 Lucas J. Lenzi1
Lucas J. Lenzi1 Paula M. A. Lucchesi
Paula M. A. Lucchesi Alejandra Krüger
Alejandra Krüger- 1Laboratorio de Inmunoquímica y Biotecnología, Facultad de Ciencias Veterinarias, Universidad Nacional del Centro de la Provincia de Buenos Aires, Tandil, Argentina
- 2Centro de Investigación Veterinaria de Tandil, CONICET-CIC-UNCPBA, Tandil, Argentina
Induction and propagation of bacteriophages along the food production chain can represent a significant risk when bacteriophages carry genes for potent toxins. The aim of this study was to evaluate the effect of different compounds used in the food industry on the growth of Shiga toxin-producing Escherichia coli (STEC) and the production of stx-phage particles and Shiga toxin. We tested the in vitro effect of lactic acid, acetic acid, citric acid, disodium phosphate, and sodium citrate on STEC growth. A bacteriostatic effect was observed in most of treated cultures. The exceptions were those treated with sodium citrate and disodium phosphate in which similar growth curves to the untreated control were observed, but with reduced OD600 values. Evaluation of phage production by plaque-based assays showed that cultures treated with sodium citrate and disodium phosphate released phages in similar o lower levels than untreated cultures. However, semi-quantification of Stx revealed higher levels of extracellular Stx in STEC cultures treated with 2.5% sodium citrate than in untreated cultures. Our results reinforce the importance to evaluate if additives and other treatments used to decrease bacterial contamination in food induce stx-phage and Stx production.
Introduction
Shiga toxin-producing Escherichia coli (STEC) are important pathogens that can cause human diseases, like diarrhoea, haemorrhagic colitis, and haemolytic uraemic syndrome (HUS) (Karmali et al., 1985). STEC strains are characterized by their capacity to produce Shiga toxins, which are encoded by bacteriophages, usually named stx-phages. These phages influence the pathogenicity of STEC strains, since the expression of these toxins is upregulated when the lytic cycle of these phages is induced. They also play a role in the spread of stx among E. coli as well as to other bacteria (Schmidt et al., 1999; Muniesa et al., 2004; Probert et al., 2014). Several factors have been shown to induce the lytic pathway of stx-phages [revised by Krüger and Lucchesi (2015)]. Particularly, some antibiotics have been reported to increase phage induction and Stx production and therefore treatments of human STEC infections with some antibiotics may have adverse clinical consequences (Wong et al., 2000; Zhang et al., 2000; McGannon et al., 2010).
Meat consumption has been identified as one of the risk factors strongly associated with HUS (Bentancor et al., 2012) and several studies have shown that meat is frequently contaminated with STEC strains (Parma et al., 2000; Bosilevac and Koohmaraie, 2011). Food producing animals have been recognized as the most important source for the entry of STEC in the food chain (Arthur et al., 2002; Martin and Beutin, 2011), and ruminants, especially cattle, have been identified as the major reservoir of STEC strains (Caprioli et al., 2005; Mainil and Daube, 2005). In addition to meat and dairy products, other vehicles and transmission routes of STEC strains for human infection have been reported, like person-to-person contact and drinking and swimming water [reviewed by Doyle et al. (2006) and Kaspar et al. (2010)]. Several intervention strategies have been proposed to minimize meat contamination by STEC during slaughtering and meat processing, like hide washing (Arthur et al., 2007), hot water treatment (Bosilevac et al., 2006), high hydrostatic pressure (HHP) treatment (Gola et al., 2000), and treatments with organic acids and/or their salts (Eswaranandam et al., 2004; Over et al., 2009), among others.
Infectious stx-phages have been detected in minced beef and salad samples (Imamovic and Muniesa, 2011). Moreover, stx-encoding phages can be more resistant than their host bacteria to chlorination and heat treatments (Muniesa et al., 1999) and also have a high ability to tolerate exposure to several disinfectants (Rode et al., 2011). There is scarce information about the risk of stx-phage induction and stx gene dissemination to other bacteria in foods. Imamovic et al. (2009) showed that stx transduction in food matrices is possible under appropriate conditions. Studies are required to evaluate if antimicrobial interventions, as well as other farming practices and food processing technologies, may increase the rate of induction and propagation of stx-phages.
Several substances are used in the production, processing, treatment, packaging, transportation, and storage of food. In the meat industry, additives like citric acid and sodium citrate are widely applied for pH control, metal chelating, and preservation (Sammel et al., 2006). Phosphates are used in meat and meat products to adjust pH, sequester cations, change the ionic charges distributions, change the ionic strength, and/or to function as a bacteriostatic agent (Long et al., 2011). The aim of this study was to evaluate the in vitro effect of some additives on STEC growth, and on the production of stx-phage and Stx.
Material and Methods
Bacterial Strains
STEC strains FB3 (serotype O157:H7, stx2-positive) and FB5 (serotype O145:H-, stx2-positive) isolated from feedlot cattle (Padola et al., 2004), and the reference strain E. coli EDL933 (serotype O157:H7, positive for stx1 and stx2) were selected to evaluate the effect of additives on STEC growth. In addition, supernatants from E. coli EDL933 cultures were used to evaluate phage and Stx production. E. coli laboratory strain DH5α was used as host strain for stx phages in double-agar-layer plaque assays.
