 Meerambika Mishra
Meerambika Mishra Ananta P. Arukha
Ananta P. Arukha Tufail Bashir3
Tufail Bashir3 Dhananjay Yadav
Dhananjay Yadav G. B. K. S. Prasad
G. B. K. S. Prasad- 1School of Life Sciences, Sambalpur University, Burla, India
- 2Department of Infectious Diseases and Pathology, University of Florida, Gainesville, FL, United States
- 3School of Biotechnology, Yeungnam University, Gyeongsan, South Korea
- 4Department of Medical Biotechnology, Yeungnam University, Gyeongsan, South Korea
- 5School of Biochemistry, Jiwaji University, Gwalior, India
Nature’s silicon marvel, the diatoms have lately astounded the scientific community with its intricate designs and lasting durability. Diatoms are a major group of phytoplanktons involved in the biogeochemical cycling of silica and are virtually inherent in every environment ranging from water to ice to soil. The usage of diatoms has proved prudently cost effective and its handling neither requires costly materials nor sophisticated instruments. Diatoms can easily be acquired from the environment, their culture requires ambient condition and does not involve any costly media or expensive instruments, besides, they can be transported in small quantities and proliferated to a desirable confluence from that scratch, thus are excellent cost effective industrial raw material. Naturally occurring diatom frustules are a source of nanomaterials. Their silica bio-shells have raised curiosity among nanotechnologists who hope that diatoms will facilitate tailoring minuscule structures which are beyond the capabilities of material scientists. Additionally, there is a colossal diversity in the dimensions of diatoms as the frustule shape differs from species to species; this provides a scope for the choice of a particular species of diatom to be tailored to an exacting requisite, thus paving the way to create desired three dimensional nanocomposites. The present article explores the use of diatoms in various arenas of science, may it be in nanotechnology, biotechnology, environmental science, biophysics or biochemistry and summarizes facets of diatom biology under one umbrella. Special emphasis has been given to biosilicification, biomineralization and use of diatoms as nanomaterials’, drug delivery vehicles, optical and immune-biosensors, filters, immunodiagnostics, aquaculture feeds, lab-on-a-chip, metabolites, and biofuels.
Introduction
Diatoms are unicellular algae (∼1–500 mm length) belonging to Class Bacillariophyceae, division Bacillariophyta, either of order centrales or pennales owing to their morphology or habitat. These phytoplanktons are further categorized into centric diatoms (Coscinodiscophyceae), pennate diatoms (Fragilariophyceae; no raphe), and pennate diatoms (Bacillariophyceae; with raphe), they exist either as unicellular or colonies, filaments, ribbons (Fragilaria), fans (Meridion), zigzags (Tabellaria), or stellate (Asterionella). Diatoms are producers within the food chain; globally contributing to almost 25% of primary productivity (Scala and Bowler, 2001). Asexual reproduction in diatoms: cell division produces two daughter cells each inheriting one parental valve, subsequently grows another smaller valve within. Owing to this size reduction division, with every generation the size of the diatom cell reduces but upon reaching a minimal size; they invert the scenario by forming an auxospore which subsequently grows larger and then undergoes size-diminishing divisions.
Diatoms can easily be acquired from the environment and transported in small quantities and proliferated to a desirable confluence. They uptake silicon from the environment and deposit it in their cell walls forming frustules which are intricate, homogenous, regularly spaced, mesoporous, siliceous nanostructures and further allow genetic modification to tailor frustules shape and pore size according to requirement. Diatoms can incorporate desired material into their frustules enhancing their use in making hybrid biosensors, bioreactors and in biotechnology, nanomedicine, photonic devices, and microfluidics. Intact frustules can be obtained from live diatoms with minimal abrasive treatment; these nanomaterials can then be further processed according to their final goal. They have been successfully used as templates for the synthesis of advanced nanostructured bio-hybrids (Nassif and Livage, 2011). Understanding and modifying the processes of biomineralization in diatoms would further accentuate its applicability in nanotechnology.
In this review, attempt to conscientiously compile the multidisciplinary applicability of diatoms in the field of nanotechnology, and biotechnology, especially in biosensor design, drug delivery, immunodiagnostics, metabolite production has been done.
Biosilicification of Diatoms
Nature has blessed diatoms with an innate ability to uptake silicon from the environment and deposit in their cell walls; thereby generating silica shells which pose as nanomaterials with multifaceted applicability. Silicon is absorbed from the surroundings at low concentration (<1 μM) and is actively transported across membranes, as silicic acid through silicic acid transporters (SITs), leading to an internal soluble silicon pool, which subsequently makes insoluble silicon for incorporation into cell walls (Martin-Jézéquel et al., 2000; Knight et al., 2016). The biogenic silica for forming frustules is manufactured intracellularly by the polymerization of silicic acid monomers. Comparatively, low molecular weight amorphous silica is transported to the edge of Silica Deposition Vesicle (SDV) by silica-transport vesicles (STVs). Upon release into interior of the SDV, these particles diffuse till they come across the part of the breeding aggregate, unto which they stick. The surface consists of silanol groups [Si (OH)2 or Si–OH], facilitating them to disseminate over the surface of aggregate in a pH and temperature dependent process called ‘sintering.’ Relocation permits the molecules to restructure themselves to attain a thermodynamic stability, typically resulting in a smoothening of the aggregate surface. Silica structure formation in diatoms is normally categorized into three distinct scales progressing from the nano to the meso and finally to the microscale (Hildebrand et al., 2006, 2007). The microscale is the overall shaping of the valve and girdle bands within the SDV through active and passive molding and involves cytoskeleton, actin, and microtubules (Round et al., 1990; Van De Meene and Pickett-Heaps, 2002; Tesson and Hildebrand, 2010a; Knight et al., 2016). The organic components required for biosilica polymerization (Kröger and Wetherbee, 2000) are LCPAs (long-chain polyamines, a component of biosilica) and silaffins (Kröger et al., 2002; Poulsen and Kröger, 2004; Tesson and Hildebrand, 2010b).
High variability in shell shape from sparse skeletons of criss-crossing bars to barrels, pods, stars, triangles, and elaborate disks that look like flying saucers is evident. During replication, the two diatom halves (epitheca and hypotheca) and girdle bands separate and new ones are synthesized intracellularly inside the SDVs. Girdle bands may be split rings or continuous, encircling the cell or scale-like (Round et al., 1990; Hildebrand et al., 2009). Although the girdle bands are less ornate than valves they still have a structure that appears to be species dependent and are synthesized within SDV (Kröger and Wetherbee, 2000). In centric diatoms (Figure 1), initial valve formation occurs by the deposition of linear ribs that radiate out from the center (Round et al., 1990; Taylor et al., 2007; Hildebrand et al., 2009). Although, the basic ribbed structure of centric diatoms appears to be conserved, that being a reasonably flat ribbed structure radiating out from the center, there are variations in the nanoscale structure.
