 Kristin Schulz-Bohm
Kristin Schulz-Bohm Lara Martín-Sánchez
Lara Martín-Sánchez Paolina Garbeva
Paolina Garbeva- Department of Microbial Ecology, Netherlands Institute of Ecology (NIOO-KNAW), Wageningen, Netherlands
During the last decades, research on the function of volatile organic compounds focused primarily on the interactions between plants and insects. However, microorganisms can also release a plethora of volatiles and it appears that microbial volatile organic compounds (mVOCs) can play an important role in intra- and inter-kingdom interactions. So far, most studies are focused on aboveground volatile-mediated interactions and much less information is available about the function of volatiles belowground. This minireview summarizes the current knowledge on the biological functions of mVOCs with the focus on mVOCs-mediated interactions belowground. We pinpointed mVOCs involved in microbe-microbe and microbe–plant interactions, and highlighted the ecological importance of microbial terpenes as a largely underexplored group of mVOCs. We indicated challenges in studying belowground mVOCs-mediated interactions and opportunities for further studies and practical applications.
Introduction
Many secondary metabolites have been reported to be involved in microbial interactions. One group of secondary metabolites produced by soil and plant-associated microorganisms, but largely unexplored to date, are the volatile organic compounds (VOCs). VOCs are typically small, odorous compounds (<C15) with low molecular mass (<300 Da), high vapor pressure, low boiling point, and a lipophilic moiety. These properties facilitate evaporation and diffusion aboveground and belowground through gas- and water- filled pores in soil and rhizosphere environments (Vespermann et al., 2007; Insam and Seewald, 2010; Effmert et al., 2012). Microbial volatile organic compounds (mVOCs) belong to different chemical classes including alkenes, alcohols, ketones, benzenoids, pyrazines, sulfides, and terpenes (Schulz and Dickschat, 2007; Lemfack et al., 2014, 2017; Kanchiswamy et al., 2015; Schmidt et al., 2015). A recent meta-analysis by Schenkel et al. (2015) provided a comprehensive overview of VOCs derived from soil-borne microbes.
The production of mVOCs in soil is influenced by various factors including the growth stage of the microbes, nutrient availability, temperature, oxygen availability, pH, and soil moisture content (Wheatley, 2002; Insam and Seewald, 2010). Several recent studies reported that the production of certain mVOCs can be induced or suppressed during inter-specific microbial interactions (Garbeva et al., 2014a; Schulz-Bohm et al., 2015; Tyc et al., 2015; Piechulla et al., 2017). mVOCs were often considered to be by-products of primary metabolism, but recent findings revealed that many mVOCs demonstrate biological activity (Schmidt et al., 2015; Tyc et al., 2017a). Furthermore, in bacteria, the production of certain mVOCs is dependent on the GacS/GacA two-component regulatory system (Cheng et al., 2016; Ossowicki et al., 2017). These findings clearly disagree with the opinion that mVOCs are just waste products.
While soluble metabolites are often responsible for short distance interactions, VOCs are considered to be long-distance messengers (Tyc et al., 2017b; Westhoff et al., 2017). There are many types of microbial interactions occurring belowground such as bacteria–bacteria, fungi–fungi, fungi–bacteria, bacteria–protists, fungi–plant, bacteria–plant, and bacteria–fungi–plant interactions. However, most studies addressing belowground VOCs-mediated interactions are focused mainly on the root-emitted volatiles (recently reviewed by Delory et al., 2016).
The knowledge we have gained from research conducted over the last few years reveals that mVOCs can have both beneficial and harmful effects on other organisms (Effmert et al., 2012; Schmidt et al., 2015). mVOCs can provide organisms with rapid and precise ways to recognize neighboring organisms (both friends and foe) and to launch proper responses.
The aim of this review is to summarize the current knowledge concerning the role of mVOCs in intra- and inter-kingdom interactions, to pinpoint mVOCs (e.g., terpenes) involved in microbe–microbe and microbe–plant interactions as well as to indicate challenges in studying belowground mVOCs-mediated interactions and opportunities for further studies and practical applications.
VOCs in Microbe–Microbe Interaction
Bacteria–Bacteria
Bacterial VOCs can have direct antagonistic effects against other bacteria. For instance, the sesquiterpene albaflavenone produced by Streptomyces albidoflavus revealed activity against Bacillus subtilis (Gürtler et al., 1994) and the emission of dimethyl disulphide by two rhizospheric bacteria, Pseudomonas fluorescens and Serratia plymuthica, showed bacteriostatic effects against two plant bacterial pathogens Agrobacterium tumefaciens and Agrobacterium vitis (Dandurishvili et al., 2011). Pseudomonas fluorescens WR-1 produces volatiles such as benzothiazole and 1-methyl naphthalene with bacteriostatic effects against the tomato pathogen Ralstonia solanacearum (Raza et al., 2016a). In fact, many species of Pseudomonas and Bacillus that are used as biocontrol agents against plant pathogens, have been reported to produce VOCs with antibacterial activity (Raza et al., 2016a,b,c; Xie et al., 2016; Rajer et al., 2017; Tahir et al., 2017a,b). For instance, a recent study revealed that VOCs produced by Bacillus spp., including benzaldehyde, 1,2-benzisothiazol-3(2 H)-one and 1,3-butadiene, had strong inhibitory activity against R. solanacearum, the causal agent of bacterial wilt disease (Tahir et al., 2017a). The mVOCs altered the transcriptional expression levels of several genes involved in motility and pathogenicity (e.g., global virulence regulator PhcA, type III secretion system, and extracellular polysaccharide [EPS] production) and induced systemic resistance by plants, which resulted in a decrease of wilt disease.
Several reports describe the effect of VOCs in bacterial virulence. For instance, 2,3 butanediol and acetoin are required for full virulence in Pectobacterium carotovorum. The same compounds can increase the production of virulence factors in Pseudomonas aeruginosa (Audrain et al., 2015).
In contrast, VOCs produced by some bacteria can also have positive effects on the growth of other neighboring bacteria in the rhizosphere. For instance, VOCs from Collimonas pratensis and S. plymuthica are able to induce the growth of P. fluorescens Pf0-1 (Garbeva et al., 2014a). These VOCs induced expression of genes involved in motility in P. fluorescens Pf0-1 and provoked an increase in the production of secondary metabolites with antibacterial activity against Bacillus (Garbeva et al., 2014a). This suggests that C. pratensis and S. plymuthica may be attracting and promoting the growth of P. fluorescens in a collaborative attempt to increase their chances against different bacterial competitors or soil fungal pathogens. Another example of the growth-promoting effect of VOCs was reported recently by Schulz-Bohm et al. (2015) which showed that VOCs released by mixtures of root exudate-consuming bacteria stimulated the activity and growth of distant nutrient-limited bacteria.
In addition to exerting antagonistic effects toward other bacteria, VOCs can also modify the behavior of other bacteria and modulate their resistance to antibiotics. Bacterial volatiles such as ammonia, trimethylamine, hydrogen sulfide, nitric oxide, and 2-amino-acetophenone can alter biofilm formation or dispersal or affect motility of bacteria (Audrain et al., 2015; Raza et al., 2016a). Bacteria often make use of their motility to move to other areas with more resources and/or less competitors. In Streptomyces venezuelae, a new mode of development, so-called exploration, has been recently discovered that allows non-motile bacteria to access regions with more nutrients (Jones et al., 2017). S. venezuelae is able to produce hydrophilic fast growing non-branching vegetative hyphae, triggered by glucose depletion and a rise in pH, to presumably escape from poor nutrient areas. Interestingly, explorer cells can release signals for long distance communication with other members of the species to induce their exploratory growth. One of these signals is trimethylamine, which works not only as a signal to communicate with distantly located Streptomyces and induce exploratory growth but also displays antibacterial activity against B. subtilis and Micrococcus luteus, probably by rising the pH of the medium (Jones et al., 2017).
Fungi–Bacteria
Fungal VOCs can play an important role in long distance fungal–bacterial interactions and can lead to different phenotypical responses in the interacting partners. For example, VOCs emitted by Trichoderma atroviride increased the expression of a biocontrol gene (phlA) in P. fluorescens that encodes the biosynthesis of 2,4-diacetylphloroglucinol (Lutz et al., 2004). A few recent studies demonstrated that the growth of some bacterial species can be suppressed by fungal VOCs (Werner et al., 2016) such as the VOCs that exhibit inhibitory effects on B. cereus and B. subtilis produced by the oyster mushroom Pleurotus ostreatus (Pauliuc and Botǎu, 2013).
