
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
MINI REVIEW article
Front. Microbiol. , 17 February 2016
Sec. Microbial Physiology and Metabolism
Volume 7 - 2016 | https://doi.org/10.3389/fmicb.2016.00185
This article is part of the Research Topic Insights into microbe-microbe interactions in human microbial ecosystems: strategies to be competitive View all 11 articles
 David Ríos-Covián
David Ríos-Covián Patricia Ruas-Madiedo
Patricia Ruas-Madiedo Abelardo Margolles
Abelardo Margolles Miguel Gueimonde
Miguel Gueimonde Clara G. de los Reyes-Gavilán
Clara G. de los Reyes-Gavilán Nuria Salazar*
Nuria Salazar*The colon is inhabited by a dense population of microorganisms, the so-called “gut microbiota,” able to ferment carbohydrates and proteins that escape absorption in the small intestine during digestion. This microbiota produces a wide range of metabolites, including short chain fatty acids (SCFA). These compounds are absorbed in the large bowel and are defined as 1-6 carbon volatile fatty acids which can present straight or branched-chain conformation. Their production is influenced by the pattern of food intake and diet-mediated changes in the gut microbiota. SCFA have distinct physiological effects: they contribute to shaping the gut environment, influence the physiology of the colon, they can be used as energy sources by host cells and the intestinal microbiota and they also participate in different host-signaling mechanisms. We summarize the current knowledge about the production of SCFA, including bacterial cross-feedings interactions, and the biological properties of these metabolites with impact on the human health.
The gut microbiota influences our health and nutritional stage via multiple mechanisms, and a mounting body of evidence recognizes that microbial metabolites have a major influence on host physiology. Short chain fatty acids (SCFA) are volatile fatty acids produced by the gut microbiota in the large bowel as fermentation products from food components that are unabsorbed/undigested in the small intestine; they are characterized by containing fewer than six carbons, existing in straight, and branched-chain conformation. Acetic acid (C2), propionic acid (C3), and butyric acid (C4) are the most abundant, representing 90–95% of the SCFA present in the colon. The main sources of SCFA are carbohydrates (CHO) but amino acids valine, leucine, and isoleucine obtained from protein breakdown can be converted into isobutyrate, isovalerate, and 2-methyl butyrate, known as branched-chain SCFA (BSCFA), which contribute very little (5%) to total SCFA production. The aim of the present mini-review is to summarize the current knowledge about SCFA production, including bacterial cross-feedings interactions, and the biological properties of these metabolites with impact in human health.
The main end products resulting from the CHO catabolism of intestinal microbes are acetate, propionate, and butyrate. Lactate, although is not a SCFA, is also produced by some members of the microbiota, such as lactic acid bacteria, bifidobacteria, and proteobacteria, but under normal physiological conditions it does not accumulate in the colon due to the presence of some species, such as Eubacterium hallii, that can convert lactate into different SCFA (Flint et al., 2015).
Acetate is the most abundant SCFA in the colon and makes up more than half of the total SCFA detected in feces (Louis et al., 2007). Two main metabolic routes have been described for acetate production by the gut microbiota (Figure 1). The majority of acetate is produced by most enteric bacteria as a result of CHO fermentation. In addition, approximately one-third of the colonic acetate is coming from acetogenic bacteria, which are able to synthesize it from hydrogen and carbon dioxide or formic acid through the Wood–Ljungdahl pathway (Miller and Wolin, 1996; Louis et al., 2014).
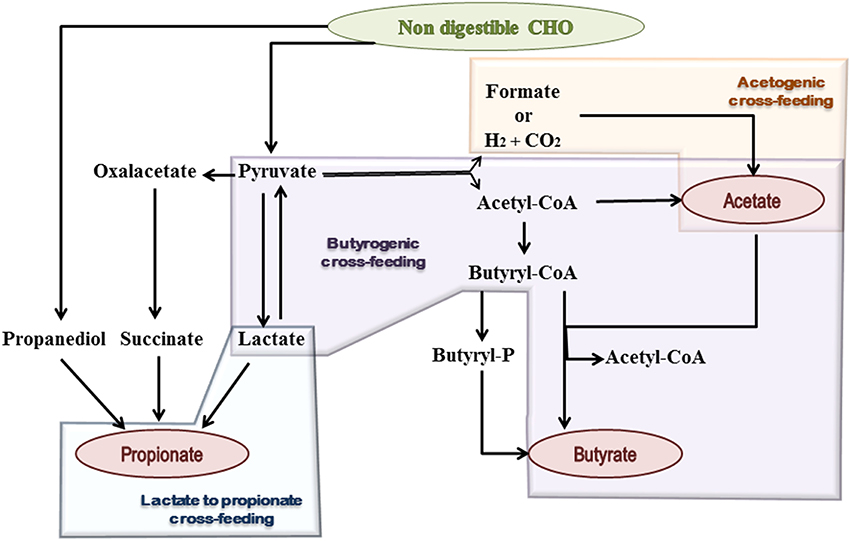
Figure 1. Schematic representation of microbial metabolic pathways and cross-feeding mechanisms, contributing to SCFA formation in the human gut. Shaded geometric shapes summarize routes of formation for each of the three main SCFA: acetate, propionate, and butyrate.
Propionate and butyrate metabolism have received much attention during the last years, mainly due to the connection between low levels of butyrate and propionate bacterial producers and some diseases in which inflammatory processes are involved. For instance, butyrate producers are normally low in ulcerative colitis (Machiels et al., 2014) and reduced levels of propionate producers have been detected in children at risk of asthma (Arrieta et al., 2015).
Three different pathways are used by colonic bacteria for propionate formation: succinate pathway, acrylate pathway, and propanodiol pathway (Reichardt et al., 2014) (Figure 1). The succinate route utilizes succinate as a substrate for propionate formation and involves the descarboxylation of methylmalonyl-CoA to propionyl-CoA. This pathway is present in several Firmicutes, belonging to the Negativicutes class, and in Bacteroidetes. In the acrylate pathway lactate is converted to propionate through the activity of the lactoyl-CoA dehydratase and downstream enzymatic reactions; this route appears to be limited to a few members of the families Veillonellaceae and Lachnospiraceae (Flint et al., 2015). In the propanodiol pathway, characterized by the conversion of deoxy-sugars to propionate, the CoA-dependent propionaldehyde dehydrogenase, that converts propionaldehyde to propionyl-CoA, has been suggested as a marker for this route. This metabolic pathway is present in bacteria which are phylogenetically distant, including proteobacteria and members of the Lachnospiraceae family (Louis et al., 2014; Reichardt et al., 2014). The relative abundance of Bacteroidetes has also been linked to the total fecal propionate concentration, suggesting that the succinate pathway is the dominant route within the gut microbiota (Salonen et al., 2014).
