 Cheng Yan
Cheng Yan Haojie Wu2
†
Haojie Wu2
†

94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Med., 10 March 2023
Sec. Translational Medicine
Volume 10 - 2023 | https://doi.org/10.3389/fmed.2023.1074878
This article is part of the Research TopicAdaptive Immunity in Local TissuesView all 12 articles
Platelets, anucleate blood components, represent the major cell type involved in the regulation of hemostasis and thrombosis. In addition to performing haemostatic roles, platelets can influence both innate and adaptive immune responses. In this review, we summarize the development of platelets and their functions in hemostasis. We also discuss the interactions between platelet products and innate or adaptive immune cells, including neutrophils, monocytes, macrophages, T cells, B cells and dendritic cells. Activated platelets and released molecules regulate the differentiation and function of these cells via platelet-derived receptors or secreting molecules. Platelets have dual effects on nearly all immune cells. Understanding the exact mechanisms underlying these effects will enable further application of platelet transfusion.
The production of platelets from megakaryocytes (MKs) is a systematic process that is thought to occur in the bone marrow (1). Thrombopoiesis occurs from common myeloid progenitor (CMP) cells in the bone marrow, which differentiate into promegakaryocytes and then into MKs. After migrating into the vascular niche, mature MKs extend many proplatelets (PPTs) through the sinusoid vessel barrier (2–4). Then, PPTs interconvert into pre-platelets, and platelets are created after the fission of pre-platelets (5). Each MK can produce 1,000–3,000 platelets after multiple divisions (6, 7). A recent study proposed the lung as the main site of platelet release (8). The average lifespan of platelets is only 8–10 days. In the circulation, each individual has 150–400 × 109 platelets per liter of peripheral blood (9).
The primary roles of platelets are hemostasis and thrombosis. Hemostasis is the process that stops blood loss from a damaged vessel (10). Hemostasis involves multiple interlinked steps: primary hemostasis, secondary hemostasis, and tertiary hemostasis (11). Platelets are mainly involved in primary hemostasis, which is also called platelet clotting. In primary hemostasis, platelets stick to the damaged tissue and become activated, which recruits more platelets to form a platelet “plug” to stop blood loss from the damaged area. Primary hemostasis may also involve constriction of the blood vessel, which can occur due to substances released by platelets (12). In addition to hemostasis, platelet activation also contributes to thrombosis, which is a blood clot within a blood vessel that limits the flow of blood. Platelets play a significant role in the development of arterial thrombosis rather than venous thrombosis (13). Atherosclerosis allows the activation of platelets, causing adhesion and aggregation, which leads to the formation of a clot. Thus, the management of arterial thrombosis predominantly involves the use of antiplatelet agents for monotherapy or dual-antiplatelet therapy (14).
For long, Platelets are small, anucleate cell debris (15). Actually, platelets possess almost every feature of cells, except the nucleus. The role of platelets in hemostasis has long been known, but they have also been shown to be involved in defense against pathogens (16, 17), as well as in the acceleration of autoimmune diseases (18). Therefore, platelets are seen as a cellular component of the innate immune system (19). In the presence of certain infectious agents or inflammatory stimuli, platelets mediate hemostasis and thrombosis and activate innate and adaptive immunity via specific receptors (CD42, CD41 CD40, CD154, etc.) and/or granule release (CXCL4, CCL5, TGFβ, serotonin, β-defensin, etc.), RNA transfer, and mitochondrial secretion (20). Moreover, it was found that platelets can also release extracellular vesicles (EVs), including ectosomes (also called microvesicles or microparticles; 100–1,000 nm) and exosomes (40–100 nm) to regulate hemostasis, thrombosis and inflammation (21). In this review, we will summarize platelet-mediated regulation of innate and adaptive immune cells.
Due to the haemostatic function of platelets, platelet transfusion is used to treat thrombocytopenia platelet function defecting disease in the clinic. Initially, platelet transfusions were thought to have no side effects, but recent findings have indicated that although the effects are not fatal, platelet transfusion can lead to febrile nonhemolytic transfusion reactions (FNHTRs), anaphylactic reactions, haemolytic transfusion reactions and other immune-mediated reactions (22). Guo et al. found that antibody-mediated immune thrombocytopenia (ITP) was resistant to allogeneic platelet transfusions, while the T-cell-mediated form of the disease was susceptible, suggesting that transfusion therapy might be beneficial for antibody-negative ITP (23). Moreover, it was reported that fresh platelets could induce transfusion-related immunomodulation (TRIM) independent of white cells (WBCs) due to their MHC antigen expression, whereas aging results in the loss of MHC and the ability to mediate TRIM (24). Ultraviolet B (UVB) radiation plus riboflavin treatment of WBC-enriched platelet-rich plasma (PRP) effectively blocks alloimmunization and modulates immune responses to subsequent exposures (25). These reports demonstrated that various reactions mediated by different WBCs led to limitations in the application of platelet transfusion. We detail the interactions between platelets and different WBCs below.
As an indispensable member of the innate immune system, neutrophils are the first leukocytes to infiltrate the site of injury (26). Platelet derived P-selectin induces neutrophils to move to sites of thrombus formation by activating P-selectin glycoprotein ligand-1 (PSGL1), a receptor of P-selectin on neutrophils (27). The platelet-derived serotonin metabolite 5-hydroxyindoleacetic acid (5-HIAA) also promotes neutrophil recruitment to inflamed tissue via the G-protein-coupled receptor 35 (GPR35) (28). In a murine model of Klebsiella pneumoniae-induced pulmonary inflammation, Toll-like receptor 4 (TLR4) deficiency in platelets decreased the number of neutrophils in the lung (29). Inhibition of platelet p110β (the catalytic subunit of phosphatidylinositol 3-kinase) prevented platelet–neutrophil interactions, diminishing neutrophil infiltration (30). It was also reported that activated platelet-derived nanovesicles could recruit neutrophils to exert anti-tumor effects (31). C-type lectin-like receptor (CLEC-2) was recently discovered as a platelet receptor. Blocking platelet CLEC-2 signaling enhanced liver recovery from acute toxic liver injuries by increasing tumor necrosis factor-α (TNF-α) production, which then improved reparative hepatic neutrophil recruitment (32). All these findings indicated that platelets regulate the movement of neutrophils.
Platelets also regulate neutrophil activation. Mac1 and LFA1 strengthen the attachment between platelets and neutrophils via junctional adhesion molecule 3 (JAM3) (33), Intercellular adhesion molecule 2 (ICAM-2) (34), CD42 (35) and Choline transporter-like protein 2 (CTL2) (36), which bind to platelet ɑIIbβ3 integrin, enhancing neutrophil activation. Clinical observational data showed that the levels of C-X-C chemokine receptor type 4 (CXCL4; also called platelet factor 4, PF4), CXCL7 (neutrophil activating protein-2, NAP2) and myeloperoxidase (MPO) were related to platelet activation and platelet–neutrophil interactions (37). Both CXCL4 and CXCL7 secreted by platelets can initiate neutrophil activation (38, 39). Similarly, inhibition of the chemokine receptors CXCR4 and CXCR7 on platelets and polymorphonuclear neutrophils (PMNs) was shown to reduce platelet–neutrophil complex (PNC) formation (40). Leukotriene B4 (LTB4) and leukotriene A4 (LTA4) derived from platelet-derived arachidonic acid (AA) can activate neutrophils (41). In addition, platelet-derived mitochondria induce the release of human neutrophil microvesicles that recruit additional immune cells to remove pathogens (42). Platelet-derived serotonin was shown to promote neutrophil degranulation, which increased the expression of the membrane-bound leukocyte adhesion molecule CD11b, enhanced inflammation in the infarct area and reduced myocardial salvage by inducing the release of myeloperoxidase and hydrogen peroxide (H2O2) (43).
An important function of neutrophils is to release neutrophil extracellular traps (NETs), which remove pathogens from the circulation. Platelet-derived exosomal high-mobility group protein 1 (HMGB1) and/or miR-15b-5p and miR-378a-3p promote excessive NET formation through the Akt/mTOR autophagy pathway during sepsis and subsequent organ injury (44). It was reported that both HMGB1 and the C3a component released by platelets could activate neutrophils to induce the formation of NETs (45–47). These stimuli significantly enhanced PSGL-1-induced neutrophil activation. Additionally, platelets interact with C3b attached to NETs (48). The P-selectin-PSGL-1 interaction was shown to induce the release of NETs, and clearing activated platelets via platelet-derived microparticle (100–1,000 nm) mediated neutrophil activation (49). Moreover, platelets were able to induce NOX-independent NET formation in a dengue virus non-structural protein 1 (NS1)-dependent manner (50). The inhibition of nuclear factor of activated T cells (NFAT) in platelets promotes interactions with neutrophils and NET induction, which might be harnessed in the clinic (51). In addition to influencing the infiltration of neutrophils, the TLR4-ERK5 axis in platelets facilitates NET formation to promote the capture of circulating tumor cells (52). Furthermore, a neutralizing anti-CXCL4 antibody significantly inhibited NET formation induced by NCA-associated vasculitis (AAV)-derived platelets (38).
