
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Microbiol. , 13 February 2017
Sec. Evolutionary and Genomic Microbiology
Volume 8 - 2017 | https://doi.org/10.3389/fmicb.2017.00202
This article is part of the Research Topic Planctomycetes-Verrucomicrobia-Chlamydiae bacterial superphylum: New model organisms! View all 14 articles
 Patrick Rast1
Patrick Rast1 Ines Glöckner2Christian Boedeker1Olga Jeske1Sandra Wiegand1Richard Reinhardt3
Ines Glöckner2Christian Boedeker1Olga Jeske1Sandra Wiegand1Richard Reinhardt3 Peter Schumann4
Peter Schumann4 Manfred Rohde5
Manfred Rohde5 Stefan Spring6Frank O. Glöckner7
Stefan Spring6Frank O. Glöckner7 Christian Jogler1,8*
Christian Jogler1,8* Mareike Jogler1*
Mareike Jogler1*The cell wall of free-living bacteria consists of peptidoglycan (PG) and is critical for maintenance of shape as dissolved solutes cause osmotic pressure and challenge cell integrity. Surprisingly, the subdivision 4 of the phylum Verrucomicrobia appears to be exceptional in this respect. Organisms of this subdivision are described to be devoid of muramic or diaminopimelic acid (DAP), usually found as components of PG in bacterial cell walls. Here we describe three novel bacterial strains from a freshwater lake, IG15T, IG16bT, and IG31T, belonging to a new genus in the subdivision 4 of Verrucomicrobia which we found to possess PG as part of their cell walls. Biochemical analysis revealed the presence of DAP not only in these novel strains, but also in Opitutus terrae PB90-1T, the closest described relative of strains IG15T, IG16bT, and IG31T. Furthermore, we found that nearly all genes necessary for peptidoglycan synthesis are present in genomes of subdivision 4 members, as well as in the complete genome sequence of strain IG16bT. In addition, we isolated and visualized PG-sacculi for strain IG16bT. Thus, our results challenge the concept of peptidoglycan-less free-living bacteria. Our polyphasic taxonomy approach places the novel strains in a new genus within the family Opitutaceae, for which the name Lacunisphaera gen. nov. is proposed. Strain designations for IG15T, IG16bT and IG31T are Lacunisphaera parvula sp. nov. (=DSM 26814 = LMG 29468), L. limnophila sp. nov. (=DSM 26815 = LMG 29469) and L. anatis sp. nov. (=DSM 103142 = LMG 29578) respectively, with L. limnophila IG16bT being the type species of the genus.
In aquatic environments, abiotic factors such as salinity and temperature, but also intrinsic metabolism-related mechanisms challenge the cellular integrity of microorganisms and their ability to proliferate. Protective elements may be of a structural nature, such as S-layers, or the avoidance of osmotic stress by living in dependency of host organisms which provide stable conditions for survival (Miles, 1992; Engelhardt, 2007). Members of the class Mollicutes for example lack a peptidoglycan cell wall (Razin, 2006), are osmotically fragile and exhibit pleomorphism (Miles, 1992). Thus, they depend on an eukaryotic host to provide an osmotically stable environment for living.
On the other hand, free-living bacteria usually possess cell wall structures including three dimensionally cross-linked polymeric glycan strands, interconnected by short peptide elements, a structure commonly known as peptidoglycan (PG) to protect cellular integrity. Among bacteria only few exceptions are described while all controversy discussed species belong to the Planctomycetes-Verrucomicrobia-Chlamydiae (PVC) superphylum (Wagner and Horn, 2006). In many respects, this PVC-superphylum seems to challenge our concept of the prokaryotic cell (Lee et al., 2009; Fuerst and Sagulenko, 2011; Jacquier et al., 2015; Rivas-Marín et al., 2016). In particular, the suggested absence of PG in Planctomycetes (König et al., 1984), Chlamydia (Fox et al., 1990) and subdivision 4 Verrucomicrobia (Yoon, 2011) is remarkable. While the assumed lack of PG seems to be associated with the lack of the otherwise universal bacterial cell division protein FtsZ in Planctomycetes (Pilhofer et al., 2008; Jogler et al., 2012) and Chlamydia (Stephens et al., 1998), subdivision 4 Verrucomicrobia encode the tubulin homolog FtsZ (Pilhofer et al., 2008). However, Planctomycetes were recently found to possess a PG cell wall (Jeske et al., 2015; van Teeseling et al., 2015). For Chlamydia, the existence of PG was demonstrated but a canonical PG sacculus was not isolated (Liechti et al., 2014; Packiam et al., 2015). However, for some other members of the phylum Chlamydiae a PG sacculus was identified (Pilhofer et al., 2013). Chlamydia are obligate intracellular pathogens (Jacquier et al., 2015) and thus dwell in an environment isotonic to their cytoplasm, they do not necessarily require a peptidoglycan sacculus to maintain cell shape. Accordingly, recent evidence suggests that PG forms an MreB regulated ring at mid-cell to allow cell division in pathogenic Chlamydia (Liechti et al., 2016). In contrast a typical bacterial sacculus was reported for the free-living Planctomycetes that have to withstand various osmotic challenges in their natural habitats (Jeske et al., 2015; van Teeseling et al., 2015), while free-living bacteria of the verrucomicrobial subdivision 4 are still considered to lack a PG sacculus. This bacterial group belongs to the phylum Verrucomicrobia which is divided into six so-called subdivisions. Thus far, cultured representatives are available for subdivisions 1–4. Recently for subdivision 5 the new Phylum Kiritimatiellaeota was proposed, with one characterized isolate (Spring et al., 2016). Playing a crucial role in environmental nutrient cycles, members of the Verrucomicrobia have not only been found to degrade a variety of complex polymeric compounds in, e.g., soil communities (Wang et al., 2014, 2015), some were also identified as methanotrophs (Sharp et al., 2013; van Teeseling et al., 2014). Increasing efforts to extend the knowledge about this environmentally important phylum have led to the successful isolation and description of several new species in recent years (Lee et al., 2014; Kim et al., 2015). However, the majority of new strains brought into pure culture is affiliated with subdivision 1. Therefore, the scarce data existing to date leaves inconclusive results about the suspected peptidoglycan anomaly of subdivision 4 Verrucomicrobia. Furthermore, thus far only two genomes from validly described species (Opitutus terrae and Coraliomargarita akajimensis) are available. Both genomes were not yet analyzed for PG related genes with state-of-the-art bioinformatic methods (Jeske et al., 2015). Some members of this subdivision have been found to be resistant to various β-lactam antibiotics, indicating either absence of PG or an resistance mechanism such as β-lactamases. For other strains the presence of typical cellular PG building blocks was not investigated at the time of their description (Shieh and Jean, 1998; Choo et al., 2007), leaving open the question whether peptidoglycan exists in verrucomicrobial subdivision 4. Members of this subdivision have been isolated from soil communities and leafs, while most strains originate from aquatic habitats, including freshwater lakes, marine waters and extreme habitats such as hot springs (Shieh and Jean, 1998; Chin et al., 2001; Choo et al., 2007; Yoon et al., 2007c, 2010). Here we describe the targeted isolation of subdivision 4 Verrucomicrobia, using antibiotic agents as selective markers for β-lactam resistant bacteria. Our strategy led to the successful cultivation of three novel strains from surface fresh water samples. By biochemical, microscopic and computational analysis we found that the novel and previously reported members of the verrucomicrobial subdivision 4 possess PG as part of their cell walls.
Our findings challenge the proposed absence of peptidoglycan among subdivision 4 Verrucomicrobia, while at the same time extending the scarce pool of cultivated species in this environmentally important phylum.
Surface freshwater samples were collected in triplicates from a local pond (52° 9′ 38″ N, 10° 32′ 40″ E, Wolfenbüttel, Germany) on August 30th, 2012 after the observation of a massive cyanobacterial blooming event. Water was collected in sterile polypropylene bottles, immediately transferred to the laboratory, homogenized and processed within 2 h.
Cultivation medium M1H was prepared with double distilled water containing 0.25 g/l peptone (BactoTM), 0.25 g/l yeast extract (BactoTM), 2.38 g/l HEPES (Serva), 20 ml/l mineral salt solution and a pH adjusted to 8.0 with 5 M KOH. After sterilization, the medium was complemented with 10 ml/l of a 2.5% glucose solution, 5 ml/l double concentrated vitamin solution, 1 ml/l of 100 mg/ml carbenicillin and 20 mg/ml cycloheximide stock solutions, respectively. Solid medium was prepared with three times washed 12 g/l agar (BactoTM) and cooled to 55°C prior to the addition of heat sensitive solutions. Both, mineral salt solution and double concentrated vitamin solution were prepared according to DSMZ medium 621, while metal salts solution consisted of 250 mg/l Na–EDTA, 1095 mg/l ZnSO4.7H2O, 500 mg/l FeSO4.7H2O, 154 mg/l MnSO4.H2O, 39.5 mg/l CuSO4.7H2O, 20.3 mg/l CoCl2.6H2O, and 17.7 mg/l Na2B4O7.10H2O of which 50 ml were added per liter of mineral salt solution.
For initial bacterial isolation, solid M1H medium was supplemented with 100 μl of carbenicillin stock solution (100 mg/ml), dried for 30 min and inoculated with 100 μl homogenized sample material per plate in a 10–10-2 dilution series and incubated at 20°C in the dark until colony formation became visible. Single colonies were inoculated on fresh solid medium with respective antibiotics. Pure cultures were cryopreserved in M1H medium supplemented with 50% glycerol or 5% DMSO and stored at -80°C. Strains isolated and later identified as members of the verrucomicrobial subdivision 4 were designated IG15T, IG16bT and IG31T. Unless otherwise indicated, verrucomicrobial strains were cultivated at 28°C to ensure reproducibility of cultivation dependent experiments.
Cultivation medium of thin layer chromatography (TLC) reference strains, Bacillus subtilis DSM 10 and Escherichia coli DSM 498, was standard LB medium contained 10 g/l tryptone, 10g/l sodium chloride and 5 g/l yeast extract at pH 7.0 (Bertani, 1951).
For O. terrae PB90-1T cultivation was performed following the recommendations of the Leibniz Institute DSMZ (DSMZ medium no. 295).
Novel isolates were identified by direct sequencing of the 16S rRNA gene after amplification with the optimized universal primers 8f (5′–AGA GTT TGA TCM TGG CTC AG–3′) and 1492r (5′–GGY TAC CTT GTT ACG ACT T–3′) modified from (Lane, 1991). PCR reactions were performed directly on single colonies for identification or liquid cultures to check for purity, using the Taq DNA Polymerase Kit (Qiagen) with one reaction of 25 μl containing 11 μl PCR–grade H2O, 2.5 μl 10x CoralLoad buffer, 2.5 μl Q-Solution, 0.5 μl dNTPs (10 mM each), 1 μl sterile bovine serum albumin solution (20 mg/ml), 0.5 μl MgCl2 solution (25 mM), 0.125 μl Taq–Polymerase (1 U/μl) and 1 μl of each primer (10 pmol). The employed protocol consisted of two steps, the first step with an initial denaturation at 94°C, 5 min, 10 cycles of denaturation at 94°C, 30 s, annealing at 59°C, 30 s, elongation at 72°C, 1 min, followed by the second step with 20 cycles denaturation at 94°C, 30 s, annealing at 54°C, 30 s, elongation at 72°C, 1 min and a final elongation step at 72°C, 7 min. All PCRs were carried out in an Applied Biosystems® Veriti® thermal cycler (Thermo Fisher Scientific) and PCR products were stored at 4°C until Sanger sequencing.
To generate near full length 16S sequences, additional primers (compare Supplementary Table S1) were used for sequencing and assembly of the resulting sequences was performed with the ContigExpress application of the Vector NTI® Advance 10 software (Thermo Fisher Scientific).
Alignment of near full length 16S rRNA sequences was performed using the SINA web aligner (Pruesse et al., 2012), corrected manually and used for phylogenetic tree reconstruction. Tree reconstruction was performed with the ARB software package (Ludwig et al., 2004) using the Maximum Likelihood RAxML module and rate distribution model GTR GAMMA running the rapid bootstrap analysis algorithm, the Neighbor Joining tool with Felsenstein correction for DNA and Maximum Parsimony method employing the Phylip DNAPARS module. Bootstrap values for all three methods were computed with 1,000 resamplings including the E. coli 16S rRNA gene positions 101–1,371. The analysis involved 68 nucleotide sequences of described type strains and uncultured clones, related to the novel strains (compare Supplementary Table S2). 16S rRNA gene identity values of novel isolates and related type strains were calculated using neighbor joining clustering of the ARB package.
Bacterial cells were immobilized on a 1% agarose–pad in MatTek 35 mm glass-bottom dishes and imaged under phase–contrast illumination using a Nikon Eclipse Ti invers microscope at 100× magnification and the Nikon DS–Ri2 camera. To determine the cell size of the novel strains, 100 individual cells of each strain were measured using the NIS-Elements software V4.3 (Nikon Instruments).
