
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Microbiol. , 22 August 2017
Sec. Infectious Agents and Disease
Volume 8 - 2017 | https://doi.org/10.3389/fmicb.2017.01554
This article is part of the Research Topic Zika Virus Research View all 25 articles
 Juan-Carlos Saiz1
Juan-Carlos Saiz1 Miguel A. Martín-Acebes1
Miguel A. Martín-Acebes1 Rubén Bueno-Marí2
Rubén Bueno-Marí2 Oscar D. Salomón3
Oscar D. Salomón3 Luis C. Villamil-Jiménez4
Luis C. Villamil-Jiménez4 Jorg Heukelbach5,6
Jorg Heukelbach5,6 Carlos H. Alencar5
Carlos H. Alencar5 Paul K. Armstrong7
Paul K. Armstrong7 Tania M. Ortiga-Carvalho8
Tania M. Ortiga-Carvalho8 Rosalia Mendez-Otero8
Rosalia Mendez-Otero8 Paulo H. Rosado-de-Castro9,10
Paulo H. Rosado-de-Castro9,10 Pedro M. Pimentel-Coelho8*
Pedro M. Pimentel-Coelho8*Zika is a viral disease transmitted mainly by mosquitoes of the genus Aedes. In recent years, it has expanded geographically, changing from an endemic mosquito-borne disease across equatorial Asia and Africa, to an epidemic disease causing large outbreaks in several areas of the world. With the recent Zika virus (ZIKV) outbreaks in the Americas, the disease has become a focus of attention of public health agencies and of the international research community, especially due to an association with neurological disorders in adults and to the severe neurological and ophthalmological abnormalities found in fetuses and newborns of mothers exposed to ZIKV during pregnancy. A large number of studies have been published in the last 3 years, revealing the structure of the virus, how it is transmitted and how it affects human cells. Many different animal models have been developed, which recapitulate several features of ZIKV disease and its neurological consequences. Moreover, several vaccine candidates are now in active preclinical development, and three of them have already entered phase I clinical trials. Likewise, many different compounds targeting viral and cellular components are being tested in in vitro and in experimental animal models. This review aims to discuss the current state of this rapidly growing literature from a multidisciplinary perspective, as well as to present an overview of the public health response to Zika and of the perspectives for the prevention and treatment of this disease.
Since the beginning of the 21st century, a number of infectious disease threats have emerged that are deemed to be such a risk as to demand a global response. Most have been respiratory viral diseases – severe acute respiratory syndrome (2003), avian influenza in humans (2005), A(H1N1) pandemic influenza (2009), and Middle East respiratory syndrome coronavirus (MERS-CoV) (2012 onward). Ebola virus disease, transmitted to others by direct contact, bucked that trend when a large outbreak in several West African countries claimed over 11,000 lives from 2014 to 2015. Very few would have predicted the most recent infectious disease emergency would involve a vector-borne virus, Zika virus (ZIKV), causing congenital malformations and other neurological disorders.
ZIKV is a flavivirus (Flaviviridae family) transmitted by mosquitoes. The virus has been isolated from many mosquito species, although it seems that the vectors of the natural transmission cycle are mosquitoes of the genus Aedes (Diagne et al., 2015). As any other flavivirus, the viral genome is composed of a single-stranded RNA molecule of positive polarity about 10 kb in length that encodes a single open reading frame (ORF) flanked by two untranslated regions at both ends (Kuno and Chang, 2007).
ZIKV was first isolated in 1947 from the serum of a febrile sentinel monkey in the Zika Forest, hence its name, and 1 year later from Aedes africanus mosquitoes caught in the same forest (Dick et al., 1952). Since then, it was confined to Africa until it first detection in Asia in the 1980s. Subsequently, human outbreaks were reported in 2007 in the Micronesia and in 2013 in the French Polynesia (Saiz et al., 2016). However, ZIKV was an almost neglected pathogen until its recent introduction into the Americas.
It is not the direct effect that ZIKV has on those infected that is the main concern, as the vast majority will be either asymptomatic or else experience a relatively mild illness and an uneventful recovery. Rather, it is the sequelae of infection– Guillain–Barré syndrome (GBS) and microcephaly and other congenital malformations – that cause the morbidity and mortality associated with the infection. As a result, the World Health Organization (WHO) declared a public health emergency of international concern (PHEIC) (WHO, 2016e), elements of which were later integrated into risk assessments by the European Centre for Disease Prevention and Control (ECDC, 2016).
This review discusses several aspects of the biology, epidemiology, transmission and health consequences of ZIKV infection, including findings from in vitro and in vivo models. Disease control measures, such as vaccine development and the public health response to ZIKV outbreaks, are also reviewed.
The emergence of new pathogens has been the reality and a prominent feature of the 21st century. It constitutes a global challenge to public health, especially in developing countries. Arboviruses such as Dengue virus (DENV), Chikungunya and ZIKV are paradigmatic examples of such a statement.
ZIKV virus is a flavivirus first discovered in 1947 in the Zika forest of Uganda, in a captive sentinel rhesus monkey during a yellow fever (YF) surveillance disease activity (Dick, 1953). In 1952, the presence of human cases was demonstrated by a mouse protection test in the sera of indigenous residents of Uganda and Tanganika (Smithburn, 1952). During 1958, the isolation of two strains of the virus were made in mosquitoes (Aedes africanus) in Lungo forest (Weinbren and Williams, 1958). The virus was also detected during the decades of 1960–1980 in sentinel Rhesus monkeys and on mosquitoes (mainly the genus Aedes) in other countries across equatorial Africa. Sporadic human cases of a mild disease were reported (Saluzzo et al., 1981).
A number of serological studies in the 1950s provided some evidence that ZIKV was widespread throughout Asia (Wikan and Smith, 2016). The presence of ZIKV in Asia was confirmed in 1969 when the virus was isolated from an Aedes aegypti mosquito in Malaysia (Marchette et al., 1969).
In April 2007, ZIKV spread its usual geographic range and was detected outside Africa and Asia for the first time when an outbreak occurred on Yap Island in the South Western Pacific Ocean, as an emerging pathogen (Hayes, 2009). Sera from acutely ill patients were sent to the Centers for Disease Control and Prevention (CDC) Arbovirus Diagnostic and Reference Laboratory in Fort Collins, Colorado, where 10 of 71 samples (14%) were found positive for the virus, as they contained ZIKV RNA according to reverse-transcriptase-polymerase-chain-reaction (RT-PCR) assay (Hayes, 2009). It has been found that the attack rates of ZIKV infection were higher among females than males and among older persons than younger persons. In contrast, the prevalence of IgM antibody against ZIKV was higher in male participants (possibly due to their greater exposure to mosquitoes) and was relatively equally distributed across age groups (Hayes, 2009).
Although wind-blown mosquitoes can travel distances of several hundred kilometers over the open ocean, introduction of the virus by travel or trade involving an infected person or an infected mosquito is considered the most likely source of the Yap Island outbreak, especially because no monkeys were present on the island during the outbreak (Hayes, 2009; Kindhauser et al., 2016).
The fact that ZIKV caused such a widespread outbreak on Yap Island, numbering more than 100 confirmed and probable cases, is striking. The absence of any evidence that viral mutation could explain changes in the epidemiological behavior of the virus has led to several other explanations being postulated, including a lack of population immunity; that means that regular exposure to infection by populations in Africa and Asia may have prevented large outbreaks in those regions, such as those seen on the Pacific Islands and in the Americas (Kindhauser et al., 2016).
Another possible reason for this change may lay on the probable under-reporting, which could explain missing reports of previous outbreaks, especially due to the clinical similarities of mild illnesses associated with ZIKV, DENV, and Chikungunya infections, and the frequent co-circulation of all three viruses (Paixao et al., 2016).
Since 2008, the availability of information about the virus has increased, including data on epidemiology, causal associations, and possible sexual transmission. Clinical and serologic evidence suggested that a US-American scientist contracted ZIKV infections while working in Senegal in 2008, and then transmitted this arbovirus to his wife after his return home. Direct contact was implicated as the transmission route, most likely as a sexually transmitted infection (Foy et al., 2011). Additional evidence was found during a ZIKV outbreak in French Polynesia (2013): the virus was isolated from the semen of a patient in Tahiti that sought treatment for hematospermia (Musso et al., 2015).
Retrospective surveys identified an unusual and heterogeneous cluster of congenital brain malformations and brainstem dysfunction in fetuses and newborns over a limited period following a ZIKV epidemic in French Polynesia (Besnard et al., 2016), prompting the conclusion of causal association (Araujo et al., 2016).
In 2016, ZIKV spread rapidly throughout the Americas after its initial appearance in Brazil in May 2015. In 2016, 48 countries and territories in the Americas had reported more than 532,000 suspected cases of Zika, including 175,063 confirmed cases. In addition, 22 countries and territories reported 2,439 cases of a congenital syndrome associated with Zika. Five countries had reported sexually transmitted Zika cases (The Pan American Health Organization [PAHO]/World Health Organization [WHO], 2015; Ikejezie et al., 2017).
In Brazil, the most heavily affected country, a very rapid dispersion of ZIKV was identified, mainly in the Northeast region. This area has the lowest vaccine coverage for YF and notified 65.7% of all cases (De Goes Cavalcanti et al., 2016). Only from March to September 2016, which is considered the highest transmission period in Brazil, as evidenced by a large number of dengue cases observed over years, there was an incidence rate of ZIKV infections of 69.22 cases/100,000 newborns, a mortality coefficient of 5.37 deaths/100,000 newborns and a case fatality rate of 7,750 deaths/100,000 cases. The highest incidence and mortality were found in the Northeast region with 201 cases/100,000 newborns and 4 deaths/100,000 newborns, respectively (SVS/MS, 2017).
The rapid spread of ZIKV disease in the Americas was aided by the lack of effective control of vectors of other arboviruses like DENV and Chikungunya, in several countries of Latin America (Rodriguez-Morales, 2015). For decades, vector control programs have failed, due to the adaptation capacity and biological efficiency of the vectors (Al-Abdely, 2016). Additional challenges for population-based surveillance of ZIKV disease include remote locations, porous borders with neighboring countries, and underreporting by health care providers, all of which can limit the ability to ascertain all cases of ZIKV disease. Underreporting of cases would be particularly marked if they occur in areas that are not considered to be at risk for ZIKV disease, such as those at elevations of more than 2,000 m above sea level (Pacheco et al., 2016).
To summarize, the global emergence (Figure 1) of certain arboviruses, such as ZIKV, that in the past were considered to be restricted to specific geographical areas and to cause a mild disease of sporadic behavior, indicates the importance of such diseases to epidemiology and public health, from the perspective of adaptation potential, certain and probable vectors, surveillance and control, and also communication, interdisciplinary investigation and intersectoral cooperation. In other words, “new threats from infectious diseases may emerge from unexpected places, and we need strategies in place that we can roll out to rapidly gain an understanding of the transmission, pathogenesis, and control of previously little-known pathogens to protect global public health” (Lessler et al., 2016).
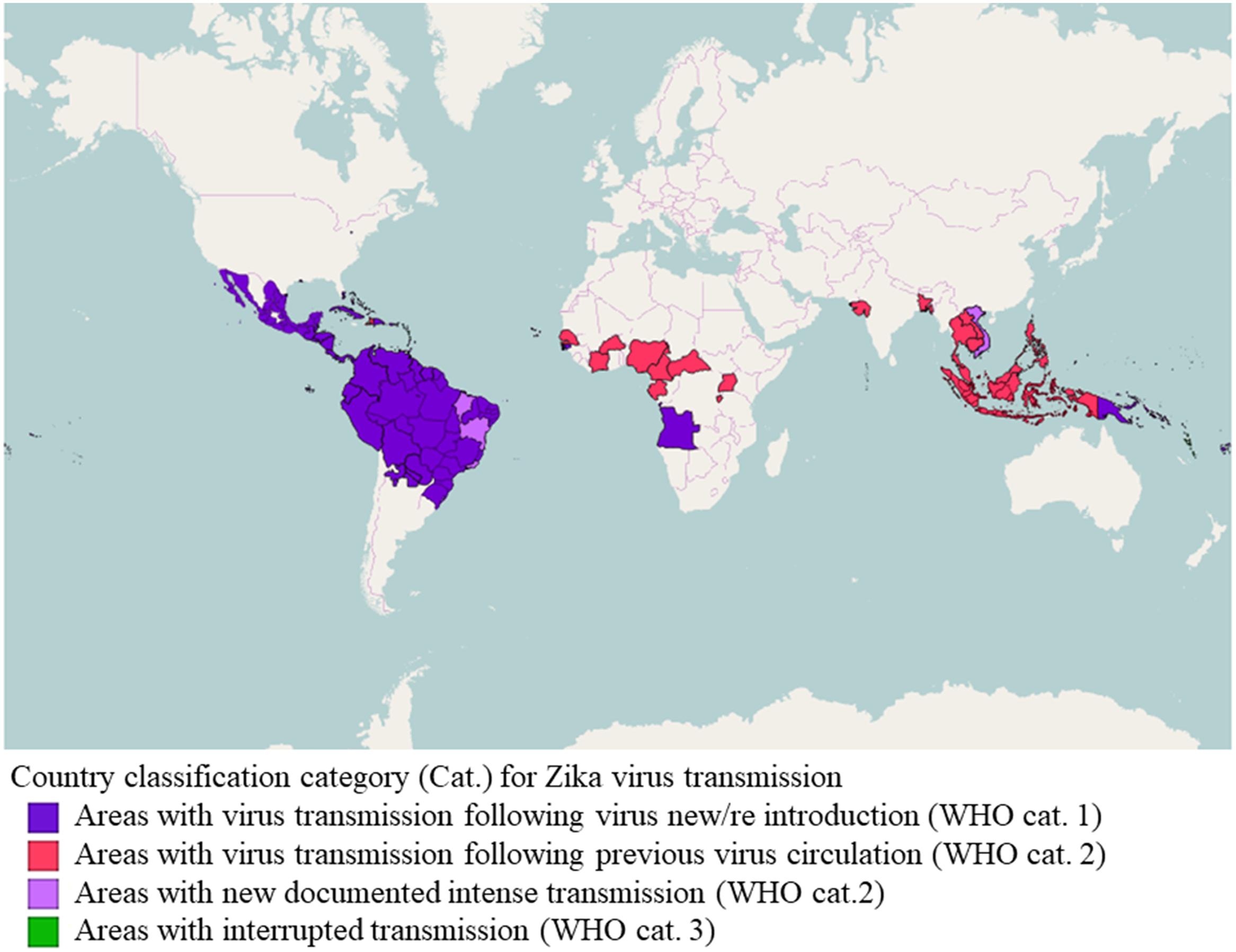
FIGURE 1. Classification of countries and territories regarding vector-borne Zika virus transmission. Map of the current Zika virus transmission based on the European Center for Disease Prevention and Control (ECDC) adaptation of the World Health Organization (WHO)’s Zika virus country classification scheme (https://ecdc.europa.eu/en/publications-data/current-zika-virus-transmission-list-countries-ecdc-adaptation-whos-zika-virus - accessed 16 July 2017). The map was generated using GADM database of Global Administrative Areas shapefiles (http://www.gadm.org/) and Openlayers plugin within QGIS 2.18.9 (Development Team, 2017, available at http://www.qgis.org/en/site).
Maybe it is time to reconsider research horizons, and to pay attention to the integral circle of host, agent, environment and vector. Health promotion approaches are still valid, and can be an alternative for the global reality of health inequities and weakening of public services (Caprara and Ridde, 2016).
Historically, ZIKV has been classified into the Spondweni serogroup, genus Flavivirus (Flaviviridae), which includes two species: ZIKV and Spondweni virus (Casals, 1957). Further molecular classifications confirmed these relationships between both species (Kuno et al., 1998; Haddow et al., 2012). Nowadays, ZIKV isolates can be grouped into two or three major lineages (Figure 2). These lineages correspond to the African lineage, the Asian lineage (that includes the American strains) and a neglected lineage circulating in Africa (designated African II) that would constitute a sister group to both African (which should be renamed to African I) and Asian lineages previously identified (Faye et al., 2014; Gong et al., 2016; Wang L. et al., 2016; Li Y. et al., 2017). All these phylogenetic analyses indicate that ZIKV originated from Africa and then spread to Asia, Pacific islands, and throughout the Americas. The introduction of ZIKV into the Americas most probably occurred by a single introduction of an Asian strain of ZIKV between May and December 2013 (Faria et al., 2016). Remarkably, despite the genetic differences between ZIKV strains, the antigenic relationships between strains support the existence of a single viral serotype which may be of crucial importance for the design of ZIKV vaccines (Dowd et al., 2016a).

FIGURE 2. Phylogram of Zika virus (ZIKV). The phylogenetic tree was based on the sequence of NS5. Multiple alignment was performed using MUSCLE (Edgar, 2004) and the tree was constructed by the Maximum Likelihood method using PhyML (Guindon et al., 2010) and Phylogeny.fr (Dereeper et al., 2008). Spondweni virus was included as the outgroup control.
The viral genome is composed of a single-stranded RNA molecule of positive polarity about 10 kb in length (Kuno and Chang, 2007). In a similar manner to cellular mRNAs, ZIKV genome includes a cap structure at its 5′ end, but in contrast to cellular mRNAs, ZIKV genome lacks a 3′ poly(A) tract and ends with CUOH (Figure 3A). The genome contains a single ORF flanked by two untranslated regions located at the 5′ and 3′ ends of the genome (Kuno and Chang, 2007). ZIKV genome also contains three conserved sequences (CS 1 to CS3) that may mediate genome cyclization between 5′ and 3′ terminal regions of the genome. Notably, the organization of the CS in the 3′ end of ZIKV is different from that of other mosquito-borne flaviviruses (Kuno and Chang, 2007). Besides genomic RNA, it has been described that due to the presence of a multi-pseudoknot structure in the genomic RNA that confounds a cellular exonuclease, ZIKV-infection also produces subgenomic flaviviral RNAs (sfRNAs) within infected cells that play relevant roles in innate immunity evasion and viral pathogenesis (Akiyama et al., 2016; Donald et al., 2016).
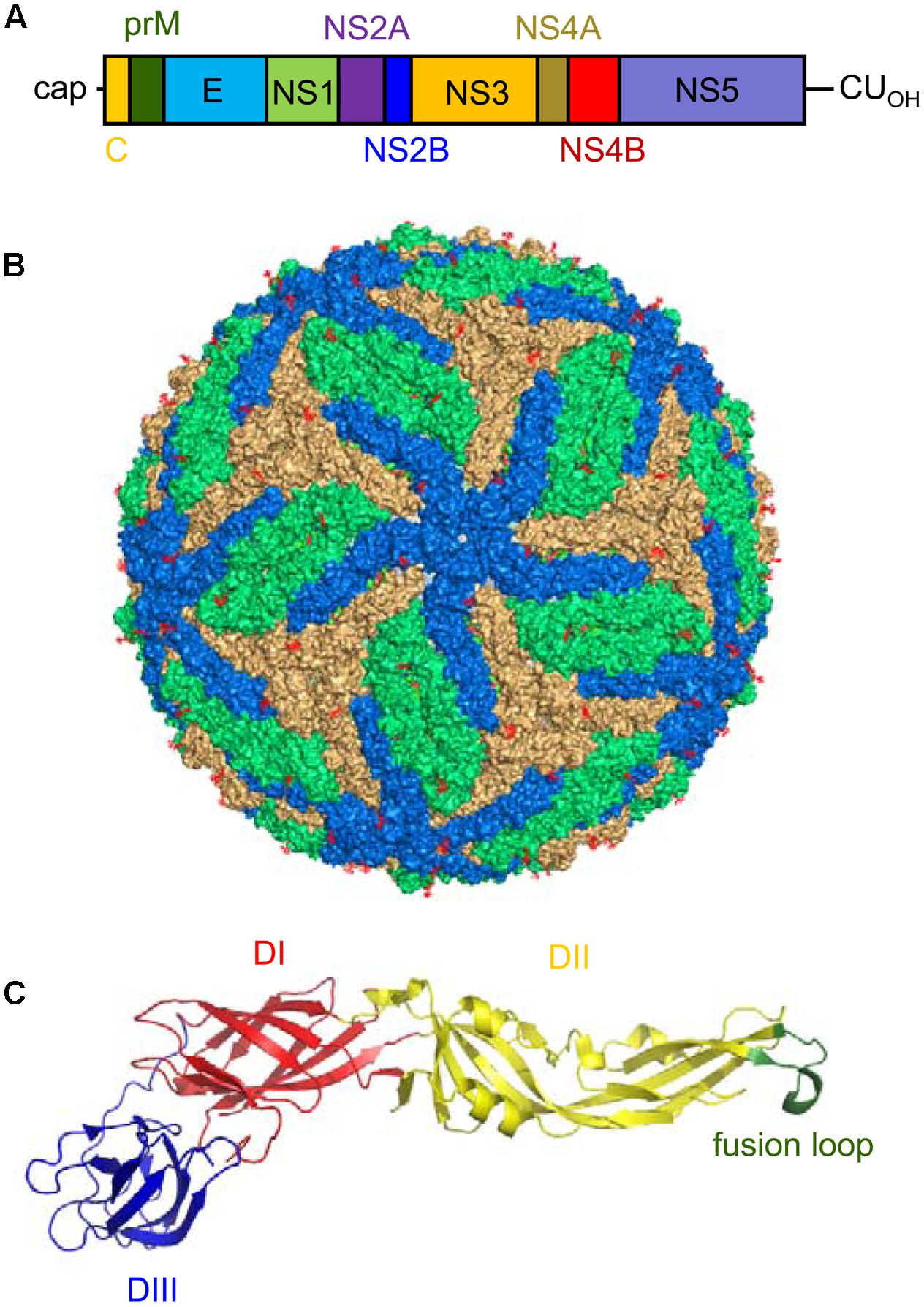
FIGURE 3. Genomic organization, virion structure, and organization of the E glycoprotein of Zika virus (ZIKV). (A) Schematic view of the genomic organization of ZIKV. The single open reading frame (ORF; boxes) that encodes both structural (C-prM/M and E) and non-structural (NS) proteins (NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5) is flanked by two untranslated regions (UTRs). Notice the presence of a 5′ cap and the lack of a poly(A) tail at the 3′ end of the genome. (B) Surface representation of a ZIKV mature particle. The E monomers are colored in blue, orange and green to facilitate the interpretation of their distribution. Image was produced using the cryo-electron microscopy data available (Protein Data Bank entry 5IRE). (C) Structure of a monomer of the soluble ectodomain of E glycoprotein of ZIKV. The ribbon diagram was based on the atomic coordinates solved by X-ray crystallography (Protein Data Bank entry 5JHM). DI in red, DII in yellow, and DIII in blue. Fusion loop is highlighted in green.