Interestingly, NETs also induce a hypercoagulable state in platelets by upregulating phosphatidylserine and P-selectin on cells in the context of gastric cancer (GC) (53). Neutrophils can activate platelets by releasing antimicrobial cathelicidins via degranulation or as part of NETs. For example, cathelicidin LL-37 and its mouse homolog cathelicidin-related antimicrobial peptide (CRAMP) can bind glycoprotein VI (GPVI) on the platelet surface, further stimulating platelet and neutrophil activation (54). Citrullinated H3 histones, key markers of ongoing NETosis, have also been shown to activate platelets (48). During ischaemic stroke, neutrophils can rapidly bind platelets through P-selectin and glycoprotein Ibα (55), and neutrophils have been shown to undergo “plucking” on megakaryocytes to accelerate platelet production via CXCR4-CXCL12 signaling (56). Additionally, neutrophils activate platelets after pneumolysin exposure by releasing extracellular vesicles (EVs; 100–1,000 nm) (57). In acute myocardial infarction, the observed increase in S100A8/A9 levels in platelets was not due to an increase in synthesis but was due to uptake of proteins secreted by neutrophils (58). This result indicated that neutrophils were able to alter the platelet proteome.
Taken together, these findings support mutual regulation between platelets and neutrophils. Platelets can regulate infiltration, activation and NET formation in neutrophils (Table 1). In the clinic, depending on the specific conditions of a disease, the mechanism of mutual regulation between these cell types could be controlled or blocked.
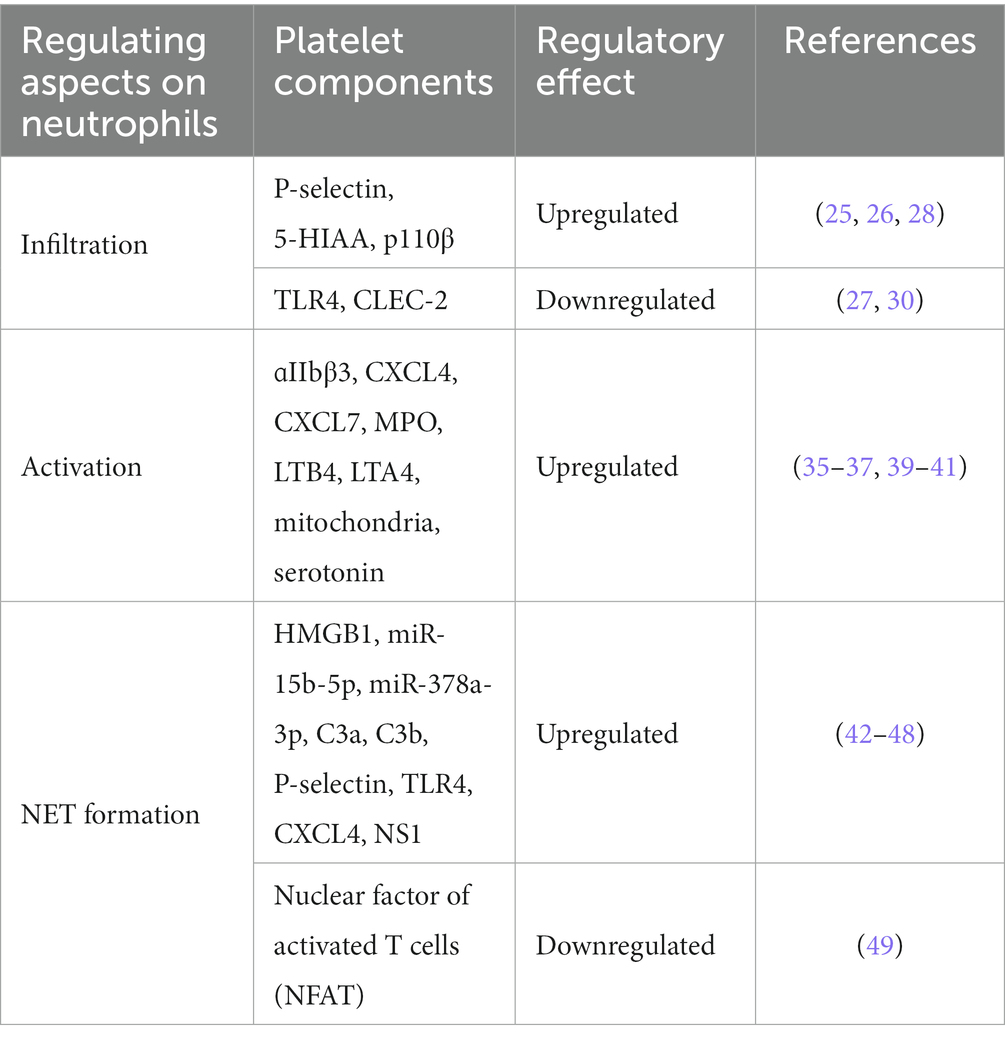
Table 1. Platelet-derived molecules affect the function of neutrophils.
Platelets mediate multiple types of immune responses, and many studies have shown that platelets can interact with innate immune cells during infection and inflammation. One study showed that platelet activation is the major initiator of platelet–monocyte aggregation (59). Platelets interact with monocytes through cluster of differentiation (CD)62p (known as P-selectin), which recognizes PSGL-1 expressed on the surface of monocytes to initiate aggregation (60). Platelet-derived hyaluronidase-2 (HYAL2) also causes aggregate formation (61). Platelets from severe COVID-19 patients were highly activated and induced the expression of tissue factor (TF) in monocytes from healthy volunteers (62). The increased expression of TF also drives platelet–monocyte aggregation (63), inflammatory activation and inflammatory cytokine secretion (64). However, activated platelet-derived EVs (40–100 nm) contribute to the suppression of TF expression by transferring hsa-miR-223-3p to monocytes, which inhibits aggregation (65). Therefore, the exact influence of platelets on platelet–monocyte aggregation needs to be further explored.
Platelets can induce the oxidative burst and inflammation in monocytes and neutrophils via direct interactions (66–70) and promote leukocyte adhesion and extravasation (71–73). In addition, platelet aggregability leads to monocyte extravasation into the infarcted myocardium and influences inflammation in patients with acute myocardial infarction (74). The SARS-CoV-2 spike protein can interact with the CD42b receptor to activate platelets and promote proinflammatory cytokine production by monocytes through the interaction of P-selectin/PGSL-1 and CD40L/CD40 (75). P-selectin was shown to contribute to the secretion of TNFα, IL-1β, IL-6, CXCL8, IL12 and CCL4 by autologous monocytes (76, 77). In addition, platelets were reported to potentiate the release of IL-8, mainly from monocytes (78). Thus, some researchers have concluded that platelet–monocyte aggregates can be used as a robust marker of platelet activation and monocyte inflammatory responses (79). Interestingly, platelets do not always activate monocytes to induce the production of proinflammatory cytokines, and platelet-monocyte interactions can actually decrease inflammation by increasing IL-10 levels and reducing TNF-α levels in monocytes through CD40L/CD40 (80).
Platelets also regulate the differentiation of monocytes. Both Sigrun Badrnya et al. and J H Phillips et al. reported that activated platelets increased CD16 expression, which induced monocytes to switch to an intermediate phenotype. CD16+ monocytes produced transcripts for the gamma subunit of the high-affinity IgE FcR and could kill anti-CD16 hybridoma cell targets in the absence of CD3 zeta (81, 82). Similarly, it was reported that platelet-derived CXCL4 induced monocyte differentiation into macrophages (83). However, another study reported that inhibition of PSGL-1 or P-selectin did not attenuate platelet-mediated monocyte activation (84). This meant that there were other pathways activating monocytes. Zachary et al. found that platelet-derived β-2 microglobulin (β2M) induced monocyte proinflammatory differentiation through a noncanonical TGFβ receptor pathway (85) and regulated age-associated monocyte polarization. β2M was shown to maintain the balance between inflammatory and reparative signals. In addition, loss of β2M increases profibrotic cardiac responses (86). Thus, platelets have both pro- and anti-inflammatory effects on monocytes (Table 2).
