), corresponding to the significant shift in C:N ratio and lower chlorophyll a content (Table 1). G. oceanica presented a more muted gene expression response compared to other form 1D Rubisco species with fewer significant changes in gene set enrichment including a decrease in fatty acid degradation pathway gene expression (Figure 3B). This muted metabolic gene expression response corresponded with a relatively minor growth rate response for G. oceanica to elevated CO2 (Table 1).
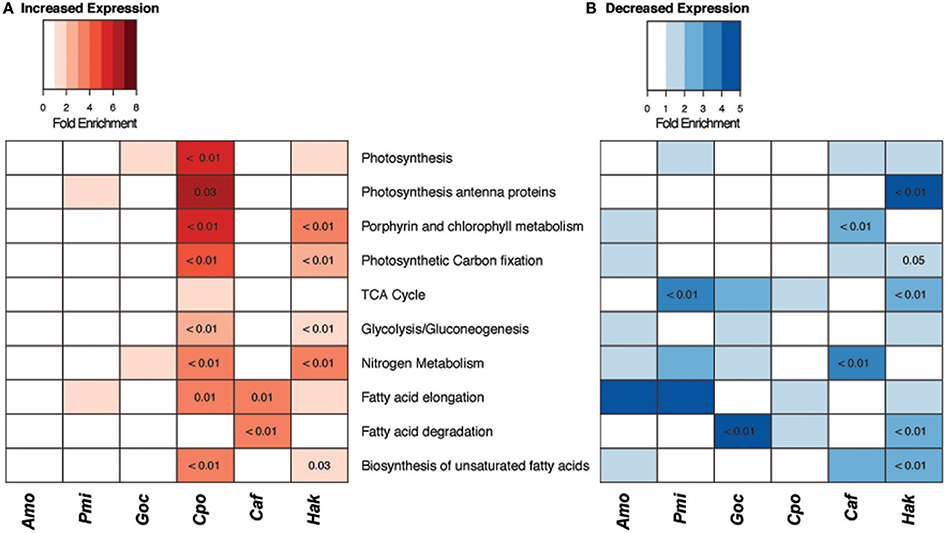
Figure 3. Changes in carbon and nitrogen metabolism gene expression under elevated CO2. Gene set enrichment of carbon and nitrogen KEGG pathways with (A) increased and (B) decreased expression (|log2FC| > 1, logCPM > −16) for six species of phytoplankton. Fold enrichment of each category is indicated by color intensity, numbers indicate p-values for gene sets with significant enrichment (p < 0.05, hypergeometric test).
The dinoflagellates, A. monilatum, and P. minimum, also displayed few significant changes in carbon and nitrogen pathway gene expression (Figure 3). The exception to this being the reduced expression of TCA cycle genes by P. minimum, which may help to explain the decrease in growth rate (Figure 1). A limited transcriptional response of the dinoflagellates has been observed in experiments with nutrient limitation (Harke et al., 2017) and elevated temperatures (Barshis et al., 2014). The limited transcriptional response to environmental perturbations may be largely explained by the underlying regulatory mechanisms of cell physiology occurring at the translational or post-translational level for an estimated ~73–90% of dinoflagellate genes (Lin, 2011; Murray et al., 2016). Regardless, the limited change in carbon and nitrogen pathway genes for this group is consistent with their lack of significant change in carbon and nitrogen partitioning (Figure 2).
To investigate whether there were any genes not classified into KEGG pathways that responded similarly to CO2 across species, the genes were also grouped into orthologous clusters. Despite similarities in physiological responses between species (Table 1), there was almost no shared expression change in orthologous genes (Figure 4). No orthologs showed an expression pattern that was shared by more than three of the six species examined, and those that did share expression patterns among three species were frequently hypothetical proteins with unidentified function or genes with general functions such as transcription (e.g., elongation factor) and oxidoreductase activity (e.g., aldo/keto reductase) (Figure 4). A few of the genes that had decreased expression across three species (Figure 4B) have putative roles in photorespiration (isocitrate lyase, ICL), C4 metabolism (phosphoenolpyruvate carboxykinase, PEPCK), and ion balance (ion channel). Decreased expression of putative CCM and photorespiration genes with elevated CO2 agrees with previous work with diatoms (Kustka et al., 2014; Hennon et al., 2015). Nevertheless, the analysis of metabolic gene sets and orthologous groups suggest limited commonality in the transcriptional response of these six species to elevated CO2.
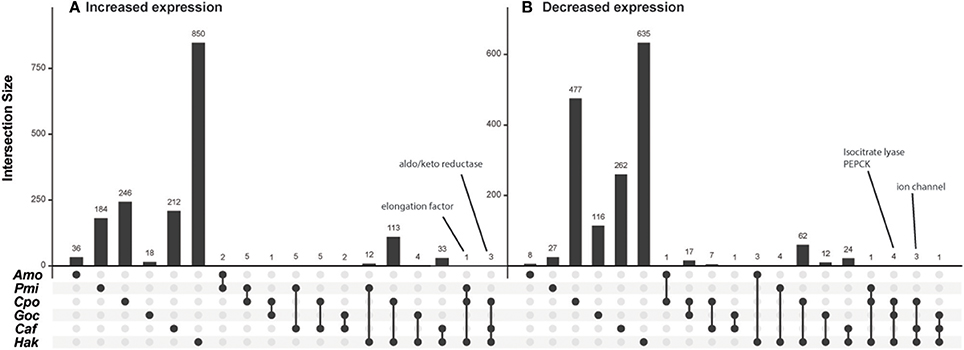
Figure 4. Intersection of orthologous genes with shared expression patterns under elevated CO2 for phytoplankton species. Intersection size (number of shared orthologous genes) is indicated by bars for each grouping of species indicated by circles below the axis. Intersections of genes with (A) increased expression (log2 Fold Change > 1) and (B) decreased expression (log2 Fold Change < −1). Intersections with no shared orthologous genes are omitted, species abbreviations are as in Figure 1.