The polyprotein encoded by the single ORF in ZIKV is cleaved by cellular and viral proteases into 10 mature proteins (three structural and seven non-structural proteins) (Figure 3A). The sequence of the cleavage sites of ZIKV follows the patterns established for other mosquito-borne viruses (Kuno and Chang, 2007). The three structural proteins [the capsid (C), premembrane/membrane (prM/M), and envelope (E) proteins] participate in the assembly of the virions. The C protein conform the core of the virions together with the RNA. The E protein should mediate the binding to the cellular receptor of the virus, promotes the fusion of the virions with the target membranes, and constitutes the main target for the induction of antibodies (Dai et al., 2016; Stettler et al., 2016; Wang Q. et al., 2016; Zhang C. et al., 2016). The E protein is N-glycosylated at Asn 154 in most ZIKV strains. This glycosylation is important for particle protein expression and secretion, viral packaging, and infectivity (Mossenta et al., 2017). The cleavage of prM into M protein promotes the maturation of the virions from “spiky” immature particles to “smooth” mature particles (Prasad et al., 2017). In a similar way to that described for other flaviviruses (Martin-Acebes and Saiz, 2012), the seven non-structural (NS) proteins (NS1, NS2A, NS2B, NS3, NS4A, NS4B, and NS5) should account for a variety of functions that go from replication of viral RNA, morphogenesis of viral particles, induction of membrane rearrangements, and viral factory development to modulation of host immune response. The NS1 protein, whose structure has been resolved (Song et al., 2016; Xu X. et al., 2016), participates in flaviviral replication and exhibits immunomodulatory activities. Since NS1 protein is secreted from infected cells, it largely induces antibodies within infected hosts that can be suitable for diagnostics purposes (Dai et al., 2016; Steinhagen et al., 2016). Regarding NS2A, to our knowledge, there are no specific studies addressing the function of this protein in ZIKV. On the other hand, NS3 is a trypsin-like serine protease involved in polyprotein processing (Gruba et al., 2016; Lei et al., 2016) that also exhibits helicase activity, which plays a pivotal role in viral RNA replication enabling RNA unwinding (Tian et al., 2016). Apart from its involvement in viral polyprotein processing, NS3 can cleave cellular factors such as FAM134B, hence modulating the autophagic response within ZIKV-infected cells (Lennemann and Coyne, 2017). NS2B acts as a cofactor necessary for the activity of this protein, and the crystal structure of NS2B-NS3 complex has been resolved under different circumstances (Lei et al., 2016; Phoo et al., 2016; Zhang C. et al., 2016). Due to the relevance of NS2B-NS3 function in the ZIKV life cycle, the search for inhibitors of the enzymatic activity of this complex is at the front line of antiviral discovery against ZIKV (Cao et al., 2016; Sahoo et al., 2016; Lee et al., 2017; Rut et al., 2017). NS4A and NS4B deregulate Akt-mTOR signaling to inhibit neurogenesis and induce autophagy (Liang et al., 2016). In addition, the expression of NS4A has been also related to activation of the cellular stress pathway involving Tor1 and type 2A phosphatase activator Tip41 (Li G. et al., 2017). NS5 is the viral RNA-dependent RNA polymerase that is in charge of genome replication constituting a major target for antiviral design (Lu et al., 2017; Xu et al., 2017). Furthermore, the analysis of the structure of the methyltransferase domain of NS5, which is responsible for capping the 5′ end of the viral genomic RNA, also provides new opportunities for the design of antiviral compounds (Coloma et al., 2016; Stephen et al., 2016; Zhang C. et al., 2016; Coutard et al., 2017; Zhou et al., 2017). Besides its function in genome replication and capping, NS5 from ZIKV also contributes to viral multiplication by inhibiting interferon signaling (Grant et al., 2016). Although a great effort has been performed to decipher the structure and function of some of the ZIKV NS proteins, many issues still remain to be analyzed, as the more detailed knowledge of their function would probably provide valuable information about the pathogenesis of ZIKV, and would also greatly contribute to the development of antiviral strategies against this pathogen.
Early filtration studies suggested that the size of ZIKV particles was about 30 to 45 nm in diameter (Dick, 1952). Further transmission electron microscopy showed that the virions were spherical particles with an overall diameter of 40 to 43 nm displaying a central electron dense core being 28 to 30 nm in diameter (Hamel et al., 2015). Nowadays, cryo-electron microscopy reconstructions of mature virions have provided a detailed view of the structure and organization of mature ZIKV particles (Kostyuchenko et al., 2016; Sirohi et al., 2016). The internal core of the particle is composed of the genomic RNA molecule complexed with multiple copies of the capsid (C) protein, enclosed within a lipid membrane derived from the endoplasmic reticulum of the host cell. The E and M proteins (180 copies of each protein) are anchored to this lipid membrane via their transmembrane regions. Since the M protein is a small protein hidden under the E protein layer, the outer layer of the viral particles consists of an icosahedral protein shell basically composed of the E protein (Figure 3B). This outer shell exhibits the characteristic herringbone structure in the virion similar to that of other flaviviruses. The E proteins are arranged as 90 anti-parallel homodimers, with three dimers lying parallel to each other forming a raft. The ectodomain of the E protein that protrudes from the lipid bilayer is organized in three different domains: DI, DII, and DIII (Figure 3C). DI acts as a bridge between DII and DIII. The tip of DII contains the hydrophobic fusion loop that interacts with cellular membranes for viral fusion (Kostyuchenko et al., 2016; Sirohi et al., 2016). DIII is the target for most neutralizing antibodies, whereas antibodies against DI/DII are poorly neutralizing (Stettler et al., 2016; Zhao et al., 2016). As mentioned above, the E protein is glycosylated at Asn 154 in most ZIKV strains, and this glycan protrudes from the surface of the particle. Remarkably, the region surrounding the glycosylation site structurally greatly differs from other related flaviviruses (Kostyuchenko et al., 2016; Sirohi et al., 2016). Also, in contrast to that described for DENV, ZIKV particles are structurally stable even when incubated at 40°C (Kostyuchenko et al., 2016). Although it has been proposed that this thermal stability may have implications for virus survival in body fluids such as saliva or semen (Kostyuchenko et al., 2016), other studies discard that the unique pathobiology of ZIKV may be only the cause of its thermal stability (Goo et al., 2016).
ZIKV was isolated for the first time from a sentinel allocthonous monkey in Uganda in 1948; just 1 year later the sylvatic Aedes africanus was found infected in the same site of the Zika Forest, and again in 1958, 1964, and 1969, caught both from the upper canopy and from ground level. Since then, during the period when the records are still mainly restricted to Africa many Aedes species reported harboring ZIKV: Ae. vitatus, Ae. hirsutus, Ae. unilineatus, Ae. metallicus. Ae. apicoergenteus, Ae. opok, Ae. dalzieli, Ae. luteocephalus, Ae. tayliri (Haddow et al., 1964; Hayes, 2009; Faye et al., 2013; Diallo et al., 2014; Vorou, 2016). Out of Africa, Ae. hensillii was incriminated in the Yap Island as the prevalent species during the outbreak and it was demonstrated experimentally capable of transmission, while in French Polynesia Ae. polynesium probably contributes to the transmission (Duffy et al., 2009; Ledermann et al., 2014; Richard et al., 2016).
The scenario of potential worldwide spread of ZIKV changed radically when Ae. aegypti and Ae. albopictus were identified as vectors during the outbreak in Brazil (Ferreira-de-Brito et al., 2016). Actually, Ae agypti had already been demonstrated competent for ZIKV in 1956 and isolated from Malaysia in 1969. In addition, it was incriminated in the French Polynesia and Indonesia outbreaks and retrospectively in Africa (Marchette et al., 1969; Richard et al., 2016). Ae. albopictus was associated with the outbreak in Gabon in 2007 as this Asian mosquito successfully replace the native Ae. aegypti in many areas of Africa (Grard et al., 2014), and it was also shown to be a potential vector in Singapore (Wong et al., 2013). These ubiquitous two mosquito species, with their dynamic adaptation to urban environments, capacity to breed in cryptic containers, to survive to adverse seasons or to be dispersed passively by humans (adults, desiccated eggs-Ae aegypti), to tolerate temperate climates and even keep sylvatic niches (Ae. albopictus), together with the increase in the last decades of social trends as those of urbanization, traveling (speed and number of people involved), migration and climate extraordinary events, generates the current epidemic risk momentum.
Further, species that belong to genera other than Aedes as Culex perfuscus, Anopheles coustani, An. gambiae s.l., Mansonia uniformis were found infected with ZIKV in Africa (Vorou, 2016), but the isolations only prove that these mosquitoes recently fed on a viremic vertebrate. On the other hand, the vector competence is the innate capability to acquire and sustain the pathogen, and transmit it to a host, so many experiments on competence were performed in alternative vectors, mainly with Cx pipiens and Cx quinquefasciatus. The results showed arguments that support (Franca et al., 2016; Guedes et al., 2016; Guo et al., 2016) or reject (Aliota et al., 2016b; Amraoui et al., 2016; Boccolini et al., 2016; Fernandes et al., 2016; Hall-Mendelin et al., 2016; Huang Y.J. et al., 2016; Weger-Lucarelli et al., 2016; Hart et al., 2017) Culex vector competence. To understand so many disparate results, the protocols should be analyzed and compared taking into account several issues that include the virus-vector origin (geographical coherence), and virus and vector past history in the laboratory (number of passages/generations) that made them genetically quite different to the complex wild circulating ones (Berthet et al., 2014; Bennett et al., 2016; Chouin-Carneiro et al., 2016; Wang L. et al., 2016).
However, even if the vector competence is assessed, it is rather different from vector capacity. The last concept attempts to explain the likelihood of effective human-vector contact and transmission, and depends not only on innate vector characteristics but also on local density, host range, and blood feeding behavior, biting rate, survival rate and colonization success (interspecific competence), as well as transovarial (Thangamani et al., 2016) and horizontal transmission. Therefore, even the presence of Ae aegypti does not confirm this species as the primary vector (Diagne et al., 2015; Di Luca et al., 2016; Weger-Lucarelli et al., 2016), and an undescribed sylvatic cycle in Asia should not be discarded (Althouse et al., 2016).
Focused in the main known urban vectors, Ae. aegypti and Ae albopictus, databases and maps of current or forecasted distribution were developed as a proxy of risk maps for ZIKV or Aedes-borne arbovirus (Kraemer et al., 2015a,b; Messina et al., 2016). The modeling of simulated risk is usually driven by temperature, precipitation, elevation, land cover and modulated by variables as seasonal or year-round abundance and density (population dynamics), vector biting and mortality rates, and extrinsic incubation period (Carlson et al., 2016; Escobar et al., 2016; Messina et al., 2016; Samy et al., 2016; Attaway et al., 2017). However, besides the accuracy and the assumptions of the model itself, some methodological matters require usually in-depth considerations from the mapping to assess the actual risk, as the space and time scale consistency between data and conclusions, accuracy and representativeness of field-collected data, and the particularities at smaller time or spatial scales (Jian et al., 2016; Misslin et al., 2016; Fischer et al., 2017; Li X. et al., 2017). Hence, the Latin America outbreak was explained by 2015–2016 ‘El Niño-Oscillation South’ 2015–2015 at continental level, but also at sub-regional level in Brazil it was explained by year to year variability (drought 2013–2015) and decadal variability followed by long-term trends as climate change (warm 2014–2015) (Munoz et al., 2016; Caminade et al., 2017). Furthermore, biological topics, as vector competence of local vectors (Gardner et al., 2016), vector competence between species (Camara et al., 2016), and the timing and location of vector or virus introduction (Robert et al., 2016; Walther et al., 2017) can change the probability and magnitude of transmission. Nevertheless, the anthropogenic factors usually are the main ones that trigger actual epidemics, even through climatic extreme events (Ahmed and Memish, 2017), and so some modeling in border areas includes also travel between borders and socioeconomic factors (Monaghan et al., 2016), while drivers of non-vectorial transmission still need better epidemiological elucidation (Guzzetta et al., 2016).
Since the first report of probable sexual transmission of ZIKV by Foy et al. (2011), many studies were published showing evidence of male-to-female, male-to-male and female-to-male sexual transmission by unprotected vaginal, oral or anal intercourse (Moreira et al., 2017). This hypothesis has been strengthened by numerous reports showing the long-term detection of ZIKV RNA and the isolation of infectious ZIKV from semen (Musso et al., 2015; Matheron et al., 2016; Nicastri et al., 2016; Moreira et al., 2017). ZIKV RNA has also been detected in female genital tract samples beyond viremia, albeit more transiently than in semen: it became undetectable around 3 weeks after symptom onset (Prisant et al., 2016, 2017; Murray et al., 2017). Moreover, Penot et al. (2017) have isolated infectious ZIKV from vaginal samples collected 3 days after the onset of symptoms from a woman with controlled HIV infection.
Remarkably, the vertical transmission of ZIKV (Besnard et al., 2014; Brasil et al., 2016; Calvet et al., 2016; Driggers et al., 2016; Oliveira Melo et al., 2016) has become a major public health challenge, as will be discussed below. It still needs to be demonstrated, however, whether ZIKV can be transmitted through breastfeeding. Infectious ZIKV particles have been isolated from the breast milk of 1 mother and ZIKV RNA was detected in the breast milk of 3 symptomatic mothers (Besnard et al., 2014; Dupont-Rouzeyrol et al., 2016). Other reported forms of non-vectorial transmission include non-sexual person-to-person contact (Swaminathan et al., 2016), and transmission by blood transfusion (Motta et al., 2016).
It has been estimated that the ZIKV infection may be symptomatic in 18–57% of cases, in which it causes a mild, self-limiting disease with an incubation period of up to 10 days (Duffy et al., 2009; Aubry et al., 2017). Symptomatic patients may develop fever and influenza-like symptoms relatively common in arboviral infections, such as rash, joint pain, conjunctivitis, headache and myalgia (Ahmad et al., 2016). These relatively mild symptoms last a few days and uncommonly result in hospitalization (Duffy et al., 2009). More recently, however, ZIKV infection has been associated with neurological and ophthalmological complications, including GBS in adults and microcephaly in fetus and newborns (Figure 4).
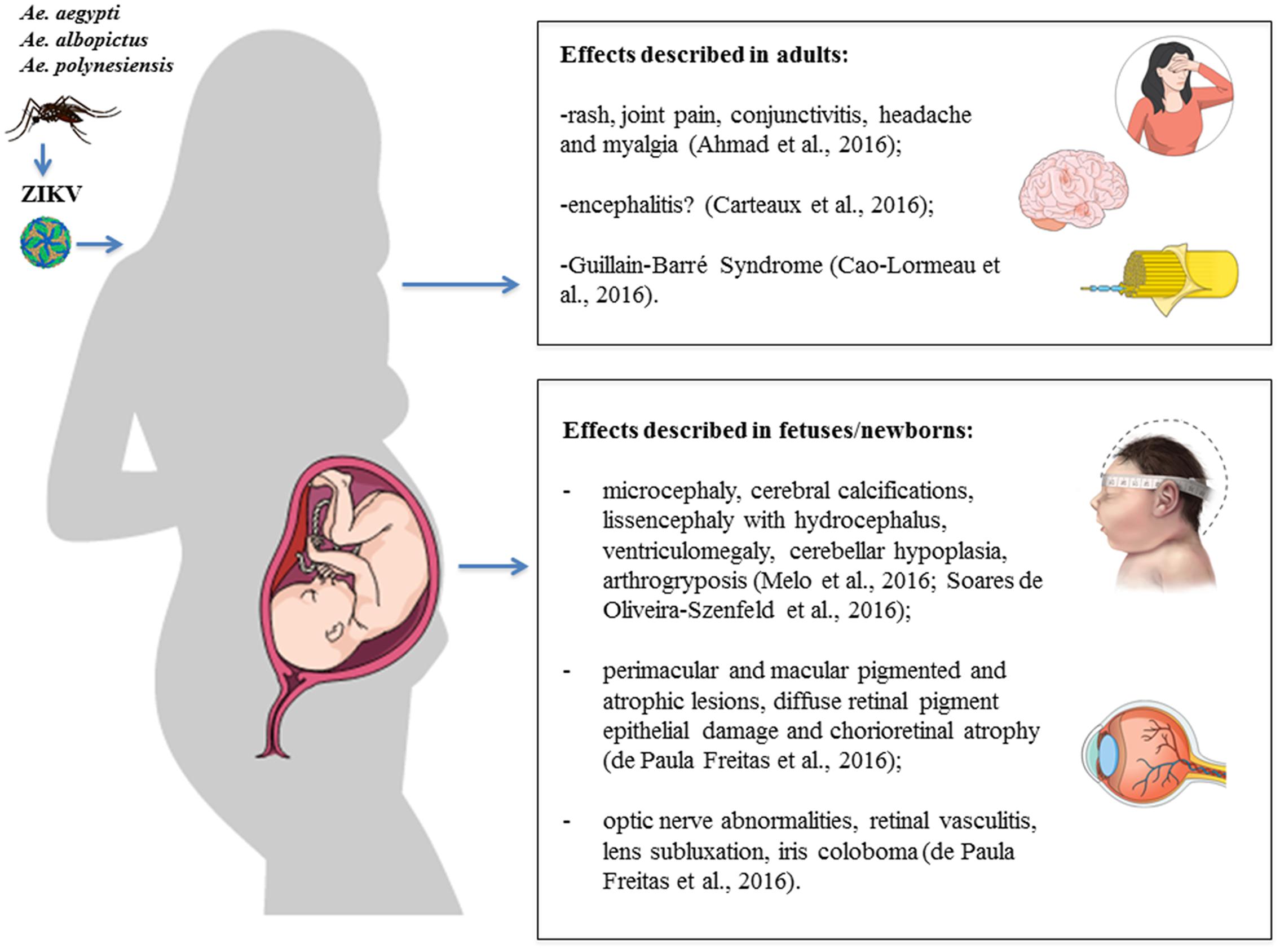
FIGURE 4. Clinical manifestations and consequences of Zika virus (ZIKV) infection. Schematic illustration of the effects described in adults and fetuses/newborns after ZIKV infection. Figure created in the Mind the Graph platform.
Two cases of encephalopathy syndromes, with seizures or electroencephalographic changes, were seen in Martinique as part of ZIKV infection, probably due to encephalitis (Roze et al., 2016). Carteaux et al. (2016) reported a case of an 81-year old man from France who was admitted to an Intensive Care Unit 10 days after a cruise to Pacific islands, with fever and decreased level of consciousness, being diagnosed with meningoencephalitis and had a positive RT-PCR for ZIKV in his cerebrospinal fluid. The patient later recovered and was discharged after 17 days, with cognitive function fully recovered after 38 days and residual weakness in his left arm. In Brazil, a 47-year old pregnant patient was admitted to an Intensive Care Unit with confusion, dysarthria and lower limb weakness, 4 days after presenting a rash and arthralgia (Soares et al., 2016). The patient was diagnosed with encephalitis and had a positive PCR for ZIKV in her urine and IgM ZIKV antibody in her cerebrospinal fluid and serum, and passed away after 11 days (Soares et al., 2016). Since these observations have only recently been described, very little is known about the frequency of a direct central nervous system (CNS) infection by ZIKV. A possible confounder is the simultaneous occurrence of various arboviruses, such as Chikungunya and DENV, since these arboviruses can cause a direct CNS invasion with myelitis, encephalitis, and meningitis (Moulin et al., 2016).
Throughout the ZIKV epidemic in French Polynesia, an increase in the number of patients presenting with GBS was seen (Paploski et al., 2016). Other arboviral infections such as dengue, chikungunya, Japanese encephalitis, and West Nile fever (WNF) have been associated with GBS (Ravi et al., 1994; Lebrun et al., 2009; Leis and Stokic, 2012; Verma et al., 2014). Cao-Lormeau et al. (2016) published a case-control study reporting the occurrence of 42 cases of GBS after a Zika outbreak between October 2013 and April 2014, in comparison to 5–10 cases in the same period in previous years. All 42 patients had neutralizing antibodies against ZIKV, whereas only 56% of neutralizing antibodies were found in the serum from a control group of 98 patients without GBS. The majority of GBS patients (93%) had detectable ZIKV IgM and 88% had a systemic febrile disease with symptoms that corresponded to ZIKV infection before the development of the neurological symptoms. This cohort of ZIKV associated GBS was classified electrophysiologically as acute motor axonal neuropathy (AMAN) with a rapid onset of disease (4 days to reach the plateau). Approximately one-third of patients required intensive medical therapy with mechanical ventilation (Cao-Lormeau et al., 2016). Anti-glycolipid antibodies were found in 31% of patients. However, anti-ganglioside antibodies typical of AMAN were rarely present (Cao-Lormeau et al., 2016). Dos Santos et al. (2016) performed an analysis of ministry of health websites and International Health Regulations channels to compare the occurrence of GBS before and after the Zika outbreak. They found an increase of 877% in Venezuela, 400% in Suriname, 211% in Colombia, 172% in the Brazilian state of Bahia, 150 % in the Dominican Republic, 144% in Honduras and 100% in El Salvador.
