 Beatriz Padilla1
Beatriz Padilla1 José V. Gil
José V. Gil Paloma Manzanares
Paloma Manzanares
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Microbiol. , 31 March 2016
Sec. Food Microbiology
Volume 7 - 2016 | https://doi.org/10.3389/fmicb.2016.00411
This article is part of the Research Topic Non-conventional yeast in the wine industry View all 15 articles
It is well established that non-Saccharomyces wine yeasts, considered in the past as undesired or spoilage yeasts, can enhance the analytical composition, and aroma profile of the wine. The contribution of non-Saccharomyces yeasts, including the ability to secret enzymes and produce secondary metabolites, glycerol and ethanol, release of mannoproteins or contributions to color stability, is species- and strain-specific, pointing out the key importance of a clever strain selection. The use of mixed starters of selected non-Saccharomyces yeasts with strains of Saccharomyces cerevisiae represents an alternative to both spontaneous and inoculated wine fermentations, taking advantage of the potential positive role that non-Saccharomyces wine yeast species play in the organoleptic characteristics of wine. In this context mixed starters can meet the growing demand for new and improved wine yeast strains adapted to different types and styles of wine. With the aim of presenting old and new evidences on the potential of non-Saccharomyces yeasts to address this market trend, we mainly review the studies focused on non-Saccharomyces strain selection and design of mixed starters directed to improve primary and secondary aroma of wines. The ability of non-Saccharomyces wine yeasts to produce enzymes and metabolites of oenological relevance is also discussed.
Wine fermentation is a complex microbiological process in which yeasts play a fundamental role. Although Saccharomyces cerevisiae is the main microorganism involved in the alcoholic fermentation of grape must, winemaking is a non-sterile process. Many other species of yeasts belonging to various non-Saccharomyces genera occur in grape juice and contribute to the first stages of fermentation and to the organoleptic characteristics of final wine (Fleet, 2008).
In the past, non-Saccharomyces yeasts were considered to be of secondary significance or undesirable spoilage yeasts; nowadays it is widely accepted that selected strains through appropriate screenings can positively impact on the winemaking process. Thus the growing demand for new and improved wine yeast strains adapted to different types and styles of wines can be met by non-Saccharomyces wine yeasts. Since these yeasts are in general poor fermenters, the design of mixed starters including selected non-Saccharomyces with optimized biotechnological characteristics and S. cerevisiae to ensure a complete fermentation has become one of the main challenges of researchers and oenologists. Moreover, proper mixed starter management during fermentation will allow winemakers to tailor wines to the changing demands of consumers.
The production of wines with particular flavor profiles has been one of the main reasons for including non-Saccharomyces yeasts in mixed starters. However, promising approaches to lowering alcohol content of wines, to control wine spoilage or to improve oenological properties are being explored, and undoubtedly they represent new opportunities for exploitation in wine production. Here we revisit the contribution of non-Saccharomyces yeasts to wine aroma complexity. First we review the ability of these yeasts to produce enzymes and metabolites of oenological relevance and finally we discuss the design of mixed starters directed to improve primary and secondary aroma of wines. Special attention was paid to update the information covered in recent reviews on the impact of non-Saccharomyces in wine production.
In the second half of the 19th century, Louis Pasteur revealed the role of yeasts during the wine fermentation process, demonstrating that yeast is the primary catalyst responsible for the conversion of grape sugars to alcohol and CO2. He noticed that in fermenting grape musts coexisted a wide variety of microorganisms, including different types of yeasts. His drawings, based on microscopic observations, showed two kinds of yeasts. The first, which was abundant in the early stages of the process, was the small, apically budding, lemon-shaped Saccharomyces apiculatus (now Hanseniaspora uvarum). The second which became the most abundant as alcoholic fermentation progressed, was a larger yeast with round cells, which Pasteur called either Saccharomyces pastorianus or Saccharomyces ellipsoideus (probably the current S. cerevisiae) (Barnett, 2000).
Despite the complex wine microbial ecology, S. cerevisiae became the wine yeast par excellence based mainly on its fermentation behavior (Reed and Peppler, 1973; Bely et al., 1990; Fleet, 1993), but also on its important role in the release of aroma precursors (Dubourdieu, 1996; Úbeda and Briones, 2000; Ugliano et al., 2006) and in the formation of secondary aroma (Fleet, 1993; Pretorius, 2003). The other yeast species occurring in musts and wines were considered as a source of potential spoilage problems during wine production. In fact, the presence or overgrowth of some of these species was often related to stuck or sluggish fermentations, or to the production of detrimental compounds to the sensory properties of wine (du Toit and Pretorius, 2000). In the context of this simplistic view of the wine fermentation process, where the most important objective was the inoculation and dominance of S. cerevisiae, the term ‘non-Saccharomyces’ yeasts referred to the wide variety of yeast genera, including more than 20 in both Ascomycota and Basidiomycota phyla, present in grape juice.
Yeasts occurring in grape musts at the early stages of fermentation originate from two main sources, the vineyard and the grapes, and the contact surfaces and equipment of the winery (Pretorius et al., 1999). The latter plays a small role as a source of non-Saccharomyces yeasts, while S. cerevisiae is the predominant yeast in such surfaces (Peynaud and Domercq, 1959; Rosini, 1984; Lonvaud-Funel, 1996; Pretorius, 2000). However, it has been recently reported for the first time the implantation in grape must of Hanseniaspora species present in the winery environment (Grangeteau et al., 2015) opening the possibility, still unexplored, that some of the non-Saccharomyces species could persist from 1 year to another in the winery environment and become dominant during fermentation, as usually described for S. cerevisiae (Santamaría et al., 2005; Le Jeune et al., 2006; Mercado et al., 2007).
The great quantitative and qualitative variability of non-Saccharomyces species found in the early stages of fermentation can be explained by the large number of factors influencing the grape microbiota such as localization, climatic conditions, cultivar, application of pesticides, and other agronomic practices, stage of ripening, health of the grapes, harvesting procedures and the specific weather conditions in each vintage year (Martini et al., 1980; Rosini et al., 1982; Querol et al., 1990; Regueiro et al., 1993; Epifanio et al., 1999; Jolly et al., 2006; Brilli et al., 2015). In spite of this wide variability of yeast species, during the first 3–4 days of a spontaneous fermentation of grape must, yeast population is numerically dominated by apiculate yeasts, Hanseniaspora/Kloeckera, and Candida species, followed by several species belonging to the genera Metschnikowia and Pichia, and occasionally to Brettanomyces, Kluyveromyces, Schizosaccharomyces, Torulaspora, Rhodotorula, Zygosaccharomyces, and Cryptococcus genera (Goto, 1980; Benda, 1982; Fleet et al., 1984; Heard and Fleet, 1985; Parish and Caroll, 1985; Martínez et al., 1989; Herraiz et al., 1990; Frezier and Dubourdieu, 1992; Schütz and Gafner, 1993; Granchi et al., 1998; Combina et al., 2005; Fleet, 2008). This scenario, with abundance of apiculate yeasts in the 1st days of alcoholic fermentation and varying amounts of other non-Saccharomyces yeasts, followed by the progressive dominance of S. cerevisiae is a common denominator in the process of elaboration of all wines, including those produced by inoculation with selected wine yeast strains (Heard and Fleet, 1985).
Industrial wine fermentations are currently conducted by starters of selected wine yeast strains of S. cerevisiae. The first reported use of a selected yeast starter for wine production dates from 1890, when Müller-Thurgau introduced this technology adapting the techniques developed by Christian Hansen for the Carlsberg Brewery (Pretorius, 2000; Barnett and Lichtenthaler, 2001). Nowadays, the use of active dry yeasts is one of the most common practices in winemaking and the market offers a wide variety of yeast strains as dehydrated cultures promising a good implantation, specific skills for different types of wines and a great list of other features such as the ability of enhancing varietal and fermentative aromas, glycerol production, tolerance to alcohol, or specific enzymatic activities. However, the main reason of selected starters is to achieve wines with uniform quality through different years avoiding the variability associated with spontaneous fermentations and the risk of spoilage (Beltran et al., 2002; Santamaría et al., 2005). In such cases, dominant growth of the inoculated strain is required. However many factors might affect the implantation/persistence of individual strains within the total population (Fleet, 2008; Blanco et al., 2012), including the variability that exists from one vintage to another at a given winery (Lange et al., 2014).
Despite the advantages of using pure cultures of S. cerevisiae with regard to the easy of control and homogeneity of fermentations, wine produced with pure yeast monocultures lacks the complexity of flavor, stylistic distinction and vintage variability caused by indigenous yeasts (Lambrechts and Pretorius, 2000; Romano et al., 2003). This fact is a never-ending debate between researchers and oenologists, and the growth of non-Saccharomyces yeasts can still be seen as an uncontrollable risk or as an opportunity of improving the quality of wine. Nevertheless it is worthwhile to note that the world’s best quality wines are produced after a fermentation process in which, in a greater or lesser extent, various species of non-Saccharomyces yeasts have played a role in the winemaking process and, therefore, have contributed to the final result. It is in this context where the inclusion of non-Saccharomyces wine yeast species as part of mixed starters together with S. cerevisiae to improve wine quality was suggested as a way of taking advantage of spontaneous fermentations without running the risks of stuck fermentations or wine spoilage (Jolly et al., 2003; Rojas et al., 2003; Romano et al., 2003; Ciani et al., 2006). However this practice is linked to new challenges for researchers and oenologists such as the selection of suitable non-Saccharomyces strains, the appropriate modality and time of inoculation, the proportion of yeasts in the culture and the potential microorganism interactions, among others. Figure 1 shows a schematic outline of spontaneous versus inoculated fermentation and the use of mixed starters of selected non-Saccharomyces yeasts with strains of S. cerevisiae as an alternative to both approaches.
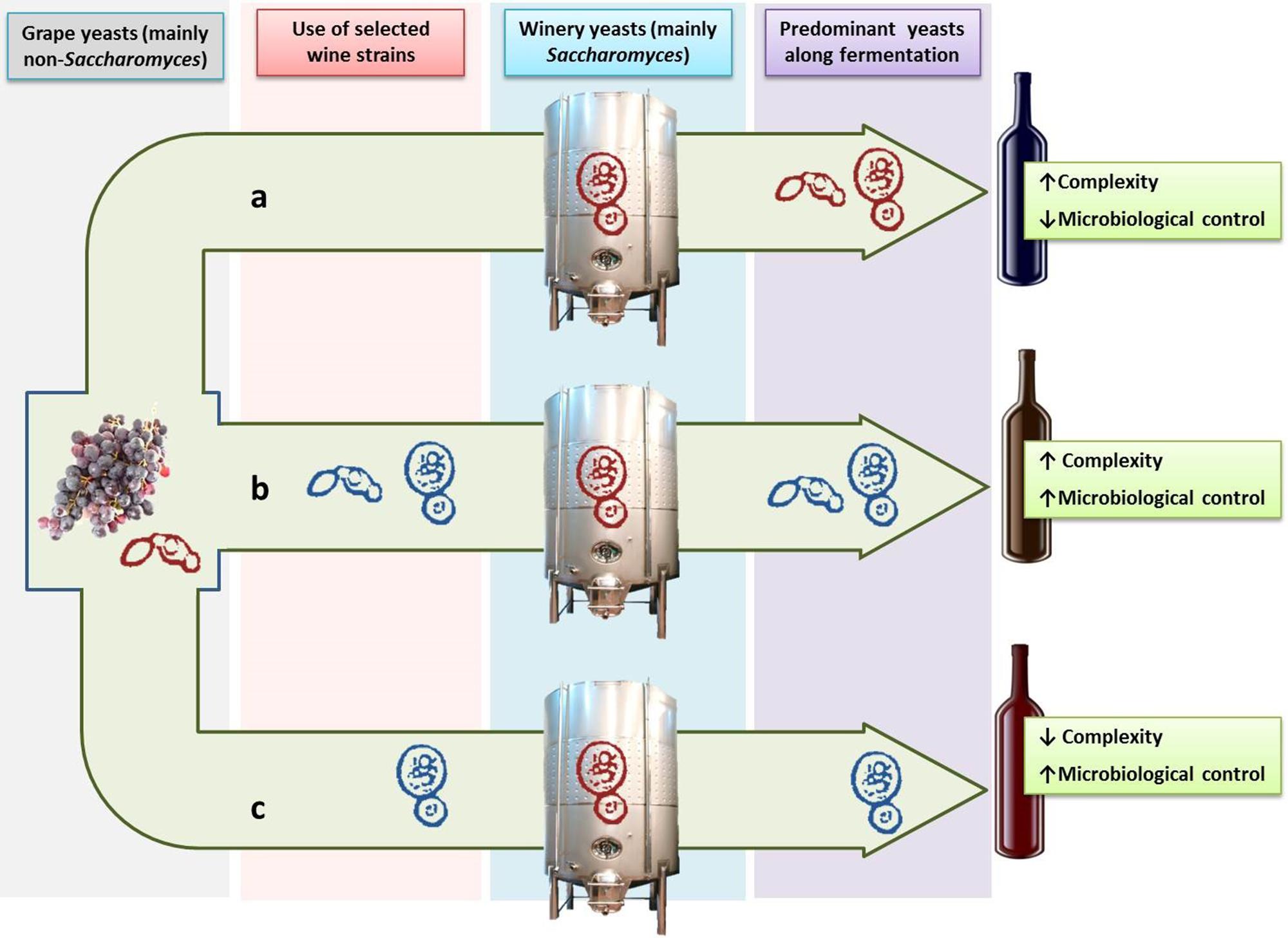
FIGURE 1. Use of selected strains of Saccharomyces and non-Saccharomyces yeasts in winemaking. Spontaneous fermentation (a) allows the development of indigenous yeasts from grapes (mainly non-Saccharomyces) and winery (mainly Saccharomyces) leading to wines with a greater aromatic complexity but with less microbiological control. Inoculation with a selected strain of Saccharomyces cerevisiae (c) leads to a greater microbiological control but can reduce the aromatic complexity of wine. The use of mixed cultures of S. cerevisiae and non-Saccharomyces selected strains (b) allows to obtain wines with both greater aromatic complexity and microbiological control of the process. Autochthonous and inoculated selected yeasts are represented in red and blue color, respectively.
Undoubtedly, aroma is one of most important characteristics that contribute to the quality of wine. As in many foods, wine aroma is composed by 100s of different compounds with concentrations that can vary between 10-1 and 10-10 g/kg (Rapp and Mandery, 1986). The balance and interaction of all of them determine the wine aromatic quality.
Wine aroma can be subdivided into three groups: the varietal or primary aroma, determined by the grape variety; the fermentation or secondary aroma; and the bouquet or tertiary aroma resulting from the transformation of aromas during aging. Non-Saccharomyces yeasts can influence both the primary and secondary aroma through the production of enzymes and metabolites, respectively.
Primary or varietal aroma is formed during the ripening of grapes and its contribution to the final wine aroma is considered an appreciated feature. The production of active compounds of primary wine odor takes place in the exocarp of the grape berry and its final concentration in wine is primarily influenced by the vine variety and secondarily by the state of ripeness and the agronomic and oenological practices (Ewart et al., 1985; Spayd et al., 2002; Hernández-Orte et al., 2008, 2015).
Compounds forming primary aroma belong to a limited number of chemical families, including methoxypyrazines, C13-norisoprenoids, volatile sulfur compounds, and terpenes (Ebeler and Thorngate, 2009). Methoxypyrazines are products of amino acid metabolism, and they have been associated to vegetal, green, and herbaceous aromas in certain vine cultivars (reviewed in Sidhu et al., 2015). C13-norisoprenoids derive from carotenoids and particularly β-ionone and β-damascenone are considered impact volatiles of non-floral grapes (Fang and Qian, 2006; Bindon et al., 2007; Pineau et al., 2007; Ristic et al., 2010; Fang and Qian, 2016). Certain organic volatile sulfur compounds such as aromatic thiols make important contributions to Sauvignon Blanc and red cultivars aroma (Darriet et al., 1995; Tominaga et al., 1996, 1998a; Bouchilloux et al., 1998), whereas terpenoids, although present in grapes of all vine varieties, occur in aromatic varieties such as Muscat, Gewürztraminer and Rhine Riesling in the highest concentrations (King and Dickinson, 2000). In grape berries and corresponding wines, approximately seventy terpenoid compounds have been identified (Mateo and Jiménez, 2000). Among them, five monoterpenoid alcohols, namely linalool, geraniol, nerol, citronellol, and α-terpineol are the most abundant and the strongest contributors to wine aroma (Rapp, 1998; Mateo and Jiménez, 2000; Carrau et al., 2005). These compounds provide floral notes and have low odor thresholds (Zalacain et al., 2007).
