
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
PERSPECTIVE article
Front. Microbiol. , 06 October 2014
Sec. Microbial Immunology
Volume 5 - 2014 | https://doi.org/10.3389/fmicb.2014.00502
This article is part of the Research Topic Why vaccines to HIV, HCV and Malaria have so far failed - challenges to developing vaccines against immunoregulating pathogens View all 14 articles
 Leonard Moise1,2Frances Terry1Andres H. Gutierrez2Ryan Tassone2Phyllis Losikoff3Stephen H. Gregory3Chris Bailey-Kellogg4William D. Martin1Anne S. De Groot1,2*
Leonard Moise1,2Frances Terry1Andres H. Gutierrez2Ryan Tassone2Phyllis Losikoff3Stephen H. Gregory3Chris Bailey-Kellogg4William D. Martin1Anne S. De Groot1,2*Despite years of research, vaccines against HIV and HCV are not yet available, due largely to effective viral immunoevasive mechanisms. A novel escape mechanism observed in viruses that cause chronic infection is suppression of viral-specific effector CD4+ and CD8+ T cells by stimulating regulatory T cells (Tregs) educated on host sequences during tolerance induction. Viral class II MHC epitopes that share a T cell receptor (TCR)-face with host epitopes may activate Tregs capable of suppressing protective responses. We designed an immunoinformatic algorithm, JanusMatrix, to identify such epitopes and discovered that among human-host viruses, chronic viruses appear more human-like than viruses that cause acute infection. Furthermore, an HCV epitope that activates Tregs in chronically infected patients, but not clearers, shares a TCR-face with numerous human sequences. To boost weak CD4+ T cell responses associated with persistent infection, vaccines for HIV and HCV must circumvent potential Treg activation that can handicap efficacy. Epitope-driven approaches to vaccine design that involve careful consideration of the T cell subsets primed during immunization will advance HIV and HCV vaccine development.
Despite modern advances in preventing disease by vaccination, persistent viral infections continue to pose a major challenge to vaccine development. The most prominent examples are HIV-1 and HCV infections, which remain two of the largest global public health challenges today. Highly effective medications are now available, but these are still inaccessible to the majority of at-risk individuals mainly due to their cost, and limitations on access to healthcare in the developing world countries where HIV and HCV are most prevalent. Unfortunately, no vaccine candidate against AIDS or hepatitis C is currently nearing market approval. Only a handful of HIV vaccine efficacy trials have been completed, and none have yet been completed for HCV (Honegger et al., 2014). While a range of strategies to treat infection and prevent transmission have been studied and implemented, it is widely believed that an effective vaccine for these global health threats is essential to stopping new infections worldwide.
Development of effective HIV and HCV vaccines is lagging because traditional strategies for developing vaccines have failed to overcome the ability of the viruses to evade the human immune response. Many obstacles to vaccine development have been uncovered: (i) extensive viral genetic diversity, enabling HIV and HCV to evade humoral as well as cell-mediated immune responses (von Hahn et al., 2007; Haaland et al., 2013), (ii) lack of suitable animal models, (iii) establishment of latent reservoirs following integration into the host genome soon after infection (in the case of HIV; Perreau et al., 2013), and (iv) lack of clear correlates of protective immunity. We recently hypothesized that viruses that tend to cause chronic diseases mutate their T cell epitopes toward greater homology with the human genome. The existence of several highly homologous T cell epitopes, some of which induce regulatory T cell (Treg) responses, has been confirmed. We postulate that this is an important means by which HIV and HCV evade effective T cell responses, and that failure to account for such epitopes may have contributed to the failure of certain vaccine approaches undertaken to date. Methods for discovering HIV and HCV Treg-activating epitopes and strategies for improving HIV and HCV vaccines are described briefly here.
No matter where one stands on the subject of the correlates of immunity to HIV and HCV infection, it is generally acknowledged that strong and broadly reactive HIV- and HCV-specific CD4+ T cell responses are required for control of acute viral infections (Rosenberg et al., 1997; Gerlach et al., 1999). Early collapse of the CD4+ T cell response impairs antibody production and CD8+ T cell responses; thus, an effective vaccine needs to induce long-lived CD4+ T cells capable of sustaining these essential components of immunity (Lichterfeld et al., 2004; Schulze Zur Wiesch et al., 2012).