For field emission scanning electron microscopy (FESEM) bacteria were fixed in 1% formaldehyde in HEPES buffer (3 mM HEPES, 0.3 mM CaCl2, 0.3 mM MgCl2, 2.7 mM sucrose, pH 6.9) for 1 h on ice and washed one time with HEPES buffer. Cover slips with a diameter of 12 mm were coated with a poly-L-lysine solution (Sigma–Aldrich) for 10 min, washed in distilled water and air-dried. 50 μl of the fixed bacteria solution was placed on a cover slip and allowed to settle for 10 min. Cover slips were then fixed in 1% glutaraldehyde in TE buffer (20 mM TRIS, 1 mM EDTA, pH 6,9) for 5 min at room temperature and subsequently washed twice with TE–buffer before dehydrating in a graded series of acetone (10, 30, 50, 70, 90, and 100%) on ice for 10 min at each concentration. Samples from the 100% acetone step were brought to room temperature before placing them in fresh 100% acetone. Samples were then subjected to critical-point drying with liquid CO2 (CPD 300, Leica). Dried samples were covered with a gold/palladium (80/20) film by sputter coating (SCD 500, Bal–Tec) before examination in a field emission scanning electron microscope (Zeiss Merlin) using the Everhart Thornley HESE2–detector and the inlens SE–detector in a 25:75 ratio at an acceleration voltage of 5 kV.
Temperature optima of the novel isolates were determined by optical density measurements of growing cultures at 600 nm (OD600nm). Strains were inoculated 1:10 from early stationary phase cultures in glass tubes with M1H medium and incubated under constant agitation in temperature controlled shakers (for exact temperatures tested, compare Supplementary Figure S1). Measurements were performed in triplicates and each tube served as its own blank prior to inoculation. Resulting growth curves were analyzed by plotting change of OD600nm during exponential growth phases (slope values), of each individual temperature against temperature values in °C.
To determine the pH optimum, M1H medium was buffered to pH values of 5.0, 6.0, 6.5, 7.0, 7.5, 8.0, 8.5, 9.0, and 10.0 using 10 mM MES, HEPES, HEPPS and CHES buffers, corresponding to their individual buffer range. OD600nm was determined in glass tubes, incubated at 28°C, with three replicates as measure of growth. Catalase activity was determined by bubble formation with fresh 3% H2O2 solution. Cytochrom oxidase activity was determined using Bactident® Oxidase test stripes (Merck Millipore) following the manufacturer’s instructions. Gram properties were determined by reaction of fresh biomass with fresh 3% KOH solution (Suslow et al., 1982).
Substrate utilization of the isolated strains was investigated using the Biolog GN2 MicroLogTM test panel for Gram-negative bacteria. Sterile glass tubes were prepared in duplicates with a basic medium mixture containing 15.7 ml IF-0a inoculation fluid (Biolog), 160 μl of 1 M HEPES buffer (pH 8.0) and 80 μl double concentrated vitamin solution. Tubes were inoculated with bacterial colony material from exponentially growing cultures to a turbidity of 56–68%. Two individual plates per strain were evaluated. To enable the comparison of the derived data, the data of each single experiment were normalized to 100. Only values corresponding to >25% utilization were considered as positive. The heat map graphic was obtained in the R environment (R Core Team, 2015) by using the heatmap.2() function of the gplots package.
Biomass of the isolated strains was obtained from liquid cultures grown in M1H medium at 28°C until stationary phase. The obtained biomasses were stored at -20°C. For fatty acid analysis, 30 mg of lyophilized biomass was processed according to the standards of the Identification Service of the German Collection of Microorganisms and Cell Cultures (DSMZ) (Miller, 1982; Kuykendall et al., 1988).
Strains were grown in liquid culture to stationary phase and biomass was obtained by centrifugation. For strains IG15T and IG31T, the molar G + C content was determined by the service facilities of the DSMZ. In brief, genomic DNA is isolated (Cashion et al., 1977), hydrolyzed, dephosphorylized (Mesbah et al., 1989) and analyzed by HPLC (Tamaoka and Komagata, 1984) in comparison to DNA standards from organisms with published genome sequences and a G + C content range from 43 to 72 mol%. G + C content of strain IG16bT was determined during genome sequencing with the Pacific Bioscience sequencer.
Tolerance of IG15T and IG16bT toward β-lactam antibiotic agents was investigated in a treatment assay using carbenicillin. Strains were inoculated as triplicates 1:10 in glass tubes with M1H medium and final concentrations of 0, 500, 1000, or 2000 mg/l carbenicillin were added. Tubes were incubated at 28°C and growth was measured as change in optical density at 600 nm. After 120 h of incubation, cell viability was investigated by FESEM and cell numbers per ml were calculated by counting with a Neubauer chamber.
To obtain high molecular weight DNA of strain IG16bT, nucleic acid was extracted from whole-cells using a tweaked Genomic DNA kit protocol with Genomic tips 100/G (Qiagen). The protocol was performed as recommended by the manufacturer with one exception: incubation time with digestive enzymes was prolonged to an overnight step to ensure complete lysis of bacterial cells. An aliquot of the extracted DNA was used to prepare 16S rRNA clone libraries (Zero Blunt® PCR Cloning kit; Invitrogen) and resulting clones were sequenced to ensure purity of the extracted DNA.
De novo genome sequencing of strain IG16bT was performed using a PacBio RS sequencer. Single molecule real-time (SMRT) bellTM libraries (Pacific Bioscience) were prepared using ∼10 μg genomic DNA. Sequencing data was processed and assembled using the SMRT analysis software. The closed and complete chromosome of strain IG16bT was annotated using the Prokka annotation tool (Seemann, 2014) and subjected to analysis for putative genomic islands and phage regions using IslandViewer3 (Dhillon et al., 2015) and PHAST (Zhou et al., 2011), respectively. The verrucomicrobial genomes for the gene content analysis were derived from NCBI and IMG (Markowitz et al., 2012) in April 2016 and had to match the following criteria upon CheckM analysis (Parks et al., 2015): completeness > 90, contamination < 5 and strain heterogeneity < 20. Orthologs were detected by Proteinortho (Lechner et al., 2011), a tool that identifies the reciprocal best hits from the given protein sequences. The genome plot was then generated with BRIG (Alikhan et al., 2011).
The presence of peptidoglycan synthesis genes was analyzed using blastp (Altschul et al., 1997), while protein sequences of Phycisphaera mikurensis FYK2301M01T or Gimesia maris 534-30T served as query and were compared with protein sequences encoded in the genomes of O. terrae PB90-1T, C. akajimensis 04OKA010-24T and strain IG16bT. β-lactamase encoding genes were detected in IG16, O. terrae and C. akajimensis as previosly described (Bush, 2013; Jeske et al., 2015). For both analysis, homologous proteins required an identity >30%, an e-value lower than 1e-6 and a conserved domain architecture.
Susceptibility to lysozyme was investigated by incubation of the novel strains in M1H medium. Since strain IG31T showed no lysis after 24 h in M1H medium, osmotic stress was increased by incubation of cells in ddH2O (negative controls as well as lysozyme treated cells). Lysozyme was added to a final concentration of 10 mg/ml and cells were incubated for up to 24 h at 37°C under constant agitation at 300 rpm. Bacterial cells were immobilized on a 1% agarose–pad in MatTek 35 mm glass–bottom dishes and imaged under phase–contrast illumination using a Nikon Eclipse Ti invers microscope at 100× magnification and the Nikon DS–Ri2 camera (Nikon Instruments). Cell viability was checked by microscopy after 1, 3, 6, and 24 h of incubation in M1H medium or ddH2O until cell lysis was observed.
The presence of diaminopimelic acid (DAP) was investigated employing thin-layer chromatography and gas chromatography/mass spectrometry (GC/MS). Thin-layer chromatography of whole-cell hydrolysates of strains IG15T, IG16bT, IG31T as well as reference strains B. subtilis DSM 10 and E. coli DSM 498 was performed as previously described (Staneck and Roberts, 1974). Novel isolates were grown in M1H medium at 28°C to stationary phase and cells were harvested by centrifugation. B. subtilis and E. coli served as organismic controls, grown in 50 ml LB medium at 37°C overnight and harvested by centrifugation, while a mixture of purified DAP isomers (Sigma) was used as detection standard.
Whole-cell hydrolysates of strains IG15T, IG16bT, IG31T as well as of O. terrae PB90-1T (DSM 11246) were analyzed using a gas chromatography/mass spectrometry (GS/MS)-based method (Schumann, 2011), previously employed to quantify the peptidoglycan marker DAP and in addition ornithine in a new proposed Verrucomicrobia related phylum (Spring et al., 2016). In brief, cell pellets were obtained from liquid cultures (grown as described above) and biomass was lyophilized. Samples were standardized for the quantification of diagnostic diamino acids by supplementing lyophilized biomass with 2 μmol of norleucine as internal standard. The hydrolysates (200 μl 4N HCl, 100°C, 16 h) of the samples were dried in a vacuum desiccator. Amino acids derivatized to N-heptafluorobutyryl isobutylesters and were resolved in ethyl acetate and analyzed by GC/MS (Singlequad 320, Varian; electron impact ionization, scan range 60 to 800 m/z). The DAP derivative was detected in extracted ion chromatograms using the characteristic fragment ion set 380, 324, 306, and 278 m/z at a retention time of 22.17 min. A fragment ion of 266 m/z with a retention time of 15.13 min was indicative of the presence of ornithine.
Cells of IG16bT were harvested from 2 l of stationary phase cultures grown in M1H medium at 28°C, by centrifugation at room temperature following a protocol established by van Teeseling et al. (2015). In brief, cells were boiled at 100°C for 1 h with 4% SDS, while being gently mixed by inverting the reaction tube several times in 15 min intervals. Lysates were transferred to Float-a-Lyzer® dialysis tubes (SpectrumLabs, DG Breda, Netherlands) and dialyzed against deionized water in a five-liter beaker over the course of 3 days (water was exchanged two times). Dialyzed samples were stored at RT until analysis by transmission electron microscopy (TEM).
Thin carbon support films were prepared by sublimation of a carbon thread onto a freshly cleaved mica surface. Lysate containing the sacculi was adsorbed onto a carbon film for 1 min and negatively stained with 1% (w/v) aqueous uranyl acetate, pH 5.0 (Valentine et al., 1968). After air-drying, samples were examined in a TEM 910 transmission electron microscope (Carl Zeiss, Oberkochen, Germany) at an acceleration voltage of 80 kV and calibrated magnifications using a line replica. Images were recorded digitally with a Slow-Scan CCD-Camera (ProScan, 1024x1024, Scheuring, Germany) with ITEM-Software (Olympus Soft Imaging Solutions, Münster, Germany).
Near full-length sequences of the 16S ribosomal RNA genes as well as the complete genome sequence of strain IG16bT were deposited with the National Center for Biotechnology Information (NCBI) and are available under KX058881 (IG15T), KX058882 (IG16bT), KX058883 (IG31T) and CP016094 (IG16bT whole genome).
Surface water samples from a local duck pond were used for the targeted isolation of novel subdivision 4 Verrucomicrobia. Given that members of subdivision 4 were thought to lack peptidoglycan, β-lactam antibiotics were used as selection pressure to enrich target bacteria. Obtained colonies of β-lactam resistant bacteria were screened by 16S rRNA gene sequencing analysis and three isolates were identified as members of the verrucomicrobial subdivision 4. Phylogenetic tree reconstruction based on near full-length 16S rRNA gene sequences (Figure 1) revealed that strains IG15T, IG16bT, and IG31T belong to the family of Opitutaceae, sharing 92.21, 92.39, and 92.90% sequence identity with the closest related species O. terrae PB90-1T, respectively (Table 1). Based on recent threshold values for 16S rRNA gene sequence comparison (Rosselló-Móra and Amann, 2015), the novel strains represent three distinct species that form a novel genus within the family Opitutaceae, with IG15T, IG16bT, and IG31T being the type strains.
