 Hasmik Hayrapetyan
Hasmik Hayrapetyan Roland Siezen2,3,4
Roland Siezen2,3,4 Tjakko Abee
Tjakko Abee Masja Nierop Groot
Masja Nierop Groot- 1Laboratory of Food Microbiology, Wageningen University, Wageningen, Netherlands
- 2Top Institute of Food and Nutrition, Wageningen, Netherlands
- 3Microbial Bioinformatics, NIZO, Ede, Netherlands
- 4Center for Molecular and Biomolecular Informatics, Radboud University Medical Centre, Nijmegen, Netherlands
- 5Wageningen UR Food and Biobased Research, Wageningen, Netherlands
Iron is an important element for bacterial viability, however it is not readily available in most environments. We studied the ability of 20 undomesticated food isolates of Bacillus cereus and two reference strains for capacity to use different (complex) iron sources for growth and biofilm formation. Studies were performed in media containing the iron scavenger 2,2-Bipyridine. Transcriptome analysis using B. cereus ATCC 10987 indeed showed upregulation of predicted iron transporters in the presence of 2,2-Bipyridine, confirming that iron was depleted upon its addition. Next, the impact of iron sources on growth performance of the 22 strains was assessed and correlations between growth stimulation and presence of putative iron transporter systems in the genome sequences were analyzed. All 22 strains effectively used Fe citrate and FeCl3 for growth, and possessed genes for biosynthesis of the siderophore bacillibactin, whereas seven strains lacked genes for synthesis of petrobactin. Hemoglobin could be used by all strains with the exception of one strain that lacked functional petrobactin and IlsA systems. Hemin could be used by the majority of the tested strains (19 of 22). Notably, transferrin, ferritin, and lactoferrin were not commonly used by B. cereus for growth, as these iron sources could be used by 6, 3, and 2 strains, respectively. Furthermore, biofilm formation was found to be affected by the type of iron source used, including stimulation of biofilms at liquid-air interphase (FeCl3 and Fe citrate) and formation of submerged type biofilms (hemin and lactoferrin). Our results show strain variability in the genome-encoded repertoire of iron-transporting systems and differences in efficacy to use complex iron sources for growth and biofilm formation. These features may affect B. cereus survival and persistence in specific niches.
Introduction
Iron is one of the essential elements required for growth and metabolism of the majority of microorganisms. Despite its important role in microbial cells, the availability of free iron in the environment is limited due to oxidation of ferrous iron to ferric ions which precipitate near neutral pH (Ratledge and Dover, 2000). Free ferrous iron can be toxic to mammals due to formation of oxygen radicals, consequently the majority of host iron is bound to transport molecules such as hemoglobin (red blood cells), transferrin (serum), and lactoferrin (milk and mucosal secretions), or to ferritin-like proteins for intracellular iron storage (Ratledge and Dover, 2000). The storage of iron in complexed form also reduces its availability for invading pathogenic microorganisms. However, many pathogens developed mechanisms to overcome iron scarcity by the expression of scavenging systems specific to complex and non-complex iron sources. Two main scavenging mechanisms for iron have been described. Bacteria may secrete specific molecules with high affinity to iron named siderophores (Ratledge and Dover, 2000; Zawadzka et al., 2009) that facilitate iron transport into the microbial cell. These siderophores sequester iron from different sources such as transferrin (Abergel et al., 2008). The second mechanism involves specific ABC-type transporters encompassing high-affinity surface receptors specific for either complex iron compounds or free iron (Brown and Holden, 2002; Daou et al., 2009). B. cereus genomes encode several putative ABC transporters for complexed iron including ferric citrate (Harvie and Ellar, 2005; Fukushima et al., 2012) and ferrichrome, and several others of unknown substrate specificity (Hotta et al., 2010). Furthermore, a possible interplay between different molecules has been suggested. For example the heme-binding surface protein IlsA in B. cereus also serves as ferritin receptor and assists in ferritin-iron sequestration by bacillibactin siderophore (Segond et al., 2014). IlsA has also been shown to transfer bound hemin to another surface iron transporting molecule of the IlsA system IsdC (Abi-Khalil et al., 2015).
For B. cereus, two different siderophores, bacillibactin (BB), and petrobactin (PB) (Wilson et al., 2006) have been identified. PB is the main siderophore for B. anthracis (Koppisch et al., 2005) and important for its virulence since it is not recognized by the innate immune system (Abergel et al., 2006). In B. cereus, BB seems to be of higher importance in virulence compared to PB based on experiments in an insect model (Segond et al., 2014).
B. cereus has been reported to use various iron sources for growth that are typically present in red blood cells such as hemoglobin (Hb), hemin, and other hemoproteins (Sato et al., 1998, 1999a,b). For B. cereus ATCC 14579, the use of ferritin as an iron source has been described (Daou et al., 2009). Concerning the use of transferrin by different B. cereus strains, contradictory reports have been published that conceivably links to strain variability (Sato et al., 1998; Park et al., 2005; Daou et al., 2009) and pointing to the importance to take strain diversity into account in studies on iron metabolism. Lactoferrin, an iron source typically present in milk, cannot be used by B. cereus and inhibits its growth when present in high concentrations (Sato et al., 1999b; Daou et al., 2009). Ferric citrate, an iron source formed from citric acid which is commonly present in milk and citrus fruits, can also be used by B. cereus (Fukushima et al., 2012). These iron sources can be encountered in different environments including soil, food and processing environments, and mammals or insects. The ability to use these sources largely determines the fitness of bacteria and capacity to adapt to specific niches.
Besides its important role as essential element for bacterial growth and virulence (Cendrowski et al., 2004; Harvie et al., 2005; Porcheron and Dozois, 2015), iron has also been reported to affect biofilm formation (Porcheron and Dozois, 2015). It was recently shown that air-liquid biofilm formation by a selection of B. cereus food isolates was stimulated by addition of FeCl3 (Hayrapetyan et al., 2015a). Biofilm formation may serve as survival mechanism in different environments and can be an important factor contributing to host colonization. To our knowledge, the impact of different (complex) iron sources on biofilm formation capacity and type of biofilms formed including submerged or surface-attached liquid-air biofilms, has not been reported for this species.