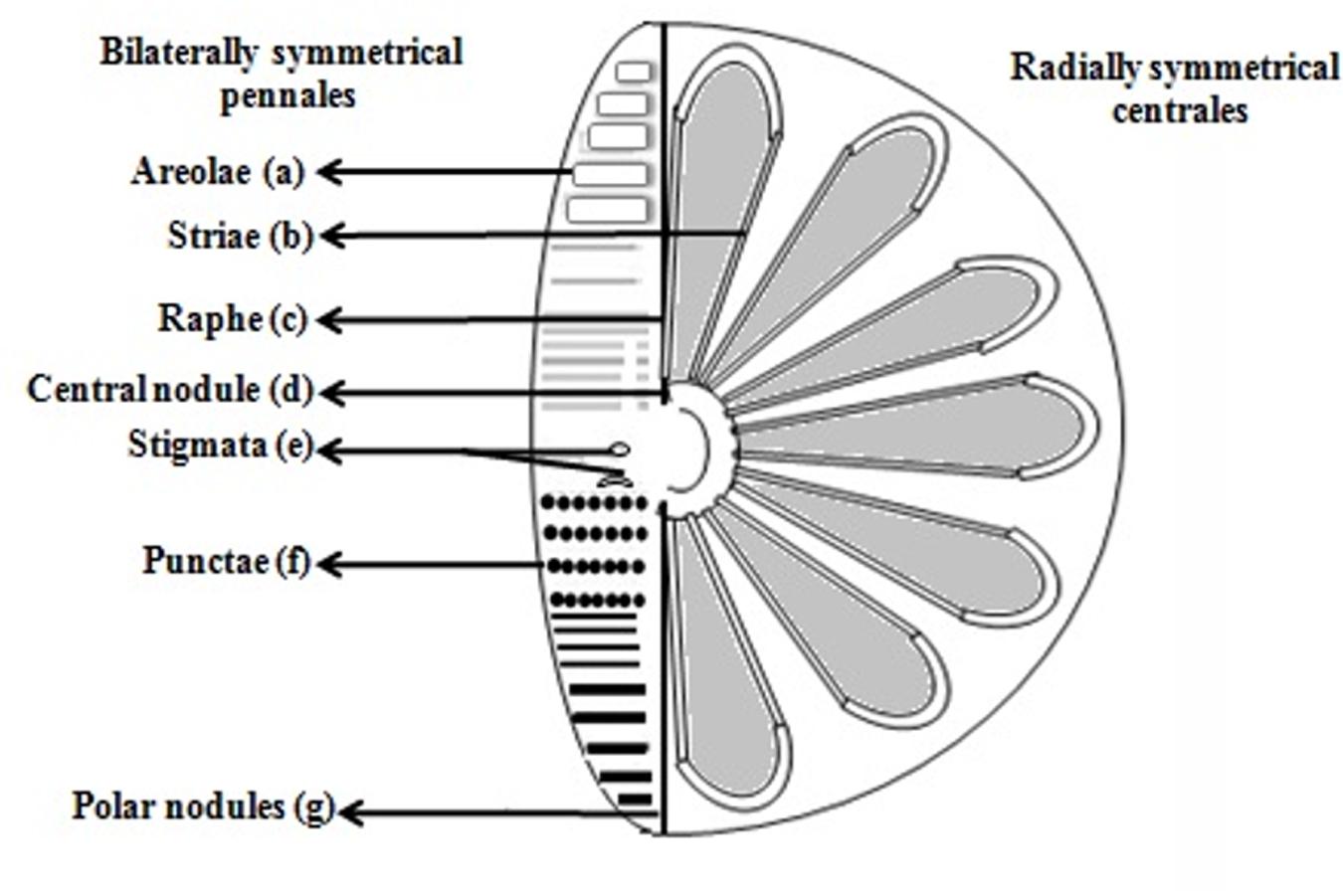
FIGURE 1. The intricate structures of the diatom. Diatom encompasses (a). Areolae (hexagonal or polygonal boxlike perforation with a sieve present on the surface of diatom, b). Striae (pores, punctae, spots or dots in a line on the surface, c). Raphe (slit in the valves, d). Central nodule (thickening of wall at the midpoint of raphe, e). Stigmata (holes through valve surface which looks rounded externally but with a slit like internal, f). Punctae (spots or small perforations on the surface, g). Polar nodules (thickening of wall at the distal ends of the raphe) diagram modified from Taylor et al. (2007).
Multipurpose Uses of Diatoms
Both live diatoms and their modified frustules have innumerable uses. Diatoms have evolved by secondary endocytobiosis, possessing atypical cell biology and genetic makeup. Advances in molecular biology and genetic engineering will unravel usage of diatoms in nanotechnology and biotechnology (Kroth, 2007). In nature, they potently remove carbon-dioxide from the atmosphere and are largely used for environmental reconstruction and audit, forensic investigation of drowning victims and water quality monitoring. The various properties leading to the use of diatoms and their frustules in different areas of technology has been summarized in Table 1.