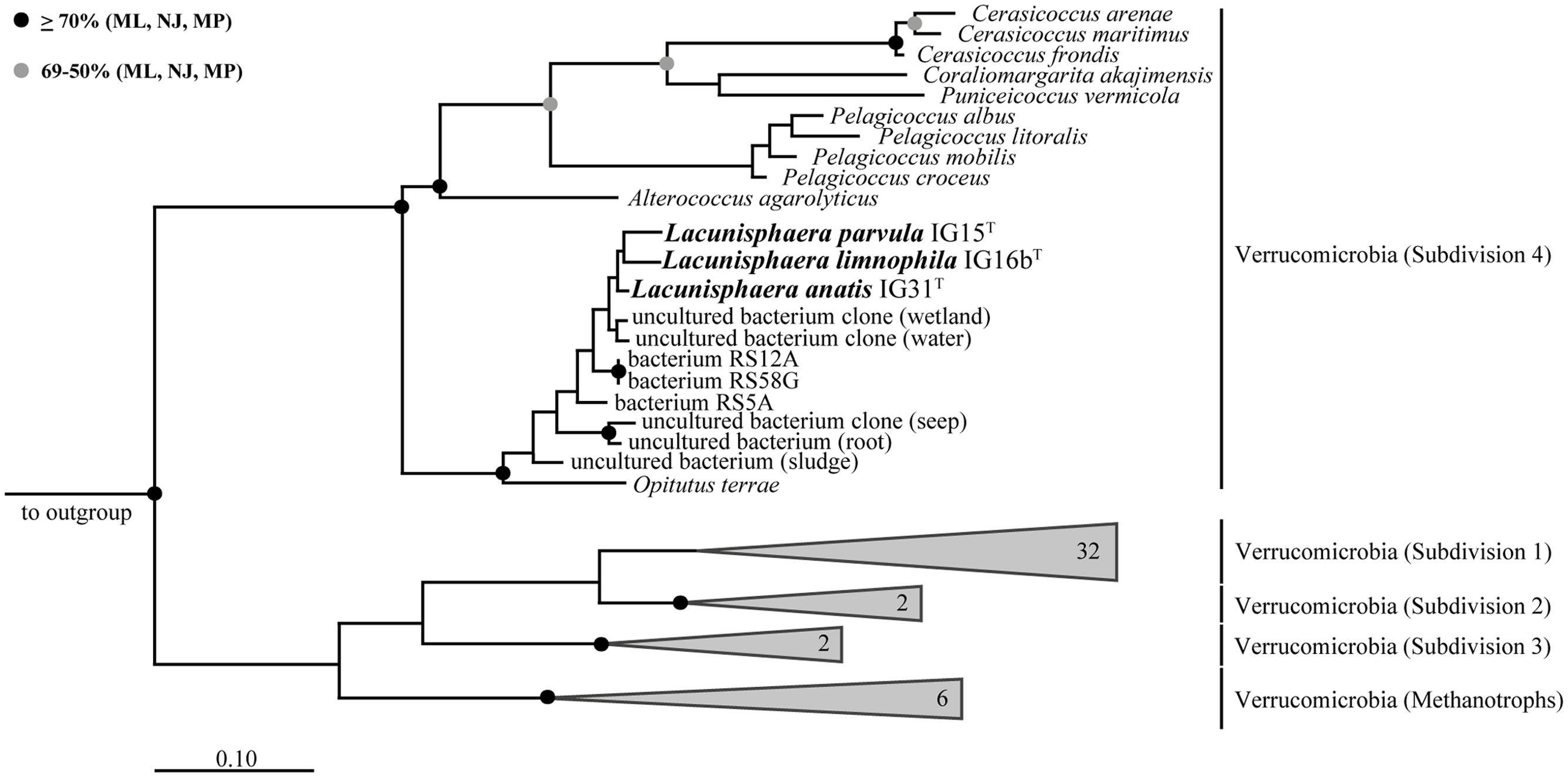
FIGURE 1. Maximum likelihood 16S rRNA gene-based phylogenetic tree. All three strains cluster within the subdivision 4 of Verrucomicrobia with Opitutus terrae as the closest representative with validly published name. Related sequences of uncultured bacteria are shown for comparison and pronounce the distinct phylogenetic position of strains IG15T, IG16bT, and IG31T. Bootstrap values based on three different tree building methods (Maximum Likelihood: ML; Neighbor Joining: NJ; Maximum Parsimony: MP). Black dots indicate support values above 70% for all three methods while gray dots show support values of more than 50% for all three methods and less than 70%, at least for one method. Branches that were not supported by all three methods show no dot. Scale bar indicates 10% estimated sequence divergence.
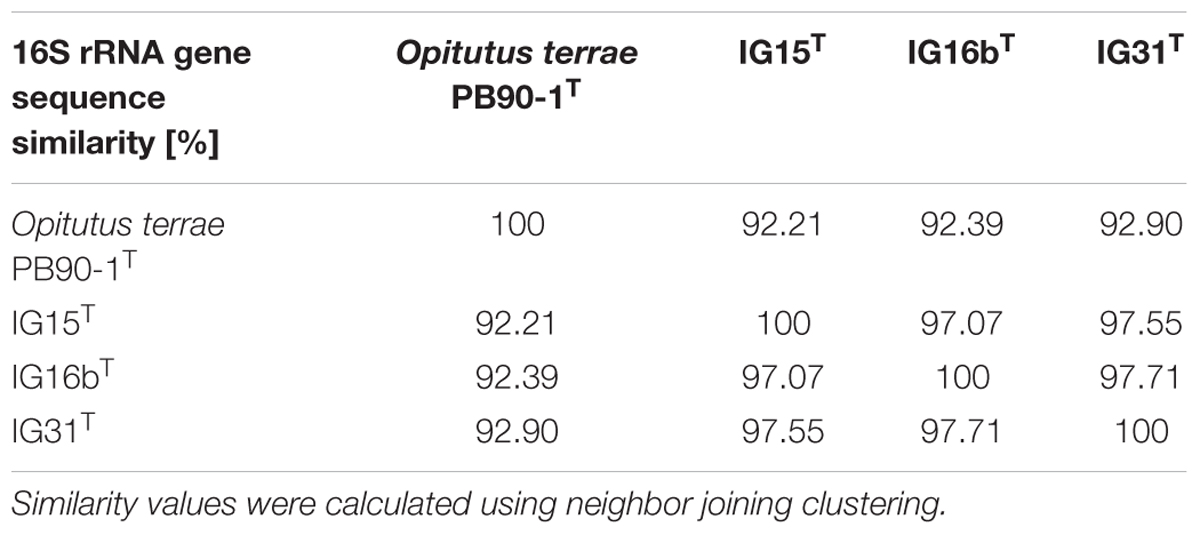
TABLE 1. 16S rRNA gene sequence identity matrix indicating similarity between the three verrucomicrobial isolates and the next relative Opitutus terrae PB90-1.
Cells of strains IG15T, IG16bT, and IG31T were investigated using light microscopic and electron microscopic techniques, revealing a coccoid cell shape with cells present as mono- or diplococci (Figures 2 and 3). No chain or rosette formation was observed. IG15T cells were the smallest of the three strains in average, measuring 0.6 ± 0.1 μm (diameter of single cocci with standard deviation; n = 100 cells) while cells of IG16bT and IG31T measured 0.9 ± 0.2 and 0.6 ± 0.1 μm in diameter, respectively (Figure 2D). In wide-field microscopy experiments, cell size variability of all three strains (compare Figure 2) became more evident than in scanning electron microscopy, where cells appeared smaller in size (compare Figure 3) due to osmotic stress during fixation. During exponential growth, cells of strain IG15T and IG31T were highly motile, while IG16bT showed only very few motile cells. While culture agitation was not necessary for growth, cells of strain IG15T produced an extracellular matrix when grown under constant agitation (90 rpm) (Figures 3A,B) with cells embedded in loose aggregates. No extracellular matrix formation was observed for strain IG16bT and IG31T (Figures 3C–F). All strains grow aerobically. Temperature and pH optima measurements revealed a mesophilic growth profile with growth temperatures from 13–38, 13–36, and 20–36°C for strains IG15T, IG16bT, and IG31T, respectively. Optical density changes during exponential growth pointed to optimum growth temperatures of 33, 32, and 30°C, respectively (Supplementary Figure S1). IG15T and IG16bT were able to grow in pH ranges from 6.0 to 9.0, with an optimum between 7.5 and 8.0. The pH optimum for strain IG31T was not determined, since its pH growth properties are likely to be similar to strains IG15T and IG16bT. Additionally, results of the oxidase assays were positive and determination of catalase activity showed negative results for all three strains. Strains were found to be Gram-negative by reaction with 3% KOH solution (Suslow et al., 1982). Substrate utilization profiles of strain IG15T and IG16bT showed similar patterns in terms of sugar and sugar acid utilization, while strain IG31T was clearly distinct, utilizing substrates such as glycyl-L-glutamic acid, L-rhamnose and succinic acid mono-methyl ester (Figure 4). Cellular fatty acid analysis identified iso-C15:0 as major component of IG15T and IG16bT cell walls with 33.3 and 48.6%, respectively, while IG31T only contained 9.1% of this particular fatty acid (Supplementary Table S3). Furthermore, IG31T possessed iso-C14:0 as major component (15.4%).
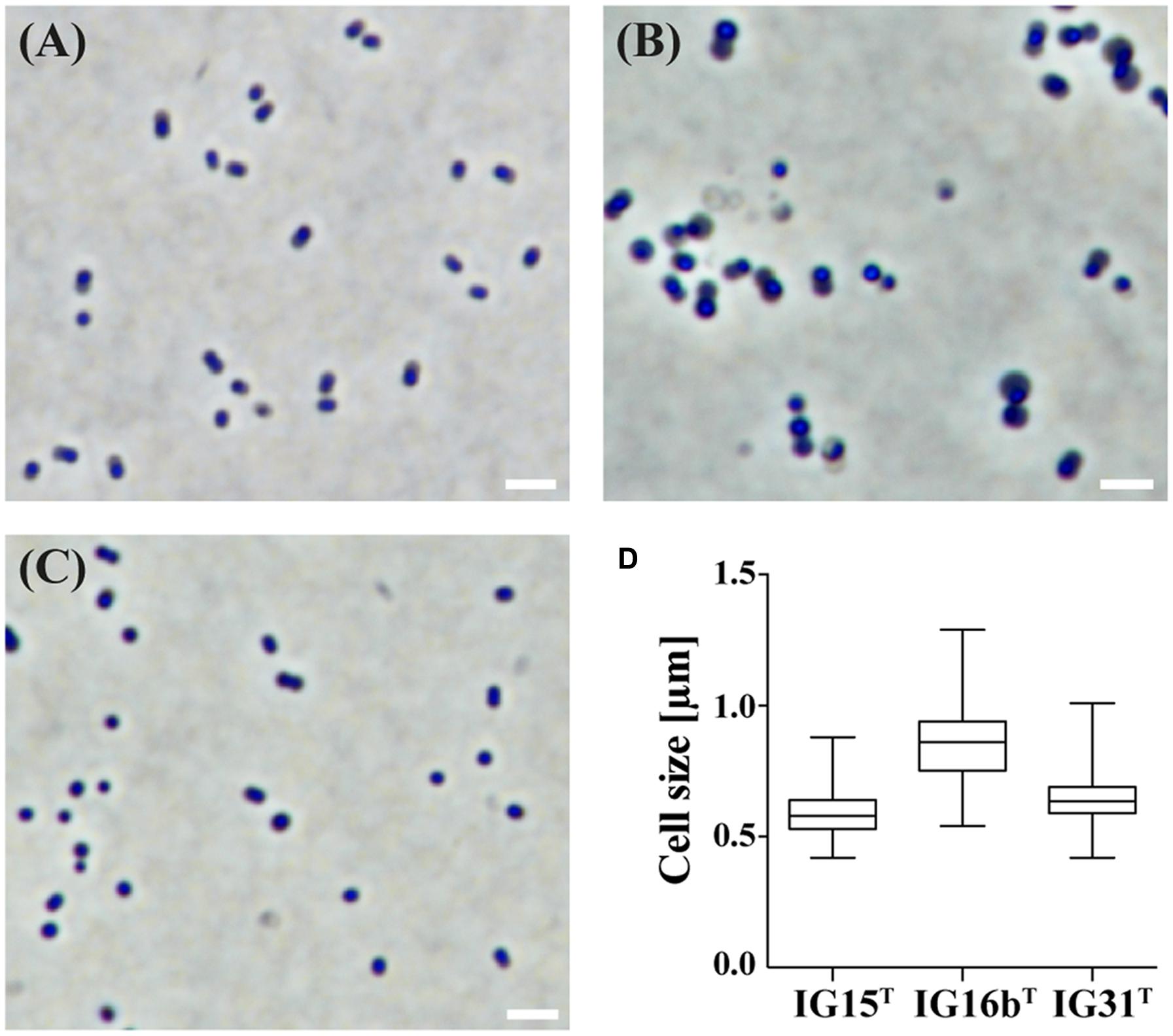
FIGURE 2. Investigation of cell morphology and size by light microscopy. The morphology and average cell size of IG15T (A), IG16bT (B), and IG31T (C) was investigated by light microscopy under phase-contrast illumination. Cells of strain IG15T (A), IG16bT (B), and IG31T (C) are of coccoid morphology and grow as mono- or diplococci. Cell size was determined by measuring 100 individual cells per strain (D) and average cell size with standard deviation differed from 0.6 ± 0.1 μm (IG15T), 0.9 ± 0.2 μm (IG16bT) to 0.6 ± 0.1 μm (IG31T). Scale bar indicates 2 μm.

FIGURE 3. Field emission scanning electron microscopy. Strains show coccoid morphology with organization as mono- or diplococci. Micrographs of IG15T cells (A, overview; B, close up) illustrate the formation of multicellular aggregates embedded in an extracellular matrix substance (white arrowheads). In contrast, cells of IG16bT (C, overview; D, close up) or IG31T (E, overview; F, close up) did not produce an extracellular matrix. Scale bar indicates 2 μm.