To more comprehensively explore the possible CO2-related shifts in CCM and photorespiration genes, Hidden Markov Model (HMM) searches were performed to identify genes that match the patterns of known homologs (Supplementary Table 2). The expression patterns of CCM and photorespiration homologs were compared to the distribution of all genes (Figure 5). Similar to the other gene expression and physiology measures, there were a large variety of responses between species. The two dinoflagellates, A. monilatum, and P. minimum, had significantly different distributions of CCM and photorespiration gene expression compared to all expressed genes (K-S test, p < 0.05, Figures 5A,B). However, few genes in the dinoflagellates had fold changes greater than the 2-fold change cut-off to be canonically considered biologically significant (Table 2). The median expression of CCM and photorespiration genes declined by 6–8% in A. monilatum and 11–16% in P. minimum, agreeing with the expectation that elevated CO2 would result in a decrease in the use of these pathways and with previous studies showing a decrease in carbonic anhydrase expression in other dinoflagellates (Van de Waal et al., 2013, 2014). These data suggest that CCM and photorespiration genes may be responding to elevated CO2. However, the growth rate effects (Table 1, Figure 1) are opposite of expectations for a decreased CCM, leaving the question open as to why dinoflagellates in this study grew more slowly under elevated CO2. One possibility is that P. minimum and A. monilatum may be sensitive to changes in the carbon dioxide to oxygen ratio rather than simply to elevated CO2 (Bagby and Chisholm, 2015), due to an unrecognized importance of photorespiration. More research is needed to determine whether photorespiration may play an important role in dinoflagellate metabolism.
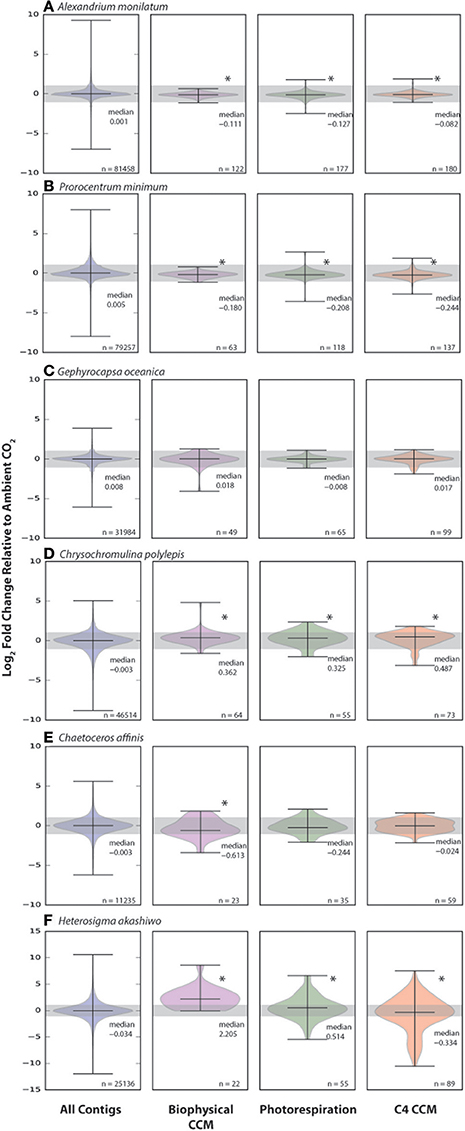
Figure 5. Change in distribution of carbon concentrating mechanism (CCM) and photorespiration genes in six eukaryotic phytoplankton under elevated CO2. Distributions of the expression (log2 Fold Change) at elevated CO2 relative to ambient CO2 of all expressed contigs, biophysical CCM, photorespiration pathway genes and C4 CCM genes for Alexandrium monilatum (A), Prorocentrum minimum (B), Gephyrocapsa oceanica (C), Chrysochromulina polylepis (D), Chaetoceros affinis (E), Heterosigma akashiwo (F). Asterisks indicate significant difference in expression distribution from that of all expressed contigs (Kolmogorov-Smirnov test, p < 0.05). Distribution median and number of genes per distribution (n) given on the figure.
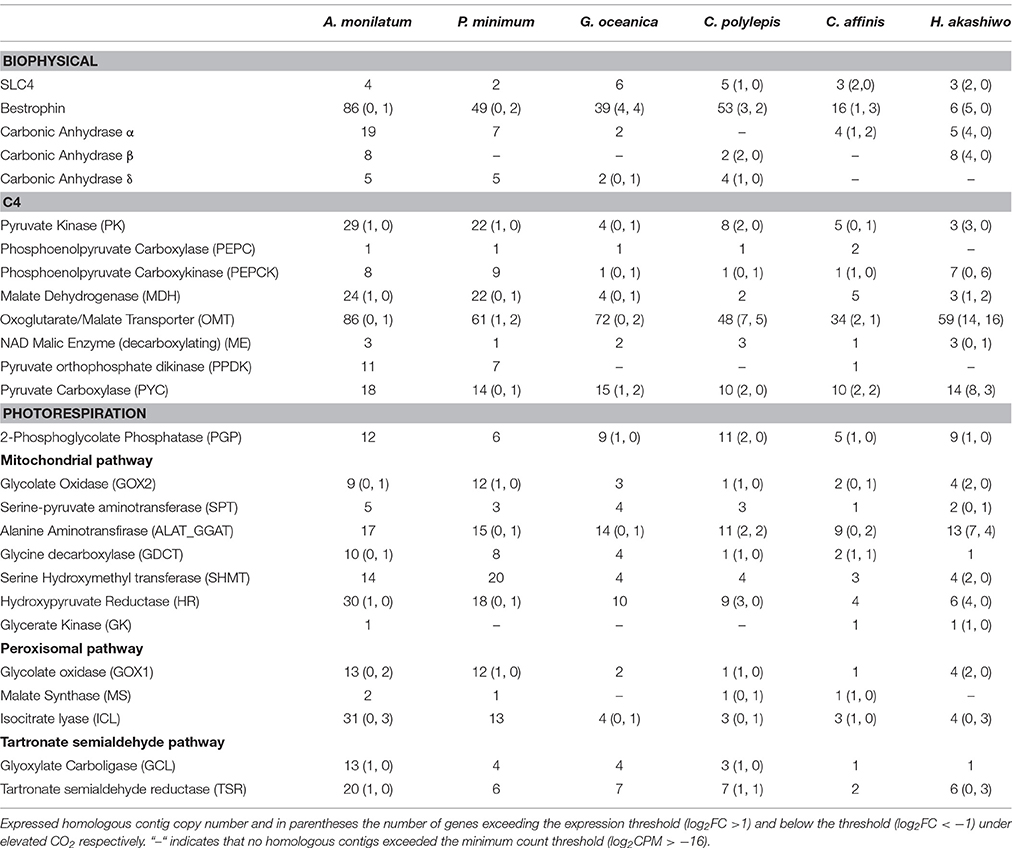
Table 2. Gene expression of putative CCM and photorespiration genes in six eukaryotic phytoplankton.