Two different pathways for butyrate production are known in butyrate-producing bacteria (Figure 1). The butyrate kinase pathway employs phosphotransbutyrylase and butyrate kinase enzymes to convert butyryl-CoA into butyrate (Louis et al., 2004). This route is not common among members of the gut microbiota and is mainly limited to some Coprococcus species (Flint et al., 2015). In contrast, the butyryl-CoA: acetate CoA-transferase pathway, in which butyryl-CoA is converted to butyrate in a single step enzymatic reaction, is used by the majority of gut butyrate-producers (Louis et al., 2010), including some of the most abundant genera of the intestinal microbiota, such as Faecalibacterium, Eubacterium, and Roseburia. Remarkably, the production of butyrate and propionate by the same bacterium is not common and only a few anaerobes, such as Roseburia inulinivorans and Coprococcus catus, are able to produce both (Louis et al., 2014).
Bacterial cross-feeding has a huge impact on the final balance of SCFA production and the efficient exploitation of the substrates that reach the human gut. These mechanisms consist either in the utilization of end products from the metabolism of a given microorganism by another one, called metabolic cross-feeding (Figure 1), and/or the utilization by one microorganism of the energy rich complex CHO breakdown products formed by another one, called substrate cross-feeding (Belenguer et al., 2006; Flint et al., 2007). A recent in silico study showed that mutualism cross-feeding interactions were promoted by anoxic conditions, which are more common in the large intestine than in the small one (Heinken and Thiele, 2015).
Microorganisms that are not capable of using complex CHO may proliferate by taking advantage of substrate cross-feeding, using breakdown compounds produced by hydrolytic bacteria. This is the case of some Bifidobacterium species that are not able to use inulin-type fructans (ITF) but can grow by cross-feeding of mono- and oligosaccharides released by primary inulin degraders in fecal cultures added with inulin as carbon source (Rossi et al., 2005; Salazar et al., 2009). Other example is the degradation of agaro-oligosaccharides (AO), which is more effective when Bacteroides uniformis and Escherichia coli are grown in co-culture than in separated monoculture (Li et al., 2014). In the same study the authors suggest the utilization of agarotriose, an intermediate in the degradation of AO, by Bifidobacterium adolescentis and Bifidobacterium infantis. In another work, it was demonstrated that Roseburia sp. strain A2-183 is unable to use lactate as carbon source, but when it is co-cultured with B. adolescentis L2-32 in the presence of FOS or starch, produces butyrate (Belenguer et al., 2006).
Although, there are a lot of in vitro studies pointing to metabolic cross-feeding it was not until recently that was demonstrated in vivo by using stable isotopes of acetate, propionate and butyrate perfused into the caecum of mice (Den Besten et al., 2013a). This study evidenced that the bacterial cross-feeding occurred mainly from acetate to butyrate, at lower extent between butyrate and propionate, and almost no metabolic flux exists between propionate and acetate. In vitro utilization of acetate by Faecalibacterium prausnitzii and Roseburia sp. has been evidenced (Duncan et al., 2002, 2004b). Prediction of metabolic fluxes between F. prausnitzii A2-165 and B. adolescentis L2-32 in co-culture has been reported in a computational model (El-Semman et al., 2014) and was recently demonstrated in vitro (Rios-Covián et al., 2015). Moreover, a recent animal study suggests that F. prausnitzii is able to use the acetate produced by Bacteroides thetaiotaomicron in vivo, this interaction having a significant impact in the modulation of the intestinal mucus barrier (Wrzosek et al., 2013). Although lactate is not a SCFA, it is used by some butyrate and propionate producing bacteria, avoiding metabolic acidosis in the host (El Aidy et al., 2013). Several in vitro studies confirm that lactate and/or acetate produced by Bifidobacterium when grown in oligofructose, is used by members of Roseburia, Eubacterium, and Anaeroestipes genera (Duncan et al., 2004a,b; Belenguer et al., 2006; Falony et al., 2006). Members of Veillonella and Propionibacterium are capable of transforming lactate to propionate in vitro (Counotte et al., 1981). H2 plays an important role in cross-feeding as well. Co-cultures of Roseburia intestinalis with the methanogen Methanobrevibacter smithii and the acetogen Blautia hydrogenotrophica, resulted in a decrease of final H2 and the production of CH4 and acetate. The acetate formed is used by R. intestinalis to produce butyrate (Chassard and Bernalier-Donadille, 2006). Ba. thetaiotaomicron bi-associated mice with Bl. hydrogenotrophica showed higher levels of acetate in caecal contents and lower NADH/NAD+ ratio; the removal of H2 by B. hydrogenotrophyca in this case allows Ba. thetaoitaomicron to regenerate NAD+ (Rey et al., 2010).
Diet affects the gut microbiota composition and activity, and therefore the profile of SCFA and BSCFA synthesized, this having a deep impact on human health (Brussow and Parkinson, 2014; Louis et al., 2014). The first work linking the long-term diet style with the so-called human “enterotypes” was published in 2011 (Wu et al., 2011) but it has been also demonstrated that short-term diets can alter the human gut microbiome (David et al., 2014). The amount and relative abundance of SCFA may be considered as biomarkers of a healthy status (Table 1A). For example, high fiber-low fat and meat diets are characterized by the presence of higher amounts of fecal SCFA than diets with reduced fiber intake (De Filippo et al., 2010; Cuervo et al., 2013; Ou et al., 2013). A reduction in fecal butyrate has been found in patients with colorectal adenocarcinoma (Chen et al., 2013), whereas obesity has been related with increases in total fecal SCFA concentration (Fernandes et al., 2014; Rahat-Rozenbloom et al., 2014) which tend to decrease following an anti-obesity treatment (Patil et al., 2012). These epidemiological data have been further supported by dietary intervention studies carried out with different human populations (Table 1B). Prebiotic substrates that selectively promote the growth of beneficial microbiota also induce changes in SCFA production of healthy individuals (Lecerf et al., 2012) and in patients with irritable bowel syndrome or those receiving enteral nutrition (Majid et al., 2011; Halmos et al., 2015). Interestingly, the consumption of dairy products fermented with beneficial bacteria also modifies the intestinal microbiota toward more butyrate producers in comparison to chemically-acidified milk (Veiga et al., 2014). Finally, dietary intervention studies carried out with different overweight and obese populations seemed to be effective in lowering the high levels of fecal SCFA associated with the obesity status (Salazar et al., 2015).