Additives
The following compounds were prepared in stock solutions and used at indicated final concentrations: lactic acid (0.5 and 2.5% v/v), acetic acid (2.5% v/v), citric acid (0.5 and 2.5% w/v), disodium phosphate (0.1, 0.5, and 1% w/v), and sodium citrate (0.5, 1.0, and 2.5% w/v).
Bacterial Growth/Lysis Curves
STEC strains were cultivated overnight in Luria Bertani (LB) medium at 37°C with shaking at 120 rpm. Aliquots (300 μl) were inoculated into 100 ml flasks containing 15 ml of fresh LB medium. The new cultures were incubated at 37°C and 120 rpm ~45 min. up to an optical density at 600 nm (OD600) ≈ 0.2-0.3 when each flask was added with the respective additive, or water (untreated control), or mitomycin C (final concentration of 0.5 μg/ml; positive control of phage induction), reaching a final culture volume of 15.5 ml. This moment was identified as 0 h of the assay. The incubation was continued at 37°C and 180 rpm for 18 h, and spectrophotometrically monitored every hour for the first 5 h. When it was necessary, the aliquots measured were previously diluted. In addition, viable bacterial count at 2 h was conducted by plating appropriate dilutions on LB agar plates. These assays were repeated at least two times.
Evaluation of Phage Production
For phage quantification, aliquots of EDL933 cultures under different treatments were assayed. This strain was selected because phages induced from it showed lysis plaques easier to be counted by visual inspection than those from FB3 and FB5 strains. Aliquots from EDL933 cultures were taken at 3 h and centrifuged for 10 min at 10,000 × g, at 4°C. Supernatants were collected, filtered through low-protein-binding 0.22 μm membrane filters (Millex-GV, Millipore) and tenfold serially diluted for titration assays using the double-agar-layer method as follows. One hundred microliters of each dilution were mixed with 500 μl of an exponential phase culture of E. coli DH5α (OD600 ≈ 0.6–0.8) and incubated for 30 min at 37°C with shaking (120 rpm) (phage adsorption step). This suspension was then mixed with 3 ml of LB soft agar (0.75% w/v) supplemented with 9 mM CaCl2 and 1.5 μg/ml ampicillin and poured onto LB agar plates supplemented with 0.5–1 μg/ml ampicillin. After 18 h incubation at 37°C, lysis plaques were examined and enumerated. Ten lysis plaques were picked out from each plate and individually evaluated by PCR for stx carriage using a multiplex assay that detects stx1, stx2, and eae genes (Paton and Paton, 1998). This last gene was used to check the absence of chromosomal DNA from the STEC strain which could otherwise lead to false positive results in stx-phage PCR detection.
Phage quantification assays were repeated at least two times. To avoid possible interferences in the double-agar-layer method related with a possible chelating effect of disodium phosphate and sodium citrate, two more assays (named 3 and 4), were performed with addition of 100 μl CaCl2 0.1 M in the phage adsorption step.
Evaluation of Extracellular Shiga toxin
Stx production was semi-quantified by using an enzyme immunoassay (EIA, Ridascreen® Verotoxin, R-Biopharm, Germany). Supernatants of cultures at 18 h were obtained by centrifugation at 12,000 ×g for 10 min, diluted 1/10 in LB and then analyzed according to manufacturer instructions. Regarding supernatants from cultures treated with acids, they were neutralized before performing the ELISA.
The results were spectrophotometrically measured at 450 nm and classified as weak positive (1+) if the extinction was >0.1–0.5 above the negative control, moderate (2+) (>0.5–1.0) and strongly positive 3+ (>1.0–2.0) to 4+ (>2.0). The assays were done twice.
Results and Discussion
Some practices used along the food chain, like addition of substances, may influence bacterial growth and could increase the rate of induction and propagation of bacteriophages. This could represent a risk when the bacteriophages carry genes for potent toxins such as Stx. In this study, we evaluated the effect of different compounds used in the meat industry on the growth of STEC strains, and on the production of stx-phages and Stx.
Cultures of FB3, FB5, and EDL933 strains (control cultures and cultures exposed to different concentration of the compounds) were incubated and monitored spectrophoto metrically. For each treatment, similar OD600 patterns were observed for each of the three strains (results for EDL933 are shown). Cultures added with mitomycin C showed an increase in the OD600 during the first hours, reaching a maximum 2 h after induction, followed by a significant decrease, corresponding to a typical pattern of host cell lysis subsequent to induction of phage lytic cycle. This growth/lysis pattern was not observed in any of the cultures treated with the additives.
Cultures treated with lactic acid, acetic acid, or citric acid, at the concentrations tested, did not show an increase in OD600 over time (Figure 1 shows the results for treatments with 2.5% citric acid, 2.5% lactic acid, and 2.5% acetic acid). Phage titration assays showed that phages in supernatants of cultures treated with acids were below the accepted range for countable phage plaques (around 3–5 plaques per plate could be observed from undiluted supernatants, which represented a ~2-log reduction in the plaque counts in comparison to the untreated control). In addition, extracellular Stx was not detected in the supernatants. Taking into account the results, those acids at tested concentrations inhibited the bacterial growth, without evidence of production of stx-phages and Stx.