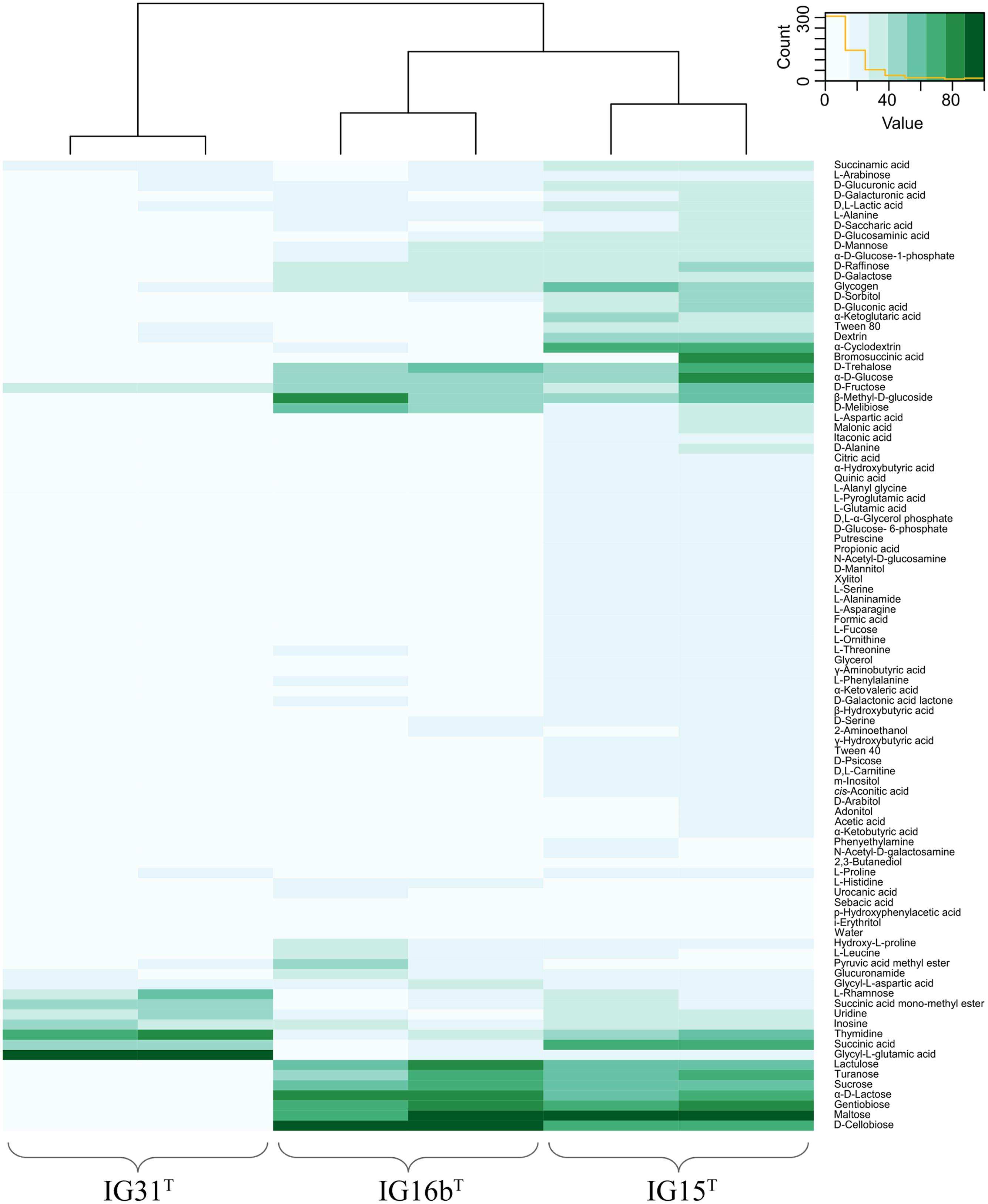
FIGURE 4. Heatmap illustration of substrate utilization. Substrate utilization was tested using the GN2 MicrologTM plate system. Substrate spectrum of IG15T was more similar to IG16bT, while some substrates such as succinic acid and α-cyclodextrin were almost solely degraded by IG15T. IG16bT in contrast was able to utilize D-cellobiose and α-D-lactose, distinguishing it from strains IG15T and IG31T. The utilization pattern of IG31T was less broad, encompassing eight of the 95 tested substrates, but included for example glycyl-L-glutamic acid, which was not utilized by IG15T or IG16bT.
Antibiotic susceptibility of strains IG15T and IG16bT toward β-lactams was investigated by treatment with carbenicillin. Optical density (OD600nm) measurements indicated growth at all tested antibiotic concentrations for both strains, as values increased over time (Supplementary Figures S2A,B). However, size measurements based on SEM micrographs revealed that treated cells of both strains were significantly increased in size when compared to untreated samples (Figure 5; p = 0.0001). Furthermore, the number of cells per ml was significantly lower (about 10-fold) in treated samples (Supplementary Figure S2C; p = 0.001). Thus, the increase of OD600nm was rather caused by swelling of the cells, than by multiplication after cell division.
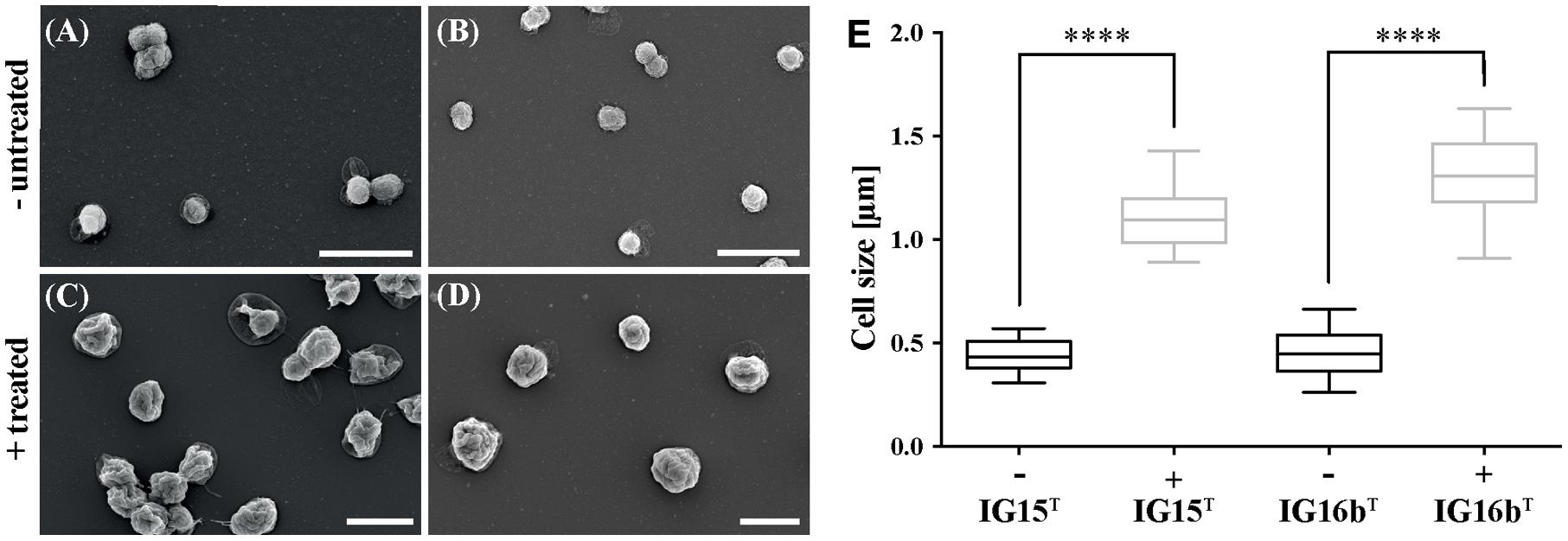
FIGURE 5. Effect of carbenicillin treatment on IG15T and IG16bT cells. Cells of IG15T and IG16bT were treated with carbenicillin for 120 h at 28°C. Untreated cells IG15T and IG16bT (A,B) showed no change in morphology, while size of treated cells (C,D: 2 mg/ml carbenicillin) was significantly increased (E). Cell size was calculated by counting 20–32 cells per treatment condition. ∗∗∗∗ = p < 0.0001. Scale bar indicates 2 μm.
The genome of strain IG16bT was obtained solely with single molecule real-time sequencing (PacBio). Sequencing read length was 3823 bp in average and yielded 616 mega bp of sequencing data from 6 SMRT cells with a coverage of ∼80× per base. Chromosome size was determined at 4,199,284 bp in length and bear a GC content of 66.5 mol%. Annotation with Prokka revealed the presence of 3575 coding sequences, 3 rRNA and 50 tRNA entries (Table 2). In Figure 6 the results of gene content analysis based on reciprocal blast are shown in a circular plot. Known genomes of subdivision 4 Verrucomicrobia are compared to the IG16bT chromosome, thereby revealing its unique genomic regions (Figure 6, gray boxes). Some of these regions were also predicted to be genomic islands (Figure 6, gray zones, outer rim), originating from horizontal gene transfer, and mainly hold hypothetical proteins or proteins with domains of unknown function. All predicted prophage regions (Figure 6, yellow zones, outer rim) were incomplete (Supplementary Table S4), thus no intact prophage exists in the chromosome of strain IG16bT.

TABLE 2. Basic genome information of the newly sequenced strain IG16bT and other described subdivision 4 Verrucomicrobia.
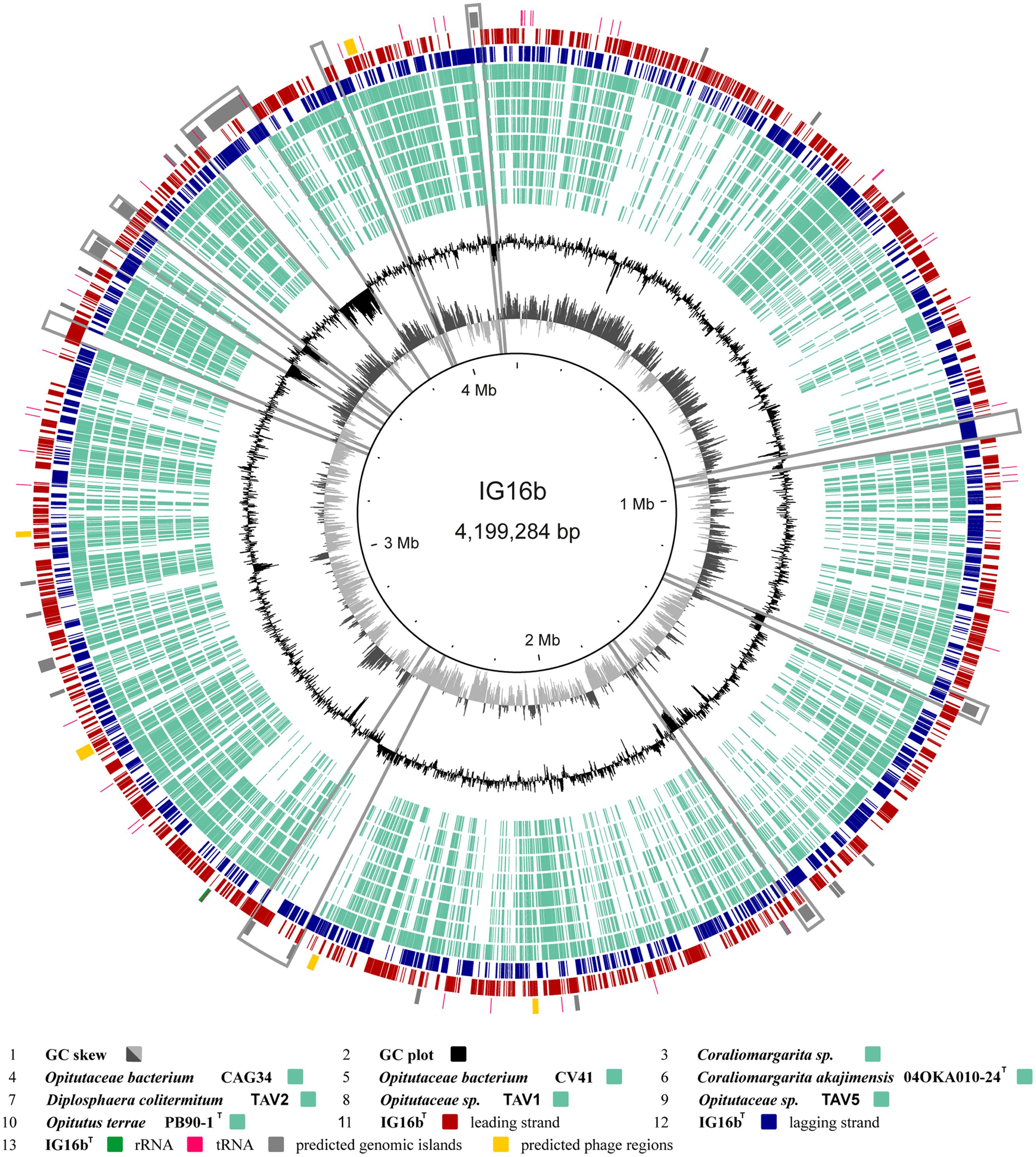
FIGURE 6. Circular plot of strain IG16bT’s 4,199,284 bp chromosome. Outer circles display protein (red and blue), tRNA (pink) and rRNA (green) encoding genes as well as predicted genomic islands (gray) and prophage regions (yellow). The inner circles show the GC plot (black) and the GC skew (dark and light gray). Ortholog genes from available genomes of subdivision 4 verrucomicrobial strains, were identified by reciprocal BLAST and are depicted in light turquoise. Notable islands of unique gene content in IG16bT are marked by gray boxes. These regions are often accompanied by a distinct change in GC composition. They mainly hold hypothetical proteins and proteins of unknown function. All predicted prophage regions are incomplete.