Interestingly most of primary aroma compounds are found in free or bound forms. The latter are not odorant compounds which hydrolysis can occur during fermentation through the action of wine yeasts (Figure 2). Particularly important are aroma precursors linked to sugar molecules, mainly terpenol and C13-norisoprenoid glycosides, and the non-volatile precursor forms of volatile thiols conjugated to cysteine or glutathione. The main yeast enzymes involved in the release of aroma compounds from odorless grape precursors are glycosidases that hydrolyze the non-volatile glycosidic precursors (Gunata et al., 1988), and carbon-sulfur lyases that release volatile thiols from aroma-inactive cysteine-bound conjugates (Tominaga et al., 1998b). Below we focus on the production of these enzymes of oenological relevance by non-Saccharomyces wine yeasts. Table 1 summarizes the yeast species described as producers of glycosidases and carbon-sulfur lyases.
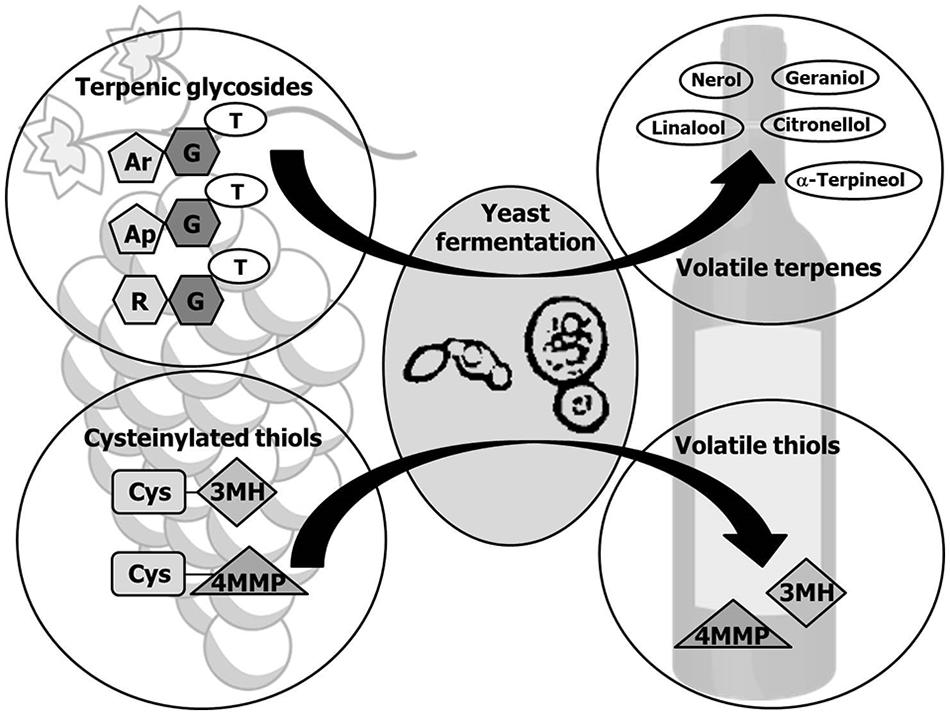
FIGURE 2. Release of primary aroma compounds by yeasts. Monoterpenes and volatile thiols occur in grape as odorless precursors which can be released by enzymatic activities of Saccharomyces and non-Saccharomyces yeasts during fermentation. Monoterpene glycosides are mainly glucosides and diglycosides, in which the glucose moiety (G) has been further substituted mainly with arabinose (Ar), apiose (Ap), or rhamnose (R). A two-step enzyme-catalyzed reaction is the main mechanism proposed for the enzymatic hydrolysis of the diglycosides and subsequent release of the free volatile terpene (T) to wine. First a specific glycosidase cleaves the linkage between both sugars and in a second step the released glucoside is hydrolyzed by a β-D-glucosidase, liberating glucose and the corresponding terpene. Volatile thiols are generated from the odourless cysteinylated precursors cysteine-3-mercaptohexan-1-ol (Cys-3MH) and cysteine-4-mercapto-4-methylpentan-2-one (Cys-4MMP) by the action of carbon-sulfur-lyases.
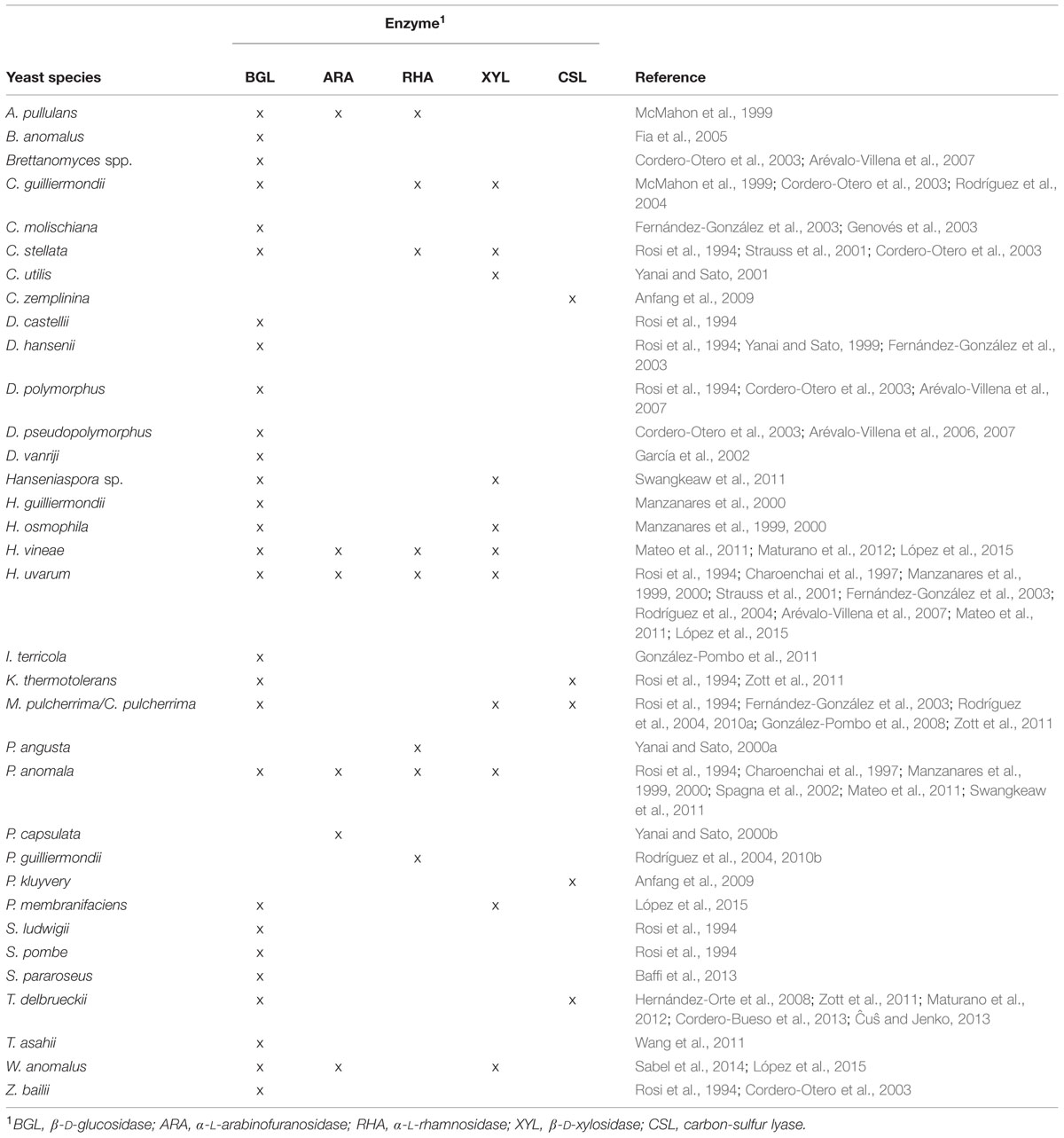
TABLE 1. Non-Saccharomyces yeast species described as producers of enzymes involved in the release of aroma compounds from grape precursors.
Since the demonstration that the aromatic components of certain grape varieties are present in the grape berry both in free form and bound to sugars as glycosides (Cordonnier and Bayonove, 1974; Williams et al., 1982), there has been a continuous research to find glycosidases able to release varietal aromas from precursors. The bound aroma fraction comprises glucosides and diglycosides, and compounds such as terpenols, terpene diols, 2-phenylethyl alcohol, benzyl alcohol and C13-norisoprenoids have been shown to be aglycons of such glycosides (Winterhalter and Skouroumounis, 1997). Diglycosides mainly include 6-O-α-L-arabinofuranosyl-β-D-glucopyranosides, 6-O-α-L-rhamnopyran-osyl-β-D-glucopyranosides and 6-O-β-D-apiofuranosyl-β-D-glucopyranosides. Due to the important role of monoterpenes in determining the aroma of grapes and wines, hydrolysis of terpene glycosides has been the main focus of research. It is now well established that the enzymatic hydrolysis occurs in two steps (Gunata et al., 1988). During the first step and depending on the conjugate, the glycosidic linkage is cleaved by either an α-L-arabinofuranosidase, an α-L-rhamnosidase or a β-D-apiosidase, and the corresponding monoterpenyl-β-D-glucosides are released. In the second step, monoterpenes are liberated by the action of a β-D-glucosidase.
Although oenological yeasts may produce glycosidases, the potential effectiveness of enzymes may be hampered by acidic wine conditions or high ethanol concentrations; another limitation of these enzymes is their weak activity in the presence of glucose in the must or wine, making it especially necessary to analyze their inhibition by these wine components. The potential effectiveness of yeast-derived glycosidases is even further reduced in most cases by the fact that some of the enzymes are intracellular and released only in very small amounts into the culture medium. The degree to which these factors inhibit glycosidase production and activity depends on the species and strains of the organisms involved, pointing out the need of enzyme screenings.
Widespread occurrence of β-D-glucosidase activity in non-Saccharomyces yeasts has been revealed in several screenings. Rosi et al. (1994) showed that yeasts of the genera Candida, Debaryomyces, Hanseniaspora/Kloeckera, Kluyveromyces, Metschnikowia, Pichia, Saccharomycodes, Schizosaccharomyces, and Zygosaccharomyces can produce β-D-glucosidases. Later on this capability was confirmed by other authors (Charoenchai et al., 1997; McMahon et al., 1999; Manzanares et al., 2000; Strauss et al., 2001; Spagna et al., 2002; Cordero-Otero et al., 2003; Fernández-González et al., 2003; Rodríguez et al., 2004; González-Pombo et al., 2008; Sabel et al., 2014; López et al., 2015) and was extended also to the genera Torulaspora (Hernández-Orte et al., 2008; Cordero-Bueso et al., 2013), Brettanomyces (Cordero-Otero et al., 2003; Fia et al., 2005; Arévalo-Villena et al., 2007), and Trichosporon (Wang et al., 2011). Some of these enzymes, selected as a result of their activity with artificial substrates, proved also effective in hydrolyzing either a grape glycoside extract or in releasing terpenols after addition to must or wine. Debaryomyces hansenii and H. uvarum β-D-glucosidases hydrolyzed terpenic glycosides isolated from grape must (Rosi et al., 1994; Fernández-González et al., 2003). Terpene release was also observed in must and wine treated with β-D-glucosidases from Hanseniaspora sp. and Pichia anomala (Swangkeaw et al., 2011). Moreover, results suggested that the enzyme from Hansenispora sp. was more efficient in releasing desirable aromas during an early stage of alcoholic fermentation while β-D-glucosidase from P. anomala was suitable at the final stage. Several yeast β-D-glucosidases have been purified and characterized. Two Debaryomyces β-D-glucosidases suitable for enhancing wine aroma have been reported. An intracellular D. hansenii β-D-glucosidase, tolerant to ethanol and glucose, efficiently released monoterpenols from the glycosides extracted from Muscat grape must. In addition, when the enzyme was added during Muscat fermentation, a considerably increase in the concentration of mainly nerol and linalool was observed (Yanai and Sato, 1999). By contrast, a Debaryomyces pseudopolymorphus strain (Cordero-Otero et al., 2003) produced an exocellular β-D-glucosidase with acidic optimal pH and not inhibited by glucose or ethanol (Arévalo-Villena et al., 2006), although the effectiveness of the purified enzyme in winemaking was not tested. Enzymatic treatment of wine with a purified ethanol tolerant β-D-glucosidase from Sporidiobolus pararoseus, a member of oenological ecosystems in the southeastern region of Brazil, considerably increased the amount of free terpenes (Baffi et al., 2011, 2013).
Several strategies for either improving β-D-glucosidase stability or enzyme yield have been described. An extracellular β-D-glucosidase from Issatchenkia terricola, active in the presence of glucose, ethanol, and metabisulfite was immobilized for improving acidic pH stability. This strategy increased the amount of monoterpenes and norisoprenoids, showing the potential of the immobilized enzyme for aroma development in wines (González-Pombo et al., 2011). With respect to yield enhancement, the usefulness of response surface methodology for optimizing the production of a Trichosporon asahii β-D-glucosidase was reported (Wang et al., 2012). T. asahii β-D-glucosidase exhibited better ability than fungal and plant commercial enzymes in hydrolyzing aromatic precursors in young wine. Also, a recombinant S. cerevisiae wine yeast expressing the Candida molischiana bgln gene encoding a β-D-glucosidase able to release terpenols and alcohols from a glycoside extract has been used to facilitate protein purification (Genovés et al., 2003).
Some of the β-D-glucosidase screenings described above also included the search of less common glycosidases, such as α-L-arabinofuranosidase, α-L-rhamnosidase and β-D-xylosidase. From more than 300 wine yeast strains, only one strain of P. anomala showed α-L-arabinofuranosidase activity whereas none of them was positive for α-L-rhamnosidase production (Spagna et al., 2002). Also the potential of certain wine yeasts from the genera Candida, Hanseniaspora, and Pichia to produce β-D-xylosidase activity active at winemaking conditions has been discussed (Manzanares et al., 1999; Yanai and Sato, 2001; Rodríguez et al., 2004; López et al., 2015). Interestingly yeast strains able to display several glycosidase activities have been reported: one strain of Aureobasidium pullulans able to hydrolyze grape glycosides displayed β-D-glucosidase, α-L-arabinofuranosidase and α-L-rhamnosidase activities, whereas Candida guilliermondii produced both β-D-glucosidase and α-L-rhamnosidase (McMahon et al., 1999). Two H. uvarum, one Hanseniaspora vineae and one P. anomala strains were described as producers of the four glycosidase activities (Mateo et al., 2011), while a Wickerhamomyces anomalus (alternative names Hansenula anomala, P. anomala and Candida pelliculosa) strain producing β-D-glucosidase, also exhibited α-L-arabinofuranosidase and β-D-xylosidase activities (Sabel et al., 2014). However, the effectiveness of purified glycosidases for terpene releasing from precursors has only been reported for the intracellular α-L-rhamnosidases from Pichia angusta (Yanai and Sato, 2000a) and Pichia guilliermondii (Rodríguez et al., 2004, 2010b), an α-L-arabinofuranosidase from Pichia capsulata (Yanai and Sato, 2000b) and a β-D-xylosidase from Candida utilis (Yanai and Sato, 2001). The latter also increased the concentration of terpenes after addition to Moscatel grape must during fermentation (Yanai and Sato, 2001).
The role of exo-glucanases in the release of aromatic compounds from glycosidically bound precursors in a single enzymatic step has been discussed (Gil et al., 2005). In this regard, a yeast isolate AS1, identified as a W. anomalus strain was selected by its capability to hydrolyze several synthetic and natural glycosides under oenological conditions (Sabel et al., 2014). Later, the enzyme responsible for the hydrolysis of selected glycosides was purified from the culture supernatant of AS1 and characterized as a multifunctional exo-β-1,3-glucanase active under typical wine related conditions (Schwentke et al., 2014).