In this study we investigated the use of different iron sources by 22 B. cereus strains in relation to their genome content. Expression of the iron transporters in iron deplete and replete conditions was studied in the reference strain ATCC 10987. Since the ability of B. cereus to form biofilms contributes to its persistence in environment and free iron availability is important for biofilm formation of B. cereus (Hayrapetyan et al., 2015a), we also studied the effect of iron sources encountered in different environments on biofilm formation.
Results
Iron Transporting Systems Presence and Expression
Genomes of 20 food isolates and 2 reference B. cereus strains ATCC 14579 and ATCC 10987 were analyzed for genes with predicted function in iron transport (Figure 1A and Table 1). Genes encoding for synthesis of siderophore BB structural components (dhbACBEF) and transporters were present in all strains, while PB biosynthesis genes (asbABCDEF) were absent in seven of the 22 strains analyzed. For five strains, PB biosynthesis genes were present but a functional fpuA/fhuB gene cluster necessary for PB uptake was lacking. However, another permease (fatCD) with a redundant function with fhuB (Dixon et al., 2012), was identified in all the strains in a cluster together with ATP- and substrate-binding proteins (BC5103–5106). Interestingly BC4416, a fhuD-like putative iron compound binding protein with unknown specificity (Hotta et al., 2010) was absent in the strains that also lacked PB siderophore biosynthesis genes, which could indicate a role for this protein in PB transport.
Figure 1. (A) Hierarchical clustering of 22 B. cereus strains based on gene repertoire encoding iron transporters. Clustering was performed using Genesis software (Sturn et al., 2002). (B) Expression of genes encoding iron transporters in B. cereus ATCC 10987 in BHI, BHI+Bip, BHI+FeCl3, and BHI+Bip+FeCl3 at exponential growth phase (t = 5 h). Up-regulated genes are presented in red, down-regulated genes in green and unaffected genes in black. The scale −10 to 10 is based on log2 values of expression ratios compared to BHI.
Table 1. Genes and their predicted function in iron transport in B. cereus.
The IlsA-system acts as a hemophore, and is encoded by the ilsA gene (BC1331) and an isd-like operon consisting of the ABC-transporter (BC4544-4546), sortase (BC4543), heme degrading monooxygenase (BC4542), and heme transport associated proteins BC4547, BC4548, and BC4549 (IsdC) in B. cereus ATCC 14579 (Daou et al., 2009). Genes encoding the IlsA system are present in all strains. In B4079 the IlsA protein appears to be truncated and non-functional due to a point mutation in the encoding gene that creates a premature stop codon. In B4147 the IlsA also appears to be ineffective due to a large internal deletion identified in the encoding gene (both verified with PCR and sequencing). The transport associated protein (BC4547) was identified as a pseudogene in eight strains. Interestingly, the other transport associated secreted component of this system BC4548, which may function as a hemophore that captures heme from Hb and has 98% identity to isdX1 of B. anthracis (Daou et al., 2009), was absent in 11 strains.
Several other known iron ABC-transporters, such as an iron (III) dicitrate-binding complex (fhuD, fecCDE), a ferrichrome-binding complex (feuA/fhuGB), a fepC/fhuGD complex and a fepBC/fhuGB complex, were present in all strains. The feuA/fhuGB complex, known to bind ferric citrate in B. cereus (Fukushima et al., 2012), was only absent in strain B4088. The putative iron-binding protein yfiY (BC2208) was identified in all strains. Three additional systems, two of which encode ferrous iron transport FeoB-FeoA proteins (Kim et al., 2012), were identified in all B. cereus strains. Besides iron uptake genes, proteins involved in iron storage in bacteria, as for example the ferritin-like di-iron-binding proteins of the Dps family (DNA protection during starvation; Tu et al., 2012) were considered. Five genes with putative function in iron storage were identified and were present in most strains with a few exceptions (Figure 1A). The global regulator of iron uptake Fur (Harvie et al., 2005) was also present in all strains.
Transcriptome analysis of ATCC 10987 in iron replete (BHI+FeCl3; BHI+Bip+FeCl3) and deplete (BHI+Bip) conditions showed significant upregulation of most of the above mentioned genes encoding iron transporters under iron starvation evoked by addition of the scavenger (Figure 1B).
Iron transport genes were upregulated from 8 up to 900 fold (Supplementary Table 1), which was most prominent for the BB biosynthesis genes. Ferritin-like proteins for storage of intracellular iron were not significantly affected. The second ferrous iron transport cluster FeoA/B (BCE4965-4966) was significantly up regulated during iron starvation, indicating that the so called “living fossil” (Hantke, 2003) might still be functional in atmospheric conditions. Upon supplementation with FeCl3, none of these genes were significantly affected, with exception of BCE3769. This was the case also in the presence of Bip together with FeCl3 (with BCE2399 as an exception), showing that addition of iron reversed the iron starvation effect of Bip and support a role in iron transport and metabolism for these genes. These results indicate that iron scavenger Bip can be used to assess the efficacy of alternative (complex) iron sources to support growth of the selected 22 strains.
Iron Sources and Growth
The ability of B. cereus strains to use different iron sources for growth was tested in LB+Bip medium (Figures 2–4). The capacity to cope with iron starvation varied highly among the different strains (Figures 3, 4). Notably, growth of all strains was restored in the presence of either Fe citrate or FeCl3 by 80–135% according to growth index (GI) values. All strains, except B4079, could grow with Hb as sole iron source and restored growth to levels ranging from 43% for strain B4078, up to 90% for strain B4117, compared to control conditions (LB medium).
Figure 2. Phenotypic hierarchical clustering of 22 B. cereus strains based on ability to grow on different iron sources. Strains were clustered using the Genesis software (Sturn et al., 2002).