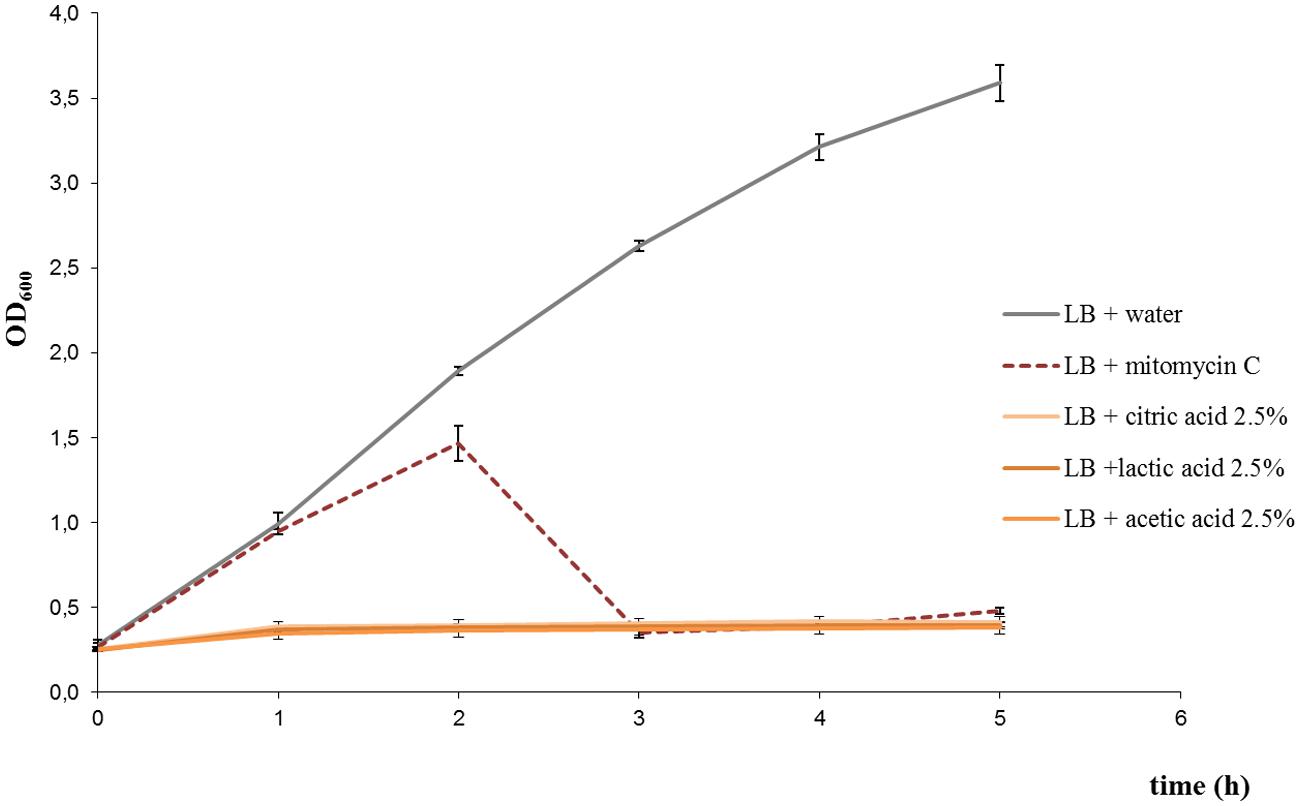
FIGURE 1. Growth curves of Escherichia coli EDL933 strain under different treatments: 2.5% citric acid, 2.5% lactic acid, 2.5% acetic acid, and 0.5 μg/ml mitomycin C. Growth curve for the untreated control is also shown. Similar results were obtained with E. coli FB5 and FB3 strains under the different treatments (not shown).
Cultures treated with disodium phosphate or sodium citrate showed growth patterns similar to untreated cultures but with a reduction in the OD600 values with increasing salt concentrations (Figures 2A,B). Besides, viable bacterial counts showed that EDL933 cultures exposed to these salts had titers similar or slightly lower than untreated control cultures, and markedly higher than cultures with mitomycin C (Figure 3). In the first two phage titration assays, the supernatants of the cultures treated with disodium phosphate or sodium citrate contained phages below the accepted range for countable phage plaques, while titers of 3 × 102 and 4 × 105 plaque forming units (pfu)/ml were observed in untreated cultures and cultures with mitomycin C, respectively. The supplementation of the supernatants with CaCl2 in the adsorption step allowed quantification of phages from most of the cultures treated with salts, and also increased the number of pfu for both untreated and mitomycin C added controls. In all cases, the plaques analyzed by PCR were confirmed to correspond to stx2-phages.