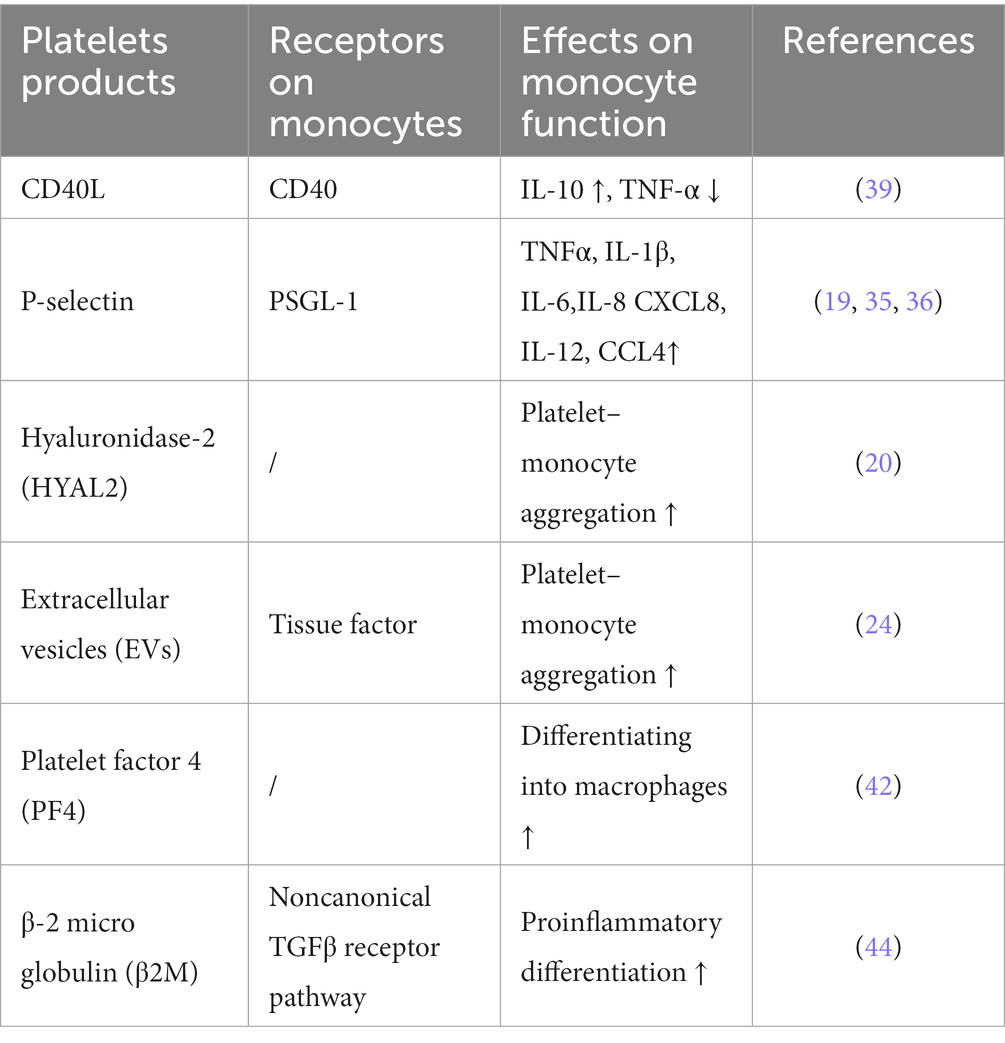
Table 2. Platelet-derived molecules affect the function and differentiation of monocytes.
The phenotype and function of macrophages are also affected by platelets. Haem-activated platelets promote the formation of macrophage extracellular traps (METs) via reactive oxygen species generation or histone citrullination, enhancing rhabdomyolysis-induced acute kidney injury (87). Platelet-conditioned medium was also shown to induce an anti-inflammatory, pro-resolving phenotype in macrophages (88). Platelet-Treg cell aggregates in the lung induce macrophage polarization toward an anti-inflammatory phenotype and promote effective resolution of pulmonary inflammation (89). Ryoka et al. reported that platelet-rich plasma (PRP) suppressed M1 macrophage polarization and promoted M2 macrophage polarization (90). Platelet-rich fibrin could shift macrophage polarization from an M1 phenotype toward an M2 phenotype to induce an anti-inflammatory response (91) and reduce IL-1β release and caspase-1 production in macrophages that underwent pyroptosis by increasing NLR family pyrin domain containing 3 (NLRP3) ubiquitination (92, 93). Interestingly, both leukocyte-poor (LP) and leukocyte-rich (LR) PRP promoted the recruitment of M1 macrophages during the process of tendon healing, while the number of M2 macrophages was high only in the LP-PRP group (94). However, the presence of platelets skewed monocytes toward an M1 phenotype via the GPIb-CD11b axis in the presence of lipopolysaccharide (LPS) (95). Thus, platelets can affect macrophage polarization via different pathways.
Natural killer cells are also regulated by platelets. PLT-ectosomes (100–1,000 nm) can inhibit NK cell effector function in a TGF-β1-dependent manner, reducing the expression of surface receptors, like natural-killer group 2, member D (NKG2D), natural-killer p30 (NKp30) and CD226, and IFN-γ production (96). This result suggested that platelets can promote tumor dissemination by coating tumor cells (97, 98). Co-incubation of NK cells with platelets was shown to reduce NK cell cytotoxicity by reducing NK cell degranulation, IFN-γ production, NKG2D and natural-killer p46 (NKp46) expression and increased Killer cell immunoglobulin-like receptor 2DL1 (KIR2DL1) expression in NK cells (99). Furthermore, NK cell cytolytic activity was shown to be attenuated via tumor cell-induced platelet secretion (100). However, platelet-derived growth factor D (PDGF-DD)-activated IL-2-induced NK cells exert anti-tumor effects by binding with the NKp44 receptor (101), and PDGF-D-PDGFRβ signaling enhances IL-15-mediated human NK cell survival (102). In addition, platelet-derived CXCL4 induces human natural killer cells to synthesize and release interleukin-8 (103). Taken together, these findings show that platelets also have dual regulatory effects on NK cells.
In addition to innate immune cells, platelets also influence T-cell and B-cell responses. Norbert et al. found that platelets enhanced the differentiation and cytokine production of CD4+ T cells via both direct cell–cell contact and multiple chemokines (platelet-derived CXCL4 and CCL5) (104). Platelets can produce many molecules, such as FasL, TNF-related apoptosis-inducing ligand (TRAIL), IL-7 and CD40L, which are all important for adaptive immune responses (105–107). In particular, platelet CD40L regulates CD8+ T-cell response. Elzey et al. reported that depletion of platelets decreased the generation of cytotoxic T lymphocytes (CTLs) (108). In platelet-depleted mice, reconstitution of platelets increased the number of CTLs in the spleen and liver after lymphocytic choriomeningitis virus (LCMV) infection (109). Thus, platelets are important for the expansion of antigen-specific CTLs. Chapman et al. also demonstrated that platelets can process and present antigens via MHC class I and directly activate naive T cells in a platelet MHC class I-dependent manner (110). However, platelets were also shown to delay the infiltration of CD8+ T cells into the liver, allowing increased viral replication via the release of serotonin, which might ultimately cause chronic hepatitis (111). In addition, Aslam et al. found that platelets suppressed CD8+ T-cell function in a transfusion-related model and that transfusion of MHC-I bearing platelets prolonged allograft survival (24). Interestingly, the expression of MHC-I in platelets was shown to be significantly increased in humans and mice, which reduced the numbers and impaired the function of antigen-specific CD8+ T cells during sepsis (112). Platelets promote protection against C. albicans airway mycosis by activating Th2 and Th17 responses via an antifungal pathway that includes candidalysin, GP1bα, and dickkopf WNT signaling pathway inhibitor 1 (Dkk-1) (113). However, activated platelets accumulate in the lung lesions of tuberculosis patients and inhibit T-cell responses and mycobacterium tuberculosis replication in macrophages (114). CD84-lacking platelets were shown to reduce cerebral CD4+ T-cell infiltration and thrombotic activity, slowing neurological damage in an experimental model of stroke. In a clinical study, a high level of platelet CD84 expression resulted in poor outcomes in patients with stroke (115). Platelets block the immunosuppressive function of Tregs directly via the P-selectin/PSGL-1 axis, which induces Syk phosphorylation and an increase in intracellular calcium in systemic lupus erythematosus (SLE) patients (116). However, Jan et al. reported that interactions between platelets and Tregs via the P-selectin/PSGL-1 axis encouraged the release of the anti-inflammatory mediators IL-10 and TGFβ. Platelet-Treg cell aggregates induce macrophage polarization toward an anti-inflammatory phenotype in pulmonary inflammation (89). Rachidi et al. found that platelets constrained T-cell immunity through a glycoprotein repetition predominant (GARP)-TGFβ axis, and platelet-specific deletion of GARP potentiated protective immunity against both melanoma and colon cancer (117). In addition, Hinterleitner et al. reported that platelet-derived GARP induced peripheral Treg cells by upregulating Foxp3 expression (118).
Platelet-derived CXCL4 was also shown to enhance Th1 cell responses and CD4+ T effector memory cell responses via Akt-PGC1α-TFAM signaling-mediated mitochondrial biogenesis (119). Platelets exert dose-dependent regulatory effects on the effector responses of naive T cells via CXCL4-TGFβ. Low concentrations of CXCL4 reinforce TGFβ signaling, but high concentrations of CXCL4 have the opposite effect (120). In addition, knocking down the expression of CXCR3, the receptor of CXCL4, was shown to abolish Th1 and Treg cell responses (121). Platelet-derived mitochondria directly upregulate central memory (TCM) CD4+ T cells and downregulate effector memory (TEM) CD4+ T cells through C-X-C motif chemokine receptor 4 (CXCR4) and its ligand stromal cell-derived factor-1 (SDF-1) (122). However, CXCL4 expression is inversely correlated with T-cell function in advanced lung adenocarcinoma (LAC), leading to accelerated development of tumors (123). Pten-deficient platelets are hyperactive and overproduce multiple Tfh-promoting cytokines via the PDK1/mTORC2-AKT-SNAP23 axis, which promotes CD4+ T-cell differentiation into Tfh cells. Pten deletion results in age-related lymphoproliferative diseases and humoral autoimmunity in mice (124).