Figure 3. Growth of strains B4079 (A) and ATCC 14579 (B) in LB and LB supplemented with iron scavenger (LB+Bip) with and without addition of different iron sources. Presented values are averages of 3 independent experiments with standard deviation. Data points for LB for both strains are very close to FeCl3 and Fe citrate. For B4079 (A) data points of LB+Bip are very close to LB+Bip+Lactoferrin, LB+Bip+Transferrin, LB+Bip+Ferritin and LB+Bip+Hemin. For ATCC 14579 (B) LB+Bip data points are very close to LB+Bip+Lactoferrin; LB+Bip+Ferritin is close to LB+Bip+Transferrin; LB+Bip+Hb is close to LB+Bip+Hemin.
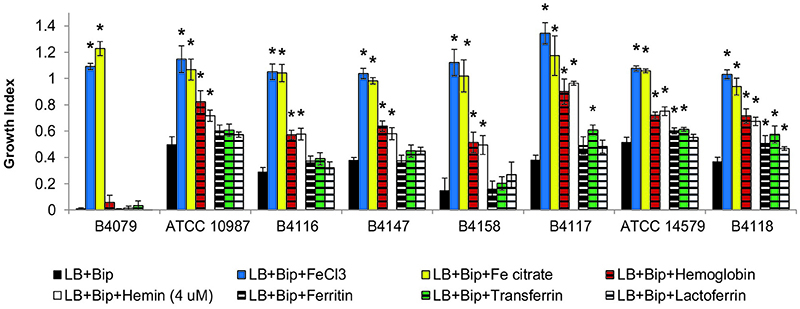
Figure 4. Growth indexes for selected strains in LB, LB supplemented with iron scavenger (LB+Bip) with and without supplementation with different iron sources. Growth indexes represent the ratio of OD(600 nm) reached after 10 h of growth with the corresponding iron source relative to the OD reached in LB. Asterix (*) indicates significant difference (p < 0.01) from iron depleted condition (LB+Bip) for each strain, indicating that the strain could grow with the supplemented iron source.
Hemin could be used by all except three strains (B4077, B4079, B4080). Notably, bacteria that use heme as an iron source also have to cope with its toxicity. This is achieved by a tight control of heme transport, biosynthesis, and degradation. All strains harbored genes to synthesize protoheme and heme, as well as genes encoding the heme efflux ABC transporter HrtA-HrtB, and the associated two-component system HssS-HssR (Stauff and Skaar, 2009; not shown). Only in strain B4158 the latter gene cluster appeared impaired due to an internal deletion, and this strain was among those most sensitive to hemin, along with B4118 and B4147 that were inhibited at higher hemin concentrations (Figure 5).
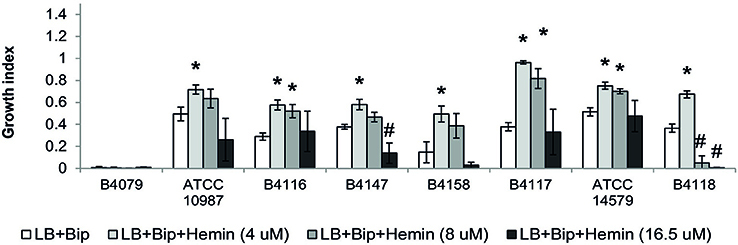
Figure 5. Growth of selected B. cereus strains on different concentrations of hemin. Growth is expressed as the growth indexes. Asterix (*) indicates significant difference (p < 0.01) from growth index in LB+Bip for each strain showing that the strain could grow on the specified concentration of hemin, while the hash (#) shows that the growth was significantly inhibited. For all of the presented strains, 4 uM hemin was the optimal concentration for growth, with the exception of strain B4079 which did not grow on this iron source with any concentration.
Transferrin and ferritin could be used by six and three strains, respectively (Figure 2), and both compounds restored growth to a maximum of 60% of the control. Lactoferrin was a poor iron source for most strains and could only be used by strains B4082 and B4118 (Figure 2) albeit that growth was restored to a maximum of 47% of the controls (not shown).
Linking Genotypes with Growth Phenotypes
The growth performance data on different iron sources and genome contents were clustered (Figures 2, 1A). Four main clusters could be distinguished but phenotypes did not match fully with predicted capacity based on gene content. B4079 showed poorest growth in iron-depleted condition and with complex iron sources. In line with this observation, B4079 lacks most functional transporters. B4079 clusters separately from the other strains (cluster 1, Figures 1A,B) and based on gene content it is most similar to the subgroup of strains lacking PB encoding genes (cluster 2, Figure 1A). The strains of cluster 2 (Figure 1A), along with the strains missing fpuA/fhuB genes for PB import (cluster 3, Figure 1A), belong to one large phenotypic cluster (cluster 2+3, Figure 2) of strains which can use FeCl3, Fe citrate, Hb, and hemin, but not transferrin, ferritin or lactoferrin. The exceptions are B4077 (no growth on hemin) and B4117 (can use transferrin) which fall out of the phenotypic cluster 2+3. The other five strains that could use more than three of the above mentioned complex iron sources group together based on phenotypes (cluster 4, Figure 2) and they harbor all or most iron transporter genes considered (genotypic cluster 4, Figure 1A). Notably, the other five strains with all the genes present did not match the expected use of complex iron sources. On the other hand, the feuA/fhuGB complex is lacking in strain B4088 which nevertheless can grow on Fe citrate. Overall, the phenotypes for 15 out of 22 strains (70%) corresponded to that predicted based on genome content.
Iron Sources and Biofilm Formation
The ability of the different strains to form biofilms with different types of iron sources was tested on polystyrene microtiter plates. 10 out of 22 tested strains formed a biofilm in LB medium without supplementation (control; Table 2). Removal of free iron with Bip eliminated the biofilm forming capacity of nine of these strains, leaving only strain B4155 positive for biofilm formation. For two strains (B4080 and B4120), biofilm formation was promoted under iron deplete condition (Table 2), even though the growth was reduced. Supplementation with Fe citrate and FeCl3 not only restored but even increased biofilm forming capacity of the above mentioned 10 strains, and additionally triggered biofilm formation by B4087 (Table 2). Hb allowed biofilm formation by 16 strains, among them 6 strains that did not form biofilm in the control condition, albeit the amount of formed biofilm was lower than that formed in presence of FeCl3 or Fe citrate for most of the strains. In the presence of hemin, six strains were able to form biofilm, similar to lactoferrin. These biofilms were completely submerged on the bottom of the well, in contrast to the air-liquid interface biofilm formed in LB, LB+Bip+FeCl3, and LB+Bip+Fe citrate (Figure 6).