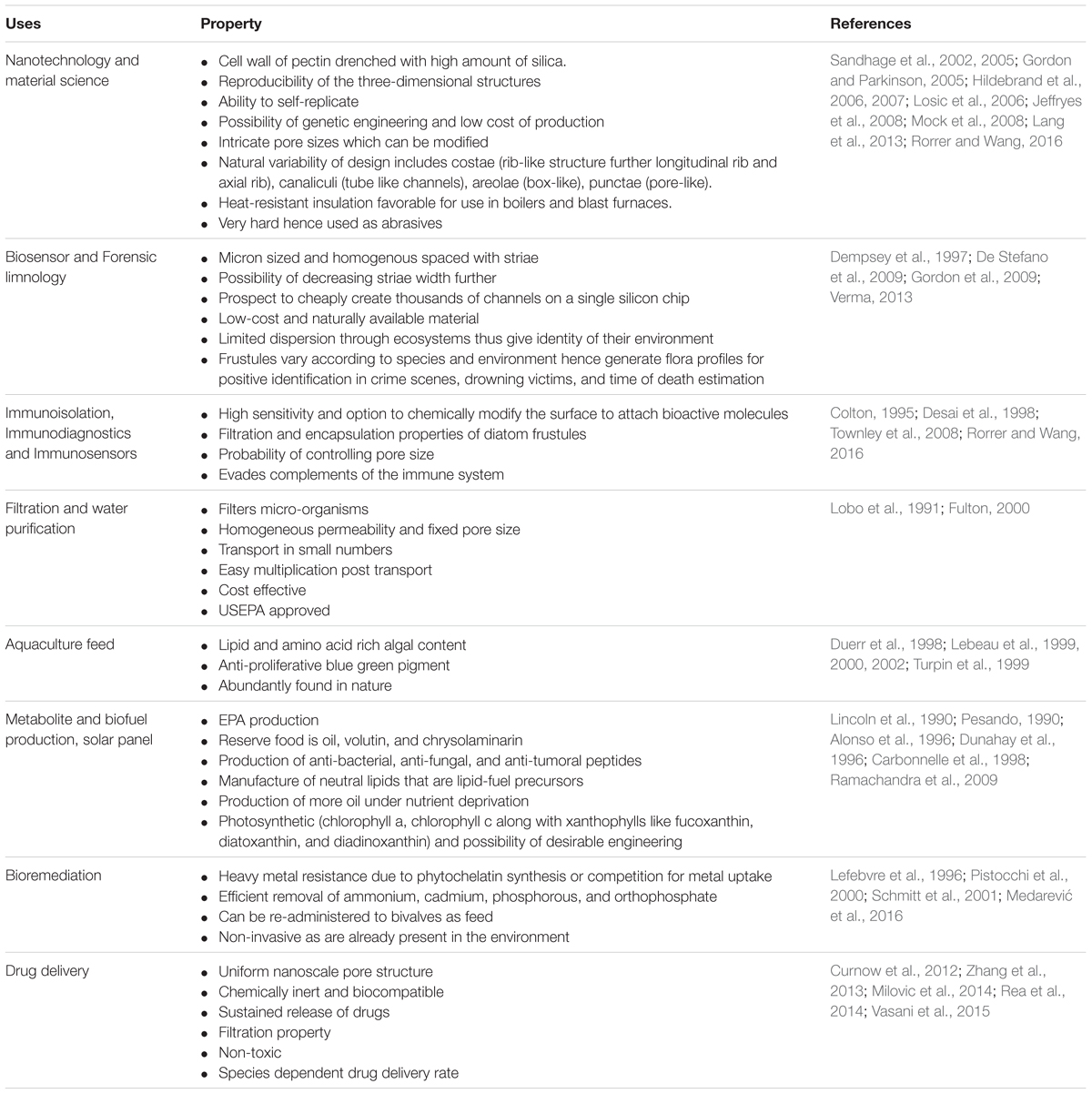
TABLE 1. Properties of diatoms which make them suitable for various uses.
As a Source of Nanomaterials
Diatoms can self-replicate and can further be engineered to provide cost-effective and programmable industrialized system. Efforts to substitute silicon with metal oxides of established optical, electrical, thermal, biological, and chemical properties as germanium, titanium; even zinc have paid off bountifully (Rorrer et al., 2005; Jeffryes et al., 2008; Jaccard et al., 2009). Rorrer et al. (2005) have used diatom to controllably fabricate semiconductor titanium dioxide nanostructured by a bottom-up self-assembly course on a massively parallel scale. They metabolically inserted nano-structured TiO2, forming a nano-composite of titanium and silicon in the diatom Pinnularia sp., by cultivating the diatom in a controlled two-stage bioreactor process. Greatly useful in dye-sensitized solar cells designed for improved light trapping efficiency and structured photocatalysts for the superior breakdown of toxic chemicals. Lang et al. (2013) have used live diatom cells to formulate organo-silica assemblies without any loss in the intricate frustule patterning. Addition of various metals to the already existant silica frustues improves their durability and usability in various nanotechnological purposes.
As Filterant in Water Purification
Diatomaceous earth (DE) is a heterogeneous concoction of the fossil residue of dead diatoms with filtration capability. The use of diatoms over DE is advantageous because; usage of a single culture will ensure homogenous permeability and fixed pore size (Hildebrand, 2008). They can be transported cost-effectively in small numbers and cultured to desired confluence, ideal for industrial processes (Lobo et al., 1991).
As Biodevices
Diatom cells have been grown on self-assembled monolayers. The surface of glass was activated with the addition of trifluoromethyl, methyl, carboxyl, and amino groups by the self-assembled monolayers (SAM) process following which diatom was cultured on the modified glass surface. Upon rinsing post adhesion, diatoms had formed a 2D array, thus aggrandizing their use in bio-devices development (Umemura et al., 2001). Freshwater diatoms have been used to make biosensors for water quality assessment using alternating current dielectrophoresis to chain live diatom cells in order to create a 2D array (Siebman et al., 2017).
Industrial Applications
Metabolite Production
Diatoms are artificially cultivated for their intracellular metabolites like eicosapentaenoic acid (EPA), essential lipids, and amino acids for pharmaceutical and cosmetic purposes (Lebeau and Robert, 2003; Hemaiswarya et al., 2011). Live diatoms as Chaetoceros and Thalassiosira species are used as larval feed (Spolaore et al., 2006), Tetrasel missuecica, Thalassiosira pseudonana, Pavlova lutheri, Isochrysis galbana, and Skeletonema costatum are used to feed bivalve molluscs (Hemaiswarya et al., 2011). The extracellular metabolites are used as chicken and fish feeds. P. tricornutum and Nitzschia laevis have been cultivated in various photobioreactors like perfusion cell bleeding, helical tubular photobioreactor, glass tank and glass tube outdoors photobioreactor for EPA production (Lebeau et al., 2002), used to thwart coronary heart disease, hyper-triglyceridemia, blood platelet aggregation and reduction in blood cholesterol level, preventing risk of arteriosclerosis and inflammation. EPA from more popular sources like fish oil products possess poor taste, instability and higher purification cost (Abedi and Sahari, 2014). Predominantly, Nitzschia inconspicia (1.9–4.7% dw EPA), Nitzschia laevis (2.5–2.76% dw EPA), Navicula saprophila and Phaeodactylum tricornutum (2.2–3.9% dw EPA) are cultured for EPA (Wen and Chen, 2001a,b; Lebeau and Robert, 2003; Abedi and Sahari, 2014; Wah et al., 2015). Nitzschia inconspicia has been reported to produce arachidonic acid around 0.6–4.7% total fatty acids (Chu et al., 1994; Lebeau and Robert, 2003). Aspartic acid and isoleucine are synthesized by Chaetoceros calcitrans and S. costatum, while leucine is synthesized only by C. calcitrans, ornithine by S. costatum, serine, glutamic acid and tyrosine by Thalassiosira (Derrien et al., 1998; Hildebrand et al., 2012). A strong neuroexcitatory adversary of glutamate, domoic acid is also produced by Nitzschia navisvaringica with about 1.7 pg cell-1 (Kotaki et al., 2000; Martin-Jézéquel et al., 2015). Domoic acid is also established as anti-helminthic and insecticidal (Lincoln et al., 1990; Lebeau and Robert, 2003). Antibacterial and antifungal activities of diatoms are attributed to a complex of fatty acids (Pesando, 1990; Thillairajasekar et al., 2009). S. costatum inhibits growth of Vibrio in aquaculture (Naviner et al., 1999). Organic extracts of S. costatum (Bergé et al., 1996) and aqueous extract of Haslea ostrearia (Rowland et al., 2001) are anti-tumoral, effective against human lung cancer and HIV (Hildebrand et al., 2012). A C25 highly branched isoprenoidpolyenes which are polyunsaturated sesterpenes oils or haslenes are responsible for anti-tumoral activities (Lebeau and Robert, 2003; Hildebrand et al., 2012).