Recently, Schmidt et al. (2015) screened the phenotypic responses of soil bacterial strains to volatiles emitted by several fungal and oomycetal soil strains under different nutrient conditions during different growth stages. Out of the phenotypical responses tested such as growth alteration, antimicrobial activity, biofilm formation or motility, motility of bacteria (both swimming and swarming) was significantly positively or negatively affected by fungal and oomycetal VOCs. This finding could, therefore, reflect a potential strategy employed by the fungus to attract mutualistic bacteria toward itself and to repel competitors by manipulating their motility through the use of VOCs (Piechulla et al., 2017). Transcriptomics and proteomics analyses of S. plymuthica PRI-2C exposed to VOCs emitted by the fungal pathogen Fusarium culmorum, showed that S. plymuthica PRI-2C responded to the fungal VOCs with changes in gene and protein expression related to motility, signal transduction, energy metabolism, cell envelope biogenesis, and secondary metabolite production (Schmidt et al., 2017). The metabolomic analyses of S. plymuthica PRI-2C exposed to the fungal VOCs, the gene cluster comparison, and the heterologous co-expression of a terpene synthase and a methyltransferase revealed the production of the unusual terpene named sodorifen (Kai et al., 2010; Von Reuß et al., 2010) in response to fungal VOCs. These findings support the suggested importance of VOCs (and in particular terpenes) as signaling molecules in fungal–bacterial interactions.
Many soil bacteria can produce VOCs with antifungal effects and thus contribute to the phenomenon known as soil fungistasis where fungal propagules are restricted in their ability to grow or germinate (Garbeva et al., 2011). Recently, Cordovez et al. (2015) revealed that VOCs produced by Streptomyces spp. exhibit antifungal properties against Rhizoctonia solani and may contribute to plant disease suppressiveness. Ossowicki et al. (2017) showed that VOCs from the tomato rhizosphere isolate Pseudomonas donghuensis P482 have strong antifungal and anti-oomycete activity which suggests that the antagonistic capabilities of this strain against plant pathogens are due to their volatile potential (Ossowicki et al., 2017). This effect of bacterial VOCs against oomycetes is not an isolated case and other Pseudomonas strains have been reported to have anti-oomycete activities (De Vrieze et al., 2015; Hunziker et al., 2015). In a recent report, VOCs produced by several Lysobacter strains growing in a protein-rich medium showed anti-oomycete activity whereas non-antagonistic VOCs were produced by these strains when grown on a sugar-rich medium. This indicates that the production of volatiles is highly dependent on growth conditions and nutrient availability (Lazazzara et al., 2017).
Fungi–Fungi
The 1-octen-3-ol, one of the most prominent fungal VOC, known as the mushroom smell, is produced by a wide range of filamentous fungi and can function as a development signal among fungi (Miyamoto et al., 2014). The same compound was described to function in Penicillium paneum as a self-inhibitor signal in spore germination (Chitarra et al., 2004). As developmental signals during population establishment, certain fungal VOCs act in a concentration-dependent manner to regulate conspecific mycelial growth and spore germination (Nemčovič et al., 2008; Stoppacher et al., 2010).
Fungal VOCs can have inhibitory effects and drive antagonistic interactions among fungi. For example, the endophytic fungi Muscodor albus and Oxyporus latemarginatus can strongly inhibit the growth of several plant pathogenic fungi, including Botrytis cinerea and Rhizoctonia solani (Strobel et al., 2001). VOCs emitted by Trichoderma spp. have a strong effect against plant pathogenic fungi such as Fusarium oxysporum, Rhizoctonia solani, Sclerotium rolfsii, Sclerotinia sclerotiorum, and Alternaria brassicicola (Amin et al., 2010). Similarly, VOCs such as 5-hexenoic acid, limonene, octanoic acid and 3,4-2H-dihydropyran produced by the non-pathogenic fungus F. oxysporum CanR-46 could inhibit mycelial growth of 14 fungal species including the pathogenic Verticillium dahlia (Zhang et al., 2015). Recently, a proteomic study demonstrated that fungal VOCs can interfere with essential metabolic pathways to prevent fungal growth (Fialho et al., 2016).
Some fungal species can detoxify the antifungal compounds produced by their microbial competitors. For example, Fusarium graminearum can detoxify the toxic compound 6-pentyl-alpha-pyrone, emitted by Trichoderma harzianum (Cooney et al., 2001). Fungal VOCs can be important carbon sources for fungi colonizing carbon-limited environments (Cale et al., 2016). Conversely, for fungi colonizing a more carbon-rich environment, VOCs may act, in a concentration-dependent manner, as semio-chemicals to mediate antagonistic and beneficial interactions between fungi.
Protists–Bacteria
A very diverse and abundant group of soil microorganisms are protists (Protozoa) (Fierer and Jackson, 2006; Geisen et al., 2015). Due to their grazing activities, protists play an important role in the soil food web and significantly affect carbon allocation and nutrient-cycling in the soil-plant-interphase (Geisen et al., 2016). Most soil protists are known to be key predators of bacteria and can shape bacterial communities by selective feeding (Griffiths et al., 1999; Bonkowski and Brandt, 2002; Rosenberg et al., 2009; Glücksman et al., 2010). Reaching suitable prey is very energy consuming (Jousset, 2012). Thus, sensing their prey over long distances in the porous soil matrix would be very beneficial for protists. A recent study by Schulz-Bohm et al. (2017) revealed that volatile organic compounds can play a key role in long-distance bacterial–protists interactions. By testing various volatile-mediated interactions between phylogenetically different soil bacteria and protists and comparing those with direct trophic interactions, they demonstrated that specific bacterial volatiles can provide early information about suitable prey. In particular, it was shown that terpenes such as β-linalool, β-pinene, germacrene D-4-ol or δ-cadinene produced by C. pratensis Ter91 (Song et al., 2015b) can stimulate protist activity and motility suggesting that terpenes can be key components in VOCs-mediated communication between protists and bacteria (Schulz-Bohm et al., 2017). Interestingly, soil protists such as Dictyostelium discoideum (Chen et al., 2016) produce volatile terpenes. These terpenes might be involved in defense mechanisms, for example, to repel nematode predators. Similarly, it was shown that soil bacteria can produce specific volatiles to repel protist predators (Kai et al., 2009; Schulz-Bohm et al., 2017).
Besides bacterivorous protists, obligate and facultative mycophageous (fungus grazing) protists are common soil inhabitants (Geisen, 2016). Mycophageous protists feed mostly on yeast and fungal spores while some specialists are able to graze directly on the hyphae of filamentous fungi (Geisen et al., 2016). It is well known that soil fungi such as yeast produce a wide set of volatile compounds involved in various belowground interactions (Effmert et al., 2012; Werner et al., 2016). Thus, although not demonstrated yet, it is plausible that fungal volatiles might play an important role in belowground communication between soil fungi and protists, as well.
VOCs in Microbe-Plant Interactions
In recent years, evidence supporting the idea that plants respond strongly to mVOCs has grown. Most of the research carried out so far has investigated the impact of microbial VOCs on the model plant Arabidopsis thaliana. This has revealed that, without physical contact, microorganisms are able to drastically alter plant root system development, plant physiology, hormonal pathways, and biomass production (Ryu et al., 2004; Blom et al., 2011; Wenke et al., 2012; Bailly et al., 2014; Bitas et al., 2015; Ditengou et al., 2015; Li et al., 2016; Piechulla et al., 2017). mVOCs can also function as a direct source of nutrients for plants (Meldau et al., 2013), induce resistance to pathogens in plants (D’Alessandro et al., 2014; Kottb et al., 2015; Song et al., 2015b; Wintermans et al., 2016), affect plant secondary metabolite production (Santoro et al., 2011), directly inhibit plant pathogens (Kai et al., 2009; Garbeva et al., 2014b; De Vrieze et al., 2015; Kottb et al., 2015) and induce soil fungistasis and suppressiveness (Garbeva et al., 2011; Van Agtmaal et al., 2015). Moreover, one single mVOC can show various functions, such as dimethyl disulfide, which improves plant growth by enhancing the availability of reduced sulfur (Meldau et al., 2013). It also protects tobacco and corn plants against Botrytis cinerea and Cochliobolus heterostrophus by directly inhibiting pathogens and inducing systemic resistance in plants (Huang C.-J. et al., 2012). Likewise, a characteristic compound of Trichoderma asperellum, 6-pentyl-pyrone, can increase plant defense reactions and at the same time decrease B. cinerea and Alternaria alternata sporulation (Kottb et al., 2015).