Table 2. Biofilm formation in the presence of different iron sources.
Figure 6. Biofilm formation for selected B. cereus strains. The biofilm was formed in polystyrene 96-well-plate in LB and LB supplemented with Bip, with or without addition of different iron sources. The biofilm was measured with the CV assay after 24 h incubation at 30°C.
Discussion
In this study we present data showing the impact of different iron sources on growth and biofilm formation capacity and type of biofilms formed for 20 Bacillus cereus food isolates and two reference strains.
Bacillibactin (BB) and petrobactin (PB) are iron-transporting siderophores produced by Bacillus cereus group members. The relevance of PB in B. anthracis growth and virulence was shown, however for B. cereus BB was suggested to be of more importance (Segond et al., 2014). Notably, BB is present in all the strains in this study, while PB is absent in seven strains.
Limitation of free iron impaired the growth of all tested B. cereus strains in LB+Bip but was most prominent for B4079, lacking both PB siderophore and functional IlsA. This also prevented efficient use of Hb and hemin by this strain, in contrast to strains missing only one of the mentioned systems. Interestingly, strains able to use ferritin or transferrin as iron source encompass the whole repertoire of iron transporters, with only minor exceptions. This is in agreement with the previously suggested cooperation between different systems such as IlsA and petrobactin siderophore in iron uptake from ferritin (Segond et al., 2014).
The ability of B. cereus strains to grow on complex iron sources does not always correspond to the presence of relevant genes. For example, B4120 and B4155 contain the full repertoire of iron transporters, however these strains could not use transferrin, ferritin or lactoferrin as iron sources. This may be explained either by differences in regulation of expression of these genes in the selected conditions, presence of transcriptional activators such as specific iron starvation ECF factors (Visca et al., 2002), or factors that affect translation or activity of the synthesized proteins.
Contradictory data have been reported previously concerning the use of transferrin by B. cereus. According to one report, B. cereus could use human transferrin as an iron source, albeit with lower efficiency compared to Staphylococcus aureus, Escherichia coli, and Pseudomonas aeruginosa (Park et al., 2005). Two other studies report inability of B. cereus to grow on transferrin (Sato et al., 1998; Daou et al., 2009), or growth inhibition of B. cereus and B. anthracis by human transferrin (Sato et al., 1998) due to iron deprivation (Rooijakkers et al., 2010). Our data show that the ability to use human transferrin is strain and concentration dependent, concentrations exceeding 2 uM displayed a bacteriostatic effect on several strains (not shown), while 1.5 uM transferrin was the optimal concentration that could be used by 6 out of 22 strains. Besides, the source of transferrin seems of importance since the S. aureus transferrin receptor was shown to bind preferentially human and rodent transferrin but not that of bovine and porcine origin (Modun and Williams, 1999). Aerobic or anaerobic growth conditions could also play a role since oxygen availability has for example been shown to affect the relative abundance of petrobactin and bacillibactin in B. anthracis (Lee et al., 2011). Furthermore, all the strains used in this study, with the exception of ATCC 14579 (isolated from air in a cow shed) were food isolates. Systemic infections caused by B. cereus (Bottone, 2010; Uchino et al., 2012) are caused by more clinically relevant strains, that likely differ in their ability to use and tolerate high levels of transferrin compared to food isolates. To test this, further studies including clinical isolates should be performed.
Lactoferrin is abundant in milk, but also in blood and secreted fluids such as tears and displays antimicrobial properties (Oram and Reiter, 1968; Sato et al., 1999b; Orsi, 2004). Lactoferrin can be used as an iron source by Pseudomonas ssp. (Xiao and Kisaalita, 1997) and several other microorganisms (Morgenthau et al., 2013), but not by B. cereus as reported previously (Sato et al., 1999b; Daou et al., 2009). The latter study used 1.5 uM of lactoferrin, which in our study also did not restore the growth of any of the 22 strains and inhibited the growth for strain B4086 (not shown). However, a concentration of 0.7 uM lactoferrin slightly restored the growth of two strains (B4082 and B4118), which could also use all other tested iron sources, indicating that these strains were in general better equipped for use of complex iron sources, in line with the full repertoire of iron transporting systems present in these strains. The low number of strains able to use lactoferrin is unexpected given the fact that B. cereus is a common contaminant in dairy products.
The capacity to use different complex iron sources could not be linked to the isolation source of the strains. However, clustering of the strains used in this study according to Guinebretière et al. (2008), revealed that all strains lacking petrobactin encoding genes belong to the phylogenetic group III (Warda et al., in press). A common habitat for strains of group III are dehydrated/starchy foods (Guinebretière et al., 2008). Interestingly, all group III strains in the current study were isolated from a starch or dairy containing food product as reported previously (Hayrapetyan et al., 2015a).
Iron Sources and Biofilm Formation
Previously, we reported that addition of free iron (FeCl3) promoted formation of air-liquid interface biofilms by B. cereus strains. In this study we show that apart from FeCl3 also Fe citrate promoted biofilm formation. Hb triggered biofilm formation for a subset of strains for which the growth was also restored and resulting in partial submerged and air-liquid biofilms. Even strain B4079, which did not show significant growth recovery with Hb, was able to form biofilm upon its addition. It showed very limited growth in the presence of Hb (to OD = 0.05, compared to LB+Bip OD = 0.01, Figure 3), which may have caused stress conceivably linked to biofilm formation as a response. Hb was previously identified as a component in nasal secretions that promoted colonization by S. aureus via repression of the agr quorum sensing system resulting in reduced production of proteases with concomitant reduction in biofilm dispersal (Pynnonen et al., 2011). Interestingly this effect was found to be exerted by the Hb protein independently of its iron content. The mechanism of Hb-induced biofilm formation in B. cereus remains to be elucidated.