Zika infection has been confirmed in newborns with microcephaly, and an up to 20-fold increase in the number of microcephaly cases in the French Polynesia, Brazil and other Latin American countries (Brasil et al., 2016; Jouannic et al., 2016; Schuler-Faccini et al., 2016; Ventura et al., 2016a). After the French Polynesia epidemic between October 2013 and April 2014, 14 cases of fetuses and newborns with brain abnormalities (six of them without microcephaly) and five cases with brainstem dysfunction were observed, much higher numbers than those expected outside the epidemic period. Symptomatic cases were related to infections acquired during the first trimester, which is consistent with other congenital infections, such as rubella, cytomegalovirus, and toxoplasmosis (Besnard et al., 2016). However, some severe forms have been described in infections acquired after 20 weeks of gestation (Brasil et al., 2016). There is not always evidence of infection of mothers or fetuses by ZIKV but its presence in the amniotic fluid has been documented (Brasil et al., 2016). The rate of fetal transmission and the actual incidence of serious congenital infections remain unclear. The virus has been detected in fetal cerebral tissues by RT-PCR and electron microscopy (Mlakar et al., 2016).
Brasil et al. (2016) included 345 pregnant women with a history of rash within 5 previous days in a cohort study from September 2015 to May 2016. A total of 182 (53%) tested positive for ZIKV in the blood, urine, or both, and outcomes were available for 125 of these patients. There were nine fetal deaths, and the remaining 116 ZIKV-positive women gave birth to 117 live infants, 42% of which had grossly altered clinical and/or neuroimaging findings, four of them with microcephaly. Melo et al. (2016) followed 11 infants with congenital ZIKV syndrome from gestation to 6 months in Brazil, identifying neurological injuries that included lissencephaly with hydrocephalus, ventriculomegaly, microcephaly, reduction in cerebral volume, cerebellar hypoplasia and arthrogryposis. Soares de Oliveira-Szejnfeld et al. (2016) performed radiological investigation in 17 patients with confirmed ZIKV infection and 18 with presumed ZIKV. The authors reported that neuroimaging abnormalities were similar between both groups, with the more frequent being ventriculomegaly, corpus callosum, infections, intracranial calcifications in the junction of the gray-white matter or in the thalamus and/or basal ganglia.
Ventura et al. (2016a) first described ocular lesions in three infants of ZIKV-infected mothers in Brazil, who presented with microcephaly and cerebral calcifications. Mothers and their children underwent ophthalmological evaluation by biomicroscopy and fundus examination. None of the infected mothers presented eye lesions; however, their infants had gross pigment mottle in the macula and no foveal reflex. ZIKV infection was not tested by RT-PCR, but this was the first warning of its deleterious effects on the developing visual system. The same group also reported on a 57-day infant without microcephaly that had chorioretinal scar on the macula of the left eye (Ventura et al., 2016b). Later, the same group published a study with 40 infants with microcephaly, and reported that eye lesions were more frequent in infants whose mothers had symptoms during the first trimester and infants who had smaller cephalic diameters (Ventura et al., 2016c).
A report by de Paula Freitas et al. (2016) evaluated 23 out of 27 women with suspected infection by ZIKV who had presented clinical manifestations during gestation. Eighteen of them presented them during the first trimester; four occurred in the second trimester and one in the third trimester of gestation. The newborns were studied by ophthalmic examination and, of 29 children (58 eyes), 10 (35.5%) ocular abnormalities were identified in 17 eyes (29.3%). The main fundus results were perimacular and macular pigmented and atrophic lesions, diffuse retinal pigment epithelial damage and chorioretinal atrophy, which in some cases presented very severe forms affecting the macula. They also reported the occurrence of optic nerve abnormalities, mainly hypoplasia, and other findings such as retinal vasculitis, lens subluxation and iris coloboma. In the study, the authors described bilateral findings in 2/3 of the children examined. The authors determined that congenital ZIKV infection was associated with potentially blinding eye disease, including bilateral irreversible macular and perimacular lesions and involvement of the optic nerve (de Paula Freitas et al., 2016).
Well before the first suspected cases of ZIKV-associated neurological disorders in humans, there were reports showing the marked neurotropism of ZIKV strains isolated in Uganda (Dick, 1952; Bell et al., 1971). These studies briefly described the neuropathological changes induced by the intracerebral inoculation of ZIKV in newborn and adult mice, such as the severe neuronal degeneration and reactive astrocytosis in the hippocampus of newborn Webster Swiss mice inoculated with ZIKV (Bell et al., 1971). Only recently, however, after the recent outbreak of ZIKV in South and Central America, the study of the vulnerability of neural cells to ZIKV infection has become a focus of intense research.
In vitro studies using human induced pluripotent stem cells (iPSC) have allowed the investigation of the consequences of ZIKV infection in different types of human neural stem cells (NSC), neural progenitor cells (NPC) and their progeny, as well as in cerebral organoids (Cugola et al., 2016; Garcez et al., 2016; Souza et al., 2016; Tang et al., 2016). Human organotypic fetal brain slices (Onorati et al., 2016; Retallack et al., 2016), human NPC derived from the fetal brain (Hanners et al., 2016; Liang et al., 2016; Onorati et al., 2016) and immortalized cells lines also have been employed to study the neurovirulence of ZIKV.
Tang et al. (2016) were the first to show that the MR766 strain of ZIKV (from Uganda) infected iPSC-derived forebrain-specific human NPC, leading to cell-cycle dysregulation and apoptosis, whereas human iPSC and immature neurons exhibited lower levels of infection. In line with these evidence, Garcez et al. (2016) showed that, while both ZIKV (MR766 strain) and DENV 2 (16681 strain) were capable of infecting human NSC, only ZIKV induced apoptosis in NSC, impaired the formation of neurospheres and decreased the growth rate of human brain organoids. Further studies revealed that brain organoids represent an interesting platform for the study of ZIKV-associated microcephaly (Cugola et al., 2016; Dang et al., 2016; Qian et al., 2016; Wells et al., 2016; Gabriel et al., 2017). For instance, Qian et al. (2016) exposed brain organoids at different stages of cortical neurogenesis to ZIKV (MR766) for 24 h. They showed that most of the infected cells were NPC, although the virus could be detected to a lesser extent in immature neurons, intermediate progenitor cells, and astrocytes. Infection of early stage organoids decreased the number of proliferating cells and induced apoptosis in infected and non-infected cells. As a consequence, the ventricular zone and the neuronal layer were thinner and the ventricles were enlarged in infected organoids, resembling some of the characteristics of microcephaly.
Studies using an Asian strain of ZIKV (FSS13025, isolated in Cambodia) (Zhang F. et al., 2016) or ZIKV isolated from recent outbreaks in Brazil (Cugola et al., 2016; Souza et al., 2016; Garcez et al., 2017; Sacramento et al., 2017), Puerto Rico (Hanners et al., 2016; Wells et al., 2016) and French Polynesia (Ghouzzi et al., 2016) have shown the ability of the virus to infect and induce apoptosis of human NPC.
Another common feature of the different strains of ZIKV is the capacity to inhibit the proliferation of NPC. In this regard, Liang et al. (2016) screened the effects of ten ZIKV-encoding potential proteins and found that the ectopic expression of two proteins (NS4A and NS4B, alone or in combination) inhibited neurosphere formation, decreased the proliferation rates of NSC derived from human fetuses, and reduced their capacity to differentiate into neurons and astrocytes. They showed that the co-expression of NS4A and NS4B induced autophagy, by impairing Akt/mTOR signaling, and suggested that the efficient replication of ZIKV requires autophagy (Liang et al., 2016). Infection with either the Brazilian ZIKV or the Cambodian ZIKV strain FSS 13025 caused mitotic abnormalities and increased the number of neural stem/progenitor cells with supernumerary centrosomes (Onorati et al., 2016; Souza et al., 2016; Garcez et al., 2017). Supernumerary foci of centriolar proteins have also been found after ZIKV infection (Polynesia strain PF-25013-18) in untransformed human retinal epithelia RPE-1 cells and human CHME3 microglial cells, but not in the NPC line ReN (Wolf et al., 2017). Onorati et al. (2016) have proposed a model in which ZIKV infection activates RIG-I-like receptors, cytoplasmic sensors of different forms of dsRNA (which are present during the replication of ssRNA virus) (Thompson and Locarnini, 2007). This would, in turn, cause the relocation of phosphorylated TANK-binding kinase 1 (pTBK1) from centrosomes to mitochondria - where pTBK1 could take part in an innate antiviral immune response – resulting in mitotic impairments in neocortical neuroepithelial stem cells and radial glial cells (Onorati et al., 2016). Interestingly, the relocation of pTBK1 to the mitochondria was also induced by human cytomegalovirus (HCMV), another TORCH syndrome pathogen, but not by DENV 2 (16681 strain), despite the fact that both viruses induced apoptosis in human neocortical neuroepithelial stem cells derived from prenatal specimens (Onorati et al., 2016). The importance of ZIKV-induced centrosomal abnormalities was reinforced by recent findings from Gabriel et al. (Gabriel et al., 2017), who suggested a link between centrosomal structural defects in ZIKV-infected human NPC and the premature differentiation of NPC, which would result in the depletion of the NPC pool.
Although the knowledge of the cell biology of ZIKV is still scarce, recent advances have provided insights on the life cycle of this pathogen. ZIKV can bind to target cells using adhesion factors such as DC-SIGN and phosphatidylserine binding receptors (Hamel et al., 2017) from which Axl appeared as the main receptor for the entry in human skin fibroblasts (Hamel et al., 2015), microglia (Meertens et al., 2017), astrocytes (Retallack et al., 2016; Meertens et al., 2017) and blood-brain barrier endothelial cells (Liu et al., 2016). Surprisingly, Axl does not seem to be necessary for the entry of ZIKV in NPC and cerebral organoids (Wells et al., 2016; Meertens et al., 2017), despite the high expression of this receptor in NSC and NPC (Cugola et al., 2016; Liu et al., 2016; Nowakowski et al., 2016; Onorati et al., 2016; Meertens et al., 2017). Replication and assembly of progeny virions of ZIKV take place onto modified membranes derived from the endoplasmic reticulum. Thus, ZIKV-infected cells exhibit the characteristic ultrastructural alterations of flavivirus-infected cells (Dick, 1952; Offerdahl et al., 2017). In addition, ZIKV infection provokes changes in the pattern of cellular and viral RNA methylation (Lichinchi et al., 2016) and induces a major impact on the transcriptome of the host cell (Rolfe et al., 2016).
It has been shown that ZIKV (MR766) infection increases the expression of Toll-like receptor 3 (TLR3) (Dang et al., 2016), another innate immune receptor that recognizes dsRNA (Thompson and Locarnini, 2007), in iPS-derived human cerebral organoids and neurospheres. Treatment with the TLR3 agonist poly(I:C) decreased the size of neurospheres in a similar fashion to the infection with ZIKV. Nevertheless, treatment with a TLR3 competitive inhibitor only provided a modest protection against the deleterious effects of ZIKV in neurospheres and cerebral organoids (Dang et al., 2016). In another study, it was shown that while poly(I:C) induced the secretion of inflammatory mediators by human NPC, infection with a ZIKV strain isolated in Puerto Rico (ZIKV-PRVABC59) was not capable of inducing such a response – despite inducing apoptosis. In addition, ZIKV did not induce a type I Interferon response in NPC (did not induce IFN-α secretion) and did not stimulate cytokine secretion in THP-1 human monocytic cells (Hanners et al., 2016). ZIKV infection (MR766) was also shown to downregulate several immune response genes in a human microglial cell line (Tiwari et al., 2017). Suppression of the innate immune response in human CHME3 microglial cells infected with ZIKV was shown to depend on the kinase activity of Axl (Meertens et al., 2017).
Other studies, however, have found that ZIKV is capable of inducing an immune response in human fetal brain microglia (Lum et al., 2017) and other cell types. Human embryonic stem cells-derived cranial neural crest cells (CNCC) were induced to secrete high concentrations of several cytokines and growth factors, such as IL-6, PAI-1, LIF, and VEGF, after the infection with ZIKV (MR766 or H/PF/2013 strains). CNCC supported ZIKV replication and only a small fraction of the cells died after infection (Bayless et al., 2016). Moreover, ZIKV (PF-25013-18) induced the upregulation of the transcription factor IRF7 and several interferon-stimulated genes in human skin fibroblasts and lung epithelial A549 cells (Hamel et al., 2015; Frumence et al., 2016). Interestingly, the pattern of antiviral response induction in human astrocytes can differ between two ZIKV strains (H/PF/2013 and African HD 78788 strains) (Hamel et al., 2017).
Taken together, current evidence indicates that ZIKV preferentially infects NPC, inducing apoptosis, autophagy and interfering with mitosis (Figure 5). ZIKV replicates in NPC and the surviving cells produce the virus for several weeks (Hanners et al., 2016). All strains of ZIKV currently tested have been shown to induce nearly the same cytopathological effects in NPC and few studies have addressed whether there are mechanistic differences among the strains. Cugola et al. (2016) showed that, while the African strain reduced the number of neurons in non-human primate cerebral organoids, the Brazilian strain failed to replicate and did not change the number of neurons, suggesting that the Brazilian virus strain underwent adaptive changes in human cells. Gabriel et al. (2017) have also found some differences in the cellular outcome by comparing the effects of two strains of ZIKV isolated during the recent outbreaks and the African strain MR766 in iPSC-derived human NPC and cerebral organoids. Furthermore, Zhang F. et al. (2016) compared the effects of infecting iPSC-derived human NPC with an Asian ZIKV isolate (FSS13025) or the African strain (MR766). Both strains induced the same alterations in NPC (cell death and decreased proliferation) and caused similar gene expression changes: downregulation of genes involved in cell cycle, DNA repair, and DNA replication, coupled to the upregulation of genes involved in cell death and unfolded protein responses. Comparing the effect of both strains, they found that TP53 (coding for the tumor suppressor protein p53) was significantly upregulated following the infection with the Asian strain, but only marginally upregulated after the infection with the African strain. Accordingly, p53 inhibitors reduced the activation of caspase-3 more efficiently in NPC infected with the Asian strain (Zhang F. et al., 2016). Activation of p53 has also been reported after the infection of iPSC-derived human NPC with a ZIKV strain isolated from French Polynesia (PF13). The activation of p53 was not restricted to NPC expressing high levels of viral antigens (Ghouzzi et al., 2016), suggesting that this might be an early event or an indirect effect of ZIKV infection in neighboring cells. Other cell types, such as microglia, astrocytes, and endothelial cells, can also be infected (Delvecchio et al., 2016; Lum et al., 2017; Meertens et al., 2017), but it is unknown how the interplay among different cell types contribute to the outcome of microcephaly.
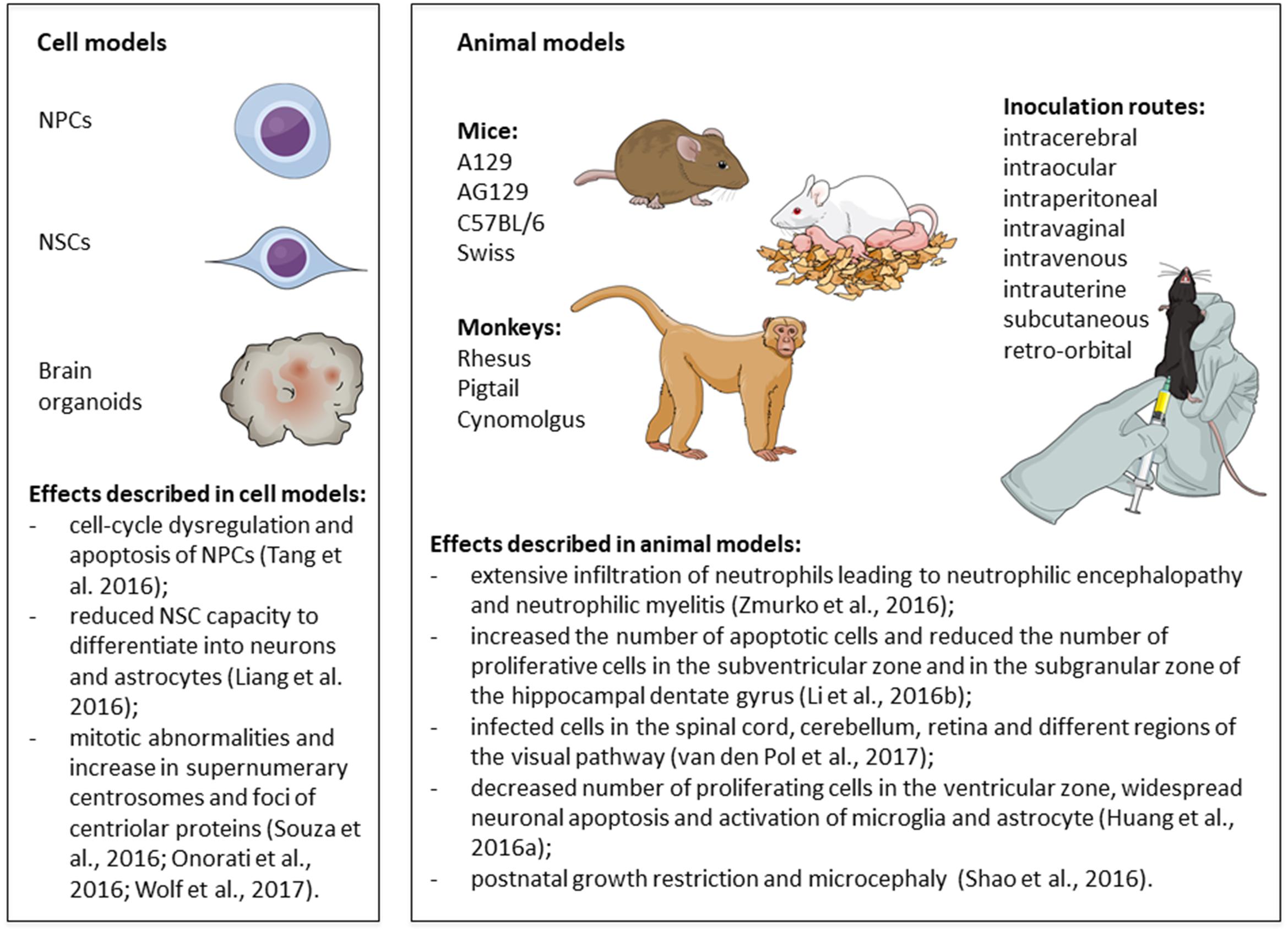
FIGURE 5. Mechanisms of Zika virus (ZIKV)-induced neural damage. Schematic illustration of the main findings from in vitro and in vivo models of Zika virus infection. Figure created in the Mind the Graph platform. NPCs: neural progenitor cells; NSCs: neural stem cells.
Several studies have used mice deficient in type I (A129 mice) or type I/II interferon receptors (AG129) to establish models of robust ZIKV infection in juvenile and adult animals. Aliota et al. (2016a) showed that the subcutaneous injection of a French Polynesian strain of ZIKV produced a lethal infection in AG129 mice. Animals exhibited early serum viremia and high viral loads in several organs, which was accompanied by signs of illness (including weight loss and lethargy) and brain degeneration. One of the most striking observations was the extensive infiltration of neutrophils in the brain. Animals had to be euthanized 7–8 days post-infection due to the rapid progression of the disease. Zmurko et al. (2016) infected AG129 mice with the African strain of ZIKV (MR766) and reported that the first signs of disease appeared at 10 days after the intraperitoneal inoculation. Mice developed acute neutrophilic encephalopathy and had to be euthanized, on average, at 14 days post-inoculation. Acute multifocal neutrophilic encephalitis, inflammatory lesions in the cerebellum and multifocal neutrophilic myelitis were also observed after the subcutaneous inoculation of AG129 mice with the Malaysian strain of ZIKV (P 6-740). In this model, all mice died within 21 days after the infection (Julander et al., 2017). Similar findings were reported after the infection of A129 mice with an African strain (MP1751) (Dowall et al., 2016), a Cambodian strain (FSS13025) (Rossi et al., 2016), or a French Polynesian strain of ZIKV (H/PF/2013) (Lazear et al., 2016). Rossi et al. (2016) compared the effects of ZIKV infection in both mouse strains and found that AG129 mice had more severe neurological symptoms than A129 mice, although there were no differences in other parameters, such as time to death and weight change, suggesting that type II interferon signaling might also influence certain aspects of the disease. Lazear et al. (2016) established a lethal infection in triple knockout mice deficient for IRF3, IRF5 and IRF7, which virtually do not produce interferon-α/β, and showed that these mice are more vulnerable to the intravenous injection of ZIKV than A129 mice, although there were no differences when the subcutaneous route was used. They also showed that the African strain MR766 was less pathogenic to A129 mice than the French Polynesian ZIKV strain (H/PF/2013). Another study using triple knockout mice (IRF3, IRF5, and IRF7 knockout) observed that ZIKV also has a tropism for NPC and immature neurons in the adult brain (Li H. et al., 2016). In this study, mice infected with a Cambodian strain of ZIKV (FSS13025) through a retro-orbital injection exhibited signs of viral illness and hindlimb weakness. The viral envelope protein was detected mainly in the subventricular zone and in the subgranular zone of the hippocampal dentate gyrus, two neurogenic regions in adults. Immunohistochemical analysis revealed that the infection increased the number of apoptotic cells and reduced the number of proliferative cells in these two neurogenic niches.