Using comparative genomics, we analyzed the genomes of strain IG16bT, O. terrae PB90-1T and C. akajimensis 04OKA010-24T (compare Table 2) with respect to genes required for the synthesis of peptidoglycan (PG). Results of our blast-based approach led to the conclusion that all investigated organisms harbor almost all genes essential for the synthesis of PG (Supplementary Table S5). Interestingly, for the penicillin binding proteins only ftsI was identified above threshold. Gene products of murB and murC were encoded polycistronic in IG16bT, O. terrae and C. akajimensis (compare Supplementary Table S5, orange boxes) leading to the identification of the same protein when investigated with the query protein sequences for MurB and MurC.
Tolerance of β-lactam-derived antibiotic agents in bacteria is often related to one of several modes of resistance, including efflux or exclusion mechanisms, alterations in target proteins or the most common cause being the presence of β-lactamases to degrade the antibiotic compound (Poole, 2004). Growth of strains IG15T, IG16bT, and IG31T on solid media supplemented with the β-lactam carbenicillin gave rise to the assumptions that these strains possess a mode of tolerance against β-lactams. Employing comparative genomics, we analyzed the presence of β-lactamase genes in the genomes of strain IG16bT, O. terrae and C. akajimensis (see Supplementary Tables S6 and S7). For IG16bT and O. terrae, three β-lactamases were identified, while for C. akajimensis no β-lactamase was found with the tested criteria. Our findings suggest that a tolerance mechanism against carbenicillin exists in strain IG16bT and is at least partially due to the presence of β-lactamases, leading to the survival of the organism until the antibiotic agent is decayed from the cultivation medium.
Treatment with lysozyme leads to the disruption of the cell envelope by hydrolytic cleavage of β-1,4-linkages in the peptidoglycan complex (Johnson et al., 1968). Untreated cells of strains IG15T, IG16bT, and IG31T maintained typical coccoid cell morphology, while all three strains displayed a loss of mobility during incubation at 37°C (Figures 7A–C, respectively). Cells that were treated with lysozyme for up to 24 h at 37°C in either culture medium (IG15T and IG16bT) or ddH2O (IG31T) showed different susceptibility levels toward the lysozyme treatment. Cells of strain IG15T showed no lysis in M1H medium after 1, 3, or 6 h, but were lysed after 24 h of incubation (Figure 7D; white arrowheads). Cells of strain IG16bT were destroyed after 3 h incubation in M1H medium (Figure 7E; white arrowheads). Since strain IG31T showed no lysis after 24 h in M1H medium, osmotic stress was increased by incubation of cells in ddH2O and cells were disrupted in ddH2O after 24 h (Figures 7D,F; white arrowheads).

FIGURE 7. Effect of lysozyme treatment on strains IG15T, IG16bT, and IG31T. Cells of all three strains were incubated with lysozyme. Untreated cells served as negative controls. Cell morphology was investigated by light microscopy. All strains were susceptible to lysozyme treatment, leading to disruption of the cells shape (D–F; white arrowheads), while cells in negative controls remained healthy during the whole incubation time (A–C). The isolates showed different resistance to the lysozyme treatment. Cells of isolate IG15T (D) and IG31T (F) were disrupted after 24 h of incubation in M1H medium or ddH2O, respectively. In contrast, cells of strain IG16bT (E) lysed after 3 h of incubation in M1H medium. Scale bar indicates 5 μm.
First, the presence of DAP was investigated for strains IG15T, IG16bT, and IG31T by TLC and no DL-DAP was detected. In contrast, Gram-negative and Gram-positive reference strains, E. coli DSM498 and B. subtilis DSM10, respectively, showed signals for DAP (Supplementary Figure S3), with E. coli giving only a weak signal. However, we analyzed whole-cell hydrolysates of IG15T, IG16bT, and IG31T using a more sensitive GC/MS method that previously revealed DAP in Planctomycetes. Despite negative results in TLC, we found the specific ion peaks, characteristic for DAP (compare Figure 8B), indicating the presence of peptidoglycan in IG15T, IG16bT, and IG31T and O. terrae PB90-1T. The same ion peaks were previously detected for E. coli DSM 498 (Spring et al., 2016), the identical E. coli strain we here used in our TLC experiment. In addition, ornithine was detected in the whole cell hydrolysates of all three novel strains and the closest related type strain, O. terrae (Figure 8A). A quantitative estimation, based on the internal standard used, revealed that DAP and ornithine occurred in nearly equivalent, albeit low amounts in strains IG15T and IG16bT while ornithine was the dominant substance detected for O. terrae and strain IG31T (Table 3). However, quantities of DAP for strains IG15T (7 nmol), IG16bT (6 nmol), IG31T (3 nmol) and O. terrae (4 nmol) were nearly 10-fold lower than those detected for the control E. coli strain (63 nmol), investigated in the study of Spring et al. (2016), which explains why no signal of DAP was visible in TLC experiments for strains IG15T, IG16bT, and IG31T, but a weak signal for E. coli (compare Supplementary Figure S3). Furthermore, proteins essential for DAP biosynthesis via the aminotransferase pathway are present in the genome of strain IG16bT (LysC :WP_069962807.1, WP_069963418.1; Asd: WP_069963129.1; DapA: WP_069962952.1; DapB: WP_069962953.1; DapL: WP_069960938.1; DapF: WP_069963382.1) as well as a alanine racemase (WP_069962553.1).
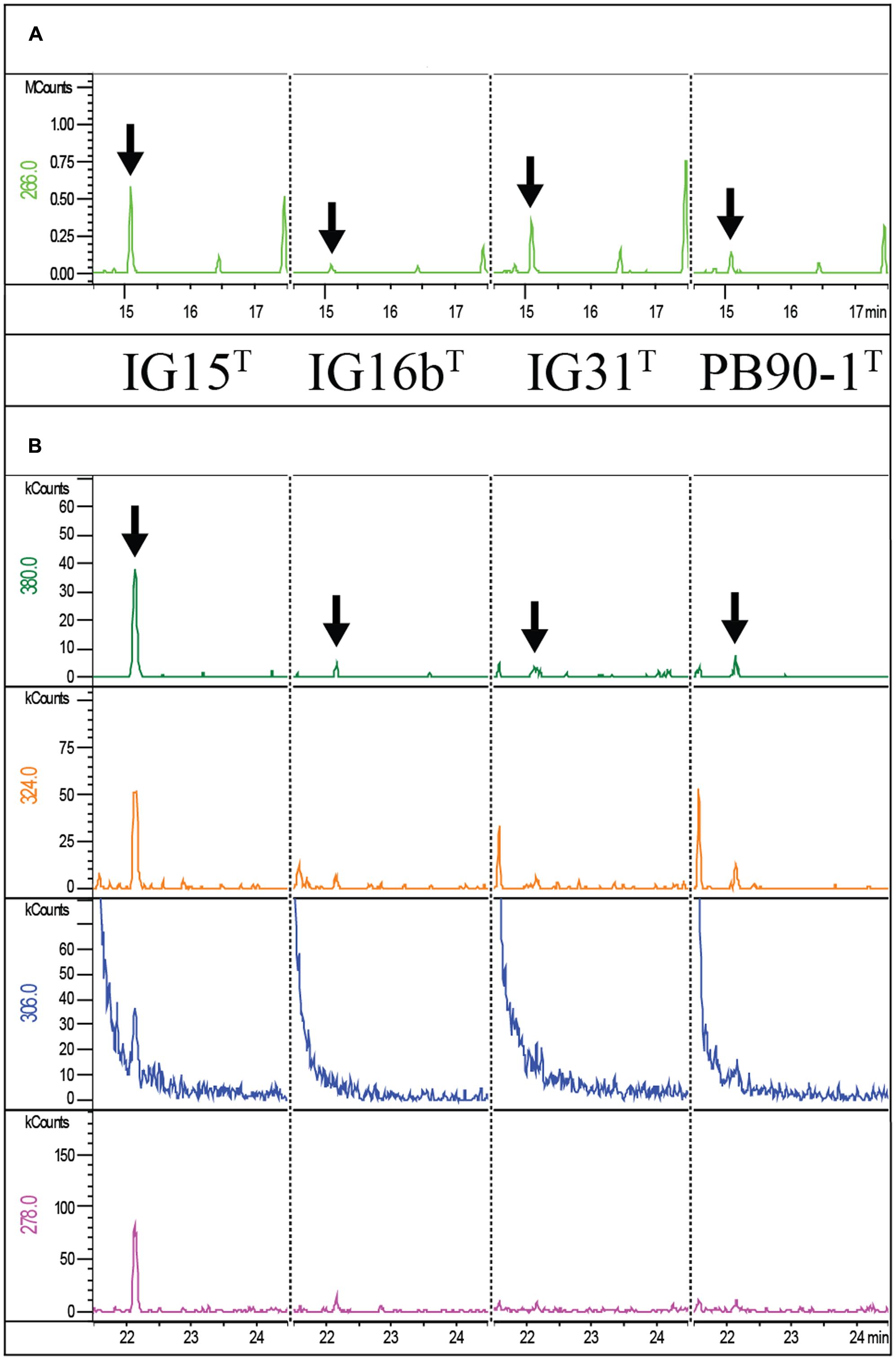
FIGURE 8. Mass spectrometric detection of diaminopimelic acid and ornithine in IG15T, IG16bT, IG31T and O. terrae PB90-1T. Extracted Ion chromatograms of ornithine (A) and the DAP derivative (N-heptafluorobutyryl DAP isobutylester) (B) from whole-cell hydrolysates of strains IG15T, IG16bT, IG31T and O. terrae PB90-1T are shown. Masses of the ornithine fragment (266 m/z) were detected for IG15T, IG16bT, IG31T, O. terrae PB90-1T at 15.13 min retention time. Masses of DAP fragments (380, 324, 306, and 278 m/z) were detected for IG15T, IG16bT, IG31T and O. terrae PB90-1T at 22.17 min retention time. Peaks confirming the presence of amino acids are highlighted for ornithine (A; black arrows) and DAP (B; black arrows) for all strains analyzed.

TABLE 3. Content of diagnostic diamino acids of peptidoglycan in whole-cell hydrolysates of Opitutus terrae PB90-1T and strains IG15T, IG16bT, and IG31T.
Thus, we conclude despite negative results in TLC, that all analyzed strains contain DAP as diagnostic diamino acid of peptidoglycan. Additionally, ornithine was detected which is a part of the peptidoglycan backbone of certain gram-negative bacteria (Yanagihara et al., 1984; Spring et al., 2016).
To give the ultimate proof that PG exists in the novel strains isolated in this study cell sacculi were extracted from strain IG16bT and investigated by TEM. TEM imaging revealed the presence of cell sacculi (Figure 9; Supplementary Figure S4) with remaining protein accumulations (white arrowheads) being present in the sample investigated.