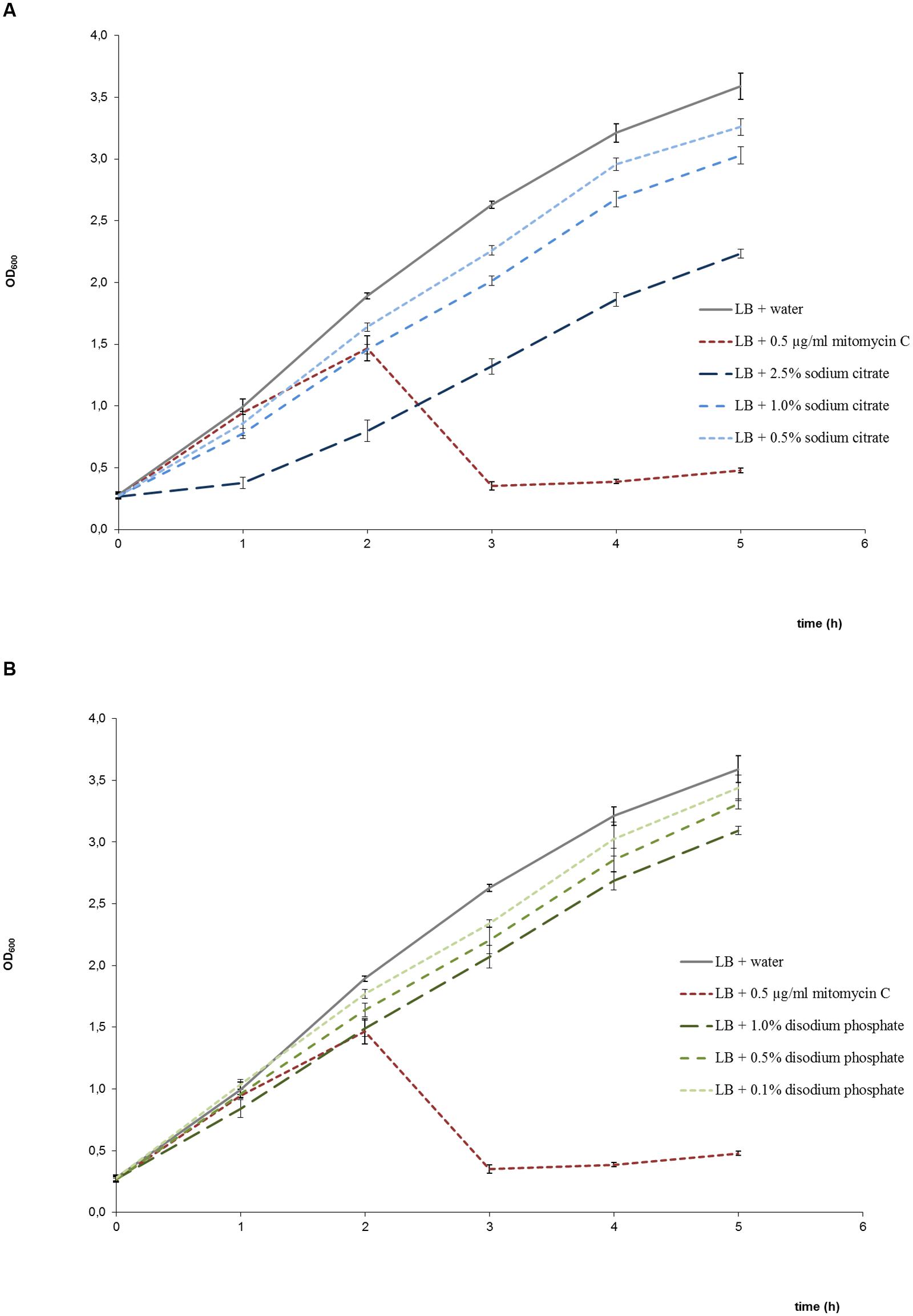
FIGURE 2. Growth curves of E. coli EDL933 in LB medium in the presence and absence of sodium citrate (A), and in the presence and absence of disodium phosphate (B). Bacterial growth was determined by measuring the OD600. Each point corresponds to the average of four determinations ± standard error. Growth in the presence of mitomycin C was also included in both graphs as positive control of bacteriolysis due to phage lytic cycle induction.
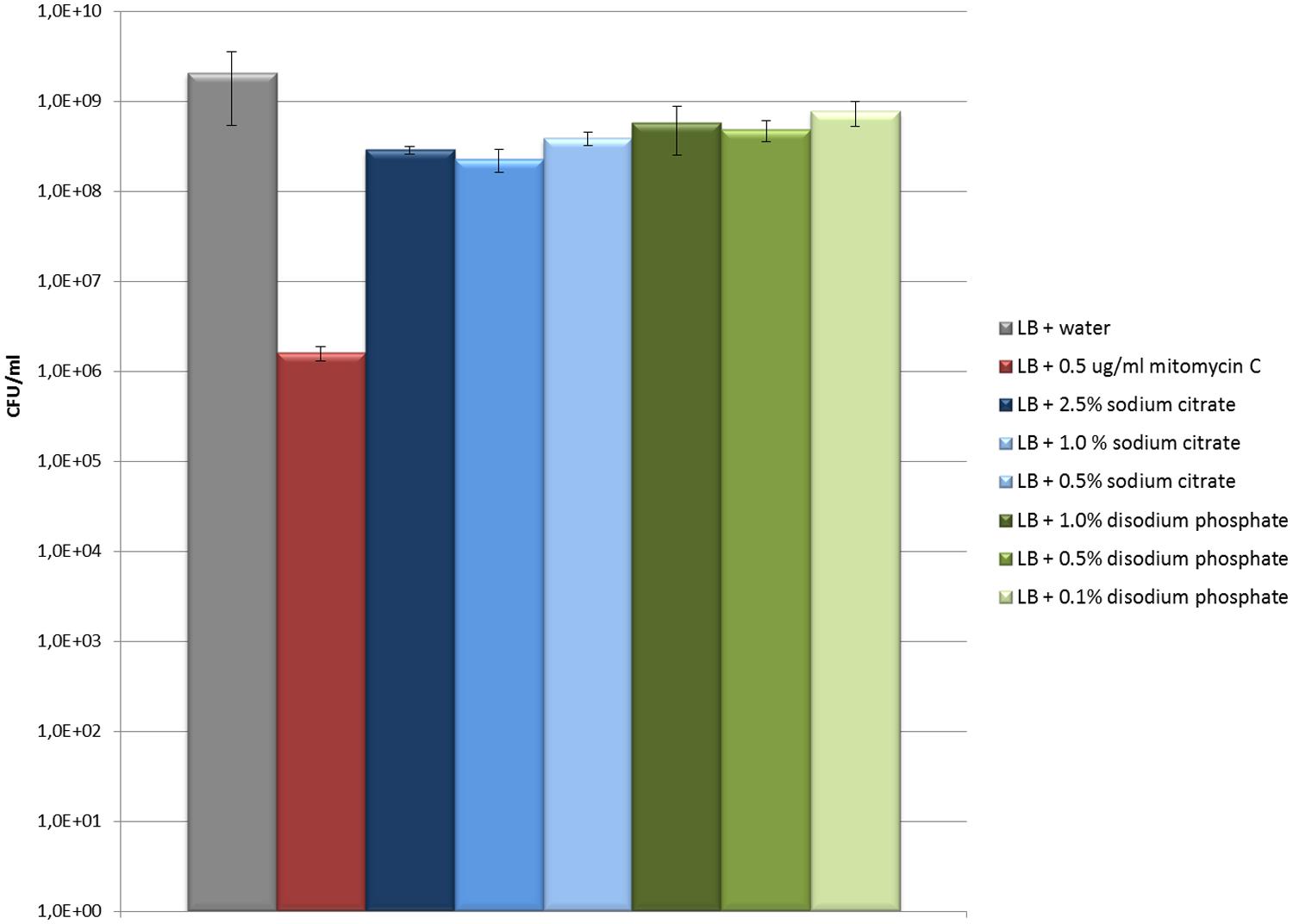
FIGURE 3. Evaluation of the effect of sodium citrate and disodiun phosphate on E. coli EDL933 growth (CFU/ml). The values and error bars on the graph are averages of three independent experiments.
Phage titers observed for the supernatants of cultures treated with disodium phosphate and those with 0.5 and 1% sodium citrate were lower than those observed for untreated cultures (Figure 4). The exception was the culture treated with 2.5 % sodium citrate, which showed titers similar to the untreated control (Figure 4). Regarding extracellular Stx production, cultures treated with disodium phosphate presented similar results to the untreated control (Table 1). The cultures supplemented with 0.5% and 1.0% sodium citrate showed similar or slightly higher levels of Stx than the water control, respectively. Interestingly, cultures with 2.5% sodium citrate showed Stx levels considerable higher than those of untreated cultures.
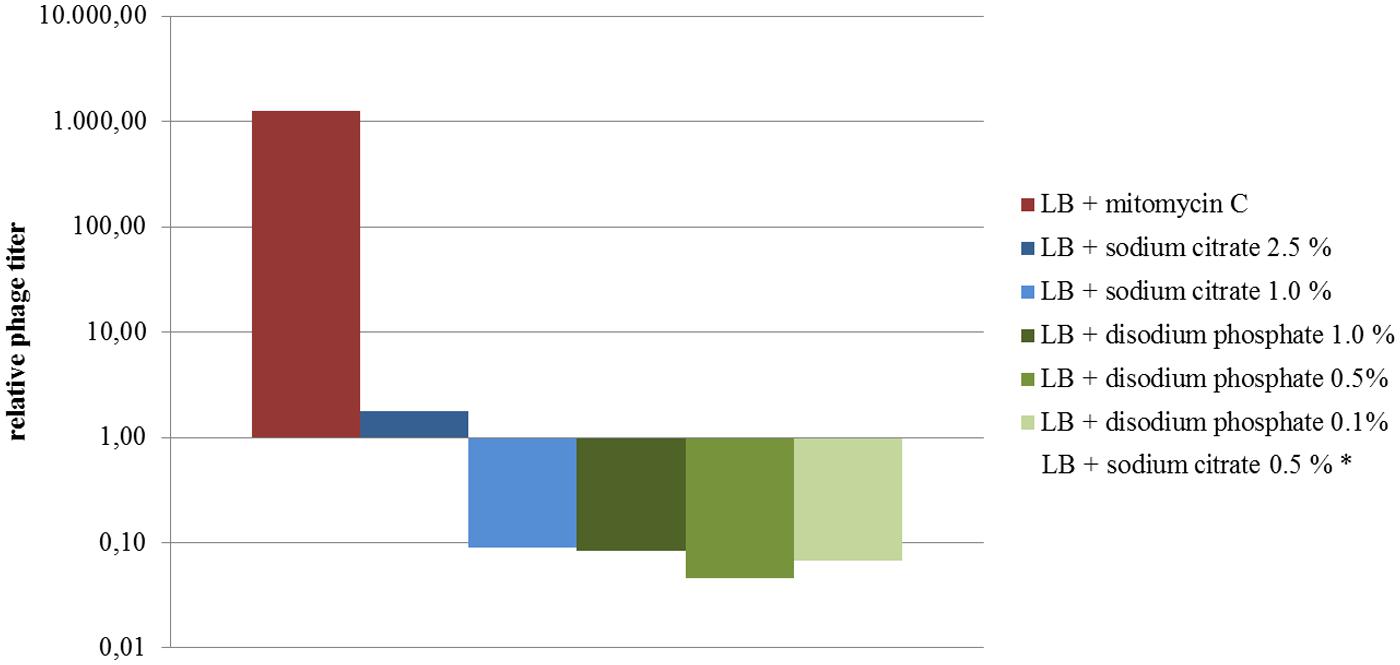
FIGURE 4. Levels of phages expressed as the ratio of phage titers in supernatants of treated cultures relative to the untreated culture (results from assay 4 are shown). The relative titers were plotted on a semilogarithmic scale. *Not shown (phage numbers below the accepted range for countable lysis plaques).