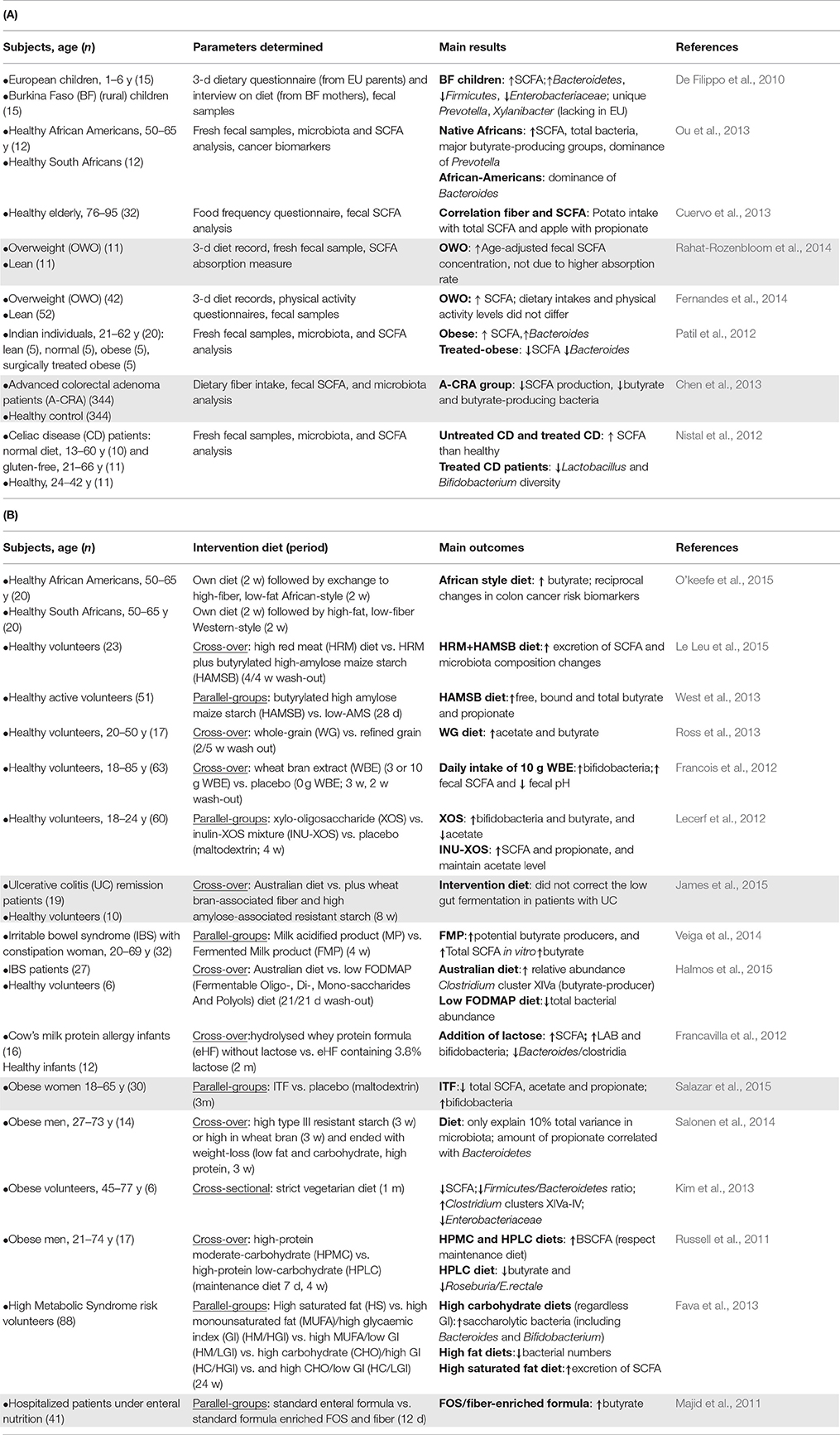
Table 1. (A) Epidemiological studies, carried out since 2010, showing the impact of diet on SCFA produced by the gut microbiota. The shaded areas indicate a change in the populations analyzed in terms of their health status. D, days; y, year. (B) Intervention studies, carried out since 2010, showing the impact of diet on SCFA produced by the gut microbiota. The shaded areas indicate a change in the populations analyzed in terms of their health status. D, day; w, week; m, month; y, year.
Although animal and human trials provide the best models for studying the influence of diet on the gut microbiota, in vitro fecal cultures constitute simpler approaches for investigating the interactions of diet and food components with the intestinal microbiota. Available in vitro models range from simple batch fermentation (Salazar et al., 2009; Arboleya et al., 2013b) to complex multi-stage continuous culture systems. The SHIME (Van Den Abbeele et al., 2010) and SIMGI models (Barroso et al., 2015) simulate the digestion from stomach to colon whereas the EnteroMix (Makivuokko et al., 2005) and the Lacroix models mimic the entire colonic process. TIM-2 reproduces the proximal colon and incorporates a dialysis membrane that simulates absorption of microbial metabolites by the body (Minekus et al., 1999). A microbial bias regarding butyrate and propionate producers occurs with some of these models (Van Den Abbeele et al., 2010), that could be alleviated by incorporating a simulation of the intestinal mucosa surface (Van Den Abbeele et al., 2013a). Labelling substrates with the stable isotope 13C makes possible to link the fermentation with specific members of the microbiota and to quantify production of metabolites (Maathuis et al., 2012) whilst the mathematical modeling is becoming a useful tool to study microbe-diet-host interactions (Shoaie et al., 2015).
When studying in vitro the influence of dietary components on microbial composition, the main aim usually is to increase beneficial bacteria and to enhance the production of SCFA whereas minimizing the synthesis of BSCFA. The fermentation of different substrates has been evaluated, ITF being the most studied (Sivieri et al., 2014). Starch (Fassler et al., 2006), arabinans, arabinoxylans (Van Den Abbeele et al., 2013b), galactooligosaccharides (Rodriguez-Colinas et al., 2013), xylitol (Makelainen et al., 2007), and lactulose (Cardelle-Cobas et al., 2009) have been also considered. The influence of polyphenols on the gut microbiota metabolism is currently receiving considerable attention (Valdés et al., 2015). Different microbial fermentation patterns can be obtained depending on physico-chemical characteristics of the substrates, speed of fermentation and the microbial populations involved in the process (initial breakdown of long polymers, direct fermentation of substrates, and cross-feeding interactions; Hernot et al., 2009; Zhou et al., 2013; Puertollano et al., 2014). Probiotics and their extracellular components (exopolysaccharides), can also act as modulators of SCFA microbial formation (Salazar et al., 2009; Van Zanten et al., 2012). In addition, a large number of studies highlight the influence of different foods and long-term diets on the intestinal microbiota activity and specifically, over the pattern of SCFA (Yang and Rose, 2014; Costabile et al., 2015).
The basal microbiota composition has also a profound influence on the final effects exerted in vitro by diet on microbial populations and metabolic activity (Arboleya et al., 2013a; Souza et al., 2014). In this regard, it has been found a different response to probiotics and prebiotics by the microbiota of individuals from different groups of age (Arboleya et al., 2013a; Likotrafiti et al., 2014), or between obese and lean people (Yang et al., 2013).
One of the health effects attributed to the production of SCFA is the concomitant reduction of the luminal pH, which by itself inhibits pathogenic microorganisms and increases the absorption of some nutrients (Macfarlane and Macfarlane, 2012). Acetate has been found to be a key player in the ability of bifidobacteria to inhibit enteropathogens (Fukuda et al., 2011). Moreover, butyrate fuels the intestinal epithelial cells and increases mucin production which may result in changes on bacterial adhesion (Jung et al., 2015) and improved tight-junctions integrity (Peng et al., 2009). Thus, the production of SCFA seems to play an important role in the maintenance of the gut barrier function.