Many independent studies revealed that mVOCs emitted by beneficial soil microorganisms can affect plant growth but only few studies focused on how VOCs produced by soil-borne plant pathogens affect plant growth and development. These studies suggest that mVOCs from plant pathogens may modulate the trade-off between plant growth, development and defense. Bitas et al. (2015) showed that VOCs emitted by pathogenic F. oxysporum promoted the growth of A. thaliana and Nicotiana tabacum and affected auxin transport and signaling. VOCs emitted by the pathogen Alternaria alternaria enhanced growth, early flowering and photosynthesis rates of Arabidopsis, maize and pepper by affecting the levels of plastidic cytocinin (Sanchez-Lopez et al., 2016). A more recent study showed that the soil-borne pathogen Rhizoctonia solani produced an array of mVOCs that promote plant growth, accelerate development, change plant VOCs emission and reduce insect resistance (Cordovez et al., 2017). This must be a successful strategy for the pathogenic fungi since with increased root biomass and stimulation of lateral root formation there is a greater surface area for fungal colonization and infection.
When analyzing mVOCs effects on plant growth, it is important to take into account, that microorganisms can produce high amounts of CO2 that can promote plant growth (Kai and Piechulla, 2009; Piechulla et al., 2017). Hence, a good experimental setup with appropriate controls are required to avoid artifacts in the results (Piechulla et al., 2017).
Alternatively, plants are able to mediate the belowground plant–microbe interactions via root-emitted VOCs (Wenke et al., 2010). Root-derived VOCs may serve multiple roles such as carbon sources, defense metabolites and chemo-attractants (Van Dam et al., 2016). Rhizobacteria such as Pseudomonas fluorescens and Alcaligenes xylosoxidans have been shown to metabolize α- pinene as their sole carbon source (Kleinheinz et al., 1999). Del Giudice et al. (2008) also reported that bacteria associated with the roots of vetiver grass (Vetiveria zizanioides) use sesquiterpenes as a carbon source. Undoubtedly, plants and soil microorganisms are engaged via VOCs in long-distance interactions (Van Dam et al., 2016). However, so far, limited knowledge exists concerning the role of plant VOCs in attracting beneficial organisms and how plant-associated microorganisms affect the quantity and quality of plant volatile emission. Only recently, using a glass olfactometer system, the attraction of distant soil bacteria by VOCs emitted by plant roots was revealed (Schulz-Bohm et al., 2017). Olfactometer systems have been used successfully to study aboveground plant–herbivores interactions (Ballhorn and Kautz, 2013) or belowground plant–nematode interactions (Rasmann et al., 2005). However, this is the first case to apply an olfactometer to study plant–microbe interactions. Moreover, the same study revealed that upon fungal infection, the blend of root VOCs changed and specific bacteria with antifungal properties were attracted (Schulz-Bohm et al., 2017).
mVOCs-Mediated Dialog
Several reports describe the chemical dialog between microbes, plants, and other organisms by the exchange of soluble compounds (Badri et al., 2009; Lira et al., 2015; Song et al., 2015a; Liu et al., 2016). Most of the studies reporting mVOCs-mediated communication belowground focus on the uni-directional responses and only a few studies reported on bi-directional mVOCs-mediated interactions. For instance, the importance of mVOCs in the dialog between the fungal plant pathogen Verticillium longisporum and its bacterial antagonist Paenibacillus polymyxa was recently revealed in both in vitro and in planta experiments (Rybakova et al., 2017). Both microorganisms responded to one another’s VOCs and this specific mVOCs-mediated interaction resulted in the inhibition of cellular metabolism and growth reduction of the fungal pathogen.
A VOCs-mediated dialog between bacteria and fungi was also reported by Spraker et al. (2014) where VOCs of the fungal plant pathogen Aspergillus flavus reduced the production of the major virulence factor EPS of the bacterial plant pathogen R. solanacearum. In parallel, A. flavus responded to VOCs of R. solanacearum by reducing conidia production and increasing aflatoxin production.
Conclusion and Outlook
Over the last decades, our understanding of the chemical complexity of mVOCs produced by many different soil microorganisms has grown. It is clear that these small and odorous molecules can modify the behavior and promote or inhibit growth of neighboring organisms (Figure 1).
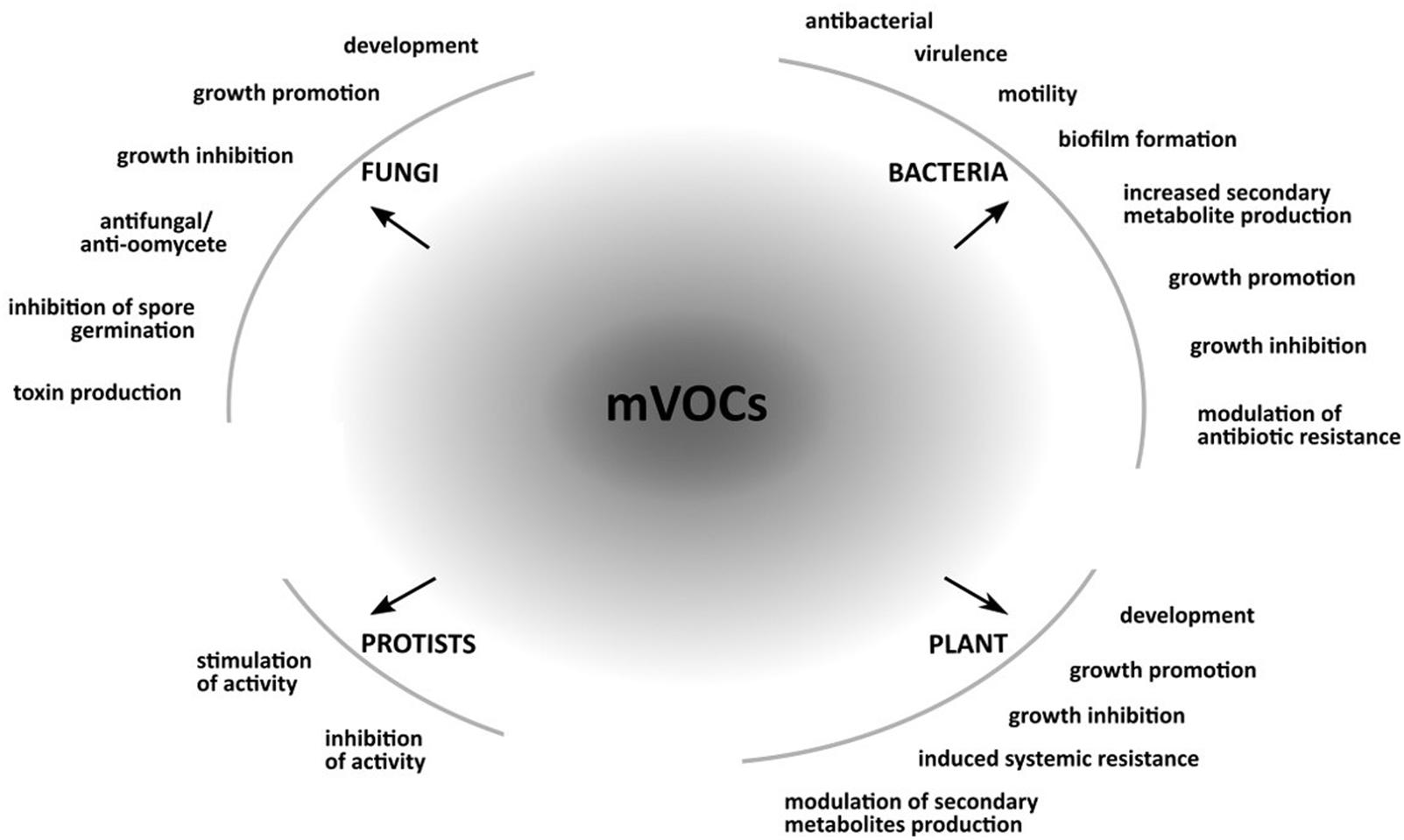
FIGURE 1. Responses in bacteria, fungi, protists and plants caused by microbial volatile organic compounds (mVOCs). The figure shows examples of responses caused my mVOCs in bacteria, fungi, protists, and plants.
Most existing studies on mVOCs are focused on describing the uni-directional effect of mVOCs produced by a single organism and the responses of the organisms perceiving them without considering mVOCs-mediated dialog and the bi-directional responses to one another. Furthermore, microbial interactions taking place belowground are far more complex than single one-to-one interactions and involve more organisms, which can significantly affect mVOCs emission. For example, fungal-associated bacteria have been shown to affect the production of VOCs in fungi (Schulz-Bohm et al., 2015; Splivallo et al., 2015) and in addition, they can affect the fungal plant-pathogenicity and repress the expression of fungal virulence genes (Minerdi et al., 2009). Therefore, a holistic approach considering the effect of mVOCs on belowground soil community is needed. For instance, using a metagenomics approach Yuan et al. (2017) revealed that mVOCs could alter the composition of soil bacterial and fungal communities and significantly increased the relative abundance of Proteobacteria, Bacteroidetes, Firmicutes, and Ascomycota. Furthermore, mVOCs influenced genes involved in important soil functions such as N-fixation (nifH), nitrification (amoA), denitrification (nirS) and antibiotic production (NRPS) (Yuan et al., 2017).
