 Alain Hartmann1,2*
Alain Hartmann1,2* Aude Locatelli1
Aude Locatelli1 Lucie Amoureux3 Géraldine Depret1,2
Lucie Amoureux3 Géraldine Depret1,2 Claudy Jolivet4 Eric Gueneau5
Claudy Jolivet4 Eric Gueneau5 Catherine Neuwirth3
Catherine Neuwirth3
- 1 INRA, UMR Microbiologie des Sols et de l’Environnement (MSE), The Institut National de la Recherche Agronomique, Dijon, France
- 2 Université de Bourgogne, Dijon, France
- 3 Laboratory of Bacteriology, University Hospital of Dijon, Dijon, France
- 4 Unité InfoSol, The Institut National de la Recherche Agronomique, Orléans, France
- 5 Laboratoire Départemental de la Côte d’Or, Dijon, France
CTX-M [a major type of extended-spectrum beta-lactamase (ESBL)] producing Escherichia coli are increasingly involved in human infections worldwide. The aim of this study was to investigate potential reservoirs for such strains: soils, cattle, and farm environment. The prevalence of blaCTX-M genes was determined directly from soil DNA extracts obtained from 120 sites in Burgundy (France) using real-time PCR. blaCTX-M targets were found in 20% of the DNA extracts tested. Samples of cattle feces (n = 271) were collected from 182 farms in Burgundy. Thirteen ESBL-producing isolates were obtained from 12 farms and further characterized for the presence of bla genes. Of the 13 strains, five and eight strains carried blaTEM-71 genes and blaCTX-M-1 genes respectively. Ten strains of CTX-M-1 producing E. coli were isolated from cultivated and pasture soils as well as from composted manure within two of these farms. The genotypic analysis revealed that environmental and animal strains were clonally related. Our study confirms the occurrence of CTX-M producing E. coli in cattle and reports for the first time the occurrence of such strains in cultivated soils. The environmental competence of such strains has to be determined and might explain their long term survival since CTX-M isolates were recovered from a soil that was last amended with manure 1 year before sampling.
Introduction
The production of extended-spectrum beta-lactamases (ESBLs) is one of the most significant mechanisms of resistance to oxyiminocephalosporin antibiotics in E. coli (Pitout and Laupland, 2008). Among these enzymes, the CTX-M type ESBLs have emerged worldwide, they have progressively replaced the TEM and SHV families (Bonnet, 2004). To date, 123 blaCTX-M genes have been reported1, the corresponding CTX-M enzymes are clustered in five groups, group 1, 9, and 2 being predominant. The producing organisms are sometimes involved in nosocomial infections but are widely encountered in community settings (Arpin et al., 2009; Woerther et al., 2010). Their rate of dissemination might suggest the occurrence of environmental reservoirs potentially leading to human contamination through water, food consumption or direct contact with animals (Leverstein-Van Hall et al., 2011). There are many descriptions of fecal carriage of such organisms among food-producing animals especially poultry (broilers) and pigs (Costa et al., 2009; Bortolaia et al., 2010; Cortes et al., 2010). The reports concerning livestock cattle are much less abundant (Horton et al., 2011), and there is no published data about the prevalence of CTX-M producing E. coli in soils. Nevertheless cultivated soils are frequently fertilized with agricultural or urban organic residues that may contain antibiotic resistant microorganisms (Moodley and Guardabassi, 2009; Reinthaler et al., 2010) and thus might act as environmental reservoirs.
The aim of this study was to develop an integrated approach encompassing soils, livestock, and farm environment in a whole region (Burgundy, France). A systematic large scale study was conducted using a molecular detection approach to detect blaCTX-M directly from soil DNA extracts. Conventional bacteriological methods were used to isolate ESBL-producing E. coli from cattle feces from 182 farms. When positive animals were detected, the corresponding farm environment, i.e., cultivated and pasture soils as well as cattle manure was sampled and analyzed by bacteriological methods. All environmental and animal isolates recovered during the study have been subjected to genotypic characterization and blaCTX-M genes have been sequenced.
Materials and Methods
Soil Sampling
Soils were sampled in the “Réseau de Mesures de la Qualité des Sols” (RMQS = French Soil Quality Monitoring Network) which is a network based on the sampling of soil with a 16 × 16 km systematic grid covering the whole French territory (Arrouays et al., 2002). The RMQS consisted in 2200 monitoring sites, which are located close to the center of each 16 × 16 km cell. Corresponding land covers were recorded and categorized as: large scale crops, pastures, orchards, vineyards, natural vegetation such as forests or meadows. Each site is geo-positioned with a precision <0.5 m. Twenty five individual core samples were collected from the topsoil (0–30 cm) using a stratified random sampling design within a 20 m × 20 m area. Core samples were bulked to obtain a composite sample for each site. Soil samples were air-dried and sieved to 2 mm before analysis. For this study 120 soil samples corresponding to the Burgundy region (four departments Côte d’Or, Saône et Loire, Yonne, and Nièvre) were analysed.
Bovine Feces Sampling
From April 2009 to June 2009, a total of 271 fecal swabs of cattle were collected from 182 farms located in three departments of the Burgundy region, namely Côte d’Or, Nièvre, and Saône et Loire. Three groups of animals were sampled: (i) healthy adults (218), (ii) enteritic diseased calves (35), and healthy adults linked with sick calves (18). Swabs (COPAN, CML, France) were immediately transferred into tubes containing Amies agar gel transport medium. Bacteriological analyses of the fecal swabs were performed within 3 days after sampling. In farm 2 where positive animals had been detected in 2009, feces samples of the whole cow herd were analyzed (90 animals) in July 2010 as described above.