The calcifying haptophyte, G. oceanica, displayed no significant difference in CCM or photorespiration gene expression distribution with CO2 (Figure 5C). This differed markedly from the non-calcifying haptophyte, C. polylepis (Figure 5D), which had significantly different distributions of CCM and photorespiration gene sets with a higher median expression of 25–40%, contrary to expectations of a decreased need for CCM and photorespiration genes under elevated CO2. In previous studies with coccolithophores, changes in CCM gene expression and growth rates were only stimulated at much higher CO2 levels (Rickaby et al., 2010; Lohbeck et al., 2014). In another study, only a few of the many putative CCM genes were down-regulated in coccolithophores under similar CO2 concentrations (Rokitta et al., 2012). Thus, it is possible that coccolithophores may have a threshold response to changing CO2 that is only triggered by much larger changes in CO2 or that the CCM is controlled by a smaller subset of genes, whose expression is obscured by examining the distribution of all putative CCMs. We speculate that the process of calcification could also uncouple coccolithophores from the impacts of changing CO2 by maintaining the need for an energy consuming bicarbonate pump under a wider range of CO2 concentrations. This hypothesis is supported by the lack of biologically significant change in the six SLC4 bicarbonate transporter copies for G. oceanica (Table 2), but warrants further study.
The diatom, C. affinis, significantly shifted the expression of biophysical CCM genes, with the median decreasing by 35% (Figure 5E). The decrease in C. affinis biophysical CCM genes with elevated CO2 agrees with previous work on the diatom, T. pseudonana (Hennon et al., 2014). It may be that the energy saved by decreased expression of biophysical CCM genes (Figure 5, Table 2) allowed for the increase in nitrogen assimilation per cell by C. affinis (Table 1), similar to observations of the effects of energy flux on diatom nitrate assimilation (Parker and Armbrust, 2005) and dinoflagellate nitrate assimilation (Eberlein et al., 2016). H. akashiwo showed the greatest relative changes in CCM, photorespiration and C4 CCM genes, with a greater than 4-fold increase in median biophysical CCM and a 42% increase in photorespiration, coupled to a 20% decrease in the median C4 CCM gene set expression (Figure 5F). Both CCM gene sets were significantly different in expression distribution when compared to the total gene set. The increase in expression of putative biophysical CCM and photorespiration genes for both H. akashiwo and C. polylepis is surprising, however it is important to note that the genes in these categories may have multiple functions including acid-base balance of the cell and functions in other metabolic pathways. This re-shuffling of H. akashiwo gene expression away from putative C4 genes may have the net impact of increasing energy equivalents for growth and carbon fixation, as evidenced by significantly increased growth rate (Table 1).
Few CCM and photorespiration genes were found to share expression patterns even among the more closely related species (Table 2). All phytoplankton differentially expressed at least one copy of pyruvate kinase (PK), an enzyme central to many metabolic processes including C4 metabolism. There were also changes in phosphoenolpyruvate carboxykinase (PEPCK) and pyruvate carboxylase (PYC) gene expression that exceeded our thresholds (Table 2), however the direction of change was not consistent within a species or among species within the same phyla. These results provide little support for a coherent change in C4 CCM in response to elevated CO2 in these phytoplankton species. Within the photorespiration pathway, at least one copy of the gene encoding the enzyme that catalyzes the first step of photorespiration, 2-phosphoglycolate phosphatase (PGP), had increased expression in all four species with form 1D Rubisco (Table 2). This is counter to expectation, as rising CO2 should decrease the probability of oxygen being fixed in place of carbon and therefore the need for expressing photorespiration pathways. Other photorespiration genes did have decreased expression including alanine amino transferase (ALAT_GGAT) in the mitochondrial pathway and isocitrate lyase (ICL) in the peroxisomal pathway (Table 2). These gene expression patterns reveal a mixed metabolic pathway response to CO2 even within a single species that does not fit the simple picture of how phytoplankton are expected to respond to elevated CO2.
Several putative CCM genes displayed a shared response across species but are poorly characterized (Table 2). For example, a family of gated ion channels (bestrophins), closely associated with the expression of other CCM genes (Hennon et al., 2015), met our threshold for differential expression in all six species examined in this study in response to CO2 (Table 2). Another gene that was widely shared and regulated in response to CO2 was the oxoglutarate/malate transporter (OMT) (Table 2). This gene is also poorly characterized, yet is thought to be essential for C4 metabolic processes, as all eukaryotic phytoplankton have many organelle compartments between the site of C4 fixation and decarboxylation (Kroth et al., 2008). These examples highlight the need for further research on the transporters and ion channels that may serve vital functions in shuttling metabolites between intracellular compartments. These results also suggest that these genes may be more highly-conserved between taxa than other CCM genes.
Section 3: Survey of CCM and Photorespiration Gene Complement of Four Eukaryotic Phytoplankton Phyla
To assess whether the changes in CCM and photorespiration genes within the six phytoplankton species examined were representative of other phytoplankton within the same phyla, we surveyed genes in these pathways across the consensus assemblies of related species in the MMETSP data set and available genomes (Figure 6, Supplementary Tables 3, 4). A total of 77 species belonging to four phyla were examined (Figure 6). At least one complete photorespiration pathway was expressed in all 77 species (Supplementary Table 3), emphasizing the importance of this pathway for phytoplankton with both forms of Rubisco. Genes for biophysical CCMs were expressed in all species, although the gene type and copy number varied between and within each group (Figure 6). The better-studied CCM gene families like carbonic anhydrases (CA) varied in the type expressed and copy number between phyla (Figure 6). Beta-CAs were prevalent in raphidophytes and dinoflagellates, while delta-CAs were most prevalent in diatoms and haptophytes (Figure 6). Even within diatoms, there was large variation in expression of CA types, with many lacking expression of a beta-CA and expressing only alpha and delta (Supplementary Table 3). No CA family was universally expressed by all groups, suggesting high diversity in the genes used for biophysical CCM, and supporting convergent evolution of some biophysical CCM genes.