Biofuels
Oil as food reserve is produced by diatoms during vegetative phase which keeps them afloat while awaiting favorable conditions. Using these oils glands they also produce neutral lipids which are lipid-fuel precursors; yield a lot more oil than soybean, oil seeds and palm. Ramachandra et al. (2009) professed that diatom substantially produces more oil under stress as lesser silica or nitrogen in the culture. Micro spectrometry comparative analysis of diatom oil compared with known crude oil revealed that the former has 60–70% more saturated fatty acid than the latter. A lion’s share of the existent petrol has arisen from the fossilized diatoms. Diatoms imbibe CO2 and sink on the ocean floor, gets preserved to yield petroleum (Ramachandra et al., 2009; Vinayak et al., 2015).
Ramachandra et al. (2009) also established a time-saving method of producing diatom oil which reduces the production time. They have successfully modified diatom to secrete oil as contrary to storage, which facilitates daily extraction of oil. Diatoms are adhered to a solar panel on an angiosperm leaf wherein the photosynthetic diatom substitutes mesophyll. Thus stomata facilitate gaseous exchange and leaf provides a humid growth environment for diatom while it photosynthesizes. Subsequently, they have genetically engineered diatoms to directly secrete gasoline which averts additional processing (Ramachandra et al., 2009). Diatom fuels may substitute fossil fuels thus substantially reducing greenhouse gases burden. Cyclotella cryptica has been genetically engineered for biodiesel production (Dunahay et al., 1996). Phaeodactylum tricornutum Bohlin UTEX 640 was mutated to exhibit 44% higher EPA production (Alonso et al., 1996; Lebeau and Robert, 2003).
Nanomedicine and Medical Applications
Nanomedicine employs nanomaterials, nanoelectric biosensors and molecular nanotechnology with drug delivery vehicles, diagnostic devices and physical therapy applications being equally pivotal in it. However, the major shortcoming faced by nanomedicine is toxicity, biodegradability, and environmental impact. Using diatoms or their derived frustules instead provides intricate homogeneity while also surpassing the shortcomings as they are non-toxic, biodegradable, and readily available in the environment (Bradbury, 2004; Dolatabadi and de la Guardia, 2011; Jamali et al., 2012; Li et al., 2016).
Biosensors
The striae (Figure 1) in pennales are microscopic and are constantly spaced which can further be decreased using the compustat approach. The possibility of cheaply making such arrays of channels leading to Lab-on-a-chip (numerous channels on a single silicon chip) and the filtration ability of diatoms are favorable for numerous biosensor designs (Dempsey et al., 1997; Gordon et al., 2009; Siebman et al., 2017). These sensitive devices possess a biological molecular recognition constituent allied to a transducer, proficient of inducing a signal relative to the changing concentration of the molecule being sensed (Collings and Caruso, 1997). The flaw in extant biosensors is interference due to clustering of biomolecules in the circumference of the sensor. Frustules can filter; pore size is controllable, thus by incorporating a specific frustule in specific sensing chamber of biosensor, selective trafficking of the molecule can be achieved. Due to their extremely refractive nature, frustules amplify signal and thus can be used as fluorescent probe.
Immunodiagnostics
Immunoisolating bio-encapsulation benefits from the filtration and encapsulation features of frustules. Lately, a biocapsule competent of selectively immune-isolating transplants was fashioned. The researchers used UV lithography, silicon thin film deposition and selective etching techniques (Desai et al., 1998). These capsules are adept in shielding its enclosure from defensive components of the immune system while concomitantly permitting the ample inflow of nutrients and oxygen to the transplanted tissue. Since frustules are naturally mesoporous, they are ideal vehicles for transporting nutrients to the girdled cells. In order to armor the frustules to filter immunoglobulins and complement system apparatus, the pore size is constrained in dimensions (30 nm) impenetrable to C1q and IgM (Colton, 1995). Furthermore, controlling the dimension of the pores, overall dimensions of frustule can also be altered so that hefty biocapsules adroit of enclosing several mammalian cells can be designed.
The diatom frustule can be chemically tailored for artificially tethering antibodies and bioactive molecules to it. The attached antibodies or molecules retain their inherent biological activity. These customized structures are crucial in antibody arrays and also form the basis of immunodiagnostics. As diatom biosilica requires only light and nominal nutrients hence they spawn an outstandingly low-priced and renewable starting matter (Townley et al., 2008).
Optical Biosensors
The frustules of the central Coscinodiscus concinnus Wm. Smith have been chemically modified to bind to an exceedingly selective bio-probe as an antibody. Measuring the photoluminescence emission of these modified diatoms frustules, reveal the degree of antibody–ligand interaction. Diatom frustules are nanostructured, inexpensive, abundantly available naturally and also exhibit extreme sensitivity, therefore, are the ultimate entrant for the lab-on-a-chip applications (De Stefano et al., 2009).
Drug Delivery
Homogenous pore size, constant spacing of striae, hard biosilica, genetically modifiable, chemically inert and biocompatibility are the decisive features facilitating the use of frustules as drug delivery vehicles. Pore size and rate at which the drug would be released from the diatom frustules is species-specific which gives investigators ample choices. Drug-laden diatoms can be directed to the site of release by integrating ferromagnetic elements into the frustules and then using a magnet. Currently, diatom nanotechnology is an exceedingly interdisciplinary yet a rapidly growing research front with extremely divergent applicability (Gordon and Parkinson, 2005). High-resolution imaging techniques establish a baseline for investigating biomineralization in diatoms that ultimately impact device manufacturing capabilities. Zhang et al. (2013) have efficiently used diatom for the oral delivery of drugs for gastrointestinal diseases. Usage of diatom microparticles has no toxicity rather effectively enhanced the permeability of prednisone and mesalamine while also enabling their sustained release. The use of diatom as a solid carrier for BCS Class II drugs notorious for their low water solubility for oral administration through self-emulsifying drug delivery system (SEDDS) has been reported. Two approaches using diverse self-emulsifying phospholipid suspension of carbamazepine (CBZ) first by directly mixing with diatoms, second by dispersing diatoms into its ethanolic preparation was employed. While the physical mixture procedure was more efficient, mixing with the ethanolic extract deemed faster. Both processes, however, showed prolonged longevity (Milovic et al., 2014). Diatom has also been used for transport of siRNA into tumor cells (Rea et al., 2014). Besides, diatom frustules have also been used for antibiotic delivery (Vasani et al., 2015). The genome sequences of two diatom species, Thalassiosira pseudonana and Phaeodactylum tricornutum, has already been deciphered, works on others is in progress (Armbrust et al., 2004; Bowler et al., 2008; Hildebrand et al., 2012) to effectively identify the proteins involved in fabrication of diatom skeleton features enhancing expression or direct production of desired products.