Farm Environment Sampling
For three farms (farms 1, 2 and 3), where CTX-M producing E. coli occurred in animals, we conducted further analysis of several environmental samples during the autumn 2009. These farms were chosen on the basis of the willingness of the farmers to cooperate to the study. In each farm one cropped and one pasture soils were sampled by taking three individual cores in the top soil (using a 7 cm width disinfected auger) that were pooled to obtain a composite sample. Manure samples were collected in the three farms using the same equipment (disinfected auger). All samples were kept moist in single use plastic bags at room temperature. Bacteriological analyses were performed on these samples within 3 days after sampling.
Soil Microbial Community DNA Extraction
Microbial DNA was extracted from bulk RMQS soil samples according to the method described by Ranjard et al. (2003). Briefly, 1.5 g of each soil was mixed with 5 ml of a solution containing 100 mM Tris (pH 8.0), 100 mM EDTA (pH 8.0), 100 mM NaCl, and 2% (w/v) sodium dodecyl sulfate. Two grams of 106 μm diameter glass beads and eight glass beads of 2-mm diameter were added in a bead-beater tube. The samples were then homogenized for 30 s at 1,600 rpm in a mini bead-beater cell disruptor (Mikrodismembrator; S.B. Braun Biotech International) and centrifuged at 7,000 × g for 5 min at 4°C after 30 min incubation at 70°C. The collected supernatants were incubated for 10 min on ice after adding 1/10 volume of 3 M potassium acetate (pH 5.5) and centrifuged at 14,000 × g for 5 min. After precipitation with one volume of ice-cold isopropanol, the nucleic acids were washed with 70% ethanol. DNA extracts were purified on polyvinyl polypyrrolydone minicolumns (Bio-Rad, France) by centrifugation at 1,000 × g for 2 min at 10°C. Residual impurities from DNA extracts were finally removed by using a Geneclean Turbo kit as recommended by the manufacturer (Q Biogene®, Illkirch, France). DNA’s were quantified by agarose gel electrophoresis (1% agarose in TBE buffer) using calf thymus DNA dilutions as standards and the ImageQuant software (Applied Biosytems). Five nanograms of DNA were used per PCR reaction.
Development and Use of a Real-Time PCR Assay for blaCTX-M Detection from Soil Extracted DNA
Twenty seven blaCTX-M genes sequences were aligned using the ClustalW program. Sequences were chosen to be representative from the five distinct groups of CTX-M enzymes described by Bonnet (2004). Seven sequences from CTX-M-1 group were used (GenBank accession numbers: DQ658221, X92506, EU921825, NC004464, Y10278, NC005327, and AY238472) corresponding to CTX-M-12, CTX-M-1, CTX-M-32, CTX-M-3, CTX-M-15, CTX-M-33, respectively. Four sequences from CTX-M-9 group (GenBank accession numbers: AF174129, EU916273, EU921824, and EU921826) corresponding to CTX-M-9, CTX-M-27, CTX-M-24, and CTX-M-65, respectively. Three sequences from CTX-M-2 group were used (GenBank accession numbers: X92504, EF374097, and AM982521) corresponding to CTX-M-2, CTX-M-56, and CTX-M-77 respectively. Three sequences from CTX-M-25 group were used (GenBank accession numbers: AF518567, DQ023162, and AM982522) corresponding to CTX-M-25, CTX-M-41, and CTX-M-78 respectively. Two sequences from CTX-M-8 group were used (GenBank accession number: AF189721 and AY750914) corresponding to CTX-M-8 and CTX-M-40. From these alignments, primers and Taqman type probes specific for either blaCTX-M genes encoding CTX-M-1 or CTX-M-9 groups were selected using Primer Express software (Applied Biosystems). For CTX-M-1 group, primers were F469 (5′-CAGCTGGGAGACGAAACGTT-3′) and R532 (5′-CCGGAATGGCGGTGTTTA-3′) and the probe was S490 (5′ -6FAM-CGTCTCGACCGTACCGAGCCGAC-TAMRA-3′). For the CTX-M-9 group, primers were F446 (5′-GAGGCGTGACGGCTTTTG-3′) and R513 (5′-CGTAGGTTCAGTGCGATCCA-3′), and the probe was S470 (5′-6FAM-CGATCGGCGATGAGACGTTTCGT-TAMRA-3′). Duplicate real-time PCRs were run with the ABI Prism 7900 sequence detection system (Applied Biosystems, France) and Absolute QPCR ROX master mix (Thermo Scientific, France) in 25-μl reaction mixtures and under reaction conditions of 95°C for 15 min (enzyme activation), and 40 two-step cycles consisting of 95°C for 15 s and 60°C for 1 min. Primers concentrations were 400 nM and Taqman type probe concentration was 200 nM, 1-μl of T4 GP32 (MP Biomedicals, France) was added per reaction. Five microliters of template DNA was added per reaction. PCR targets were cloned in pCR II-TOPO plasmid and recombinant linearized plasmids were used as standards. Gene copy numbers were calculated by amplifying six serial dilutions of the standard (101–105 copies per reaction mixture) in parallel with the samples. The specificity of the CTX-M detection systems has been checked in silico using the BLASTN algorithm across the GenBank nucleotide database (NCBI website). The specificity of the CTX-M detection systems has also been validated by using human clinical isolates harboring known blaCTX-M genes (Table 1). In order to determine the efficiency of these detection systems in soil, soil samples (2 g) were spiked with known titers of E. coli strain EC 913 SE carrying blaCTX-M-1, soil DNA was extracted immediately as described above and subjected to real-time PCR using the primers and probe specific for CTX-M-1 group (Figure 1).