CD4+ T cells are activated by virus-specific epitopes presented in the context of class II MHC by antigen presenting cells. Identification of class II MHC epitopes has been an active area of research for characterization of antigen-specific HCV and HIV CD4+ T cell responses in infection and vaccination and for construction of epitope-driven vaccines (De Groot et al., 2004; El-Awady et al., 2013; Karpenko et al., 2014; Mishra et al., 2014; Takei et al., 2014). Long-standing criteria for characterizing class II MHC epitopes include allele coverage in the human population and virus coverage among circulating strains, measured by how well an epitope represents a virus-induced response in the human host. Our group has published and validated methods by which this might be accomplished (De Groot et al., 2004; Koita et al., 2006; Mishra et al., 2014).
A significant epitope property that is beginning to gain wider attention is homology with host sequences. Viral epitopes with substantial homology to self are, at best, inert because of clonal deletion in the development of central tolerance; at worst, they may activate Tregs that suppress protective inflammatory responses and thereby enable viral persistence (Rolland et al., 2007; Frankild et al., 2008; Calis et al., 2012). It is well established that HCV-induced Treg activation is associated with extended chronic infections (Losikoff et al., 2012). Several studies of chronic HCV subjects have shown increased frequencies of natural CD4+ Tregs that express high levels of CD25, produce IL-10 (Cabrera et al., 2004), TGF-β (Bolacchi et al., 2006), and FoxP3 (Li et al., 2007), and suppress IFN-γ production (Sugimoto et al., 2003) and proliferation of HCV-specific CD8+ T cells (Boettler et al., 2005; Rushbrook et al., 2005). In the case of HIV, the role of Tregs in infection is currently being debated and requires further investigation to identify Treg subsets that may be responsible for suppressing non-specific T cell activation (beneficial) and of HIV-specific effector T cell responses (detrimental; Chevalier and Weiss, 2013).
We believe that vaccines for HIV and HCV must account for potential Treg activation that can diminish efficacy, particularly when a strong CD4+ T helper immune response is required. Ideally, vaccines should be carefully designed to reduce or eliminate potential Treg-activating sequences. That is possible today with the availability of immunoinformatic tools to predict class II MHC epitopes that may stimulate Tregs.
For over 20 years and until very recently, T cell epitope-mapping algorithms have focused on the MHC-facing side of epitopes and ignored the T cell receptor (TCR) face; thus, their usefulness was limited to identifying MHC ligands. While MHC binding is necessary to stimulate a T cell response, it is not sufficient. Hence, immunoinformatic-identified MHC ligands have been screened experimentally for T cell activation to validate predictions. Because Tregs are responsive to HIV and HCV epitopes, T cell assays should be performed to ascertain which T cell subsets are activated. Indeed, Treg-activating epitopes have been discovered using overlapping peptide arrays and tetramers in HCV core, NS3, NS4, and NS5 antigens (Li et al., 2007, 2009; Ebinuma et al., 2008; Langhans et al., 2010) and HIV Gag (Angin et al., 2012). These approaches are extremely cumbersome, however, when screens are conducted on a genomic scale with the intent to broadly cover human MHC diversity.
An informatic tool that rapidly screens thousands of candidate epitopes could address the problem of viral immune escape, but also needs to consider the vast variability and degeneracy of TCRs, making prediction extremely challenging. Fortunately, the problem can be significantly reduced by searching directly for virus-encoded human homologs that potentially stimulate natural and inducible Tregs, even if some Treg-activating epitopes are not necessarily human homologs. Specifically, pathogen sequences that bind MHC and share the same TCR-face with human MHC ligands may stimulate pre-existing Tregs that emerged from development of central and peripheral tolerance. Shared sequence patterns on the TCR-face are easily searchable.
To better define and rapidly assess viral camouflage epitopes, i.e., those epitopes homologous to human, we developed the JanusMatrix algorithm, which leverages our existing algorithm (EpiMatrix) to define MHC-binding peptide epitopes while searching for cross-conservation at the TCR-face. JanusMatrix can be applied to any viral or bacterial target protein to compare its TCR-faces to others in any genomes of interest. JanusMatrix analyzes the two faces of peptide sequences of pathogen origin for T cell activation potential (Moise et al., 2013). MHC-facing residues are analyzed for MHC binding potential using the EpiMatrix epitope-mapping algorithm (De Groot et al., 1997). We have examined TCR-facing residues for conservation against a variety of sequence databases, including the complete human proteome, the human microbiome, and human pathogens (Moise et al., 2013). For example, we screened a wide range of human-host viruses for TCR-face similarity to self and discovered that chronic viruses generally appear more human-like than viruses that cause acute infection (He et al., 2014), and that H7N9 influenza may evade immune response in a similar way (De Groot et al., 2013, 2014).