Future Prospects
Diatoms make gargantuan variety of shapes. Some of these structures are dependent on microtubules and possibly are sensitive to microgravity. The NASA Single Loop for Cell Culture (SLCC) for culturing and observing microbes authorizes economical, low labor in-space experiments. Three diatom species were sent to the International Space Station, together with the huge (6 mm length) diatoms of Antarctica and the exclusive colonial diatom, Bacillaria paradoxa. The cells of Bacillaria moved next to each other in partial but opposite synchrony by a microfluidics method. Swift, directed evolution is achievable by using the SLCC as a compustat. Since the structural details are well conserved in hard silica, the development of normal and deviant morphogenesis can be achieved by drying the samples on a moving diatom filter paper. Owing to the massive biodiversity of diatoms, its nanotechnology will present a condensed and portable diatom nanotechnology toolkit for space exploration (Gordon and Parkinson, 2005).
Diatoms pose a novel example of a natural enigma which has been unfolded recently. There are still many unanswered questions, as the equation amid the genotype and phenotype of diatom, its further manipulation without breaking the balance of its 3D shape and pattern, methods of genetic engineering applicable. Other speculations are about the limits for diatom evolution, how can we make the most out of them and in what other fields can diatoms find use. As our comprehension of genetic composition of diatoms gets enlightened, the possibility of designing molecularly explicit architectures of large (mm) and minute (nm) dimensions would be more feasible. Genetically engineered diatoms are employed as vectors for vaccine delivery and used for enhancing the nutritional quality of the feedstuff for crustaceans and aqua-cultured fish, few diatom based vaccines have been successfully used and patented as well (Gladue and Maxey, 1994; Hempel et al., 2011; Corbeil et al., 2015; Doron et al., 2016). Various researches have been structured to find novel diatoms even in unconventional places to decipher these siliceous mysteries (Amspoker, 2016; Noga et al., 2016). The future harbors promising challenges endowed with great rewards for diatomists and nanotechnologists eventually as the research on diatoms gets more illumined.
Author Contributions
All authors listed have made a substantial, direct and intellectual contribution to the work, and approved it for publication.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgment
The authors are thankful to Dr. Nanda Kishore Mishra (Department of English, Gangadhar Meher University, Odisha) for improving the English language quality.
References
Abedi, E., and Sahari, M. A. (2014). Long-chain polyunsaturated fatty acid sources and evaluation of their nutritional and functional properties. Food Sci. Nutr. 2, 443–463. doi: 10.1002/fsn3.121
Alonso, D. L., Segura del Castillo, C. I., Grima, E. M., and Cohen, Z. (1996). First insights into improvement of eicosapentaenoic acid content in Phaeodactylum tricornutum (Bacillariophyceae) by induced mutagenesis. J. Phycol. 32, 339–345. doi: 10.1111/j.0022-3646.1996.00339.x
Amspoker, M. C. (2016). Eunotogramma litorale sp. nov., a marine epipsammic diatom from Southern California, USA. Diatom Res. 31, 389–395. doi: 10.1080/0269249X.2016.1256350
Armbrust, E. V., Berges, J. A., Bowler, C., Green, B. R., Martinez, D., Putnam, N. H., et al. (2004). The genome of the diatom Thalassiosira pseudonana: ecology, evolution, and metabolism. Science 306, 79–86. doi: 10.1126/science.1101156
Bergé, J., Bourgougnon, N., Carbonnelle, D., Le Bert, V., Tomasoni, C., Durand, P., et al. (1996). Antiproliferative effects of an organic extract from the marine diatom Skeletonema costatum (Grev.) Cleve. Against a non-small-cell bronchopulmonary carcinoma line (NSCLC-N6). Anticancer Res. 17, 2115–2120.
Bowler, C., Allen, A. E., Badger, J. H., Grimwood, J., Jabbari, K., Kuo, A., et al. (2008). The Phaeodactylum genome reveals the evolutionary history of diatom genomes. Nature 456, 239–244. doi: 10.1038/nature07410
Bradbury, J. (2004). Nature’s nanotechnologists: unveiling the secrets of diatoms. PLoS Biol. 2:e306. doi: 10.1371/journal.pbio.0020306
Carbonnelle, D., Pondaven, P., Morançais, M., Massé, G., Bosch, S., Jacquot, C., et al. (1998). Antitumor and antiproliferative effects of an aqueous extract from the marine diatom Haslea ostrearia (Simonsen) against solid tumors: lung carcinoma (NSCLC-N6), kidney carcinoma (E39) and melanoma (M96) cell lines. Anticancer Res. 19, 621–624.
Chu, W.-L., Phang, S.-M., and Goh, S.-H. (1994). “Studies on the production of useful chemicals, especially fatty acids in the marine diatom Nitzschia conspicua Grunow,” in Ecology and Conservation of Southeast Asian Marine and Freshwater Environments including Wetlands, eds A. Sasekumar, N. Marshall, and D. J. Macintosh (Berlin: Springer), 33–40.
Collings, A., and Caruso, F. (1997). Biosensors: recent advances. Rep. Prog. Phys. 60, 1397–1445. doi: 10.1088/0034-4885/60/11/005
Colton, C. K. (1995). Implantable biohybrid artificial organs. Cell Transplant. 4, 415–436. doi: 10.1016/0963-6897(95)00025-S
Corbeil, L. B., Hildebrand, M., Shrestha, R., Davis, A., Schrier, R., Oyler, G. A., et al. (2015). Diatom-based vaccines. U.S. Patent 20150037370 A1.