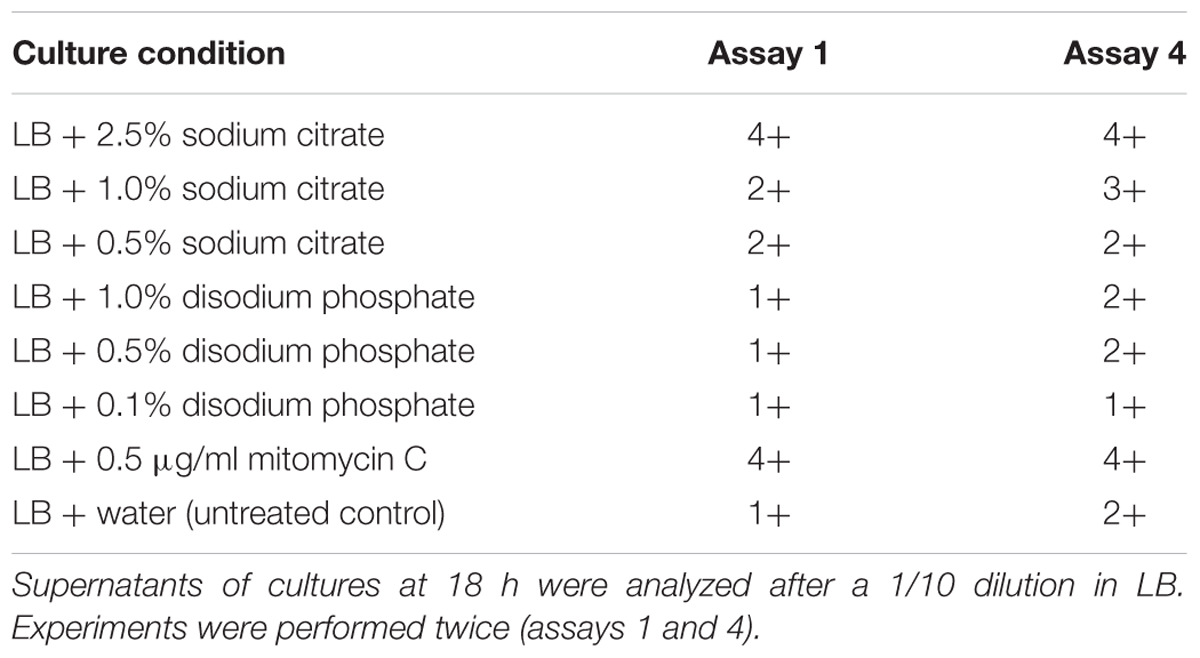
TABLE 1. Semi-quantification of extracellular Shiga toxin by Ridascreen® Verotoxin EIA (R-Biopharm, Germany).
Altogether, the assays showed that sodium citrate and disodium phosphate at the tested concentrations slightly diminished the growth rate of the analyzed STEC strains without evidence of an increment of phage production. However, induction of stx-phage production cannot be ruled out, as phage can be produced but not released, or produced as non-infectious particles. Imamovic and Muniesa (2012) showed that chelation and the increase in stx2-phage induction are linked. In that study, culture treatment with 0.2 M sodium citrate showed effect on stx2 phage induction. Interestingly, our results showed an increase in Stx production in cultures treated with 2.5% sodium citrate in relation to the untreated control. The fact that production of Stx was increased in that condition could reinforce the previous arguments that stx phage production could be induced but not detected, or it could suggest Stx production independent of a complete phage production. It is important to note that EDL933 carries stx1 and stx2 genes, and both Stx1 and Stx2 toxin types can be detected by the Ridascreen kit. In consequence, we were not able to discriminate between Stx1 and Stx2 production, which are known to have differences in transcription control (Wagner and Waldor, 2002).
In the present study, we have shown that an additive, such as sodium citrate, although not having a strong effect on bacterial growth and phage production, can induce the production of Shiga toxin. Although we evaluated few STEC strains, and these results may not accurately represent the behavior of other strains, the present study alerts for a possible increase of Stx production by STEC in presence of some food additives.
Therefore, we consider that when testing the use of additives or other treatments applied in the food industry to decrease bacterial contamination, it is important to take into account the kind of bacteria that can be present. Particularly, in cases in which the bacteria can harbor phages encoding toxins that could be induced with the treatment.
Author Contributions
LL, LM, JB: performed the experiments, participated in the acquisition, analysis and interpretation of the data, approved the final version of the paper. PL, AK: supervised the laboratory work, participated in the analysis and interpretation of the data, drafted the manuscript, and approved the final version of the paper.