Several studies have demonstrated that immunocompetent mouse strains (CD1, C57BL/6 and 129Sv/Ev mice) are resistant to the subcutaneous or intraperitoneal inoculation of ZIKV and do not develop signs of illness (Dowall et al., 2016; Lazear et al., 2016; Rossi et al., 2016).
Immunocompetent mice only developed the disease when infected during the neonatal period (Lazear et al., 2016; Fernandes et al., 2017). For instance, van den Pol et al. (2017) inoculated C57BL/6 mice on the day of birth with an Asian strain of ZIKV (FSS13025) via the intraperitoneal route. They found that astrocytes were the first targets of the virus in the brain, which was followed by the infection of neurons. Infected cells were also found in the spinal cord, cerebellum, retina and different regions of the visual pathway. Infection was often lethal and caused growth restriction and motor dysfunction.
Fernandes et al. (2017) inoculated newborn Swiss mice with ZIKV (Brazilian strain SPH 2015), through the intracerebral or the subcutaneous routes. Animals from both groups showed neurological symptoms and severe illness, but the disease progressed faster in the intracerebral group. Histopathological findings were similar to what had been described in A129 and AG129 juvenile and adult animals. An interesting observation was that animals infected through the subcutaneous route additionally exhibited myelopathy, although the brain injury was less severe. Huang W. C. et al. (2016) infected C57BL/6 mice with ZIKV (MR766) at two different time points (either at postnatal day 7 or 21) through the intracerebral route. Infection of postnatal day 7 mice decreased the number of proliferating cells in the ventricular zone and resulted in widespread neuronal apoptosis and in the activation of microglia and astrocytes throughout the brain at 4 days post-infection, when the animals already exhibited neurological symptoms. At this time point, 21-day-old infected mice had severe paralysis, but the regional pattern of neuronal apoptosis was different and less prominent.
Three studies have also reported the effects of injecting ZIKV into the lateral ventricles of C57BL/6 and 129S1/SvImJ mouse embryos. Shao et al. (2016) observed postnatal growth restriction and microcephaly in newborn pups infected with a Mexican strain of ZIKV (MEX1-44) at embryonic day 14.5. Microscopic analysis of their brains revealed cortical thinning, massive neuronal death, glial activation and abnormal vascular density and permeability. Li C. et al. (2016) injected an Asian strain of ZIKV (SZ01) in the lateral ventricles of embryonic day 13.5 mice. They confirmed the tropism of ZIKV for NPC, although at a later time-point (5 days post-infection) almost all cells in the brain were positive for ZIKV. They also found that ZIKV induced the death of immature and mature neurons and reduced NPC proliferation and differentiation, which resulted in a thinner cortical layer. In both studies, there was no evidence of disruption of cortical lamination and the global transcriptome analyses of infected brains showed the upregulation of genes involved in immune-response-related and apoptosis pathways (Li C. et al., 2016; Shao et al., 2016). Wu et al. (2016), however, found the virus mainly in the ventricular zone and striatum of postnatal day 1 mice that were infected at embryonic day 13.5.
Thus far, animal models corroborate the evidence from in vitro and ex vivo studies, indicating that NPC are highly susceptible to ZIKV infection (Figure 5). Postmitotic neurons were also shown to be targets of infection in the developing brain and spinal cord of immunocompetent animals and in the brain of young and adult transgenic mice that do not mount an antiviral interferon response. Hindlimb paralysis and neurological symptoms were found in almost all models. Moreover, these models allowed the investigation of important questions raised by clinical and epidemiological observations, such as the link between ZIKV infection during pregnancy and the occurrence of microcephaly, and the possibility of sexual transmission of ZIKV.
The vertical transmission of ZIKV requires the ability of the virus to cross the placental barrier, as well as the developing or developed blood-brain barrier (BBB). Cugola et al. (2016) infected C57BL/6 or SJL pregnant mice on day 10–13 of gestation with a Brazilian strain of ZIKV. Newborns from the SJL ZIKV-infected mice had a high viral load in the brain, intra-uterine growth restriction (IUGR), cortical malformations and ocular defects. These alterations, however, were not observed in pups from C57BL/6 ZIKV-infected mice, indicating that the virus was not able to cross the placenta in this mouse strain, a finding that was supported by a recent study by van den Pol et al. (2017). This is in contrast with the findings of Wu et al. (2016), who injected ZIKV isolated from a patient contaminated in Samoa in the peritoneal cavity of pregnant C57BL/6 mice at embryonic day 13.5. They detected viral RNA in 5 out of 9 placentas and found the virus in radial glial cells localized in the ventricular zone of the fetal brains, 3 days after infection. ZIKV infection reduced the number of proliferating cells in the ventricular zone and intermediate zone and decreased the outer perimeter of the cortex of mice fetuses, although there was no difference in relative thickness of cortical layers. Recently, Xavier-Neto et al. (2017) showed a window of susceptibility between 5.5 and 9.5 days post coitum for ZIKV-induced teratogenesis in FVB/NJ and C57BL/6J WT mice. Their model of ZIKV (HS-2015-BA-01, isolated from a Brazilian patient) injection through the jugular vein of pregnant mice caused gross and generalized malformations, IUGR and neural tube defects in the embryos/fetuses.
Another strategy was employed by Miner et al. (2016) who crossed A129 female mice with wild-type (WT) males. Pregnant mice were infected with ZIKV (H/PF/2013) through the subcutaneous route on embryonic days 6.5 or 7.5. This model resulted in high levels of ZIKV RNA in the placenta (1000-fold greater than in the blood), placental abnormalities (vascular injury and apoptosis of trophoblasts) and fetal demise and resorption. IUGR and a large number of apoptotic cells in the brain were found in the remaining fetuses. Alternatively, they treated pregnant mice with a blocking anti-interferon alpha/beta receptor subunit 1 (MAR1-5A3) 24 or 48 h before the inoculation o ZIKV at embryonic days 6.5 or 7.5. This protocol did not cause fetal demise but caused mild IUGR and placental and fetal infection.
Another interesting observation from animal studies is that the brain and testes are the main sites of ZIKV replication in juvenile A129 and AG129 mice (Rossi et al., 2016) and that the virus can persist in the brain and testes for up to 28 days after infection in adult A129 mice (Lazear et al., 2016). Importantly, ZIKV infection (African strain Dakar 41519, and Asian strain H/PF/2013), although at lower levels, caused testis damage, oligospermia and decreased sex hormone production in WT mice treated with anti-Ifnar1 blocking antibodies (Govero et al., 2016). Testicular damage was also observed after the intraperitoneal inoculation of ZIKV (Asian strain SMGC-1) in A129 mice and after the intratesticular injection of ZIKV in WT mice (Ma et al., 2016). Future studies are warranted to determine whether the testicles are affected in ZIKV-infected men.
The possibility of ZIKV transmission through sexual contact was investigated by Yockey et al. (2016), who demonstrated that ZIKV (Cambodian strain FSS13025) replicates in the vaginal tract of WT mice after the intravaginal inoculation. However, while WT mice did not develop disease, A129 mice developed hindlimb paralysis and died within 9 days. However, it was shown that this model of intravaginal infection in WT mice can lead to IUGR if the maternal infection occurs during early pregnancy, even in the absence of viremia. The presence of ZIKV in neurons and glial cells in the fetal brain was also demonstrated. On the other hand, vaginal ZIKV exposure of pregnant A129 mice resulted in viremia, placental infection and fetal demise (early infection at embryonic day 4.5) or severe IUGR (infection at embryonic day 8.5). In conclusion, it has been shown that ZIKV can infect the fetal brain and cause developmental abnormalities (IUGR, for instance) in immunocompetent fetuses if there is a high viral load in the maternal vagina and/or blood, or if the virus is directly inoculated in the brain. Finally, Vermillion et al. (2017) developed a model of transplacental transmission in immunocompetent CD1 mice, in which ZIKV was directly inoculated into the uterine wall at embryonic day 10. At postnatal day 0, they found evidence of cortical thinning and microglial activation in the brain of pups from ZIKV-infected dams.
Although the models that use animals that do not mount an interferon response might not be useful for the screening of compounds that target the components of this anti-viral pathway, these models might be useful for testing other therapeutic strategies. Indeed, there is evidence that the ZIKV NS5 protein inhibits type I interferon response in human cells in a species-specific fashion, which might explain why WT mice are more resistant to the infection (Grant et al., 2016). Studies using animal models, therefore, represent an important step for the development of new therapies, especially when in conjunction with in vitro and ex vivo models (Delvecchio et al., 2016; Retallack et al., 2016; Zmurko et al., 2016; Julander et al., 2017; Sacramento et al., 2017). This includes the utilization of non-human primate models in preclinical translational studies. It has been shown that the infection of Indian-origin rhesus macaques with French Polynesian, South American, Puerto Rican or Thai ZIKV strains, via the subcutaneous route, results in transient viremia and uremia, prolonged presence of ZIKV RNA in cerebrospinal fluid, lymph nodes and colorectal tissue, as well as in the persistent presence of infectious ZIKV in the semen (Dudley et al., 2016; Li X.F. et al., 2016; Osuna et al., 2016; Aid et al., 2017). ZIKV infection induced T-cell responses and protected non-human primates from ZIKV re-infection or from heterologous ZIKV infection (Dudley et al., 2016; Osuna et al., 2016). Pregnant macaques infected at mid-first semester, however, exhibited persistent viremia, although the amniotic fluid was negative for ZIKV (Dudley et al., 2016). Moreover, Adams Waldorf et al. (2016) showed the vertical transmission of ZIKV (strain FSS13025, from Cambodia) after the subcutaneous inoculation of a pregnant pigtail macaque at 119 days of gestation. This model caused fetal brain lesions, including white matter injury and gliosis, brain growth arrest and an increase in the number of apoptotic cells in the subependymal zone, a neurogenic niche. ZIKV RNA was detected in both maternal and fetal brains and in the placenta at 6 weeks after inoculation, which might explain the persistent maternal viremia observed in other studies. Models of ZIKV infection in Cynomolgus monkeys have also been developed (Koide et al., 2016; Osuna et al., 2016), showing that these animals are susceptible to infection to ZIKV isolates from Cambodia and Puerto Rico, but not to an African ZIKV strain (IBH30656). In search of alternative and less expensive models, Goodfellow et al. (2016) have demonstrated that ZIKV induced a microcephaly like phenotype in chicken embryos. Finally, a model of intraocular ZIKV inoculation in mice (van den Pol et al., 2017) has already been developed and will permit the investigation of ZIKV-induced dysfunction of the retina and visual pathway.
Taken together, the large amount of data generated over the last years indicate that ZIKV can cause neural damage in an age-dependent manner. The causal relationship between ZIKV and birth defects has been inferred by using the Shepard’s criteria for proof of human teratogenicity and the Bradford Hill’s criteria for evidence of causation (Rasmussen et al., 2016). The publication of several experimental studies in animals showing the maternal-fetal transmission of ZIKV and the consequences of congenital infection have reinforced this conclusion. By analyzing data from epidemiologic and experimental studies, Krauer et al. (2017) concluded that ZIKV is a trigger of GBS and found evidence of causality between ZIKV and congenital abnormalities. However, the full spectrum of neurological complications and the long-term sequelae of ZIKV infection, even in the absence of microcephaly, remain to be determined. The mechanisms of neural damage at different developmental stages and the contribution of potential cofactors (nutritional status, viral load, previous infection with DENV, etc.) to the development of adverse outcomes also deserve further investigation.
Since the beginning of the 21st century, a number of infectious disease threats have emerged that were deemed to be such a risk that they demanded a global response. Although the transmission dynamics of the various pathogens causing these global emergencies differed, the overarching principles of the public health response were the same – development of policies and procedures, risk communication, effective surveillance, and use of disease control measures to mitigate the risk of infection in the population. These principles formed the bulk of the advice by the Emergency Committee on ZIKV to the Director-General of the WHO when the association between ZIKV and microcephaly and other neurological disorders (e.g., GBS) were declared to be a PHEIC on 1 February 2016 (WHO, 2016e). In addition to the public health measures, other key elements of that advice were that Zika-affected countries should prepare health services; research and development efforts should be increased; national authorities should ensure the rapid and timely reporting and sharing of information of public health importance relevant to this PHEIC; and there should be no restrictions on travel and trade as a result of the outbreak.
Because of the novel and unanticipated nature of the public health emergency relating to ZIKV, there were no specific pre-existing policies and protocols to guide the public health response. In December 2015, the European Centre for Disease Prevention and Control published a rapid risk assessment on the unfolding epidemic (ECDC, 2015) and, soon after, disease control agencies around the world developed public health and clinical guidelines. These guidelines were largely focused on women of reproductive age, their partners, and their clinicians, providing advice on how infection could be prevented, and on testing and management in the event of possible exposure (Petersen et al., 2016; WHO, 2016b). Microbiological testing for ZIKV is not straightforward and was not widely available at the outset of the epidemic, so the development of laboratory guidelines was another early focus.
Guidelines development accelerated following the WHO announcement that the world was experiencing a PHEIC. And as the scientific understanding of the risk and clinical manifestations of ZIKV infection on unborn children was rapidly evolving, guidelines required frequent updating. Over time, the guidelines for various agencies became more consistent with one another.
Surveillance activities for ZIKV in areas where transmission is occurring focus on recording case numbers, complications of the infection, and vectors. The objectives of the surveillance for human cases are to monitor the geographical distribution and temporal trend of infection; characterize disease presentation; identify complications related to the infection; identify non-vector borne routes of transmission; and monitor the effectiveness of containment measures (WHO, 2016d). In countries which are receptive to the virus but without local transmission, the key surveillance objectives are to ascertain imported cases and undertake vector monitoring. In countries where there is no chance of local transmission by vectors, the focus is on identifying imported cases.
The transmission dynamics of ZIKV mean that when it is introduced to an immunologically naive population, an epidemic ensues until a level of population immunity is reached that sees the end of the outbreak. Modeling shows that this period will last approximately 3 years, with a further decade or more before large epidemics are once again possible (Ferguson et al., 2016). It is important for a country to know where on the epidemic curve it is at any point in time, so that the risk to the population is understood and communicated to them, control measures can be instituted, and planning health services undertaken. Although the mild nature of ZIKV disease means that few will come to the attention of health authorities, an understanding of trends can be observed if a robust surveillance system is in place that allows counting of the relatively small proportion of cases that do seek medical help. The more robust systems are those where notification of ZIKV infection to public health authorities is included in the list of reportable infectious diseases mandated by law. Population-based serological studies can be undertaken in Zika-affected countries to better understand the level of population immunity at a point in time.
Estimating the risk of complications from ZIKV is difficult as baseline data for benchmarking are often unavailable and neither accurate denominator data (number of mothers infected), nor numerator data (number of congenital malformations), is available in most of those countries directly affected by ZIKV. Robust surveillance systems for measuring rates of congenital anomalies do exist but tend to occur in countries where ZIKV is not endemic. One such system is the US Zika Pregnancy Registry (USZPR), which provides perhaps the most accurate estimate of risk – of 442 women with completed pregnancies and laboratory evidence of recent Zika infection, 6% of fetuses or infants overall had one or more brain abnormalities and 4% had a finding of microcephaly (Honein et al., 2017). The rate was the same for symptomatic as for asymptomatic women and by far the greatest risk was for women who were infected in their first trimester.
The complication of GBS is rare, estimated to be 2.4 cases per 10,000 people infected during the 2013–2014 outbreak in French Polynesia (Cao-Lormeau et al., 2016). Estimates of the incidence of GBS from a number of Central and South American countries affected by ZIKV range from 2.0 to 9.8-fold higher than pre-epidemic baselines, which pose a significant burden to communities and health systems (Dos Santos et al., 2016).
The aims of vector surveillance are to determine when and where competent vectors are active in a country, where vector control efforts should be focused, and the effectiveness of vector control programs. As a consequence of dengue fever epidemics, most affected countries can rely on existing vector control programs. However, such programs are resource-intensive and need to be well-planned to ensure an efficient use of resources. Trapping sites should be placed to ensure representativeness of the geographical area, and fixed in place to allow determination of changes in mosquito densities over time (WHO, 2016f). Sampling should involve counting of both larval and adult forms of known mosquito vectors to help inform the use of larvicides, adulticides or both. Ideally, synthesis of all the information should be combined with geographic information systems (GIS) data to allow prediction of disease transmission scenarios and focus risk communication to the public. Another important component is the periodic determination of insecticide sensitivity to help selection of authorized biocides used in control efforts.
Raising awareness of the risk of ZIKV infection to those groups at risk is of critical importance. The population groups to be targeted will vary according to whether they are in a region or country where there is ZIKV activity. In those areas where ZIKV transmission is occurring, the entire population needs to be aware of the disease and vigilant of the risk in order for them to be motivated to undertake risk mitigation measures. Apart from the general public, specific stakeholder groups in the risk communication strategy should include pregnant women, women of reproductive age and their partners, community organizations, schools, health care workers, the media, local and international organizations involved in reproductive health, and local policymakers (WHO, 2016c). Key messages should include prevention of unintended pregnancies by the use of reversible contraception methods; using insect repellents, mosquito nets and other mosquito avoidance measures; and assisting in local vector control activities, such as reducing the mosquito breeding sites on private property. This last issue is essential, because the majority of breeding sites of Ae. albopictus and Ae. aegypti in urbanized areas are usually found on private property, where the simple preventive measure of water source elimination by owners can significantly reduce the risk.
The advice of health ministries of some Zika-countries for women to defer pregnancy for considerable periods to lessen the risk to their newborns is unprecedented and controversial. Apart from the attendant population planning risks of a distorted population profile resulting from a diminished birth cohort, some of these countries have high rates of unplanned pregnancies, strict abortion laws, a lack of sexuality education programs in schools, and poor access to contraception, leading to difficulties in implementation of this policy (Ahmed, 2016).
Because of the rarity of the complication of GBS, raising awareness among clinicians is likely to be the more effective risk communication strategy. However, the fact that cases may have mild, or no, symptoms of ZIKV infection preceding it means that clinicians won’t necessarily be aided by a clinical prompt, hence delays in diagnosis are likely.
In countries where ZIKV is not active and not receptive to the virus, the focus of awareness-raising is on those citizens planning to travel to Zika-affected areas, in particular, to individuals or couples who are pregnant or planning to become pregnant. Avoiding or deferring travel is advised, and if the person does decide to travel, the messages focus on mosquito avoidance and contraception advice, and symptoms to be aware of on return. Again, the high rate of asymptomatic infection means that the advice needs to relate to anyone traveling to endemic areas, not just those who develop symptoms. Based on available evidence of risk of sexual transmission of ZIKV, the WHO recommends men and women returning from areas where transmission of ZIKV is known to occur to abstain from unprotected sex for at least 6 months upon return to prevent ZIKV infection through sexual transmission and that women who are planning a pregnancy wait at least 6 months before trying to conceive to ensure that possible ZIKV infection has cleared (WHO, 2016b). Disease control agencies from individual countries advise a lesser risk period for women to defer pregnancy following their (or their partner’s) return from Zika-affected areas (CDC, 2017).
A third category includes those countries where ZIKV infection is not endemic but are receptive to the virus as they harbor competent vectors. The key population health messages in these countries relate to raising awareness of the symptoms of ZIKV infection so that cases are ascertained early and disease control measures rapidly instigated. Travelers from these countries to endemic areas need to be especially aware of risk mitigation measures whilst traveling to endemic areas, as well as symptoms of the infection, as they are at risk of triggering an outbreak on their return.
Disease control methods available for the fight against ZIKV are limited. There is neither an effective vaccine against the virus, nor anti-viral drugs to reduce the viremic period during which a competent vector with hematophagic behavior can amplify the disease. Vector control measures are, therefore, crucial and reduction of contact between hosts and vectors form the basis of disease control strategies worldwide.
Two broad types of complementary vector control strategies can be applied. First, individuals must take personal responsibility for avoiding mosquito bites and, second, government and non-government organizations must implement vector surveillance and control programs at global, national and local levels.
Recently, innovative methods to reduce mosquito populations have been shown great promise in laboratory and field conditions. Examples include the employment of insect sterile techniques (Alphey et al., 2010), introduction of Wolbachia strains (Jeffries and Walker, 2016) and genetically modified organisms (Beisel and Boëte, 2013), and these may be introduced in future years as adjuncts to conventional mosquito control programs.
In countries that do not have Zika activity but harbor competent vectors of the disease, disinsection of arriving airplanes is an important consideration. The WHO recommends that these countries undertake a risk assessment, and if it concludes that a disinsection program is indicated, that it should be conducted according to standard WHO recommendations (WHO, 2016a).
A study of blood donors during the outbreak in French Polynesia, which found that 3% were positive for ZIKV by PCR while asymptomatic (Musso et al., 2014), has led to concerns about the risk of transmission of ZIKV infection during the transfusion of blood products. The risk is small, however, with only one report of infection caused by transfusion of platelets (Motta et al., 2016).
Some countries require blood donors who have traveled to Zika-affected areas defer their donation for a period after their return or after any sexual contact with a known case (CDNA, 2016). Tests to screen blood donations for ZIKV are available and, despite them not being licensed by the United States Food and Drug Administration (US FDA), it recommends screening all donations in the US and the removal of any positive samples from the blood supply (FDA, 2016).