The feasibility of using β-D-glucosidase-producing yeasts in fermentation instead of adding purified enzymes represents an interesting option. Either alone or in a mixed starter with S. cerevisiae, the ability of non-Saccharomyces yeasts to contribute to the aromatic wine profile has been assessed. Different strains of Torulaspora delbrueckii contributed to the aroma profile with flowery and fruity aroma (Maturano et al., 2012; Cordero-Bueso et al., 2013). Secreted H. vineae and T. delbrueckii β-D-glucosidases were detected throughout the fermentation process, although activity diminished with increasing fermentation time, suggesting the adverse effect of ethanol (Maturano et al., 2012). Also, isolates of Pichia membranifaciens, H. vineae, H. uvarum, and W. anomalus showing β-D-glucosidase activity provoked a moderated overall terpene increase when inoculated to final wines (López et al., 2015). The first mixed starters based on non-Saccharomyces yeasts able to produce β-D-glucosidase activity were described for Debaryomyces vanriji and D. pseudopolymorphus (García et al., 2002; Cordero-Otero et al., 2003), and later on for Candida pulcherrima (alternative name Metschnikowia pulcherrima) (Rodríguez et al., 2010a) and T. delbrueckii (Ĉuŝ and Jenko, 2013). Detailed information of these mixed starters will be found in next sections.
Some sulfur containing compounds, the so-called volatile or varietal thiols, can contribute to positive fragances such as tropical, passion fruit and guava-like nuances. These compounds considered to be impact odorants in Sauvignon Blanc wines are 4-mercapto-4-methylpentan-2-one (4MMP), reminiscent of box tree, passion fruit, broom, and black current bud; and 3-mercaptohexan-1-ol (3MH) and 3-mercaptohexyl acetate (3MHA), responsible for the passion fruit, grapefruit, and citrus aroma. Volatile thiols are not unique to Sauvignon Blanc wines. They have been also found to contribute significantly to the aroma profiles of wines made from other varieties such as Riesling, Colombard, Semillon, Cabernet Sauvignon, and Merlot (revised in Coetzee and du Toit, 2012).
Volatile thiols are mostly non-existent in grape juice and they are generated during the fermentation process by yeasts from odorless, non-volatile precursors initially present in must (revised in Cordente et al., 2012). It has been shown that 4MMP and 3MH exist in grapes in their non-volatile precursor form, conjugated to cysteine, or glutathione. The key enzyme for cleaving cysteinylated precursors is the S. cerevisiae β-lyase IRC7, with a substrate preference for cys-4MMP over cys-3MH (Roncoroni et al., 2011). The mechanism by which the glutathionated thiol precursors are degraded has not been fully elucidated, but is likely to involve a multi-step pathway with the production of the cysteinylated form as an intermediate (Grant-Preece et al., 2010). No precursor of 3MHA has been identified in grapes; this compound is formed during fermentation through esterification of 3MH by the alcohol acetyltransferase ATF1 (Swiegers et al., 2006).
Undoubtedly the main factor in volatile thiol release during alcoholic fermentation is the yeast strain (Dubourdieu et al., 2006). It was found that S. cerevisiae strains varied significantly in terms of their capabilities to produce volatile thiols and to modulate the varietal characters of Sauvignon Blanc wine (Swiegers et al., 2009). With regard to non-Saccharomyces species, only two screenings have addressed their feasibility to release volatile thiols. The first screening by Anfang et al. (2009) showed that most of the eleven non-Saccharomyces isolates tested were able to produce concentrations of 3MH above the perception threshold, but only two isolates of Pichia kluyvery and Candida zemplinina (alternative names Candida stellata and Starmerella bacillaris) produced concentrations of 3MH and 3MHA comparable with those produced by S. cerevisiae. In contrast to that found for S. cerevisiae, results showed an inverse correlation between the concentrations of 3MH and 3MHA produced by the P. kluyvery and C. zemplinina isolates, suggesting a decreased ability to convert 3MH to 3MHA, or possibly alternate metabolic routes for its formation (Anfang et al., 2009). In a second screening, the potential impact of 15 non-Saccharomyces strains from seven species on 4MMP and 3MH release in model medium and Sauvignon Blanc must was evaluated after partial fermentation (Zott et al., 2011). In general, non-Saccharomyces strains had greater ability to release 3MH than 4MMP in both media. Only M. pulcherrima and H. uvarum strains in model medium and Kluyveromyces thermotolerans in must were able to produce significant amounts of 4MMP. With respect to 3MH release, M. pulcherrima and T. delbrueckii strains released large amounts of this compound in model medium whereas M. pulcherrima and K. thermotolerans stood out as good producers in natural must. C. zemplinina isolates included in the screening did not produce volatile thiols, in contrast to previous results (Anfang et al., 2009). This can be explained by the strain dependent capacity to release 3MH as showed for M. pulcherrima (Zott et al., 2011). Undoubtedly additional screening experiments including numerous non-Saccharomyces strains are required to obtain a clear image of species-associated behavior or strain effects. Mixed fermentations with volatile thiol releasing yeasts will be discussed in later sections.
Most of the compounds that determine wine aroma arise from the fermentation process. Their concentrations are mainly dependent on the predominant yeasts and the fermentation conditions (Egli et al., 1998; Henick-Kling et al., 1998; Steger and Lambrechts, 2000). Although ethanol, glycerol, and CO2 are quantitatively the most abundant of these compounds, their contribution to the secondary aroma is relatively limited. Volatile fatty acids, higher alcohols, esters, and, to a lesser extent, aldehydes, have a greater contribution to secondary aroma (Rapp and Versini, 1991), although volatiles derived from fatty acids and from nitrogen- or sulfur-containing compounds also contribute (Boulton et al., 1996). The biosynthesis of these compounds has been reviewed in greater detail by Lambrechts and Pretorius (2000). It is worthwhile to note that the biosynthesis of these compounds is species- and strain-dependent, allowing the selection of those strains of biotechnological interest. Moreover, and depending on the concentration reached in wine, those compounds arising from yeast metabolism have a positive or negative impact on wine aroma and quality. Below we describe the contribution of non-Saccharomyces yeast species to wine secondary aroma. Table 2 shows the yeast species described as high- or low-producers of secondary aroma compounds.
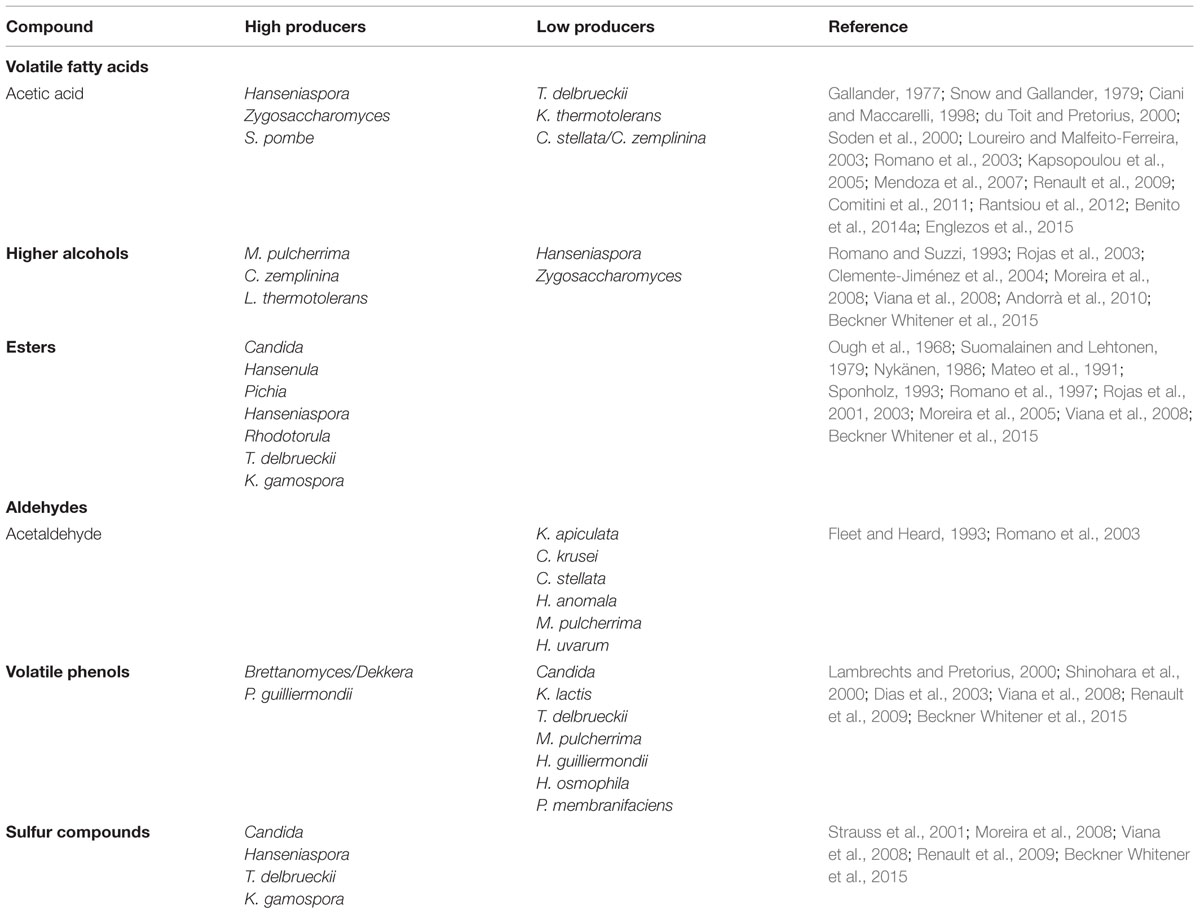
TABLE 2. Secondary aroma compounds produced by non-Saccharomyces wine yeasts.
Acetic acid is responsible for 90% of the volatile acidity of wines while the remaining fatty acids, such as propanoic and butanoic acid, are present in small quantities (Radler, 1993). Their production is also associated with bacterial growth (Ribereau-Gayon et al., 1998). Acetic acid becomes unpleasant at concentrations near its flavor threshold of 0.7–1.1 g/L and usually values between 0.2 and 0.7 g/L are considered optimal (Lambrechts and Pretorius, 2000).
Studies of acetic acid production by non-Saccharomyces yeasts have generated highly variable results. Some non-Saccharomyces genera such as Hanseniaspora and Zygosaccharomyces have been traditionally described as producers of excessive amounts of acetic acid (du Toit and Pretorius, 2000; Loureiro and Malfeito-Ferreira, 2003; Romano et al., 2003; Mendoza et al., 2007) and, for this reason, they have been considered for long time as spoilage yeasts. Also the species Schizosaccharomyces pombe is commonly associated with high levels of acetic acid (Gallander, 1977; Snow and Gallander, 1979). However, this compound is produced with a considerably strain variability. For instance, levels of acetic acid ranging from about 0.6 g/L to more than 3.4 g/L have been described for H. uvarum strains (Romano et al., 2003) while a screening of S. pombe allowed the selection of strains producing less than 0.4 g/L of acetic acid (Benito et al., 2014a).
By contrast, different screenings of T. delbrueckii strains for desirable oenological properties pointed out differences in fermentative capability but always a low production of volatile acidity when compared to S. cerevisiae (Ciani and Maccarelli, 1998; Renault et al., 2009; Comitini et al., 2011). This feature is also a characteristic of Lachancea thermotolerans (previously known as K. thermotolerans) together with the high production of L-lactic acid (Kapsopoulou et al., 2005). C. stellata/C. zemplinina presents a strong fructophilic character (Soden et al., 2000), which may be an advantage during the fermentation of sweet wines, since this species do not produce excessive levels of acetic acid as a response to the osmotic stress in comparison to S. cerevisiae (Rantsiou et al., 2012). Recently the strong fructophilic character of C. zemplinina and its ability to produce low quantities of ethanol and acetic acid and high amounts of glycerol were confirmed (Englezos et al., 2015).
They are the largest group of aromatic compounds (Amerine et al., 1980). Higher alcohols contribute to the aromatic complexity of wine at concentrations below 300 mg/L. However when their concentrations exceed 400 mg/L, they are considered to have a negative effect on aroma (Rapp and Mandery, 1986). The importance of higher alcohols is also related to their role as ester precursors (Soles et al., 1982).
In general, studies of higher alcohol production in non-Saccharomyces yeasts highlight the influence that these yeasts can have on the chemical composition and quality of wine (Herraiz et al., 1990; Mateo et al., 1991; Gil et al., 1996). In fermented musts, the total production of higher alcohols by pure cultures of Hanseniaspora species is lower than that found with S. cerevisiae (Rojas et al., 2003; Moreira et al., 2008; Viana et al., 2008). Also, Zygosaccharomyces strains isolated from grape musts have been described as producers of low amounts of higher alcohols (Romano and Suzzi, 1993). By contrast, C. zemplinina wines contained huge amounts of higher alcohols, which concentrations clearly exceeded 400 mg/L (Andorrà et al., 2010).
Regarding specific alcohols, increased production of 2-phenylethyl alcohol, compound associated with pleasant aromas, has been described as a characteristic of M. pulcherrima (Clemente-Jiménez et al., 2004), L. thermotolerans (Beckner Whitener et al., 2015), and C. zemplinina (Andorrà et al., 2010).
Esters are the most abundant compounds found in wine, with around 160 identified to date. Although various esters can be formed during fermentation, the most abundant are those derived from acetic acid (ethyl acetate, isoamyl acetate, isobutyl acetate, and 2-phenylethyl acetate) and ethyl esters of saturated fatty acids (ethyl butanoate, ethyl caproate, ethyl caprylate, and ethyl caprate). The main ester in wine is ethyl acetate, and it can impart spoilage character at levels of 150–200 mg/L (Lambrechts and Pretorius, 2000).
Non-Saccharomyces wine yeasts, known as good producers of esters, have been traditionally associated with the negative effects of high ethyl acetate formation, whereas the levels of ethyl esters produced by these yeasts are generally much lower than those detected in S. cerevisiae wines (Rojas et al., 2001, 2003).
Species belonging to the genera Candida, Hansenula, and Pichia were described as having a greater capacity to produce ethyl acetate than wine strains of S. cerevisiae (Ough et al., 1968; Nykänen, 1986). Also, in a study where ester production was grouped by yeast genera, Hanseniaspora and Pichia stood out by the production of ethyl acetate (Viana et al., 2008). Both genera produced similar ethyl acetate levels, but Hanseniaspora was also a potent producer of specific fruity acetate esters such as 2-phenylethyl acetate and isoamyl acetate (Rojas et al., 2001; Moreira et al., 2005; Viana et al., 2008), whereas the genera Pichia and Rhodotorula produced remarkable levels of isoamyl acetate (Suomalainen and Lehtonen, 1979; Viana et al., 2008). Among Hanseniaspora species, specifically H. uvarum is reported to be a good producer of esters in general (Mateo et al., 1991; Sponholz, 1993; Romano et al., 1997) whereas Hanseniaspora guilliermondii and Hanseniaspora osmophila are strong producers of 2-phenylethyl acetate (Rojas et al., 2001, 2003; Viana et al., 2008).
Regarding ethyl esters, production of ethyl caprylate seems to be a characteristic of T. delbrueckii (Viana et al., 2008). The aroma profile of the newly discovered yeast Kazachstania gamospora showed that this species produced more esters than the S. cerevisiae control strain, but specially phenylethyl propionate, an ester desirable in wine due to its floral aroma (Beckner Whitener et al., 2015).
These compounds with apple-like odors are important to the aroma and bouquet of wine due to their low sensory threshold values. Among aldehydes, acetaldehyde constitutes more than 90% of the total content of wines, and its amount can vary from 10 mg/L up to 300 mg/L (Lambrechts and Pretorius, 2000).
Saccharomyces cerevisiae strains usually produce higher acetaldehyde levels (5–120 mg/L) than non-Saccharomyces species (up to 40 mg/L) such as Kloeckera apiculata, Candida krusei, C. stellata, H. anomala, and M. pulcherrima (Fleet and Heard, 1993). A mean acetaldehyde concentration of around 25 mg/L was described for H. uvarum strains, although significant differences in production among strains were observed (Romano et al., 2003).
Among volatile phenols, the most important are vinylphenols in white wines and ethylphenols in red wines. Their presence is always undesirable, since even at concentrations below the perception threshold they are reported to mask the fruity notes of white wines. These compounds are produced from the non-volatile ferulic and p-coumaric acids. Traditionally, ethylphenol producers have been ascribed to the genus Brettanomyces/Dekkera (Lambrechts and Pretorius, 2000). However, several studies also identified Candida species, Kluyveromyces lactis, T. delbrueckii, M. pulcherrima, and P. guilliermondii strains as volatile phenol producers, although only P. guilliermondii displayed the same conversion capacity as Dekkera species (Shinohara et al., 2000; Dias et al., 2003; Renault et al., 2009; Beckner Whitener et al., 2015). By contrast H. guilliermondii, H. osmophila, and P. membranifaciens were not able to decarboxylate either ferulic or p-coumaric acids (Viana et al., 2008).