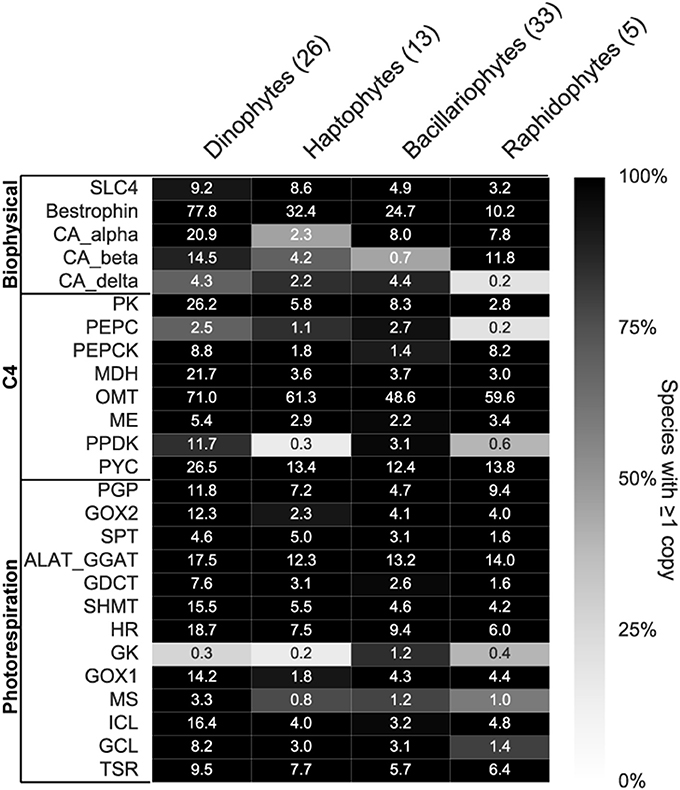
Figure 6. Carbon concentrating mechanism (CCM) and photorespiration expressed gene complement in four phytoplankton phyla. Custom HMM searches for Biophysical (BP) and C4 CCM (C4) and photorespiration (PR) genes within consensus contigs or gene models of each phyla (Supplementary Tables 3, 4). Numbers in parentheses next to phyla name indicates total number of species within each group. Shading indicates percentage of species with at least one copy within each phyla and the numbers indicate mean copy number of each gene. Gene abbreviations are as follows; SLC4, SLC4 bicarbonate transporter; CA, carbonic anhydrase; PK, pyruvate kinase; PEPC, phosphoenolpyruvate carboxylase; PEPCK, phosphoenolpyruvate carboxykinase; MDH, malate dehydrogenase; OMT, oxoglutarate/malate transporter; ME, malic enzyme; PPDK, pyruvate orthophosphate dikinase; PYC, pyruvate carboxylase; PGP, phosphoglycolate phosphatase; GOX, glycolate oxidase; SPT, serine pyruvate aminotransferase; ALAT-GGAT, alanine aminotransferase; GDCT, glycine decarboxylase T-protein; SHMT, serine hydroxymethyl transferase; HR, hydroxypyruvate reductase; GK, glycerate kinase; MS, malate synthase; ICL, isocitrate lyase; GCL, glycolate carboligase; TSR, tartronate semialdehyde reductase.
Seventy-five out of 77 species expressed a SLC4 bicarbonate transporter gene. The SLC4 transporter has been shown in a diatom to transport bicarbonate into the cell (Nakajima et al., 2013) and is thought to be an integral part of the phytoplankton biophysical CCM. A putative ion channel (bestrophin), identified as a potentially conserved biophysical CCM gene (Hennon et al., 2015, this study), was present in high copy numbers in all 77 species examined (Figure 6). The bestrophin gene family is much less well-studied than SLC4 bicarbonate transporters, yet may serve vital functions for the biophysical CCM (Hennon et al., 2015). Bestrophin genes were likely missed by our orthologous group analysis (Figure 4) because of the abundant paralogous gene copies of bestrophins within each organism (Figure 6, Supplementary Table 3) making it difficult to classify into a single orthologous group. Because bestrophin genes have only been studied in the context of human eye diseases (Yang et al., 2014), their role in phytoplankton CCM remains uncertain.
Putative C4 pathway genes also varied in gene copy and type (Figure 6), with 48 out of 77 species expressing a full complement of C4 genes (Kroth et al., 2008). Similar to the bestrophin gene, the putative OMT transporter for C4 CCMs was universally conserved across the dataset and expressed in high copy numbers (~60 per species, Figure 6). The gene typically considered rate-limiting for the C4 pathway, pyruvate orthophosphate dikinase (PPDK), was not detected in 85% of haptophytes and 60% of raphidophytes (Supplementary Table 3). These patterns indicate that C4 may not be a widespread CCM strategy in haptophytes and raphidophytes, or perhaps that it is expressed at such low levels that it cannot be resolved in the available transcriptomes. The results from the transcriptome survey warrant caution in attributing changes in C4 genes to a change in CCM, particularly given that the localization of these genes is unknown, yet crucial to the functioning of a C4 CCM (Kroth et al., 2008).