Funding
This work was supported by Fondo para la Investigación Científica y Tecnológica (FONCYT) under Grant PICT N° 2012-2438; and Consejo Nacional de Investigaciones Científicas y Técnicas (CONICET) under Grant PIP 939. LL and LM were fellowship holders from Comisión de Investigaciones Científicas-Provincia de Buenos Aires (CICPBA) and Consejo Interuniversitario Nacional (CIN), respectively. JB is a fellowship holder from CONICET. PL and AK are members of the Research Career of CONICET.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgment
Authors thank M. R. Ortiz and Vet. G. H. Arroyo for their technical assistance.
References
Arthur, T. M., Barkocy-Gallagher, G. A., Rivera-Betancourt, M., and Koohmaraie, M. (2002). Prevalence and characterization of non-O157 Shiga toxin-producing Escherichia coli on carcasses in commercial beef cattle processing plants. Appl. Environ. Microbiol. 68, 4847–4852. doi: 10.1128/AEM.68.10.4847-4852.2002
Arthur, T. M., Bosilevac, J. M., Brichta-Harhay, D. M., Kalchayanand, N., Shackelford, S. D., Wheeler, T. L., et al. (2007). Effects of a minimal hide wash cabinet on the levels and prevalence of Escherichia coli O157:H7 and Salmonella on the hides of beef cattle at slaughter. J. Food Prot. 70, 1076–1079.
Bentancor, A. B., Ameal, L. A., Calviño, M. F., Martinez, M. C., Miccio, L., and Degregorio, O. J. (2012). Risk factors for Shiga toxin-producing Escherichia coli infections in preadolescent school children in Buenos Aires, Argentina. J. Infect. Dev. Ctries. 6, 378–386.
Bosilevac, J. M., and Koohmaraie, M. (2011). Prevalence and characterization of non-O157 Shiga toxin-producing Escherichia coli isolates from commercial ground beef in the United States. Appl. Environ. Microbiol. 77, 2103–2112. doi: 10.1128/AEM.02833-10
Bosilevac, J. M., Nou, X., Barkocy-Gallagher, G. A., Arthur, T. M., and Koohmaraie, M. (2006). Treatments using hot water instead of lactic acid reduce levels of aerobic bacteria and Enterobacteriaceae and reduce the prevalence of Escherichia coli O157:H7 on preevisceration beef carcasses. J. Food Prot. 69, 1808–1813.
Caprioli, A., Morabito, S., Brugère, H., and Oswald, E. (2005). Enterohaemorrhagic Escherichia coli: emerging issues on virulence and modes of transmission. Vet. Res. 36, 289–311. doi: 10.1051/vetres:2005002
Doyle, M. E., Archer, J., Kaspar, C. W., and Weiss, R. (2006). Human Illness Caused by E. coli O157:H7 from Food and Non-Food Sources FRI Briefings Available at: https://fri.wisc.edu/files/Briefs_File/FRIBrief_EcoliO157H7humanillness.pdf.
Eswaranandam, S., Hettiarachchy, N., and Johnson, M. (2004). Antimicrobial activity of citric, lactic, malic, or tartaric acids and nisin-incorporated soy protein film against Listeria monocytogenes, Escherichia coli O157:H7, and Salmonella gaminara. J. Food Sci. 69, 79–84.
Gola, S., Mutti, P., Manganelli, E., Squarcina, N., and Rovere, P. (2000). Behaviour of E. coli O157:H7 strains in model system and in raw meat by HPP: microbial and technological aspects. High Pres. Res. 19, 91–97. doi: 10.1080/08957950008202541
Imamovic, L., Jofre, J., Schmidt, H., Serra-Moreno, R., and Muniesa, M. (2009). Phage-mediated Shiga toxin 2 gene transfer in food and water. Appl. Environ. Microbiol. 75, 1764–1768. doi: 10.1128/AEM.02273-08
Imamovic, L., and Muniesa, M. (2011). Quantification and evaluation of infectivity of Shiga toxin-encoding bacteriophages in beef and salad. Appl. Environ. Microbiol. 77, 3536–3540. doi: 10.1128/AEM.02703-10
Imamovic, L., and Muniesa, M. (2012). Characterizing RecA-independent induction of Shiga toxin 2-encoding phages by EDTA treatment. PLoS ONE 7:e32393. doi: 10.1371/journal.pone.0032393
Karmali, M. A., Petric, M., Lim, C., Fleming, P. C., Arbus, G. S., and Lior, H. (1985). The association between idiopathic hemolytic uremic syndrome and infection by verotoxin-producing Escherichia coli. J. Infect. Dis. 151, 775–782. doi: 10.1093/infdis/151.5.775
Kaspar, C., Doyle, M.E., and Archer, J. (2010). White Paper On Non-O157:H7 Shiga Toxin-Producing E. coli from Meat and Non-Meat Sources. Fri food Saf Rev. Available at: http://www.namif.org/namif/wp-content/uploads/08-402.pdf.
Krüger, A., and Lucchesi, P. M. A. (2015). Shiga toxins and stx phages: highly diverse entities. Microbiology 161, 451–462. doi: 10.1099/mic.0.000003
Long, N. H. B. S., Gál, R., and Buňka, F. (2011). Use of phosphates in meat products. Afr. J. Biotechnol. 10, 19874–19882.