Ferritin and transferrin only slightly supported biofilm formation, mostly for strains already able to form biofilm in iron limited conditions (B4080, B4120, and B4155, Table 2). A role for the surface protein IsdC in cell-cell attachment and biofilm formation under iron deplete conditions was shown for Staphylococcus lugdunensis (Missineo et al., 2014). Interestingly, this protein is a homolog of BC4549, encoding a component of the IlsA iron transporting system. Since iron starvation most likely triggers the upregulation of such proteins this may be linked to biofilm-promoting effect of iron depletion for strains B4080 and B4120 (Table 2).
The iron-chelating properties combined with a direct bactericidal effect of lactoferrin has led to its proposed role as potential anti-biofilm compound (Ammons and Copié, 2013). In our study, lactoferrin triggered submerged biofilm formation by B. cereus strains B4158 and ATCC 10987, even though growth was not restored. The underlying mechanism remains to be elucidated.
This study shows that ferric citrate and FeCl3 could be used by all B. cereus strains and were preferred iron sources. Hemoglobin, hemin, transferrin, ferritin and lactoferrin could also act as iron sources but their use appeared to be highly strain-dependent. The ability of B. cereus strains to grow on complex iron sources correlated largely with the genome content, but could not always be linked to specific iron transporter genes present. The ability to use complex iron sources seems to be dictated by the combined presence or absence of more than one functional iron transporting system, rather than one single system. Furthermore, biofilm formation was found to be affected by the type of iron source used, including stimulation of biofilms at liquid-air interphase (FeCl3 and Fe citrate) and formation of submerged type biofilms (hemin and lactoferrin). Notably, generation of submerged biofilms was in some cases linked to lack of growth stimulation by the complex iron source tested. To conclude, our results show strain variability in the genome repertoire of iron-transporting systems and differences in efficacy to use complex iron sources for growth and biofilm formation. These features may affect B. cereus survival and persistence in specific niches including food processing environments and the human host.
Materials and Methods
Strains and Culturing Conditions
Twenty Bacillus cereus food isolates from the NIZO culture collection were used in this study (Hayrapetyan et al., 2015a) along with two reference strains B. cereus ATCC 10987 and ATCC 14579. To obtain overnight cultures, a loop full with stock cultures stored at −80°C was inoculated into 10 ml LB broth (Miller, MERCK), supplemented with 100 μM 2,2-Bipyridine (Bip) (MERCK) to induce iron starvation, and incubated for 18 h at 30°C with shaking at 200 rpm.
The twenty B. cereus food isolates were sequenced by next-generation whole genome sequencing. For eight strains (B4077, B4078, B4080, B4086, B4087, B4147, B4153, B4158), total DNA isolation and sequencing details are described elsewhere (Krawczyk et al., 2015), for the remaining 12 isolates (B4081, B4082, B4083, B4084, B4085, B4088, B4116, B4117 [recently re-classified by NCBI as Bacillus mycoides based on ANI typing (Federhen et al., 2016)], B4118, B4120, B4155, B4079) draft genomes were obtained and deposited as described in Hayrapetyan et al. (2016).
Searching for Iron-Transporting Systems in B. cereus Genomes
Orthologous groups (OGs; i.e., gene families) were determined using OrthoMCL (Enright et al., 2002). This program uses all-against-all protein BLAST where it groups proteins with more homology within the species than homology with proteins outside the species. In this way orthologs (genes in different species that evolved from a common ancestral gene by speciation) are separated from paralogs (genes related by duplication within a genome). In addition to the 20 newly sequenced genomes of food isolates (Krawczyk et al., 2015; Hayrapetyan et al., 2016), the circular genomes of the two reference strains B. cereus ATCC 14579 and ATCC 10987 obtained from the NCBI database, were included. Contigs of the 20 newly sequenced genomes were scaffolded into their presumed correct order using the circular reference genomes as templates.
A database (in MS Excel) was built encompassing information about the location and length of orthologous proteins. Multiple sequence alignment files (MSA) were made (MUSCLE, version 3.8; Edgar, 2004), where the protein sequences within ortholog groups were aligned, to facilitate identification of pseudogenes (encoding incomplete proteins).
A literature search was performed to find known iron-uptake systems for B. cereus (Daou et al., 2009; Zawadzka et al., 2009; Hotta et al., 2010). Orthologous groups (OGs) containing the locus tags of these known genes were searched for in the OG table. Furthermore, a key word search was done to find additional iron uptake and storage systems, by searching in the annotation of all genomes for keywords: iron, ferric, ferrous, ferritin.
For relevant identified OGs containing pseudogenes, which are fragments of genes (i.e. truncated, frame-shifted or at the end of contigs), which had been classified by OrthoMCL into separate OGs adjacent on the chromosome, were combined into single OGs representing all the fragments of a single pseudogene.
The RAST automatic annotation of the encoded proteins was manually improved using InterproScan (http://www.ebi.ac.uk/Tools/pfa/iprscan/), NCBI-BLAST (http://blast.ncbi.nlm.nih.gov/http://blast.ncbi.nlm.nih.gov/) and NCBI/Genbank database for the comparison of genes with other species (http://www.ncbi.nlm.nih.gov/).
Growth and Biofilm Formation
The growth and biofilm formation on different iron sources was tested in LB (as control), LB supplemented with 600 μM 2,2-Bipyridine (LB+Bip) as iron depleted condition, and in iron-replete conditions using LB+Bip with addition of the following iron sources in final concentrations: FeCl3 (250 μM; LB+Bip+FeCl3), ferric citrate (250 μM; LB+Bip+Fe citrate), hemoglobin (human, 2.5 μM; LB+Bip+Hb), hemin (4, 8, and 16.5 μM; LB+Bip+Hemin), ferritin (from equine spleen, 0.9 μM; LB+Bip+Ferritin), transferrin (human, partially saturated, 1.5 μM; LB+Bip+Transferrin), and lactoferrin (bovine milk, 0.7 μM; LB+Bip+Lactoferrin). 2,2-Bipyridine, FeCl3 and ferric citrate were from MERCK and the remaining iron sources used were obtained from SIGMA. Selected concentrations were adapted from previously reported concentrations used for B. cereus (Daou et al., 2009), (Segond et al., 2014), with some optimization for the culturing conditions and strains of this study.