PD-L1 is widely known as an inhibitor of the adaptive immune system. It can be transferred from tumor cells to platelets in a fibronectin 1-, integrin α5β1- and GPIbα-dependent manner in non-small cell lung cancer (NSCLC), and platelet PD-L1 possesses the ability to inhibit the function of CD4+ and CD8+ T cells (118). Christina et al. demonstrated that platelets decrease PD-1 and PD-L1 expression, T-cell proliferation and IFN-γ and TNF-α production (125). PD-L1-overexpressing platelets can rescue β-cells by suppressing the activity of pancreatic autoreactive T cells and increasing the percentage of Tregs in type 1 diabetes (T1D) (126). However, high expression of PD-L1 was found in platelets from COVID-19 patients, which inhibited the upregulation of CD25 expression and TNF-α and IFN-γ production by CD4+ T cells (127). Activated platelet-derived IL-33, Dkk-1, and 5-HT or CD40L induce type 2 immune response or interact with TSLP-stimulated myeloid DCs to tune the sensitization stage of allergic asthma through RANK/RANKL signaling (128). In addition, platelet-CD4+ T-cell aggregate frequency was positively correlated with HIV-1 viral load and was related to immune activation during HIV-1 infection (129). HIV-containing platelets result in dysfunction of glycolysis-mediated energy production in CD4+ T cells. This result indicates that platelets might be a therapeutic target for immunological non-responders (130) (Figure 1).
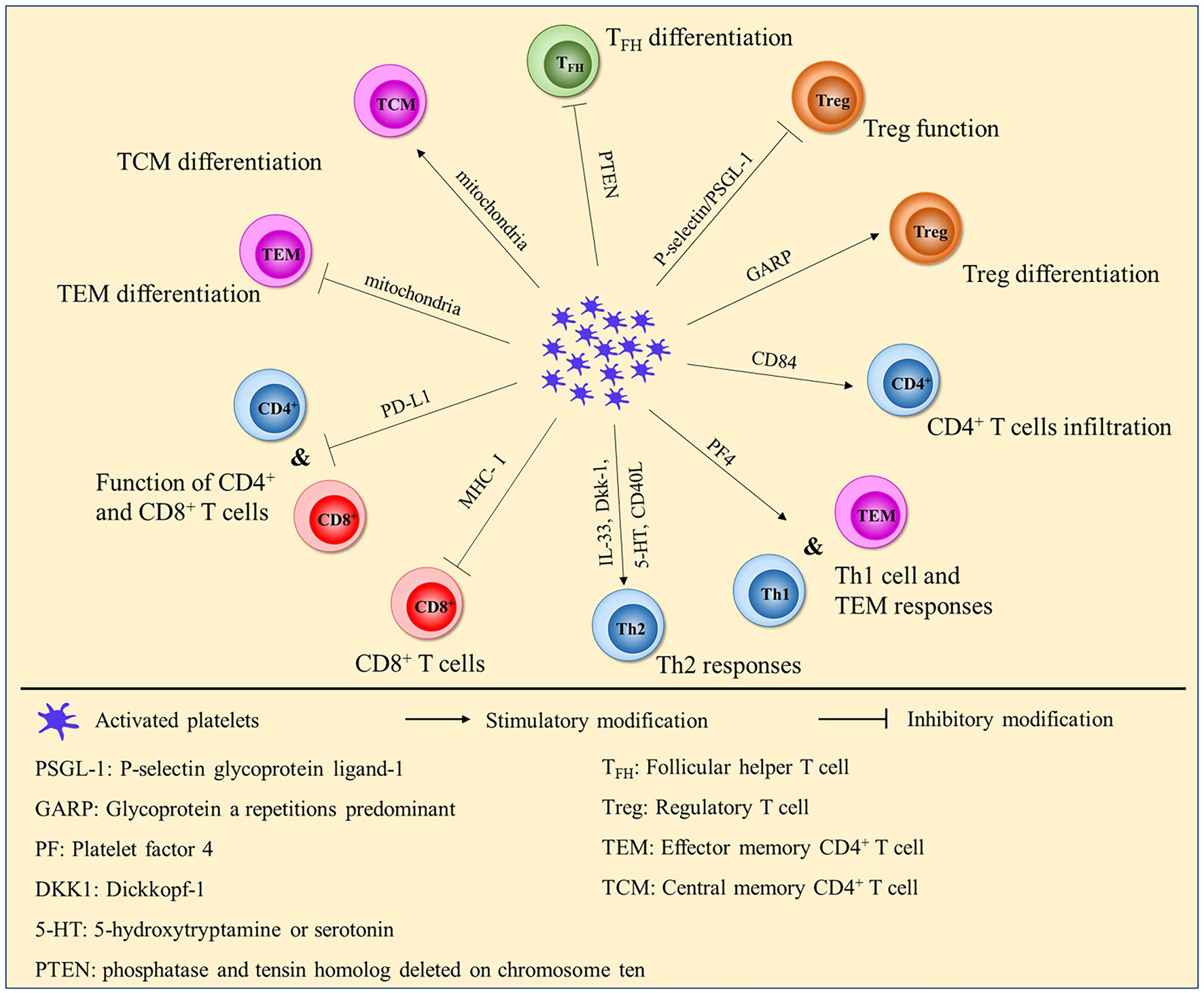
Figure 1. Platelets affect the function and differentiation of CD4+ and CD8+ T cells.
Compared with those on T cells, the effects of platelets on B cells are less well studied. It was reported that the transfer of normal platelets into CD40L-deficient mice could transiently increase antigen-specific IgG production (131, 132). Fabrice et al. also reported that platelets could activate peripheral blood B cells and increase the production of immunoglobulins (133). These results indicated that platelet CD40L contributed to B-cell responses when CD4+ T-cell-derived CD40L was absent. CD40L released from platelets contributes to the proliferation of tumor cells in intravascular large B-cell lymphoma (134). In addition, platelets secreting PF4 increase B-cell differentiation in the bone marrow environment by inducing the phosphorylation of STAT5 (135).
Since the start of the COVID-19 pandemic, vaccines for SARS-CoV-2 have been developed. In addition, numerous researchers have found that vaccine-induced immune thrombotic thrombocytopenia (VITT) occurs in individuals exposed to vaccines, especially adenoviral vector vaccines (136). VITT is an autoimmune condition characterized by antibodies that directly activate platelets, triggering thrombosis in the arterial and venous circulation. The pathophysiology of VITT is still incompletely understood (137). One hypothesis suggested that vaccines might bind to PF4, creating a novel antigen that is subsequently taken up by monocytes and trafficked to the lymph nodes, where it stimulates the proliferation of anti-PF4 memory B cells. Then, antibody binding to FcγRIII-A contributes to platelet clearance and thrombocytopenia (138). Additionally, platelets express human Fc receptors. The receptors for IgG, the Fc-γ receptor, and IgE, the Fc-ε receptor, are expressed on the platelet surface (139). FcγRIIA-expressing platelets activated by IgG immune complexes contribute to the severity of anaphylaxis (140). IgE antibody binding to platelets via the low-affinity IgE receptor (Fc epsilon RII/CD23) or high-affinity IgE receptor (Fc epsilon RI) led to immediate-type allergic reactions (141, 142). Thus, platelets and their secreted molecules can influence B cells in adaptive immunity. In addition, antibodies released by B cells are also able to regulate the numbers and activation of platelets.
Although many studies have indicated that platelets play critical roles in T-cell and B-cell adaptive immunity, the mechanism is still unknown. In addition to their ability to mediate T-cell and B-cell immune responses directly, platelets might also regulate these responses indirectly through dendritic cells (DCs). DCs are the primary antigen-presenting cells and can cross-present antigens to T cells to induce antigen-specific cell responses. Thus, changes in the number or phenotype of DCs influence cell immunity.
Platelets were reported to significantly inhibit proinflammatory (IL-12, IL-6, and TNFα) cytokine production and increase anti-inflammatory (IL-10) cytokine production in moDCs. In addition, platelet-derived soluble mediators inhibit T-cell priming and T helper differentiation toward an IFNγ+ Th1 phenotype induced by moDCs (143). Platelet concentrates also downregulate the expression of CD40, CD80, CD83, and CD86 and IL-8, IL-12 and IL-6 secretion by BDCA+ DCs (144, 145). Similarly, platelet-monocyte complex (PMC)-derived DCs were shown to exhibit reduced levels of critical molecules for DC function (CD206, CD80, CD86, and CCR7) and reduced antigen uptake capacity (146). Conversely, thrombin-activated platelets increased CD80 expression in DCs and induced DCs to produce tumor necrosis factor alpha (TNF-α), interleukin 12 (IL-12), and IL-6 after coculture of BMDCs and staphylococcus aureus in vitro (147). Furthermore, platelets have the ability to enhance the DC-mediated Th2 response and contribute to allergic inflammation through the RANK ligand (148). Platelets are necessary for efficient host sensitization to allergens and increase the allergen sensitization of CD11c+ DCs (149).