The sensory properties of sulfur compounds vary extensively and although most of them are associated with negative aromatic descriptors, they can have a positive contribution to wine aroma through the introduction of fruity notes (reviewed in Swiegers and Pretorius, 2005). The main compound in this group is hydrogen sulfide. Production by non-Saccharomyces yeasts includes Candida and Hanseniaspora species (Strauss et al., 2001; Viana et al., 2008) as well as T. delbrueckii (Renault et al., 2009). The contribution of H. uvarum and H. guilliermondii to the sulfur compound profile of wines was evaluated by Moreira et al. (2008) and it was concluded that the growth of the apiculate yeasts might not have a negative influence. It should also be noted that T. delbrueckii and K. gamospora are able to produce the sulfur compound 3-methylthio-1-propanol in higher concentration than S. cerevisiae, although it was dependent on the must variety (Beckner Whitener et al., 2015). The formation of volatile thiols by non-Saccharomyces yeasts has been described in the primary aroma section.
The use of mixed starters of selected non-Saccharomyces yeasts to exploit their positive abilities combined with S. cerevisiae to avoid stuck fermentations represents a feasible alternative to both spontaneous and inoculated fermentations (Figure 1). These combinations could be used to produce wines with unique aromatic characteristics. Based on their capability to produce flavor enhancing enzymes or to modify the concentration of secondary metabolites, different mixed starters have been designed and proposed as a tool to enhance wine quality (Table 3). Some of them were designed with the aim of modifying a specific target such as the terpenic profile or final ester concentrations while others show a general impact on wine aroma complexity. The increasing interest in the use of non-Saccharomyces yeasts in winemaking has even prompted commercial production of several species including L. thermotolerans, M. pulcherrima, T. delbrueckii, P. kluyvery, and S. pombe.
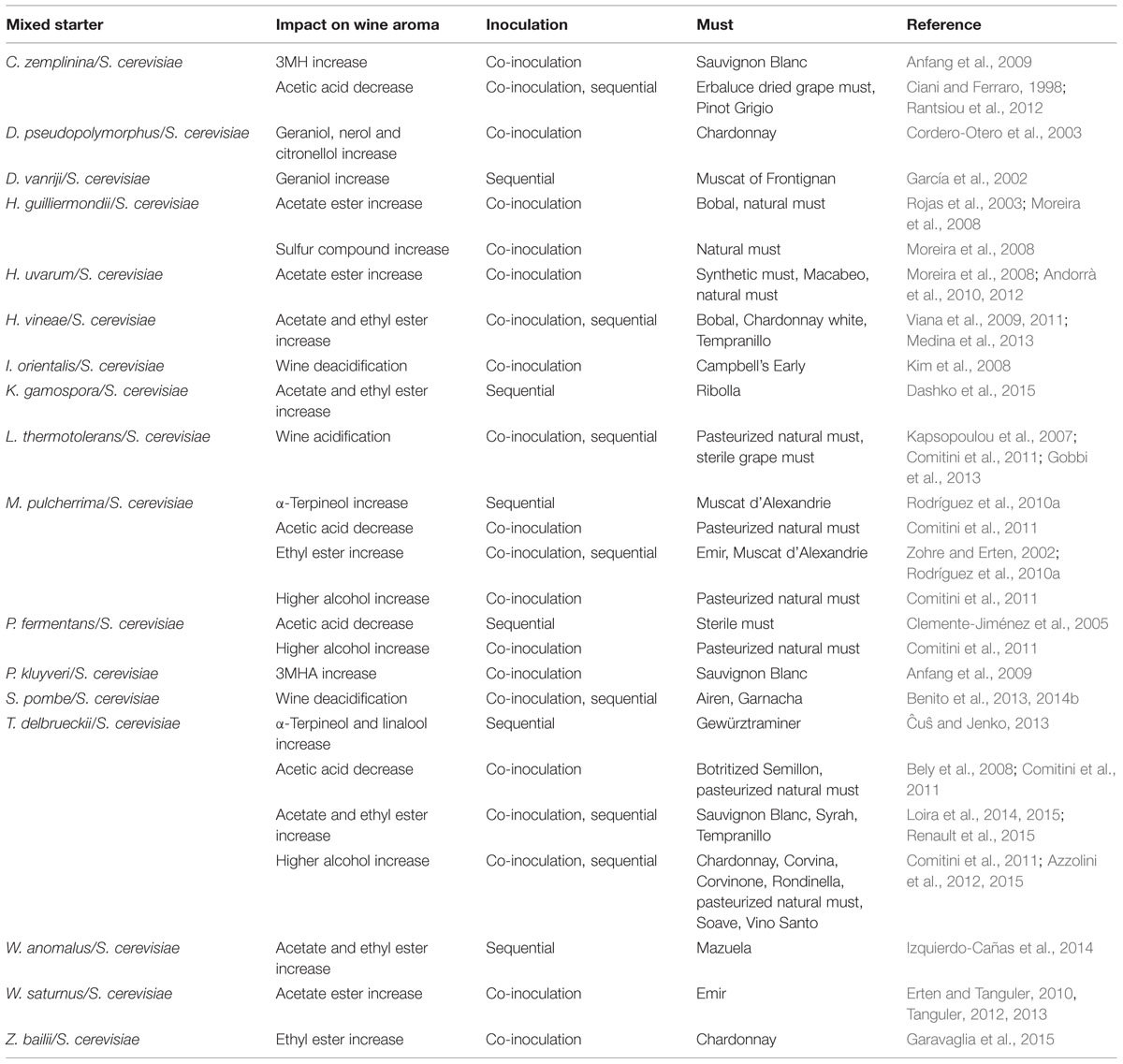
TABLE 3. Mixed starters designed to improve primary and secondary wine aroma.
When using non-Saccharomyces yeasts in mixed starters, there are two general practices of inoculation. The first, known as co-inoculation, involves the inoculation of the selected non-Saccharomyces yeasts at high cell concentration together with S. cerevisiae, while the second, sequential inoculation, implies that the selected non-Saccharomyces yeasts are first inoculated at high levels and allowed to ferment on their own for a given amount of time before S. cerevisiae is added to take over the fermentation. Both are feasible practices although potential interactions between yeasts could determine which inoculation strategy is more appropriate.
With the aim of obtaining terpene-enriched wines, T. delbrueckii, M. pulcherrima, D. hansenii, and D. pseudopolymorphus strains able to produce β-D-glucosidase activity were combined with S. cerevisiae.
Higher concentrations of α-terpineol and linalool were found in Gewürztraminer wine fermented with the combination T. delbrueckii/S. cerevisiae, although more nerol and geraniol were detected in the control fermentation conducted with S. cerevisiae alone (Ĉuŝ and Jenko, 2013). Moreover those chemical changes enhanced the overall quality of Gewürztraminer wine.
Metschnikowia pulcherrima is known to produce β-D-glucosidase activity able to increase the α-terpineol, nerol as well as geraniol concentrations in monoculture wines (Rodríguez et al., 2010a). However, in wines obtained by mixed fermentation, in either simultaneous or sequential inoculation, nerol and geraniol concentrations were significantly lower than those observed in grape must, and only α-terpineol concentration was higher. This fact was related to the S. cerevisiae capability to transform nerol and geraniol into α-terpineol at must pH (Di Stefano et al., 1992; Mateo and Jiménez, 2000), pointing out the relevance of yeast interactions.
With respect to Debaryomyces species, a D. vanriji strain isolated from grape berry flora was found to influence wine volatiles of the cv. Muscat of Frontignan when co-cultured with native or selected strains of S. cerevisiae. The concentrations of several volatiles including terpenols were significantly different between the control and the wines inoculated with D. vanriji. The increase in geraniol concentration was attributed to the hydrolysis of the corresponding glucosidic precursor by D. vanriji β-D-glucosidase since musts inoculated with the non-Saccharomyces yeast showed higher levels of enzymatic activity throughout the fermentation compared to the control sample. Moreover bound geraniol concentration was found to be lower in D. vanriji inoculated wines compared to the control one (García et al., 2002). The high terpene concentrations of wines obtained with mixed cultures D. vanriji/S. cerevisiae was recently confirmed and associated with the production of pectinase, amylase, and xylanase activities along the fermentation (Maturano et al., 2015). Also, a β-D-glucosidase producing D. pseudopolymorphus strain when co-cultured with S. cerevisiae VIN13 significantly increased concentrations of citronellol, nerol, and geraniol during the fermentation of Chardonnay juice (Cordero-Otero et al., 2003).
To take advantage of the capability of C. zemplinina to produce volatile thiols, mixed starters of commercially available S. cerevisiae strains with C. zemplinina isolates were employed in Sauvignon Blanc fermentations. Inoculation with equal amounts or with a ratio that initially favored the non-Saccharomyces isolates, produced wines with the greatest increase of the volatile thiol 3MH when compared with the S. cerevisiae single ferment. However, the co-ferments with the C. zemplinina isolates had significantly lower concentrations of 3MHA (Anfang et al., 2009). By contrast, an elevation in thiol production, especially 3MHA, was found in Sauvignon Blanc wines co-fermented with P. kluyvery isolates and different S. cerevisiae strains (Anfang et al., 2009). Although both species were able to produce 3MHA in monoculture, the increase in 3MHA concentration observed in co-fermented wines could not be explained under simple additive assumptions suggesting an interaction between the co-fermenting partners. Moreover the elevation in thiols was only seen in co-ferments with certain S. cerevisiae strains suggesting that the nature of this particular interaction may not be generalized to the species level. However, the mechanism behind this interaction is still unknown.
Nowadays, selected non-Saccharomyces strains for the improvement of wine primary aroma are on the market. A M. pulcherrima strain selected for its specific property to release enzymes with α-L-arabinofuranosidase activity is now available. M. pulcherrima, with a suitably paired S. cerevisiae strain sequentially inoculated, impacts on the expression of terpenes and thiols and it is recommended for Riesling and Sauvignon Blanc wines. Also a commercial product based on a selected strain of P. kluyveri is recommended due to its ability to boost fruit flavors through a more efficient conversion of flavor precursors into volatile thiols.
Different strategies based on non-Saccharomyces yeasts have been described for reducing volatile acidity or either for acidifying or deacidifying wines.
To solve the problem of excessive volatile acidity due to high acetic acid concentrations, non-Saccharomyces yeasts such as T. delbrueckii, M. pulcherrima, and C. stellata/C. zemplinina can be employed. T. delbrueckii, often described as a low acetic acid producer under standard conditions, retains this quality even fermenting high-sugar media. A mixed culture of T. delbrueckii and S. cerevisiae was shown to be the best combination for improving the analytical profile of wines produced from botrytized musts, particularly volatile acidity, and acetaldehyde production. Specifically, the mixed T. delbrueckii/S. cerevisiae culture produced 53% less in volatile acidity and 60% less acetaldehyde than a pure culture of S. cerevisiae (Bely et al., 2008). Interestingly, the mixed culture was only effective in simultaneous inoculation since the sequential one resulted in stuck fermentation. Significant reductions in volatile acidity in mixed fermentations of T. delbrueckii/S. cerevisiae were also reported by other authors (Comitini et al., 2011), who observed the same effect when using M. pulcherrima/S. cerevisiae starters independently of the inoculum ratios.
The high fermentative capacity of C. stellata/C. zemplinina has been explored in mixed starters. In a fermentation conducted by a mixture of C. stellata and S. cerevisiae in Pinot Grigio must (270 g sugars/L), yeast cells were able to completely consume all glucose and fructose while reducing the levels of acetic acid (Ciani and Ferraro, 1998). Similarly, specific strains of C. zemplinina when co-inoculated with S. cerevisiae were able to reduce the content of acetic acid while maintaining high glycerol and ethanol levels (Rantsiou et al., 2012).
In grape must, different combinations of Pichia fermentans with S. cerevisiae produced less acetic acid than S. cerevisiae in single cultures. Moreover the decrease in acetic acid was accompanied by a substantial increase in aromatic compounds such as acetaldehyde, ethyl acetate, 1-propanol, n-butanol, 1-hexanol, ethyl caprilate, 2,3-butanediol, and glycerol (Clemente-Jiménez et al., 2005).
The ability of non-Saccharomyces yeasts to act as acidifying agents is of increasing interest, as global climate change and variations in viticulture and oenology practices have resulted in a trend toward the reduction of the total acidity of wines. L. thermotolerans, through the production of L-lactic acid, is a potential acidifying microorganism during must fermentation which could compensate the insufficient acidity of specific grape varieties (Mora et al., 1990; Kapsopoulou et al., 2007). L. thermotolerans, in both simultaneous and sequential inoculations with S. cerevisiae, provided an effective acidification during alcoholic fermentation although the production of L-lactic acid was dependent on the time of inoculation of the S. cerevisiae strain (Kapsopoulou et al., 2007). Also the L. thermotolerans/S. cerevisiae consortium provoked a pH reduction associated with a significant enhancement in the total acidity and reduction in the volatile acidity, as compared to pure S. cerevisiae cultures (Comitini et al., 2011). The mixed fermentations were characterized as well by increases in glycerol and main esters. In agreement with previous results, pH reductions and enhancement of glycerol and 2-phenylethyl alcohol contents were shown in wines fermented with the co-culture. Moreover sensory analysis tests showed significant increases in the spicy notes and in terms of total acidity increases (Gobbi et al., 2013).
Also the deacidifying capacity of S. pombe and Issatchenkia orientalis (alternative name Pichia kudriavzevii) due to the consumption of malic acid has been explored in mixed starters. The combination S. pombe/S. cerevisiae has proved successful in biological deacidification of white and red wines (Benito et al., 2013, 2014b). In all wines obtained with S. pombe either alone or together with S. cerevisiae, nearly all the malic acid was consumed, and moderate acetic acid concentrations were formed. Moreover the urea content of these wines was notably lower when compared with those that were made with S. cerevisiae alone. White wines obtained with mixed cultures received the best overall scores after sensory evaluation (Benito et al., 2013) but in red wine fermentations, the maximum aroma intensity and quality corresponded to those obtained with S. pombe in monoculture (Benito et al., 2014b). Similarly, wines co-fermented by I. orientalis and S. cerevisiae showed decreased malic acid concentrations and the highest score in sensory evaluation. The co-fermentation also decreased the contents of acetaldehyde, 1-propanol, 2-butanol, and isoamyl alcohol but increased the methanol content (Kim et al., 2008). Recently the capability of P. kudriavzevii to degrade malic acid in microvinifications, increasing the pH 0.2–0.3 units, was confirmed (del Mónaco et al., 2014).
Additionally, the combined use of selected S. pombe and L. thermotolerans strains has been described as a feasible alternative to the traditional malolactic fermentation (Benito et al., 2015). With this approach, malic acid is totally consumed by S. pombe, while lactic acid produced by L. thermotolerans maintains or increases the acidity of wines produced from low acidity musts. Final wines had more fruity character and they contained less acetic acid and biogenic amines than the traditional malolactic fermentation controls (Benito et al., 2015).
The increase of fruity acetate esters has been the main target of mixed starters designed with Hanseniaspora species. H. guilliermondii and H. uvarum grown as mixed cultures with S. cerevisiae in grape must increased the 2-phenylethyl acetate and isoamyl acetate content of wines, respectively (Rojas et al., 2003; Moreira et al., 2008). However, the excessive production of ethyl acetate limited the applicability of both mixed starters. Similarly, although a desirable increase of acetate esters in mixed fermentations of Macabeo and synthetic must with H. uvarum/S. cerevisiae was reported, the high concentration of acetic acid hampered the industrial application of the mixed starter (Andorrà et al., 2010, 2012). However, since a large strain variability is associated to metabolite production (Plata et al., 2003; Romano et al., 2003; Ciani et al., 2006), the excessive concentration of both ethyl acetate and acetic acid can be avoided by means of specific screenings. In this regard a H. vineae strain yielding high levels of 2-phenylethyl acetate while producing levels of acetic acid and ethyl acetate within the optimal ranges described for wine was selected (Viana et al., 2008). Moreover the potential of using that selected strain in a mixed starter with S. cerevisiae to increase the levels of 2-phenylethyl acetate in wines without compromising quality was demonstrated (Viana et al., 2009). Besides, the ratio of both yeast strains in the mixed culture modulated ester concentrations leading to wines with a wide range of flavor compounds. Further studies showed that the selected H. vineae strain inoculated as a part of a sequential mixed starter was able to compete with native yeasts present in a non-sterile must and modify the wine aroma profile, specifically 2-phenylethyl acetate concentration (Viana et al., 2011). Recently, wines obtained from industrial Chardonnay white grape vinifications conducted by sequential H. vineae/S. cerevisiae inoculation showed a significant increase in fruity intensity described as banana, pear, apple, citric fruits, and guava, in comparison to spontaneous and pure S. cerevisiae fermentations. Fruity intensity was mainly correlated to higher concentrations of acetyl and ethyl esters and relative decreases in alcohols and fatty acids (Medina et al., 2013).