After their production, SCFA will be absorbed and used in different biosynthetic routes by the host (Den Besten et al., 2013b). During the intestinal absorption process part of the SCFA, mainly butyrate, will be metabolized by the colonocytes (Pryde et al., 2002) whilst the rest will be transported by the hepatic vein and go into the liver, where they will be metabolized (Den Besten et al., 2013b). These SCFA will enter diverse CHO and lipid metabolic routes; propionate will mainly incorporate into gluconeogenesis whilst acetate and butyrate will be mostly introduced into the lipid biosynthesis. The involvement of SCFA in energy and lipid metabolism attracted the attention of researchers toward the potential role of SCFA in the control of metabolic syndrome. A reduction in obesity and insulin resistance in experimental animals on high-fat diet after dietary supplementation with butyrate has been observed (Gao et al., 2009). This protective effect of SCFA on the high-fat diet-induced metabolic alterations seems to be dependent on down-regulation of the peroxisome proliferator-activated receptor gamma (PPARγ), therefore promoting a change from lipid synthesis to lipids oxidation (Den Besten et al., 2015). Interestingly although the three main intestinal SCFA have a protective effect on diet-induced obesity, butyrate and propionate seem to exert larger effects than acetate (Lin et al., 2012). Different mechanisms have been proposed to explain these effects, the activation of signaling pathways mediated by protein kinases, such as AMP-activated protein kinase (Gao et al., 2009; Peng et al., 2009; Den Besten et al., 2015) or mitogen-activated protein kinases (MAPK; Jung et al., 2015), being a common observation. Butyrate and propionate, but not acetate, have been reported to induce the production of gut hormones, thus reducing food intake (Lin et al., 2012). Acetate has also been found to reduce the appetite, in this case through the interaction with the central nervous system (Frost et al., 2014). However, in spite of these promising animal data, controlled human intervention studies are still needed before drawing firm conclusions (Canfora et al., 2015).
It has also been observed that SCFA protect against the development of colorectal cancer (CRC), with most studies focusing on butyrate (Canani et al., 2011; Keku et al., 2015). Butyrate promotes colon motility, reduces inflammation, increases visceral irrigation, induces apoptosis, and inhibits tumor cell progression (Zhang et al., 2010; Canani et al., 2011; Leonel and Alvarez-Leite, 2012; Keku et al., 2015), all of these properties being beneficial in CRC prevention. In cancerous colonocytes, due to the Warburg effect, butyrate accumulates, which increases its activity as inhibitor of histone deacetylation, promoting apoptosis of CRC cells. Interestingly, a recent animal study suggests that the protective effect of dietary fiber upon CRC is dependent on the production of butyrate by the microbiota (Donohoe et al., 2014).
In addition, butyrate and propionate have also been reported to induce the differentiation of T-regulatory cells, assisting to control intestinal inflammation; this effect seems to be mediated via inhibition of histone deacetylation (Donohoe et al., 2014; Louis et al., 2014). This control of intestinal inflammation may result beneficial in terms of gut barrier maintenance, reducing the risk of inflammatory bowel disease or CRC. Unlike what happens with the three main intestinal SCFA, acetate, propionate, and butyrate, little is known about the potential health effects of other SCFA.
The main role of diet is to provide enough macro- and micronutrients to fulfill daily requirements and well-being. However, during the last decades the association between dietary intake and physiology has been increasingly-recognized, although many of the molecular and immunological aspects by which dietary components could influence human health remain still largely unknown. Bacterial fermentation of CHO and proteins produces SCFA which emerge as major mediators in linking nutrition, gut microbiota, physiology and pathology. Many biological effects seem to be mediated by these bacterial metabolites but a conclusive proof is not available for many of the health claims made for SCFA. Promising in vitro and animal studies have been published but they cannot be easily extrapolated to the human situation. The design of improved approaches combining in vitro, in vivo, and “omics” technologies should be carried out, with emphasis in human intervention trials, to explore the mechanisms of production and action of SCFA, thus opening the possibility to find strategies for developing personalized nutrition.
All authors listed, have made substantial, direct and intellectual contribution to the work, and approved it for publication.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer BJ and handling Editor declared their shared affiliation, and the handling Editor states that the process nevertheless met the standards of a fair and objective review
This work was financed by project AGL2013-43770-R from Plan Nacional/Plan Estatal de I+D+I (Spanish Ministry of Economy and Competitiveness, MINECO). The activity of Probiotics and Prebiotics Group is being partly supported through the Grant GRUPIN14-043 from “Plan Regional de Investigación del Principado de Asturias.” Both, national and regional grants received cofounding from European Union FEDER funds. DR was the recipient of predoctoral FPI fellowship whereas NS benefits from a Clarín postdoctoral contract (Marie Curie European CoFund Program) cofinanced by Plan Regional de Investigación del Principado de Asturias, Spain.
Arboleya, S., Salazar, N., Solís, G., Fernández, N., Gueimonde, M., and De Los Reyes-Gavilán, C. G. (2013a). In vitro evaluation of the impact of human background microbiota on the response to Bifidobacterium strains and fructo-oligosaccharides. Br. J. Nutr. 110, 2030–2036. doi: 10.1017/S0007114513001487
Arboleya, S., Salazar, N., Solís, G., Fernández, N., Hernández-Barranco, A. M., Cuesta, I., et al. (2013b). Assessment of intestinal microbiota modulation ability of Bifidobacterium strains in in vitro fecal batch cultures from preterm neonates. Anaerobe 19, 9–16. doi: 10.1016/j.anaerobe.2012.11.001
Arrieta, M. C., Stiemsma, L. T., Dimitriu, P. A., Thorson, L., Russell, S., Yurist-Doutsch, S., et al. (2015). Early infancy microbial and metabolic alterations affect risk of childhood asthma. Sci. Transl. Med. 7, 307ra152. doi: 10.1126/scitranslmed.aab2271
Barroso, E., Cueva, C., Pelaez, C., Martinez-Cuesta, M. C., and Requena, T. (2015). Development of human colonic microbiota in the computer-controlled dynamic SIMulator of the GastroIntestinal tract SIMGI. Lwt Food Sci. Technol. 61, 283–289. doi: 10.1016/j.lwt.2014.12.014
Belenguer, A., Duncan, S. H., Calder, A. G., Holtrop, G., Louis, P., Lobley, G. E., et al. (2006). Two routes of metabolic cross-feeding between Bifidobacterium adolescentis and butyrate-producing anaerobes from the human gut. Appl. Environ. Microbiol. 72, 3593–3599. doi: 10.1128/AEM.72.5.3593-3599.2006
Brussow, H., and Parkinson, S. J. (2014). You are what you eat. Nat. Biotechnol. 32, 243–245. doi: 10.1038/nbt.2845
Canani, R. B., Costanzo, M. D., Leone, L., Pedata, M., Meli, R., and Calignano, A. (2011). Potential beneficial effects of butyrate in intestinal and extraintestinal diseases. World J. Gastroenterol. 17, 1519–1528. doi: 10.3748/wjg.v17.i12.1519
Canfora, E. E., Jocken, J. W., and Blaak, E. E. (2015). Short-chain fatty acids in control of body weight and insulin sensitivity. Nat. Rev. Endocrinol. 11, 577–591. doi: 10.1038/nrendo.2015.128
Cardelle-Cobas, A., Fernández, M., Salazar, N., Martínez-Villaluenga, C., Villamiel, M., Ruas-Madiedo, P., et al. (2009). Bifidogenic effect and stimulation of short chain fatty acid production in human faecal slurry cultures by oligosaccharides derived from lactose and lactulose. J. Dairy Res. 76, 317–325. doi: 10.1017/S0022029909004063
Chassard, C., and Bernalier-Donadille, A. (2006). H2 and acetate transfers during xylan fermentation between a butyrate-producing xylanolytic species and hydrogenotrophic microorganisms from the human gut. FEMS Microbiol. Lett. 254, 116–122. doi: 10.1111/j.1574-6968.2005.00016.x
Chen, H. M., Yu, Y. N., Wang, J. L., Lin, Y. W., Kong, X., Yang, C. Q., et al. (2013). Decreased dietary fiber intake and structural alteration of gut microbiota in patients with advanced colorectal adenoma. Am. J. Clin. Nutr. 97, 1044–1052. doi: 10.3945/ajcn.112.046607
Costabile, A., Walton, G. E., Tzortzis, G., Vulevic, J., Charalampopoulos, D., and Gibson, G. R. (2015). Effects of orange juice formulation on prebiotic functionality using an in vitro colonic model system. PLoS ONE 10:e0121955. doi: 10.1371/journal.pone.0121955
Counotte, G. H., Prins, R. A., Janssen, R. H., and Debie, M. J. (1981). Role of Megasphaera elsdenii in the Fermentation of dl-[2-C]lactate in the rumen of dairy cattle. Appl. Environ. Microbiol. 42, 649–655.