From the current scientific literature, it is clear that the most studied belowground mVOCs-mediated interactions are the interactions between bacteria, fungi and plants (Figure 1). There is a lack of knowledge relating to the emission of VOCs by protists, archaea or other rhizosphere organisms, such as nematodes or earthworms, indicating that these groups are currently understudied with regards to this aspect.
Several VOCs are commonly produced and emitted by both plant roots, fungi, bacteria and protists and it is possible that these compounds act as a ‘lingua franca’ for intra- and inter-kingdom communication between these organisms. Let us take as an example only one chemical class, the terpenes. Terpenes are the largest and most diverse class of metabolites known to date. They are best known to humans as plants metabolites. However, recent studies revealed that terpenes can be produced by all kingdoms of life including prokaryotes (Takamatsu et al., 2011; Yamada et al., 2012, 2015; Song et al., 2015b; Chen et al., 2016). Recently, Yamada et al. (2015) described a powerful bioinformatics method based on the use of Hidden Markov Models (HMMs) and Protein Families Database (PFAM) search that has allowed the discovery of terpene synthases of bacterial origin and showed that phylogenetically different bacteria can be a rich source of terpenes. Both the number, the wide distribution, and the structural diversity of terpenes provide enormous potential for mediating significant chemical interactions and communication belowground. Examples of terpene-mediated microbial interactions are presented in Figure 2 and Table 1, indicating the ecological importance of terpenes in interactions between soil micro- and macro-organisms, including plant roots.
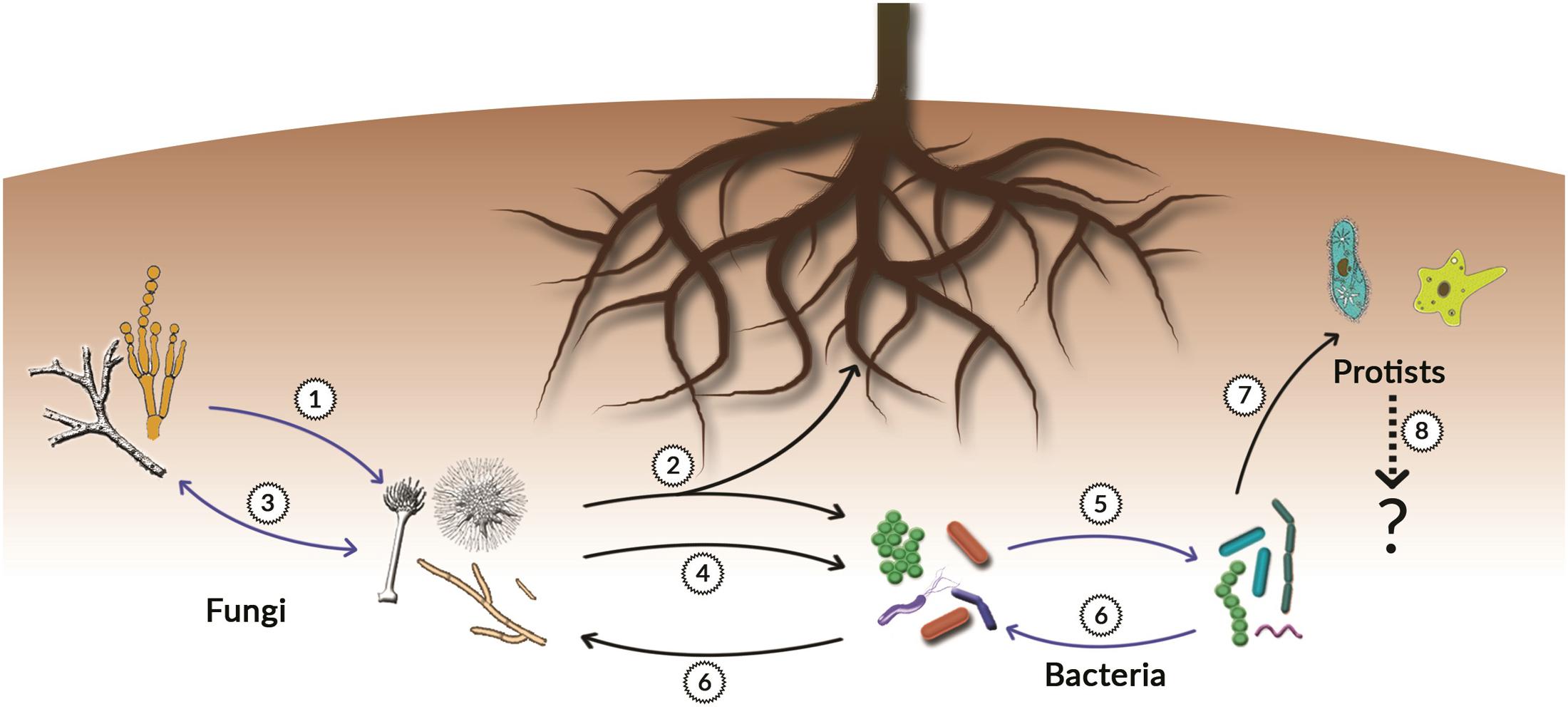
FIGURE 2. Terpenes-mediated belowground interactions. The figure shows examples of interactions between different organisms in the rhizosphere that are mediated by microbial terpenes. Blue arrows indicate intra-kingdom interactions while black arrows indicate inter-kingdom interactions. The numbers in the figure correspond with the numbers in Table 1.

TABLE 1. Examples of terpenes involved in belowground microbial interactions.
Despite the rapid increasing numbers of studies showing the importance of mVOCs in the long-distance belowground chemical interactions, we still do not know exactly how VOCs are recognized and perceived. VOCs receptors or other perception mechanisms have not been identified in any of the described cases. The big challenge is to determine whether VOCs are internalized and transduced by receptor-mediated processes, whether they interact with the cell membrane to initiate signal transduction cascades or whether they are simply taken up by the cell and metabolized (Widhalm et al., 2015; Adebesin et al., 2017; Tissier et al., 2017). For plants, the current view is that due to their lipophilic nature, VOCs such as mono- and sesquiterpenes may interfere with membrane structures, thereby causing depolarization of the membranes and triggering Ca2+-signaling in plants (Maffei et al., 2001; Heil and Land, 2014). For further deciphering of mVOC-mediated microbe-microbe interactions, the mVOCs microbial perception mechanism needs to be elucidated. The application of methods for screening of mutant strains may be useful for that purpose, to identify microbial genes and proteins that are required for VOCs perception.
Another big challenge is to determine what concentrations of mVOCs are produced in soil and at what distances these mVOCs are eliciting a biological response in other organisms. There is the possibility that, similar to the roles of antibiotics in nature (Davies et al., 2006; Yim et al., 2006; Romero et al., 2011), mVOCs could have concentration-dependent function either as weapons in intercellular chemical warfare or as signaling compounds when they are present in low concentrations.
Concerning the implementations of mVOCs, our knowledge on the potential use of those compounds in large-scale agriculture and horticulture is still limited. In agriculture systems, mVOCs have to be applied under open-field conditions, which are very different from the in vitro conditions currently used in most studies. There are very few studies assessing the effects of mVOCs application under open conditions and they have been summarized in a recent review from Chung et al. (2016). Since it was discovered that the 2,3-butanediol elicited plant growth and induced systemic resistance (Ryu et al., 2003, 2004), several studies have applied this compound or the producing strains to the soil of open fields to test its effects under agricultural conditions and have revealed promising results (Velivelli et al., 2015). Dimethyl disulfide, frequently emitted by many bacteria, is another compound used in recent years in the novel soil fumigant PALADIN® that targets nematodes and soil-borne pathogens. However, the research concerning the application of other mVOCs in agriculture is still in its infancy. We now live in a time in which the old methods of using chemicals to protect crops need to be replaced with and, in some cases, complemented by green solutions. The traditional harmful synthetic fungicides currently used could be replaced with the so far under-explored and unique mVOCs for which significant proof of plant growth promoting effects and plant protection ability already exists. In spite of the obvious potential of mVOCs in agriculture, the field suffers from the common ‘translational gap’ because of a lack of studies evaluating other unexpected effects of those bioactive molecules on non-target beneficial soil organisms.
Author Contributions
All authors contributed and approved the manuscript.