Conclusions
With the recent increase in availability of whole transcriptomes for eukaryotic phytoplankton species (Keeling et al., 2014), it is now possible to ask whether expressed gene complements can inform us about the potential for physiological responses to rising CO2. This study focused on characterizing the physiological and gene expression changes of six ecologically important phytoplankton species to elevated CO2 in the context of the expressed gene complements from the four phyla they represent. Although some similarities were observed in growth rate and C:N ratio related to phylogeny, changes in gene expression, even those that underpin CCM activity and photorespiration, were largely unique to each species. Many phytoplankton express CCM and photorespiration genes, but the lack of consistent CO2 response in the four phyla studied here suggests that responses to CO2 perturbations are more diverse than their physiological responses would indicate. As such, extrapolation of detailed genetic mechanisms from a few well-studied phytoplankton to other species, even within the same phylum, may be problematic with regards to CCM and photorespiration pathways. This study demonstrates the value of broad taxonomic comparisons in gene expression studies. Several broadly-defined gene groups or families were highlighted that responded similarly to changing CO2 across diverse taxa. Though the functions of those highlighted gene groups, such as bestrophins and oxoglutarate/malate transporters, are poorly understood, identifying such broadly-conserved responses may be required to gauge the response of natural phytoplankton communities to CO2 perturbations. The cross-species comparison also revealed divergence in gene complement and regulation between species, providing insight into the physiological and genetic flexibility with respect to changing CO2 inherent within phytoplankton communities. Understanding the diversity in genetic complement and gene expression response to elevated CO2 can improve predictions of the relative success of phytoplankton taxa in future ecosystems, their impacts on biogeochemical cycles, and harmful algal bloom frequency.
Author Contributions
AJ and SD designed the experiments, AJ and SH performed the experiments, GH and MH analyzed the data, GH wrote the manuscript with input from all authors.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This research was made possible by funding from the National Science Foundation Biological Oceanography Program #OCE13-14336 (SD) and the Gordon and Betty Moore Foundation Marine Microbial Eukaryote Transcriptome Sequencing Project, with partial support also provided by the Paul M. Angell Family Foundation and WSL PURE in partnership with Columbia University's Center for Climate and Life. This work is LDEO contribution 8171 and was also made possible by resources provided by the JP Sulzberger Columbia Genome Center and the Mason high performance computing cluster at the University of Indiana (NSF Extreme Science and Engineering Discovery Environment project). We would also like to thank Alexandra Bausch for productive discussions, Dr. Craig Aumack for conducting the carbohydrate analyses, and Senjie Lin and two reviewers for valuable feedback.
Supplementary Material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2017.02547/full#supplementary-material
References
Anderson, D. M., Alpermann, T. J., Cembella, A. D., Collos, Y., Masseret, E., and Montresor, M. (2012). The globally distributed genus Alexandrium: multifaceted roles in marine ecosystems and impacts on human health. Harmful Algae 14, 10–35. doi: 10.1016/j.hal.2011.10.012
Badger, M. R., Andrews, T. J., Whitney, S. M., Ludwig, M., Yellowlees, D. C., Leggat, W., et al. (1998). The diversity and coevolution of Rubisco, plastids, pyrenoids, and chloroplast-based CO2-concentrating mechanisms in algae. Can. J. Bot. 76, 1052–1071. doi: 10.1139/b98-074
Bagby, S. C., and Chisholm, S. W. (2015). Response of Prochlorococcus to varying CO2:O2 ratios. ISME J. 9, 2232–2245. doi: 10.1038/ismej.2015.36
Barshis, D. J., Ladner, J. T., Oliver, T. A., and Palumbi, S. R. (2014). Lineage-specific transcriptional profiles of Symbiodinium spp. Unaltered by heat stress in a coral host. Mol. Biol. Evol. 31, 1343–1352. doi: 10.1093/molbev/msu107
Bauwe, H., Hagemann, M., and Fernie, A. R. (2010). Photorespiration: players, partners and origin. Trends Plant Sci. 15, 330–336. doi: 10.1016/j.tplants.2010.03.006
Berge, T., Daugbjerg, N., Andersen, B. B., and Hansen, P. J. (2010). Effect of lowered pH on marine phytoplankton growth rates. Mar. Ecol. Prog. Ser. 416, 79–91. doi: 10.3354/meps08780
Branson, J. P., Nezic, M., Jitrapakdee, S., Wallace, J. C., and Attwood, P. V. (2004). Kinetic characterization of yeast pyruvate carboxylase isozyme Pyc1 and the Pyc1 mutant, C249A. Biochemistry 43, 1075–1081. doi: 10.1021/bi035575y
Cai, W. J., Hu, X., Huang, W. J., Jiang, L. Q., Wang, Y., Peng, T. H., et al. (2010). Alkalinity distribution in the western North Atlantic Ocean margins. J. Geophys. Res. Ocean. 115, 1–15. doi: 10.1029/2009JC005482
Chi, S., Wu, S., Yu, J., Wang, X., Tang, X., and Liu, T. (2014). Phylogeny of C4-photosynthesis enzymes based on algal transcriptomic and genomic data supports an archaeal/proteobacterial origin and multiple duplication for most C4-related genes. PLoS ONE 9:e110154. doi: 10.1371/journal.pone.0110154
Ciais, P., Sabine, C., Bala, G., Bopp, L., Brovkin, V., Canadell, J., et al. (2013). “Carbon and other biogeochemical cycles,” in Climate Change 2013: The Physical Science Basis (Cambridge).
Dahl, E., Lindahl, O., Paasche, E., and Throndsen, J. (1989). “The Chrysochromulina polylepis bloom in Scandinavian waters during spring 1988,” in Novel Phytoplankton Blooms: Causes and Impacts of Recurrent Brown Tides and Other Unusual Blooms, eds E. M. Cosper, V. M. Bricelj, and E. J. Carpenter (Berlin; Heidelberg: Springer), 383–405.