Platelet-derived P-selectin interacts with PSGL-1 on the surface of monocytes and induces monocyte differentiation into DCs, which are more potent than cytokine-derived DCs during tumor-specific T-cell immune responses (150). Additionally, the interaction between platelets and DCs is mediated by Mac-1, which is upregulated on DCs by activated platelets in a PSGL-1-dependent manner (151). Moreover, platelet-derived CD40L can induce monocyte differentiation into DCs, promote DC maturation, increase the expression of costimulatory molecules (152, 153) and enhance interferon-α secretion by plasmacytoid dendritic cells in systemic lupus erythematosus (154). Sharmeen et al. also reported that platelets enhanced dendritic cell responses through CD40-CD40L during staphylococcus aureus infection (147).
Serotonin, another platelet molecule, had opposite effects on DC differentiation. As the expression of costimulatory molecules on DCs was reduced and IL-10 production was increased by serotonin, the antigen presentation function of DCs was repressed (155). Similarly, CXCL4 inhibited monocyte differentiation into CD1α+ DCs and increased the number of CD1α− DCs, but CD1α− DCs were not as effective as CD1α+ DCs in activating T cells (156). Moreover, CXCL4 enhanced monocyte-derived DCs to promote autologous CD4+ T-cell and CD8+ T-cell proliferation and the production of IFN-γ and IL-4 (157). In summary, platelets and their secreted molecules have different effects on the development or differentiation of DCs, and the exact mechanism still needs further exploration (Table 3).
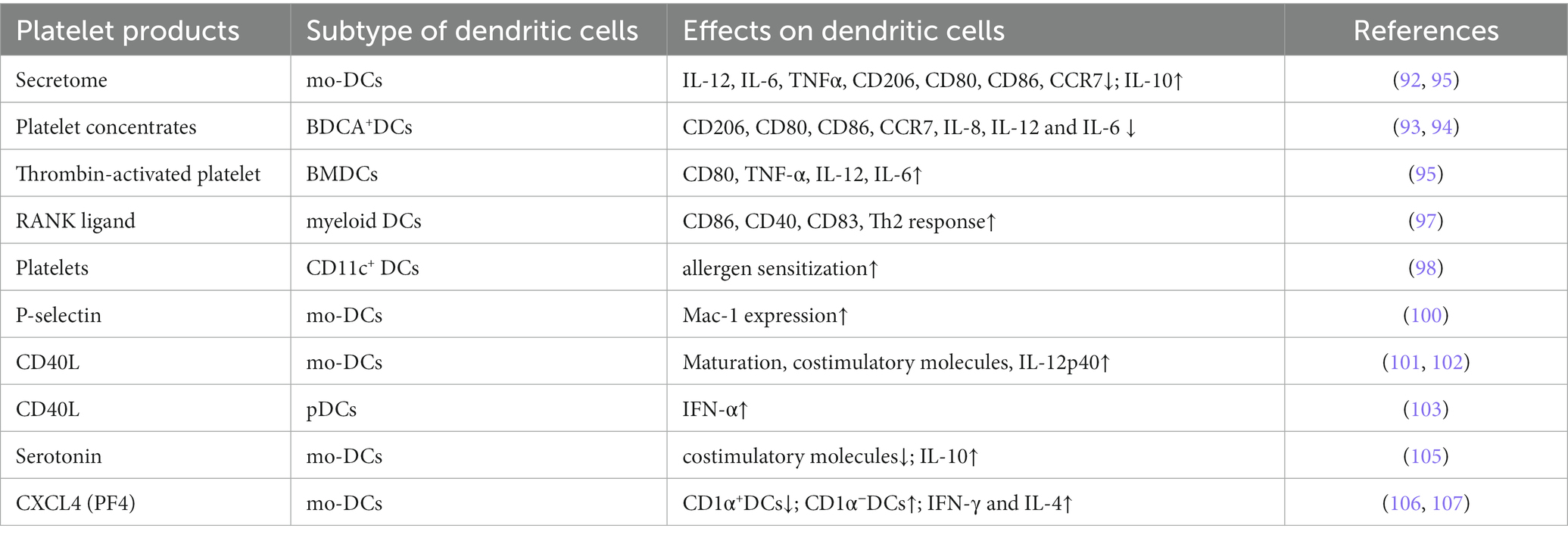
Table 3. Platelet-derived molecules affect the function and differentiation of dendritic cells.
In addition to supporting thrombosis, platelets release a number of mediators that regulate both innate and adaptive immunity. Due to the large number of platelets in the circulation, they and their products can efficiently interact with peripheral circulating cells directly, such as neutrophils, monocytes, T cells, B cells, DCs, macrophages, and NK cells, which modulates their differentiation. Platelets affect the functions of monocytes and neutrophils, including their receptors and soluble mediators. Moreover, platelet interactions with monocytes induce their differentiation into macrophages and regulate cytokine release. Depending on the severity of the disease, platelets can enhance or reduce leukocyte cytokine production, which maintains a balance to limit excessive inflammation during infection. Platelet-derived CD40L or other ligands can also modulate adaptive immunity. Additionally, both in vitro and in vivo evidence suggests that platelets also impact the development and functions of DCs to regulate T-cell and B-cell responses. According to previous reports, different products of platelets have different effects on DCs, thus changing their antigen presentation capacity.
All these reports indicated that in addition to hemostasis, platelets also play critical roles in the immune system, but the exact mechanism is still not clear. Given that platelet concentrates are widely used in clinical treatment and given the side effects of platelet transfusion, we need to consider the effects of platelets and their secreted molecules on immune cells, such as neutrophils, monocytes, B cells, T cells and DCs. Elucidating how platelets interact with these cells will contribute to broader application of platelet products and avoid adverse reactions.
CY and HW mainly drafted and revised this review. XF and JH conceived and provided lots of advises about this review. FZ conducted the overall writing and previewed the draft and revised the manuscript. All authors contributed to the article and approved the submitted version.
The present study was supported by grants Scientific Research and Development Fund of Kangda College, Nanjing Medical University (grant no. KD2019KYJJZD019) and the Jiangning District Social Undertakings Science and Technology Development Project (grant no. 2020SHSY0101).
The authors would like to thank Hongmei Zhang for the scientific editing to improve the manuscript writing. This review could not be performed without her help.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
1. Boscher, J, Guinard, I, Eckly, A, Lanza, F, and Léon, C. Blood platelet formation at a glance. J Cell Sci. (2020) 133:jcs244731. doi: 10.1242/jcs.244731
2. Bornert, A, Boscher, J, Pertuy, F, Eckly, A, Stegner, D, Strassel, C, et al. Cytoskeletal-based mechanisms differently regulate in vivo and in vitro proplatelet formation. Haematologica. (2021) 106:1368–80. doi: 10.3324/haematol.2019.239111
3. Junt, T, Schulze, H, Chen, Z, Massberg, S, Goerge, T, Krueger, A, et al. Dynamic visualization of thrombopoiesis within bone marrow. Science. (2007) 317:1767–70. doi: 10.1126/science.1146304
4. Kowata, S, Isogai, S, Murai, K, Ito, S, Tohyama, K, Ema, M, et al. Platelet demand modulates the type of intravascular protrusion of megakaryocytes in bone marrow. Thromb Haemost. (2014) 112:743–56. doi: 10.1160/TH14-02-0123
5. Machlus, KR, and Italiano, JE Jr. The incredible journey: from megakaryocyte development to platelet formation. J Cell Biol. (2013) 201:785–96. doi: 10.1083/jcb.201304054
7. Kellie, R.M., and Joseph, E.I., 2—megakaryocyte development and platelet formation. (2019): p. 25–46. doi: 10.1016/B978-0-12-813456-6.00002-3
8. Lefrançais, E, Ortiz-Muñoz, G, Caudrillier, A, Mallavia, B, Liu, F, Sayah, DM, et al. The lung is a site of platelet biogenesis and a reservoir for haematopoietic progenitors. Nature. (2017) 544:105–9. doi: 10.1038/nature21706
9. Thon, JN, and Italiano, JE. Platelets: production, morphology and ultrastructure. Handb Exp Pharmacol. (2012) 210:3–22. doi: 10.1007/978-3-642-29423-5_1
10. Antonio, B., and Gustavo, B., “Hemostasis,” in Medical Biochemistry. (2017): Academic Press. p. 781–789.
11. Morayma, S., “Overview of the coagulation system,” in Transfusion Medicine and Hemostasis. (2019): Elsevier. p. 559–564.