Curnow, P., Senior, L., Knight, M. J., Thamatrakoln, K., Hildebrand, M., and Booth, P. J. (2012). Expression, purification, and reconstitution of a diatom silicon transporter. Biochemistry 51, 3776–3785. doi: 10.1021/bi3000484
De Stefano, L., Rotiroti, L., De Stefano, M., Lamberti, A., Lettieri, S., Setaro, A., et al. (2009). Marine diatoms as optical biosensors. Biosens. Bioelectron. 24, 1580–1584. doi: 10.1016/j.bios.2008.08.016
Dempsey, E., Diamond, D., Smyth, M. R., Urban, G., Jobst, G., Moser, I., et al. (1997). Design and development of a miniaturised total chemical analysis system for on-line lactate and glucose monitoring in biological samples. Anal. Chim. Acta 346, 341–349. doi: 10.1016/S0003-2670(97)90075-1
Derrien, A., Coiffard, L. J., Coiffard, C., and De Roeck-Holtzhauer, Y. (1998). Free amino acid analysis of five microalgae. J. Appl. Phycol. 10, 131–134. doi: 10.1023/A:1008003016458
Desai, T. A., Chu, W. H., Tu, J. K., Beattie, G. M., Hayek, A., and Ferrari, M. (1998). Microfabricated immunoisolating biocapsules. Biotechnol. Bioeng. 57, 118–120. doi: 10.1002/(SICI)1097-0290(19980105)57:1<118::AID-BIT14>3.0.CO;2-G
Dolatabadi, J. E. N., and de la Guardia, M. (2011). Applications of diatoms and silica nanotechnology in biosensing, drug and gene delivery, and formation of complex metal nanostructures. TrAC Trends Anal. Chem. 30, 1538–1548. doi: 10.1016/j.trac.2011.04.015
Doron, L., Segal, N. A., and Shapira, M. (2016). Transgene expression in microalgae—from tools to applications. Front. Plant Sci. 7:505. doi: 10.3389/fpls.2016.00505
Duerr, E. O., Molnar, A., and Sato, V. (1998). Cultured microalgae as aquaculture feeds. J. Mar. Biotechnol. 6, 65–70.
Dunahay, T. G., Jarvis, E. E., Dais, S. S., and Roessler, P. G. (1996). “Manipulation of microalgal lipid production using genetic engineering,” in Proceedings of the Seventeenth Symposium on Biotechnology for Fuels and Chemicals (Berlin: Springer), 223–231. doi: 10.1007/978-1-4612-0223-3_20
Fulton, G. P. (2000). Diatomaceous Earth Filtration for Safe Drinking Water. Reston, VA: American Society of Civil Engineers. doi: 10.1061/9780784404294
Gladue, R. M., and Maxey, J. E. (1994). Microalgal feeds for aquaculture. J. Appl. Phycol. 6, 131–141. doi: 10.1007/BF02186067
Gordon, R., Losic, D., Tiffany, M. A., Nagy, S. S., and Sterrenburg, F. A. (2009). The glass menagerie: diatoms for novel applications in nanotechnology. Trends Biotechnol. 27, 116–127. doi: 10.1016/j.tibtech.2008.11.003
Gordon, R., and Parkinson, J. (2005). Potential roles for diatomists in nanotechnology. J. Nanosci. Nanotechnol. 5, 35–40. doi: 10.1166/jnn.2005.002
Hemaiswarya, S., Raja, R., Kumar, R. R., Ganesan, V., and Anbazhagan, C. (2011). Microalgae: a sustainable feed source for aquaculture. World J. Microbiol. Biotechnol. 27, 1737–1746. doi: 10.1007/s11274-010-0632-z
Hempel, F., Bozarth, A. S., Lindenkamp, N., Klingl, A., Zauner, S., Linne, U., et al. (2011). Microalgae as bioreactors for bioplastic production. Microb. Cell Fact. 10:81. doi: 10.1186/1475-2859-10-81
Hildebrand, M. (2008). Diatoms, biomineralization processes, and genomics. Chem. Rev. 108, 4855–4874. doi: 10.1021/cr078253z
Hildebrand, M., Davis, A. K., Smith, S. R., Traller, J. C., and Abbriano, R. (2012). The place of diatoms in the biofuels industry. Biofuels 3, 221–240. doi: 10.4155/bfs.11.157
Hildebrand, M., Frigeri, L. G., and Davis, A. K. (2007). Synchronized growth of Thalassiosira pseudonana (Bacillariophyceae) provides novel insights into cell-wall synthesis processes in relation to the cell cycle. J Phycol. 43, 730–740. doi: 10.1111/j.1529-8817.2007.00361.x
Hildebrand, M., Kim, S., Shi, D., Scott, K., and Subramaniam, S. (2009). 3D imaging of diatoms with ion-abrasion scanning electron microscopy. J. Struct. Biol. 166, 316–328. doi: 10.1016/j.jsb.2009.02.014
Hildebrand, M., York, E., Kelz, J. I., Davis, A. K., Frigeri, L. G., Allison, D. P., et al. (2006). Nanoscale control of silica morphology and three-dimensional structure during diatom cell wall formation. J. Mater. Res. 21, 2689–2698. doi: 10.1557/jmr.2006.0333
Jaccard, T., Ariztegui, D., and Wilkinson, K. J. (2009). Incorporation of zinc into the frustule of the freshwater diatom Stephanodiscus hantzschii. Chem. Geol. 265, 381–386. doi: 10.1016/j.chemgeo.2009.04.016
Jamali, A. A., Akbari, F., Ghorakhlu, M. M., De La Guardia, M., and Yari Khosroushahi, A. (2012). Applications of diatoms as potential microalgae in nanobiotechnology. Bioimpacts 2, 83–89. doi: 10.5681/bi.2012.012
Jeffryes, C., Gutu, T., Jiao, J., and Rorrer, G. L. (2008). Metabolic insertion of nanostructured TiO2 into the patterned biosilica of the diatom Pinnularia sp. by a two-stage bioreactor cultivation process. ACS Nano 2, 2103–2112. doi: 10.1021/nn800470x
Knight, M. J., Senior, L., Nancolas, B., Ratcliffe, S., and Curnow, P. (2016). Direct evidence of the molecular basis for biological silicon transport. Nat. Commun. 7:11926. doi: 10.1038/ncomms11926
Kotaki, Y., Koike, K., Yoshida, M., Van Thuoc, C., Huyen, N. T. M., Hoi, N. C., et al. (2000). Domoic acid production in Nitzschia sp. (Bacillariophyceae) isolated from a shrimp-culture pond in Do Son, Vietnam. J. Phycol. 36, 1057–1060. doi: 10.1046/j.1529-8817.2000.99209.x
Kröger, N., Lorenz, S., Brunner, E., and Sumper, M. (2002). Self-assembly of highly phosphorylated silaffins and their function in biosilica morphogenesis. Science 298, 584–586. doi: 10.1126/science.1076221
Kröger, N., and Wetherbee, R. (2000). Pleuralins are involved in theca differentiation in the diatom Cylindrotheca fusiformis. Protist 151, 263–273. doi: 10.1078/1434-4610-00024
Kroth, P. G. (2007). Genetic transformation: a tool to study protein targeting in diatoms. Methods Mol. Biol. 390, 257–267. doi: 10.1007/978-1-59745-466-7_17
Lang, Y., Del Monte, F., Collins, L., Rodriguez, B. J., Thompson, K., Dockery, P., et al. (2013). Functionalization of the living diatom Thalassiosira weissflogii with thiol moieties. Nat. Commun. 4:2683. doi: 10.1038/ncomms3683
Lebeau, T., Gaudin, P., Junter, G.-A., Mignot, L., and Robert, J.-M. (2000). Continuous marennin production by agar-entrapped Haslea ostrearia using a tubular photobioreactor with internal illumination. Appl. Microbiol. Biotechnol. 54, 634–640. doi: 10.1007/s002530000380
Lebeau, T., Gaudin, P., Moan, R., and Robert, J.-M. (2002). A new photobioreactor for continuous marennin production with a marine diatom: influence of the light intensity and the immobilised-cell matrix (alginate beads or agar layer). Appl. Microbiol. Biotechnol. 59, 153–159. doi: 10.1007/s00253-002-0993-9
Lebeau, T., Junter, G.-A., Jouenne, T., and Robert, J.-M. (1999). Marennine production by agar-entrapped Haslea ostrearia Simonsen. Bioresour. Technol. 67, 13–17. doi: 10.1016/S0960-8524(99)00096-6
Lebeau, T., and Robert, J. M. (2003). Diatom cultivation and biotechnologically relevant products. Part I: cultivation at various scales. Appl. Microbiol. Biotechnol. 60, 612–623. doi: 10.1007/s00253-002-1176-4
Lefebvre, S., Hussenot, J., and Brossard, N. (1996). Water treatment of land-based fish farm effluents by outdoor culture of marine diatoms. J. Appl. Phycol. 8, 193–200. doi: 10.1007/BF02184971
Li, A., Zhang, W., Ghaffarivardavagh, R., Wang, X., Anderson, S. W., and Zhang, X. (2016). Towards uniformly oriented diatom frustule monolayers: experimental and theoretical analyses. Microsyst. Nanoeng. 2:16064. doi: 10.1038/micronano.2016.64
Lincoln, R. A., Strupinski, K., and Walker, J. M. (1990). Biologically active compounds from diatoms. Diatom Res. 5, 337–349. doi: 10.1080/0269249X.1990.9705124
Lobo, E. A., Oliveira, M. A., Neves, M., and Schuler, S. (1991). Caracterização de ambientes de terras úmidas, no Estado do Rio Grande do Sul, onde ocorrem espécies de anatídeos com valor cinegético. Acta Biol. Leopoldensia 13, 19–60.