Doney, S. C., Fabry, V. J., Feely, R. A., and Kleypas, J. A. (2009). Ocean acidification: the other CO2 problem. Annu. Rev. Mar. Sci. 1, 169–192. doi: 10.1146/annurev.marine.010908.163834
DuBois, M., Gilles, K. A., Hamilton, J. K., Rebers, P. A., and Smith, F. (1956). Colorimetric method for determination of sugars and related substances. Anal. Chem. 28, 350–356. doi: 10.1021/ac60111a017
Ducklow, H. W., Steinberg, D. K., and Buesseler, K. O. (2001). Upper ocean carbon export and the biological pump. Oceanography 14, 50–58. doi: 10.5670/oceanog.2001.06
Dutkiewicz, S., Morris, J. J., Follows, M. J., Scott, J., Levitan, O., Dyhrman, S. T., et al. (2015). Impact of ocean acidification on the structure of future phytoplankton communities. Nat. Clim. Chang. 5, 1002–1006. doi: 10.1038/nclimate2722
Eberlein, T., Van de Waal, D. B., Brandenburg, K. M., John, U., Voss, M., Achterberg, E. P., et al. (2016). Interactive effects of ocean acidification and nitrogen limitation on two bloom-forming dinoflagellate species. Mar. Ecol. Prog. Ser. 543, 127–140. doi: 10.3354/meps11568
Eddy, S. R. (2011). Accelerated profile HMM searches. PLoS Comput. Biol. 7:e1002195. doi: 10.1371/journal.pcbi.1002195
Elser, J. J., Fagan, W. F., Denno, R. F., Dobberfuhl, D. R., Folarin, A., Huberty, A., et al. (2000). Nutritional constraints in terrestrial and freshwater food webs. Nature 408, 578–580. doi: 10.1038/35046058
Emms, D. M., and Kelly, S. (2015). OrthoFinder: solving fundamental biases in whole genome comparisons dramatically improves orthogroup inference accuracy. Genome Biol. 16:157. doi: 10.1186/s13059-015-0721-2
Feely, R. A., Sabine, C. L., Hernandez-Ayon, J. M., Ianson, D., and Hales, B. (2008). Evidence for upwelling of corrosive “acidified” water onto the continental shelf. Science 320, 1490–1492. doi: 10.1126/science.1155676
Feely, R. A., Sabine, C. L., Lee, K., Berelson, W., Kleypas, J., Fabry, V. J., et al. (2004). Impact of anthropogenic CO2 on the CaCO3 system in the oceans. Science 305, 362–366. doi: 10.1126/science.1097329
Hackenberg, C., Kern, R., Hüge, J., Stal, L. J., Tsuji, Y., Kopka, J., et al. (2011). Cyanobacterial lactate oxidases serve as essential partners in N2 fixation and evolved into photorespiratory glycolate oxidases in plants. Plant Cell 23, 2978–2990. doi: 10.1105/tpc.111.088070
Hallegraeff, G. M. (2010). Ocean climate change, phytoplankton community responses, and harmful algal blooms: a formidable predictive challenge. J. Phycol. 46, 220–235. doi: 10.1111/j.1529-8817.2010.00815.x
Harke, M. J., Juhl, A. R., Haley, S. T., Alexander, H., and Dyhrman, S. T. (2017). Conserved transcriptional responses to nutrient stress in bloom-forming algae. Front. Microbiol. 8:1279. doi: 10.3389/fmicb.2017.01279
Heil, C. A., Glibert, P. M., and Fan, C. (2005). Prorocentrum minimum (Pavillard) Schiller: a review of a harmful algal bloom species of growing worldwide importance. Harmful Algae 4, 449–470. doi: 10.1016/j.hal.2004.08.003
Hennon, G. M. M., Ashworth, J., Groussman, R. D., Berthiaume, C., Morales, R. L., Baliga, N. S., et al. (2015). Diatom acclimation to elevated CO2 via cAMP signalling and coordinated gene expression. Nat. Clim. Chang. 5, 761–765. doi: 10.1038/nclimate2683
Hennon, G. M. M., Quay, P., Morales, R. L., Swanson, L. M., and Armbrust, E. V. (2014). Acclimation conditions modify physiological response of the diatom Thalassiosira pseudonana to elevated CO2 concentrations in a nitrate-limited chemostat. J. Phycol. 253, 243–253. doi: 10.1111/jpy.12156
Honjo, T. (1992). “Harmful red tides of Heterosigma akashiwo,” in Control of Disease in Aquaculture, ed R. S. Svrjecek (Springfield, VA: NOAA Technical Report NMFS 111), 27–32.
Hopkinson, B. M., Dupont, C. L., Allen, A. E., and Morel, F. M. M. (2011). Efficiency of the CO2-concentrating mechanism of diatoms. Proc. Natl. Acad. Sci. U.S.A. 108, 3830–3837. doi: 10.1073/pnas.1018062108
Huang, X., and Madan, A. (1999). CAP3: a DNA sequence assembly program. Genome Res. 9, 868–877. doi: 10.1101/gr.9.9.868
Jitrapakdee, S., St Maurice, M., Rayment, I., Cleland, W. W., Wallace, J. C., and Attwood, P. V. (2008). Structure, mechanism and regulation of pyruvate carboxylase. Biochem. J. 413, 369–387. doi: 10.1042/BJ20080709
Juhl, A. R. (2005). Growth rates and elemental composition of Alexandrium monilatum, a red-tide dinoflagellate. Harmful Algae 4, 287–295. doi: 10.1016/j.hal.2004.05.003
Juhl, A. R., and Latz, M. I. (2002). Mechanisms of fluid shear-induced inhibition of population growth in a red-tide dinoflagellate. J. Phycol. 38, 683–694. doi: 10.1046/j.1529-8817.2002.00165.x
Kanehisa, M., Sato, Y., and Morishima, K. (2016). BlastKOALA and GhostKOALA: KEGG tools for functional characterization of genome and metagenome sequences. J. Mol. Biol. 428, 726–731. doi: 10.1016/j.jmb.2015.11.006
Katoh, K., and Standley, D. M. (2013). MAFFT multiple sequence alignment software version 7: improvements in performance and usability. Mol. Biol. Evol. 30, 772–780. doi: 10.1093/molbev/mst010
Keeling, P. J., Burki, F., Wilcox, H. M., Allam, B., Allen, E. E., Amaral-Zettler, L. A., et al. (2014). The Marine Microbial Eukaryote Transcriptome Sequencing Project (MMETSP): illuminating the functional diversity of eukaryotic life in the oceans through transcriptome sequencing. PLoS Biol. 12:e1001889. doi: 10.1371/journal.pbio.1001889
Kroth, P. G., Chiovitti, A., Gruber, A., Martin-Jezequel, V., Mock, T., Parker, M. S., et al. (2008). A model for carbohydrate metabolism in the diatom Phaeodactylum tricornutum deduced from comparative whole genome analysis. PLoS ONE 3:e1426. doi: 10.1371/journal.pone.0001426
Kustka, A. B., Milligan, A. J., Zheng, H., New, A. M., Gates, C., Bidle, K. D., et al. (2014). Low CO2 results in a rearrangement of carbon metabolism to support C4 photosynthetic carbon assimilation in Thalassiosira pseudonana. New Phytol. 204, 507–520. doi: 10.1111/nph.12926