An increase in isoamyl acetate content can also be achieved by fermenting Emir must with Williopsis saturnus/S. cerevisiae cultures. Furthermore the mixed culture did not produce any off-flavors, although the changes observed in the aromatic profile of mixed wines were inoculum and temperature dependent (Erten and Tanguler, 2010; Tanguler, 2012, 2013). Wines elaborated by sequential fermentation of W. anomalus and S. cerevisiae presented higher levels of acetate and ethyl esters and of linear alcohols, which contribute to increase the aromatic quality with floral and fruity notes (Izquierdo-Cañas et al., 2014). With the aim of obtaining white wines with an enhanced aromatic complexity, a Zygosaccharomyces bailii strain characterized as producer of several esters (Garavaglia et al., 2014) was inoculated together with S. cerevisiae. In all trials that contained the non-Saccharomyces yeast the production of ethyl esters was increased in comparison to the vinification control (Garavaglia et al., 2015).
In mixed cultures T. delbrueckii/S. cerevisiae the formation of specific esters was reported. For instance, the mixed culture produced Tempranillo wines with larger quantities of 2-phenylethyl acetate and ethyl lactate than single S. cerevisiae fermentations (Loira et al., 2014), or larger amounts of isoamyl acetate, hexyl acetate, ethyl hexanoate, and ethyl octanoate in Syrah wines (Loira et al., 2015). Also, ethyl propanoate, ethyl isobutanoate, ethyl dihydrocinnamate, isobutyl acetate, and isoamyl acetate concentrations were increased in wines obtained by mixed fermentations T. delbrueckii/S. cerevisiae, either in sequential or simultaneous inoculation (Renault et al., 2015). Favoring T. delbrueckii development when performing sequential inoculation enhanced the concentration of the above mentioned ethyl esters, which were linked to T. delbrueckii activity. On the contrary, simultaneous inoculation restricted the growth of T. delbrueckii, limiting the production of its activity markers. However, simultaneous inoculation involved a high production of numerous esters due to more important positive interactions between yeast species. These results suggested that the ester concentration enhancement via interactions during mixed modalities was due to S. cerevisiae production in response to the presence of T. delbrueckii (Renault et al., 2015). A pure culture of T. delbrueckii selected for its properties to enhance wine aromatic and mouthfeel complexity is available on the market. When used in sequential inoculation with compatible selected S. cerevisiae, it favors the perception of certain esters without overwhelming the wines.
Finally, aromatic complexity of Ribolla wines can be improved by sequential fermentation with K. gamospora and S. cerevisiae due to the enhanced production of esters such as 2-phenylethyl acetate and ethyl propionate and also of 2-phenylethyl alcohol (Dashko et al., 2015).
The presence of T. delbrueckii in mixed starters has been associated with increases in the production of 2-phenylethyl alcohol in different kinds of wine. Mixed starters T. delbrueckii/S. cerevisiae were proposed for the production of Amarone wine, a high-alcohol dry red wine obtained from withered grapes. The most significant changes caused by the presence of T. delbrueckii were observed among alcohols, specifically benzyl alcohol and 2-phenylethyl alcohol, but also in fermentative esters, fatty acids and lactones, which are important in the Amarone wine flavor (Azzolini et al., 2012). Interestingly the increase in the levels of 2-phenylethyl alcohol seemed to be related to the β-glucosidase activity of the T. delbrueckii strain employed, although other factors could not be discarded. Also in the fermentation of dry and sweet wines, mixed cultures T. delbrueckii/S. cerevisiae affected the content of several important volatile compounds, including 2-phenylethyl alcohol, isoamyl acetate, fatty acid esters, C4–C10 fatty acids and vinylphenols (Azzolini et al., 2015). In addition to T. delbrueckii mixed starters, M. pulcherrima/S. cerevisiae, L. thermotolerans/S. cerevisiae, and K. gamospora/S. cerevisiae fermentations resulted in higher productions of 2-phenylethyl alcohol (Comitini et al., 2011; Dashko et al., 2015).
Finally, and with the aim of mimicking the complex yeast microbiota present in fermenting musts, wine mixed cultures composed by more than one non-Saccharomyces species in combination with S. cerevisiae have been also developed. However, the number of studies focused on wine aroma development is still low and results somehow controversial.
In this regard, fermentations of natural grape musts with a Saccharomyces strain together with a C. zemplinina and/or a H. uvarum strains showed the preferential use of some groups of amino acids (aliphatic, aromatic, and sulphur amino acids) in the mixed fermentations compared with the pure cultures. These results suggested that the presence of several yeast species might improve the uptake or consumption of some amino acids by some kind of synergistic mechanism (Andorrà et al., 2010). However the preferential use of amino acids did not have a clear consequence on aroma production as it would be expected: fermentations with the triple mixed culture only stood out for ethyl lactate production and significant differences were only observed with the pure S. cerevisiae fermentation, while fermentations including one or two non-Saccharomyces strains were comparable to each other. Moreover the amount of acetic acid was well above the admissible levels and thus compromising the immediate application of these mixed cultures (Andorrà et al., 2010). Later on, the same authors reported significant differences in the above results when a synthetic grape must was used (Andorrà et al., 2012).
Production of white wine by sequential inoculation of H. anomala, T. delbrueckii, and S. cerevisiae has been proposed (Izquierdo-Cañas et al., 2011) Resulting wines were chemically different to those produced by S. cerevisiae alone, by H. anomala/S. cerevisiae, or by T. delbrueckii/S. cerevisiae. Surprisingly, the two last combinations produced wines exhibiting more complexity than the one including the two non-Saccharomyces species, which in fact was comparable to the pure S. cerevisiae fermentation.
Nowadays, a blend of three yeasts, S. cerevisiae, K. thermotolerans, and T. delbrueckii, is commercialized. The mixture gives tropical fruitiness and an overall aromatic intensity in white wines and more pronounced fruity and spicy notes in red wines.
Based on numerous studies showing the positive influence of non-Saccharomyces yeasts in winemaking, the wine industry has been directed toward the use of controlled mixed fermentations. Indeed designed mixed starters with selected non-Saccharomyces strains and S. cerevisiae can enhance, as pointed out in this review, primary and secondary wine aroma, but also they are involved in reductions of the ethanol content of wine (González et al., 2013; Contreras et al., 2015; Morales et al., 2015), control of the spoilage wine microflora (Oro et al., 2014), release of mannoproteins (Domizio et al., 2014) or wine color stabilization (Morata et al., 2012; Loira et al., 2015). Moreover they can exert a positive effect in base wines for sparkling wine production improving foaming properties (González-Royo et al., 2015). Remarkably, a new red winemaking technology based on the combined use of two non-Saccharomyces yeast strains has been developed as an alternative to the traditional malolactic fermentation (Benito et al., 2015).
In addition to the mandatory strain selection, the benefits of mixed cultures should be tested in different grape musts since different nutritional characteristics and limitations might modify the impact of the individual components of the starter on the final wine. Moreover mixed cultures should be tested at industrial or semi-industrial scales because it has been reported that the production of different metabolites can vary depending on the fermentation volume and the oxygen conditions (Beltran et al., 2008; Viana et al., 2009). Certainly the study of the impact of common oenological practices on the dynamics of non-Saccharomyces yeasts will be also useful for a better management of mixed fermentations (Albertin et al., 2014).
Considering that the main reason for re-evaluating non-Saccharomyces yeasts and for introducing mixed cultures in the winemaking process was to get differentiated wines reflecting the characteristic of a given wine region, the commercial assortment of non-Saccharomyces cultures is still reduced. In this context the continuous ecological studies as well as the oenological and sensory characterization of autochthonous non-Saccharomyces and even S. cerevisiae isolates will provide appropriate candidates to be included as a part of commercial mixed starter cultures for the production of typical wines (Canonico et al., 2015; Teixeira et al., 2015).
Finally, rational design of mixed cultures should take into account not just results from smart screenings that allow exploiting positive features of non-Saccharomyces yeasts but also potential interactions among microorganisms. Some yeast interactions reported to occur in mixed starters have been briefly discussed in this review, but little is known about the mechanisms involved. In fact, positive, negative, and neutral interactions in mixed fermentations of non-Saccharomyces and Saccharomyces yeasts for the formation of aromatic compounds have been identified (Sadoudi et al., 2012). These interactions seem to be strain-dependent for both non-Saccharomyces and S. cerevisiae strains (Anfang et al., 2009; Canonico et al., 2015) and might affect the entire metabolic pathway. Current knowledge on wine yeast interactions has been recently revised (Ciani and Comitini, 2015) but it is an area that requires in-depth studies. Undoubtedly the application of high throughput techniques will offer a powerful approach for unraveling microbial interactions and thus it will allow a better design of mixed cultures and also an increased control over mixed culture fermentations.
BP, JG, and PM made contributions to conception and design of the review. BP and JG draft the manuscript. PM supervised and edited the manuscript. All authors commented on the manuscript at all stages.
Our work on non-Saccharomyces yeasts was supported by grants VIN03-007-C2-1, AGL2003-01295, AGL2004-00978, Consolider-Ingenio 2010 CSD2007-0063, AGL2010-21009, and AGL2014-58205-REDC from the Spanish Government-FEDER.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
We extend special thanks to our collaborators and laboratory members for help and thoughtful discussions. We apologize to all the investigators whose research could not be appropriately cited because of space limitations.
Albertin, W., Miot-Sertier, C., Bely, M., Marullo, P., Coulon, J., Moine, V., et al. (2014). Oenological prefermentation practices strongly impact yeast population dynamics and alcoholic fermentation kinetics in Chardonnay grape must. Int. J. Food Microbiol. 178, 87–97. doi: 10.1016/j.ijfoodmicro.2014.03.009
Amerine, A. M., Berg, H. V., Kunkee, R. E., Ough, C. S., Singleton, V. L., and Webb, A. D. (1980). The Technology of Winemaking. Wesport, CT: AVI Technical Books Inc.
Andorrà, I., Berradre, M., Mas, A., Esteve-Zarzoso, B., and Guillamón, J. M. (2012). Effect of mixed culture fermentations on yeast populations and aroma profile. LWT Food Sci. Technol. 49, 8–13. doi: 10.1016/j.lwt.2012.04.008
Andorrà, I., Berradre, M., Rozès, N., Mas, A., Guillamón, J. M., and Esteve-Zarzoso, B. (2010). Effect of pure and mixed cultures of the main wine yeast species on grape must fermentations. Eur. Food Res. Technol. 231, 215–224. doi: 10.1007/s00217-010-1272-0
Anfang, N., Brajkovich, M., and Goddard, M. R. (2009). Co-fermentation with Pichia kluyveri increases varietal thiol concentrations in Sauvignon Blanc. Aust. J. Grape Wine Res. 15, 1–8. doi: 10.1111/j.1755-0238.2008.00031.x
Arévalo-Villena, M., Úbeda-Iranzo, J. F., and Briones-Pérez, A. I. (2007). β-Glucosidase activity in wine yeasts: application in enology. Enzyme Microb. Technol. 40, 420–425. doi: 10.1016/j.enzmictec.2006.07.013
Arévalo-Villena, M., Úbeda-Iranzo, J. F., Gundllapalli, S. B., Cordero-Otero, R. R., and Briones-Pérez, A. I. (2006). Characterization of an exocellular β-glucosidase from Debaryomyces pseudopolymorphus. Enzyme Microb. Technol. 39, 229–234. doi: 10.1016/j.enzmictec.2005.10.018
Azzolini, M., Fedrizzi, B., Tosi, E., Finato, F., Vagnoli, P., Scrinzi, C., et al. (2012). Effects of Torulaspora delbrueckii and Saccharomyces cerevisiae mixed cultures on fermentation and aroma of Amarone wine. Eur. Food Res. Technol. 235, 303–313. doi: 10.1007/s00217-012-1762-3
Azzolini, M., Tosi, E., Lorenzini, M., Finato, F., and Zapparoli, G. (2015). Contribution to the aroma of white wines by controlled Torulaspora delbrueckii cultures in association with Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 31, 277–293. doi: 10.1007/s11274-014-1774-1
Baffi, M. A., Dos Santos Bezerra, C., Arévalo-Villena, M., Briones-Pérez, A. I., Gomes, E., and Da Silva, R. (2011). Isolation and molecular identification of wine yeasts from a Brazilian vineyard. Ann. Microbiol. 61, 75–78. doi: 10.1007/s13213-010-0099-z
Baffi, M. A., Martin, N., Tobal, T. M., Ferrarezi, A. L., Ghilardi Lago, J. H., Boscolo, M., et al. (2013). Purification and characterization of an ethanol-tolerant β-glucosidase from Sporidiobolus pararoseus and its potential for hydrolysis of wine aroma precursors. Appl. Biochem. Biotechnol. 171, 1681–1691. doi: 10.1007/s12010-013-0471-0
Barnett, J. A. (2000). A history of research on yeasts 2: Louis Pasteur and his contemporaries, 1850 – 1880. Yeast 16, 755–771. doi: 10.1002/1097-0061(20000615)16:8<755::AID-YEA587>3.0.CO;2-4
Barnett, J. A., and Lichtenthaler, F. W. (2001). A history of research on yeasts 3: emil Fischer Eduard Buchner and their contemporaries, 1880–1900. Yeast 18, 363–388. doi: 10.1002/1097-0061(20010315)18:4<363::AID-YEA677>3.0.CO;2-R
Beckner Whitener, M. E., Carlin, S., Jacobson, D., Weighill, D., Divol, B., Conterno, L., et al. (2015). Early fermentation volatile metabolite profile of non-Saccharomyces yeasts in red and white grape must: a targeted approach. LWT Food Sci. Technol. 64, 412–422. doi: 10.1016/j.lwt.2015.05.018
Beltran, G., Novo, M., Guillamón, J. M., Mas, A., and Rozès, N. (2008). Effect of fermentation temperature and culture media on the yeast lipid composition and wine volatile vompounds. Int. J. Food Microbiol. 121, 169–177. doi: 10.1016/j.ijfoodmicro.2007.11.030
Beltran, G., Torija, M. J., Novo, M., Ferrer, N., Poblet, M., Guillamón, J. M., et al. (2002). Analysis of yeast populations during alcoholic fermentation: a six year follow-up study. Syst. Appl. Microbiol. 25, 287–293. doi: 10.1078/0723-2020-00097
Bely, M., Sablayrolles, J. M., and Barre, P. (1990). Description of alcoholic fermentation kinetics: its variability and significance. Am. J. Enol. Vitic. 41, 319–324.
Bely, M., Stoeckle, P., Masneuf-Pomarède, I., and Dubourdieu, D. (2008). Impact of mixed Torulaspora delbrueckii-Saccharomyces cerevisiae culture on high-sugar fermentation. Int. J. Food Microbiol. 122, 312–320. doi: 10.1016/j.ijfoodmicro.2007.12.023
Benda, I. (1982). “Wine and Brandy,” in Prescott and Dunn’s Industrial Microbiology, ed. G. Reed (Westport, CT: AVI Technical Books Inc), 293–402.
Benito, A., Calderón, F., Palomero, F. E., and Benito, S. (2015). Combine use of selected Schizosaccharomyces pombe and Lachancea thermotolerans yeast strains as an alternative to the traditional malolactic fermentation in red wine production. Molecules 20, 9510–9523. doi: 10.3390/molecules20069510
Benito, S., Palomero, F., Calderón, F., Palmero, D., and Suárez-Lepe, J. A. (2014a). Selection of appropriate Schizosaccharomyces strains for winemaking. Food Microbiol. 42, 218–224. doi: 10.1016/j.fm.2014.03.014
Benito, S., Palomero, F., Gálvez, L., Morata, A., Calderón, F., Palmero, D., et al. (2014b). Quality and composition of red wine fermented with Schizosaccharomyces pombe as sole fermentative yeast, and in mixed and sequential fermentations with Saccharomyces cerevisiae. Food Technol. Biotechnol. 52, 376–382.