Cuervo, A., Salazar, N., Ruas-Madiedo, P., Gueimonde, M., and Gonzalez, S. (2013). Fiber from a regular diet is directly associated with fecal short-chain fatty acid concentrations in the elderly. Nutr. Res. 33, 811–816. doi: 10.1016/j.nutres.2013.05.016
David, L. A., Maurice, C. F., Carmody, R. N., Gootenberg, D. B., Button, J. E., Wolfe, B. E., et al. (2014). Diet rapidly and reproducibly alters the human gut microbiome. Nature 505, 559–563. doi: 10.1038/nature12820
De Filippo, C., Cavalieri, D., Di Paola, M., Ramazzotti, M., Poullet, J. B., Massart, S., et al. (2010). Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc. Natl. Acad. Sci. U.S.A. 107, 14691–14696. doi: 10.1073/pnas.1005963107
Den Besten, G., Bleeker, A., Gerding, A., Van Eunen, K., Havinga, R., Van Dijk, T. H., et al. (2015). Short-chain fatty acids protect against high-fat diet-induced obesity via a PPARgamma-Dependent Switch from lipogenesis to fat oxidation. Diabetes 64, 2398–2408. doi: 10.2337/db14-1213
Den Besten, G., Lange, K., Havinga, R., Van Dijk, T. H., Gerding, A., Van Eunen, K., et al. (2013a). Gut-derived short-chain fatty acids are vividly assimilated into host carbohydrates and lipids. Am. J. Physiol. Gastrointest. Liver Physiol. 305, G900–G910. doi: 10.1152/ajpgi.00265.2013
Den Besten, G., Van Eunen, K., Groen, A. K., Venema, K., Reijngoud, D. J., and Bakker, B. M. (2013b). The role of short-chain fatty acids in the interplay between diet, gut microbiota and host energy metabolism. J. Lipid Res. 54, 2325–2340. doi: 10.1194/jlr.R036012
Donohoe, D. R., Holley, D., Collins, L. B., Montgomery, S. A., Whitmore, A. C., Hillhouse, A., et al. (2014). A gnotobiotic mouse model demonstrates that dietary fiber protects against colorectal tumorigenesis in a microbiota- and butyrate-dependent manner. Cancer Discov. 4, 1387–1397. doi: 10.1158/2159-8290.CD-14-0501
Duncan, S. H., Barcenilla, A., Stewart, C. S., Pryde, S. E., and Flint, H. J. (2002). Acetate utilization and butyryl coenzyme A (CoA):acetate-CoA transferase in butyrate-producing bacteria from the human large intestine. Appl. Environ. Microbiol. 68, 5186–5190. doi: 10.1128/AEM.68.10.5186-5190.2002
Duncan, S. H., Holtrop, G., Lobley, G. E., Calder, A. G., Stewart, C. S., and Flint, H. J. (2004a). Contribution of acetate to butyrate formation by human faecal bacteria. Br. J. Nutr. 91, 915–923. doi: 10.1079/BJN20041150
Duncan, S. H., Louis, P., and Flint, H. J. (2004b). Lactate-utilizing bacteria, isolated from human feces, that produce butyrate as a major fermentation product. Appl. Environ. Microbiol. 70, 5810–5817. doi: 10.1128/AEM.70.10.5810-5817.2004
El Aidy, S., Van Den Abbeele, P., Van De Wiele, T., Louis, P., and Kleerebezem, M. (2013). Intestinal colonization: how key microbial players become established in this dynamic process: microbial metabolic activities and the interplay between the host and microbes. Bioessays 35, 913–923. doi: 10.1002/bies.201300073
El-Semman, I. E., Karlsson, F. H., Shoaie, S., Nookaew, I., Soliman, T. H., and Nielsen, J. (2014). Genome-scale metabolic reconstructions of Bifidobacterium adolescentis L2-32 and Faecalibacterium prausnitzii A2-165 and their interaction. BMC Syst. Biol. 8:41. doi: 10.1186/1752-0509-8-41
Falony, G., Vlachou, A., Verbrugghe, K., and De Vuyst, L. (2006). Cross-feeding between Bifidobacterium longum BB536 and acetate-converting, butyrate-producing colon bacteria during growth on oligofructose. Appl. Environ. Microbiol. 72, 7835–7841. doi: 10.1128/AEM.01296-06
Fassler, C., Arrigoni, E., Venema, K., Brouns, F., and Amado, R. (2006). In vitro fermentability of differently digested resistant starch preparations. Mol. Nutr. Food Res. 50, 1220–1228. doi: 10.1002/mnfr.200600106
Fava, F., Gitau, R., Griffin, B. A., Gibson, G. R., Tuohy, K. M., and Lovegrove, J. A. (2013). The type and quantity of dietary fat and carbohydrate alter faecal microbiome and short-chain fatty acid excretion in a metabolic syndrome 'at-risk' population. Int. J. Obes. (Lond.) 37, 216–223. doi: 10.1038/ijo.2012.33
Fernandes, J., Su, W., Rahat-Rozenbloom, S., Wolever, T. M., and Comelli, E. M. (2014). Adiposity, gut microbiota and faecal short chain fatty acids are linked in adult humans. Nutr. Diabetes 4, e121. doi: 10.1038/nutd.2014.23
Flint, H. J., Duncan, S. H., Scott, K. P., and Louis, P. (2007). Interactions and competition within the microbial community of the human colon: links between diet and health. Environ. Microbiol. 9, 1101–1111. doi: 10.1111/j.1462-2920.2007.01281.x
Flint, H. J., Duncan, S. H., Scott, K. P., and Louis, P. (2015). Links between diet, gut microbiota composition and gut metabolism. Proc. Nutr. Soc. 74, 13–22. doi: 10.1017/s0029665114001463
Francavilla, R., Calasso, M., Calace, L., Siragusa, S., Ndagijimana, M., Vernocchi, P., et al. (2012). Effect of lactose on gut microbiota and metabolome of infants with cow's milk allergy. Pediatr. Allergy Immunol. 23, 420–427. doi: 10.1111/j.1399-3038.2012.01286.x
Francois, I. E., Lescroart, O., Veraverbeke, W. S., Marzorati, M., Possemiers, S., Evenepoel, P., et al. (2012). Effects of a wheat bran extract containing arabinoxylan oligosaccharides on gastrointestinal health parameters in healthy adult human volunteers: a double-blind, randomised, placebo-controlled, cross-over trial. Br. J. Nutr. 108, 2229–2242. doi: 10.1017/S0007114512000372