12. Broos, K, Feys, HB, de Meyer, SF, Vanhoorelbeke, K, and Deckmyn, H. Platelets at work in primary hemostasis. Blood Rev. (2011) 25:155–67. doi: 10.1016/j.blre.2011.03.002
13. Insull, W Jr. The pathology of atherosclerosis: plaque development and plaque responses to medical treatment. Am J Med. (2009) 122:S3–S14. doi: 10.1016/j.amjmed.2008.10.013
15. Pease, DC. An electron microscopic study of red bone marrow. Blood. (1956) 11:501–26. doi: 10.1182/blood.V11.6.501.501
16. Koupenova, M, Vitseva, O, MacKay, CR, Beaulieu, LM, Benjamin, EJ, Mick, E, et al. Platelet-TLR7 mediates host survival and platelet count during viral infection in the absence of platelet-dependent thrombosis. Blood. (2014) 124:791–802. doi: 10.1182/blood-2013-11-536003
17. Gaertner, F, Ahmad, Z, Rosenberger, G, Fan, S, Nicolai, L, Busch, B, et al. Migrating platelets are Mechano-scavengers that collect and bundle bacteria. Cell. (2017) 171:1368–1382.e23. doi: 10.1016/j.cell.2017.11.001
18. Linge, P, Fortin, PR, Lood, C, Bengtsson, AA, and Boilard, E. The non-haemostatic role of platelets in systemic lupus erythematosus. Nat Rev Rheumatol. (2018) 14:195–213. doi: 10.1038/nrrheum.2018.38
19. Garraud, O, and Cognasse, F. Are platelets cells? And if yes, are they immune cells? Front Immunol. (2015) 6:70. doi: 10.3389/fimmu.2015.00070
20. Koupenova, M, Clancy, L, Corkrey, HA, and Freedman, JE. Circulating platelets as mediators of immunity, inflammation, and thrombosis. Circ Res. (2018) 122:337–51. doi: 10.1161/CIRCRESAHA.117.310795
21. Puhm, F, Boilard, E, and Machlus, KR. Platelet extracellular vesicles: beyond the blood. Arterioscler Thromb Vasc Biol. (2021) 41:87–96. doi: 10.1161/ATVBAHA.120.314644
22. Kiefel, V. Reactions induced by platelet transfusions. Transfus Med Hemother. (2008) 35:354–8. doi: 10.1159/000151350
23. Guo, L, Yang, L, Speck, ER, Aslam, R, Kim, M, McKenzie, CGJ, et al. Allogeneic platelet transfusions prevent murine T-cell-mediated immune thrombocytopenia. Blood. (2014) 123:422–7. doi: 10.1182/blood-2013-08-523308
24. Aslam, R, et al. Transfusion-related immunomodulation by platelets is dependent on their expression of MHC class I molecules and is independent of white cells. Transfusion. (2008) 48:1778–86. doi: 10.1111/j.1537-2995.2008.01791.x
25. Jackman, RP, Muench, MO, Heitman, JW, Inglis, HC, Law, JP, Marschner, S, et al. Immune modulation and lack of alloimmunization following transfusion with pathogen-reduced platelets in mice. Transfusion. (2013) 53:2697–709. doi: 10.1111/trf.12133
26. Silvestre-Roig, C, Braster, Q, Ortega-Gomez, A, and Soehnlein, O. Neutrophils as regulators of cardiovascular inflammation. Nat Rev Cardiol. (2020) 17:327–40. doi: 10.1038/s41569-019-0326-7
27. Smyth, SS, Reis, ED, Zhang, W, Fallon, JT, Gordon, RE, and Coller, BS. Beta(3)-integrin-deficient mice but not P-selectin-deficient mice develop intimal hyperplasia after vascular injury: correlation with leukocyte recruitment to adherent platelets 1 hour after injury. Circulation. (2001) 103:2501–7. doi: 10.1161/01.CIR.103.20.2501
28. de Giovanni, M, Tam, H, Valet, C, Xu, Y, Looney, MR, and Cyster, JG. GPR35 promotes neutrophil recruitment in response to serotonin metabolite 5-HIAA. Cell. (2022) 185:1103–4. doi: 10.1016/j.cell.2022.03.003
29. Revenstorff, J, Ludwig, N, Hilger, A, Mersmann, S, Lehmann, M, Grenzheuser, JC, et al. Role of S100A8/A9 in platelet-neutrophil complex formation during acute inflammation. Cells. (2022) 11:3944. doi: 10.3390/cells11233944
30. Schrottmaier, WC, Kral-Pointner, JB, Salzmann, M, Mussbacher, M, Schmuckenschlager, A, Pirabe, A, et al. Platelet p110beta mediates platelet-leukocyte interaction and curtails bacterial dissemination in pneumococcal pneumonia. Cell Rep. (2022) 41:111614. doi: 10.1016/j.celrep.2022.111614
31. Shang, Y, Sun, J, Wu, X, and Wang, Q. Activated platelet membrane nanovesicles recruit neutrophils to exert the antitumor efficiency. Front Chem. (2022) 10:955995. doi: 10.3389/fchem.2022.955995
32. Chauhan, A, Sheriff, L, Hussain, MT, Webb, GJ, Patten, DA, Shepherd, EL, et al. The platelet receptor CLEC-2 blocks neutrophil mediated hepatic recovery in acetaminophen induced acute liver failure. Nat Commun. (2020) 11:1939. doi: 10.1038/s41467-020-15584-3
33. Santoso, S, Sachs, UJH, Kroll, H, Linder, M, Ruf, A, Preissner, KT, et al. The junctional adhesion molecule 3 (JAM-3) on human platelets is a counterreceptor for the leukocyte integrin mac-1. J Exp Med. (2002) 196:679–91. doi: 10.1084/jem.20020267
34. Kuijper, PH, Torres, HIG, Lammers, JWJ, Sixma, JJ, Koenderman, L, and Zwaginga, JJ. Platelet associated fibrinogen and ICAM-2 induce firm adhesion of neutrophils under flow conditions. Thromb Haemost. (1998) 80:443–8. doi: 10.1055/s-0037-1615227
35. Li, J, Kim, K, Jeong, SY, Chiu, J, Xiong, B, Petukhov, PA, et al. Platelet protein disulfide isomerase promotes glycoprotein Ibalpha-mediated platelet-neutrophil interactions under Thromboinflammatory conditions. Circulation. (2019) 139:1300–19. doi: 10.1161/CIRCULATIONAHA.118.036323
36. Constantinescu-Bercu, A, Grassi, L, Frontini, M, Salles-Crawley, II, Woollard, K, and Crawley, JTB. Activated alpha(IIb)beta(3) on platelets mediates flow-dependent NETosis via SLC44A2. elife. (2020) 9:9. doi: 10.7554/eLife.53353
37. Kollikowski, AM, Pham, M, März, AG, Papp, L, Nieswandt, B, Stoll, G, et al. Platelet activation and chemokine release are related to local neutrophil-dominant inflammation during Hyperacute human stroke. Transl Stroke Res. (2022) 13:364–9. doi: 10.1007/s12975-021-00938-w
38. Matsumoto, K, Yasuoka, H, Yoshimoto, K, Suzuki, K, and Takeuchi, T. Platelet CXCL4 mediates neutrophil extracellular traps formation in ANCA-associated vasculitis. Sci Rep. (2021) 11:222. doi: 10.1038/s41598-020-80685-4
39. Bdeir, K, Gollomp, K, Stasiak, M, Mei, J, Papiewska-Pajak, I, Zhao, G, et al. Platelet-specific chemokines contribute to the pathogenesis of acute lung injury. Am J Respir Cell Mol Biol. (2017) 56:261–70. doi: 10.1165/rcmb.2015-0245OC
40. Ngamsri, KC, Putri, RA, Jans, C, Schindler, K, Fuhr, A, Zhang, Y, et al. CXCR4 and CXCR7 inhibition ameliorates the formation of platelet-neutrophil complexes and neutrophil extracellular traps through Adora2b signaling. Int J Mol Sci. (2021) 22:13576. doi: 10.3390/ijms222413576
41. Li, ZS, and Smyth, SS. Interactions between platelets, leukocytes, and the endothelium. Platelets. (2019) 8:295–310. doi: 10.1016/B978-0-12-813456-6.00016-3
42. Léger, JL, Soucy, MN, Veilleux, V, Foulem, RD, Robichaud, GA, Surette, ME, et al. Functional platelet-derived mitochondria induce the release of human neutrophil microvesicles. EMBO Rep. (2022) 23:e54910. doi: 10.15252/embr.202254910
43. Mauler, M, Herr, N, Schoenichen, C, Witsch, T, Marchini, T, Härdtner, C, et al. Platelet serotonin aggravates myocardial ischemia/reperfusion injury via neutrophil degranulation. Circulation. (2019) 139:918–31. doi: 10.1161/CIRCULATIONAHA.118.033942
44. Jiao, Y, Li, W, Wang, W, Tong, X, Xia, R, Fan, J, et al. Platelet-derived exosomes promote neutrophil extracellular trap formation during septic shock. Crit Care. (2020) 24:380. doi: 10.1186/s13054-020-03082-3