Benito, S., Palomero, F., Morata, A., Calderón, F., Palmero, D., and Suárez-Lepe, J. A. (2013). Physiological features of Schizosaccharomyces pombe of interest in making of white wines. Eur. Food Res. Technol. 236, 29–36. doi: 10.1007/s00217-012-1836-2
Bindon, K. A., Dry, P. R., and Loveys, B. R. (2007). Influence of plant water status on the production of C13-norisoprenoid precursors in Vitis vinifera L. Cv. Cabernet Sauvignon grape berries. J. Agric. Food Chem. 55, 4493–4500. doi: 10.1021/jf063331p
Blanco, P., Mirás-Avalos, J. M., and Orriols, I. (2012). Effect of must characteristics on the diversity of Saccharomyces strains and their prevalence in spontaneous fermentations. J. Appl. Microbiol. 112, 936–944. doi: 10.1111/j.1365-2672.2012.05278.x
Bouchilloux, P., Darriet, P., Henry, R., Lavigne-Cruege, V., and Dubourdieu, D. (1998). Identification of volatile and powerful odorous thiols in Bordeaux red wine varieties. J. Agric. Food Chem. 46, 3095–3099. doi: 10.1021/jf971027d
Boulton, R. B., Singleton, V. L., Bisson, L. F., and Kunkee, R. E. (1996). Principles and Practices of Winemaking. New York, NY: Chapman.
Brilli, L., Buscioni, G., Moriondo, M., Bindi, M., and Vicenzini, M. (2015). Influence of interannual meteorological variability on yeast content and composition in Sangiovese grapes. Am. J. Enol. Vitic. 65, 375–380. doi: 10.5344/ajev.2014.13116
Canonico, L., Comitini, F., and Ciani, M. (2015). Influence of vintage and selected starter on Torulaspora delbrueckii/Saccharomyces cerevisiae sequential fermentation. Eur. Food Res. Technol. 241, 827–833. doi: 10.1007/s00217-015-2507-x
Carrau, F. M., Medina, K., Boido, E., Farina, L., Gaggero, C., Dellacassa, E., et al. (2005). De novo synthesis of monoterpenes by Saccharomyces cerevisiae wine yeasts. FEMS Microbiol. Lett. 243, 107–115. doi: 10.1016/j.femsle.2004.11.050
Charoenchai, C., Fleet, G. H., Henschke, P. A., and Todd, B. E. N. (1997). Screening of non-Saccharomyces wine yeasts for the presence of extracellular hydrolytic enzymes. Aust. J. Grape Wine Res. 3, 2–8. doi: 10.1111/j.1755-0238.1997.tb00109.x
Ciani, M., Beco, L., and Comitini, F. (2006). Fermentation behaviour and metabolic interactions of multistarter wine yeast fermentations. Int. J. Food Microbiol. 108, 239–245. doi: 10.1016/j.ijfoodmicro.2005.11.012
Ciani, M., and Comitini, F. (2015). Yeast interactions in multi-starter wine fermentation. Curr. Opin. Food Sci. 1, 1–6. doi: 10.1016/j.cofs.2014.07.001
Ciani, M., and Ferraro, L. (1998). Combined use of immobilized Candida stellata cells and Saccharomyces cerevisiae to improve the quality of wines. J. Appl. Microbiol. 85, 247–254. doi: 10.1046/j.1365-2672.1998.00485.x
Ciani, M., and Maccarelli, F. (1998). Oenological properties of non-Saccharomyces yeasts associated with wine-making. World J. Microbiol. Biotechnol. 14, 199–203. doi: 10.1023/A:1008825928354
Clemente-Jiménez, J. M., Mingorance-Cazorla, L., Martínez-Rodríguez, S., Las Heras-Vázquez, F. J., and Rodríguez-Vico, F. (2004). Molecular characterization and oenological properties of wine yeasts isolated during spontaneous fermentation of six varieties of grape must. Food Microbiol. 21, 149–155. doi: 10.1016/S0740-0020(03)00063-7
Clemente-Jiménez, J. M., Mingorance-Cazorla, L., Martínez-Rodríguez, S., Las Heras-Vázquez, F. J., and Rodríguez-Vico, F. (2005). Influence of sequential yeast mixtures on wine fermentation. Int. J. Food Microbiol. 98, 301–308. doi: 10.1016/j.ijfoodmicro.2004.06.007
Coetzee, C., and du Toit, W. J. (2012). A comprehensive review on Sauvignon Blanc aroma with a focus on certain positive volatile thiols. Food Res. Int. 45, 287–298. doi: 10.1016/j.foodres.2011.09.017
Combina, M., Elía, A., Mercado, L., Catania, C., Ganga, A., and Martinez, C. (2005). Dynamics of indigenous yeast populations during spontaneous fermentation of wines from Mendoza, Argentina. Int. J. Food Microbiol. 99, 237–243. doi: 10.1016/j.ijfoodmicro.2004.08.017
Comitini, F., Gobbi, M., Domizio, P., Romani, C., Lencioni, L., Mannazzu, I., et al. (2011). Selected non-Saccharomyces wine yeasts in controlled multistarter fermentations with Saccharomyces cerevisiae. Food Microbiol. 28, 873–882. doi: 10.1016/j.fm.2010.12.001
Contreras, A., Hidalgo, C., Schmidt, S., Henschke, P. A., Curtin, C., and Varela, C. (2015). The application of non-Saccharomyces yeast in fermentations with limited aeration as a strategy for the production of wine with reduced alcohol content. Int. J. Food Microbiol. 205, 7–15. doi: 10.1016/j.ijfoodmicro.2015.03.027
Cordente, A. G., Curtin, C. D., Varela, C., and Pretorius, I. S. (2012). Flavour-active wine yeasts. Appl. Microbiol. Biotechnol. 96, 601–618. doi: 10.1007/s00253-012-4370-z
Cordero-Bueso, G., Esteve-Zarzoso, B., Cabellos, J. M., Gil-Díaz, M., and Arroyo, T. (2013). Biotechnological potential of non-Saccharomyces yeasts isolated during spontaneous fermentations of Malvar (Vitis vinifera Cv. L.). Eur. Food Res. Technol. 236, 193–207. doi: 10.1007/s00217-012-18749
Cordero-Otero, R. R., Úbeda-Iranzo, J. F., Briones-Pérez, A. I., Potgieter, N. M., Villena, A., Pretorius, I. S., et al. (2003). Characterization of the β-glucosidase activity produced by enological strains of non-Saccharomyces yeasts. J. Food. Sci. 68, 2564–2569. doi: 10.1111/j.1365-2621.2003.tb07062.x
Cordonnier, C., and Bayonove, R. (1974). Mise en évidence dans le baie de raisin, variété Muscat d’Alexandrie, de monoterpènes liés révélables par une ou plusieurs enzymes du fruit. C. R. Acad. Sci. Ser. D 278, 3387–3390.
Ĉuŝ, F., and Jenko, M. (2013). The influence of yeast strains on the composition and sensory quality of Gewürztraminer wine. Food Technol. Biotechnol. 51, 547–553.
Darriet, P., Tominaga, T., Lavigne, V., Boidron, J.-N., and Dubourdieu, D. (1995). Identification of a powerful aromatic component of Vitis vinifera L. Var. Sauvignonwines: 4-mercapto-4-methylpentan-2-one. Flavour Fragr. J. 10, 385–392. doi: 10.1002/ffj.2730100610
Dashko, S., Zhou, N., Tinta, T., Sivilotti, P., Sternad Lemut, M., Trost, K., et al. (2015). Use of non-conventional yeast improves the wine aroma profile of Ribolla Gialla. J. Ind. Microbiol. Biotechnol. 42, 997–1010. doi: 10.1007/s10295-015-1620-y
del Mónaco, S. M., Barda, N. B., Rubio, N. C., and Caballero, A. C. (2014). Selection and characterization of a patagonian Pichia kudriavzevii for wine deacidification. J. Appl. Microbiol. 117, 451–464. doi: 10.1111/jam.12547
Di Stefano, R., Maggiorotto, G., and Gianotti, S. (1992). Bioconversion of nerol and geraniol during fermentation. Riv. Vitic. Enol. 45, 43–49.
Dias, L., Dias, S., Sancho, T., Stender, H., Querol, A., Malfeito-Ferreira, M., et al. (2003). Identification of yeasts isolated from wine-related environments and capable of producing 4-ethylphenol. Food Microbiol. 20, 567–574. doi: 10.1016/S0740-0020(02)00152-1
Domizio, P., Liu, Y., Bisson, L. F., and Barile, D. (2014). Use of non-Saccharomyces wine yeasts as novel sources of mannoproteins in wine. Food Microbiol. 43, 5–15. doi: 10.1016/j.fm.2014.04.005
du Toit, M., and Pretorius, I. S. (2000). Microbial spoilage and preservations of wine: using weapons from nature’s own arsenal-a review. S. Afr. J. Enol. Vitic. 21, 74–96.
Dubourdieu, D. (1996). The aroma of Sauvignon: improvement by winemaking. Rev. Oenol. Tech. Vitivin. Oenol. 79, 18–20. doi: 10.1111/j.1750-3841.2012.02705.x
Dubourdieu, D., Tominaga, T., Masneuf, I., Peyrot des Gachons, C., and Murat, M. L. (2006). The role of yeasts in grape flavor development during fermentation: the example of Sauvignon Blanc. Am. J. Enol. Vitic. 57, 81–88.
Ebeler, S. E., and Thorngate, J. H. (2009). Wine chemistry and flavor: looking into the crystal glass. J. Agric. Food Chem. 57, 8098–8108. doi: 10.1021/jf9000555
Egli, C. M., Edinger, W. D., Mitrakul, C. M., and Henick-Kling, T. (1998). Dynamics of indigenous and inoculated yeast populations and their effect on the sensory character of Riesling and Chardonnay wines. J. Appl. Microbiol. 85, 779–789. doi: 10.1046/j.1365-2672.1998.00521.x
Englezos, V., Rantsiou, K., Torchio, F., Rolle, L., Gerbi, V., and Cocolin, L. (2015). Exploitation of the non-Saccharomyces yeast Starmerella bacillaris (synonym Candida zemplinina) in wine fermentation: physiological and molecular characterizations. Int. J. Food Microbiol. 199, 33–40. doi: 10.1016/j.ijfoodmicro.2015.01.009
Epifanio, S. I., Gutiérrez, A. R., Santamaría, M. P., and López, R. (1999). The influence of enological practices on the selection of wild yeast strains in spontaneous fermentation. Am. J. Enol. Vitic. 50, 219–224.
Erten, H., and Tanguler, H. (2010). Influence of Williopsis saturnus yeasts in combination with Saccharomyces cerevisiae on wine fermentation. Lett. Appl. Microbiol. 50, 474–479. doi: 10.1111/j.1472-765X.2010.02822.x
Ewart, A. J. W., Brien, C. J., Soderlund, R., and Smart, R. E. (1985). The effects of light pruning, irrigation and improved soil-management on wine quality of the Vitis vinifera CV Riesling. Vitis J. Grapevine Res. 24, 209–217.
Fang, Y., and Qian, M. C. (2006). Quantification of selected aroma-active compounds in Pinot Noir wines from different grape maturities. J. Agric. Food Chem. 54, 8567–8573. doi: 10.1021/jf061396m
Fang, Y., and Qian, M. C. (2016). Development of C13-norisoprenoids, carotenoids and other volatile compounds in Vitis vinifera L. Cv. Pinot Noir grapes. Food Chem. 192, 633–641. doi: 10.1016/j.foodchem.2015.07.050
Fernández-González, M., Di Stefano, R., and Briones, A. I. (2003). Hydrolysis and transformation of terpene glycosides from Muscat must by different yeast species. Food Microbiol. 20, 35–41. doi: 10.1016/S0740-0020(02)00105-3
Fia, G., Giovani, G., and Rosi, I. (2005). Study of β-glucosidase production by wine-related yeasts during alcoholic aermentation. A new rapid fluorimetric method to determine enzymatic activity. J. Appl. Microbiol. 99, 509–517. doi: 10.1111/j.1365-2672.2005.02657.x
Fleet, G. H. (1993). “The microorganisms of winemaking-isolation enumeration and identification,” in Wine Microbiology and Biotechnology, ed. G. H. Fleet (Chur: Harwood Academic Publishers), 1–25.
Fleet, G. H. (2008). Wine yeasts for the future. FEMS Yeast Res. 8, 979–995. doi: 10.1111/j.1567-1364.2008.00427.x
Fleet, G. H., and Heard, G. M. (1993). “Yeast-growth during winemaking,” in Wine Microbiology and Biotechnology, ed. G. H. Fleet (Chur: Harwood Academic Publishers), 27–54.
Fleet, G. H., Lafon-Lafourcade, S., and Ribereau-Gayon, P. (1984). Evolution of yeasts and lactic acid bacteria during fermentation and storage of Bordeaux wines. Appl. Environ. Microbiol. 48, 1034–1038.
Frezier, V., and Dubourdieu, D. (1992). Ecology of yeast strain Saccharomyces cerevisiae during spontaneous fermentation in a Bordeaux winery. Am. J. Enol. Vitic. 4, 375–380.
Gallander, J. F. (1977). Deacidification of eastern table wines with Schizosaccharomyces pombe. Am. J. Enol. Vitic. 28, 65–72.
Garavaglia, J., Andressa, H., Rodrigues Bjerk, T., De Cassia de Souza Schneider, R., Welke, J. E., and Alcaraz Zini, C. (2014). A new method for rapid screening of ester-producing yeasts using in situ HS-SPME. J. Microbiol. Methods 103, 1–2. doi: 10.1016/j.mimet.2014.05.001
Garavaglia, J., De Cassia De Souza Schneider, R., Camargo Mendes, S. D., Welke, J. E., Alcaraz Zini, C., Bastos Caramão, E., et al. (2015). Evaluation of Zygosaccharomyces bailii BCV 08 as a co-starter in wine fermentation for the improvement of ethyl esters production. Microbiol. Res. 173, 59–65. doi: 10.1016/j.micres.2015.02.002
García, A., Carcel, C., Dulau, L., Samson, A., Aguera, E., Agosin, E., et al. (2002). Influence of a mixed culture with Debaryomyces vanriji and Saccharomyces cerevisiae on the volatiles of a Muscat wine. J. Food Sci. 67, 1138–1143. doi: 10.1111/j.1365-2621.2002.tb09466.x
Genovés, S., Gil, J. V., Manzanares, P., Aleixandre, J. L., and Vallés, S. (2003). Production by Saccharomyces cerevisiae and its application in winemaking. J. Food Sci. 68, 2096–2100. doi: 10.1111/j.1365-2621.2003.tb07025.x
Gil, J. V., Manzanares, P., Genovés, S., Vallés, S., and González-Candelas, L. (2005). Over-production of the major exoglucanase of Saccharomyces cerevisiae leads to an increase in the aroma of wine. Int. J. Food Microbiol. 103, 57–68. doi: 10.1016/j.ijfoodmicro.2004.11.026
Gil, J. V., Mateo, J. J., Jiménez, M., Pastor, A., and Huerta, T. (1996). Aroma compounds in wine as influenced by apiculate yeasts. J. Food Sci. 61, 1247–1250. doi: 10.1111/j.1365-2621.1996.tb10971.x
Gobbi, M., Comitini, F., Domizio, P., Romani, C., Lencioni, L., Mannazzu, I., et al. (2013). Lachancea thermotolerans and Saccharomyces cerevisiae in simultaneous and sequential co-fermentation: a strategy to enhance acidity and improve the overall quality of wine. Food Microbiol. 33, 271–281. doi: 10.1016/j.fm.2012.10.004
González, R., Quirós, M., and Morales, P. (2013). Yeast respiration of sugars by non-Saccharomyces yeast species: a promising and barely explored approach to lowering alcohol content of wines. Trends Food Sci. Technol. 29, 55–61. doi: 10.1016/j.tifs.2012.06.015
González-Pombo, P., Farina, L., Carrau, F., Batista-Viera, F., and Brena, B. M. (2011). A novel extracellular β-glucosidase from Issatchenkia terricola: isolation, immobilization and application for aroma enhancement of white Muscat wine. Process Biochem. 46, 385–389. doi: 10.1016/j.procbio.2010.07.016
González-Pombo, P., Pérez, G., Carrau, F., Guisán, J. M., Batista-Viera, F., and Brena, B. M. (2008). One-step purification and characterization of an intracellular β-glucosidase from Metschnikowia pulcherrima. Biotechnol. Lett. 30, 1469–1475. doi: 10.1007/s10529-008-9708-3
González-Royo, E., Pascual, O., Kontoudakis, N., Esteruelas, M., Esteve-Zarzoso, B., Mas, A., et al. (2015). Oenological consequences of sequential inoculation with non-Saccharomyces yeasts (Torulaspora delbrueckii or Metschnikowia pulcherrima) and Saccharomyces cerevisiae in base wine for sparkling wine production. Eur. Food Res. Technol. 240, 999–1012. doi: 10.1007/s00217-014-2404-8
Goto, S. (1980). Changes in the wild yeast flora of sulfited grape musts. J. Inst. Enol. Vitic. Yamanashi Univ. 15, 29–32.