Funding
This work is financed by Netherlands Organization for Scientific Research (NWO) VIDI personal grant 864.11.015.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
The authors want to thank Adam Ossowicki, for his great help in making the figure. This is publication 6429 of the NIOO-KNAW.
References
Adebesin, F., Widhalm, J. R., Boachon, B., Lefèvre, F., Pierman, B., Lynch, J. H., et al. (2017). Emission of volatile organic compounds from petunia flowers is facilitated by an ABC transporter. Science 356, 1386–1388. doi: 10.1126/science.aan0826
Amin, F., Razdan, V., Mohiddin, F., Bhat, K., and Sheikh, P. (2010). Effect of volatile metabolites of Trichoderma species against seven fungal plant pathogens in-vitro. J. Phytol. 2, 34–37.
Audrain, B., Farag, M. A., Ryu, C. M., and Ghigo, J. M. (2015). Role of bacterial volatile compounds in bacterial biology. FEMS Microbiol. Rev. 39, 222–233. doi: 10.1093/femsre/fuu013
Badri, D. V., Weir, T. L., van der Lelie, D., and Vivanco, J. M. (2009). Rhizosphere chemical dialogues: plant–microbe interactions. Curr. Opin. Biotechnol. 20, 642–650. doi: 10.1016/j.copbio.2009.09.014
Bailly, A., Groenhagen, U., Schulz, S., Geisler, M., Eberl, L., and Weisskopf, L. (2014). The inter-kingdom volatile signal indole promotes root development by interfering with auxin signalling. Plant J. 80, 758–771. doi: 10.1111/tpj.12666
Ballhorn, D. J., and Kautz, S. (2013). How useful are olfactometer experiments in chemical ecology research? Commun. Integr. Biol. 6:e24787. doi: 10.4161/cib.24787
Bitas, V., McCartney, N., Li, N., Demers, J., Kim, J.-E., Kim, H.-S., et al. (2015). Fusarium oxysporum volatiles enhance plant growth via affecting auxin transport and signaling. Front. Microbiol. 6:1248. doi: 10.3389/fmicb.2015.01248
Blom, D., Fabbri, C., Eberl, L., and Weisskopf, L. (2011). Volatile-mediated killing of Arabidopsis thaliana by bacteria is mainly due to hydrogen cyanide. Appl. Environ. Microbiol. 77, 1000–1008. doi: 10.1128/aem.01968-10
Bonkowski, M., and Brandt, F. (2002). Do soil protozoa enhance plant growth by hormonal effects? Soil Biol. Biochem. 34, 1709–1715. doi: 10.1016/S0038-0717(02)00157-8
Cale, J. A., Collignon, R. M., Klutsch, J. G., Kanekar, S. S., Hussain, A., and Erbilgin, N. (2016). Fungal volatiles can act as carbon sources and semiochemicals to mediate interspecific interactions among bark beetle-associated fungal symbionts. PLOS ONE 11:e0162197. doi: 10.1371/journal.pone.0162197
Chen, X., Kollner, T. G., Jia, Q., Norris, A., Santhanam, B., Rabe, P., et al. (2016). Terpene synthase genes in eukaryotes beyond plants and fungi: occurrence in social amoebae. Proc. Natl. Acad. Sci. U.S.A. 113, 12132–12137. doi: 10.1073/pnas.1610379113
Cheng, X., Cordovez, V., Etalo, D. W., van der Voort, M., and Raaijmakers, J. M. (2016). Role of the GacS sensor kinase in the regulation of volatile production by plant growth-promoting Pseudomonas fluorescens SBW25. Front. Plant Sci. 7:1706. doi: 10.3389/fpls.2016.01706
Chitarra, G. S., Abee, T., Rombouts, F. M., Posthumus, M. A., and Dijksterhuis, J. (2004). Germination of Penicillium paneum conidia is regulated by 1-octen-3-ol, a volatile self-inhibitor. Appl. Environ. Microbiol. 70, 2823–2829. doi: 10.1128/AEM.70.5.2823-2829.2004
Chung, J.-H., Song, G. C., and Ryu, C.-M. (2016). Sweet scents from good bacteria: case studies on bacterial volatile compounds for plant growth and immunity. Plant Mol. Biol. 90, 677–687. doi: 10.1007/s11103-015-0344-8
Cooney, J. M., Lauren, D. R., and di Menna, M. E. (2001). Impact of competitive fungi on trichothecene production by Fusarium graminearum. J. Agric. Food Chem. 49, 522–526. doi: 10.1021/jf0006372
Cordovez, V., Carrion, V. J., Etalo, D. W., Mumm, R., Zhu, H., van Wezel, G. P., et al. (2015). Diversity and functions of volatile organic compounds produced by Streptomyces from a disease-suppressive soil. Front. Microbiol. 6:1081. doi: 10.3389/fmicb.2015.01081
Cordovez, V., Mommer, L., Moisan, K., Lucas-Barbosa, D., Pierik, R., Mumm, R., et al. (2017). Plant phenotypic and transcriptional changes induced by volatiles from the fungal root pathogen Rhizoctonia solani. Front. Plant Sci. 8:1262. doi: 10.3389/fpls.2017.01262
D’Alessandro, M., Erb, M., Ton, J., Brandenburg, A., Karlen, D., Zopfi, J., et al. (2014). Volatiles produced by soil-borne endophytic bacteria increase plant pathogen resistance and affect tritrophic interactions. Plant Cell Environ. 37, 813–826. doi: 10.1111/pce.12220
Dandurishvili, N., Toklikishvili, N., Ovadis, M., Eliashvili, P., Giorgobiani, N., Keshelava, R., et al. (2011). Broad-range antagonistic rhizobacteria Pseudomonas fluorescens and Serratia plymuthica suppress Agrobacterium crown gall tumours on tomato plants. J. Appl. Microbiol. 110, 341–352. doi: 10.1111/j.1365-2672.2010.04891.x
Davies, J., Spiegelman, G. B., and Yim, G. (2006). The world of subinhibitory antibiotic concentrations. Curr. Opin. Microbiol. 9, 445–453. doi: 10.1016/j.mib.2006.08.006
De Vrieze, M., Pandey, P., Bucheli, T. D., Varadarajan, A. R., Ahrens, C. H., Weisskopf, L., et al. (2015). Volatile organic compounds from native potato-associated Pseudomonas as potential anti-oomycete agents. Front. Microbiol. 6:1295. doi: 10.3389/fmicb.2015.01295
Del Giudice, L., Massardo, D. R., Pontieri, P., Bertea, C. M., Mombello, D., Carata, E., et al. (2008). The microbial community of Vetiver root and its involvement into essential oil biogenesis. Environ. Microbiol. 10, 2824–2841. doi: 10.1111/j.1462-2920.2008.01703.x
Delory, B. M., Delaplace, P., Fauconnier, M.-L., and du Jardin, P. (2016). Root-emitted volatile organic compounds: can they mediate belowground plant-plant interactions? Plant Soil 402, 1–26. doi: 10.1007/s11104-016-2823-3
Ditengou, F. A., Muller, A., Rosenkranz, M., Felten, J., Lasok, H., van Doorn, M. M., et al. (2015). Volatile signalling by sesquiterpenes from ectomycorrhizal fungi reprogrammes root architecture. Nat. Commun. 6:6279. doi: 10.1038/ncomms7279
Effmert, U., Kalderas, J., Warnke, R., and Piechulla, B. (2012). Volatile mediated interactions between bacteria and fungi in the soil. J. Chem. Ecol. 38, 665–703. doi: 10.1007/s10886-012-0135-5
Fialho, M. B., de Andrade, A., Bonatto, J. M. C., Salvato, F., Labate, C. A., and Pascholati, S. F. (2016). Proteomic response of the phytopathogen Phyllosticta citricarpa to antimicrobial volatile organic compounds from Saccharomyces cerevisiae. Microbiol. Res. 183(Suppl. C), 1–7. doi: 10.1016/j.micres.2015.11.002
Fierer, N., and Jackson, R. B. (2006). The diversity and biogeography of soil bacterial communities. Proc. Natl. Acad. Sci. U.S.A. 103, 626–631. doi: 10.1073/pnas.0507535103
Garbeva, P., Hol, W. H. G., Termorshuizen, A. J., Kowalchuk, G. A., and de Boer, W. (2011). Fungistasis and general soil biostasis - a new synthesis. Soil Biol. Biochem. 43, 469–477. doi: 10.1016/j.soilbio.2010.11.020
Garbeva, P., Hordijk, C., Gerards, S., and de Boer, W. (2014a). Volatile-mediated interactions between phylogenetically different soil bacteria. Front. Microbiol. 5:289. doi: 10.3389/fmicb.2014.00289
Garbeva, P., Hordijk, C., Gerards, S., and de Boer, W. (2014b). Volatiles produced by the mycophagous soil bacterium Collimonas. FEMS Microbiol. Ecol. 87, 639–649. doi: 10.1111/1574-6941.12252
Geisen, S. (2016). The bacterial-fungal energy channel concept challenged by enormous functional versatility of soil protists. Soil Biol. Biochem. 102, 22–25. doi: 10.1016/j.soilbio.2016.06.013
Geisen, S., Koller, R., Hunninghaus, M., Dumack, K., Urich, T., and Bonkowski, M. (2016). The soil food web revisited: diverse and widespread mycophagous soil protists. Soil Biol. Biochem. 94, 10–18. doi: 10.1016/j.soilbio.2015.11.010
Geisen, S., Tveit, A. T. I., Clark, M., Richter, A., Svenning, M. M., Bonkowski, M., et al. (2015). Metatranscriptomic census of active protists in soils. ISME J. 9, 2178–2190. doi: 10.1038/ismej.2015.30
Glücksman, E., Bell, T., Griffiths, R. I., and Bass, D. (2010). Closely related protist strains have different grazing impacts on natural bacterial communities. Environ. Microbiol. 12, 3105–3113. doi: 10.1111/j.1462-2920.2010.02283.x
Griffiths, B. S., Bonkowski, M., Dobson, G., and Caul, S. (1999). Changes in soil microbial community structure in the presence of microbial-feeding nematodes and protozoa. Pedobiologia 43, 297–304.