45. Koupenova, M, Corkrey, HA, Vitseva, O, Manni, G, Pang, CJ, Clancy, L, et al. The role of platelets in mediating a response to human influenza infection. Nat Commun. (2019) 10:1780. doi: 10.1038/s41467-019-09607-x
46. Vogel, S, Bodenstein, R, Chen, Q, Feil, S, Feil, R, Rheinlaender, J, et al. Platelet-derived HMGB1 is a critical mediator of thrombosis. J Clin Invest. (2015) 125:4638–54. doi: 10.1172/JCI81660
47. Denorme, F, Portier, I, Rustad, JL, Cody, MJ, de Araujo, CV, Hoki, C, et al. Neutrophil extracellular traps regulate ischemic stroke brain injury. J Clin Invest. (2022) 132:e154225. doi: 10.1172/JCI154225
48. van der Maten, E, de Bont, CM, de Groot, R, de Jonge, MI, Langereis, JD, and van der Flier, M. Alternative pathway regulation by factor H modulates Streptococcus pneumoniae induced proinflammatory cytokine responses by decreasing C5a receptor crosstalk. Cytokine. (2016) 88:281–6. doi: 10.1016/j.cyto.2016.09.025
49. Manfredi, AA, Ramirez, GA, Godino, C, Capobianco, A, Monno, A, Franchini, S, et al. Platelet phagocytosis via P-selectin glycoprotein ligand 1 and accumulation of microparticles in systemic sclerosis. Arthritis Rheumatol. (2022) 74:318–28. doi: 10.1002/art.41926
50. Garishah, FM, Rother, N, Riswari, SF, Alisjahbana, B, Overheul, GJ, van Rij, RP, et al. Neutrophil extracellular traps in dengue are mainly generated NOX-independently. Front Immunol. (2021) 12:629167. doi: 10.3389/fimmu.2021.629167
51. Poli, V, di Gioia, M, Sola-Visner, M, Granucci, F, Frelinger, AL III, Michelson, AD, et al. Inhibition of transcription factor NFAT activity in activated platelets enhances their aggregation and exacerbates gram-negative bacterial septicemia. Immunity. (2022) 55:224–236.e5. doi: 10.1016/j.immuni.2021.12.002
52. Ren, J, He, J, Zhang, H, Xia, Y, Hu, Z, Loughran, P, et al. Platelet TLR4-ERK5 Axis facilitates NET-mediated capturing of circulating tumor cells and distant metastasis after surgical stress. Cancer Res. (2021) 81:2373–85. doi: 10.1158/0008-5472.CAN-20-3222
53. Li, JC, Zou, XM, Yang, SF, Jin, JQ, Zhu, L, Li, CJ, et al. Neutrophil extracellular traps participate in the development of cancer-associated thrombosis in patients with gastric cancer. World J Gastroenterol. (2022) 28:3132–49. doi: 10.3748/wjg.v28.i26.3132
54. Pircher, J, Czermak, T, Ehrlich, A, Eberle, C, Gaitzsch, E, Margraf, A, et al. Cathelicidins prime platelets to mediate arterial thrombosis and tissue inflammation. Nat Commun. (2018) 9:1523. doi: 10.1038/s41467-018-03925-2
55. Denorme, F, Rustad, JL, and Campbell, RA. Brothers in arms: platelets and neutrophils in ischemic stroke. Curr Opin Hematol. (2021) 28:301–7. doi: 10.1097/MOH.0000000000000665
56. Petzold, T, Zhang, Z, Ballesteros, I, Saleh, I, Polzin, A, Thienel, M, et al. Neutrophil "plucking" on megakaryocytes drives platelet production and boosts cardiovascular disease. Immunity. (2022) 55:2285–2299.e7. doi: 10.1016/j.immuni.2022.10.001
57. Letsiou, E, Teixeira Alves, LG, Felten, M, Mitchell, TJ, Müller-Redetzky, HC, Dudek, SM, et al. Neutrophil-derived extracellular vesicles activate platelets after Pneumolysin exposure. Cells. (2021) 10:3581. doi: 10.3390/cells10123581
58. Joshi, A, Schmidt, LE, Burnap, SA, Lu, R, Chan, MV, Armstrong, PC, et al. Neutrophil-derived protein S100A8/A9 alters the platelet proteome in acute myocardial infarction and is associated with changes in platelet reactivity. Arterioscler Thromb Vasc Biol. (2022) 42:49–62. doi: 10.1161/ATVBAHA.121.317113
59. Singh, MV, Davidson, DC, Kiebala, M, and Maggirwar, SB. Detection of circulating platelet-monocyte complexes in persons infected with human immunodeficiency virus type-1. J Virol Methods. (2012) 181:170–6. doi: 10.1016/j.jviromet.2012.02.005
60. Fu, G, Deng, M, Neal, MD, Billiar, TR, and Scott, MJ. Platelet-monocyte aggregates: understanding mechanisms and functions in sepsis. Shock. (2021) 55:156–66. doi: 10.1097/SHK.0000000000001619
61. Vinci, R, Pedicino, D, D’Aiello, A, Ciampi, P, Ponzo, M, Bonanni, A, et al. Platelet hyaluronidase 2 enrichment in acute coronary syndromes: a conceivable role in monocyte-platelet aggregate formation. J Enzyme Inhib Med Chem. (2021) 36:785–9. doi: 10.1080/14756366.2021.1900159
62. Manne, BK, Denorme, F, Middleton, EA, Portier, I, Rowley, JW, Stubben, C, et al. Platelet gene expression and function in patients with COVID-19. Blood. (2020) 136:1317–29. doi: 10.1182/blood.2020007214
63. Hottz, ED, Azevedo-Quintanilha, IG, Palhinha, L, Teixeira, L, Barreto, EA, Pão, CRR, et al. Platelet activation and platelet-monocyte aggregate formation trigger tissue factor expression in patients with severe COVID-19. Blood. (2020) 136:1330–41. doi: 10.1182/blood.2020007252
64. Hottz, ED, Martins-Gonçalves, R, Palhinha, L, Azevedo-Quintanilha, IG, de Campos, MM, Sacramento, CQ, et al. Platelet-monocyte interaction amplifies thromboinflammation through tissue factor signaling in COVID-19. Blood Adv. (2022) 6:5085–99. doi: 10.1182/bloodadvances.2021006680
65. Collier, MEW, Ambrose, AR, and Goodall, AH. Does hsa-miR-223-3p from platelet-derived extracellular vesicles regulate tissue factor expression in monocytic cells? Platelets. (2022) 33:1031–42. doi: 10.1080/09537104.2022.2027903
66. Hottz, ED, Medeiros-de-Moraes, IM, Vieira-de-Abreu, A, de Assis, EF, Vals-de-Souza, R, Castro-Faria-Neto, HC, et al. Platelet activation and apoptosis modulate monocyte inflammatory responses in dengue. J Immunol. (2014) 193:1864–72. doi: 10.4049/jimmunol.1400091
67. Weyrich, AS, Elstad, MR, McEver, RP, McIntyre, TM, Moore, KL, Morrissey, JH, et al. Activated platelets signal chemokine synthesis by human monocytes. J Clin Invest. (1996) 97:1525–34. doi: 10.1172/JCI118575
68. Rong, MY, Wang, CH, Wu, ZB, Zeng, W, Zheng, ZH, Han, Q, et al. Platelets induce a proinflammatory phenotype in monocytes via the CD147 pathway in rheumatoid arthritis. Arthritis Res Ther. (2014) 16:478. doi: 10.1186/s13075-014-0478-0
69. Gros, A, Syvannarath, V, Lamrani, L, Ollivier, V, Loyau, S, Goerge, T, et al. Single platelets seal neutrophil-induced vascular breaches via GPVI during immune-complex-mediated inflammation in mice. Blood. (2015) 126:1017–26. doi: 10.1182/blood-2014-12-617159
70. Clark, SR, Ma, AC, Tavener, SA, McDonald, B, Goodarzi, Z, Kelly, MM, et al. Platelet TLR4 activates neutrophil extracellular traps to ensnare bacteria in septic blood. Nat Med. (2007) 13:463–9. doi: 10.1038/nm1565
71. Armstrong, PC, Kirkby, NS, Chan, MV, Finsterbusch, M, Hogg, N, Nourshargh, S, et al. Novel whole blood assay for phenotyping platelet reactivity in mice identifies ICAM-1 as a mediator of platelet-monocyte interaction. Blood. (2015) 126:e11–8. doi: 10.1182/blood-2015-01-621656
72. Diacovo, TG, Roth, SJ, Buccola, JM, Bainton, DF, and Springer, TA. Neutrophil rolling, arrest, and transmigration across activated, surface-adherent platelets via sequential action of P-selectin and the beta 2-integrin CD11b/CD18. Blood. (1996) 88:146–57. doi: 10.1182/blood.V88.1.146.146
73. Stone, PCW, and Nash, GB. Conditions under which immobilized platelets activate as well as capture flowing neutrophils. Br J Haematol. (1999) 105:514–22. doi: 10.1111/j.1365-2141.1999.01375.x