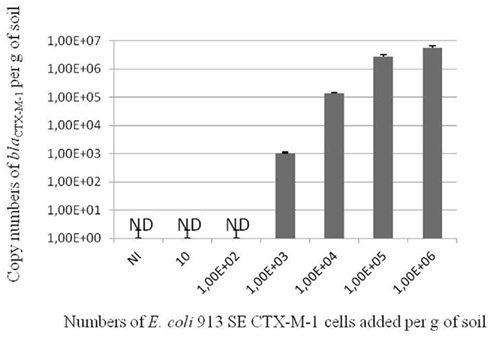
Figure 1. Detection levels of blaCTX-M-1 genes by real-time PCR (CTX-M-1 group detection system) in soil microcosms spiked with increasing amounts of E. coli strain 913SE CTX-M-1. ND, not detected. Results are the means of two replicates and bars represent standard deviations.
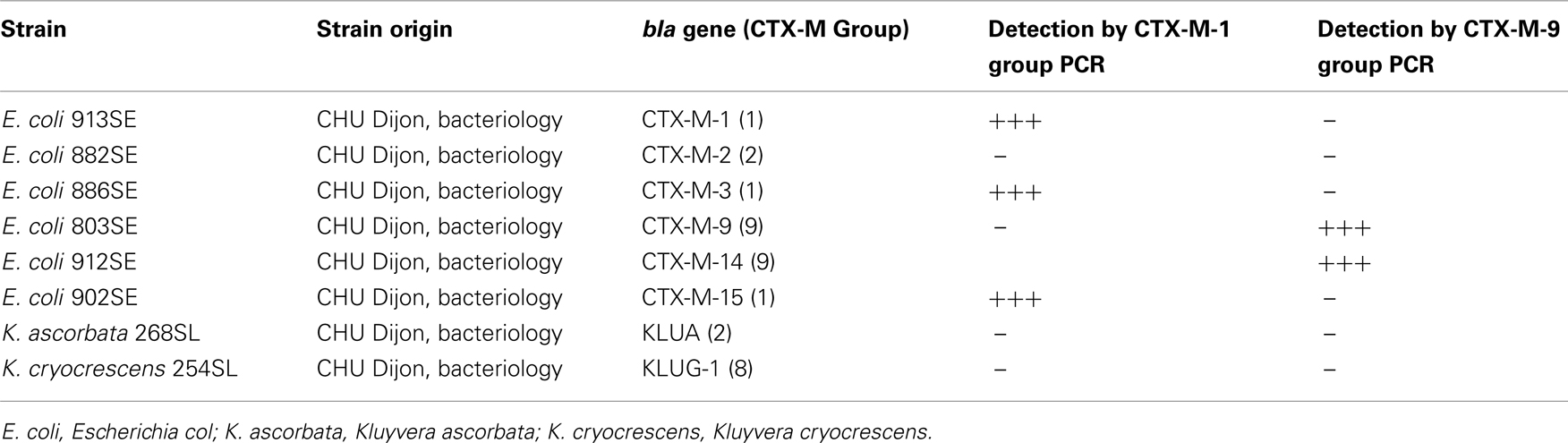
Table 1. Human clinical strains used to validate the CTX-M-1 group and CTX-M-9 group detection systems (clinical strains were collected between 2006 and 2007 at CHU Dijon, France).
Bacteriological Methods and Antibiotic Resistance Testing
All fecal samples from the cows and all environmental samples have been inoculated on ESBL screening agar plates containing Drigalsky medium supplemented with either cefotaxime (4 mg/l) or ceftazidime (4 mg/l). The antibiotic susceptibility testings have been performed by the disk diffusion method. A range of antibiotics including penicillins, cephalosporins, carbapenem (imipenem), aminoglycosides (kanamycin, tobramycin, gentamycin, streptomycin, amikacin, netilmicin), chloramphenicol, quinolones (ciprofloxacin, ofloxacin), doxycycline, cotrimoxazol, and colistin was used to determine antibiotic susceptibility patterns of the isolates and the production of ESBL was assessed by the double-disk synergy test (Jarlier et al., 1988). Guidelines for the interpretation of antibiotic susceptibility testing were from the Clinical and Laboratory Standards Institute (CLSI, 2010).
Identification of ESBL Types
The characterization of the bla gene was performed by PCR on isolates with a positive double-disk synergy test by using primers specific for the genes encoding ESBL from TEM, SHV, and CTX-M families (Chanal et al., 1992; Neuwirth et al., 1995; Sabate et al., 2002). PCR products were sequenced on both strands using ABI PRISM 3100 (Applied Biosystems, France).