Gürtler, H., Pedersen, R., Anthoni, U., Christophersen, C., Nielsen, P. H., Wellington, E. M., et al. (1994). Albaflavenone, a sesquiterpene ketone with a zizaene skeleton produced by a streptomycete with a new rope morphology. J. Antibiot. 47, 434–439. doi: 10.7164/antibiotics.47.434
Heil, M., and Land, W. G. (2014). Danger signals – damaged-self recognition across the tree of life. Front. Plant Sci. 5:578. doi: 10.3389/fpls.2014.00578
Hornby, J. M., Jensen, E. C., Lisec, A. D., Tasto, J. J., Jahnke, B., Shoemaker, R., et al. (2001). Quorum sensing in the dimorphic fungus Candida albicans is mediated by farnesol. Appl. Environ. Microbiol. 67, 2982–2992. doi: 10.1128/AEM.67.7.2982-2992.2001
Huang, C.-J., Tsay, J.-F., Chang, S.-Y., Yang, H.-P., Wu, W.-S., and Chen, C.-Y. (2012). Dimethyl disulfide is an induced systemic resistance elicitor produced by Bacillus cereus C1L. Pest Manage. Sci. 68, 1306–1310. doi: 10.1002/ps.3301
Huang, M., Sanchez-Moreiras, A. M., Abel, C., Sohrabi, R., Lee, S., Gershenzon, J., et al. (2012). The major volatile organic compound emitted from Arabidopsis thaliana flowers, the sesquiterpene (E)-beta-caryophyllene, is a defense against a bacterial pathogen. New Phytol. 193, 997–1008. doi: 10.1111/j.1469-8137.2011.04001.x
Hunziker, L., Bonisch, D., Groenhagen, U., Bailly, A., Schulz, S., and Weisskopf, L. (2015). Pseudomonas strains naturally associated with potato plants produce volatiles with high potential for inhibition of Phytophthora infestans. Appl. Environ. Microbiol. 81, 821–830. doi: 10.1128/AEM.02999-14
Insam, H., and Seewald, M. S. A. (2010). Volatile organic compounds (VOCs) in soils. Biol. Fertil. Soils 46, 199–213. doi: 10.1007/s00374-010-0442-3
Jones, S. E., Ho, L., Rees, C. A., Hill, J. E., Nodwell, J. R., and Elliot, M. A. (2017). Streptomyces exploration is triggered by fungal interactions and volatile signals. Elife 6:e21738. doi: 10.7554/eLife.21738
Jousset, A. (2012). Ecological and evolutive implications of bacterial defences against predators. Environ. Microbiol. 14, 1830–1843. doi: 10.1111/j.1462-2920.2011.02627.x
Kai, M., Crespo, E., Cristescu, S. M., Harren, F. J. M., Francke, W., and Piechulla, B. (2010). Serratia odorifera: analysis of volatile emission and biological impact of volatile compounds on Arabidopsis thaliana. Appl. Microbiol. Biotechnol. 88, 965–976. doi: 10.1007/s00253-010-2810-1
Kai, M., Haustein, M., Molina, F., Petri, A., Scholz, B., and Piechulla, B. (2009). Bacterial volatiles and their action potential. Appl. Microbiol. Biotechnol. 81, 1001–1012. doi: 10.1007/s00253-008-1760-3
Kai, M., and Piechulla, B. (2009). Plant growth promotion due to rhizobacterial volatiles – An effect of CO2? FEBS Lett. 583, 3473–3477. doi: 10.1016/j.febslet.2009.09.053
Kanchiswamy, C. N., Mainoy, M., and Maffei, M. E. (2015). Chemical diversity of microbial volatiles and their potential for plant growth and productivity. Front. Plant Sci. 6:151. doi: 10.3389/fpls.2015.00151
Kleinheinz, G. T., Bagley, S. T., St. John, W. P., Rughani, J. R., and McGinnis, G. D. (1999). Characterization of alpha-pinene-degrading microorganisms and application to a bench-scale biofiltration system for VOC degradation. Arch. Environ. Contam. Toxicol. 37, 151–157. doi: 10.1007/s002449900500
Kottb, M., Gigolashvili, T., Grosskinsky, D. K., and Piechulla, B. (2015). Trichoderma volatiles effecting Arabidopsis: from inhibition to protection against phytopathogenic fungi. Front. Microbiol. 6:995. doi: 10.3389/fmicb.2015.00995
Lazazzara, V., Perazzolli, M., Pertot, I., Biasioli, F., Puopolo, G., and Cappellin, L. (2017). Growth media affect the volatilome and antimicrobial activity against Phytophthora infestans in four Lysobacter type strains. Microbiol. Res. 201, 52–62. doi: 10.1016/j.micres.2017.04.015
Lemfack, M. C., Gohlke, B.-O., Toguem, S. M. T., Preissner, S., Piechulla, B., and Preissner, R. (2017). mVOC 2.0: a database of microbial volatiles. Nucleic Acids Res. doi: 10.1093/nar/gkx1016 [Epub ahead of print].
Lemfack, M. C., Nickel, J., Dunkel, M., Preissner, R., and Piechulla, B. (2014). mVOC: a database of microbial volatiles. Nucleic Acids Res. 42, D744–D748. doi: 10.1093/nar/gkt1250
Li, N. X., Alfiky, A., Vaughan, M. M., and Kang, S. (2016). Stop and smell the fungi: fungal volatile metabolites are overlooked signals involved in fungal interaction with plants. Fungal Biol. Rev. 30, 134–144. doi: 10.1016/j.fbr.2016.06.004
Lira, M. A. Jr., Nascimento, L. R., and Fracetto, G. G. (2015). Legume-rhizobia signal exchange: promiscuity and environmental effects. Front. Microbiol. 6:945. doi: 10.3389/fmicb.2015.00945
Liu, Y., Chen, L., Zhang, N., Li, Z., Zhang, G., Xu, Y., et al. (2016). Plant-microbe communication enhances auxin biosynthesis by a root-associated bacterium, Bacillus amyloliquefaciens SQR9. Mol. Plant Microbe Interact. 29, 324–330. doi: 10.1094/MPMI-10-15-0239-R
Lutz, M. P., Wenger, S., Maurhofer, M., Défago, G., and Duffy, B. (2004). Signaling between bacterial and fungal biocontrol agents in a strain mixture. FEMS Microbiol. Ecol. 48, 447–455. doi: 10.1016/j.femsec.2004.03.002
Maffei, M., Camusso, W., and Sacco, S. (2001). Effect of Mentha × piperita essential oil and monoterpenes on cucumber root membrane potential. Phytochemistry 58, 703–707. doi: 10.1016/S0031-9422(01)00313-2
Martins, M., Henriques, M., Azeredo, J., Rocha, S. M., Coimbra, M. A., and Oliveira, R. (2007). Morphogenesis control in Candida albicans and Candida dubliniensis through signaling molecules produced by planktonic and biofilm cells. Eukaryot. Cell 6, 2429–2436. doi: 10.1128/EC.00252-07
Meldau, D. G., Meldau, S., Hoang, L. H., Underberg, S., Wunsche, H., and Baldwin, I. T. (2013). Dimethyl disulfide produced by the naturally associated bacterium Bacillus sp B55 promotes Nicotiana attenuata growth by enhancing sulfur nutrition. Plant Cell 25, 2731–2747. doi: 10.1105/tpc.113.114744
Minerdi, D., Bossi, S., Gullino, M. L., and Garibaldi, A. (2009). Volatile organic compounds: a potential direct long-distance mechanism for antagonistic action of Fusarium oxysporum strain MSA 35. Environ. Microbiol. 11, 844–854. doi: 10.1111/j.1462-2920.2008.01805.x
Minerdi, D., Bossi, S., Maffei, M. E., Gullino, M. L., and Garibaldi, A. (2011). Fusarium oxysporum and its bacterial consortium promote lettuce growth and expansin A5 gene expression through microbial volatile organic compound (MVOC) emission. FEMS Microbiol. Ecol. 76, 342–351. doi: 10.1111/j.1574-6941.2011.01051.x
Miyamoto, K., Murakami, T., Kakumyan, P., Keller, N. P., and Matsui, K. (2014). Formation of 1-octen-3-ol from Aspergillus flavus conidia is accelerated after disruption of cells independently of Ppo oxygenases, and is not a main cause of inhibition of germination. PeerJ 2:e395. doi: 10.7717/peerj.395
Nemčovič, M., Jakubíková, L., Víden, I., and Farkaš, V. (2008). Induction of conidiation by endogenous volatile compounds in Trichoderma spp. FEMS Microbiol. Lett. 284, 231–236. doi: 10.1111/j.1574-6968.2008.01202.x
Ossowicki, A., Jafra, S., and Garbeva, P. (2017). The antimicrobial volatile power of the rhizospheric isolate Pseudomonas donghuensis P482. PLOS ONE 12:e0174362. doi: 10.1371/journal.pone.0174362
Pauliuc, I., and Botǎu, D. (2013). Antibacterial activity of Pleurotus ostreatus gemmotherapic extract. J. Hort. For. Biotechnol. 17, 242–245.