Losic, D., Triani, G., Evans, P. J., Atanacio, A., Mitchell, J. G., and Voelcker, N. H. (2006). Controlled pore structure modification of diatoms by atomic layer deposition of TiO2. J. Mater. Chem. 16, 4029–4034. doi: 10.1039/b610188g
Martin-Jézéquel, V., Calu, G., Candela, L., Amzil, Z., Jauffrais, T., Séchet, V., et al. (2015). Effects of organic and inorganic nitrogen on the growth and production of domoic acid by Pseudo-nitzschia multiseries and P. australis (Bacillariophyceae) in Culture. Mar. Drugs 13, 7067–7086. doi: 10.3390/md13127055
Martin-Jézéquel, V., Hildebrand, M., and Brzezinski, M. A. (2000). Silicon metabolism in diatoms: implications for growth. J. Phycol. 36, 821–840. doi: 10.1046/j.1529-8817.2000.00019.x
Medarević, D. P., Lošić, D., and Ibrić, S. R. (2016). Diatoms-nature materials with great potential for bioapplications. Hemijska Industrija 70, 613–627. doi: 10.2298/HEMIND150708069M
Milovic, M., Simovic, S., Losic, D., Dashevskiy, A., and Ibric, S. (2014). Solid self-emulsifying phospholipid suspension (SSEPS) with diatom as a drug carrier. Eur. J. Pharm. Sci. 63, 226–232. doi: 10.1016/j.ejps.2014.07.010
Mock, T., Samanta, M. P., Iverson, V., Berthiaume, C., Robison, M., Holtermann, K., et al. (2008). Whole-genome expression profiling of the marine diatom Thalassiosira pseudonana identifies genes involved in silicon bioprocesses. Proc. Natl. Acad. Sci. U.S.A. 105, 1579–1584. doi: 10.1073/pnas.0707946105
Nassif, N., and Livage, J. (2011). From diatoms to silica-based biohybrids. Chem. Soc. Rev. 40, 849–859. doi: 10.1039/c0cs00122h
Naviner, M., Bergé, J.-P., Durand, P., and Le Bris, H. (1999). Antibacterial activity of the marine diatom Skeletonema costatum against aquacultural pathogens. Aquaculture 174, 15–24. doi: 10.1016/S0044-8486(98)00513-4
Noga, T., Stanek-Tarkowska, J., Kochman-Kędziora, N., Pajączek, A., and Peszek, Ł (2016). The inside of a dam as an unusual habitat for two rare species of Gomphosphenia–G. fontinalis and G. holmquistii. Diatom Res. 31, 379–387. doi: 10.1080/0269249X.2016.1247019
Pesando, D. (1990). “Antibacterial and antifungal activities of marine algae,” in Introduction to Applied Phycology, ed. I. Akatsuka (The Hague: SPB Academic Publishing B.V), 3–26.
Pistocchi, R., Mormile, M., Guerrini, F., Isani, G., and Boni, L. (2000). Increased production of extra-and intracellular metal-ligands in phytoplankton exposed to copper and cadmium. J. Appl. Phycol. 12, 469–477. doi: 10.1023/A:1008162812651
Poulsen, N., and Kröger, N. (2004). Silica morphogenesis by alternative processing of silaffins in the diatom Thalassiosira pseudonana. J. Biol. Chem. 279, 42993–42999. doi: 10.1074/jbc.M407734200
Ramachandra, T. V., Mahapatra, D. M., and Gordon, R. (2009). Milking diatoms for sustainable energy: biochemical engineering versus gasoline-secreting diatom solar panels. Ind. Eng. Chem. Res. 48, 8769–8788. doi: 10.1021/ie900044j
Rea, I., Martucci, N. M., De Stefano, L., Ruggiero, I., Terracciano, M., Dardano, P., et al. (2014). Diatomite biosilica nanocarriers for siRNA transport inside cancer cells. Biochim. Biophys. Acta 1840, 3393–3403. doi: 10.1016/j.bbagen.2014.09.009
Rorrer, G. L., Chang, C.-H., Liu, S.-H., Jeffryes, C., Jiao, J., and Hedberg, J. A. (2005). Biosynthesis of silicon–germanium oxide nanocomposites by the marine diatom Nitzschia frustulum. J. Nanosci. Nanotechnol. 5, 41–49. doi: 10.1166/jnn.2005.005
Rorrer, G. L., and Wang, A. X. (2016). Nanostructured diatom frustule immunosensors. Front. Nanosci. Nanotechnol. 2:128–130. doi: 10.15761/FNN.1000122
Round, F. E., Crawford, R. M., and Mann, D. G. (1990). Diatoms: Biology and Morphology of the Genera. New York, NY: Cambridge University Press.