Genotyping by the Diversilab System (bioMérieux)
The Diversilab system is a typing technique which is based on the repetitive-sequence-based PCR (rep-PCR) and proved to be a useful method for genotyping of E. coli (Fluit et al., 2010). All reagents, automates and software used for this study were provided by bioMérieux, France. DNA was extracted from E. coli colonies using an UltraClean Microbial DNA isolation kit and following the manufacturer’s instructions. The extracted DNA was amplified using a DiversiLab Escherichia DNA fingerprinting kit. Electrophoresis of the amplified fragments using a microfluidics LabChip with an Agilent 2100 Bioanalyzer and analysis were performed according to the protocol of the manufacturer. Isolates with a similarity of <95% were considered different, and isolates with a similarity of >98% were considered indistinguishable.
MLST Typing of E. coli Strains
MLST typing of E. coli strains was done according to recommendations found at http://mlst.ucc.ie/mlst/dbs/Ecoli/documents/primersColi_html. The E. coli MLST scheme uses internal fragments of the following seven house-keeping genes: adk (adenylate kinase), fumC (fumarate hydratase), gyrB (DNA gyrase), icd (isocitrate/isopropylmalate dehydrogenase), mdh (malate dehydrogenase), purA (adenylosuccinate dehydrogenase), recA (ATP/GTP binding motif). MLST typing was done at the Institute of Microbiology and Epizootics, Freie Universität Berlin, Berlin, Germany, by Dr. Sebastian Guenther.
Results
blaCTX-M Detection in Soils
The method of real-time PCR detection of blaCTX-M has been validated on the basis of amplification of DNA from pure strains or from inoculated soil DNA. The two detection systems proved to be sensitive and specific of blaCTX-M genes encoding group 1 and group 9 enzymes. Detection threshold was estimated to be one copy per PCR reaction. Detection limit in soil samples was estimated to be 103 copies per gram of soil (Figure 1). Out of the 120 soil extracted DNA’s from Burgundy, 22 were found to be positive using one or two detection systems (Figure 2). The proportion of positive soil for occurrence of blaCTX-M 1 or 9 was of 3.3% in Yonne, 6.2% in Côte d’Or, 16.0% in Nièvre, and 24.2% in Saône et Loire. These results demonstrate the wide distribution of some blaCTX-M genes in soils from Burgundy. No obvious correlation was found between soil physical and chemical properties and the occurrence of blaCTX-M genes [analysis by principal component analysis (PCA), data not shown]. The causes of the higher prevalence of blaCTX-M genes in soils from Saône et Loire remain to be deciphered.
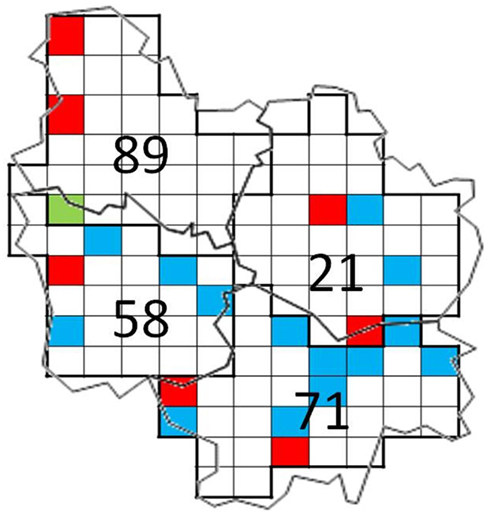
Figure 2. Distribution of RMQS soils where blaCTX-M genes were detected (colored squares) in the four departments of Burgundy (21: Côte d’Or, 58: Nièvre, 89: Yonne, and 71: Saône et Loire). Red square: detection of blaCTX-M-1, blue square: detection of blaCTX-M-9, green square: detection of both gene types.
Detection of ESBL-Producing E. coli from Livestock
Out of the 271 feces samples analyzed in 2009, 13 proved to contain ESBL-producing E. coli (Table 2). Five isolates harbored the TEM-71-ESBL. Interestingly all these isolates were originated from “Nièvre.” Four isolates were genotypically indistinguishable. Among them isolates 32 and 101 were originated from two farms located in the same village. The others (92 and 107) were recovered in animals from farms located 16 km apart. The eight remaining isolates harbored CTX-M-1. The CTX-M-1 producing isolates have been isolated in three departments (Nièvre, Saône et Loire, and Côte d’Or). The genotypical analysis performed by rep-PCR with Diversilab revealed a wide diversity among the strains isolated from different farms. However, the strains 234 and 235 originated from two animals sampled in a same farm (farm 2) were clonally related (not distinguishable).