Using JanusMatrix, we discovered a promiscuous class II epitope located within non-structural HCV protein p7 that exhibits homology with hundreds of human sequences (Figure 1). The epitope induces an increase in CD4+CD25+FoxP3+ Treg number and function in peripheral blood leukocyte cultures derived from an HLA-diverse cohort of HCV-infected patients, but not in cultures derived from patients who spontaneously cleared HCV or from non-infected individuals (Losikoff et al., 2014). A human analog of the HCV epitope stimulates Tregs in both HCV-infected and non-infected people, suggesting that tolerance to HCV is promoted by activating Tregs that recognize a common TCR-face. It is well known that HCV and HIV CD8+ and CD4+ T cell epitopes mutate over the course of infection, decreasing MHC binding as a mechanism of viral escape (Harcourt et al., 1998; Norris et al., 2006; Petrovic et al., 2012). The p7 HCV epitope exhibits a novel escape mechanism, evolving a TCR-face similar to that found in autologous T cell epitopes, thus stimulating Treg responses and suppressing immune clearance.
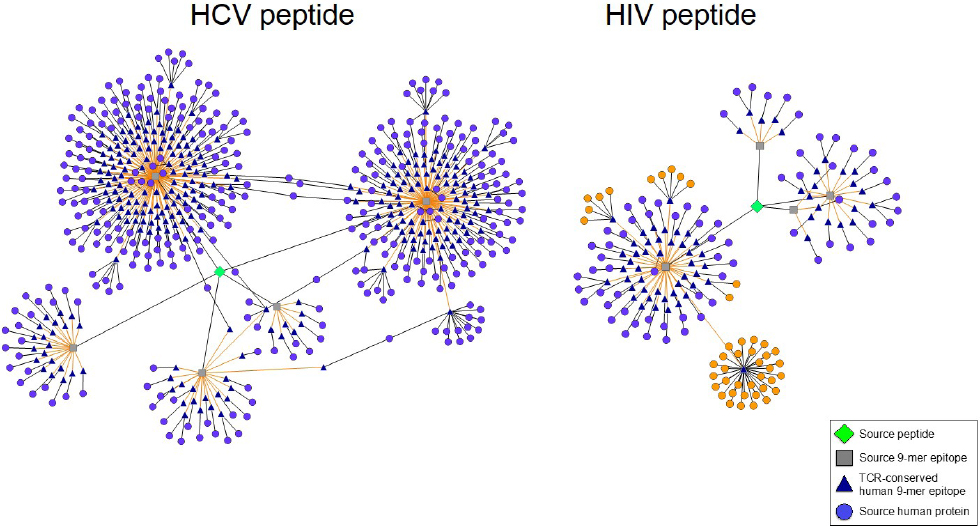
Figure 1. Predicted Treg-activating HIV and HCV sequences possess TCR faces shared by numerous human proteins. Epitope networks are shown, illustrating the abundance of TCR faces one HCV and one HIV peptide share with the human genome as determined by JanusMatrix analysis. The HIV and HCV source peptides are represented by green diamonds, their constituent 9-mer epitopes by gray squares, their cross-conserved partners in the human genome by blue triangles, and the source human proteins by light purple circles. In the HIV peptide (right) a single cross-conserved epitope can be found in 32 different HLA class I alleles; several additional 9-mer epitopes are cross-conserved with 12 other HLA sequences (source protein orange highlight).
At first glimpse, JanusMatrix analysis indicates that HIV also exhibits curious patterns of potential T cell cross-reactivity. While searching HIV envelope sequences for TCR-face conservation in the human genome, we recently uncovered a high frequency of human MHC (HLA) molecule sequences that share a TCR-face with a highly conserved epitope located in the HIV envelope protein (orange circles, Figure 1). Because HLA is highly variable in the human population, conservation of this sequence across HLA subtypes is noteworthy. The phenotype of T cells responding to this epitope has not yet been evaluated in our laboratory but an Immune Epitope Database search identified a closely related epitope capable of stimulating CD4+ T cell recognition and proliferation (Atassi and Atassi, 1992). If these CD4+ T cells are T-effector in nature, their activation could restrict HIV expansion (Sanjuán et al., 2013). We believe that it is more likely, however, that T cells that recognize this epitope possess a Treg phenotype, which may promote HIV expansion and/or persistence instead.