Le Quéré, C., Andres, R. J., Boden, T., Conway, T., Houghton, R. A., House, J. I., et al. (2012). The global carbon budget 1959–2011. Earth Syst. Sci. Data Discuss. 5, 1107–1157. doi: 10.5194/essdd-5-1107-2012
Lewis, E., and Wallace, D. (1998). Program Developed for CO2 System Calculations. Oak Ridge, TN. Available online at: http://cdiac.ornl.gov/oceans/co2rprt.html. doi: 10.2172/639712
Li, H., and Durbin, R. (2009). Fast and accurate short read alignment with Burrows-Wheeler transform. Bioinformatics 25, 1754–1760. doi: 10.1093/bioinformatics/btp324
Li, R., Li, Y., Kristiansen, K., and Wang, J. (2008). SOAP: Short oligonucleotide alignment program. Bioinformatics 24, 713–714. doi: 10.1093/bioinformatics/btn025
Li, W., and Godzik, A. (2006). CD-HIT: a fast program for clustering and comparing large sets of protein or nucleotide sequences. Bioinformatics 22, 1658–1659. doi: 10.1093/bioinformatics/btl158
Lin, S. (2011). Genomic understanding of dinoflagellates. Res. Microbiol. 162, 551–569. doi: 10.1016/j.resmic.2011.04.006
Lohbeck, K. T., Riebesell, U., and Reusch, T. B. H. (2014). Gene expression changes in the coccolithophore Emiliania huxleyi after 500 generations of selection to ocean acidification. Proc. R. Soc. 281:20140003. doi: 10.1098/rspb.2014.0003
McGinn, P. J., and Morel, F. M. M. (2007). Expression and inhibition of the carboxylating and decarboxylating enzymes in the photosynthetic C4 pathway of marine diatoms. Plant Physiol. 146, 300–309. doi: 10.1104/pp.107.110569
Milenkovic, V. M., Langmann, T., Schreiber, R., Kunzelmann, K., and Weber, B. H. F. (2008). Molecular evolution and functional divergence of the bestrophin protein family. BMC Evol. Biol. 8:72. doi: 10.1186/1471-2148-8-72
Milliman, J. D. (1993). Production and accumulation of calcium carbonate in the ocean: budget of a nonsteady state. Global Biogeochem. Cycles 7, 927–957. doi: 10.1029/93GB02524
Mock, T., Samanta, M. P., Iverson, V., Berthiaume, C., Robison, M., Holtermann, K., et al. (2008). Whole-genome expression profiling of the marine diatom Thalassiosira pseudonana identifies genes involved in silicon bioprocesses. Proc. Natl. Acad. Sci. U.S.A. 105, 1579–1584. doi: 10.1073/pnas.0707946105
Morse, D., Salois, P., Markovic, P., and Hastings, J. W. (1995). A nuclear-encoded form II RuBisCO in dinoflagellates. Science 268, 1622-1624.
Murray, S. A., Suggett, D. J., Doblin, M. A., Kohli, G. S., Seymour, J. R., Fabris, M., et al. (2016). Unravelling the functional genetics of dinoflagellates: a review of approaches and opportunities. Perspect. Phycol. 3, 37–52. doi: 10.1127/pip/2016/0039
Nakajima, K., Tanaka, A., and Matsuda, Y. (2013). SLC4 family transporters in a marine diatom directly pump bicarbonate from seawater. Proc. Natl. Acad. Sci. U.S.A. 110, 1767–1772. doi: 10.1073/pnas.1216234110
Nelson, D. M., Treguer, P., Brzezinski, M. A., Leynaert, A., and Queguiner, B. (1995). Production and dissolution of biogenic silica in the ocean: revised global estimates, comparison with regional data and relationship to biogenic sedimentation. Glob. Biogeochem. Cycle 9, 359–372. doi: 10.1029/95GB01070
Parker, M. S., and Armbrust, E. V. (2005). Synergistic effects of light, temperature, and nitrogen source on transcription of genes for carbon and nitrogen metabolism in the centric diatom Thalassiosira pseudonana (Bacillariophyceae). J. Phycol. 41, 1142–1153. doi: 10.1111/j.1529-8817.2005.00139.x
Reinfelder, J. R., Kraepiel, A. M., and Morel, F. M. (2000). Unicellular C4 photosynthesis in a marine diatom. Nature 407, 996–999. doi: 10.1038/35039612
Rickaby, R. E. M., Henderiks, J., and Young, J. N. (2010). Perturbing phytoplankton: response and isotopic fractionation with changing carbonate chemistry in two coccolithophore species. Clim. Past 6, 771–785. doi: 10.5194/cp-6-771-2010
Riebesell, U., Schulz, K. G., Bellerby, R. G. J., Botros, M., Fritsche, P., Meyerhöfer, M., et al. (2007). Enhanced biological carbon consumption in a high CO2 ocean. Nature 450, 545–548. doi: 10.1038/nature06267
Roberts, K., Granum, E., Leegood, R. C., and Raven, J. A. (2007). C3 and C4 pathways of photosynthetic carbon assimilation in marine diatoms are under genetic, not environmental, control. Plant Physiol. 145, 230–235. doi: 10.1104/pp.107.102616
Rokitta, S. D., John, U., and Rost, B. (2012). Ocean acidification affects redox-balance and ion-homeostasis in the life-cycle stages of Emiliania huxleyi. PLoS ONE 7:e52212. doi: 10.1371/journal.pone.0052212
Romero, M. F., Chen, A.-P., Parker, M. D., and Boron, W. F. (2013). The SLC4 family of bicarbonate (HCO3−) transporters. Mol. Aspects Med. 34, 159–182. doi: 10.1016/j.mam.2012.10.008
Rost, B., Riebesell, U., Burkhardt, S., and Sültemeyer, D. (2003). Carbon acquisition of bloom-forming marine phytoplankton. Limnol. Oceanogr. 48, 55–67. doi: 10.4319/lo.2003.48.1.0055
Ryther, J. H. (1969). Photosynthesis and fish production in the sea. Science 166, 72–76. doi: 10.1126/science.166.3901.72
Sabine, C. L., Feely, R. A., Gruber, N., Key, R. M., Lee, K., Bullister, J. L., et al. (2004). The oceanic sink for anthropogenic CO2. Science 305, 367–371. doi: 10.1126/science.1097403
Shrestha, R. P., Tesson, B., Norden-Krichmar, T., Federowicz, S., Hildebrand, M., and Allen, A. E. (2012). Whole transcriptome analysis of the silicon response of the diatom Thalassiosira pseudonana. BMC Genomics 13:499. doi: 10.1186/1471-2164-13-499
Simpson, J. T., and Durbin, R. (2012). Efficient de novo assembly of large genomes using compressed data structures. Genome Res. 22, 549–556. doi: 10.1101/gr.126953.111
Simpson, J. T., Wong, K., Jackman, S. D., Schein, J. E., Jones, S. J. M., and Birol, I. (2009). ABySS: a parallel assembler for short read sequence data. Genome Res. 19, 1117–1123. doi: 10.1101/gr.089532.108
Strickland, J. D. H., and Parsons, T. R. (1972). A Practical Handbook of Seawater Analyses, 2nd Edn. Ottawa, ON: Fisheries Research Board of Canada. p. 310.