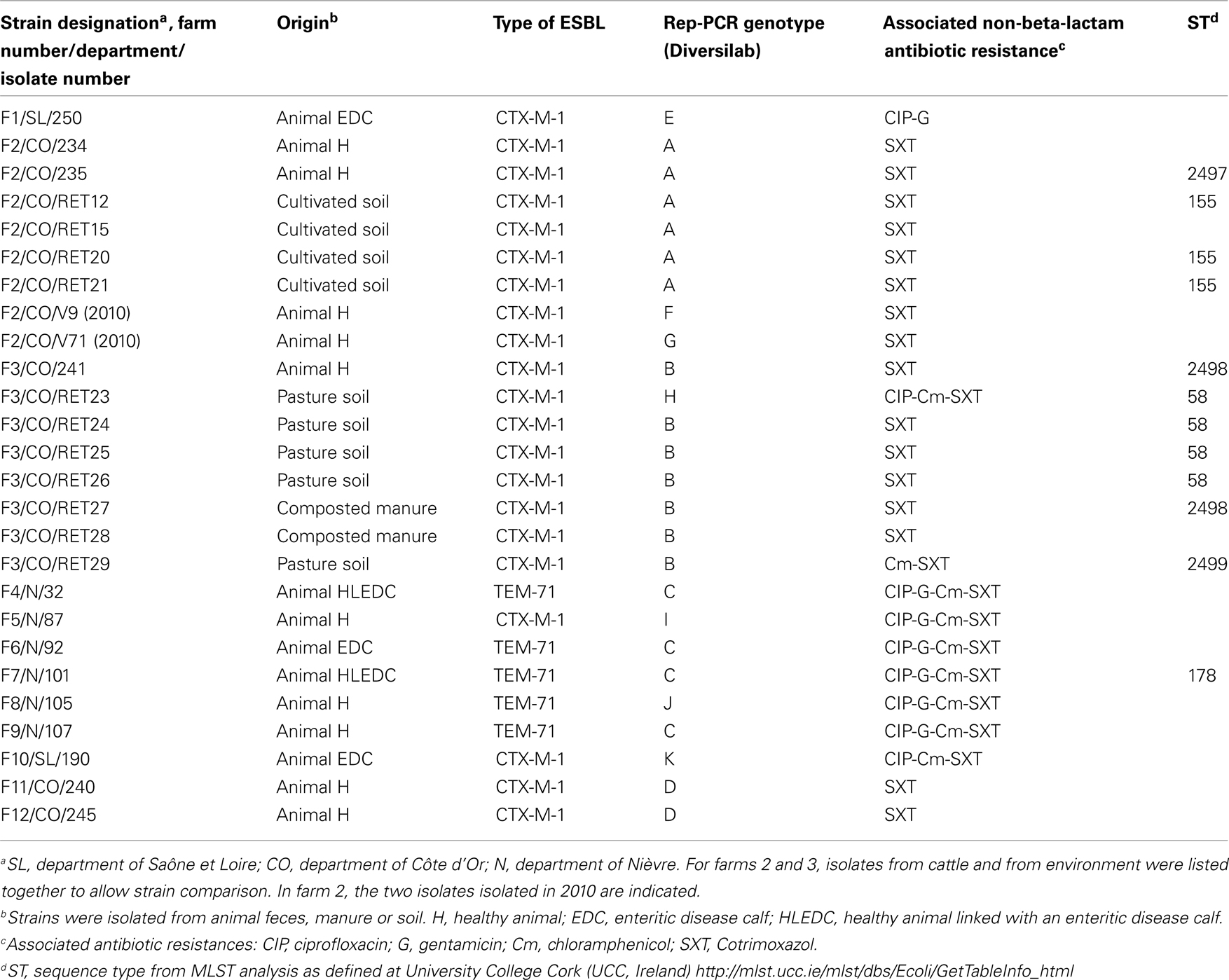
Table 2. Characteristics of the E. coli strains isolated from animals, manure and soils.
One year later (July 2010), feces samples of the whole cow herd from farm 2 were analyzed (90 animals), and CTX-M-1 producing E.coli isolates were detected from two animals. Strains V71 and V9 were genotyped and were different from the strains 234 and 235 isolated in June 2009 (Table 2).
Detection of ESBL E. coli Isolates in Environmental Samples
The environments of three farms where positive animals have been detected were chosen for further investigations. In the farm 1, no ESBL-producing E. coli were found in the farm environment. On the contrary, 4 and 6 CTX-M-1 producing E. coli were detected in farms 2 and 3 respectively (Table 2). In farm 2, a cultivated soil amended one year before with liquid cow manure from the farm was found to contain CTX-M harboring E. coli strains. These isolates were genotypically indistinguishable from the animal strains F2/CO/234 and F2/CO/235. Interestingly the soil sampling site was located 3 km away from the cattle barn. In farm 3 the positive samples were: pasture soil, and composted manure. In that case, composted manure isolate was identical to the animal isolate, whereas one soil isolate had a different genotype.
Discussion
The worldwide emergence of CTX-M producing E. coli in human clinical samples is a public health concern (Pitout and Laupland, 2008) and raises several interrogations regarding their high dissemination rate. These strains are also widely described in animals (pets, farm animals; Costa et al., 2009; Bortolaia et al., 2010; Cortes et al., 2010). It can be hypothesized that there is a cross-transmission between the human being and the animals (Leverstein-Van Hall et al., 2011), or that there are common environmental sources leading to human and animal contaminations. The treatment regimes for eradication of infections caused by such strains are sometimes very limited, i.e., when the CTX-M production is associated with the production of aminoglycosides modifying enzymes and gyrase mutations (fluoroquinolones resistance). We report in this study our findings concerning the prevalence of fecal carriage of E. coli CTX-M producing in cattle as well as the contamination of the farm environment.
Detection of ESBL-Producing E. coli in Livestock
We have detected ESBL-producing E. coli in livestock (5% of the animals tested). In Europe such strains have been only sporadically described in cattle: in Germany (Guerra et al., 2007), England (Horton et al., 2011), and Spain (Brinas et al., 2005). To our best knowledge this is the second report of ESBL-producing E. coli in cattle in France where Meunier et al. (2006) reported three isolates (carrying either CTX-M-1 or CTX-M-15 ESBL) responsible for infections in cow. Our results demonstrate that ESBL-producing E. coli strain are carried indifferently by sick and healthy animals. Our results are in agreement with previously reported data: CTX-M-1 is the ESBL which is the most frequently encountered in animals (Girlich et al., 2007; Bonnedahl et al., 2009; Bortolaia et al., 2010).