The strains were grown in a 96-well-plate filled with 200 μl LB with or without supplements inoculated with 1% overnight culture. The growth was monitored by measuring the OD at 600 nm in SPECTRAmax (model PLUS384) at 30°C, with shaking for 60 s every 5 min. The growth index (GI) for each iron source was calculated as described elsewhere (Daou et al., 2009), by dividing the OD at 600 nm reached in LB after 10 h of growth by OD reached when grown with the specific iron source.
The biofilms formed in 96-wells-plates inoculated as described above, were measured after 24 h of static incubation at 30°C using the Crystal Violet (CV) assay as described previously (Hayrapetyan et al., 2015a). Washing, staining and de-staining steps were performed using 250 μl of de-mineralized water, 0.1% crystal violet and 70% ethanol, respectively. After de-staining the OD was measured at 595 nm. The strain was considered to form a biofilm if in a given condition the OD value was higher than 0.1, a threshold value as defined in (Hayrapetyan et al., 2015a).
Transcriptome Analysis to Identify Iron-Responsive Genes
For transcriptome analysis RNA was isolated from static liquid cultures of B. cereus ATCC 10987 grown in BHI (control), BHI supplemented with 450 μM Bip (BHI+Bip) for iron deplete condition, BHI supplemented with 250 μM FeCl3 (BHI+FeCl3) and BHI with both Bip and FeCl3 (BHI+Bip+FeCl3) for iron replete conditions, and the latter to test whether iron supplementation could restore effects evoked by iron starvation induced by Bip. These conditions were based on a previous study in our laboratory showing the role of free iron in biofilm formation (Hayrapetyan et al., 2015a). The samples were taken at exponential growth phase (5 h). RNA was isolated as previously described (Hayrapetyan et al., 2015b). Labeling and hybridization were performed as described elsewhere (Mols et al., 2013). Two independent biological replicates were hybridized on the arrays, each sample was used three times and was labeled with the swapped dyes Cy3 and Cy5.
Custom-made array design for B. cereus ATCC 10987 developed by Agilent Technologies (GEO accession number GPL7681; Mols et al., 2010) was used in this study. Microarray scanning and data normalization were performed as previously described (Hayrapetyan et al., 2015b). Genes with more than two fold change in expression and p < 0.05 were considered significantly affected. The processed and raw microarray data is deposited in GEO database under accession number GSE74045.
Statistical Analysis
Presented values are averages of at least three independent experiments with standard deviations. The growth was considered recovered if the growth index of the strain on a specific iron source was significantly different from the growth index of the same strain when grown in LB+Bip without iron supplementation. Significance of the growth differences was concluded based on a two-sided student's t-test, assuming equal variances and a P < 0.01.
Author Contributions
Conceived and designed experiments: TA, MG, and HH. Performed the experiments: HH. Analyzed the data: HH. Performed genomic comparisons: RS. Wrote the paper: HH. All authors read and approved the final manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We thank Jos Boekhorst (NIZO food research) for his assistance with genome database and comparative genomic activities and Michiel Wells (NIZO food research) for the array hybridization design and data normalization. The project was funded by TI Food and Nutrition, a public-private partnership on precompetitive research in food and nutrition. The public partners are responsible for the study design, data collection and analysis, decision to publish, and preparation of the manuscript. The private partners have contributed to the project through regular discussion.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2016.00842
References
Abergel, R. J., Wilson, M. K., Arceneaux, J. E., Hoette, T. M., Strong, R. K., Byers, B. R., et al. (2006). Anthrax pathogen evades the mammalian immune system through stealth siderophore production. Proc. Natl. Acad. Sci. U.S.A. 103, 18499–18503. doi: 10.1073/pnas.0607055103
Abergel, R. J., Zawadzka, A. M., and Raymond, K. N. (2008). Petrobactin-mediated iron transport in pathogenic bacteria: coordination chemistry of an unusual 3,4-catecholate/citrate siderophore. J. Am. Chem. Soc. 130, 2124–2125. doi: 10.1021/ja077202g
Abi-Khalil, E., Segond, D., Terpstra, T., André-Leroux, G., Kallassy, M., Lereclus, D., et al. (2015). Heme interplay between IlsA and IsdC: Two structurally different surface proteins from Bacillus cereus. Biochim. Biophys. Acta 1850, 1930–1941. doi: 10.1016/j.bbagen.2015.06.006
Ammons, M. C., and Copié, V. (2013). Mini-review: lactoferrin: a bioinspired, anti-biofilm therapeutic. Biofouling 29, 443–455. doi: 10.1080/08927014.2013.773317
Bottone, E. J. (2010). Bacillus cereus, a volatile human pathogen. Clin. Microbiol. Rev. 23, 382–398. doi: 10.1128/CMR.00073-09