These HIV and HCV epitope examples are consistent with our previously published observation that “hit-and-stay” viruses escape protective immune responses by stimulating cross-reactive Tregs (He et al., 2014). This suggests that Treg-activating HCV and HIV sequences may affect HCV and HIV vaccine efficacy. We find similar patterns of cross reactivity in EBV, CMV, and HSV, all viruses that establish chronic infection and for which no vaccine exists.
Homology with the human genome represents a novel means by which viruses that seek to establish chronic infections escape human immunity and ensure their survival. Better classification of viral epitopes as either effector-or Treg-activating will improve the design of HIV and HCV vaccines. Knowledge of which epitopes to include or exclude makes it possible to generate virus-specific T cell responses that are essential for protection and that sidestep suppression. One potential solution to the challenge of HCV and HIV vaccine design is to develop epitope-driven subunit vaccines, either as whole antigen protein vaccines, using a structure-based approach, or alternatively, as platform-neutral epitope-based vaccines that do not contain Treg-activating epitopes. Such vaccines would have major advantages over conventional, but as yet unsuccessful approaches because they would simultaneously account for viral and human diversity for the purpose of broad reactivity and promote protective virus-specific T cell responses.
Importantly, the impact of Treg-activating epitopes may have different inhibitory effects depending on the level of viral replication and immune activation in acute or chronic HIV infection. Thus, Treg epitopes to exclude may differ for prophylactic and therapeutic HIV vaccines. With respect to HCV, we believe the maximum effectiveness of an HCV vaccine used either prophylac-tically or therapeutically would be achieved by always excluding Treg-activating epitopes because (i) unlike HIV, which infects and replicates in Tregs, HCV is primarily hepatotropic and (ii) a better understanding of Treg function in chronic HCV infection could lead to treatments that are capable of balancing the competing needs for sustained effector T cell-mediated immunity and limited tissue damage moderated by Tregs (Self et al., 2013).
Finally, we are exploring means to fine-tune the epitope content of HIV and HCV vaccines to induce nuanced T cell responses associated with protection. We believe that careful design is needed to improve efficacy. Cross-reactivity at the individual level owing to HLA and HIV or HCV sequence variation may necessitate the development of personalized vaccines that contain T effector, but not Treg epitopes. While personalized vaccines may seem futuristic, tools are available to design such vaccines. As this technology becomes ever more accessible, there will be an even greater incentive to define the means of personalizing vaccines.
Several of the coauthors on this manuscript are employees of EpiVax (Anne S. De Groot, Frances Terry, Leonard Moise, William D. Martin). Anne S. De Groot and William D. Martin are majority stockholders and Leonard Moise holds stock options. These authors recognize the presence of a potential conflict of interest and affirm that the information represented in this paper is original and unbiased observations.