Frost, G., Sleeth, M. L., Sahuri-Arisoylu, M., Lizarbe, B., Cerdan, S., Brody, L., et al. (2014). The short-chain fatty acid acetate reduces appetite via a central homeostatic mechanism. Nat. Commun. 5, 3611. doi: 10.1038/ncomms4611
Fukuda, S., Toh, H., Hase, K., Oshima, K., Nakanishi, Y., Yoshimura, K., et al. (2011). Bifidobacteria can protect from enteropathogenic infection through production of acetate. Nature 469, 543–547. doi: 10.1038/nature09646
Gao, Z., Yin, J., Zhang, J., Ward, R. E., Martin, R. J., Lefevre, M., et al. (2009). Butyrate improves insulin sensitivity and increases energy expenditure in mice. Diabetes 58, 1509–1517. doi: 10.2337/db08-1637
Halmos, E. P., Christophersen, C. T., Bird, A. R., Shepherd, S. J., Gibson, P. R., and Muir, J. G. (2015). Diets that differ in their FODMAP content alter the colonic luminal microenvironment. Gut 64, 93–100. doi: 10.1136/gutjnl-2014-307264
Heinken, A., and Thiele, I. (2015). Anoxic conditions promote species-specific mutualism between gut microbes in silico. Appl. Environ. Microbiol. 81, 4049–4061. doi: 10.1128/AEM.00101-15
Hernot, D. C., Boileau, T. W., Bauer, L. L., Middelbos, I. S., Murphy, M. R., Swanson, K. S., et al. (2009). In vitro fermentation profiles, gas production rates, and microbiota modulation as affected by certain fructans, galactooligosaccharides, and polydextrose. J. Agric. Food Chem. 57, 1354–1361. doi: 10.1021/jf802484j
James, S. L., Christophersen, C. T., Bird, A. R., Conlon, M. A., Rosella, O., Gibson, P. R., et al. (2015). Abnormal fibre usage in UC in remission. Gut 64, 562–570. doi: 10.1136/gutjnl-2014-307198
Jung, T. H., Park, J. H., Jeon, W. M., and Han, K. S. (2015). Butyrate modulates bacterial adherence on LS174T human colorectal cells by stimulating mucin secretion and MAPK signaling pathway. Nutr. Res. Pract. 9, 343–349. doi: 10.4162/nrp.2015.9.4.343
Keku, T. O., Dulal, S., Deveaux, A., Jovov, B., and Han, X. (2015). The gastrointestinal microbiota and colorectal cancer. Am. J. Physiol. Gastrointest. Liver Physiol. 308, G351–G363. doi: 10.1152/ajpgi.00360.2012
Kim, M. S., Hwang, S. S., Park, E. J., and Bae, J. W. (2013). Strict vegetarian diet improves the risk factors associated with metabolic diseases by modulating gut microbiota and reducing intestinal inflammation. Environ. Microbiol. Rep. 5, 765–775. doi: 10.1111/1758-2229.12079
Le Leu, R. K., Winter, J. M., Christophersen, C. T., Young, G. P., Humphreys, K. J., Hu, Y., et al. (2015). Butyrylated starch intake can prevent red meat-induced O6-methyl-2-deoxyguanosine adducts in human rectal tissue: a randomised clinical trial. Br. J. Nutr. 114, 220–230. doi: 10.1017/S0007114515001750
Lecerf, J. M., Depeint, F., Clerc, E., Dugenet, Y., Niamba, C. N., Rhazi, L., et al. (2012). Xylo-oligosaccharide (XOS) in combination with inulin modulates both the intestinal environment and immune status in healthy subjects, while XOS alone only shows prebiotic properties. Br. J. Nutr. 108, 1847–1858. doi: 10.1017/S0007114511007252
Leonel, A. J., and Alvarez-Leite, J. I. (2012). Butyrate: implications for intestinal function. Curr. Opin. Clin. Nutr. Metab. Care 15, 474–479. doi: 10.1097/MCO.0b013e32835665fa
Li, M., Li, G., Zhu, L., Yin, Y., Zhao, X., Xiang, C., et al. (2014). Isolation and characterization of an agaro-oligosaccharide (AO)-hydrolyzing bacterium from the gut microflora of Chinese individuals. PLoS ONE 9:e91106. doi: 10.1371/journal.pone.0091106
Likotrafiti, E., Tuohy, K. M., Gibson, G. R., and Rastall, R. A. (2014). An in vitro study of the effect of probiotics, prebiotics and synbiotics on the elderly faecal microbiota. Anaerobe 27, 50–55. doi: 10.1016/j.anaerobe.2014.03.009
Lin, H. V., Frassetto, A., Kowalik, E. J. Jr., Nawrocki, A. R., Lu, M. M., Kosinski, J. R., et al. (2012). Butyrate and propionate protect against diet-induced obesity and regulate gut hormones via free fatty acid receptor 3-independent mechanisms. PLoS ONE 7:e35240. doi: 10.1371/journal.pone.0035240PONE-D-11-22997
Louis, P., Duncan, S. H., Mccrae, S. I., Millar, J., Jackson, M. S., and Flint, H. J. (2004). Restricted distribution of the butyrate kinase pathway among butyrate-producing bacteria from the human colon. J. Bacteriol. 186, 2099–2106. doi: 10.1128/JB.186.7.2099-2106.2004
Louis, P., Hold, G. L., and Flint, H. J. (2014). The gut microbiota, bacterial metabolites and colorectal cancer. Nat. Rev. Microbiol. 12, 661–672. doi: 10.1038/nrmicro3344
Louis, P., Scott, K. P., Duncan, S. H., and Flint, H. J. (2007). Understanding the effects of diet on bacterial metabolism in the large intestine. J. Appl. Microbiol. 102, 1197–1208. doi: 10.1111/j.1365-2672.2007.03322.x
Louis, P., Young, P., Holtrop, G., and Flint, H. J. (2010). Diversity of human colonic butyrate-producing bacteria revealed by analysis of the butyryl-CoA:acetate CoA-transferase gene. Environ. Microbiol. 12, 304–314. doi: 10.1111/j.1462-2920.2009.02066.x
Maathuis, A. J. H., van den Heuvel, E. C., Schoterman, M. H. C., and Venema, K. (2012). Galacto-oligosaccharides have prebiotic activity in a dynamic in vitro colon model using a C-13-labeling technique. J. Nutr. 142, 1205–1212. doi: 10.3945/jn.111.157420
Macfarlane, G. T., and Macfarlane, S. (2012). Bacteria, colonic fermentation, and gastrointestinal health. J. AOAC Int. 95, 50–60.