Brown, J. S., and Holden, D. W. (2002). Iron acquisition by Gram-positive bacterial pathogens. Microbes Infect. 4, 1149–1156. doi: 10.1016/S1286-4579(02)01640-4
Cendrowski, S., Macarthur, W., and Hanna, P. (2004). Bacillus anthracis requires siderophore biosynthesis for growth in macrophages and mouse Virulence 51, 407–417. doi: 10.1046/j.1365-2958.2003.03861.x
Daou, N., Buisson, C., Gohar, M., Vidic, J., Bierne, H., Kallassy, M., et al. (2009). IlsA, a unique surface protein of Bacillus cereus required for iron acquisition from heme, hemoglobin and ferritin. PLoS Pathog. 5:e1000675. doi: 10.1371/journal.ppat.1000675
Dixon, S. D., Janes, B. K., Bourgis, A., Carlson, P. E. Jr., and Hanna, P. C. (2012). Multiple ABC transporters are involved in the acquisition of petrobactin in Bacillus anthracis. Mol. Microbiol. 84, 370–382. doi: 10.1111/j.1365-2958.2012.08028.x
Edgar, R. C. (2004). MUSCLE: a multiple sequence alignment method with reduced time and space complexity. BMC Bioinform. 5:113. doi: 10.1186/1471-2105-5-113
Enright, A. J., Van Dongen, S., and Ouzounis, C. A. (2002). An efficient algorithm for large-scale detection of protein families. Nucleic Acids Res. 30, 1575–1584. doi: 10.1093/nar/30.7.1575
Federhen, S., Rossello-Mora, R., Klenk, H.-P., Tindall, B. J., Konstantinidis, K. T., Whitman, W. B., et al. (2016). Meeting report: GenBank microbial genomic taxonomy workshop (12–13 May, 2015). Stand. Genomic Sci. 11, 15. doi: 10.1186/s40793-016-0134-1
Fukushima, T., Sia, A. K., Allred, B. E., Nichiporuk, R., Zhou, Z., Andersen, U. N., et al. (2012). Bacillus cereus iron uptake protein fishes out an unstable ferric citrate trimer. PNAS 109, 16829–16834. doi: 10.1073/pnas.1210131109
Guinebretière, M. H., Thompson, F. L., Sorokin, A., Normand, P., Dawyndt, P., Ehling-Schulz, M., et al. (2008). Ecological diversification in the Bacillus cereus Group. Environ. Microbiol. 10, 851–865. doi: 10.1111/j.1462-2920.2007.01495.x
Hantke, K. (2003). Is the bacterial ferrous iron transporter FeoB a living fossil? Trends Microbiol. 11, 192–195. doi: 10.1016/S0966-842X(03)00100-8
Harvie, D. R., and Ellar, D. J. (2005). A ferric dicitrate uptake system is required for the full virulence of Bacillus cereus. Curr. Microbiol. 50, 246–250. doi: 10.1007/s00284-004-4442-0
Harvie, D. R., Vílchez, S., Steggles, J. R., and Ellar, D. J. (2005). Bacillus cereus Fur regulates iron metabolism and is required for full virulence. Microbiology 151, 569–577. doi: 10.1099/mic.0.27744-0
Hayrapetyan, H., Muller, L., Tempelaars, M., Abee, T., and Nierop Groot, M. (2015a). Comparative analysis of biofilm formation by Bacillus cereus reference strains and undomesticated food isolates and the effect of free iron. Int. J. Food Microbiol. 200, 72–79. doi: 10.1016/j.ijfoodmicro.2015.02.005
Hayrapetyan, H., Tempelaars, M., Nierop Groot, M., and Abee, T. (2015b). Bacillus cereus ATCC 14579 RpoN (Sigma 54) is a pleiotropic regulator of growth, carbohydrate metabolism, motility, biofilm formation and toxin production. PLoS ONE 10:e0134872. doi: 10.1371/journal.pone.0134872
Hayrapetyan, H., Boekhorst, J., De Jong, A., Kuipers, O., and Nierop Groot M. Abee, T. (2016). Draft whole-genome sequences of eleven Bacillus cereus food isolates. Genome Announc. 4:e00485–16. doi: 10.1128/genomeA.00485-16
Hotta, K., Kim, C. Y., Fox, D. T., and Koppisch, A. T. (2010). Siderophore-mediated iron acquisition in Bacillus anthracis and related strains. Microbiology 156, 1918–1925. doi: 10.1099/mic.0.039404-0
Kim, H., Lee, H., and Shin, D. (2012). The FeoA protein is necessary for the FeoB transporter to import ferrous iron. Biochem. Biophys. Res. Commun. 423, 733–738. doi: 10.1016/j.bbrc.2012.06.027
Koppisch, A. T., Browder, C. C., Moe, A. L., Shelley, J. T., Kinkel, B. A., Hersman, L. E., et al. (2005). Petrobactin is the primary siderophore synthesized by Bacillus anthracis str. Sterne under conditions of iron starvation. Biometals 18, 577–585. doi: 10.1007/s10534-005-1782-6
Krawczyk, A. O., De Jong, A., Eijlander, R. T., Berendsen, E. M., Holsappel, S., Wells-Bennik, M. H., et al. (2015). Next-generation whole-genome sequencing of eight strains of Bacillus cereus, isolated from food. Genome Announc. 3:e01480–15. doi: 10.1128/genomeA.01480-15
Lee, J. Y., Passalacqua, K. D., Hanna, P. C., and Sherman, D. H. (2011). Regulation of petrobactin and bacillibactin biosynthesis in Bacillus anthracis under iron and oxygen variation. PLoS ONE 6:e20777. doi: 10.1371/journal.pone.0020777
Missineo, A., Poto, A. D., Geoghegan, J. A., Rindi, S., Heilbronner, S., Gianotti, V., et al. (2014). IsdC from Staphylococcus lugdunensis induces biofilm formation under low-iron growth conditions. Infect. Immun. 82, 2448–2459. doi: 10.1128/IAI.01542-14
Modun, B., and Williams, P. (1999). The staphylococcal transferrin-binding protein is a cell wall glyceraldehyde-3-phosphate dehydrogenase. Infect. Immun. 67, 1086–1092.