Angin, M., King, M., Altfeld, M., Walker, B. D., Wucherpfennig, K. W., and Addo, M. M. (2012). Identification of HIV-1-specific regulatory T-cells using HLA class II tetramers. AIDS 26, 2112–2115. doi: 10.1097/QAD.0b013e328358cc75
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Atassi, H., and Atassi, M. Z. (1992). HIV envelope protein is recognized as an alloantigen by human DR-specific alloreactive T cells. Hum. Immunol. 34, 31–38. doi: 10.1016/0198-8859(92)90082-X
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Boettler, T., Spangenberg, H. C., Neumann-Haefelin, C., Panther, E., Urbani, S., Ferrari, C., et al. (2005). T cells with a CD4+CD25+ regulatory phenotype suppress in vitro proliferation of virus-specific CD8+ T cells during chronic hepatitis C virus infection. J. Virol. 79, 7860–7867. doi: 10.1128/JVI.79.12.7860-7867.2005
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Bolacchi, F., Sinistro, A., Ciaprini, C., Demin, F., Capozzi, M., Carducci, F. C., et al. (2006). Increased hepatitis C virus (HCV)-specific CD4+CD25+ regulatory T lymphocytes and reduced HCV-specific CD4+ T cell response in HCV-infected patients with normal versus abnormal alanine aminotransferase levels. Clin. Exp. Immunol. 144, 188–196. doi: 10.1111/j.1365-2249.2006.03048.x
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Cabrera, R., Tu, Z., Xu, Y., Firpi, R. J., Rosen, H. R., Liu, C., et al. (2004). An immunomodulatory role for CD4(+)CD25(+) regulatory T lymphocytes in hepatitis C virus infection. Hepatology 40, 1062–1071. doi: 10.1002/hep.20454
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Calis, J. J., de Boer, R. J., and Keşmir, C. (2012). Degenerate T-cell recognition of peptides on MHC molecules creates large holes in the T-cell repertoire. PLoS Comput. Biol. 8:e1002412. doi: 10.1371/journal.pcbi.1002412
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Chevalier, M. F., and Weiss, L. (2013). The split personality of regulatory T cells in HIV infection. Blood 121, 29–37. doi: 10.1182/blood-2012-07-409755
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
De Groot, A. S., Ardito, M., Terry, F., Levitz, L., Ross, T., Moise, L., et al. (2013). Low immunogenicity predicted for emerging avian-origin H7N9: implication for influenza vaccine design. Hum. Vaccin. Immunother. 9, 950–956. doi: 10.4161/hv.24939
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
De Groot, A. S., Bishop, E. A., Khan, B., Lally, M., Marcon, L., Franco, J., et al. (2004). Engineering immunogenic consensus T helper epitopes for a cross-clade HIV vaccine. Methods 34, 476–487. doi: 10.1016/j.ymeth.2004.06.003
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
De Groot, A. S., Jesdale, B. M., Szu, E., Schafer, J. R., Chicz, R. M., and Deocampo, G. (1997). An interactive Web site providing major histocompatibility ligand predictions: application to HIV research. AIDS Res. Hum. Retroviruses 13, 529–531. doi: 10.1089/aid.1997.13.529
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
De Groot, A. S., Moise, L., Liu, R., Gutierrez, A. H., Terry, F., Koita, O. A., et al. (2014). Cross-conservation of T-cell epitopes: now even more relevant to (H7N9) influenza vaccine design. Hum. Vaccin. Immunother. 10, 256–262. doi: 10.4161/hv.28135
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Ebinuma, H., Nakamoto, N., Li, Y., Price, D. A., Gostick, E., Levine, B. L., et al. (2008). Identification and in vitro expansion of functional antigen-specific CD25+ FoxP3+ regulatory T cells in hepatitis C virus infection. J. Virol. 82, 5043–5053. doi: 10.1128/JVI.01548-07
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
El-Awady, M. K., El Gendy, M., Waked, I., Tabll, A. A., El Abd, Y., Bader El Din, N., et al. (2013). Immunogenicity and safety of HCV E1E2 peptide vaccine in chronically HCV-infected patients who did not respond to interferon based therapy. Vaccine. doi: 10.1016/j.vaccine.2013.07.074 [Epub ahead of print].
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Frankild, S., de Boer, R. J., Lund, O., Nielsen, M., and Kesmir, C. (2008). Amino acid similarity accounts for T cell cross-reactivity and for “holes” in the T cell repertoire. PLoS ONE 3:e1831. doi: 10.1371/journal.pone.0001831
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Gerlach, J. T., Diepolder, H. M., Jung, M. C., Gruener, N. H., Schraut, W. W., Zachoval, R., et al. (1999). Recurrence of hepatitis C virus after loss of virus-specific CD4(+) T-cell response in acute hepatitis C. Gastroenterology 117, 933–941. doi: 10.1016/S0016-5085(99)70353-7