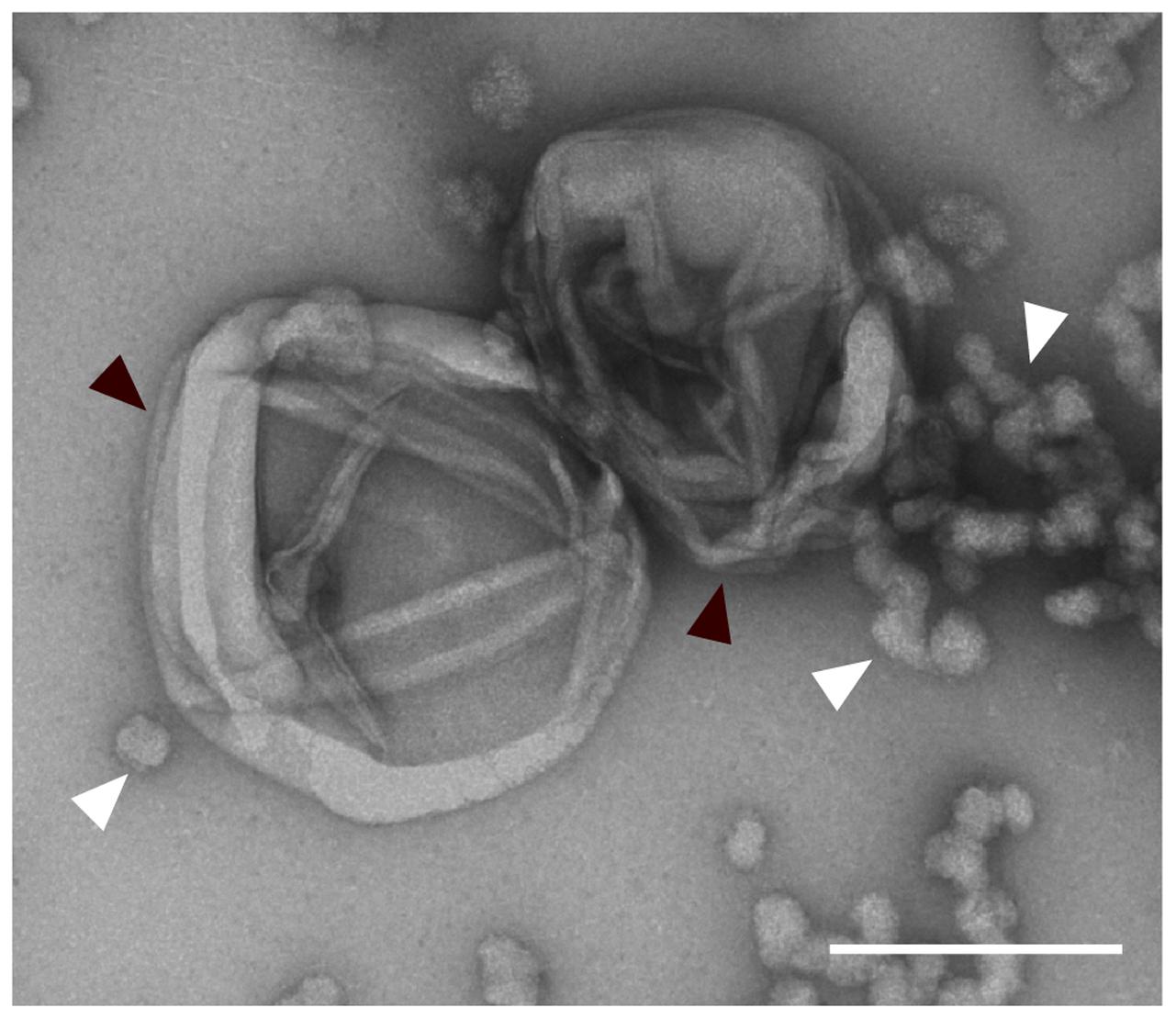
FIGURE 9. Extracted peptidoglycan sacculus of strain IG16bT. Cells were boiled in 4% SDS for 1 h and unbound SDS was dialyzed against ddH2O over the course of 3 days. Sacculi were negatively stained with 1% aqueous uranyl acetate and imaged by transmission electron microscopy (black arrowheads). Protein-bound SDS is seen in the sample (white arrowheads). Scale bar indicates 0.2 μm.
The isolation of PG sacculi together with the presence of DAP and PG synthesis genes, suggests that the claim of verrucomicrobial subdivision 4 lacking peptidoglycan, is not entirely justifiable (compare Table 4).
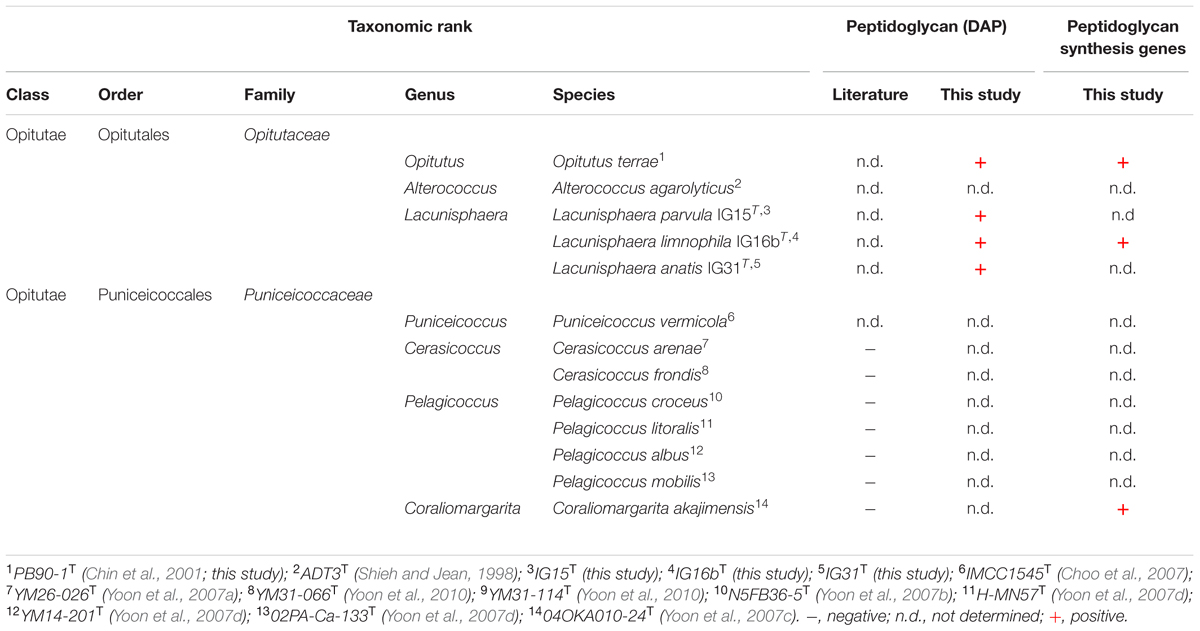
TABLE 4. Taxonomic affiliation and detected presence of peptidoglycan (DAP) in strains IG15T, IG16bT, IG31T and validly published type strains within subdivision 4 Verrucomicrobia.
All free-living bacteria possess a peptidoglycan cell wall (PG) to withstand environmental osmotic challenges and to maintain cell shape (Vollmer et al., 2008), with subdivision 4 Verrucomicrobia being described as one of the few exceptions (Yoon, 2011). Since it was recently demonstrated that Planctomycetes possess a PG cell wall (Jeske et al., 2015) despite oppositional previous reports (König et al., 1984), we revisited the question if subdivision 4 Verrucomicrobia are indeed an exception to this otherwise universal cell biological bacterial trait. Given that only a few representatives of the verrucomicrobial subdivision 4 are available in axenic culture, we applied a selective β-lactam-based cultivation approach considering the putative lack of PG to specifically enrich subdivision 4 Verrucomicrobia from a limnic water sample. As β-lactam antibiotics prevent PG formation and remodeling during cell division by irreversible interaction with penicillin-binding proteins involved in the final step of PG synthesis (Waxman and Strominger, 1983), subdivision 4 Verrucomicrobia should comprise intrinsic resistance if no PG per se exists. Accordingly, all three strains described in this study were obtained from plates initially containing carbenicillin. However, all novel verrucomicrobial strains grew only after 4 months of incubation, indicating rather antibiotic degradation through hydrolysis than an intrinsic resistance against β-lactam antibiotics. Thus, we analyzed the genome of the novel strain IG16bT in more detail to reveal the nature of its cell wall architecture and possible resistance mechanism to β-lactam antibiotics. First, we employed bioinformatics and found strain IG16bT to encode β-lactamase proteins that can confer resistance against β-lactam antibiotics such as carbenicillin. Second, we incubated cultures of strains IG16bT and IG15T with carbenicillin concentrations of 500–2000 mg/l, which were far above the 100 mg/l working concentration usually used as selection pressure for β-lactamase mediated resistance in molecular laboratory approaches (Green and Sambrook, 2012). Accordingly, SEM analysis revealed that carbenicillin treated cells were inhibited in cell division and increased in size. However, they withstood the antibiotic reagent and resumed growth, once carbenicillin was depleted from the cultivation medium, as happened through hydrolysis over time once they were initially isolated from the environment. Similar behavior has been observed for Chlamydia psittaci, where presence of penicillin led to swelling of reticulate bodies and incomplete cell division, while cells transferred to penicillin-free medium resumed division (Matsumoto and Manire, 1970). Our observations thus rather suggest a mode of tolerance, possibly enabled by β-lactamases, than a mode of intrinsic resistance due to the absence of PG in the novel isolates. In case of intrinsic resistance, increase of carbenicillin concentration would have had no effect on cell division. However, the degradation capability of β-lactamases can be titrated to a point, where the enzyme cannot confer resistance anymore and the cell becomes affected as observed in changes of morphology in this study. Thus, this finding provided us with the ample motivation to further analyze PG in our strains. To do this comprehensively, we analyzed the genome of strain IG16bT (obtained in this study) along with the published genomes of O. terrae (van Passel et al., 2011) and C. akajimensis (Mavromatis et al., 2010) employing complementary blast methods that were previously used to identify PG synthesis related genes in Planctomycetes (Jeske et al., 2015). We found that O. terrae. C. akajimensis and strain IG16bT harbor nearly all genes essential for the synthesis of PG. Thus, from a genomic perspective based on all available type strain genomes, it is likely that subdivision 4 Verrucomicrobia can synthesize PG.
Employing a previously described procedure (Jeske et al., 2015), we next demonstrated that all three novel strains, IG15T, IG16bT, and IG31T, are susceptible to the treatment with lysozyme, an enzyme that destroys beta-1,4 glycosidic bonds in the peptidoglycan structure, leading to disruption of the bacterial cell envelope (Johnson et al., 1968). Our results indicate different tolerance levels of strains IG15T, IG16bT, and IG31T against lysozyme, under laboratory culture conditions (in M1H medium) or under osmotic stress (IG31T in ddH2O).
Even though all evidence so far points toward the existence of an peptidoglycan cell wall in strains IG15T, IG16bT, and IG31T, we obtained no signals for the diagnostic peptidoglycan-specific structural element DAP when performing TLC experiments. This result is consistent with previous reports that led to the conclusion that subdivision 4 Verrucomicrobia lack PG (Yoon et al., 2007a). However, we analyzed whole-cell hydrolysates of our strains and O. terrae PB90-1T using a modified version –capable of quantification- of a highly sensitive method based on gas chromatography and mass spectrometry (GC/MS) detection that previously revealed DAP in Planctomycetes (Jeske et al., 2015). We found the specific ion set characteristic for DAP, while quantification of DAP in whole-cell hydrolysates of our strains and O. terrae revealed that this marker was only present in low quantities, possibly explaining why less sensitive methods such as TLC failed to detect DAP. Furthermore, all proteins essential for DAP synthesis were detected within the genome of strain IG16bT. In addition, we surprisingly detected the non-proteinogenic diamino acid ornithine that was, until recently, thought to be an exception in PG among Gram-negative bacteria limited to Spirochaetaceae (Schleifer and Joseph, 1973; Yanagihara et al., 1984). At this point, it cannot be excluded that ornithine could have been extracted from certain amino lipids or other cell components instead of peptidoglycan, because only whole-cell hydrolysates were analyzed. However, ornithine was recently identified in whole-cell hydrolysates of both, the proposed phylum Kiritimatiellaeota -formally known as verrucomicrobial subdivision 5- and representatives of the phylum Lentisphaerae (Spring et al., 2016), indicating that more Gram-negative bacteria display such alterations in their PG cell walls. Spring et al. additionally analyzed whole-cell hydrolysates of E. coli DSM 498, the same strain we used for TLC analysis, and found much higher quantities of DAP (63 nmol) then we did for our strains (compare Table 3), consequently supporting the observation of TLC being a method unfit to detect DAP in cases were only low quantities are present in the cell walls of the investigated organism.
To ultimately proof the existence of peptidoglycan sacculi, we isolated them from strain IG16bT and visualized them employing TEM (Figure 9).
Based on our findings, we conclude that subdivision 4 Verrucomicrobia do possess PG sacculi. Contrary previous reports used methods such as TLC (Yoon et al., 2007c) that did not detect DAP in subdivision 4 Verrucomicrobia in our hands as well (Supplementary Figure S3). Thus, future analyses must meet a new standard in PG detection, set by others and us, to justify the claim that a certain free-living bacterial strain lacks PG (Pilhofer et al., 2013; Jeske et al., 2015; Packiam et al., 2015; van Teeseling et al., 2015).
Based on recent results (Pilhofer et al., 2013; Jeske et al., 2015; Packiam et al., 2015; van Teeseling et al., 2015) and the outcome of this study we further postulate -applying the lex parsimoniae- that all free-living bacteria require a PG cell wall to maintain cell shape integrity in habitats with osmotic conditions different from their cytosol.
Lacunisphaera (La.cu.ni.sphae.ra N.L. fem. n. lacuna, a little lake, referring to the origin of the organism; N.L. fem. n. sphaera, a ball, globe, sphere; N.L. fem. n. Lacunisphaera, a spherical microorganism from a lake).
Cells are Gram-negative, aerobic cocci. Mono- or diplococcic are formed, but no chains or rosettes. Cells are motile during exponential growth phase, but not in late stationary phase. No spore formation was observed. Members test positive for cytochrome oxidase activity, but show no catalase activity in reaction with H2O2. Extracellular matrix formation in liquid culture is observed for some members when cultured under constant agitation. This is not true for the type species. The molar G + C content is between 65 and 67 mol%. Members contain peptidoglycan with DAP and ornithine as diamino acids. The predominant cellular fatty acid of the type species is iso-C15:0. Members belong to the phylum Verrucomicrobia, class Opitutae, order Optitutales, family Opitutaceae. The type species of the genus is Lacunisphaera limnophila.