Granchi, L., Ganucci, D., Messini, A., Rosellini, D., and Vicenzini, M. (1998). Dynamics of yeast populations during the early stages of natural fermentations for the production of Brunello de Montalcino wines. Food Technol. Biotechnol. 36, 313–318.
Grangeteau, C., Gerhards, D., Rousseaux, S., von Wallbrunn, C., Alexandre, H., and Guilloux-Benatier, M. (2015). Diversity of yeast strains of the genus Hanseniaspora in the winery environment: what is their involvement in grape must fermentation? Food Microbiol. 50, 70–77. doi: 10.1016/j.fm.2015.03.009
Grant-Preece, P. A., Pardon, K. H., Capone, D. L., Cordente, A. G., Sefton, M. A., Jeffery, D. W., et al. (2010). Synthesis of wine thiol conjugates and labeled analogues: fermentation of the glutathione conjugate of 3-mercaptohexan-1-ol yields the corresponding cysteine conjugate and free thiol. J. Agric. Food Chem. 58, 1383–1389. doi: 10.1021/jf9037198
Gunata, Z., Bitteur, S., Brillouet, J. M., Bayonove, C., and Cordonnier, R. (1988). Sequential enzymatic hydrolisis of potentially aromatic glycosides form grapes. Carbohydr. Res. 184, 139–149. doi: 10.1016/0008-6215(88)80012-0
Heard, G. M., and Fleet, G. H. (1985). Growth of natural yeast flora during the fermentation of inoculated wines. Appl. Environ. Microbiol. 50, 727–728.
Henick-Kling, T., Edinger, W., Daniel, P., and Monk, P. (1998). Selective effects of sulfur dioxide and yeast starter culture addition on indigenous yeast populations and sensory characteristics of wine. J. Appl. Microbiol. 84, 865–876. doi: 10.1046/j.1365-2672.1998.00423.x
Hernández-Orte, P., Cersosimo, M., Loscos, N., Cacho, J., García-Moruno, E., and Ferreira, V. (2008). The development of varietal aroma from non-floral grapes by yeasts of different genera. Food Chem. 107, 1064–1077. doi: 10.1016/j.foodchem.2007.09.032
Hernández-Orte, P., Concejero, B., Astrain, J., Lacau, B., Cacho, J., and Ferreira, V. (2015). Influence of viticulture practices on grape aroma precursors and their relation with wine aroma. J. Sci. Food Agric. 95, 688–701. doi: 10.1002/jsfa.6748
Herraiz, T., Reglero, G., Herraiz, M., Martín-Álvarez, P. J., and Cabezudo, M. D. (1990). The influence of the yeast and type of culture on the volatile composition of wines fermented without sulfur dioxide. Am. J. Enol. Vitic. 41, 313–318.
Izquierdo-Cañas, P. M., García-Romero, E., Heras Manso, J. M., and Fernández-González, M. (2014). Influence of sequential inoculation of Wickerhamomyces anomalus and Saccharomyces cerevisiae in the quality of red wines. Eur. Food Res. Technol. 239, 279–286. doi: 10.1007/s00217-014-2220-1
Izquierdo-Cañas, P. M., Palacios-García, A. T., and García-Romero, E. (2011). Enhancement of flavour properties in wines using sequential inoculations of non-Saccharomyces (Hansenula and Torulaspora) and Saccharomyces yeast starter. Vitis J. Grapevine Res. 50, 177–182.
Jolly, N. P., Augustyn, O. H. P., and Pretorius, I. S. (2003). The effect of non-Saccharomyces yeasts on fermentation and wine quality. S. Afr. J. Enol. Vitic. 24, 55–62.
Jolly, N. P., Augustyn, O. H. P., and Pretorius, I. S. (2006). The role and use of non- Saccharomyces yeasts in wine production. S. Afr. J. Enol. Vitic. 27, 15–39.
Kapsopoulou, K., Kapaklis, A., and Spyropoulos, H. (2005). Growth and fermentation characteristics of a strain of the wine yeast Kluyveromyces thermotolerans isolated in Greece. World J. Microbiol. Biotechnol. 21, 1599–1602. doi: 10.1007/s11274-005-8220-3
Kapsopoulou, K., Mourtzini, A., Anthoulas, M., and Nerantzis, E. (2007). Biological acidification during grape must fermentation using mixed cultures of Kluyveromyces thermotolerans and Saccharomyces cerevisiae. World J. Microbiol. Biotechnol. 23, 735–739. doi: 10.1007/s11274-006-9283-5
Kim, D. H., Hong, Y. A., and Park, H. D. (2008). Co-fermentation of grape must by Issatchenkia orientalis and Saccharomyces cerevisiae reduces the malic acid content in wine. Biotechnol. Lett. 30, 1633–1638. doi: 10.1007/s10529-008-9726-1
King, A., and Dickinson, J. R. (2000). Biotransformation of monoterpene alcohols by Saccharomyces cerevisiae, Torulaspora delbrueckii and Kluyveromyces lactis. Yeast 16, 499–506.
Lambrechts, M. G., and Pretorius, I. S. (2000). Yeast and its importance to wine aroma – a review. S. Afr. J. Enol. Vitic. 21, 97–129.
Lange, J. N., Faasse, E., Tantikachornkiat, M., Gustafsson, F. S., Halvorsen, L. C., Kluftinger, A., et al. (2014). Implantation and persistence of yeast inoculum in Pinot Noir fermentations at three Canadian wineries. Int. J. Food Microbiol. 180, 56–61. doi: 10.1016/j.ijfoodmicro.2014.04.003
Le Jeune, C., Erny, C., Demuyter, C., and Lollier, M. (2006). Evolution of the population of Saccharomyces cerevisiae from grape to wine in a spontaneous fermentation. Food Microbiol. 23, 709–716. doi: 10.1016/j.fm.2006.02.007
Loira, I., Morata, A., Comuzzo, P., Callejo, M. J., González, C., Calderón, F., et al. (2015). Use of Schizosaccharomyces pombe and Torulaspora delbrueckii strains in mixed and sequential fermentations to improve red wine sensory quality. Food Res. Int. 76, 325–333. doi: 10.1016/j.foodres.2015.06.030
Loira, I., Vejarano, R., Bañuelos, M. A., Morata, A., Tesfaye, W., Uthurry, C., et al. (2014). Influence of sequential fermentation with Torulaspora delbrueckii and Saccharomyces cerevisiae on wine quality. LWT Food Sci. Technol. 59, 915–922. doi: 10.1016/j.lwt.2014.06.019
López, M. C., Mateo, J. J., and Maicas, S. (2015). Screening of β-glucosidase and β-xylosidase activities in four non-Saccharomyces yeast isolates. J. Food Sci. 80, 1696–1704. doi: 10.1111/1750-3841.12954
Loureiro, V., and Malfeito-Ferreira, M. (2003). Spoilage yeasts in the wine industry. Int. J. Food Microbiol. 86, 23–50. doi: 10.1016/S0168-1605(03)00246-0
Manzanares, P., Ramon, D., and Querol, A. (1999). Screening of non-Saccharomyces yeasts for the production of β-D-xylosidase activity. Int. J. Food Microbiol. 46, 105–112. doi: 10.1016/S0168-1605(98)00186-X
Manzanares, P., Rojas, V., Genovés, S., and Vallés, S. (2000). A preliminary search for anthocyanin-β-D-glucosidase activity in non-Saccharomyces wine yeast. Int. J. Food Sci. Technol. 35, 95–103. doi: 10.1046/j.1365-2621.2000.00364.x
Martínez, J., Millán, C., and Ortega, J. M. (1989). Growth of natural flora during the fermentation of inoculated musts from ‘Pedro Ximenez’ Grapes. S. Afr. J. Enol. Vitic. 10, 31–35.
Martini, A., Frederici, F., and Rosini, G. (1980). A new approach to the study of yeast ecology of natural substances. Can. J. Microbiol. 26, 856–859. doi: 10.1139/m80-149
Mateo, J. J., and Jiménez, M. (2000). Monoterpenes in grape juice and wines. J. Chromatogr. A 881, 557–567. doi: 10.1016/S0021-9673(99)01342-4
Mateo, J. J., Jiménez, M., Huerta, T., and Pastor, A. (1991). Contribution of different yeasts isolated from musts of Monastrell grapes to the aroma of wine. Int. J. Food Microbiol. 14, 153–160. doi: 10.1016/0168-1605(91)90102-U
Mateo, J. J., Peris, L., Ibáñez, C., and Maicas, S. (2011). Characterization of glycolytic activities from non-Saccharomyces yeasts isolated from Bobal musts. J. Ind. Microbiol. Biotechnol. 38, 347–354. doi: 10.1007/s10295-010-0780-z
Maturano, Y. P., Assof, M., Fabani, M. P., Nally, M. C., Jofré, V., Rodríguez Assaf, L. A., et al. (2015). Enzymatic activities produced by mixed Saccharomyces and non-Saccharomyces cultures: relationship with wine volatile composition. Antonie Van Leeuwenhoek 108, 1239–1256. doi: 10.1007/s10482-015-0578-0
Maturano, Y. P., Rodríguez Assaf, L. A., Toro, M. E., Nally, M. C., Vallejo, M., Castellanos de Figueroa, L. I., et al. (2012). Multi-enzyme production by pure and mixed cultures of Saccharomyces and non-Saccharomyces yeasts during wine fermentation. Int. J. Food Microbiol. 155, 43–50. doi: 10.1016/j.ijfoodmicro.2012.01.015
McMahon, H., Zoecklein, B. W., Fugelsang, K., and Jasinski, Y. (1999). Quantification of glycosidase activities in selected yeasts and lactic acid bacteria. J. Ind. Microbiol. Biotechnol. 23, 198–203. doi: 10.1038/sj.jim.2900720
Medina, K., Boido, E., Fariña, L., Gioia, O., Gómez, M. E., Barquet, M., et al. (2013). Increased flavour diversity of Chardonnay wines by spontaneous fermentation and co-fermentation with Hanseniaspora vineae. Food Chem. 141, 2513–2521. doi: 10.1016/j.foodchem.2013.04.056
Mendoza, L. M., De Nadra, M. C. M., and Farías, M. E. (2007). Kinetics and metabolic behavior of a composite culture of Kloeckera apiculata and Saccharomyces cerevisiae wine related strains. Biotechnol. Lett. 29, 1057–1063. doi: 10.1007/s10529-007-9355-0
Mercado, L., Dalcero, A., Masuelli, R., and Combina, M. (2007). Diversity of Saccharomyces strains on grapes and winery surfaces: analysis of their contribution to fermentative flora of Malbec wine from Mendoza (Argentina) during two consecutive years. Food Microbiol. 24, 403–412. doi: 10.1016/j.fm.2006.06.005
Mora, J., Barbas, J. I., and Mulet, A. (1990). Growth of yeast species during the fermentation of musts inoculated with Kluyveromyces thermotolerans and Saccharomyces cerevisiae. Am. J. Enol. Vitic. 41, 156–159.
Morales, P., Rojas, V., Quirós, M., and González, R. (2015). The impact of oxygen on the final alcohol content of wine fermented by a mixed starter culture. Appl. Microbiol. Biotechnol. 99, 3993–4003. doi: 10.1007/s00253-014-6321-3
Morata, A., Benito, S., Loira, I., Palomero, F., González, M. C., and Suárez-Lepe, J. A. (2012). Formation of pyranoanthocyanins by Schizosaccharomyces pombe during the fermentation of red must. Int. J. Food Microbiol. 159, 47–53. doi: 10.1016/j.ijfoodmicro.2012.08.007
Moreira, N., Mendes, F., Guedes de Pinho, P., Hogg, T., and Vasconcelos, I. (2008). Heavy sulphur compounds, higher alcohols and esters production profile of Hanseniaspora uvarum and Hanseniaspora guilliermondii grown as pure and mixed cultures in grape must. Int. J. Food Microbiol. 124, 231–238. doi: 10.1016/j.ijfoodmicro.2008.03.025
Moreira, N., Mendes, F., Hogg, T., and Vasconcelos, I. (2005). Alcohols, esters and heavy sulphur compounds production by pure and mixed cultures of apiculate wine yeasts. Int. J. Food Microbiol. 103, 285–294. doi: 10.1016/j.ijfoodmicro.2004.12.029
Nykänen, L. (1986). Formation and occurrence of flavor compounds in wine and distilled alcoholic beverages. Am. J. Enol. Vitic. 37, 84–96.
Oro, L., Ciani, M., and Comitini, F. (2014). Antimicrobial activity of Metschnikowia pulcherrima on wine yeasts. J. Appl. Microbiol. 116, 1209–1217. doi: 10.1111/jam.12446
Ough, C. S., Cook, J. A., and Lider, L. A. (1968). Rootstock-scion interactions concerning wine making. II. Wine compositional and sensory changes attributed to rootstock and fertilizer differences. Am. J. Enol. Vitic. 19, 254–265.
Parish, M. E., and Caroll, D. E. (1985). Indigenous yeasts associated with Muscadine (Vitis rotundifolia) grapes and musts. Am. J. Enol. Vitic. 36, 165–169.
Peynaud, E., and Domercq, S. (1959). A review of microbiological problems in winemaking in France. Am. J. Enol. Vitic. 10, 69–77.
Pineau, B., Barbe, J. C., Van Leeuwen, C., and Dubordieu, D. (2007). Which impact for β-Damascenone on red wines aroma? J. Agric. Food Chem. 55, 4103–4108. doi: 10.1021/jf070120r
Plata, C., Millán, C., Mauricio, J. C., and Ortega, J. M. (2003). Formation of ethyl acetate and isoamyl acetate by various species of wine yeasts. Food Microbiol. 20, 217–224. doi: 10.1016/S0740-0020(02)00101-6
Pretorius, I. S. (2000). Tailoring wine yeast for the new millennium: novel approaches to the ancient art of winemaking. Yeast 16, 675–729. doi: 10.1002/1097-0061(20000615)16:8<675::AID-YEA585>3.0.CO;2-B
Pretorius, I. S. (2003). “The genetic analysis and tailoring of wine yeasts,” in Functional Genetics of Industrial Yeasts, Vol. 2, Topics in Current Genetics, ed. J. H. DeWinde (Berlin: Springer-Verlag), 99–142.
Pretorius, I. S., Van Der Westhuizen, T. J., and Augustyn, O. P. H. (1999). Yeast biodiversity in vineyards and wineries and its importance to the South African wine industry-A review. S. Afr. J. Enol. Vitic. 20, 61–75.
Querol, A., Jiménez, M., and Huerta, T. (1990). Microbiological and enological parameters during fermentation of musts from poor and normal grape harvests in the region of Alicante. J. Food Sci. 55, 1603–1606. doi: 10.1111/j.1365-2621.1990.tb03580.x
Radler, F. (1993). “Yeasts-metabolism of organic acids,” in Wine Microbiology and Biotechnology, ed. G. H. Fleet (Chur: Harwood Academic Publishers), 165–182.
Rantsiou, K., Dolci, P., Giacosa, S., Torchio, F., Tofalo, R., Torriani, S., et al. (2012). Candida zemplinina can reduce acetic acid produced by Saccharomyces cerevisiae in sweet wine fermentations. Appl. Environ. Microbiol. 78, 1987–1994. doi: 10.1128/AEM.06768-11
Rapp, A. (1998). Volatile flavour of wine: correlation between instrumental nalysis and sensory perception. Nahrung 42, 351–363. doi: 10.1002/(SICI)1521-3803(199812)42:06<351::AID-FOOD351>3.3.CO;2-U
Rapp, A., and Versini, G. (1991). “Influence of nitrogen compounds in grapes on aroma compounds of wine,” in International Symposium on Nitrogen in Grapes and Wines, ed. J. M. Rantz (Davis, CA: American Society for Enology and Viticulture), 156–164.
Reed, G., and Peppler, H. J. (1973). Yeast Technology. Westport, CT: The AVI Publishing Company, Inc.
Regueiro, L. A., Costas, C. L., and Rubio, J. E. L. (1993). Influence of viticultural and enological practices on the development of yeast populations during winemaking. Am. J. Enol. Vitic. 44, 405–408.