Vaccines for several flaviviruses have been produced during the past decades, some of them being already in the market, such as those for YF, tick-borne encephalitis (TBE), or WNF. These vaccines have been produced using different strategies: inactivated or live-attenuated viruses, recombinant proteins and recombinant subviral particles expressed in different heterologous systems, chimeric backbone viruses, or naked cDNA, among others (Martin-Acebes and Saiz, 2012). Thus, after the explosive spread of ZIKV in the Americas that quickly raised social and health worldwide concern because of its possible association with severe neurological pathologies (Blazquez and Saiz, 2016), it was reasonable to think that similar strategies can be applied to ZIKV (Saiz et al., 2016).
In this way, an alum-adjuvanted whole inactivated ZIKV (ZPIV) vaccine candidate recently showed complete protection against detectable viremia in challenged mice (Larocca et al., 2016) and later on in rhesus macaques (Abbink et al., 2016). On the other hand, although live attenuated vaccines are in widespread use for several viral infections, they are usually contraindicated for pregnant women, and in some instances for children, and therefore, they are not a primary target for ZIKV, even though WHO has reported no evidence of increased adverse pregnancy outcomes when licensed vaccines of this kind have been used (WHO, 2014). Similar issues apply for recombinant heterologous viral vectored vaccines and, thus, none of them have been licensed to date, even though a vaccine candidate that incorporates ZIKV pre-membrane and envelope (prM-E) proteins into a rhesus adenovirus serotype 52 viral vector has recently shown complete protection in challenged monkeys 4 weeks after vaccination with a single dose (Abbink et al., 2016).
On the other hand, until now, and though DNA vaccine technology has been available for many years, no such a human vaccine has been licensed; however, a DNA-ZIKV expressing the full-length ZIKV prM-E proteins induced complete protection against viremia both in mice (Larocca et al., 2016) and rhesus macaques (Abbink et al., 2016). Likewise, a recombinant vaccine ZIKV/JEV prM-E DNA constructs, in which the ZIKV prM signal sequence was replaced with the analogous JEV sequence to improve expression, has shown high levels of protection against viremia in challenged rhesus macaques (Dowd et al., 2016b). Even more, adoptive transfer of purified IgG from mice vaccinated with a ZIKV plasmid DNA vaccine conferred passive protection (Larocca et al., 2016), as did those from mice and rhesus macaques inoculated with an inactivated ZIKV vaccine (Abbink et al., 2016). Similarly, mice treated with a monoclonal antibody against the Domain III of ZIKV-E protein were protected from lethal ZIKV infection (Stettler et al., 2016).
As today, more than 30 vaccine candidates are in active preclinical development, and three have been already approved by the FDA to enter phase I clinical trials (WHO, 2017). NCT02809443 (GLS-5700, Inovio Pharmaceuticals and GeneOne Life Sciences) is a synthetic DNA plasmid vaccine which encodes for the pM-E regions of ZIKV that is being tested in healthy volunteers. This candidate is also in a phase I clinical trial in endemic areas in DENV seropositive adults (NCT02963909). NCT02840487 (VRC-ZKADNA085-00-VP, Vaccine Research Center, NIAID) is also composed of a single closed-circular DNA plasmid that encodes the prM-E proteins from ZIKV. NCT02963909 (NIAID) is an alum adjuvanted ZIKV purified inactivated vaccine (ZPIV) that has entered clinical trial in healthy flavivirus-naïve and flavivirus-primed subjects.
However, due to the characteristics of ZIKV infection, several specific concerns should be kept in mind when developing vaccine candidates against the virus. Thereby, given the possible association of the viral infection with congenital abnormalities (Blazquez and Saiz, 2016), the primary target of the vaccine would be pregnant women and women of childbearing age, although men may also be a target, as ZIKV has been detected in semen pointing to a sexual transmission route. In addition, there are also concerns over the possible interaction of preexisting flavivirus immunity with neutralization and/or enhancement, since several flavivirus cocirculate in the ZIKV endemic areas, including DENV for which an antibody dependent enhancement (ADE) effect has been described. Indeed, the relationships between the immune response to ZIKV and previous DENV infection has been recently demonstrated (Dejnirattisai et al., 2016; Stettler et al., 2016). However, no such ADE effect has been observed in ZIKV infected animal models challenge with WNV (Vázquez-Calvo et al., 2017a). Based on an ecological study, a possible protective effect of YZ fever vaccination has been discussed (De Goes Cavalcanti et al., 2016).
Furthermore, as there is no guarantee that experimental promising results will be reflected in the clinical evaluation of vaccine candidates, and based on the previous experience from the Ebola epidemic, where substantial delays occurred before the stakeholders established the necessary agreements, WHO has launched a target product profile (TPP) describing the preferred and minimal product characteristics for a vaccine targeted to the proposed priority populations (Vannice et al., 2016), which certainly will have to be updated in the coming months once new data are available. In fact, there are tests that take months to be evaluated (stability, neurovirulence, toxicity, etc.), but that should be addressed even under emergency circumstances. Moreover, as noted by Dittmer (2016), besides the technical and ethical aspects, several questions arise regarding ZIKV vaccine campaign implementation that should be considered: Do we need a sophisticated expensive vaccine? Is it economically worthy to develop a vaccine and large-scale clinical trials? Moreover, although vaccines potentially provide powerful tools for the control of viral pathogens, as mentioned before, the development of possible adverse effects derived from ADE of infection of related flavivirus (i.e., DENV) should also be extensively considered before starting large-scale vaccination campaigns.
As mentioned before, nowadays there are no approved specific antiviral agents against any flavivirus (Menendez-Arias and Richman, 2014), and treatment is generally directed to symptom relief with analgesics and antipyretic. However, in the past months, several drugs have been tested in vitro and in vivo as antiviral candidates (Saiz and Martin-Acebes, 2017), including the screening of different compounds libraries, such those already approved by the FDA, and the repurposing of drugs already used in clinic for other diseases, many of which are broad spectrum molecules.
For instances, different nucleoside analogs/derivatives that target viral polymerases, such as 2′-C-methylated nucleosides, have shown to inhibit ZIKV multiplication in cell culture (Eyer et al., 2016; Zmurko et al., 2016; Hercik et al., 2017), as did Sofosbuvir and BCX4430 (Bullard-Feibelman et al., 2017; Julander et al., 2017; Sacramento et al., 2017), which also induced greater survival rates in treated experimental immunodeficient mice (Bullard-Feibelman et al., 2017; Julander et al., 2017). Likewise, the pyrimidine synthesis inhibitors NITD008, CID 91632869, finasteride, brequinar, 6-azauridine, gemcitabine, and 5-fluorouracil reduced viral multiplication to different levels (Pascoalino et al., 2016; Adcock et al., 2017; Kuivanen et al., 2017). Even more, NITD008 improved survival rates in treated mice infected with ZIKV (Deng et al., 2016). In addition, by means of a high-throughput screening of over 40,000 compounds, it has been shown that non-peptidic small molecules targeting other viral proteins, such as the N2B-NS3 trypsin-like serine-protease, were capable of inhibiting ZIKV multiplication in cell culture (Lee et al., 2017).
On the other hand, it has been recently reported that passive transfer of human neutralizing antibodies to pregnant mice suppressed ZIKV replication and prevent microcephaly (Sapparapu et al., 2016; Wang et al., 2017), as did a monoclonal antibody against the Domain III of ZIKV-E protein protected against lethal ZIKV infection in a murine model (Stettler et al., 2016). Even more, compounds present in many natural products that also target the viral particle, such as the polyphenols epigallocatechin gallate and delphinidin chloride, also exhibit anti-ZIKV activity, probably through a virucidal effect (Carneiro et al., 2016; Vázquez-Calvo et al., 2017b).
Besides drugs targeting viral components, those directed against cellular factors implicated in the viral life cycle have also been assayed, as they are less prone to induce the emergence of resistant virus. In this line, by screening different libraries and bioactive molecules and by drug repurposing, different inhibitors of ZIKV infection were uncover, including the fusion inhibitors SaliPhe, monesin, and niclosamide (Xu M. et al., 2016; Adcock et al., 2017; Kuivanen et al., 2017), as well as others molecules that affect different cellular pathways, like bortzetomib, sertraline (Barrows et al., 2016), palonosetron (Pascoalino et al., 2016), tenovir-1 (Kuivanen et al., 2017), obatoclax (Rausch et al., 2017), PHA-690509, and emirascan (Xu M. et al., 2016). The immunosuppressants cyclosporine A, mycophenolic acid, and azathiophine have also been tested with promising results (Barrows et al., 2016). Likewise, hypolipidemic drugs like lovastatin, PF-429242, fatostatin, nordihydroguaiaretic acid, and tetra-O methyl nordihydroguaiaretic acid have demonstrated inhibitory activity against ZIKV in cell culture (Pascoalino et al., 2016; Merino-Ramos et al., 2017).
Finally, antiparasitics and antimalarials, such as ivermectin, chloroquine, quinacrine, mefloquine, GSK-36796, and pyrimethamine (Barrows et al., 2016; Delvecchio et al., 2016; Balasubramanian et al., 2017), as well as antibiotics like nanchagmycin, daptomycin and kitasmycin (Barrows et al., 2016; Pascoalino et al., 2016; Rausch et al., 2017) have also been shown to reduce ZIKV multiplication. Nevertheless, and despite the great effort made by the scientific community, it will take time until any drug against ZIKV will be commercially available.
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
e-26/201.341/2016 from FAPERJ to TO-C, PR-d-C and PP-C ZIKA-BIO-2016-2, RTA2013-0013-C04-01-E, and RTA2015-00009 from INIA to J-CS. AGL2014-56518-JIN from MINECO to MM-A.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
JH and RM-O are research fellows from the Brazilian Research Council Conselho Nacional de Desenvolvimento Científico e Tecnológico (CNPq). RM-O is a research fellow from Fundação Carlos Chagas Filho de Amparo à Pesquisa do Estado do Rio de Janeiro (FAPERJ). Figures 4, 5 were created using the Mind the Graph platform (www.mindthegraph.com).
Abbink, P., Larocca, R. A., De La Barrera, R. A., Bricault, C. A., Moseley, E. T., Boyd, M., et al. (2016). Protective efficacy of multiple vaccine platforms against Zika virus challenge in rhesus monkeys. Science 353, 1129–1132. doi: 10.1126/science.aah6157
Adams Waldorf, K. M., Stencel-Baerenwald, J. E., Kapur, R. P., Studholme, C., Boldenow, E., Vornhagen, J., et al. (2016). Fetal brain lesions after subcutaneous inoculation of Zika virus in a pregnant nonhuman primate. Nat. Med. 22, 1256–1259. doi: 10.1038/nm.4193
Adcock, R. S., Chu, Y. K., Golden, J. E., and Chung, D. H. (2017). Evaluation of anti-Zika virus activities of broad-spectrum antivirals and NIH clinical collection compounds using a cell-based, high-throughput screen assay. Antiviral Res. 138, 47–56. doi: 10.1016/j.antiviral.2016.11.018
Ahmad, S. S., Amin, T. N., and Ustianowski, A. (2016). Zika virus: management of infection and risk. BMJ 352:i1062. doi: 10.1136/bmj.i1062
Ahmed, A. (2016). El Salvador’s Advice on Zika Virus: Don’t Have Babies. The New York Times. Available at: http://www.nytimes.com/2016/01/26/world/americas/el-salvadors-advice-on-zika-dont-have-babies.html (accessed January 25, 2016).
Ahmed, Q. A., and Memish, Z. A. (2017). The public health planners’ perfect storm: hurricane Matthew and Zika virus. Travel Med. Infect. Dis. 15, 63–66. doi: 10.1016/j.tmaid.2016.12.004
Aid, M., Abbink, P., Larocca, R. A., Boyd, M., Nityanandam, R., Nanayakkara, O., et al. (2017). Zika virus persistence in the central nervous system and lymph nodes of rhesus monkeys. Cell 169, 610–620.e614. doi: 10.1016/j.cell.2017.04.008
Akiyama, B. M., Laurence, H. M., Massey, A. R., Costantino, D. A., Xie, X., Yang, Y., et al. (2016). Zika virus produces noncoding RNAs using a multi-pseudoknot structure that confounds a cellular exonuclease. Science 354, 1148–1152. doi: 10.1126/science.aah3963
Al-Abdely, H. M. (2016). Zika. An emerging teratogenic virus. Saudi Med. J. 37, 831–833. doi: 10.15537/smj.2016.8.15676
Aliota, M. T., Caine, E. A., Walker, E. C., Larkin, K. E., Camacho, E., and Osorio, J. E. (2016a). Characterization of lethal zika virus infection in AG129 Mice. PLoS Negl. Trop. Dis. 10:e0004682. doi: 10.1371/journal.pntd.0004682
Aliota, M. T., Peinado, S. A., Osorio, J. E., and Bartholomay, L. C. (2016b). Culex pipiens and Aedes triseriatus mosquito susceptibility to Zika virus. Emerg. Infect. Dis. 22, 1857–1859. doi: 10.3201/eid2210.161082
Alphey, L., Benedict, M., Bellini, R., Clark, G. G., Dame, D. A., Service, M. W., et al. (2010). Sterile-insect methods for control of mosquito-borne diseases: an analysis. Vector Borne Zoonotic Dis. 10, 295–311. doi: 10.1089/vbz.2009.0014
Althouse, B. M., Vasilakis, N., Sall, A. A., Diallo, M., Weaver, S. C., and Hanley, K. A. (2016). Potential for Zika virus to establish a sylvatic transmission cycle in the Americas. PLoS Negl. Trop. Dis. 10:e0005055. doi: 10.1371/journal.pntd.0005055
Amraoui, F., Atyame-Nten, C., Vega-Rua, A., Lourenco-de-Oliveira, R., Vazeille, M., and Failloux, A. B. (2016). Culex mosquitoes are experimentally unable to transmit Zika virus. Euro. Surveill. 21:30333. doi: 10.2807/1560-7917.es.2016.21.35.30333
Araujo, A. Q., Silva, M. T., and Araujo, A. P. (2016). Zika virus-associated neurological disorders: a review. Brain 139(Pt 8), 2122–2130. doi: 10.1093/brain/aww158
Attaway, D. F., Waters, N. M., Geraghty, E. M., and Jacobsen, K. H. (2017). Zika virus: endemic and epidemic ranges of Aedes mosquito transmission. J. Infect. Public Health 10, 120–123. doi: 10.1016/j.jiph.2016.09.008
Aubry, M., Teissier, A., Huart, M., Merceron, S., Vanhomwegen, J., Roche, C., et al. (2017). Zika virus seroprevalence, French Polynesia, 2014-2015. Emerg. Infect. Dis. 23, 669–672. doi: 10.3201/eid2304.161549
Balasubramanian, A., Teramoto, T., Kulkarni, A. A., Bhattacharjee, A. K., and Padmanabhan, R. (2017). Antiviral activities of selected antimalarials against dengue virus type 2 and Zika virus. Antiviral Res. 137, 141–150. doi: 10.1016/j.antiviral.2016.11.015
Barrows, N. J., Campos, R. K., Powell, S. T., Prasanth, K. R., Schott-Lerner, G., Soto-Acosta, R., et al. (2016). A screen of FDA-Approved drugs for inhibitors of Zika virus infection. Cell Host Microbe 20, 259–270. doi: 10.1016/j.chom.2016.07.004
Bayless, N. L., Greenberg, R. S., Swigut, T., Wysocka, J., and Blish, C. A. (2016). Zika virus infection induces cranial neural crest cells to produce cytokines at levels detrimental for neurogenesis. Cell Host Microbe 20, 423–428. doi: 10.1016/j.chom.2016.09.006
Beisel, U., and Boëte, C. (2013). The flying public health tool: genetically modified mosquitoes and malaria control. Sci. Cult. 22, 38–60. doi: 10.1080/09505431.2013.776364
Bell, T. M., Field, E. J., and Narang, H. K. (1971). Zika virus infection of the central nervous system of mice. Arch. Gesamte Virusforsch. 35, 183–193. doi: 10.1007/BF01249709
Bennett, K. L., Shija, F., Linton, Y. M., Misinzo, G., Kaddumukasa, M., Djouaka, R., et al. (2016). Historical environmental change in Africa drives divergence and admixture of Aedes aegypti mosquitoes: a precursor to successful worldwide colonization? Mol. Ecol. 25, 4337–4354. doi: 10.1111/mec.13762
Berthet, N., Nakoune, E., Kamgang, B., Selekon, B., Descorps-Declere, S., Gessain, A., et al. (2014). Molecular characterization of three Zika flaviviruses obtained from sylvatic mosquitoes in the Central African Republic. Vector Borne Zoonotic Dis. 14, 862–865. doi: 10.1089/vbz.2014.1607
Besnard, M., Eyrolle-Guignot, D., Guillemette-Artur, P., Lastere, S., Bost-Bezeaud, F., Marcelis, L., et al. (2016). Congenital cerebral malformations and dysfunction in fetuses and newborns following the 2013 to 2014 Zika virus epidemic in French Polynesia. Euro Surveill. 21:30181. doi: 10.2807/1560-7917.es.2016.21.13.30181
Besnard, M., Lastere, S., Teissier, A., Cao-Lormeau, V., and Musso, D. (2014). Evidence of perinatal transmission of Zika virus, French Polynesia, December 2013 and February 2014. Euro Surveill. 19:20751. doi: 10.2807/1560-7917.ES2014.19.13.20751
Blazquez, A. B., and Saiz, J. C. (2016). Neurological manifestations of Zika virus infection. World J. Virol. 5, 135–143. doi: 10.5501/wjv.v5.i4.135
Boccolini, D., Toma, L., Di Luca, M., Severini, F., Romi, R., Remoli, M. E., et al. (2016). Experimental investigation of the susceptibility of Italian Culex pipiens mosquitoes to Zika virus infection. Euro Surveill. 21:30328. doi: 10.2807/1560-7917.es.2016.21.35.30328
Brasil, P., Pereira, J. P. Jr., Moreira, M. E., Ribeiro Nogueira, R. M., Damasceno, L., Wakimoto, M., et al. (2016). Zika virus infection in pregnant women in Rio de Janeiro. N. Engl. J. Med. 375, 2321–2334. doi: 10.1056/NEJMoa1602412
Bullard-Feibelman, K. M., Govero, J., Zhu, Z., Salazar, V., Veselinovic, M., Diamond, M. S., et al. (2017). The FDA-approved drug sofosbuvir inhibits Zika virus infection. Antiviral Res. 137, 134–140. doi: 10.1016/j.antiviral.2016.11.023
Calvet, G., Aguiar, R. S., Melo, A. S., Sampaio, S. A., de Filippis, I., Fabri, A., et al. (2016). Detection and sequencing of Zika virus from amniotic fluid of fetuses with microcephaly in Brazil: a case study. Lancet Infect. Dis. 16, 653–660. doi: 10.1016/s1473-3099(16)00095-5
Camara, D. C., Codeco, C. T., Juliano, S. A., Lounibos, L. P., Riback, T. I., Pereira, G. R., et al. (2016). Seasonal differences in density but similar competitive impact of Aedes albopictus (Skuse) on Aedes aegypti (L.) in Rio de Janeiro, Brazil. PLoS ONE 11:e0157120. doi: 10.1371/journal.pone.0157120
Caminade, C., Turner, J., Metelmann, S., Hesson, J. C., Blagrove, M. S., Solomon, T., et al. (2017). Global risk model for vector-borne transmission of Zika virus reveals the role of El Nino 2015. Proc. Natl. Acad. Sci. U.S.A. 114, 119–124. doi: 10.1073/pnas.1614303114
Cao, X., Li, Y., Jin, X., Li, Y., Guo, F., and Jin, T. (2016). Molecular mechanism of divalent-metal-induced activation of NS3 helicase and insights into Zika virus inhibitor design. Nucleic Acids Res. 44, 10505–10514. doi: 10.1093/nar/gkw941
Cao-Lormeau, V. M., Blake, A., Mons, S., Lastere, S., Roche, C., Vanhomwegen, J., et al. (2016). Guillain-Barre syndrome outbreak associated with Zika virus infection in French Polynesia: a case-control study. Lancet 387, 1531–1539. doi: 10.1016/s0140-6736(16)00562-6
Caprara, A., and Ridde, V. (2016). Zika: nuevo revelador de la necesidad de promocion de la salud en America Latina. Glob. Health Promot. 23, 89–91. doi: 10.1177/1757975916673965
Carlson, C. J., Dougherty, E. R., and Getz, W. (2016). An ecological assessment of the pandemic threat of Zika virus. PLoS Negl. Trop. Dis. 10:e0004968. doi: 10.1371/journal.pntd.0004968
Carneiro, B. M., Batista, M. N., Braga, A. C., Nogueira, M. L., and Rahal, P. (2016). The green tea molecule EGCG inhibits Zika virus entry. Virology 496, 215–218. doi: 10.1016/j.virol.2016.06.012
Carteaux, G., Maquart, M., Bedet, A., Contou, D., Brugieres, P., Fourati, S., et al. (2016). Zika Virus Associated with Meningoencephalitis. N. Engl. J. Med. 374, 1595–1596. doi: 10.1056/NEJMc1602964
Casals, J. (1957). Viruses: the versatile parasites; the arthropod-borne group of animal viruses. Trans. N. Y. Acad. Sci. 19, 219–235. doi: 10.1111/j.2164-0947.1957.tb00526.x
CDC (2017). Zika Virus: Women & Their Partners Trying to Become Pregnant [Online]. Available at: https://www.cdc.gov/zika/pregnancy/women-and-their-partners.html (accessed March 27, 2017).
CDNA (2016). Zika Virus Infection. National Guidelines for Public Health Units [Online]. Available at: http://www.health.gov.au/internet/main/publishing.nsf/Content/cdna-song-zika.htm (accessed March 30, 2017).