Lacunisphaera parvula (par.vu.la, L. adj. parvula small, referring to the size of individual cells).
Main attributes are as given for the genus. Colonies grown on M1H agar were round, smooth and cream colored, while aging colonies became translucent. An extracellular matrix compound is produced in liquid cultures when kept under constant agitation, but formation was not observed on solid media. Cells are present as mono- or diplococci, but form aggregates when embedded in the extracellular matrix compound. Single cells measured 0.6 ± 0.1 μm in diameter. Substrates utilized were D-cellobiose, maltose, gentiobiose, α-D-lactose, sucrose, turanose, lactulose, succinic acid, thymidine, inosine, uridine, succinic acid mono-methyl ester, L-rhamnose, D-alanine, malonic acid, L-aspartic acid, D-melibiose, β-methyl-D-glucoside, D-fructose, α-D-glucose, DD-trehalose, bromosuccinic acid, α-cyclodextrin, dextrin, tween 80, α-ketoglutaric acid, D-gluconic acid, D-sorbitol, glycogen, D-galactose, D-raffinose, α-D-glucose-1-phosphate, D-mannose, D-glucosaminic acid, D-saccharic acid, L-alanine, D,L-lactic acid, D-galacturonic acid, D-glucuronic acid and succinamic acid. Cells grew in M1H medium at temperatures between 12 and 38°C, while 33°C was the optimum. Cells did not grow below 10°C and above 38°C. pH values between 6.0 and 9.0 were tolerated for growth, while the optimum was between 7.5 and 8.0. Major cellular fatty acids were iso-C15:0 (33.3%), C16:0 (10.2%), iso-C13:0 3-OH (8.7%), C16:1 ω5c (8.4%) and iso-C11:0 (4.9%). The G + C content of the DNA of the type strain is 65.9 mol%. The type strain is IG15T (=DSM 26814 = LMG 29468) and was isolated from the surface water column of a freshwater lake during a cyanobacterial blooming event.
Lacunisphaera limnophila (lim.no’ phi.la Gr. n. limnos lake; Gr. adj. philus loving; N.L. adj. limnophila lake loving).
Overall characteristics are as described for the genus. Colonies grown on M1H agar were round, smooth and cream colored, while aging colonies became translucent. Liquid cultures appeared pale yellowish. Cells are present as mono- or diplococci and form no chains or rosettes. Single cells measured 0.9 ± 0.2 μm in diameter. Substrates utilized were D-cellobiose, maltose, gentiobiose, α-D-lactose, sucrose, turanose, lactulose, thymidine, inosine, glycyl-L-aspartic acid, glucuronamide, pyruvic acid methyl ester, L-leucin, hydroxyl-L-proline, D-melibiose, β-methyl-D-glucoside, D-fructose, α-D-glucose, D-trehalose, glycogen, D-galactose, D-raffinose, α-D-glucose-1-phosphate and D-mannose. Cells grew in M1H medium at temperatures between 13 and 36°C, while 32°C was the optimum. Cells did not grow below 10°C and above 36°C. pH values between 6.0 and 9.0 were tolerated for growth, while the optimum was between 7.5 and 8.0. Major cellular fatty acids were iso-C15:0 (48.6%), Anteiso-C15:0 (12.1%), iso-C15:1 ω9c (10.3%), iso-C13:0 3-OH (6.6%) and iso-C13:0 (5.0%). The genome based G + C content of coding sequences is 66.5 mol%. The type strain is IG16bT (=DSM 26815 = LMG 29469) and was isolated from the particle-containing fraction of surface water from a freshwater lake. The 4,199,284 bp genome of L. limnophila IG16bT was similar, yet distinct from other sequenced verrucomicrobial species in terms of gene content (Figure 6). In most cases, such differences were associated with genomic islands which indicate frequent horizontal gene transfer.
Lacunisphaera anatis (a.na.tis L. fem. n. anatis with the ducks, referring to the term ‘duck pond’ describing a lake or pond inhabited by ducks).
Overall characteristics are as described for the genus. Colonies grown on M1H agar were round, smooth and cream colored, while aging colonies became translucent. Cells are present as mono- or diplococci and form no chains or aggregates. Single cells measured 0.6 ± 0.1 μm in diameter. Substrates utilized were glycyl-L-glutamic acid, succinic acid, thymidine, inosine, uridine, succinic acid mono-methyl ester, L-rhamnose, and D-fructose. Cells grew in M1H medium at temperatures between 15 and 36°C, while 30°C was the optimum. Cells did not grow below 12°C and above 36°C. pH values between 6.0 and 9.0 were tolerated for growth, while the optimum was between 7.5 and 8.0. Major cellular fatty acids were iso-C14:0 (15.4%), iso-C14:0 (15.4%), C16:0 (12.1%), Anteiso-C15:0 (10.6%) and iso-C16:0 (10.1%). The G + C content of the DNA of the type strain is 67.2 mol%. The type strain is IG31T (=DSM 103142 = LMG 29578) and was isolated from surface freshwater containing biomass of a cyanobacterial bloom.
PR did most of the experimental laboratory work and wrote the main part of the manuscript body and functions as first author. IG helped with the isolation of the novel strains and with cultivation measurements. CB performed all light microscopic work and cell size measurements. OJ conducted blast analysis for the detection of peptidoglycan synthesis genes and ß-lactamases. SW analyzed the gene content and generated substrate utilization heatmaps. RR coordinated sequencing of the IG16b genome sequence with Pacific Bioscience. PS did the gas chromatographic analysis of PG in the new strains. MR performed scanning electron microscopy experiments. SS cultivated reference strains for GC MS and fatty acid analysis and contributed in literature research toward ornithin in PG layers. FG analyzed sequencing data and was involved in genome assembly of IG16b’s genome sequence. CJ is PI and together with MJ functions as corresponding author. MJ and CJ, along with PR designed the study and helped with experimental setups and design.
This work was kindly funded by the Deutsche Forschungsgemeinschaft (JO 893/3-1). We thank Pacific Bioscience for genome sequencing.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We thank Anja Heuer for versatile and skillful technical assistance. Gabriele Pötter we thank for technical assistance in fatty acid analysis and thin layer chromatography of the novel strains.
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2017.00202/full#supplementary-material
Alikhan, N. F., Petty, N. K., Ben Zakour, N. L., and Beatson, S. A. (2011). BLAST Ring Image Generator (BRIG): simple prokaryote genome comparisons. BMC Genomics 12:402. doi: 10.1186/1471-2164-12-402
Altschul, S. F., Madden, T. L., Schaffer, A. A., Zhang, J., Zhang, Z., Miller, W., et al. (1997). Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nucleic Acids Res. 25, 3389–3402. doi: 10.1093/nar/25.17.3389
Bertani, G. (1951). Studies on lysogenesis. I. The mode of phage liberation by lysogenic Escherichia coli. J. Bacteriol. 62, 293–300.
Bush, K. (2013). The ABCD’s of beta-lactamase nomenclature. J. Infect. Chemother. 19, 549–559. doi: 10.1007/s10156-013-0640-7
Cashion, P., Hodler-Franklin, M. A., McCully, J., and Franklin, M. (1977). A rapid method for base ratio determination of bacterial DNA. Anal. Biochem. 81, 461–466. doi: 10.1016/0003-2697(77)90720-5
Chin, K. J., Liesack, W., and Janssen, P. H. (2001). Opitutus terrae gen. nov., sp. nov., to accommodate novel strains of the division ‘Verrucomicrobia’ isolated from rice paddy soil. Int. J. Syst. Evol. Microbiol. 51(Pt 6), 1965–1968. doi: 10.1099/00207713-51-6-1965
Choo, Y. J., Lee, K., Song, J., and Cho, J. C. (2007). Puniceicoccus vermicola gen. nov., sp. nov., a novel marine bacterium, and description of Puniceicoccaceae fam. nov., Puniceicoccales ord. nov., Opitutaceae fam. nov., Opitutales ord. nov. and Opitutae classis nov. in the phylum ‘Verrucomicrobia’. Int. J. Syst. Evol. Microbiol. 57(Pt 3), 532–537. doi: 10.1099/ijs.0.64616-0
Dhillon, B. K., Laird, M. R., Shay, J. A., Winsor, G. L., Lo, R., Nizam, F., et al. (2015). IslandViewer 3: more flexible, interactive genomic island discovery, visualization and analysis. Nucleic Acids Res. 43, W104–W108. doi: 10.1093/nar/gkv401
Engelhardt, H. (2007). Are S-layers exoskeletons? The basic function of protein surface layers revisited. J. Struct. Biol. 160, 115–124. doi: 10.1016/j.jsb.2007.08.003
Fox, A., Rogers, J. C., Gilbart, J., Morgan, S., Davis, C. H., Knight, S., et al. (1990). Muramic acid is not detectable in Chlamydia psittaci or Chlamydia trachomatis by gas chromatography-mass spectrometry. Infect. Immun. 58,k835–837.
Fuerst, J. A., and Sagulenko, E. (2011). Beyond the bacterium: planctomycetes challenge our concepts of microbial structure and function. Nat. Rev. Microbiol. 9, 403–413. doi: 10.1038/nrmicro2578
Green, M. R., and Sambrook, J. (2012). Molecular Cloning: A Laboratory Manual. New York, NY: Cold Spring Harbor Laboratory Press.
Jacquier, N., Viollier, P. H., and Greub, G. (2015). The role of peptidoglycan in chlamydial cell division: towards resolving the chlamydial anomaly. FEMS Microbiol. Rev. 39, 262–275. doi: 10.1093/femsre/fuv001
Jeske, O., Schüler, M., Schumann, P., Schneider, A., Boedeker, C., Jogler, M., et al. (2015). Planctomycetes do possess a peptidoglycan cell wall. Nat. Commun. 6:7116. doi: 10.1038/ncomms8116
Jogler, C., Waldmann, J., Huang, X., Jogler, M., Glöckner, F. O., Mascher, T., et al. (2012). Identification of proteins likely to be involved in morphogenesis, cell division, and signal transduction in Planctomycetes by comparative genomics. J. Bacteriol. 194, 6419–6430. doi: 10.1128/JB.01325-12
Johnson, L. N., Phillips, D. C., and Rupley, J. A. (1968). The activity of lysozyme: an interim review of crystallographic and chemical evidence. Brookhaven Symp. Biol. 21, 120–138.
Kim, M., Pak, S., Rim, S., Ren, L., Jiang, F., Chang, X., et al. (2015). Luteolibacter arcticus sp. nov., isolated from high Arctic tundra soil, and emended description of the genus Luteolibacter. Int. J. Syst. Evol. Microbiol. 65(Pt 6), 1922–1928. doi: 10.1099/ijs.0.000202
König, E., Schlesner, H., and Hirsch, P. (1984). Cell wall studies on budding bacteria of the Planctomyces/Pasteuria group and on a Prosthecomicrobium sp. Arch. Microbiol. 138, 200–205. doi: 10.1007/BF00402120
Kuykendall, L. D., Roy, M. A., Neill, J. J., and Devine, T. E. (1988). Fatty acids, antibiotic resistance, and deoxyribonucleic acid homology groups of Bradyrhizobium japonicum. Int. J. Syst. Evol. Microbiol. 38, 358–361. doi: 10.1099/00207713-38-4-358
Lechner, M., Findeiss, S., Steiner, L., Marz, M., Stadler, P. F., and Prohaska, S. J. (2011). Proteinortho: detection of (co-)orthologs in large-scale analysis. BMC Bioinformatics 12:124. doi: 10.1186/1471-2105-12-124
Lee, J., Park, B., Woo, S. G., Lee, J., and Park, J. (2014). Prosthecobacter algae sp. nov., isolated from activated sludge using algal metabolites. Int. J. Syst. Evol. Microbiol. 64(Pt 2), 663–667. doi: 10.1099/ijs.0.052787-0
Lee, K. C., Webb, R. I., Janssen, P. H., Sangwan, P., Romeo, T., Staley, J. T., et al. (2009). Phylum Verrucomicrobia representatives share a compartmentalized cell plan with members of bacterial phylum Planctomycetes. BMC Microbiol. 9:5. doi: 10.1186/1471-2180-9-5
Liechti, G., Kuru, E., Packiam, M., Hsu, Y. P., Tekkam, S., Hall, E., et al. (2016). Pathogenic chlamydia lack a classical sacculus but synthesize a narrow, mid-cell peptidoglycan ring, regulated by MreB, for cell division. PLoS Pathog. 12:e1005590. doi: 10.1371/journal.ppat.1005590
Liechti, G. W., Kuru, E., Hall, E., Kalinda, A., Brun, Y. V., van Nieuwenhze, M., et al. (2014). A new metabolic cell-wall labelling method reveals peptidoglycan in Chlamydia trachomatis. Nature 506, 507–510. doi: 10.1038/nature12892
Ludwig, W., Strunk, O., Westram, R., Richter, L., Meier, H., Yadhukumar, et al. (2004). ARB: a software environment for sequence data. Nucleic Acids Res. 32, 1363–1371. doi: 10.1093/nar/gkh293
Markowitz, V. M., Chen, I. M., Palaniappan, K., Chu, K., Szeto, E., Grechkin, Y., et al. (2012). IMG: the integrated microbial genomes database and comparative analysis system. Nucleic Acids Res. 40, D115–D122. doi: 10.1093/nar/gkr1044
Matsumoto, A., and Manire, G. P. (1970). Electron microscopic observations on the effects of penicillin on the morphology of Chlamydia psittaci. J. Bacteriol. 101, 278–285.