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Haaland, R. E., Johnson, J. A., and Tang, J. (2013). Recent advances in research of HIV infection: implications of viral and host genetics on treatment and prevention. Public Health Genomics 16, 31–36. doi: 10.1159/000345935
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Harcourt, G. C., Garrard, S., Davenport, M. P., Edwards, A., and Phillips, R. E. (1998). HIV-1 variation diminishes CD4 T lymphocyte recognition. J. Exp. Med. 188, 1785–1793. doi: 10.1084/jem.188.10.1785
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
He, L., De Groot, A. S., Gutierrez, A. H., Martin, W. D., Moise, L., and Bailey-Kellogg, C. (2014). Integrated assessment of predicted MHC binding and cross-conservation with self reveals patterns of viral camouflage. BMC Bioinformatics 15:S1. doi: 10.1186/1471-2105-15-S4-S1
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Honegger, J. R., Zhou, Y., and Walker, C. M. (2014). Will there be a vaccine to prevent HCV infection? Semin. Liver Dis. 34, 79–88. doi: 10.1055/s-0034-1371081
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Karpenko, L. I., Bazhan, S. I., Antonets, D. V., and Belyakov, I. M. (2014). Novel approaches in polyepitope T-cell vaccine development against HIV-1. Expert Rev. Vaccines 13, 155–173. doi: 10.1586/14760584.2014.861748
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Koita, O. A., Dabitao, D., Mahamadou, I., Tall, M., Dao, S., Tounkara, A., et al. (2006). Confirmation of immunogenic consensus sequence HIV-1 T-cell epitopes in Bamako, Mali and Providence, Rhode Island. Hum. Vaccin. 2, 119–128. doi: 10.4161/hv.2869
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Langhans, B., Braunschweiger, I., Arndt, S., Schulte, W., Satoguina, J., Layland, L. E., et al. (2010). Core-specific adaptive regulatory T-cells in different outcomes of hepatitis C. Clin. Sci. (Lond.) 119, 97–109. doi: 10.1042/CS20090661
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Li, S., Floess, S., Hamann, A., Gaudieri, S., Lucas, A., Hellard, M., et al. (2009). Analysis of FOXP3+ regulatory T cells that display apparent viral antigen specificity during chronic hepatitis C virus infection. PLoS Pathog. 5:e1000707. doi: 10.1371/journal.ppat.1000707
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Li, S., Jones, K. L., Woollard, D. J., Dromey, J., Paukovics, G., Plebanski, M., et al. (2007). Defining target antigens for CD25+ FOXP3+ IFN-gamma-regulatory T cells in chronic hepatitis C virus infection. Immunol. Cell Biol. 85, 197–204. doi: 10.1038/sj.icb.7100020
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Lichterfeld, M., Kaufmann, D. E., Yu, X. G., Mui, S. K., Addo, M. M., Johnston, M. N., et al. (2004). Loss of HIV-1-specific CD8+ T cell proliferation after acute HIV-1 infection and restoration by vaccine-induced HIV-1-specific CD4+ T cells. J. Exp. Med. 200, 701–712. doi: 10.1084/jem.20041270
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Losikoff, P. T., Mishra, S., Terry, F., Gutierrez, A., Ardito, M., et al. (2014). HCV epitope, homologous to multiple human protein sequences, induces a regulatory T cell response in infected patients. J. Hepatol. doi: 10.1016/j.jhep.2014.08.026 [Epub ahead of print].
Losikoff, P. T., Self, A. A., and Gregory, S. H. (2012). Dendritic cells, regulatory T cells and the pathogenesis of chronic hepatitis C. Virulence 3, 610–620. doi: 10.4161/viru.21823
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Mishra, S., Losikoff, P. T., Self, A. A., Terry, F., Ardito, M. T., Tassone, R., et al. (2014). Peptide-pulsed dendritic cells induce the hepatitis C viral epitope-specific responses of naïve human T cells. Vaccine 32, 3285–3292. doi: 10.1016/j.vaccine.2014.03.083
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Moise, L., Gutierrez, A. H., Bailey-Kellogg, C., Terry, F., Leng, Q., Abdel Hady, K. M., et al. (2013). The two-faced T cell epitope: examining the host-microbe interface with JanusMatrix. Hum. Vaccin. Immunother. 9, 1577–1586. doi: 10.4161/hv.24615
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Norris, P. J., Stone, J. D., Anikeeva, N., Heitman, J. W., Wilson, I. C., Hirschkorn, D. F., et al. (2006). Antagonism of HIV-specific CD4+ T cells by C-terminal truncation of a minimum epitope. Mol. Immunol. 43, 1349–1357. doi: 10.1016/j.molimm.2005.09.004