Piechulla, B., Lemfack, M. C., and Kai, M. (2017). Effects of discrete bioactive microbial volatiles on plants and fungi. Plant Cell Environ. 40, 2042–2067. doi: 10.1111/pce.13011
Rajer, F. U., Wu, H., Xie, Y., Xie, S., Raza, W., Tahir, H. A. S., et al. (2017). Volatile organic compounds produced by a soil-isolate, Bacillus subtilis FA26 induce adverse ultra-structural changes to the cells of Clavibacter michiganensis ssp. sepedonicus, the causal agent of bacterial ring rot of potato. Microbiology 163, 523–530. doi: 10.1099/mic.0.000451
Rasmann, S., Köllner, T. G., Degenhardt, J., Hiltpold, I., Toepfer, S., Kuhlmann, U., et al. (2005). Recruitment of entomopathogenic nematodes by insect-damaged maize roots. Nature 434, 732–737. doi: 10.1038/nature03451
Raza, W., Ling, N., Liu, D., Wei, Z., Huang, Q., and Shen, Q. (2016a). Volatile organic compounds produced by Pseudomonas fluorescens WR-1 restrict the growth and virulence traits of Ralstonia solanacearum. Microbiol. Res. 192, 103–113. doi: 10.1016/j.micres.2016.05.014
Raza, W., Ling, N., Yang, L., Huang, Q., and Shen, Q. (2016b). Response of tomato wilt pathogen Ralstonia solanacearum to the volatile organic compounds produced by a biocontrol strain Bacillus amyloliquefaciens SQR-9. Sci. Rep. 6:24856. doi: 10.1038/srep24856
Raza, W., Wang, J., Wu, Y., Ling, N., Wei, Z., Huang, Q., et al. (2016c). Effects of volatile organic compounds produced by Bacillus amyloliquefaciens on the growth and virulence traits of tomato bacterial wilt pathogen Ralstonia solanacearum. Appl. Microbiol. Biotechnol. 100, 7639–7650. doi: 10.1007/s00253-016-7584-7
Romero, D., Traxler, M. F., López, D., and Kolter, R. (2011). Antibiotics as signal molecules. Chem. Rev. 111, 5492–5505. doi: 10.1021/cr2000509
Rosenberg, K., Bertaux, J., Krome, K., Hartmann, A., Scheu, S., and Bonkowski, M. (2009). Soil amoebae rapidly change bacterial community composition in the rhizosphere of Arabidopsis thaliana. ISME J. 3, 675–684. doi: 10.1038/ismej.2009.11
Rybakova, D., Rack-Wetzlinger, U., Cernava, T., Schaefer, A., Schmuck, M., and Berg, G. (2017). Aerial warfare: a volatile dialogue between the plant pathogen Verticillium longisporum and its antagonist Paenibacillus polymyxa. Front. Plant Sci. 8:1294. doi: 10.3389/fpls.2017.01294
Ryu, C. M., Farag, M. A., Hu, C. H., Reddy, M. S., Kloepper, J. W., and Pare, P. W. (2004). Bacterial volatiles induce systemic resistance in Arabidopsis. Plant Physiol. 134, 1017–1026. doi: 10.1104/pp.103.026583
Ryu, C. M., Farag, M. A., Hu, C. H., Reddy, M. S., Wei, H. X., Pare, P. W., et al. (2003). Bacterial volatiles promote growth in Arabidopsis. Proc. Natl. Acad. Sci. U.S.A. 100, 4927–4932. doi: 10.1073/pnas.0730845100
Sanchez-Lopez, A. M., Baslam, M., De Diego, N., Munoz, F. J., Bahaji, A., Almagro, G., et al. (2016). Volatile compounds emitted by diverse phytopathogenic microorganisms promote plant growth and flowering through cytokinin action. Plant Cell Environ. 39, 2592–2608. doi: 10.1111/pce.12759
Santoro, M. V., Zygadlo, J., Giordano, W., and Banchio, E. (2011). Volatile organic compounds from rhizobacteria increase biosynthesis of essential oils and growth parameters in peppermint (Mentha piperita). Plant Physiol. Biochem. 49, 1177–1182. doi: 10.1016/j.plaphy.2011.07.016
Schenkel, D., Lemfack, M., Piechulla, B., and Splivallo, R. (2015). A meta-analysis approach for assessing the diversity and specificity of belowground root and microbial volatiles. Front. Plant Sci. 6:707. doi: 10.3389/fpls.2015.00707
Schmidt, R., Cordovez, V., de Boer, W., Raaijmakers, J., and Garbeva, P. (2015). Volatile affairs in microbial interactions. ISME J. 9, 2329–2335. doi: 10.1038/ismej.2015.42
Schmidt, R., Jager, V., Zuhlke, D., Wolff, C., Bernhardt, J., Cankar, K., et al. (2017). Fungal volatile compounds induce production of the secondary metabolite Sodorifen in Serratia plymuthica PRI-2C. Sci. Rep. 7:862. doi: 10.1038/s41598-017-00893-3
Schulz, S., and Dickschat, J. S. (2007). Bacterial volatiles: the smell of small organisms. Nat. Prod. Rep. 24, 814–842. doi: 10.1039/b507392h
Schulz-Bohm, K., Gerards, S., Hundscheid, M. P. J., Melenhorst, J., De Boer, W., and Garbeva, P. V. (2017). Calling from distance: Attraction of soil bacteria by plant root volatiles. ISME J. (accepted)
Schulz-Bohm, K., Geisen, S., Wubs, E. R., Song, C., de Boer, W., and Garbeva, P. (2017). The prey’s scent - volatile organic compound mediated interactions between soil bacteria and their protist predators. ISME J. 11, 817–820. doi: 10.1038/ismej.2016.144
Schulz-Bohm, K., Zweers, H., de Boer, W., and Garbeva, P. (2015). A fragrant neighborhood: volatile mediated bacterial interactions in soil. Front. Microbiol. 6:1212. doi: 10.3389/fmicb.2015.01212
Song, C., Mazzola, M., Cheng, X., Oetjen, J., Alexandrov, T., Dorrestein, P., et al. (2015a). Molecular and chemical dialogues in bacteria-protozoa interactions. Sci. Rep. 5:12837. doi: 10.1038/srep12837
Song, C., Schmidt, R., de Jager, V., Krzyzanowska, D., Jongedijk, E., Cankar, K., et al. (2015b). Exploring the genomic traits of fungus-feeding bacterial genus Collimonas. BMC Genomics 16:1103. doi: 10.1186/s12864-015-2289-3
Splivallo, R., Deveau, A., Valdez, N., Kirchhoff, N., Frey-Klett, P., and Karlovsky, P. (2015). Bacteria associated with truffle-fruiting bodies contribute to truffle aroma. Environ. Microbiol. 17, 2647–2660. doi: 10.1111/1462-2920.12521
Spraker, J. E., Jewell, K., Roze, L. V., Scherf, J., Ndagano, D., Beaudry, R., et al. (2014). A volatile relationship: profiling an inter-kingdom dialogue between two plant pathogens, Ralstonia solanacearum and Aspergillus flavus. J. Chem. Ecol. 40, 502–513. doi: 10.1007/s10886-014-0432-2
Stoppacher, N., Kluger, B., Zeilinger, S., Krska, R., and Schuhmacher, R. (2010). Identification and profiling of volatile metabolites of the biocontrol fungus Trichoderma atroviride by HS-SPME-GC-MS. J. Microbiol. Methods 81, 187–193. doi: 10.1016/j.mimet.2010.03.011
Strobel, G. A., Dirkse, E., Sears, J., and Markworth, C. (2001). Volatile antimicrobials from Muscodor albus, a novel endophytic fungus. Microbiology 147, 2943–2950. doi: 10.1099/00221287-147-11-2943