Machiels, K., Joossens, M., Sabino, J., De Preter, V., Arijs, I., Eeckhaut, V., et al. (2014). A decrease of the butyrate-producing species Roseburia hominis and Faecalibacterium prausnitzii defines dysbiosis in patients with ulcerative colitis. Gut 63, 1275–1283. doi: 10.1136/gutjnl-2013-304833
Majid, H. A., Emery, P. W., and Whelan, K. (2011). Faecal microbiota and short-chain fatty acids in patients receiving enteral nutrition with standard or fructo-oligosaccharides and fibre-enriched formulas. J. Hum. Nutr. Diet 24, 260–268. doi: 10.1111/j.1365-277X.2011.01154.x
Makelainen, H. S., Makivuokko, H. A., Salminen, S. J., Rautonen, N. E., and Ouwehand, A. C. (2007). The effects of polydextrose and xylitol on microbial community and activity in a 4-stage colon simulator. J. Food Sci. 72, M153–M159. doi: 10.1111/j.1750-3841.2007.00350.x
Makivuokko, H., Nurmi, J., Nurminen, P., Stowell, J., and Rautonen, N. (2005). In vitro effects on polydextrose by colonic bacteria and caco-2 cell cyclooxygenase gene expression. Nutr. Cancer 52, 94–104. doi: 10.1207/s15327914nc5201_12
Miller, T. L., and Wolin, M. J. (1996). Pathways of acetate, propionate, and butyrate formation by the human fecal microbial flora. Appl. Environ. Microbiol. 62, 1589–1592.
Minekus, M., Smeets-Peeters, M., Bernalier, A., Marol-Bonnin, S., Havenaar, R., Marteau, P., et al. (1999). A computer-controlled system to simulate conditions of the large intestine with peristaltic mixing, water absorption and absorption of fermentation products. Appl. Microbiol. Biotechnol. 53, 108–114.
Nistal, E., Caminero, A., Vivas, S., Ruiz De Morales, J. M., Saenz De Miera, L. E., Rodriguez-Aparicio, L. B., et al. (2012). Differences in faecal bacteria populations and faecal bacteria metabolism in healthy adults and celiac disease patients. Biochimie 94, 1724–1729. doi: 10.1016/j.biochi.2012.03.025
O'keefe, S. J., Li, J. V., Lahti, L., Ou, J., Carbonero, F., Mohammed, K., et al. (2015). Fat, fibre and cancer risk in African Americans and rural Africans. Nat. Commun. 6, 6342. doi: 10.1038/ncomms7342
Ou, J., Carbonero, F., Zoetendal, E. G., Delany, J. P., Wang, M., Newton, K., et al. (2013). Diet, microbiota, and microbial metabolites in colon cancer risk in rural Africans and African Americans. Am. J. Clin. Nutr. 98, 111–120. doi: 10.3945/ajcn.112.056689
Patil, D. P., Dhotre, D. P., Chavan, S. G., Sultan, A., Jain, D. S., Lanjekar, V. B., et al. (2012). Molecular analysis of gut microbiota in obesity among Indian individuals. J. Biosci. 37, 647–657. doi: 10.1007/s12038-012-9244-0
Peng, L., Li, Z. R., Green, R. S., Holzman, I. R., and Lin, J. (2009). Butyrate enhances the intestinal barrier by facilitating tight junction assembly via activation of AMP-activated protein kinase in Caco-2 cell monolayers. J. Nutr. 139, 1619–1625. doi: 10.3945/jn.109.104638
Pryde, S. E., Duncan, S. H., Hold, G. L., Stewart, C. S., and Flint, H. J. (2002). The microbiology of butyrate formation in the human colon. FEMS Microbiol. Lett. 217, 133–139. doi: 10.1111/j.1574-6968.2002.tb11467.x
Puertollano, E., Kolida, S., and Yaqoob, P. (2014). Biological significance of short-chain fatty acid metabolism by the intestinal microbiome. Curr. Opin. Clin. Nutr. Metab. Care 17, 139–144. doi: 10.1097/MCO.0000000000000025
Rahat-Rozenbloom, S., Fernandes, J., Gloor, G. B., and Wolever, T. M. (2014). Evidence for greater production of colonic short-chain fatty acids in overweight than lean humans. Int. J. Obes. (Lond.) 38, 1525–1531. doi: 10.1038/ijo.2014.46
Reichardt, N., Duncan, S. H., Young, P., Belenguer, A., Mcwilliam Leitch, C., Scott, K. P., et al. (2014). Phylogenetic distribution of three pathways for propionate production within the human gut microbiota. ISME J. 8, 1323–1335. doi: 10.1038/ismej.2014.14
Rey, F. E., Faith, J. J., Bain, J., Muehlbauer, M. J., Stevens, R. D., Newgard, C. B., et al. (2010). Dissecting the in vivo metabolic potential of two human gut acetogens. J. Biol. Chem. 285, 22082–22090. doi: 10.1074/jbc.M110.117713
Rios-Covián, D., Gueimonde, M., Duncan, S. H., Flint, H. J., and De Los Reyes-Gavilán, C. G. (2015). Enhanced butyrate formation by cross-feeding between Faecalibacterium prausnitzii and Bifidobacterium adolescentis. FEMS Microbiol. Lett. 362:fnv176. doi: 10.1093/femsle/fnv176
Rodriguez-Colinas, B., Kolida, S., Baran, M., Ballesteros, A. O., Rastall, R. A., and Plou, F. J. (2013). Analysis of fermentation selectivity of purified galacto-oligosaccharides by in vitro human faecal fermentation. Appl. Microbiol. Biotechnol. 97, 5743–5752. doi: 10.1007/s00253-013-4892-z
Ross, A. B., Pere-Trépat, E., Montoliu, I., Martin, F.-P. J., Collino, S., Moco, S., et al. (2013). A whole-grain-rich diet reduces urinary excretion of markers of protein catabolism and gut microbiota metabolism in healthy men after one week. J. Nutr. 143, 766–777. doi: 10.3945/jn.112.172197
Rossi, M., Corradini, C., Amaretti, A., Nicolini, M., Pompei, A., Zanoni, S., et al. (2005). Fermentation of fructooligosaccharides and inulin by bifidobacteria: a comparative study of pure and fecal cultures. Appl. Environ. Microbiol. 71, 6150–6158. doi: 10.1128/AEM.71.10.6150-6158.2005
Russell, W. R., Gratz, S. W., Duncan, S. H., Holtrop, G., Ince, J., Scobbie, L., et al. (2011). High-protein, reduced-carbohydrate weight-loss diets promote metabolite profiles likely to be detrimental to colonic health. Am. J. Clin. Nutr. 93, 1062–1072. doi: 10.3945/ajcn.110.002188