Chouin-Carneiro, T., Vega-Rua, A., Vazeille, M., Yebakima, A., Girod, R., Goindin, D., et al. (2016). Differential susceptibilities of Aedes aegypti and Aedes albopictus from the Americas to Zika virus. PLoS Negl. Trop. Dis. 10:e0004543. doi: 10.1371/journal.pntd.0004543
Coloma, J., Jain, R., Rajashankar, K. R., Garcia-Sastre, A., and Aggarwal, A. K. (2016). Structures of NS5 methyltransferase from Zika virus. Cell Rep. 16, 3097–3102. doi: 10.1016/j.celrep.2016.08.091
Coutard, B., Barral, K., Lichiere, J., Selisko, B., Martin, B., Aouadi, W., et al. (2017). Zika virus methyltransferase: structure and functions for drug design perspectives. J. Virol. 91:e02202–16. doi: 10.1128/jvi.02202-16
Cugola, F. R., Fernandes, I. R., Russo, F. B., Freitas, B. C., Dias, J. L., Guimaraes, K. P., et al. (2016). The Brazilian Zika virus strain causes birth defects in experimental models. Nature 534, 267–271. doi: 10.1038/nature18296
Dai, L., Song, J., Lu, X., Deng, Y. Q., Musyoki, A. M., Cheng, H., et al. (2016). Structures of the Zika virus envelope protein and its complex with a flavivirus broadly protective antibody. Cell Host Microbe 19, 696–704. doi: 10.1016/j.chom.2016.04.013
Dang, J., Tiwari, S. K., Lichinchi, G., Qin, Y., Patil, V. S., Eroshkin, A. M., et al. (2016). Zika virus depletes neural progenitors in human cerebral organoids through activation of the innate immune receptor TLR3. Cell Stem Cell 19, 258–265. doi: 10.1016/j.stem.2016.04.014
De Goes Cavalcanti, L. P., Tauil, P. L., Alencar, C. H., Oliveira, W., Teixeira, M. M., and Heukelbach, J. (2016). Zika virus infection, associated microcephaly, and low yellow fever vaccination coverage in Brazil: is there any causal link? J. Infect. Dev. Ctries. 10, 563–566. doi: 10.3855/jidc.8575
de Paula Freitas, B., de Oliveira Dias, J. R., Prazeres, J., Sacramento, G. A., Ko, A. I., Maia, M., et al. (2016). Ocular findings in infants with microcephaly associated with presumed Zika virus congenital infection in Salvador, Brazil. JAMA Ophthalmol. doi: 10.1001/jamaophthalmol.2016.0267 [Epub ahead of print].
Dejnirattisai, W., Supasa, P., Wongwiwat, W., Rouvinski, A., Barba-Spaeth, G., Duangchinda, T., et al. (2016). Dengue virus sero-cross-reactivity drives antibody-dependent enhancement of infection with zika virus. Nat. Immunol. 17, 1102–1108. doi: 10.1038/ni.3515
Delvecchio, R., Higa, L. M., Pezzuto, P., Valadao, A. L., Garcez, P. P., Monteiro, F. L., et al. (2016). Chloroquine, an endocytosis blocking agent, inhibits Zika virus infection in different cell models. Viruses 8:322. doi: 10.3390/v8120322
Deng, Y. Q., Zhang, N. N., Li, C. F., Tian, M., Hao, J. N., Xie, X. P., et al. (2016). Adenosine Analog NITD008 is a potent inhibitor of Zika virus. Open Forum Infect. Dis. 3:ofw175. doi: 10.1093/ofid/ofw175
Dereeper, A., Guignon, V., Blanc, G., Audic, S., Buffet, S., Chevenet, F., et al. (2008). Phylogeny.fr: robust phylogenetic analysis for the non-specialist. Nucleic Acids Res. 36, W465–W469. doi: 10.1093/nar/gkn180
Di Luca, M., Severini, F., Toma, L., Boccolini, D., Romi, R., Remoli, M. E., et al. (2016). Experimental studies of susceptibility of Italian Aedes albopictus to Zika virus. Euro Surveill. 21:30223. doi: 10.2807/1560-7917.es.2016.21.18.30223
Diagne, C. T., Diallo, D., Faye, O., Ba, Y., Faye, O., Gaye, A., et al. (2015). Potential of selected Senegalese Aedes spp. mosquitoes (Diptera: Culicidae) to transmit Zika virus. BMC Infect. Dis. 15:492. doi: 10.1186/s12879-015-1231-2
Diallo, D., Sall, A. A., Diagne, C. T., Faye, O., Faye, O., Ba, Y., et al. (2014). Zika virus emergence in mosquitoes in southeastern Senegal, 2011. PLoS ONE 9:e109442. doi: 10.1371/journal.pone.0109442
Dick, G. W. (1952). Zika virus. II. Pathogenicity and physical properties. Trans. R. Soc. Trop. Med. Hyg. 46, 521–534. doi: 10.1016/0035-9203(52)90043-6
Dick, G. W. (1953). Epidemiological notes on some viruses isolated in uganda; yellow fever, rift valley fever, bwamba fever, West Nile, Mengo, Semliki forest, Bunyamwera, Ntaya, Uganda S and Zika viruses. Trans. R. Soc. Trop. Med. Hyg. 47, 13–48. doi: 10.1016/0035-9203(53)90021-2
Dick, G. W., Kitchen, S. F., and Haddow, A. J. (1952). Zika virus. I. Isolations and serological specificity. Trans. R. Soc. Trop. Med. Hyg. 46, 509–520. doi: 10.1016/0035-9203(52)90042-4
Dittmer, D. P. (2016). Zika vaccine: clinical trial and error? Science 353:1375. doi: 10.1126/science.aai8117
Donald, C. L., Brennan, B., Cumberworth, S. L., Rezelj, V. V., Clark, J. J., Cordeiro, M. T., et al. (2016). Full genome sequence and sfRNA interferon antagonist activity of Zika virus from recife, Brazil. PLoS Negl. Trop. Dis. 10:e0005048. doi: 10.1371/journal.pntd.0005048
Dos Santos, T., Rodriguez, A., Almiron, M., Sanhueza, A., Ramon, P., de Oliveira, W. K., et al. (2016). Zika virus and the guillain-barre syndrome - case series from seven countries. N. Engl. J. Med. 375, 1598–1601. doi: 10.1056/NEJMc1609015
Dowall, S. D., Graham, V. A., Rayner, E., Atkinson, B., Hall, G., Watson, R. J., et al. (2016). A susceptible mouse model for Zika virus infection. PLoS Negl. Trop. Dis. 10:e0004658. doi: 10.1371/journal.pntd.0004658
Dowd, K. A., DeMaso, C. R., Pelc, R. S., Speer, S. D., Smith, A. R., Goo, L., et al. (2016a). Broadly neutralizing activity of Zika virus-immune sera identifies a single viral serotype. Cell Rep. 16, 1485–1491. doi: 10.1016/j.celrep.2016.07.049
Dowd, K. A., Ko, S. Y., Morabito, K. M., Yang, E. S., Pelc, R. S., DeMaso, C. R., et al. (2016b). Rapid development of a DNA vaccine for Zika virus. Science 354, 237–240. doi: 10.1126/science.aai9137
Driggers, R. W., Ho, C. Y., Korhonen, E. M., Kuivanen, S., Jaaskelainen, A. J., Smura, T., et al. (2016). Zika virus infection with prolonged maternal viremia and fetal brain abnormalities. N. Engl. J. Med. 374, 2142–2151. doi: 10.1056/NEJMoa1601824
Dudley, D. M., Aliota, M. T., Mohr, E. L., Weiler, A. M., Lehrer-Brey, G., Weisgrau, K. L., et al. (2016). A rhesus macaque model of Asian-lineage Zika virus infection. Nat. Commun. 7:12204. doi: 10.1038/ncomms12204
Duffy, M. R., Chen, T. H., Hancock, W. T., Powers, A. M., Kool, J. L., Lanciotti, R. S., et al. (2009). Zika virus outbreak on yap island, federated states of micronesia. N. Engl. J. Med. 360, 2536–2543. doi: 10.1056/NEJMoa0805715
Dupont-Rouzeyrol, M., Biron, A., O’Connor, O., Huguon, E., and Descloux, E. (2016). Infectious Zika viral particles in breastmilk. Lancet 387, 1051. doi: 10.1016/s0140-6736(16)00624-3
ECDC (2015). Rapid Risk Assessment. Zika virus Epidemic in the Americas: Potential Association with Microcephaly and Guillain-Barré Syndrome [Online]. Available at: http://ecdc.europa.eu/en/publications/Publications/zika-virus-americas-association-with-microcephaly-rapid-risk-assessment.pdf
ECDC (2016). Zika virus Epidemic in the Americas: Potential Association with Microcephaly and Guillain-Barré Syndrome (First Update) [Online]. Available at: http://ecdc.europa.eu/en/publications/Publications/rapid-risk-assessment-zika-virus-first-update-jan-2016.pdf (accessed March 20, 2017).
Edgar, R. C. (2004). MUSCLE: multiple sequence alignment with high accuracy and high throughput. Nucleic Acids Res. 32, 1792–1797. doi: 10.1093/nar/gkh340
Escobar, L. E., Romero-Alvarez, D., Leon, R., Lepe-Lopez, M. A., Craft, M. E., Borbor-Cordova, M. J., et al. (2016). Declining prevalence of disease vectors under climate change. Sci. Rep. 6:39150. doi: 10.1038/srep39150
Eyer, L., Nencka, R., Huvarova, I., Palus, M., Joao Alves, M., Gould, E. A., et al. (2016). Nucleoside inhibitors of Zika virus. J. Infect. Dis. 214, 707–711. doi: 10.1093/infdis/jiw226
Faria, N. R., Azevedo Rdo, S., Kraemer, M. U., Souza, R., Cunha, M. S., Hill, S. C., et al. (2016). Zika virus in the Americas: early epidemiological and genetic findings. Science 352, 345–349. doi: 10.1126/science.aaf5036
Faye, O., Faye, O., Diallo, D., Diallo, M., Weidmann, M., and Sall, A. A. (2013). Quantitative real-time PCR detection of Zika virus and evaluation with field-caught mosquitoes. Virol. J. 10:311. doi: 10.1186/1743-422x-10-311
Faye, O., Freire, C. C., Iamarino, A., Faye, O., de Oliveira, J. V., Diallo, M., et al. (2014). Molecular evolution of Zika virus during its emergence in the 20(th) century. PLoS Negl. Trop. Dis. 8:e2636. doi: 10.1371/journal.pntd.0002636
FDA (2016). FDA News Release. FDA Advises Testing for Zika virus in all Donated Blood and Blood Components in the US [Online]. Available at: https://www.fda.gov/NewsEvents/Newsroom/PressAnnouncements/ucm518218.htm (accessed March 30, 2017).
Ferguson, N. M., Cucunuba, Z. M., Dorigatti, I., Nedjati-Gilani, G. L., Donnelly, C. A., Basanez, M. G., et al. (2016). EPIDEMIOLOGY. Countering the Zika epidemic in Latin America. Science 353, 353–354. doi: 10.1126/science.aag0219
Fernandes, N. C., Nogueira, J. S., Ressio, R. A., Cirqueira, C. S., Kimura, L. M., Fernandes, K. R., et al. (2017). Experimental Zika virus infection induces spinal cord injury and encephalitis in newborn Swiss mice. Exp. Toxicol. Pathol. 69, 63–71. doi: 10.1016/j.etp.2016.11.004
Fernandes, R. S., Campos, S. S., Ferreira-de-Brito, A., Miranda, R. M., Barbosa da Silva, K. A., Castro, M. G., et al. (2016). Culex quinquefasciatus from Rio de Janeiro is not competent to transmit the local Zika Virus. PLoS Negl. Trop. Dis. 10:e0004993. doi: 10.1371/journal.pntd.0004993
Ferreira-de-Brito, A., Ribeiro, I. P., Miranda, R. M., Fernandes, R. S., Campos, S. S., Silva, K. A., et al. (2016). First detection of natural infection of Aedes aegypti with Zika virus in Brazil and throughout South America. Mem. Inst. Oswaldo Cruz 111, 655–658. doi: 10.1590/0074-02760160332
Fischer, S., De Majo, M. S., Quiroga, L., Paez, M., and Schweigmann, N. (2017). Long-term spatio-temporal dynamics of the mosquito Aedes aegypti in temperate Argentina. Bull. Entomol. Res. 107, 225–233. doi: 10.1017/s0007485316000869
Foy, B. D., Kobylinski, K. C., Chilson Foy, J. L., Blitvich, B. J., Travassos da Rosa, A., Haddow, A. D., et al. (2011). Probable non-vector-borne transmission of Zika virus, Colorado, USA. Emerg. Infect. Dis. 17, 880–882. doi: 10.3201/eid1705.101939
Franca, R. F., Neves, M. H., Ayres, C. F., Melo-Neto, O. P., and Filho, S. P. (2016). First international workshop on Zika virus held by Oswaldo Cruz foundation FIOCRUZ in northeast Brazil March 2016 - A meeting report. PLoS Negl. Trop. Dis. 10:e0004760. doi: 10.1371/journal.pntd.0004760
Frumence, E., Roche, M., Krejbich-Trotot, P., El-Kalamouni, C., Nativel, B., Rondeau, P., et al. (2016). The south pacific epidemic strain of Zika virus replicates efficiently in human epithelial A549 cells leading to IFN-beta production and apoptosis induction. Virology 493, 217–226. doi: 10.1016/j.virol.2016.03.006
Gabriel, E., Ramani, A., Karow, U., Gottardo, M., Natarajan, K., Gooi, L. M., et al. (2017). Recent Zika Virus isolates induce premature differentiation of neural progenitors in human brain organoids. Cell Stem Cell 20, 397–406.e5. doi: 10.1016/j.stem.2016.12.005
Garcez, P. P., Loiola, E. C., Madeiro da Costa, R., Higa, L. M., Trindade, P., Delvecchio, R., et al. (2016). Zika virus impairs growth in human neurospheres and brain organoids. Science 352, 816–818. doi: 10.1126/science.aaf6116
Garcez, P. P., Nascimento, J. M., de Vasconcelos, J. M., Madeiro da Costa, R., Delvecchio, R., Trindade, P., et al. (2017). Zika virus disrupts molecular fingerprinting of human neurospheres. Sci. Rep. 7:40780. doi: 10.1038/srep40780
Gardner, L. M., Chen, N., and Sarkar, S. (2016). Global risk of Zika virus depends critically on vector status of Aedes albopictus. Lancet Infect. Dis. 16, 522–523. doi: 10.1016/s1473-3099(16)00176-6
Ghouzzi, V. E., Bianchi, F. T., Molineris, I., Mounce, B. C., Berto, G. E., Rak, M., et al. (2016). Zika virus elicits P53 activation and genotoxic stress in human neural progenitors similar to mutations involved in severe forms of genetic microcephaly and p53. Cell Death Dis. 7:e2440. doi: 10.1038/cddis.2016.266
Gong, Z., Gao, Y., and Han, G. Z. (2016). Zika virus: two or three lineages? Trends Microbiol. 24, 521–522. doi: 10.1016/j.tim.2016.05.002
Goo, L., Dowd, K. A., Smith, A. R., Pelc, R. S., DeMaso, C. R., and Pierson, T. C. (2016). Zika virus is not uniquely stable at physiological temperatures compared to other flaviviruses. mBio 7:e01396-16. doi: 10.1128/mBio.01396-16
Goodfellow, F. T., Tesla, B., Simchick, G., Zhao, Q., Hodge, T., Brindley, M. A., et al. (2016). Zika virus induced mortality and microcephaly in chicken embryos. Stem Cells Dev. 25, 1691–1697. doi: 10.1089/scd.2016.0231
Govero, J., Esakky, P., Scheaffer, S. M., Fernandez, E., Drury, A., Platt, D. J., et al. (2016). Zika virus infection damages the testes in mice. Nature 540, 438–442. doi: 10.1038/nature20556
Grant, A., Ponia, S. S., Tripathi, S., Balasubramaniam, V., Miorin, L., Sourisseau, M., et al. (2016). Zika virus targets human STAT2 to inhibit Type I interferon signaling. Cell Host Microbe 19, 882–890. doi: 10.1016/j.chom.2016.05.009
Grard, G., Caron, M., Mombo, I. M., Nkoghe, D., Mboui Ondo, S., Jiolle, D., et al. (2014). Zika virus in Gabon (Central Africa)–2007: a new threat from Aedes albopictus? PLoS Negl. Trop. Dis. 8:e2681. doi: 10.1371/journal.pntd.0002681
Gruba, N., Rodriguez Martinez, J. I., Grzywa, R., Wysocka, M., Skorenski, M., Burmistrz, M., et al. (2016). Substrate profiling of Zika virus NS2B-NS3 protease. FEBS Lett. 590, 3459–3468. doi: 10.1002/1873-3468.12443
Guedes, D. R. D., Paiva, M. H. S., Donato, M. M. A., Barbosa, P. P., Krokovsky, L., Rocha, S. W. D. S., et al. (2016). Zika Virus Replication in the Mosquito Culex Quinquefasciatus in Brazil. Available at: http://www.biorxiv.org/content/early/2016/09/02/073197 doi: 10.1101/073197
Guindon, S., Dufayard, J. F., Lefort, V., Anisimova, M., Hordijk, W., and Gascuel, O. (2010). New algorithms and methods to estimate maximum-likelihood phylogenies: assessing the performance of PhyML 3.0. Syst. Biol. 59, 307–321. doi: 10.1093/sysbio/syq010
Guo, X. X., Li, C. X., Deng, Y. Q., Xing, D., Liu, Q. M., Wu, Q., et al. (2016). Culex pipiens quinquefasciatus: a potential vector to transmit Zika virus. Emerg. Microbes Infect. 5:e102. doi: 10.1038/emi.2016.102
Guzzetta, G., Poletti, P., Montarsi, F., Baldacchino, F., Capelli, G., Rizzoli, A., et al. (2016). Assessing the potential risk of Zika virus epidemics in temperate areas with established Aedes albopictus populations. Euro. Surveill. 21:30199. doi: 10.2807/1560-7917.es.2016.21.15.30199
Haddow, A. D., Schuh, A. J., Yasuda, C. Y., Kasper, M. R., Heang, V., Huy, R., et al. (2012). Genetic characterization of Zika virus strains: geographic expansion of the Asian lineage. PLoS Negl. Trop. Dis. 6:e1477. doi: 10.1371/journal.pntd.0001477
Haddow, A. J., Williams, M. C., Woodall, J. P., Simpson, D. I., and Goma, L. K. (1964). Twelve isolations of Zika virus from aedes (stegomyia) africanus (theobald) taken in and above a uganda forest. Bull. World Health Organ. 31, 57–69.