Nevertheless we have detected for the first time E. coli TEM-71-producing in five animals whereas they have been exceptionally reported in human infection (Wong-Beringer et al., 2001; Rasheed et al., 2002; De Champs et al., 2004). This finding is interesting from the epidemiological point of view because four isolates are genotypically related whereas they were originated from different farms (F4, F6, F7, and F9) This indicates that some clonal isolates are widespread or might circulate (through animal transfer) at a regional scale or that there is a common reservoir in the region. At the farm level, this might suggest a potential cross-contamination among cattle or the presence of a reservoir within the farm. Finally, this is the first description of TEM-71 producing E. coli in animals. It is noteworthy that the TEM-71 producing isolates are multiresistant to ciprofloxacin, gentamicin, chloramphenicol, and cotrimoxazol. On the opposite, most of the CTX-M-1 producing isolates harbor a single associated resistance to cotrimoxazol (Table 2). Therefore, cefotaxim resistant E. coli recovered from animals originated from different farms show significantly different antibiotic resistance patterns. This might reflect different exposition of the animals to antimicrobial agents or different sources of contamination.
In farm 2, strains isolated from cattle in 2010 have a different genotype compared to those isolated in 2009. Several hypothesis might explain this result: (i) horizontal gene transfer may occur between E. coli genotypes carried by cattle, since blaCTX-M genes are carried by plasmids, (ii) new exogenous E. coli strains may have been disseminated in the farm environment.
Detection of ESBL-Producing E. coli in Farm Environment
Among the 12 farms where ESBL-producing E. coli were detected from animals, we chose three farms to further investigate the dissemination of these strains in the farm environment. Farms were chosen on the basis of which farmers will decide to cooperate to our study.
In two of the three farms where positive animals were reported, ESBL E. coli isolates were detected in environmental samples. Interestingly, in farm 2 one of the soil sampling sites was a crop field that has been amended 1 year before (autumn 2008) with liquid manure collected in the barn. This crop field was located 3 km away from the cattle barn. In autumn 2009, we were able to isolate several CTX-M producing E. coli strains from this soil, indicating that such strains have the ability to survive at least 1 year after soil amendment under environmental conditions. In farm 3, the positive samples were: pasture soil, and composted manure. In that case, composted manure isolate was identical to the animal isolate, whereas one soil isolate had a different genotype.
Further Genotypic Comparison of E. coli Strains
MLST typing of E. coli strains isolated from this study has been partially done. Preliminary results indicate that some E. coli strains isolated from animal and soil belong to previously described ST58, ST155, and ST178 as defined in the MLST database hosted at UCC2. Four strains appear to belong to new genotypes namely ST2497, ST2498, and ST2499. blaCTX-M-1 genes were found in E. coli isolates with variable ST. ST8 and ST155 grouped strains from animal and human origin, some strains belonging to these ST are pathogenic for human or animals. Plasmids of four strains were sequenced (454 pyrosequencing), blaCTX-M genes were found on plasmids and located close to one copy of ISEcp1 in each strain (data not shown).
Molecular Detection of blaCTX-M Genes in Soils from Burgundy
The origin of the blaCTX-M genes detected in soils remains to be elucidated, several hypotheses might be investigated. E. coli strains harboring blaCTX-M genes might have been disseminated in soils through manure application on cultivated soils (animal origin), sewage sludge application (human origin), or through irrigation with treated or untreated waste-water. As the progenitor blaCTX-M gene is the chromosomal bla genes of different species of Kluyvera (Decousser et al., 2001; Poirel et al., 2002) we can also hypothesize that bla genes detected in the studied soils might be harbored by bacterial species other than E. coli. These bacteria might thus act as potential environmental bla genes reservoirs. To our best knowledge, this is the first report about occurrence of blaCTX-M genes in soil.
The detection of CTX-M producing E. coli in soil was the major finding of this study. Our work demonstrated the survival of CTX-M producing E. coli in soil at least for one year. Future work will aim at linking blaCTX-M occurrence with the capacity of E. coli strains to survive under environmental conditions. Finally, potential risks for public health through water or vegetables contamination will have to be determined.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was co-funded by the Conseil Régional de Bourgogne (Regional Council of Burgundy), and by MERIAL SA. We wish to thank the Groupement Technique Vétérinaire (GTV) of Saône et Loire for animal feces sampling. Finally, we wish to thank Dr. Sebastian Guenther (Institute of Microbiology and Epizootics, Freie Universität Berlin, Berlin, Germany) for his invaluable help in MLST typing of the E. coli strains. The authors wish to thank the GenoSol Platform (particularly M. Lelièvre and S. Dequiedt) of UMR Agroecology, for supplying soil DNA extracts from Burgundy and for data-mining.
Footnotes
sReferences
Arpin, C., Quentin, C., Grobost, F., Cambau, E., Robert, J., Dubois, V., Coulange, L., and Andre, C. (2009). Nationwide survey of extended-spectrum beta-lactamase-producing Enterobacteriaceae in the French community setting. J. Antimicrob. Chemother. 63, 1205–1214.
Arrouays, D., Jolivet, C., Boulonne, L., Bodineau, G., Saby, N., and Grolleau, E. (2002). A new initiative in France: a multi-institutional soil quality monitoring network. Une initiative nouvelle en France: la mise en place d’un reseau multi-institutionnel de mesure de la qualite des sols (RMQS). C. R. Acad. Agric. Fr. 88, 93–103.