Tahir, H. A., Gu, Q., Wu, H., Niu, Y., Huo, R., and Gao, X. (2017a). Bacillus volatiles adversely affect the physiology and ultra-structure of Ralstonia solanacearum and induce systemic resistance in tobacco against bacterial wilt. Sci. Rep. 7:40481. doi: 10.1038/srep40481
Tahir, H. A., Gu, Q., Wu, H., Raza, W., Hanif, A., Wu, L., et al. (2017b). Plant growth promotion by volatile organic compounds produced by Bacillus subtilis SYST2. Front. Microbiol. 8:171. doi: 10.3389/fmicb.2017.00171
Takamatsu, S., Lin, X., Nara, A., Komatsu, M., Cane, D. E., and Ikeda, H. (2011). Characterization of a silent sesquiterpenoid biosynthetic pathway in Streptomyces avermitilis controlling epi-isozizaene albaflavenone biosynthesis and isolation of a new oxidized epi-isozizaene metabolite. Microb. Biotechnol. 4, 184–191. doi: 10.1111/j.1751-7915.2010.00209.x
Tissier, A., Morgan, J. A., and Dudareva, N. (2017). Plant volatiles: going ‘in’ but not ‘out’ of trichome cavities. Trends Plant Sci. 22, 930–938. doi: 10.1016/j.tplants.2017.09.001
Tyc, O., de Jager, V. C. L., van den Berg, M., Gerards, S., Janssens, T. K. S., Zaagman, N., et al. (2017a). Exploring bacterial interspecific interactions for discovery of novel antimicrobial compounds. Microb. Biotechnol. 10, 910–925. doi: 10.1111/1751-7915.12735
Tyc, O., Song, C. X., Dickschat, J. S., Vos, M., and Garbeva, P. (2017b). The ecological role of volatile and soluble secondary metabolites produced by soil bacteria. Trends Microbiol. 25, 280–292. doi: 10.1016/j.tim.2016.12.002
Tyc, O., Wolf, A. B., and Garbeva, P. (2015). The effect of phylogenetically different bacteria on the fitness of Pseudomonas fluorescens in sand microcosms. PLOS ONE 10:e0119838. doi: 10.1371/journal.pone.0119838
Van Agtmaal, M., Van Os, G., Hol, G., Hundscheid, M., Runia, W., Hordijk, C., et al. (2015). Legacy effects of anaerobic soil disinfestation on soil bacterial community composition and production of pathogen-suppressing volatiles. Front. Microbiol. 6:701. doi: 10.3389/fmicb.2015.00701
Van Dam, N. M., Weinhold, A., and Garbeva, P. (2016). “Calling in the dark: the role of volatiles for communication in the rhizosphere,” in Deciphering Chemical Language of Plant Communication, eds J. D. Blande and R. Glinwood (Cham: Springer International Publishing), 175–210.
Velivelli, S. L. S., Kromann, P., Lojan, P., Rojas, M., Franco, J., Suarez, J. P., et al. (2015). Identification of mVOCs from Andean rhizobacteria and field evaluation of bacterial and mycorrhizal inoculants on growth of potato in its center of origin. Microb. Ecol. 69, 652–667. doi: 10.1007/s00248-014-0514-2
Vespermann, A., Kai, M., and Piechulla, B. (2007). Rhizobacterial volatiles affect the growth of fungi and Arabidopsis thaliana. Appl. Environ. Microbiol. 73, 5639–5641. doi: 10.1128/aem.01078-07
Von Reuß, S. H., Kai, M., Piechulla, B., and Francke, W. (2010). Octamethylbicyclo[3.2.1]octadienes from the rhizobacterium Serratia odorifera. Angew. Chem. Int. Ed. 49, 2009–2010. doi: 10.1002/anie.200905680
Wenke, K., Kai, M., and Piechulla, B. (2010). Belowground volatiles facilitate interactions between plant roots and soil organisms. Planta 231, 499–506. doi: 10.1007/s00425-009-1076-2
Wenke, K., Wanke, D., Kilian, J., Berendzen, K., Harter, K., and Piechulla, B. (2012). Volatiles of two growth-inhibiting rhizobacteria commonly engage AtWRKY18 function. Plant J. 70, 445–459. doi: 10.1111/j.1365-313X.2011.04891.x
Werner, S., Polle, A., and Brinkmann, N. (2016). Belowground communication: impacts of volatile organic compounds (VOCs) from soil fungi on other soil-inhabiting organisms. Appl. Microbiol. Biotechnol. 100, 8651–8665. doi: 10.1007/s00253-016-7792-1
Westhoff, S., van Wezel, G. P., and Rozen, D. E. (2017). Distance-dependent danger responses in bacteria. Curr. Opin. Microbiol. 36, 95–101. doi: 10.1016/j.mib.2017.02.002
Wheatley, R. E. (2002). The consequences of volatile organic compound mediated bacterial and fungal interactions. Antonie Van Leeuwenhoek 81, 357–364. doi: 10.1023/A:1020592802234
Widhalm, J. R., Jaini, R., Morgan, J. A., and Dudareva, N. (2015). Rethinking how volatiles are released from plant cells. Trends Plant Sci. 20, 545–550. doi: 10.1016/j.tplants.2015.06.009
Wintermans, P. C. A., Bakker, P. A. H. M., and Pieterse, C. M. J. (2016). Natural genetic variation in Arabidopsis for responsiveness to plant growth-promoting rhizobacteria. Plant Mol. Biol. 90, 623–634. doi: 10.1007/s11103-016-0442-2
Xie, S., Zang, H., Wu, H., ddin Rajer, F. U., and Gao, X. (2016). Antibacterial effects of volatiles produced by Bacillus strain D13 against Xanthomonas oryzae pv. oryzae. Mol. Plant Pathol. doi: 10.1111/mpp.12494 [Epub ahead of print].
Yamada, Y., Cane, D. E., and Ikeda, H. (2012). Diversity and analysis of bacterial terpene synthases. Methods Enzymol. 515, 123–162. doi: 10.1016/B978-0-12-394290-6.00007-0
Yamada, Y., Kuzuyama, T., Komatsu, M., Shin-Ya, K., Omura, S., Cane, D. E., et al. (2015). Terpene synthases are widely distributed in bacteria. Proc. Natl. Acad. Sci. U.S.A. 112, 857–862. doi: 10.1073/pnas.1422108112
Yim, G., Huimi Wang, H., and Davies, J. (2006). The truth about antibiotics. Int. J. Med. Microbiol. 296, 163–170. doi: 10.1016/j.ijmm.2006.01.039
Yuan, J., Zhao, M., Li, R., Huang, Q., Raza, W., Rensing, C., et al. (2017). Microbial volatile compounds alter the soil microbial community. Environ. Sci. Pollut. Res. 24, 22485–22493. doi: 10.1007/s11356-017-9839-y
Keywords: volatile organic compounds, microbial interactions, bacteria, fungi, protists, plant–microbe interactions
Citation: Schulz-Bohm K, Martín-Sánchez L and Garbeva P (2017) Microbial Volatiles: Small Molecules with an Important Role in Intra- and Inter-Kingdom Interactions. Front. Microbiol. 8:2484. doi: 10.3389/fmicb.2017.02484
Received: 26 June 2017; Accepted: 29 November 2017;
Published: 12 December 2017.
Edited by:
Etienne Yergeau, Institut National de la Recherche Scientifique (INRS), CanadaReviewed by:
Stéphane Hacquard, Max Planck Institute for Plant Breeding Research (MPG), GermanyBirgit Piechulla, University of Rostock, Germany
Copyright © 2017 Schulz-Bohm, Martín-Sánchez and Garbeva. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Paolina Garbeva, cC5nYXJiZXZhQG5pb28ua25hdy5ubA==
†These authors have contributed equally to this work.