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Perreau, M., Levy, Y., and Pantaleo, G. (2013). Immune response to HIV. Curr. Opin. HIV AIDS 8, 333–340. doi: 10.1097/COH.0b013e328361faf4
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Petrovic, D., Dempsey, E., Doherty, D. G., Kelleher, D., and Long, A. (2012). Hepatitis C virus-T-cell responses and viral escape mutations. Eur. J. Immunol. 42, 17–26. doi: 10.1002/eji.201141593
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Rolland, M., Nickle, D. C., Deng, W., Frahm, N., Brander, C., Learn, G. H., et al. (2007). Recognition of HIV-1 peptides by host CTL is related to HIV-1 similarity to human proteins. PLoS ONE 2:e823. doi: 10.1371/journal.pone. 0000823
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Rosenberg, E. S., Billingsley, J. M., Caliendo, A. M., Boswell, S. L., Sax, P. E., Kalams, S. A., et al. (1997). Vigorous HIV-1-specific CD4+ T cell responses associated with control of viremia. Science 278, 1447–1450. doi: 10.1126/science.278.5342. 1447
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Rushbrook, S. M., Ward, S. M., Unitt, E., Vowler, S. L., Lucas, M., Klenerman, P., et al. (2005). Regulatory T cells suppress in vitro proliferation of virus-specific CD8+ T cells during persistent hepatitis C virus infection. J. Virol. 79, 7852–7859. doi: 10.1128/JVI.79.12.7852-7859.2005
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Sanjuán, R., Nebot, M. R., Peris, J. B., and Alcamí, J. (2013). Immune activation promotes evolutionary conservation of T-cell epitopes in HIV-1. PLoS Biol. 11:e1001523. doi: 10.1371/journal.pbio.1001523
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Schulze Zur Wiesch, J., Ciuffreda, D., Lewis-Ximenez, L., Kasprowicz, V., Nolan, B. E., Streeck, H., et al. (2012). Broadly directed virus-specific CD4+ T cell responses are primed during acute hepatitis C infection, but rapidly disappear from human blood with viral persistence. J. Exp. Med. 209, 61–75. doi: 10.1084/jem.20100388
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Self, A. A., Losikoff, P. T., and Gregory, S. H. (2013). Divergent contributions of regulatory T cells to the pathogenesis of chronic hepatitis C. Hum. Vaccin. Immunother. 9, 1569–1576. doi: 10.4161/hv.24726
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Sugimoto, K., Ikeda, F., Stadanlick, J., Nunes, F. A., Alter, H. J., and Chang, K. M. (2003). Suppression of HCV-specific T cells without differential hierarchy demonstrated ex vivo in persistent HCV infection. Hepatology 38, 1437–1448. doi: 10.1016/j.hep.2003.09.026
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Takei, S., Omoto, C., Kitagawa, K., Morishita, N., Katayama, T., Shigemura, K., et al. (2014). Oral administration of genetically modified Bifidobacterium displaying HCV-NS3 multi-epitope fusion protein could induce an HCV-NS3-specific systemic immune response in mice. Vaccine 32, 3066–3074. doi: 10.1016/j.vaccine.2014.03.022
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
von Hahn, T., Yoon, J. C., Alter, H., Rice, C. M., Rehermann, B., Balfe, P., et al. (2007). Hepatitis C virus continuously escapes from neutralizing antibody and T-cell responses during chronic infection in vivo. Gastroenterology 132, 667–678. doi: 10.1053/j.gastro.2006.12.008
Pubmed Abstract | Pubmed Full Text | CrossRef Full Text | Google Scholar
Keywords: HIV, HCV, T cell epitope, immunoinformatics, vaccines, cross-reactivity, regulatory T cells, MHC class II
Citation: Moise L, Terry F, Gutierrez AH, Tassone R, Losikoff P, Gregory SH, Bailey-Kellogg C, Martin WD and De Groot AS (2014) Smarter vaccine design will circumvent regulatory T cell-mediated evasion in chronic HIV and HCV infection. Front. Microbiol. 5:502. doi: 10.3389/fmicb.2014.00502
Received: 02 July 2014; Accepted: 08 September 2014;
Published online: 06 October 2014.
Edited by:
Shuo Li, Burnet Institute and Monash University, AustraliaReviewed by:
Katie L. Flanagan, Monash University, AustraliaCopyright © 2014 Moise, Terry, Gutierrez, Tassone, Losikoff, Gregory, Bailey-Kellogg, Martin and De Groot. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Anne S. De Groot, EpiVax, Inc., 146 Clifford Street, Providence, RI 02903, USA e-mail:ZHIuYW5uaWUuZGVncm9vdEBnbWFpbC5jb20=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.