Rowland, S., Belt, S., Wraige, E., Massé, G., Roussakis, C., and Robert, J.-M. (2001). Effects of temperature on polyunsaturation in cytostatic lipids of Haslea ostrearia. Phytochemistry 56, 597–602. doi: 10.1016/S0031-9422(00)00434-9
Sandhage, K. H., Allan, S. M., Dickerson, M. B., Gaddis, C. S., Shian, S., Weatherspoon, M. R., et al. (2005). Merging biological self-assembly with synthetic chemical tailoring: the potential for 3-D genetically engineered micro/nano-devices (3-D GEMS). Int. J. Appl. Ceram. Technol. 2, 317–326. doi: 10.1111/j.1744-7402.2005.02035.x
Sandhage, K. H., Dickerson, M. B., Huseman, P. M., Caranna, M. A., Clifton, J. D., Bull, T. A., et al. (2002). Novel, bioclastic route to self-assembled, 3D, chemically tailored meso/nanostructures: shape-preserving reactive conversion of biosilica (diatom) microshells. Adv. Mater. 14, 429–433. doi: 10.1002/1521-4095(20020318)14:6<429::AID-ADMA429>3.0.CO;2-C
Scala, S., and Bowler, C. (2001). Molecular insights into the novel aspects of diatom biology. Cell. Mol. Life Sci. 58, 1666–1673. doi: 10.1007/PL00000804
Schmitt, D., Müller, A., Csögör, Z., Frimmel, F. H., and Posten, C. (2001). The adsorption kinetics of metal ions onto different microalgae and siliceous earth. Water Res. 35, 779–785. doi: 10.1016/S0043-1354(00)00317-1
Siebman, C., Velev, O. D., and Slaveykova, V. I. (2017). Alternating current-dielectrophoresis collection and chaining of phytoplankton on chip: comparison of individual species and artificial communities. Biosensors 7:4. doi: 10.3390/bios7010004
Spolaore, P., Joannis-Cassan, C., Duran, E., and Isambert, A. (2006). Commercial applications of microalgae. J. Biosci. Bioeng. 101, 87–96. doi: 10.1263/jbb.101.87
Taylor, J. C., Harding, W. R., and Archibald, C. (2007). An Illustrated Guide to Some Common Diatom Species from South Africa. Gezina: Water Research Commission.
Tesson, B., and Hildebrand, M. (2010a). Dynamics of silica cell wall morphogenesis in the diatom Cyclotella cryptica: substructure formation and the role of microfilaments. J. Struct. Biol. 169, 62–74. doi: 10.1016/j.jsb.2009.08.013
Tesson, B., and Hildebrand, M. (2010b). Extensive and intimate association of the cytoskeleton with forming silica in diatoms: control over patterning on the meso-and micro-scale. PLoS ONE 5:e14300. doi: 10.1371/journal.pone.0014300
Thillairajasekar, K., Duraipandiyan, V., Perumal, P., and Ignacimuthu, S. (2009). Antimicrobial activity of Trichodesmium erythraeum (Ehr)(microalga) from south East coast of Tamil Nadu, India. Int. J. Integr. Biol. 5, 167–170.
Townley, H. E., Parker, A. R., and White-Cooper, H. (2008). Exploitation of diatom frustules for nanotechnology: tethering active biomolecules. Adv. Funct. Mater. 18, 369–374. doi: 10.1002/adfm.200700609
Turpin, V., Robert, J.-M., and Goulletquer, P. (1999). Limiting nutrients of oyster pond seawaters in the Marennes-Oléron region for Haslea ostrearia: applications to the mass production of the diatom in mesocosm experiments. Aquat. Living Resour. 12, 335–342. doi: 10.1016/S0990-7440(99)00114-X
Umemura, K., Ishikawa, M., and Kuroda, R. (2001). Controlled immobilization of DNA molecules using chemical modification of mica surfaces for atomic force microscopy: characterization in air. Anal. Biochem. 290, 232–237. doi: 10.1006/abio.2001.4996
Van De Meene, A. M., and Pickett-Heaps, J. D. (2002). Valve morphogenesis in the centric diatom Proboscia alata Sundstrom. J. Phycol. 38, 351–363. doi: 10.1046/j.1529-8817.2002.01124.x
Vasani, R., Losic, D., Cavallaro, A., and Voelcker, N. (2015). Fabrication of stimulus-responsive diatom biosilica microcapsules for antibiotic drug delivery. J. Mater. Chem. B 3, 4325–4329. doi: 10.1039/C5TB00648A
Verma, K. (2013). Role of diatoms in the world of forensic science. J. Forensic Res. 4:181. doi: 10.4172/2157-7145.1000181
Vinayak, V., Manoylov, K. M., Gateau, H., Blanckaert, V., Hérault, J., Pencréac’h, G., et al. (2015). Diatom milking: a review and new approaches. Mar. Drugs 13, 2629–2665. doi: 10.3390/md13052629
Wah, N. B., Ahmad, A. L. B., Chieh, D. C. J., and Hwai, A. T. S. (2015). Changes in lipid profiles of a tropical benthic diatom in different cultivation temperature. Asian J. Appl. Sci. Eng. 4, 91–101.
Wen, Z.-Y., and Chen, F. (2001a). A perfusion–cell bleeding culture strategy for enhancing the productivity of eicosapentaenoic acid by Nitzschia laevis. Appl. Microbiol. Biotechnol. 57, 316–322.
Wen, Z. Y., and Chen, F. (2001b). Application of statistically-based experimental designs for the optimization of eicosapentaenoic acid production by the diatom Nitzschia laevis. Biotechnol. Bioeng. 75, 159–169.
Zhang, H., Shahbazi, M. A., Makila, E. M., Da Silva, T. H., Reis, R. L., Salonen, J. J., et al. (2013). Diatom silica microparticles for sustained release and permeation enhancement following oral delivery of prednisone and mesalamine. Biomaterials 34, 9210–9219. doi: 10.1016/j.biomaterials.2013.08.035
Keywords: biosensors, diatoms, drug delivery, nanomaterials, nanocomposites, diatom nanotechnology
Citation: Mishra M, Arukha AP, Bashir T, Yadav D and Prasad GBKS (2017) All New Faces of Diatoms: Potential Source of Nanomaterials and Beyond. Front. Microbiol. 8:1239. doi: 10.3389/fmicb.2017.01239
Received: 29 March 2017; Accepted: 19 June 2017;
Published: 05 July 2017.
Edited by:
Spiros Paramithiotis, Agricultural University of Athens, GreeceReviewed by:
Qazi Mohd Rizwanul Haq, Jamia Millia Islamia, IndiaAbdul Qader Abbady, Department of Molecular Biology and Biotechnology, Atomic Energy Commission of Syria, Syria
Copyright © 2017 Mishra, Arukha, Bashir, Yadav and Prasad. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: G. B. K. S. Prasad, Z2Jrc3ByYXNhZEBnbWFpbC5jb20= Dhananjay Yadav, ZGhhbnlhZGF2MTY0ODFAZ21haWwuY29t