Bonnedahl, J., Drobni, M., Gauthier-Clerc, M., Hernandez, J., Granholm, S., Kayser, Y., Melhus, A., Kahlmeter, G., Waldenstrom, J., Johansson, A., and Olsen, B. (2009). Dissemination of Escherichia coli with CTX-M type ESBL between humans and yellow-legged gulls in the south of France. PLoS One 4, e5958. doi:10.1371/journal.pone.0005958
Bonnet, R. (2004). Growing group of extended-spectrum beta-lactamases: the CTX-M enzymes. Antimicrob. Agents Chemother. 48, 1–14.
Bortolaia, V., Guardabassi, L., Trevisani, M., Bisgaard, M., Venturi, L., and Bojesen, A. M. (2010). High diversity of extended-spectrum beta-lactamases in Escherichia coli isolates from Italian broiler flocks. Antimicrob. Agents Chemother. 54, 1623–1626.
Brinas, L., Moreno, M. A., Teshager, T., Saenz, Y., Porrero, M. C., Dominguez, L., and Torres, C. (2005). Monitoring and characterization of extended-spectrum beta-lactamases in Escherichia coli strains from healthy and sick animals in Spain in 2003. Antimicrob. Agents Chemother. 49, 1262–1264.
Chanal, C., Poupart, M. C., Sirot, D., Labia, R., Sirot, J., and Cluzel, R. (1992). Nucleotide sequences of CAZ-2, CAZ-6, and CAZ-7 beta-lactamase genes. Antimicrob. Agents Chemother. 36, 1817–1820.
Clinical and Laboratory Standards Institute (CLSI). (2010). “Performance standards for antimicrobial susceptibility testing,” in Twentieth Informational Supplement M100-S20 (Wayne, PA: CLSI).
Cortes, P., Blanc, V., Mora, A., Dahbi, G., Blanco, J. E., Blanco, M., Lopez, C., Andreu, A., Navarro, F., Alonso, M. P., Bou, G., Blanco, J., and Llagostera, M. (2010). Isolation and characterization of potentially pathogenic antimicrobial-resistant Escherichia coli strains from chicken and pig farms in Spain. Appl. Environ. Microbiol. 76, 2799–2805.
Costa, D., Vinue, L., Poeta, P., Coelho, A. C., Matos, M., Saenz, Y., Somalo, S., Zarazaga, M., Rodrigues, J., and Torres, C. (2009). Prevalence of extended-spectrum beta-lactamase-producing Escherichia coli isolates in faecal samples of broilers. Vet. Microbiol. 138, 339–344.
De Champs, C., Chanal, C., Sirot, D., Baraduc, R., Romaszko, J. P., Bonnet, R., Plaidy, A., Boyer, M., Carroy, E., Gbadamassi, M. C., Laluque, S., Oules, O., Poupart, M. C., Villemain, M., and Sirot, J. (2004). Frequency and diversity of Class A extended-spectrum beta-lactamases in hospitals of the Auvergne, France: a 2 year prospective study. J. Antimicrob. Chemother. 54, 634–639.
Decousser, J. W., Poirel, L., and Nordmann, P. (2001). Characterization of a chromosomally encoded extended-spectrum class A beta-lactamase from Kluyvera cryocrescens. Antimicrob. Agents Chemother. 45, 3595–3598.
Fluit, A. C., Terlingen, A. M., Andriessen, L., Ikawaty, R., Van Mansfeld, R., Top, J., Stuart, J. W. C., Leverstein-Van Hall, M. A., and Boel, C. H. E. (2010). Evaluation of the DiversiLab system for detection of hospital outbreaks of infections by different bacterial species. J. Clin. Microbiol. 48, 3979–3989.
Girlich, D., Poirel, L., Carattoli, A., Kempf, I., Lartigue, M. F., Bertini, A., and Nordmann, P. (2007). Extended-spectrum beta-Lactamase CTX-M-1 in Escherichia coli isolates from healthy poultry in France. Appl. Environ. Microbiol. 73, 4681–4685.
Guerra, B., Avsaroglu, M. D., Junker, E., Schroeter, A., Beutin, L., and Helmuth, R. (2007). Detection and characterisation of ESBLs in German Escherichia coli, isolated from animal, foods, and human origin between 2001-2006. Int. J. Antimicrob. Agents 29, S270–S271.
Horton, R. A., Randall, L. P., Snary, E. L., Cockrem, H., Lotz, S., Wearing, H., Duncan, D., Rabie, A., Mclaren, I., Watson, E., La Ragione, R. M., and Coldham, N. G. (2011). Fecal carriage and shedding density of CTX-M extended-spectrum beta-lactamase-producing Escherichia coli in cattle, chickens, and pigs: Implications for environmental contamination and food production. Appl. Environ. Microbiol. 77, 3715–3719.
Jarlier, V., Nicolas, M. H., Fournier, G., and Philippon, A. (1988). Extended broad-spectrum beta-lactamases conferring transferable resistance to newer beta-lactam agents in Enterobacteriaceae – hospital prevalence and susceptibility patterns. Rev. Infect. Dis. 10, 867–878.