Renault, P., Coulon, J., de Revel, G., Barbe, J. C., and Bely, M. (2015). Increase of fruity aroma during mixed T. delbrueckii/S. Cerevisiae wine fermentation is linked to specific esters enhancement. Int. J. Food Microbiol. 207, 40–48. doi: 10.1016/j.ijfoodmicro.2015.04.037
Renault, P., Miot-Sertier, C., Marullo, P., Hernández-Orte, P., Lagarrigue, L., Lonvaud-Funel, A., et al. (2009). Genetic characterization and phenotypic variability in Torulaspora delbrueckii species: potential applications in the wine industry. Int. J. Food Microbiol. 134, 201–210. doi: 10.1016/j.ijfoodmicro.2009.06.008
Ribereau-Gayon, P., Glories, Y., Maugean, A., and Dubordieu, D. (1998). Chimie Du Vin, Stabilisation et Traitements. Paris: Dunod.
Ristic, R., Bindon, K., Francis, L. I., Herderich, M. J., and Iland, P. G. (2010). Flavonoids and C13-norisoprenoids in Vitis vinifera L. Cv. Shiraz: relationships between grape and wine composition, wine colour and wine sensory properties. Aust. J. Grape Wine Res. 16, 369–388. doi: 10.1111/j.1755-0238.2010.00099.x
Rodríguez, M. E., Lopes, C. A., Barbagelata, R. J., Barda, N. B., and Caballero, A. C. (2010a). Influence of Candida pulcherrima Patagonian strain on alcoholic fermentation behaviour and wine aroma. Int. J. Food Microbiol. 138, 19–25. doi: 10.1016/j.ijfoodmicro.2009.12.025
Rodríguez, M. E., Lopes, C. A., Vallés, S., and Caballero, A. C. (2010b). Characterization of α-rhamnosidase activity from a Patagonian Pichia guilliermondii wine strain. J. Appl. Microbiol. 109, 2206–2213. doi: 10.1111/j.1365-2672.2010.04854.x
Rodríguez, M. E., Lopes, C. A., Van Broock, M., Vallés, S., Ramón, D., and Caballero, A. C. (2004). Screening and typing of Patagonian wine yeasts for glycosidase activities. J. Appl. Microbiol. 96, 84–95. doi: 10.1046/j.1365-2672.2003.02032.x
Rojas, V., Gil, J. V., Piñaga, F., and Manzanares, P. (2001). Studies on acetate ester production by non-Saccharomyces wine yeasts. Int. J. Food Microbiol. 70, 283–289. doi: 10.1016/S0168-1605(01)00552-9
Rojas, V., Gil, J. V., Piñaga, F., and Manzanares, P. (2003). Acetate ester formation in wine by mixed cultures in laboratory fermentations. Int. J. Food Microbiol. 86, 181–188. doi: 10.1016/S0168-1605(03)00255-1
Romano, P., Fiore, C., Paraggio, M., Caruso, M., and Capece, A. (2003). Function of yeast species and strains in wine flavour. Int. J. Food Microbiol. 86, 169–180. doi: 10.1016/S0168-1605(03)00290-3
Romano, P., and Suzzi, G. (1993). Higher alcohol and acetoin production by Zygosaccharomyces wine yeasts. J. Appl. Bacteriol. 75, 541–545. doi: 10.1111/j.1365-2672.1993.tb01592.x
Romano, P., Suzzi, G., Domizio, P., and Fatichenti, F. (1997). Secondary products formation as a tool for discriminating non-Saccharomyces wine strains. Strain diversity in non-Saccharomyces wine yeasts. Antonie van Leeuwenhoek 71, 239–242. doi: 10.1023/A:1000102006018
Roncoroni, M., Santiago, M., Hooks, D. O., Moroney, S., Harsch, M. J., Lee, S. A., et al. (2011). The yeast IRC7 gene encodes a β-lyase responsible for production of the varietal thiol 4-mercapto-4-methylpentan-2-one in wine. Food Microbiol. 28, 926–935. doi: 10.1016/j.fm.2011.01.002
Rosi, I., Vinella, M., and Domizio, P. (1994). Characterization of β-glucosidase activity in yeasts of oenological origin. J. Appl. Bacteriol. 77, 519–527. doi: 10.1111/j.1365-2672.1994.tb04396.x
Rosini, G. (1984). Assessment of dominance of added yeast in wine fermentation and origin of Saccharomyces cerevisiae in wine-making. J. Gen. Appl. Microbiol. 30, 249–256. doi: 10.2323/jgam.30.249
Rosini, G., Federici, F., and Martini, A. (1982). Yeast flora of grape berries during ripening. Microb. Ecol. 8, 83–89. doi: 10.1007/BF02011464
Sabel, A., Martens, S., Petri, A., König, H., and Claus, H. (2014). Wickerhamomyces anomalus AS1: a new strain with potential to improve wine aroma. Ann. Microbiol. 64, 483–491. doi: 10.1007/s13213-013-0678-x
Sadoudi, M., Tourdot-Maréchal, R., Rousseaux, S., Steyer, D., Gallardo-Chacón, J. J., Ballester, J., et al. (2012). Yeast-yeast interactions revealed by aromatic profile analysis of Sauvignon Blanc wine fermented by single or co-culture of non-Saccharomyces and Saccharomyces yeasts. Food Microbiol. 32, 243–253. doi: 10.1016/j.fm.2012.06.006
Santamaría, P., Garijo, P., López, R., Tenorio, C., and Gutiérrez, A. R. (2005). Analysis of yeast population during spontaneous alcoholic fermentation: effect of the age of the cellar and the practice of inoculation. Int. J. Food Microbiol. 103, 49–56. doi: 10.1016/j.ijfoodmicro.2004.11.024
Schütz, M., and Gafner, J. (1993). Analysis of yeast diversity during spontaneous and induced alcoholic fermentations. J. Appl. Bacteriol. 75, 551–558. doi: 10.1111/j.1365-2672.1993.tb01594.x
Schwentke, J., Sabel, A., Petri, A., Konig, H., and Claus, H. (2014). The yeast Wickerhamomyces anomalus ASI secretes a multtifunctional exo-β-1,3-glucanase with implications for winemaking. Yeast 31, 349–359. doi: 10.1002/yea.3029
Shinohara, T., Kubodera, S., and Yanagida, F. (2000). Distribution of phenolic yeasts and production of phenolic off-flavors in wine fermentation. J. Biosci. Bioeng. 90, 90–97. doi: 10.1016/S1389-1723(00)80040-7
Sidhu, D., Lund, J., Kotseridis, Y., and Saucier, C. (2015). Methoxypyrazine analysis and influence of viticultural and enological procedures on their levels in grapes, musts, and wines. Crit. Rev. Food Sci. Nutr. 55, 485–502. doi: 10.1080/10408398.2012.658587
Snow, P. G., and Gallander, J. F. (1979). Deacidification of white table wines trough partial fermentation with Schizosaccharomyces pombe. Am. J. Enol. Vitic. 30, 45–48.
Soden, A., Francis, I. L., Oakey, H., and Henschke, P. A. (2000). Effects of co-fermentation with Candida stellata and Saccharomyces cerevisiae on the aroma and composition of Chardonnay wine. Aust. J. Grape Wine Res. 6, 21–30. doi: 10.1111/j.1755-0238.2000.tb00158.x
Soles, R. M., Ough, C. S., and Kunkee, R. E. (1982). Ester concentration differences in wine fermented by various species and strains of yeasts. Am. J. Enol. Vitic. 33, 94–98.
Spagna, G., Barbagallo, R. N., Palmeri, R., Restuccia, C., and Giudici, P. (2002). Properties of endogenous β-glucosidase of a Saccharomyces cerevisiae strain isolated from Sicilian musts and wines. Enzyme Microb. Technol. 31, 1030–1035. doi: 10.1016/S0141-0229(02)00233-8
Spayd, S. E., Tarara, J. M., Mee, D. L., and Ferguson, J. C. (2002). Separation of sunlight and temperature effects on the composition of Vitis vinifera Cv. Am. J. Enol. Vitic. 53, 171–182.
Sponholz, W. R. (1993). “Wine spoilage by microorganisms,” in Wine Microbiology and Biotechnology, ed. G. H. Fleet (Chur: Harwood Academic Publishers), 395–420.
Steger, C. L. C., and Lambrechts, M. G. (2000). The selection of yeast strains for the production of premium quality South African brandy base products. J. Ind. Microbiol. Biotechnol. 24, 431–440. doi: 10.1038/sj.jim.7000005
Strauss, M. L. A., Jolly, N. P., Lambrechts, M. G., and Van Rensburg, P. (2001). Screening for the production of extracellular hydrolytic enzymes by non- Saccharomyces wine yeasts. J. Appl. Microbiol. 91, 182–190. doi: 10.1046/j.1365-2672.2001.01379.x
Suomalainen, H., and Lehtonen, M. (1979). The production of aroma compounds by yeast. J. Inst. Brew. 85, 149–156. doi: 10.1002/j.2050-0416.1979.tb06846.x
Swangkeaw, J., Vichitphan, S., Christian, E., Butzke, C. E., and Vichitphan, K. (2011). The characterisation of a novel Pichia anomala β-glucosidase with potentially aroma-enhancing capabilities in wine. Ann. Microbiol. 59, 335–343. doi: 10.1007/BF03178336
Swiegers, J. H., Kievit, R. L., Siebert, T., Lattey, K. A., Bramley, B. R., Francis, I. L., et al. (2009). The influence of yeast on the aroma of Sauvignon Blanc wine. Food Microbiol. 26, 204–211. doi: 10.1016/j.fm.2008.08.004
Swiegers, J. H., and Pretorius, I. S. (2005). Yeast modulation of wine flavor. Adv. Appl. Microbiol. 57, 131–175. doi: 10.1016/S0065-2164(05)57005-9
Swiegers, J. H., Willmott, R. L., Hill-Ling, A., Capone, D. L., Pardon, K. H., Elsey, G. M., et al. (2006). “Modulation of volatile thiol and ester aromas in wine by modified wine yeast,” in Developments in Food Science. Flavour Science Recent Advances and Trends, eds W. Bredie and M. Petersen (Amsterdam: Elsevier), 113–116.
Tanguler, H. (2012). Evaluation of Williopsis saturnus inoculum level on fermentation and flavor compounds of white wines made from Emir (Vitis Vinifera L.) grown in Anatolia. Food Biotechnol. 26, 351–368. doi: 10.1080/08905436.2012.724038
Tanguler, H. (2013). Influence of temperatures and fermentation behaviour of mixed cultures of Williopsis saturnus var. saturnus and Saccharomyces cerevisiae associated with winemaking. Food Sci. Technol. Res. 19, 781–793. doi: 10.3136/fstr.19.781
Teixeira, A., Caldeira, I., and Duarte, F. L. (2015). Molecular and oenological characterization of Touriga Nacional non-Saccharomyces yeasts. J. Appl. Microbiol. 118, 658–671. doi: 10.1111/jam.12727
Tominaga, T., Darriet, P., and Dubourdieu, D. (1996). Identification de l’acétate de 3-mercaptohexanol, composé à forte odeur de buis, intervenant dans l’arome des vins de Sauvignon. Vitis J. Grapevine Res. 35, 207–210.
Tominaga, T., Furrer, A., Henry, R., and Dubourdieu, D. (1998a). Identification of new volatile thiols in the aroma of Vitis vinifera L. Var. Sauvignon Blanc wines. Flavour Fragr. J. 13, 159–162. doi: 10.1002/(SICI)1099-1026(199805/06)13:3<159::AID-FFJ709>3.0.CO;2-7
Tominaga, T., Peyrot des Gachons, C., and Dubourdieu, D. (1998b). A new type of flavor precursors in Vitis vinifera L. Cv. Sauvignon Blanc: S-cysteine conjugates. J. Agric. Food Chem. 46, 5215–5219. doi: 10.1021/jf980481u
Úbeda, J., and Briones, A. I. (2000). Characterization of differences in the formation of volatiles during fermentation within synthetic and grape musts by wild Saccharomyces strains. LWT Food Sci. Technol. 33, 408–414. doi: 10.1006/fstl.2000.0680
Ugliano, M., Bartowsky, E. J., McCarthy, J., Moio, L., and Henschke, P. A. (2006). Hydrolysis and transformation of grape glycosidically bound volatile compounds during fermentation with three Saccharomyces yeast strains. J. Agric. Food Chem. 54, 6322–6331. doi: 10.1021/jf0607718
Viana, F., Belloch, C., Vallés, S., and Manzanares, P. (2011). Monitoring a mixed starter of Hanseniaspora vineae-Saccharomyces cerevisiae in natural must: impact on 2-phenylethyl acetate production. Int. J. Food Microbiol. 151, 235–240. doi: 10.1016/j.ijfoodmicro.2011.09.005
Viana, F., Gil, J. V., Genovés, S., Vallés, S., and Manzanares, P. (2008). Rational selection of non-Saccharomyces wine yeasts for mixed starters based on ester formation and enological traits. Food Microbiol. 25, 778–785. doi: 10.1016/j.fm.2008.04.015
Viana, F., Gil, J. V., Vallés, S., and Manzanares, P. (2009). Increasing the levels of 2-phenylethyl acetate in wine through the use of a mixed culture of Hanseniaspora osmophila and Saccharomyces cerevisiae. Int. J. Food Microbiol. 135, 68–74. doi: 10.1016/j.ijfoodmicro.2009.07.025
Wang, Y., Kang, W., Xu, Y., and Li, J. (2011). Effect of different indigenous yeast β-glucosidases on the liberation of bound aroma compounds. J. Inst. Brew. 117, 230–237. doi: 10.1002/j.2050-0416.2011.tb00466.x
Wang, Y., Xu, Y., and Li, J. (2012). A novel extracellular β-Glucosidase from Trichosporon asahii: yield prediction, evaluation and application for aroma enhancement of Cabernet Sauvignon. J. Food Sci. 77, 505–515. doi: 10.1111/j.1750-3841.2012.02705.x
Williams, P. J., Strauss, C. R., Wilson, B., and Massy-Westropp, R. A. (1982). Novel monoterpene disacharide glycosides of Vitis vinifera grapes and wines. Phytochemistry 21, 2013–2020. doi: 10.1016/0031-9422(82)83034-3
Winterhalter, P., and Skouroumounis, G. K. (1997). Glycoconjugated aroma compounds: occurrence, role and biotechnological transformation. Adv. Biochem. Eng. Biotechnol. 55, 73–105.
Yanai, T., and Sato, M. (1999). Isolation and properties of β-glucosidase produced by Debaryomyces hansenii and its application in winemakig. Am. J. Enol. Vitic. 50, 231–235.
Yanai, T., and Sato, M. (2000a). Purifiction and characteriztion of an α-L-rhamnosidase from Pichia angusta X349. Biosci. Biotechnol. Biochem. 64, 2179–2185. doi: 10.1271/bbb.64.2179
Yanai, T., and Sato, M. (2000b). Purification and characterization of a novel α-L-arabinofuranosidase from Pichia capsulata X91. Biosci. Biotechnol. Biochem. 64, 1181–1188. doi: 10.1271/bbb.64.1181
Yanai, T., and Sato, M. (2001). Purification and characterization of an β-D-xylosidase from Candida utilis IFO 0639. Biosci. Biotechnol. Biochem. 65, 527–533. doi: 10.1271/bbb.65.527
Zalacain, A., Marín, J., Alonso, G. L., and Salinas, M. R. (2007). Analysis of wine primary aroma compounds by stir bar sorptive extraction. Talanta 71, 1610–1615. doi: 10.1016/j.talanta.2006.07.051
Zohre, D. E., and Erten, H. (2002). The influence of Kloeckera apiculata and Candida pulcherrima yeasts on wine fermentation. Process Biochem. 38, 319–324. doi: 10.1016/S0032-9592(02)00086-9
Keywords: non-Saccharomyces yeasts, enzymes, secondary metabolites, primary aroma, secondary aroma, mixed starters, aroma complexity
Citation: Padilla B, Gil JV and Manzanares P (2016) Past and Future of Non-Saccharomyces Yeasts: From Spoilage Microorganisms to Biotechnological Tools for Improving Wine Aroma Complexity. Front. Microbiol. 7:411. doi: 10.3389/fmicb.2016.00411
Received: 12 January 2016; Accepted: 14 March 2016;
Published: 31 March 2016.
Edited by:
Andrea Gomez-Zavaglia, Center for Research and Development in Food Cryotechnology (CIDCA), ArgentinaReviewed by:
Maurizio Ciani, Università Politecnica delle Marche, ItalyCopyright © 2016 Padilla, Gil and Manzanares. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Paloma Manzanares, cG1hbnpAaWF0YS5jc2ljLmVz
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.