Hall-Mendelin, S., Pyke, A. T., Moore, P. R., Mackay, I. M., McMahon, J. L., Ritchie, S. A., et al. (2016). Assessment of local mosquito species incriminates Aedes aegypti as the potential vector of Zika virus in Australia. PLoS Negl. Trop. Dis. 10:e0004959. doi: 10.1371/journal.pntd.0004959
Hamel, R., Dejarnac, O., Wichit, S., Ekchariyawat, P., Neyret, A., Luplertlop, N., et al. (2015). Biology of Zika virus infection in human skin cells. J. Virol. 89, 8880–8896. doi: 10.1128/jvi.00354-15
Hamel, R., Ferraris, P., Wichit, S., Diop, F., Talignani, L., Pompon, J., et al. (2017). African and Asian Zika virus strains differentially induce early antiviral responses in primary human astrocytes. Infect. Genet. Evol. 49, 134–137. doi: 10.1016/j.meegid.2017.01.015
Hanners, N. W., Eitson, J. L., Usui, N., Richardson, R. B., Wexler, E. M., Konopka, G., et al. (2016). Western Zika virus in human fetal neural progenitors persists long term with partial cytopathic and limited immunogenic effects. Cell Rep. 15, 2315–2322. doi: 10.1016/j.celrep.2016.05.075
Hart, C. E., Roundy, C. M., Azar, S. R., Huang, J. H., Yun, R., Reynolds, E., et al. (2017). Zika virus vector competency of mosquitoes, gulf coast, United States. Emerg. Infect. Dis. 23, 559–560. doi: 10.3201/eid2303.161636
Hayes, E. B. (2009). Zika virus outside Africa. Emerg. Infect. Dis. 15, 1347–1350. doi: 10.3201/eid1509.090442
Hercik, K., Kozak, J., Sala, M., Dejmek, M., Hrebabecky, H., Zbornikova, E., et al. (2017). Adenosine triphosphate analogs can efficiently inhibit the Zika virus RNA-dependent RNA polymerase. Antiviral Res. 137, 131–133. doi: 10.1016/j.antiviral.2016.11.020
Honein, M. A., Dawson, A. L., Petersen, E. E., Jones, A. M., Lee, E. H., Yazdy, M. M., et al. (2017). Birth defects among fetuses and infants of US women with evidence of possible Zika virus infection during pregnancy. JAMA 317, 59–68. doi: 10.1001/jama.2016.19006
Huang, W. C., Abraham, R., Shim, B. S., Choe, H., and Page, D. T. (2016). Zika virus infection during the period of maximal brain growth causes microcephaly and corticospinal neuron apoptosis in wild type mice. Sci. Rep. 6:34793. doi: 10.1038/srep34793
Huang, Y. J., Ayers, V. B., Lyons, A. C., Unlu, I., Alto, B. W., Cohnstaedt, L. W., et al. (2016). Culex species mosquitoes and Zika virus. Vector Borne Zoonotic Dis. 16, 673–676. doi: 10.1089/vbz.2016.2058
Ikejezie, J., Shapiro, C. N., Kim, J., Chiu, M., Almiron, M., Ugarte, C., et al. (2017). Zika virus transmission - Region of the Americas, May 15, 2015-December 15, 2016. MMWR Morb. Mortal. Wkly. Rep. 66, 329–334. doi: 10.15585/mmwr.mm6612a4
Jeffries, C. L., and Walker, T. (2016). Wolbachia biocontrol strategies for arboviral diseases and the potential influence of resident Wolbachia strains in mosquitoes. Curr. Trop. Med. Rep. 3, 20–25. doi: 10.1007/s40475-016-0066-2
Jian, Y., Silvestri, S., Brown, J., Hickman, R., and Marani, M. (2016). The predictability of mosquito abundance from daily to monthly timescales. Ecol. Appl. 26, 2609–2620. doi: 10.1002/eap.1405
Jouannic, J. M., Friszer, S., Leparc-Goffart, I., Garel, C., and Eyrolle-Guignot, D. (2016). Zika virus infection in French Polynesia. Lancet 387, 1051–1052. doi: 10.1016/s0140-6736(16)00625-5
Julander, J. G., Siddharthan, V., Evans, J., Taylor, R., Tolbert, K., Apuli, C., et al. (2017). Efficacy of the broad-spectrum antiviral compound BCX4430 against Zika virus in cell culture and in a mouse model. Antiviral Res. 137, 14–22. doi: 10.1016/j.antiviral.2016.11.003
Kindhauser, M. K., Allen, T., Frank, V., Santhana, R. S., and Dye, C. (2016). Zika: the origin and spread of a mosquito-borne virus. Bull. World Health Organ. 94, 675–686. doi: 10.2471/blt.16.171082
Koide, F., Goebel, S., Snyder, B., Walters, K. B., Gast, A., Hagelin, K., et al. (2016). Development of a Zika virus infection model in cynomolgus macaques. Front. Microbiol. 7:2028. doi: 10.3389/fmicb.2016.02028
Kostyuchenko, V. A., Lim, E. X., Zhang, S., Fibriansah, G., Ng, T. S., Ooi, J. S., et al. (2016). Structure of the thermally stable Zika virus. Nature 533, 425–428. doi: 10.1038/nature17994
Kraemer, M. U., Sinka, M. E., Duda, K. A., Mylne, A. Q., Shearer, F. M., Barker, C. M., et al. (2015a). The global distribution of the arbovirus vectors Aedes aegypti and Ae. albopictus. Elife 4:e08347. doi: 10.7554/eLife.08347
Kraemer, M. U., Sinka, M. E., Duda, K. A., Mylne, A., Shearer, F. M., Brady, O. J., et al. (2015b). The global compendium of Aedes aegypti and Ae. albopictus occurrence. Sci. Data 2:150035. doi: 10.1038/sdata.2015.35
Krauer, F., Riesen, M., Reveiz, L., Oladapo, O. T., Martinez-Vega, R., Porgo, T. V., et al. (2017). Zika virus infection as a cause of congenital brain abnormalities and Guillain-Barre syndrome: systematic review. PLoS Med. 14:e1002203. doi: 10.1371/journal.pmed.1002203
Kuivanen, S., Bespalov, M. M., Nandania, J., Ianevski, A., Velagapudi, V., De Brabander, J. K., et al. (2017). Obatoclax, saliphenylhalamide and gemcitabine inhibit Zika virus infection in vitro and differentially affect cellular signaling, transcription and metabolism. Antiviral Res. 139, 117–128. doi: 10.1016/j.antiviral.2016.12.022
Kuno, G., and Chang, G. J. (2007). Full-length sequencing and genomic characterization of Bagaza, Kedougou, and Zika viruses. Arch. Virol. 152, 687–696. doi: 10.1007/s00705-006-0903-z
Kuno, G., Chang, G. J., Tsuchiya, K. R., Karabatsos, N., and Cropp, C. B. (1998). Phylogeny of the genus flavivirus. J. Virol. 72, 73–83.
Larocca, R. A., Abbink, P., Peron, J. P., Zanotto, P. M., Iampietro, M. J., Badamchi-Zadeh, A., et al. (2016). Vaccine protection against Zika virus from Brazil. Nature 536, 474–478. doi: 10.1038/nature18952
Lazear, H. M., Govero, J., Smith, A. M., Platt, D. J., Fernandez, E., Miner, J. J., et al. (2016). A mouse model of Zika virus pathogenesis. Cell Host Microbe 19, 720–730. doi: 10.1016/j.chom.2016.03.010
Lebrun, G., Chadda, K., Reboux, A. H., Martinet, O., and Gauzere, B. A. (2009). Guillain-Barre syndrome after chikungunya infection. Emerg. Infect. Dis. 15, 495–496. doi: 10.3201/eid1503.071482
Ledermann, J. P., Guillaumot, L., Yug, L., Saweyog, S. C., Tided, M., Machieng, P., et al. (2014). Aedes hensilli as a potential vector of Chikungunya and Zika viruses. PLoS Negl. Trop. Dis. 8:e3188. doi: 10.1371/journal.pntd.0003188
Lee, H., Ren, J., Nocadello, S., Rice, A. J., Ojeda, I., Light, S., et al. (2017). Identification of novel small molecule inhibitors against NS2B/NS3 serine protease from Zika virus. Antiviral Res. 139, 49–58. doi: 10.1016/j.antiviral.2016.12.016
Lei, J., Hansen, G., Nitsche, C., Klein, C. D., Zhang, L., and Hilgenfeld, R. (2016). Crystal structure of Zika virus NS2B-NS3 protease in complex with a boronate inhibitor. Science 353, 503–505. doi: 10.1126/science.aag2419
Leis, A. A., and Stokic, D. S. (2012). Neuromuscular manifestations of west nile virus infection. Front. Neurol. 3:37. doi: 10.3389/fneur.2012.00037
Lennemann, N. J., and Coyne, C. B. (2017). Dengue and Zika viruses subvert reticulophagy by NS2B3-mediated cleavage of FAM134B. Autophagy 13, 322–332. doi: 10.1080/15548627.2016.1265192
Lessler, J., Chaisson, L. H., Kucirka, L. M., Bi, Q., Grantz, K., Salje, H., et al. (2016). Assessing the global threat from Zika virus. Science 353:aaf8160. doi: 10.1126/science.aaf8160
Li, C., Xu, D., Ye, Q., Hong, S., Jiang, Y., Liu, X., et al. (2016). Zika virus disrupts neural progenitor development and leads to microcephaly in mice. Cell Stem Cell 19, 120–126. doi: 10.1016/j.stem.2016.04.017
Li, G., Poulsen, M., Fenyvuesvolgyi, C., Yashiroda, Y., Yoshida, M., Simard, J. M., et al. (2017). Characterization of cytopathic factors through genome-wide analysis of the Zika viral proteins in fission yeast. Proc. Natl. Acad. Sci. U.S.A. 114, E376–E385. doi: 10.1073/pnas.1619735114
Li, H., Saucedo-Cuevas, L., Regla-Nava, J. A., Chai, G., Sheets, N., Tang, W., et al. (2016). Zika virus infects neural progenitors in the adult mouse brain and alters proliferation. Cell Stem Cell 19, 593–598. doi: 10.1016/j.stem.2016.08.005
Li, X., Liu, T., Lin, L., Song, T., Du, X., Lin, H., et al. (2017). Application of the analytic hierarchy approach to the risk assessment of Zika virus disease transmission in Guangdong Province, China. BMC Infect. Dis. 17:65. doi: 10.1186/s12879-016-2170-2
Li, X. F., Dong, H. L., Huang, X. Y., Qiu, Y. F., Wang, H. J., Deng, Y. Q., et al. (2016). Characterization of a 2016 clinical isolate of Zika virus in non-human primates. EBioMedicine 12, 170–177. doi: 10.1016/j.ebiom.2016.09.022
Li, Y., He, L., He, R. L., and Yau, S. S. (2017). Zika and Flaviviruses phylogeny based on the alignment-free natural vector method. DNA Cell Biol. 36, 109–116. doi: 10.1089/dna.2016.3532
Liang, Q., Luo, Z., Zeng, J., Chen, W., Foo, S. S., Lee, S. A., et al. (2016). Zika Virus NS4A and NS4B proteins deregulate Akt-mTOR signaling in human fetal neural stem cells to inhibit neurogenesis and induce autophagy. Cell Stem Cell 19, 663–671. doi: 10.1016/j.stem.2016.07.019
Lichinchi, G., Zhao, B. S., Wu, Y., Lu, Z., Qin, Y., He, C., et al. (2016). Dynamics of human and viral RNA methylation during Zika virus infection. Cell Host Microbe 20, 666–673. doi: 10.1016/j.chom.2016.10.002
Liu, S., DeLalio, L. J., Isakson, B. E., and Wang, T. T. (2016). AXL-mediated productive infection of human endothelial cells by Zika virus. Circ. Res. 119, 1183–1189. doi: 10.1161/circresaha.116.309866
Lu, G., Bluemling, G. R., Collop, P., Hager, M., Kuiper, D., Gurale, B. P., et al. (2017). Analysis of ribonucleotide 5′-triphosphate analogs as potential inhibitors of Zika virus RNA-dependent RNA polymerase by using nonradioactive polymerase assays. Antimicrob. Agents Chemother. 61:e01967-16. doi: 10.1128/aac.01967-16
Lum, F. M., Low, D. K., Fan, Y., Tan, J. J., Lee, B., Chan, J. K., et al. (2017). Zika virus infects human fetal brain microglia and induces inflammation. Clin. Infect. Dis. 64, 914–920. doi: 10.1093/cid/ciw878
Ma, W., Li, S., Ma, S., Jia, L., Zhang, F., Zhang, Y., et al. (2016). Zika virus causes testis damage and leads to male infertility in mice. Cell 167, 1511.e10–1524.e10. doi: 10.1016/j.cell.2016.11.016
Marchette, N. J., Garcia, R., and Rudnick, A. (1969). Isolation of Zika virus from Aedes aegypti mosquitoes in Malaysia. Am. J. Trop. Med. Hyg. 18, 411–415. doi: 10.4269/ajtmh.1969.18.411
Martin-Acebes, M. A., and Saiz, J. C. (2012). West Nile virus: a re-emerging pathogen revisited. World J. Virol. 1, 51–70. doi: 10.5501/wjv.v1.i2.51
Matheron, S., d’Ortenzio, E., Leparc-Goffart, I., Hubert, B., de Lamballerie, X., and Yazdanpanah, Y. (2016). Long-lasting persistence of Zika virus in semen. Clin. Infect. Dis. 63:1264. doi: 10.1093/cid/ciw509
Meertens, L., Labeau, A., Dejarnac, O., Cipriani, S., Sinigaglia, L., Bonnet-Madin, L., et al. (2017). Axl mediates ZIKA virus entry in human glial cells and modulates innate immune responses. Cell Rep. 18, 324–333. doi: 10.1016/j.celrep.2016.12.045
Melo, A. S., Aguiar, R. S., Amorim, M. M., Arruda, M. B., Melo, F. O., Ribeiro, S. T., et al. (2016). Congenital Zika virus infection: beyond neonatal microcephaly. JAMA Neurol. 73, 1407–1416. doi: 10.1001/jamaneurol.2016.3720
Menendez-Arias, L., and Richman, D. D. (2014). Editorial overview: antivirals and resistance: advances and challenges ahead. Curr. Opin. Virol. 8, iv–vii. doi: 10.1016/j.coviro.2014.08.002
Merino-Ramos, T., Jimenez de Oya, N., Saiz, J. C., and Martin-Acebes, M. A. (2017). Antiviral activity of nordihydroguaiaretic acid and its derivative tetra-O-methyl nordihydroguaiaretic acid against West Nile virus and Zika virus. Antimicrob. Agents Chemother. 61:e00376-17. doi: 10.1128/aac.00376-17
Messina, J. P., Kraemer, M. U., Brady, O. J., Pigott, D. M., Shearer, F. M., Weiss, D. J., et al. (2016). Mapping global environmental suitability for Zika virus. Elife 5:e15272. doi: 10.7554/eLife.15272
Miner, J. J., Cao, B., Govero, J., Smith, A. M., Fernandez, E., Cabrera, O. H., et al. (2016). Zika virus infection during pregnancy in mice causes placental damage and fetal demise. Cell 165, 1081–1091. doi: 10.1016/j.cell.2016.05.008
Misslin, R., Telle, O., Daude, E., Vaguet, A., and Paul, R. E. (2016). Urban climate versus global climate change-what makes the difference for dengue? Ann. N. Y. Acad. Sci. 1382, 56–72. doi: 10.1111/nyas.13084
Mlakar, J., Korva, M., Tul, N., Popovic, M., Poljsak-Prijatelj, M., Mraz, J., et al. (2016). Zika virus associated with microcephaly. N. Engl. J. Med. 374, 951–958. doi: 10.1056/NEJMoa1600651
Monaghan, A. J., Morin, C. W., Steinhoff, D. F., Wilhelmi, O., Hayden, M., Quattrochi, D. A., et al. (2016). On the seasonal occurrence and abundance of the Zika virus vector mosquito Aedes aegypti in the contiguous United States. PLoS Curr. Outbreaks. doi: 10.1371/currents.outbreaks.50dfc7f46798675fc63e7d7da563da76
Moreira, J., Peixoto, T. M., Siqueira, A. M., and Lamas, C. C. (2017). Sexually acquired Zika virus: a systematic review. Clin. Microbiol. Infect. 23, 296–305. doi: 10.1016/j.cmi.2016.12.027
Mossenta, M., Marchese, S., Poggianella, M., Slon Campos, J. L., and Burrone, O. R. (2017). Role of N-glycosylation on Zika virus E protein secretion, viral assembly and infectivity. Biochem. Biophys. Res. Commun. doi: 10.1016/j.bbrc.2017.01.022 [Epub ahead of print].
Motta, I. J., Spencer, B. R., Cordeiro, da Silva, S. G., Arruda, M. B., Dobbin, J. A., et al. (2016). Evidence for transmission of Zika virus by platelet transfusion. N. Engl. J. Med. 375, 1101–1103. doi: 10.1056/NEJMc1607262
Moulin, E., Selby, K., Cherpillod, P., Kaiser, L., and Boillat-Blanco, N. (2016). Simultaneous outbreaks of dengue, Chikungunya and Zika virus infections: diagnosis challenge in a returning traveller with nonspecific febrile illness. New Microbes New Infect. 11, 6–7. doi: 10.1016/j.nmni.2016.02.003
Munoz, A. G., Thomson, M. C., Goddard, L., and Aldighieri, S. (2016). Analyzing climate variations at multiple timescales can guide Zika virus response measures. Gigascience 5:41. doi: 10.1186/s13742-016-0146-1
Murray, K. O., Gorchakov, R., Carlson, A. R., Berry, R., Lai, L., Natrajan, M., et al. (2017). Prolonged detection of Zika virus in vaginal secretions and whole blood. Emerg. Infect. Dis. 23, 99–101. doi: 10.3201/eid2301.161394
Musso, D., Nhan, T., Robin, E., Roche, C., Bierlaire, D., Zisou, K., et al. (2014). Potential for Zika virus transmission through blood transfusion demonstrated during an outbreak in French Polynesia, November 2013 to February 2014. Euro Surveill. 19:20761. doi: 10.2807/1560-7917.ES2014.19.14.20761
Musso, D., Roche, C., Robin, E., Nhan, T., Teissier, A., and Cao-Lormeau, V. M. (2015). Potential sexual transmission of Zika virus. Emerg. Infect. Dis. 21, 359–361. doi: 10.3201/eid2102.141363
Nicastri, E., Castilletti, C., Liuzzi, G., Iannetta, M., Capobianchi, M. R., and Ippolito, G. (2016). Persistent detection of Zika virus RNA in semen for six months after symptom onset in a traveller returning from Haiti to Italy, February 2016. Euro Surveill. 21:30314. doi: 10.2807/1560-7917.es.2016.21.32.30314
Nowakowski, T. J., Pollen, A. A., Di Lullo, E., Sandoval-Espinosa, C., Bershteyn, M., and Kriegstein, A. R. (2016). Expression analysis highlights AXL as a candidate Zika virus entry receptor in neural stem cells. Cell Stem Cell 18, 591–596. doi: 10.1016/j.stem.2016.03.012
Offerdahl, D. K., Dorward, D. W., Hansen, B. T., and Bloom, M. E. (2017). Cytoarchitecture of Zika virus infection in human neuroblastoma and Aedes albopictus cell lines. Virology 501, 54–62. doi: 10.1016/j.virol.2016.11.002
Oliveira Melo, A. S., Malinger, G., Ximenes, R., Szejnfeld, P. O., Alves Sampaio, S., Bispo, et al. (2016). Zika virus intrauterine infection causes fetal brain abnormality and microcephaly: tip of the iceberg? Ultrasound Obstet. Gynecol. 47, 6–7. doi: 10.1002/uog.15831
Onorati, M., Li, Z., Liu, F., Sousa, A. M., Nakagawa, N., Li, M., et al. (2016). Zika virus disrupts phospho-TBK1 localization and mitosis in human neuroepithelial stem cells and radial glia. Cell Rep. 16, 2576–2592. doi: 10.1016/j.celrep.2016.08.038
Osuna, C. E., Lim, S. Y., Deleage, C., Griffin, B. D., Stein, D., Schroeder, L. T., et al. (2016). Zika viral dynamics and shedding in rhesus and cynomolgus macaques. Nat. Med. 22, 1448–1455. doi: 10.1038/nm.4206
Pacheco, O., Beltran, M., Nelson, C. A., Valencia, D., Tolosa, N., Farr, S. L., et al. (2016). Zika virus disease in Colombia - preliminary report. N. Engl. J. Med. doi: 10.1056/NEJMoa1604037 [Epub ahead of print].
Paixao, E. S., Barreto, F., Teixeira Mda, G., Costa Mda, C., and Rodrigues, L. C. (2016). History, epidemiology, and clinical manifestations of Zika: a systematic review. Am. J. Public Health 106, 606–612. doi: 10.2105/ajph.2016.303112
Paploski, I. A., Prates, A. P., Cardoso, C. W., Kikuti, M., Silva, M. M., Waller, L. A., et al. (2016). Time lags between exanthematous illness attributed to Zika virus, Guillain-Barre syndrome, and microcephaly, Salvador, Brazil. Emerg. Infect. Dis. 22, 1438–1444. doi: 10.3201/eid2208.160496
Pascoalino, B. S., Courtemanche, G., Cordeiro, M. T., Gil, L. H., and Freitas-Junior, L. (2016). Zika antiviral chemotherapy: identification of drugs and promising starting points for drug discovery from an FDA-approved library. F1000Res. 5:2523. doi: 10.12688/f1000research.9648.1
Penot, P., Brichler, S., Guilleminot, J., Lascoux-Combe, C., Taulera, O., Gordien, E., et al. (2017). Infectious Zika virus in vaginal secretions from an HIV-infected woman, France, August 2016. Euro Surveill. 22:30444. doi: 10.2807/1560-7917.es.2017.22.3.30444
Petersen, E. E., Staples, J. E., Meaney-Delman, D., Fischer, M., Ellington, S. R., Callaghan, W. M., et al. (2016). Interim guidelines for pregnant women during a Zika virus outbreak–United States, 2016. MMWR Morb. Mortal. Wkly. Rep. 65, 30–33. doi: 10.15585/mmwr.mm6502e1
Phoo, W. W., Li, Y., Zhang, Z., Lee, M. Y., Loh, Y. R., Tan, Y. B., et al. (2016). Structure of the NS2B-NS3 protease from Zika virus after self-cleavage. Nat. Commun. 7:13410. doi: 10.1038/ncomms13410
Prasad, V. M., Miller, A. S., Klose, T., Sirohi, D., Buda, G., Jiang, W., et al. (2017). Structure of the immature Zika virus at 9 Å resolution. Nat. Struct. Mol. Biol. 24, 184–186. doi: 10.1038/nsmb.3352
Prisant, N., Breurec, S., Moriniere, C., Bujan, L., and Joguet, G. (2017). Zika virus genital tract shedding in infected women of childbearing age. Clin. Infect. Dis. 64, 107–109. doi: 10.1093/cid/ciw669
Prisant, N., Bujan, L., Benichou, H., Hayot, P. H., Pavili, L., Lurel, S., et al. (2016). Zika virus in the female genital tract. Lancet Infect. Dis. 16, 1000–1001. doi: 10.1016/s1473-3099(16)30193-1
Qian, X., Nguyen, H. N., Song, M. M., Hadiono, C., Ogden, S. C., Hammack, C., et al. (2016). Brain-region-specific organoids using mini-bioreactors for modeling ZIKV exposure. Cell 165, 1238–1254. doi: 10.1016/j.cell.2016.04.032
Rasmussen, S. A., Jamieson, D. J., Honein, M. A., and Petersen, L. R. (2016). Zika virus and birth defects–reviewing the evidence for causality. N. Engl. J. Med. 374, 1981–1987. doi: 10.1056/NEJMsr1604338
Rausch, K., Hackett, B. A., Weinbren, N. L., Reeder, S. M., Sadovsky, Y., Hunter, C. A., et al. (2017). Screening bioactives reveals nanchangmycin as a broad spectrum antiviral active against Zika virus. Cell Rep. 18, 804–815. doi: 10.1016/j.celrep.2016.12.068
Ravi, V., Taly, A. B., Shankar, S. K., Shenoy, P. K., Desai, A., Nagaraja, D., et al. (1994). Association of Japanese encephalitis virus infection with Guillain-Barre syndrome in endemic areas of south India. Acta Neurol. Scand. 90, 67–72. doi: 10.1111/j.1600-0404.1994.tb02681.x
Retallack, H., Di Lullo, E., Arias, C., Knopp, K. A., Laurie, M. T., Sandoval-Espinosa, C., et al. (2016). Zika virus cell tropism in the developing human brain and inhibition by azithromycin. Proc. Natl. Acad. Sci. U.S.A. 113, 14408–14413. doi: 10.1073/pnas.1618029113
Richard, V., Paoaafaite, T., and Cao-Lormeau, V. M. (2016). Vector competence of French polynesian Aedes aegypti and Aedes polynesiensis for Zika Virus. PLoS Negl. Trop. Dis. 10:e0005024. doi: 10.1371/journal.pntd.0005024
Robert, M. A., Christofferson, R. C., Silva, N. J., Vasquez, C., Mores, C. N., and Wearing, H. J. (2016). Modeling mosquito-borne disease spread in U.S. urbanized areas: the case of dengue in Miami. PLoS ONE 11:e0161365. doi: 10.1371/journal.pone.0161365
Rodriguez-Morales, A. J. (2015). Zika: the new arbovirus threat for Latin America. J. Infect. Dev. Ctries 9, 684–685. doi: 10.3855/jidc.7230
Rolfe, A. J., Bosco, D. B., Wang, J., Nowakowski, R. S., Fan, J., and Ren, Y. (2016). Bioinformatic analysis reveals the expression of unique transcriptomic signatures in Zika virus infected human neural stem cells. Cell Biosci. 6:42. doi: 10.1186/s13578-016-0110-x
Rossi, S. L., Tesh, R. B., Azar, S. R., Muruato, A. E., Hanley, K. A., Auguste, A. J., et al. (2016). Characterization of a novel murine model to study Zika virus. Am. J. Trop. Med. Hyg. 94, 1362–1369. doi: 10.4269/ajtmh.16-0111
Roze, B., Najioullah, F., Signate, A., Apetse, K., Brouste, Y., Gourgoudou, S., et al. (2016). Zika virus detection in cerebrospinal fluid from two patients with encephalopathy, Martinique, February 2016. Euro Surveill. 21:30205. doi: 10.2807/1560-7917.ES.2016.21.16.30205
Rut, W., Zhang, L., Kasperkiewicz, P., Poreba, M., Hilgenfeld, R., and Drag, M. (2017). Extended substrate specificity and first potent irreversible inhibitor/activity-based probe design for Zika virus NS2B-NS3 protease. Antiviral Res. 139, 88–94. doi: 10.1016/j.antiviral.2016.12.018
Sacramento, C. Q., de Melo, G. R., de Freitas, C. S., Rocha, N., Hoelz, L. V., Miranda, M., et al. (2017). The clinically approved antiviral drug sofosbuvir inhibits Zika virus replication. Sci. Rep. 7:40920. doi: 10.1038/srep40920
Sahoo, M., Jena, L., Daf, S., and Kumar, S. (2016). Virtual screening for potential inhibitors of NS3 protein of Zika virus. Genomics Inform. 14, 104–111. doi: 10.5808/gi.2016.14.3.104
Saiz, J. C., and Martin-Acebes, M. A. (2017). The race to find antivirals for Zika virus. Antimicrob. Agents Chemother. 61:e00411-17. doi: 10.1128/aac.00411-17
Saiz, J. C., Vazquez-Calvo, A., Blazquez, A. B., Merino-Ramos, T., Escribano-Romero, E., and Martin-Acebes, M. A. (2016). Zika virus: the latest newcomer. Front. Microbiol. 7:496. doi: 10.3389/fmicb.2016.00496
Saluzzo, J. F., Gonzalez, J. P., Herve, J. P., and Georges, A. J. (1981). Serological survey for the prevalence of certain arboviruses in the human population of the south-east area of Central African Republic (author’s transl). Bull. Soc. Pathol. Exot. Filiales 74, 490–499.