Mols, M., Mastwijk, H., Nierop Groot, M., and Abee, T. (2013). Physiological and transcriptional response of Bacillus cereus treated with low-temperature nitrogen gas plasma. J. Appl. Microbiol. 115, 689–702. doi: 10.1111/jam.12278
Mols, M., Van Kranenburg, R., Van Melis, C. C. J., Moezelaar, R., and Abee, T. (2010). Analysis of acid-stressed Bacillus cereus reveals a major oxidative response and inactivation-associated radical formation. Environ. Microbiol. 12, 873–885. doi: 10.1111/j.1462-2920.2009.02132.x
Morgenthau, A., Pogoutse, A., Adamiak, P., Moraes, T. F., and Schryvers, A. B. (2013). Bacterial receptors for host transferrin and lactoferrin: molecular mechanisms and role in host-microbe interactions. Future Microbiol. 8, 1575–1585. doi: 10.2217/fmb.13.125
Oram, J. D., and Reiter, B. (1968). Inhibition of bacteria by lactoferrin and other iron-chelating agents. Biochim. Biophys. Acta 170, 351–365. doi: 10.1016/0304-4165(68)90015-9
Orsi, N. (2004). The antimicrobial activity of lactoferrin: current status and perspectives. Biometals 17, 189–196. doi: 10.1023/B:BIOM.0000027691.86757.e2
Park, R. Y., Choi, M. H., Sun, H. Y., and Shin, S. H. (2005). Production of catechol-siderophore and utilization of transferrin-bound iron in Bacillus cereus. Biol. Pharm. Bull. 28, 1132–1135. doi: 10.1248/bpb.28.1132
Porcheron, G., and Dozois, C. M. (2015). Interplay between iron homeostasis and virulence: Fur and RyhB as major regulators of bacterial pathogenicity. Veterinary Microbiol. 179, 2–14. doi: 10.1016/j.vetmic.2015.03.024
Pynnonen, M., Stephenson, R. E., Schwartz, K., Hernandez, M., and Boles, B. R. (2011). Hemoglobin promotes Staphylococcus aureus nasal colonization. PLoS Pathog. 7:e1002104. doi: 10.1371/journal.ppat.1002104
Ratledge, C., and Dover, L. G. (2000). Iron metabolism in pathogenic bacteria. Annu. Rev. Microbiol. 54, 881–941. doi: 10.1146/annurev.micro.54.1.881
Rooijakkers, S. H. M., Rasmussen, S. L., Mcgillivray, S. M., Bartnikas, T. B., Mason, A. B., Friedlander, A. M., et al. (2010). Human transferrin confers serum resistance against Bacillus anthracis. J. Biol. Chem. 285, 27609–27613. doi: 10.1074/jbc.M110.154930
Sato, N., Ikeda, S., Mikami, T., and Matsumoto, T. (1999a). Bacillus cereus dissociates hemoglobin and uses released heme as an iron source. Biol. Pharm. Bull. 22, 1118–1121.
Sato, N., Kurotaki, H., Ikeda, S., Daio, R., Nishinome, N., Mikami, T., et al. (1999b). Lactoferrin inhibits Bacillus cereus growth and heme analogs recover its Growth. Biol. Pharm. Bull. 22, 197–199.
Sato, N., Kurotaki, H., Watanabe, T., Mikami, T., and Matsumoto, T. (1998). Use of hemoglobin as an iron source by Bacillus cereus. Biol. Pharm. Bull. 21, 311–314. doi: 10.1248/bpb.21.311
Segond, D., Abi Khalil, E., Buisson, C., Daou, N., Kallassy, M., Lereclus, D., et al. (2014). Iron acquisition in Bacillus cereus: the roles of ilsa and bacillibactin in exogenous ferritin iron mobilization. PLoS Pathog. 10:e1003935. doi: 10.1371/journal.ppat.1003935
Stauff, D. L., and Skaar, E. P. (2009). Bacillus anthracis HssRS signalling to HrtAB regulates haem resistance during infection. Mol. Microbiol. 72, 763–778. doi: 10.1111/j.1365-2958.2009.06684.x
Sturn, A., Quackenbush, J., and Trajanoski, Z. (2002). Genesis: cluster analysis of microarray data. Bioinformatics 18, 207–208. doi: 10.1093/bioinformatics/18.1.207
Tu, W. Y., Pohl, S., Gray, J., Robinson, N. J., Harwood, C. R., and Waldron, K. J. (2012). Cellular iron distribution in Bacillus anthracis. J. Bacteriol. 194, 932–940. doi: 10.1128/JB.06195-11
Uchino, Y., Iriyama, N., Matsumoto, K., Hirabayashi, Y., Miura, K., Kurita, D., et al. (2012). A case series of Bacillus cereus septicemia in patients with hematological disease. Intern. Med. 51, 2733–2738. doi: 10.2169/internalmedicine.51.7258
Visca, P., Leoni, L., Wilson, M. J., and Lamont, I. L. (2002). Iron transport and regulation, cell signalling and genomics: lessons from Escherichia coli and Pseudomonas. Mol. Microbiol. 45, 1177–1190. doi: 10.1046/j.1365-2958.2002.03088.x
Warda, A. K., Siezen, R. J., Boekhorst, J., Wells-Bennik, M. H. J., De Jong, A., Kuipers, O. P., et al. (in press). Linking Bacillus cereus genotypes carbohydrate utilization capacity. PLoS ONE.
Wilson, M. K., Abergel, R. J., Raymond, K. N., Arceneaux, J. E., and Byers, B. R. (2006). Siderophores of Bacillus anthracis, Bacillus cereus, and Bacillus thuringiensis. Biochem. Biophys. Res. Commun. 348, 320–325. doi: 10.1016/j.bbrc.2006.07.055
Xiao, R., and Kisaalita, W. S. (1997). Iron acquisition from transferrin and lactoferrin by Pseudomonas aeruginosa pyoverdin. Microbiology 143, 2509–2515. doi: 10.1099/00221287-143-7-2509
Keywords: Bacillus cereus, iron transport, genotypes, growth, biofilm formation, complex iron sources
Citation: Hayrapetyan H, Siezen R, Abee T and Nierop Groot M (2016) Comparative Genomics of Iron-Transporting Systems in Bacillus cereus Strains and Impact of Iron Sources on Growth and Biofilm Formation. Front. Microbiol. 7:842. doi: 10.3389/fmicb.2016.00842
Received: 24 February 2016; Accepted: 20 May 2016;
Published: 08 June 2016.
Edited by:
Romain Briandet, Institut National de la Recherche Agronomique, FranceReviewed by:
Anne-Brit Kolstø, University of Oslo, NorwayMichel Gohar, Institut National de la Recherche Agronomique, France
Copyright © 2016 Hayrapetyan, Siezen, Abee and Nierop Groot. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tjakko Abee, dGpha2tvLmFiZWVAd3VyLm5s