Salazar, N., Dewulf, E. M., Neyrinck, A. M., Bindels, L. B., Cani, P. D., Mahillon, J., et al. (2015). Inulin-type fructans modulate intestinal Bifidobacterium species populations and decrease fecal short-chain fatty acids in obese women. Clin. Nutr. 34, 501–507. doi: 10.1016/j.clnu.2014.06.001
Salazar, N., Ruas-Madiedo, P., Kolida, S., Collins, M., Rastall, R., Gibson, G., et al. (2009). Exopolysaccharides produced by Bifidobacterium longum IPLA E44 and Bifidobacterium animalis subsp. lactis IPLA R1 modify the composition and metabolic activity of human faecal microbiota in pH-controlled batch cultures. Int. J. Food Microbiol. 135, 260–267. doi: 10.1016/j.ijfoodmicro.2009.08.017
Salonen, A., Lahti, L., Salojarvi, J., Holtrop, G., Korpela, K., Duncan, S. H., et al. (2014). Impact of diet and individual variation on intestinal microbiota composition and fermentation products in obese men. ISME J. 8, 2218–2230. doi: 10.1038/ismej.2014.63
Shoaie, S., Ghaffari, P., Kovatcheva-Datchary, P., Mardinoglu, A., Sen, P., Pujos-Guillot, E., et al. (2015). Quantifying Diet-induced metabolic changes of the human gut microbiome. Cell Metab. 22, 320–331. doi: 10.1016/j.cmet.2015.07.001
Sivieri, K., Morales, M. L. V., Saad, S. M. I., Adorno, M. A. T., Sakamoto, I. K., and Rossi, E. A. (2014). Prebiotic effect of Fructooligosaccharide in the Simulator of the Human Intestinal Microbial Ecosystem (SHIME (R) Model). J. Med. Food 17, 894–901. doi: 10.1089/jmf.2013.0092
Souza, C. B. D., Roeselers, G., Troost, F., Jonkers, D., Koenen, M. E., and Venema, K. (2014). Prebiotic effects of cassava bagasse in TNO's in vitro model of the colon in lean versus obese microbiota. J. Funct. Foods 11, 210–220. doi: 10.1016/j.jff.2014.09.019
Valdés, L., Cuervo, A., Salazar, N., Ruas-Madiedo, P., Gueimonde, M., and González, S. (2015). The relationship between phenolic compounds from diet and microbiota: impact on human health. Food Funct. 6, 2424–2439. doi: 10.1039/c5fo00322a
Van Den Abbeele, P., Belzer, C., Goossens, M., Kleerebezem, M., De Vos, W. M., Thas, O., et al. (2013a). Butyrate-producing Clostridium cluster XIVa species specifically colonize mucins in an in vitro gut model. ISME J. 7, 949–961. doi: 10.1038/ismej.2012.158
Van Den Abbeele, P., Grootaert, C., Marzorati, M., Possemiers, S., Verstraete, W., Gerard, P., et al. (2010). Microbial community development in a dynamic gut model is reproducible, colon region specific, and selective for Bacteroidetes and Clostridium cluster IX. Appl. Environ. Microbiol. 76, 5237–5246. doi: 10.1128/AEM.00759-10
Van Den Abbeele, P., Venema, K., Van De Wiele, T., Verstraete, W., and Possemiers, S. (2013b). Different human gut models reveal the distinct fermentation patterns of Arabinoxylan versus inulin. J. Agric. Food Chem. 61, 9819–9827. doi: 10.1021/jf4021784
Van Zanten, G. C., Knudsen, A., Roytio, H., Forssten, S., Lawther, M., Blennow, A., et al. (2012). The effect of selected synbiotics on microbial composition and short-chain fatty acid production in a model system of the human colon. PLoS ONE 7:e47212. doi: 10.1371/journal.pone.0047212
Veiga, P., Pons, N., Agrawal, A., Oozeer, R., Guyonnet, D., Brazeilles, R., et al. (2014). Changes of the human gut microbiome induced by a fermented milk product. Sci. Rep. 4:6328. doi: 10.1038/srep06328
West, N. P., Christophersen, C. T., Pyne, D. B., Cripps, A. W., Conlon, M. A., Topping, D. L., et al. (2013). Butyrylated starch increases colonic butyrate concentration but has limited effects on immunity in healthy physically active individuals. Exerc. Immunol. Rev. 19, 102–119.
Wrzosek, L., Miquel, S., Noordine, M. L., Bouet, S., Joncquel Chevalier-Curt, M., Robert, V., et al. (2013). Bacteroides thetaiotaomicron and Faecalibacterium prausnitzii influence the production of mucus glycans and the development of goblet cells in the colonic epithelium of a gnotobiotic model rodent. BMC Biol. 11:61. doi: 10.1186/1741-7007-11-61
Wu, G. D., Chen, J., Hoffmann, C., Bittinger, K., Chen, Y. Y., Keilbaugh, S. A., et al. (2011). Linking long-term dietary patterns with gut microbial enterotypes. Science 334, 105–108. doi: 10.1126/science.1208344
Yang, J., Keshavarzian, A., and Rose, D. J. (2013). Impact of dietary fiber fermentation from cereal grains on metabolite production by the fecal microbiota from normal weight and obese individuals. J. Med. Food 16, 862–867. doi: 10.1089/jmf.2012.0292
Yang, J., and Rose, D. J. (2014). Long-term dietary pattern of fecal donor correlates with butyrate production and markers of protein fermentation during in vitro fecal fermentation. Nutr. Res. 34, 749–759. doi: 10.1016/j.nutres.2014.08.006
Zhang, Y., Zhou, L., Bao, Y. L., Wu, Y., Yu, C. L., Huang, Y. X., et al. (2010). Butyrate induces cell apoptosis through activation of JNK MAP kinase pathway in human colon cancer RKO cells. Chem. Biol. Interact. 185, 174–181. doi: 10.1016/j.cbi.2010.03.035
Keywords: short chain fatty acids, diet, human health, intestinal microbiota, cross feeding
Citation: Ríos-Covián D, Ruas-Madiedo P, Margolles A, Gueimonde M, de los Reyes-Gavilán CG and Salazar N (2016) Intestinal Short Chain Fatty Acids and their Link with Diet and Human Health. Front. Microbiol. 7:185. doi: 10.3389/fmicb.2016.00185
Received: 04 December 2015; Accepted: 02 February 2016;
Published: 17 February 2016.
Edited by:
Ivan Mijakovic, Chalmers University of Technology, SwedenReviewed by:
Biswarup Sen, Amity University Haryana, IndiaCopyright © 2016 Ríos-Covián, Ruas-Madiedo, Margolles, Gueimonde, de los Reyes-Gavilán and Salazar. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nuria Salazar, bnVyaWFzZ0BpcGxhLmNzaWMuZXM=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.