Leverstein-Van Hall, M. A., Dierikx, C. M., Stuart, J. C., Voets, G. M., Van Den Munckhof, M. P., Van Essen-Zandbergen, A., Platteel, T., Fluit, A. C., Van De Sande-Bruinsma, N., Scharinga, J., Bonten, M. J. M., and Mevius, D. J. (2011). Dutch patients, retail chicken meat and poultry share the same ESBL genes, plasmids and strains. Clin. Microbiol. Infect. 17, 873–880.
Meunier, D., Jouy, E., Lazizzera, C., Kobisch, M., and Madec, J. Y. (2006). CTX-M-1- and CTX-M-15-type beta-lactamases in clinical Escherichia coli isolates recovered from food-producing animals in France. Int. J. Antimicrob. Agents 28, 402–407.
Moodley, A., and Guardabassi, L. (2009). Transmission of IncN plasmids carrying bla(CTX-M-1) between commensal Escherichia coli in pigs and farm workers. Antimicrob. Agents Chemother. 53, 1709–1711.
Neuwirth, C., Siebor, E., Duez, J. M., Pechinot, A., and Kazmierczak, A. (1995). Imipenem resistance in clinical isolates of Proteus mirabilis associated with alterations in penicillin binding proteins. J. Antimicrob. Chemother. 36, 335–342.
Pitout, J. D. D., and Laupland, K. B. (2008). Extended-spectrum beta-lactamase-producing enterobacteriaceae: an emerging public-health concern. Lancet Infect. Dis. 8, 159–166.
Poirel, L., Kampfer, P., and Nordmann, P. (2002). Chromosome-encoded ambler class A beta-lactamase of Kluyvera georgiana, a probable progenitor of a subgroup of CTX-M extended-spectrum beta-lactamases. Antimicrob. Agents Chemother. 46, 4038–4040.
Ranjard, L., Lejon, D. P. H., Mougel, C., Schehrer, L., Merdinoglu, D., and Chaussod, R. (2003). Sampling strategy in molecular microbial ecology: influence of soil sample size on DNA fingerprinting analysis of fungal and bacterial communities. Environ. Microbiol. 5, 1111–1120.
Rasheed, J. K., Anderson, G. J., Queenan, A. M., Biddle, J. W., Oliver, A., Jacoby, G. A., Bush, K., and Tenover, F. C. (2002). TEM-71, a novel plasmid-encoded, extended-spectrum beta-lactamase produced by a clinical isolate of Klebsiella pneumoniae. Antimicrob. Agents Chemother. 46, 2000–2003.
Reinthaler, F. F., Feierl, G., Galler, H., Haas, D., Leitner, E., Mascher, F., Melkes, A., Posch, J., Winter, I., Zarfel, G., and Marth, E. (2010). ESBL-producing E. coli in Austrian sewage sludge. Water Res. 44, 1981–1985.
Sabate, M., Miro, E., Navarro, F., Verges, C., Aliaga, R., Mirelis, B., and Prats, G. (2002). beta-Lactamases involved in resistance to broad-spectrum cephalosporins in Escherichia coli and Klebsiella spp. clinical isolates collected between 1994 and 1996, in Barcelona (Spain). J. Antimicrob. Chemother. 49, 989–997.
Woerther, P. L., Angebault, C., Lescat, M., Ruppe, E., Skurnik, D., El Mniai, A., Clermont, O., Jacquier, H., Da Costa, A., Renard, M., Bettinger, R. M., Epelboin, L., Dupont, C., Guillemot, D., Rousset, F., Arlet, G., Denamur, E., Djossou, F., and Andremont, A. (2010). Emergence and dissemination of extended-spectrum beta-lactamase-producing Escherichia coli in the community: lessons from the study of a remote and controlled population. J. Infect. Dis. 202, 515–523.
Wong-Beringer, A., Hindler, J., Loeloff, M., Queenan, A. M., Lee, N., Pegues, D. A., Quinn, J. P., and Bush, K. (2001). Molecular correlation for the treatment outcomes in bloodstream infections caused by Escherichia coli and Klebsiella pneumoniae with reduced susceptibility to ceftazidime. Clin. Infect. Dis. 34, 135–146.
Keywords: extended-spectrum beta-lactamase, CTX-M, cattle, soil, Burgundy, farm environment
Citation: Hartmann A, Locatelli A, Amoureux L, Depret G, Jolivet C, Gueneau E and Neuwirth C (2012) Occurrence of CTX-M producing Escherichia coli in soils, cattle, and farm environment in France (Burgundy region). Front. Microbio. 3:83. doi: 10.3389/fmicb.2012.00083
Received: 15 November 2011; Accepted: 19 February 2012;
Published online: 09 March 2012.
Edited by:
Rustam I. Aminov, University of Aberdeen, UKReviewed by:
Luisa Peixe, Faculdade de Farmácia da Universidade do Porto, PortugalTimothy LaPara, University of Minnesota, USA
Yoshimi Matsumoto, Osaka University, Japan
Copyright: © 2012 Hartmann, Locatelli, Amoureux, Depret, Jolivet, Gueneau and Neuwirth. This is an open-access article distributed under the terms of the Creative Commons Attribution Non Commercial License, which permits non-commercial use, distribution, and reproduction in other forums, provided the original authors and source are credited.
*Correspondence: Alain Hartmann, UMR Agroécologie 1347, INRA, The Institut National de la Recherche Agronomique Centre, 17 rue Sully, BP 86510, Dijon 21065, France. e-mail:YWxhaW4uaGFydG1hbm5AZGlqb24uaW5yYS5mcg==