Mavromatis, K., Abt, B., Brambilla, E., Lapidus, A., Copeland, A., Deshpande, S., et al. (2010). Complete genome sequence of Coraliomargarita akajimensis type strain (04OKA010-24). Stand. Genomic. Sci. 2, 290–299. doi: 10.4056/sigs.952166
Mesbah, M., Premachandran, U., and Whitman, W. B. (1989). Precise measurement of the G+C content of deoxyribonucleic acid by high-performance liquid chromatography. Int. J. Syst. Bacteriol. 39, 159–167. doi: 10.1099/00207713-39-2-159
Miles, R. J. (1992). Catabolism in mollicutes. J. Gen. Microbiol. 138, 1773–1783. doi: 10.1099/00221287-138-9-1773
Miller, L. T. (1982). Single derivatization method for routine analysis of bacterial whole-cell fatty acid methyl esters, including hydroxy acids. J. Clin. Microbiol. 16, 584–586.
Packiam, M., Weinrick, B., Jacobs, W. R. Jr., and Maurelli, A. T. (2015). Structural characterization of muropeptides from Chlamydia trachomatis peptidoglycan by mass spectrometry resolves “chlamydial anomaly”. Proc. Natl. Acad. Sci. U.S.A. 112, 11660–11665. doi: 10.1073/pnas.1514026112
Parks, D. H., Imelfort, M., Skennerton, C. T., Hugenholtz, P., and Tyson, G. W. (2015). CheckM: assessing the quality of microbial genomes recovered from isolates, single cells, and metagenomes. Genome Res. 25, 1043–1055. doi: 10.1101/gr.186072.114
Pilhofer, M., Aistleitner, K., Biboy, J., Gray, J., Kuru, E., Hall, E., et al. (2013). Discovery of chlamydial peptidoglycan reveals bacteria with murein sacculi but without FtsZ. Nat. Commun. 4:2856. doi: 10.1038/ncomms3856
Pilhofer, M., Rappl, K., Eckl, C., Bauer, A. P., Ludwig, W., Schleifer, K. H., et al. (2008). Characterization and evolution of cell division and cell wall synthesis genes in the bacterial phyla Verrucomicrobia, Lentisphaerae, Chlamydiae, and Planctomycetes and phylogenetic comparison with rRNA genes. J. Bacteriol. 190, 3192–3202. doi: 10.1128/JB.01797-07
Poole, K. (2004). Resistance to beta-lactam antibiotics. Cell Mol. Life Sci. 61, 2200–2223. doi: 10.1007/s00018-004-4060-9
Pruesse, E., Peplies, J., and Glöckner, F. O. (2012). SINA: accurate high-throughput multiple sequence alignment of ribosomal RNA genes. Bioinformatics 28, 1823–1829. doi: 10.1093/bioinformatics/bts252
Razin, S. (2006). “The genus mycoplasma and related genera (Class Mollicutes),” in The Prokaryotes, eds M. Dworkin, S. Falkow, E. Rosenberg, K.-H. Schleifer, and E. Stackebrandt (Berlin: Springer), 836–904.
R Core Team. (2015). R: A Language Environment for Statistical Computing. Available at: https://www.r-project.org/
Rivas-Marín, E., Canosa, I., and Devos, D. P. (2016). Evolutionary cell biology of division mode in the bacterial planctomycetes-verrucomicrobia-chlamydiae superphylum. Front. Microbiol. 7:1964. doi: 10.3389/fmicb.2016.01964
Rosselló-Móra, R., and Amann, R. (2015). Past and future species definitions for Bacteria and Archaea. Syst. Appl. Microbiol. 38, 209–216. doi: 10.1016/j.syapm.2015.02.001
Schleifer, K. H., and Joseph, R. (1973). A directly cross-linked L-ornithine-containing peptidoglycan in cell walls of Spirochaeta stenostrepta. FEBS Lett. 36, 83–86. doi: 10.1016/0014-5793(73)80342-4
Schumann, P. (2011). “5 - Peptidoglycan Structure,” in Methods in Microbiology, eds R. Fred and O. Aharon (Cambridge, MA: Academic Press), 101–129.
Seemann, T. (2014). Prokka: rapid prokaryotic genome annotation. Bioinformatics 30, 2068–2069. doi: 10.1093/bioinformatics/btu153
Sharp, C. E., Op den Camp, H. J., Tamas, I., and Dunfield, P. F. (2013). “Unusual members of the PVC superphylum: the methanotrophic Verrucomicrobia genus “Methylacidiphilum”,” in Planctomycetes: Cell Structure, Origins and Biology, ed. J. A. Fuerst (New York, NY: Humana Press),k211–227.
Shieh, W. Y., and Jean, W. D. (1998). Alterococcus agarolyticus, gen.nov., sp.nov., a halophilic thermophilic bacterium capable of agar degradation. Can. J. Microbiol. 44, 637–645. doi: 10.1139/cjm-44-7-637
Spring, S., Bunk, B., Sproer, C., Schumann, P., Rohde, M., Tindall, B. J., et al. (2016). Characterization of the first cultured representative of Verrucomicrobia subdivision 5 indicates the proposal of a novel phylum. ISME J. 10, 2801–2816. doi: 10.1038/ismej.2016.84
Staneck, J. L., and Roberts, G. D. (1974). Simplified approach to identification of aerobic actinomycetes by thin-layer chromatography. Appl. Microbiol. 28, 226–231.
Stephens, R. S., Kalman, S., Lammel, C., Fan, J., Marathe, R., Aravind, L., et al. (1998). Genome sequence of an obligate intracellular pathogen of humans: Chlamydia trachomatis. Science 282, 754–759. doi: 10.1126/science.282.5389.754
Suslow, T. V., Schroth, M. N., and Isaka, M. (1982). Application of a rapid method for gram differentiation of plant pathogenic saprophytic bacteria without staining. Am. Phytopathol. Soc. 72, 917–918. doi: 10.1094/Phyto-72-917
Tamaoka, J., and Komagata, K. (1984). Determination of DNA base composition by reversed-phase high-performance liquid chromatography. FEMS Microbiol. Lett. 25, 125–128. doi: 10.1111/j.1574-6968.1984.tb01388.x
Valentine, R. C., Shapiro, B. M., and Stadtman, E. R. (1968). Regulation of glutamine synthetase. XII. Electron microscopy of the enzyme from Escherichia coli. Biochemistry 7, 2143–2152. doi: 10.1021/bi00846a017
van Passel, M. W., Kant, R., Palva, A., Copeland, A., Lucas, S., Lapidus, A., et al. (2011). Genome sequence of the verrucomicrobium Opitutus terrae PB90-1, an abundant inhabitant of rice paddy soil ecosystems. J. Bacteriol. 193, 2367–2368. doi: 10.1128/JB.00228-11
van Teeseling, M. C., Mesman, R. J., Kuru, E., Espaillat, A., Cava, F., Brun, Y. V., et al. (2015). Anammox Planctomycetes have a peptidoglycan cell wall. Nat. Commun. 6:6878. doi: 10.1038/ncomms7878
van Teeseling, M. C., Pol, A., Harhangi, H. R., van der Zwart, S., Jetten, M. S. M., Op den Camp, H. J., et al. (2014). Expanding the verrucomicrobial methanotrophic world: description of three novel species of Methylacidimicrobium gen. nov. Appl. Environ. Microbiol. 80:6782. doi: 10.1128/AEM.01838-14
Vollmer, W., Blanot, D., and de Pedro, M. A. (2008). Peptidoglycan structure and architecture. FEMS Microbiol. Rev. 32, 149–167. doi: 10.1111/j.1574-6976.2007.00094.x
Wagner, M., and Horn, M. (2006). The Planctomycetes. Verrucomicrobia. Chlamydiae and sister phyla comprise a superphylum with biotechnological and medical relevance. Curr. Opin. Biotechnol. 17, 241–249. doi: 10.1016/j.copbio.2006.05.005
Wang, G., Huang, X., Ng, T. B., Lin, J., and Ye, X. Y. (2014). High phylogenetic diversity of glycosyl hydrolase family 10 and 11 xylanases in the sediment of Lake Dabusu in China. PLoS ONE 9:e112798. doi: 10.1371/journal.pone.0112798
Wang, X., Sharp, C. E., Jones, G. M., Grasby, S. E., Brady, A. L., and Dunfield, P. F. (2015). Stable-isotope probing identifies uncultured planctomycetes as primary degraders of a complex heteropolysaccharide in soil. Appl. Environ. Microbiol. 81, 4607–4615. doi: 10.1128/AEM.00055-15
Waxman, D. J., and Strominger, J. L. (1983). Penicillin-binding proteins and the mechanism of action of beta-lactam antibiotics. Annu. Rev. Biochem. 52, 825–869. doi: 10.1146/annurev.bi.52.070183.004141
Yanagihara, Y., Kamisango, K., Yasuda, S., Kobayashi, S., Mifuchi, I., Azuma, I., et al. (1984). Chemical compositions of cell walls and polysaccharide fractions of spirochetes. Microbiol. Immunol. 28, 535–544. doi: 10.1111/j.1348-0421.1984.tb00706.x
Yoon, J. (2011). Phylogenetic studies on the bacterial phylum ‘Verrucomicrobia’. Microbiol. Cult. Coll. 27, 61–65.
Yoon, J., Matsuo, Y., Matsuda, S., Adachi, K., Kasai, H., and Yokota, A. (2007a). Cerasicoccus arenae gen. nov., sp. nov., a carotenoid-producing marine representative of the family Puniceicoccaceae within the phylum ‘Verrucomicrobia’, isolated from marine sand. Int. J. Syst. Evol. Microbiol. 57(Pt 9), 2067–2072. doi: 10.1099/ijs.0.651020
Yoon, J., Oku, N., Matsuda, S., Kasai, H., and Yokota, A. (2007b). Pelagicoccus croceus sp. nov., a novel marine member of the family Puniceicoccaceae within the phylum ‘Verrucomicrobia’ isolated from seagrass. Int. J. Syst. Evol. Microbiol. 57(Pt 12), 2874–2880. doi: 10.1099/ijs.0.65286-0
Yoon, J., Yasumoto-Hirose, M., Katsuta, A., Sekiguchi, H., Matsuda, S., Kasai, H., et al. (2007c). Coraliomargarita akajimensis gen. nov., sp. nov., a novel member of the phylum ‘Verrucomicrobia’ isolated from seawater in Japan. Int. J. Syst. Evol. Microbiol. 57(Pt 5), 959–963. doi: 10.1099/ijs.0.64755-0
Yoon, J., Yasumoto-Hirose, M., Matsuo, Y., Nozawa, M., Matsuda, S., Kasai, H., et al. (2007d). Pelagicoccus mobilis gen. nov., sp. nov., Pelagicoccus albus sp. nov. and Pelagicoccus litoralis sp. nov., three novel members of subdivision 4 within the phylum ‘Verrucomicrobia’, isolated from seawater by in situ cultivation. Int. J. Syst. Evol. Microbiol. 57(Pt 7), 1377–1385. doi: 10.1099/ijs.0.64970-0
Yoon, J., Matsuo, Y., Matsuda, S., Kasai, H., and Yokota, A. (2010). Cerasicoccus maritimus sp. nov. and Cerasicoccus frondis sp. nov., two peptidoglycan-less marine verrucomicrobial species, and description of Verrucomicrobia phyl. nov., nom. rev. J. Gen. Appl. Microbiol. 56, 213–222. doi: 10.2323/jgam.56.213
Keywords: peptidoglycan, subdivision 4, Verrucomicrobia, Lacunisphaera, ornithine
Citation: Rast P, Glöckner I, Boedeker C, Jeske O, Wiegand S, Reinhardt R, Schumann P, Rohde M, Spring S, Glöckner FO, Jogler C and Jogler M (2017) Three Novel Species with Peptidoglycan Cell Walls form the New Genus Lacunisphaera gen. nov. in the Family Opitutaceae of the Verrucomicrobial Subdivision 4. Front. Microbiol. 8:202. doi: 10.3389/fmicb.2017.00202
Received: 28 September 2016; Accepted: 27 January 2017;
Published: 13 February 2017.
Edited by:
Damien Paul Devos, Pablo de Olavide University, SpainReviewed by:
Seong Woon Roh, Korea Basic Science Institute, South KoreaCopyright © 2017 Rast, Glöckner, Boedeker, Jeske, Wiegand, Reinhardt, Schumann, Rohde, Spring, Glöckner, Jogler and Jogler. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Christian Jogler, Y2hyaXN0aWFuQGpvZ2xlci5kZQ== Mareike Jogler, bWFyZWlrZUBqb2dsZXIuZGU=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.