Tabita, F. R., Satagopan, S., Hanson, T. E., Kreel, N. E., and Scott, S. S. (2008). Distinct form I, II, III, and IV Rubisco proteins from the three kingdoms of life provide clues about Rubisco evolution and structure/function relationships. J. Exp. Bot. 59, 1515–1524. doi: 10.1093/jxb/erm361
Tachibana, M., Allen, A. E., Kikutani, S., Endo, Y., Bowler, C., and Matsuda, Y. (2011). Localization of putative carbonic anhydrases in two marine diatoms, Phaeodactylum tricornutum and Thalassiosira pseudonana. Photosynth. Res. 109, 205–221. doi: 10.1007/s11120-011-9634-4
Takahashi, T., Sutherland, S. C., Chipman, D. W., Goddard, J. G., and Ho, C. (2014). Climatological distributions of pH, pCO2, total CO2, alkalinity, and CaCO3 saturation in the global surface ocean, and temporal changes at selected locations. Mar. Chem. 164, 95–125. doi: 10.1016/j.marchem.2014.06.004
Thomas, W. H., and Gibson, C. H. (1990). Effects of small-scale turbulence on microalgae. J. Appl. Phycol. 2, 71–77. doi: 10.1007/BF02179771
Tortell, P. D. (2000). Evolutionary and ecological perspectives on carbon acquisition in phytoplankton. Limnol. Ocean. 45, 744–750. doi: 10.4319/lo.2000.45.3.0744
Trainer, V. L., and Yoshida, T. (eds.) (2014). Proceedings of the Workshop on Economic Impacts of Harmful Algal Blooms on Fisheries and Aquaculture. PICES Sci. Rep. No. 47, p. 85.
Tsuji, Y., Suzuki, I., and Shiraiwa, Y. (2012). Enzymological evidence for the function of a plastid-located pyruvate carboxylase in the haptophyte alga Emiliania huxleyi: a novel pathway for the production of C 4 compounds. Plant Cell Physiol. 53, 1043–1052. doi: 10.1093/pcp/pcs045
Van de Waal, D. B., Eberlein, T., John, U., Wohlrab, S., and Rost, B. (2014). Impact of elevated pCO2 on paralytic shellfish poisoning toxin content and composition in Alexandrium tamarense. Toxicon 78, 58–67. doi: 10.1016/j.toxicon.2013.11.011
Van de Waal, D. B., John, U., Ziveri, P., Reichart, G. J., Hoins, M., Sluijs, A., et al. (2013). Ocean acidification reduces growth and calcification in a marine dinoflagellate. PLoS ONE 8:e65987. doi: 10.1371/journal.pone.0065987
Waterhouse, A. M., Procter, J. B., Martin, D. M. A., Clamp, M., and Barton, G. J. (2009). Jalview Version 2-A multiple sequence alignment editor and analysis workbench. Bioinformatics 25, 1189–1191. doi: 10.1093/bioinformatics/btp033
Wu, Y., Campbell, D. A., Irwin, A. J., Suggett, D. J., and Finkel, Z. V. (2014). Ocean acidification enhances the growth rate of larger diatoms. Limnol. Oceanogr. 59, 1027–1034. doi: 10.4319/lo.2014.59.3.1027
Yang, T., Liu, Q., Kloss, B., Bruni, R., Kalathur, R. C., Guo, Y., et al. (2014). Structure and selectivity in bestrophin ion channels. Science 346, 355–359. doi: 10.1126/science.1259723
Keywords: algae, carbon concentrating mechanism (CCM), C4 CCM, biophysical CCM, photorespiration, transcriptomics
Citation: Hennon GMM, Hernández Limón MD, Haley ST, Juhl AR and Dyhrman ST (2017) Diverse CO2-Induced Responses in Physiology and Gene Expression among Eukaryotic Phytoplankton. Front. Microbiol. 8:2547. doi: 10.3389/fmicb.2017.02547
Received: 31 August 2017; Accepted: 08 December 2017;
Published: 19 December 2017.
Edited by:
Senjie Lin, University of Connecticut, United StatesReviewed by:
Dedmer B. Van de Waal, Netherlands Institute of Ecology (NIOO-KNAW), NetherlandsKathleen Scott, University of South Florida, United States
Copyright © 2017 Hennon, Hernández Limón, Haley, Juhl and Dyhrman. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sonya T. Dyhrman, c2R5aHJtYW5AbGRlby5jb2x1bWJpYS5lZHU=