Samy, A. M., Thomas, S. M., Wahed, A. A., Cohoon, K. P., and Peterson, A. T. (2016). Mapping the global geographic potential of Zika virus spread. Mem. Inst. Oswaldo Cruz 111, 559–560. doi: 10.1590/0074-02760160149
Sapparapu, G., Fernandez, E., Kose, N., Bin, C., Fox, J. M., Bombardi, R. G., et al. (2016). Neutralizing human antibodies prevent Zika virus replication and fetal disease in mice. Nature 540, 443–447. doi: 10.1038/nature20564
Schuler-Faccini, L., Ribeiro, E. M., Feitosa, I. M., Horovitz, D. D., Cavalcanti, D. P., Pessoa, A., et al. (2016). Possible association between Zika virus infection and microcephaly - Brazil, 2015. Morb. Mortal. Wkly. Rep. 65, 59–62. doi: 10.15585/mmwr.mm6503e2
Shao, Q., Herrlinger, S., Yang, S. L., Lai, F., Moore, J. M., Brindley, M. A., et al. (2016). Zika virus infection disrupts neurovascular development and results in postnatal microcephaly with brain damage. Development 143, 4127–4136. doi: 10.1242/dev.143768
Sirohi, D., Chen, Z., Sun, L., Klose, T., Pierson, T. C., Rossmann, M. G., et al. (2016). The 3.8 A resolution cryo-EM structure of Zika virus. Science 352, 467–470. doi: 10.1126/science.aaf5316
Smithburn, K. C. (1952). Neutralizing antibodies against certain recently isolated viruses in the sera of human beings residing in East Africa. J. Immunol. 69, 223–234.
Soares, C. N., Brasil, P., Carrera, R. M., Sequeira, P., de Filippis, A. B., Borges, V. A., et al. (2016). Fatal encephalitis associated with Zika virus infection in an adult. J. Clin. Virol. 83, 63–65. doi: 10.1016/j.jcv.2016.08.297
Soares de Oliveira-Szejnfeld, P., Levine, D., Melo, A. S., Amorim, M. M., Batista, A. G., Chimelli, L., et al. (2016). Congenital brain abnormalities and Zika virus: what the radiologist can expect to see prenatally and postnatally. Radiology 281, 203–218. doi: 10.1148/radiol.2016161584
Song, H., Qi, J., Haywood, J., Shi, Y., and Gao, G. F. (2016). Zika virus NS1 structure reveals diversity of electrostatic surfaces among flaviviruses. Nat. Struct. Mol. Biol. 23, 456–458. doi: 10.1038/nsmb.3213
Souza, B. S., Sampaio, G. L., Pereira, C. S., Campos, G. S., Sardi, S. I., Freitas, L. A., et al. (2016). Zika virus infection induces mitosis abnormalities and apoptotic cell death of human neural progenitor cells. Sci. Rep. 6:39775. doi: 10.1038/srep39775
Steinhagen, K., Probst, C., Radzimski, C., Schmidt-Chanasit, J., Emmerich, P., van Esbroeck, M., et al. (2016). Serodiagnosis of Zika virus (ZIKV) infections by a novel NS1-based ELISA devoid of cross-reactivity with dengue virus antibodies: a multicohort study of assay performance, 2015 to 2016. Euro Surveill. 21:30426. doi: 10.2807/1560-7917.es.2016.21.50.30426
Stephen, P., Baz, M., Boivin, G., and Lin, S. X. (2016). Structural insight into NS5 of Zika virus leading to the discovery of MTase inhibitors. J. Am. Chem. Soc. 138, 16212–16215. doi: 10.1021/jacs.6b10399
Stettler, K., Beltramello, M., Espinosa, D. A., Graham, V., Cassotta, A., Bianchi, S., et al. (2016). Specificity, cross-reactivity, and function of antibodies elicited by Zika virus infection. Science 353, 823–826. doi: 10.1126/science.aaf8505
SVS/MS (2017). Boletim Epidemiológico. Available at: http://portalarquivos.saude.gov.br/images/pdf/2017/fevereiro/27/2017_003.pdf (accessed 6, 48).
Swaminathan, S., Schlaberg, R., Lewis, J., Hanson, K. E., and Couturier, M. R. (2016). Fatal Zika virus infection with secondary nonsexual transmission. N. Engl. J. Med. 375, 1907–1909. doi: 10.1056/NEJMc1610613
Tang, H., Hammack, C., Ogden, S. C., Wen, Z., Qian, X., Li, Y., et al. (2016). Zika virus infects human cortical neural progenitors and attenuates their growth. Cell Stem Cell 18, 587–590. doi: 10.1016/j.stem.2016.02.016
Thangamani, S., Huang, J., Hart, C. E., Guzman, H., and Tesh, R. B. (2016). Vertical transmission of Zika virus in Aedes aegypti mosquitoes. Am. J. Trop. Med. Hyg. 95, 1169–1173. doi: 10.4269/ajtmh.16-0448
The Pan American Health Organization [PAHO]/World Health Organization [WHO] (2015). Zika Virus Infection. Available at: http://www.paho.org/hq/index.php?option=com_docman&task=doc_view&Itemid=270&gid=30075=en (accessed January 7, 2017).
Thompson, A. J., and Locarnini, S. A. (2007). Toll-like receptors, RIG-I-like RNA helicases and the antiviral innate immune response. Immunol. Cell Biol. 85, 435–445. doi: 10.1038/sj.icb.7100100
Tian, H., Ji, X., Yang, X., Zhang, Z., Lu, Z., Yang, K., et al. (2016). Structural basis of Zika virus helicase in recognizing its substrates. Protein Cell 7, 562–570. doi: 10.1007/s13238-016-0293-2
Tiwari, S. K., Dang, J., Qin, Y., Lichinchi, G., Bansal, V., and Rana, T. M. (2017). Zika virus infection reprograms global transcription of host cells to allow sustained infection. Emerg. Microbes Infect. 6, e24. doi: 10.1038/emi.2017.9
van den Pol, A. N., Mao, G., Yang, Y., Ornaghi, S., and Davis, J. N. (2017). Zika virus targeting in the developing brain. J. Neurosci. 37, 2161–2175. doi: 10.1523/jneurosci.3124-16.2017
Vannice, K. S., Giersing, B. K., Kaslow, D. C., Griffiths, E., Meyer, H., Barrett, A., et al. (2016). Meeting Report: WHO consultation on considerations for regulatory expectations of Zika virus vaccines for use during an emergency. Vaccine doi: 10.1016/j.vaccine.2016.10.034 [Epub ahead of print].
Vázquez-Calvo, A., Blázquez, A., Escribano-Romero, E., Merino-Ramos, T., Saiz, J. C., Martín-Acebes, M. A., et al. (2017a). Zika virus infection confers cross-protection against West Nile virus challenge in mice. Emerg. Microbes Infect. (in press).
Vázquez-Calvo,Á., Jiménez de Oya, N., Martín-Acebes, M. A., Garcia-Moruno, E., and Saiz, J.-C. (2017b). Antiviral properties of the natural polyphenols delphinidin and epigallocatechin gallate against the flaviviruses West Nile virus, Zika virus, and dengue virus. Front. Microbiol. 8:1314. doi: 10.3389/fmicb.2017.01314
Ventura, C. V., Maia, M., Bravo-Filho, V., Gois, A. L., and Belfort, R. Jr. (2016a). Zika virus in Brazil and macular atrophy in a child with microcephaly. Lancet 387, 228. doi: 10.1016/s0140-6736(16)00006-4
Ventura, C. V., Maia, M., Dias, N., Ventura, L. O., and Belfort, R. Jr. (2016b). Zika: neurological and ocular findings in infant without microcephaly. Lancet 387, 2502. doi: 10.1016/s0140-6736(16)30776-0
Ventura, C. V., Maia, M., Travassos, S. B., Martins, T. T., Patriota, F., Nunes, M. E., et al. (2016c). Risk factors associated with the ophthalmoscopic findings identified in infants with presumed Zika virus congenital infection. JAMA Ophthalmol. 134, 912–918. doi: 10.1001/jamaophthalmol.2016.1784
Verma, R., Sahu, R., and Holla, V. (2014). Neurological manifestations of dengue infection: a review. J. Neurol. Sci. 346, 26–34. doi: 10.1016/j.jns.2014.08.044
Vermillion, M. S., Lei, J., Shabi, Y., Baxter, V. K., Crilly, N. P., McLane, M., et al. (2017). Intrauterine Zika virus infection of pregnant immunocompetent mice models transplacental transmission and adverse perinatal outcomes. Nat. Commun. 8:14575. doi: 10.1038/ncomms14575
Vorou, R. (2016). Zika virus, vectors, reservoirs, amplifying hosts, and their potential to spread worldwide: what we know and what we should investigate urgently. Int. J. Infect. Dis. 48, 85–90. doi: 10.1016/j.ijid.2016.05.014
Walther, D., Scheuch, D. E., and Kampen, H. (2017). The invasive Asian tiger mosquito Aedes albopictus (Diptera: Culicidae) in Germany: local reproduction and overwintering. Acta Trop. 166, 186–192. doi: 10.1016/j.actatropica.2016.11.024
Wang, L., Valderramos, S. G., Wu, A., Ouyang, S., Li, C., Brasil, P., et al. (2016). From mosquitos to humans: genetic evolution of Zika virus. Cell Host Microbe 19, 561–565. doi: 10.1016/j.chom.2016.04.006
Wang, Q., Yang, H., Liu, X., Dai, L., Ma, T., Qi, J., et al. (2016). Molecular determinants of human neutralizing antibodies isolated from a patient infected with Zika virus. Sci. Transl. Med. 8:369ra179. doi: 10.1126/scitranslmed.aai8336
Wang, S., Hong, S., Deng, Y. Q., Ye, Q., Zhao, L. Z., Zhang, F. C., et al. (2017). Transfer of convalescent serum to pregnant mice prevents Zika virus infection and microcephaly in offspring. Cell Res. 27, 158–160. doi: 10.1038/cr.2016.144
Weger-Lucarelli, J., Ruckert, C., Chotiwan, N., Nguyen, C., Garcia Luna, S. M., Fauver, J. R., et al. (2016). Vector competence of American mosquitoes for three strains of Zika virus. PLoS Negl. Trop. Dis. 10:e0005101. doi: 10.1371/journal.pntd.0005101
Weinbren, M. P., and Williams, M. C. (1958). Zika virus: further isolations in the Zika area, and some studies on the strains isolated. Trans. R. Soc. Trop. Med. Hyg. 52, 263–268. doi: 10.1016/0035-9203(58)90085-3
Wells, M. F., Salick, M. R., Wiskow, O., Ho, D. J., Worringer, K. A., Ihry, R. J., et al. (2016). Genetic ablation of AXL does not protect human neural progenitor cells and cerebral organoids from Zika virus infection. Cell Stem Cell 19, 703–708. doi: 10.1016/j.stem.2016.11.011
WHO (2014). Safety of Immunization during Pregnancy. A Review of the Evidence. Available at: http://www.who.int/vaccine_safety/publications/safety_pregnancy_nov2014.pdf (accessed March 20, 2017).
WHO (2016a). Aircraft Disinsection for Mosquito Control. Available at: http://www.who.int/ihr/ports_airports/zika-aircraft-disinsection/en/ (accessed March 28, 2017).
WHO (2016b). Prevention of Sexual Transmission of Zika Virus: Interim Guidance. Available at: http://www.who.int/csr/resources/publications/zika/sexual-transmission-prevention/en/
WHO (2016c). Risk Communication in the Context of Zika Virus. Interim Guidance. Available at: http://www.who.int/csr/resources/publications/zika/risk-communication/en/ (accessed March 24, 2016).
WHO (2016d). Surveillance for Zika Virus Infection, Microcephaly and Guillain-Barré Syndrome. Interim Guidance. Available at: http://www.who.int/csr/resources/publications/zika/surveillance/en/ (accessed March 26, 2017).
WHO (2016e). WHO Statement on the First Meeting of the International Health Regulations (2005) (IHR 2005) Emergency Committee on Zika Virus and Observed Increase in Neurological Disorders and Neonatal Malformations. Available at: http://www.who.int/mediacentre/news/statements/2016/1st-emergency-committee-zika/en/ (accessed March 22, 2017).
WHO (2016f). Zika CDCD Interim Response Plan. Available at: https://www.cdc.gov/zika/pdfs/zika-draft-interim-conus-plan.pdf (accessed March 28, 2017).
WHO (2017). WHO Vaccine Pipeline Tracker. Available at: http://www.who.int/immunization/research/vaccine_pipeline_tracker_spreadsheet/en/ (accessed March 20, 2017).
Wikan, N., and Smith, D. R. (2016). Zika virus: history of a newly emerging arbovirus. Lancet Infect. Dis. 16, e119–e126. doi: 10.1016/s1473-3099(16)30010-x
Wolf, B., Diop, F., Ferraris, P., Wichit, S., Busso, C., Misse, D., et al. (2017). Zika virus causes supernumerary foci with centriolar proteins and impaired spindle positioning. Open Biol. 7:160231. doi: 10.1098/rsob.160231
Wong, P. S., Li, M. Z., Chong, C. S., Ng, L. C., and Tan, C. H. (2013). Aedes (Stegomyia) albopictus (Skuse): a potential vector of Zika virus in Singapore. PLoS Negl. Trop. Dis. 7:e2348. doi: 10.1371/journal.pntd.0002348
Wu, K. Y., Zuo, G. L., Li, X. F., Ye, Q., Deng, Y. Q., Huang, X. Y., et al. (2016). Vertical transmission of Zika virus targeting the radial glial cells affects cortex development of offspring mice. Cell Res. 26, 645–654. doi: 10.1038/cr.2016.58
Xavier-Neto, J., Carvalho, M., Pascoalino, B. D., Cardoso, A. C., Costa, A. M., Pereira, A. H., et al. (2017). Hydrocephalus and arthrogryposis in an immunocompetent mouse model of ZIKA teratogeny: a developmental study. PLoS Negl. Trop. Dis. 11:e0005363. doi: 10.1371/journal.pntd.0005363
Xu, H. T., Hassounah, S. A., Colby-Germinario, S. P., Oliveira, M., Fogarty, C., Quan, Y., et al. (2017). Purification of Zika virus RNA-dependent RNA polymerase and its use to identify small-molecule Zika inhibitors. J. Antimicrob. Chemother. 72, 727–734. doi: 10.1093/jac/dkw514
Xu, M., Lee, E. M., Wen, Z., Cheng, Y., Huang, W. K., Qian, X., et al. (2016). Identification of small-molecule inhibitors of Zika virus infection and induced neural cell death via a drug repurposing screen. Nat. Med. 22, 1101–1107. doi: 10.1038/nm.4184
Xu, X., Song, H., Qi, J., Liu, Y., Wang, H., Su, C., et al. (2016). Contribution of intertwined loop to membrane association revealed by Zika virus full-length NS1 structure. EMBO J. 35, 2170–2178. doi: 10.15252/embj.201695290
Yockey, L. J., Varela, L., Rakib, T., Khoury-Hanold, W., Fink, S. L., Stutz, B., et al. (2016). Vaginal exposure to Zika virus during pregnancy leads to fetal brain infection. Cell 166, 1247.e4–1256.e4. doi: 10.1016/j.cell.2016.08.004
Zhang, C., Feng, T., Cheng, J., Li, Y., Yin, X., Zeng, W., et al. (2016). Structure of the NS5 methyltransferase from Zika virus and implications in inhibitor design. Biochem. Biophys. Res. Commun. doi: 10.1016/j.bbrc.2016.11.098 [Epub ahead of print].
Zhang, F., Hammack, C., Ogden, S. C., Cheng, Y., Lee, E. M., Wen, Z., et al. (2016). Molecular signatures associated with ZIKV exposure in human cortical neural progenitors. Nucleic Acids Res. 44, 8610–8620. doi: 10.1093/nar/gkw765
Zhao, H., Fernandez, E., Dowd, K. A., Speer, S. D., Platt, D. J., Gorman, M. J., et al. (2016). Structural basis of Zika virus-specific antibody protection. Cell 166, 1016–1027. doi: 10.1016/j.cell.2016.07.020
Zhou, H., Wang, F., Wang, H., Chen, C., Zhang, T., Han, X., et al. (2017). The conformational changes of Zika virus methyltransferase upon converting SAM to SAH. Oncotarget 8, 14830–14834. doi: 10.18632/oncotarget.14780
Zmurko, J., Marques, R. E., Schols, D., Verbeken, E., Kaptein, S. J., and Neyts, J. (2016). The viral polymerase inhibitor 7-Deaza-2’-C-methyladenosine is a potent inhibitor of in vitro Zika virus replication and delays disease progression in a robust mouse infection model. PLoS Negl. Trop. Dis. 10:e0004695. doi: 10.1371/journal.pntd.0004695
Keywords: flavivirus, epidemiology, microcephaly, Guillain–Barré syndrome, antivirals
Citation: Saiz J-C, Martín-Acebes MA, Bueno-Marí R, Salomón OD, Villamil-Jiménez LC, Heukelbach J, Alencar CH, Armstrong PK, Ortiga-Carvalho TM, Mendez-Otero R, Rosado-de-Castro PH and Pimentel-Coelho PM (2017) Zika Virus: What Have We Learnt Since the Start of the Recent Epidemic? Front. Microbiol. 8:1554. doi: 10.3389/fmicb.2017.01554
Received: 23 May 2017; Accepted: 31 July 2017;
Published: 22 August 2017.
Edited by:
Sunil Kumar Lal, Monash University, AustraliaReviewed by:
Haider Abdul-Lateef Mousa, University of Basrah, IraqCopyright © 2017 Saiz, Martín-Acebes, Bueno-Marí, Salomón, Villamil-Jiménez, Heukelbach, Alencar, Armstrong, Ortiga-Carvalho, Mendez-Otero, Rosado-de-Castro and Pimentel-Coelho. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Pedro M. Pimentel-Coelho, cGVkcm9tcGNAYmlvZi51ZnJqLmJy
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.