Mainil, J., and Daube, G. (2005). Verotoxigenic Escherichia coli from animals, humans and foods: who’s who? J. Appl. Microbiol. 98, 1332–1344. doi: 10.1111/j.1365-2672.2005.02653.x
Martin, A., and Beutin, L. (2011). Characteristics of Shiga toxin-producing Escherichia coli from meat and milk products of different origins and association with food producing animals as main contamination sources. Int. J. Food Microbiol. 146, 99–104. doi: 10.1016/j.ijfoodmicro.2011.01.041
McGannon, C. M., Fuller, C. A., and Weiss, A. A. (2010). Different classes of antibiotics differentially influence Shiga toxin production. Antimicrob. Agents Chemother. 54, 3790–3798. doi: 10.1128/AAC.01783-09
Muniesa, M., Blanco, J. E., De Simón, M., Serra-Moreno, R., Blanch, A. R., and Jofre, J. (2004). Diversity of stx2 converting bacteriophages induced from Shiga-toxin-producing Escherichia coli strains isolated from cattle. Microbiology 150, 2959–2971. doi: 10.1099/mic.0.27188-0
Muniesa, M., Lucena, F., and Jofre, J. (1999). Comparative survival of free Shiga toxin 2-encoding phages and Escherichia coli strains outside the gut. Appl. Environ. Microbiol. 65, 5615–5618.
Over, K., Hettiarachchy, N., Johnson, M., and Davis, B. (2009). Effect of organic acids and plant extracts on Escherichia coli O157:H7, Listeria monocytogenes, and Salmonella Typhimurium in broth culture model and chicken meat systems. J. Food Sci. 74, M515-M521. doi: 10.1111/j.1750-3841.2009.01375.x
Padola, N. L., Sanz, M. E., Blanco, J. E., Blanco, M., Blanco, J., Etcheverria, A. I., et al. (2004). Serotypes and virulence genes of bovine Shigatoxigenic Escherichia coli (STEC) isolated from a feedlot in Argentina. Vet. Microbiol. 100, 3–9. doi: 10.1016/S0378-1135(03)00127-5
Parma, A., Sanz, M., Blanco, J., Blanco, J., Viñas, M., Blanco, M., et al. (2000). Virulence genotypes and serotypes of verotoxigenic Escherichia coli isolated from cattle and foods in Argentina. Eur. J. Epidemiol. 16, 757–762. doi: 10.1023/A:1026746016896
Paton, A. W., and Paton, J. C. (1998). Detection and characterization of Shigatoxigenic Escherichia coli by using multiplex PCR assays for stx1, stx2, eaeA, enterohemorrhagic E. coli hlyA, rfb0111, and rfb0157. J. Clin. Microbiol. 36, 598–602.
Probert, W. S., McQuaid, C., and Schrader, K. (2014). Isolation and identification of an Enterobacter cloacae strain producing a novel subtype of Shiga toxin type 1. J. Clin. Microbiol. 52, 2346–2351. doi: 10.1128/JCM.00338-14
Rode, T. M., Axelsson, L., Granum, P. E., Heir, E., Holck, A., and Trine, M. (2011). High stability of Stx2 phage in food and under food-processing conditions. Appl. Environ. Microbiol. 77, 5336–5341. doi: 10.1128/AEM.00180-11
Sammel, L. M., Claus, J. R., Greaser, M. L., and Richards, M. P. (2006). Investigation of mechanisms by which sodium citrate reduces the pink color defect in cooked ground turkey. Meat Sci. 72, 585–595. doi: 10.1016/j.meatsci.2005.09.008
Schmidt, H., Bielaszewska, M., and Karch, H. (1999). Transduction of enteric Escherichia coli isolates with a derivative of Shiga toxin 2-encoding bacteriophage φ3538 isolated from Escherichia coli O157:H7. Appl. Environ. Microbiol. 65, 3855–3861.
Wagner, P. L., and Waldor, M. K. (2002). Bacteriophage control of bacterial virulence. Infect. Immun. 70, 3985–3993. doi: 10.1128/IAI.70.8.3985-3993.2002
Wong, C. S., Jelacic, S., Habeeb, R. L., Watkins, S. L., and Tarr, P. I. (2000). The risk of the hemolytic–uremic syndrome after antibiotic treatment of Escherichia coli O157:H7 infections. N. Eng. J. Med. 342, 1930–1936. doi: 10.1056/NEJM200006293422601
Keywords: STEC, Shiga toxin, bacteriophages, food additives, sodium citrate, disodium phosphate
Citation: Lenzi LJ, Lucchesi PMA, Medico L, Burgán J and Krüger A (2016) Effect of the Food Additives Sodium Citrate and Disodium Phosphate on Shiga Toxin-Producing Escherichia coli and Production of stx-Phages and Shiga toxin. Front. Microbiol. 7:992. doi: 10.3389/fmicb.2016.00992
Received: 24 February 2016; Accepted: 09 June 2016;
Published: 23 June 2016.
Edited by:
Avelino Alvarez-Ordóńez, Teagasc Food Research Centre, IrelandReviewed by:
Panagiotis Skandamis, Agricultural University of Athens, GreeceJinshui Zheng, Huazhong Agricultural University, China
Copyright © 2016 Lenzi, Lucchesi, Medico, Burgán and Krüger. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Paula M. A. Lucchesi, cGF1bGFsdWNAdmV0LnVuaWNlbi5lZHUuYXI=