74. Kossmann, H, Rischpler, C, Hanus, F, Nekolla, SG, Kunze, KP, Götze, K, et al. Monocyte-platelet aggregates affect local inflammation in patients with acute myocardial infarction. Int J Cardiol. (2019) 287:7–12. doi: 10.1016/j.ijcard.2019.04.009
75. Li, TY, Yang, Y, Li, Y, Wang, Z, Ma, F, Luo, R, et al. Platelets mediate inflammatory monocyte activation by SARS-CoV-2 spike protein. J Clin Investig. (2022) 132:e150101. doi: 10.1172/JCI150101
76. Suzuki, J, Hamada, E, Shodai, T, Kamoshida, G, Kudo, S, Itoh, S, et al. Cytokine secretion from human monocytes potentiated by P-selectin-mediated cell adhesion. Int Arch Allergy Immunol. (2013) 160:152–60. doi: 10.1159/000339857
77. Christersson, C, Johnell, M, and Siegbahn, A. Tissue factor and IL8 production by P-selectin-dependent platelet-monocyte aggregates in whole blood involves phosphorylation of Lyn and is inhibited by IL10. J Thromb Haemost. (2008) 6:986–94. doi: 10.1111/j.1538-7836.2008.02956.x
78. Quach, HQ, Johnson, C, Ekholt, K, Islam, R, Mollnes, TE, and Nilsson, PH. Platelet-depletion of whole blood reveals that platelets potentiate the release of IL-8 from leukocytes into plasma in a thrombin-dependent manner. Front Immunol. (2022) 13:865386. doi: 10.3389/fimmu.2022.865386
79. Loguinova, M, Pinegina, N, Kogan, V, Vagida, M, Arakelyan, A, Shpektor, A, et al. Monocytes of different subsets in complexes with platelets in patients with myocardial infarction. Thromb Haemost. (2018) 118:1969–81. doi: 10.1055/s-0038-1673342
80. Gudbrandsdottir, S, Hasselbalch, HC, and Nielsen, CH. Activated platelets enhance IL-10 secretion and reduce TNF-alpha secretion by monocytes. J Immunol. (2013) 191:4059–67. doi: 10.4049/jimmunol.1201103
81. Badrnya, S, Schrottmaier, WC, Kral, JB, Yaiw, KC, Volf, I, Schabbauer, G, et al. Platelets mediate oxidized low-density lipoprotein-induced monocyte extravasation and foam cell formation. Arterioscler Thromb Vasc Biol. (2014) 34:571–80. doi: 10.1161/ATVBAHA.113.302919
82. Phillips, JH, Chang, CW, and Lanier, LL. Platelet-induced expression of fc gamma RIII (CD16) on human monocytes. Eur J Immunol. (1991) 21:895–9. doi: 10.1002/eji.1830210406
83. Scheuerer, B, Ernst, M, Dürrbaum-Landmann, I, Fleischer, J, Grage-Griebenow, E, Brandt, E, et al. The CXC-chemokine platelet factor 4 promotes monocyte survival and induces monocyte differentiation into macrophages. Blood. (2000) 95:1158–66. doi: 10.1182/blood.V95.4.1158.004k31_1158_1166
84. Stephen, J, Emerson, B, Fox, KAA, and Dransfield, I. The uncoupling of monocyte-platelet interactions from the induction of Proinflammatory signaling in monocytes. J Immunol. (2013) 191:5677–83. doi: 10.4049/jimmunol.1301250
85. Hilt, ZT, Pariser, DN, Ture, SK, Mohan, A, Quijada, P, Asante, AA, et al. Platelet-derived beta 2M regulates monocyte inflammatory responses. Jci Insight. (2019) 4:e122943. doi: 10.1172/jci.insight.122943
86. Hilt, ZT, et al. Platelet-derived beta 2m regulates age related monocyte/macrophage functions. Aging-Us. (2019) 11:11955–74. doi: 10.18632/aging.102520
87. Okubo, K, Kurosawa, M, Kamiya, M, Urano, Y, Suzuki, A, Yamamoto, K, et al. Macrophage extracellular trap formation promoted by platelet activation is a key mediator of rhabdomyolysis-induced acute kidney injury. Nat Med. (2018) 24:232–8. doi: 10.1038/nm.4462
88. Heffron, SP, Weinstock, A, Scolaro, B, Chen, S, Sansbury, BE, Marecki, G, et al. Platelet-conditioned media induces an anti-inflammatory macrophage phenotype through EP4. J Thromb Haemost. (2021) 19:562–73. doi: 10.1111/jth.15172
89. Rossaint, J, Thomas, K, Mersmann, S, Skupski, J, Margraf, A, Tekath, T, et al. Platelets orchestrate the resolution of pulmonary inflammation in mice by T reg cell repositioning and macrophage education. J Exp Med. (2021) 218. doi: 10.1084/jem.20201353
90. Uchiyama, R, Toyoda, E, Maehara, M, Wasai, S, Omura, H, Watanabe, M, et al. Effect of platelet-rich plasma on M1/M2 macrophage polarization. Int J Mol Sci. (2021) 22. doi: 10.3390/ijms22052336
91. Nasirzade, J, Kargarpour, Z, Hasannia, S, Strauss, FJ, and Gruber, R. Platelet-rich fibrin elicits an anti-inflammatory response in macrophages in vitro. J Periodontol. (2020) 91:244–52. doi: 10.1002/JPER.19-0216
92. Sordi, MB, Panahipour, L, Kargarpour, Z, and Gruber, R. Platelet-rich fibrin reduces IL-1beta release from macrophages undergoing Pyroptosis. Int J Mol Sci. (2022) 23:8306. doi: 10.3390/ijms23158306
93. Qian, J, Wang, X, Su, G, Shu, X, Huang, Z, Jiang, H, et al. Platelet-rich plasma-derived exosomes attenuate intervertebral disc degeneration by promoting NLRP3 autophagic degradation in macrophages. Int Immunopharmacol. (2022) 110:108962. doi: 10.1016/j.intimp.2022.108962
94. Nishio, H, Saita, Y, Kobayashi, Y, Takaku, T, Fukusato, S, Uchino, S, et al. Platelet-rich plasma promotes recruitment of macrophages in the process of tendon healing. Regen Ther. (2020) 14:262–70. doi: 10.1016/j.reth.2020.03.009
95. Carestia, A, Mena, HA, Olexen, CM, Ortiz Wilczyñski, JM, Negrotto, S, Errasti, AE, et al. Platelets promote macrophage polarization toward pro-inflammatory phenotype and increase survival of septic mice. Cell Rep. (2019) 28:896–908.e5. doi: 10.1016/j.celrep.2019.06.062
96. Sadallah, S, Schmied, L, Eken, C, Charoudeh, HN, Amicarella, F, and Schifferli, JA. Platelet-derived Ectosomes reduce NK cell function. J Immunol. (2016) 197:1663–71. doi: 10.4049/jimmunol.1502658
97. Placke, T, Kopp, HG, and Salih, HR. Modulation of natural killer cell anti-tumor reactivity by platelets. J Innate Immun. (2011) 3:374–82. doi: 10.1159/000323936
98. Kopp, HG, Placke, T, and Salih, HR. Platelet-derived transforming growth factor-beta down-regulates NKG2D thereby inhibiting natural killer cell antitumor reactivity. Cancer Res. (2009) 69:7775–83. doi: 10.1158/0008-5472.CAN-09-2123
99. Du, Y, Liu, X, and Guo, SW. Platelets impair natural killer cell reactivity and function in endometriosis through multiple mechanisms. Hum Reprod. (2017) 32:794–810. doi: 10.1093/humrep/dex014
100. Gockel, LM, Nekipelov, K, Ferro, V, Bendas, G, and Schlesinger, M. Tumour cell-activated platelets modulate the immunological activity of CD4(+), CD8(+), and NK cells, which is efficiently antagonized by heparin. Cancer Immunol Immunother. (2022) 71:2523–33. doi: 10.1007/s00262-022-03186-5
101. Sun, Y, Sedgwick, AJ, Palarasah, Y, Mangiola, S, and Barrow, AD. A transcriptional signature of PDGF-DD activated natural killer cells predicts more favorable prognosis in low-grade Glioma. Front Immunol. (2021) 12:668391. doi: 10.3389/fimmu.2021.724107
102. Ma, S, Tang, T, Wu, X, Mansour, AG, Lu, T, Zhang, J, et al. PDGF-D-PDGFRbeta signaling enhances IL-15-mediated human natural killer cell survival. Proc Natl Acad Sci U S A. (2022) 119:e2114134119. doi: 10.1073/pnas.2114134119
103. Martí, F, Bertran, E, Llucià, M, Villén, E, Peiró, M, Garcia, J, et al. Platelet factor 4 induces human natural killer cells to synthesize and release interleukin-8. J Leukoc Biol. (2002) 72:590–7. doi: 10.1189/jlb.72.3.590
104. Gerdes, N, Zhu, L, Ersoy, M, Hermansson, A, Hjemdahl, P, Hu, H, et al. Platelets regulate CD4(+) T-cell differentiation via multiple chemokines in humans. Thromb Haemost. (2011) 106:353–62. doi: 10.1160/TH11-01-0020
105. Ahmad, R, Menezes, J, Knafo, L, and Ahmad, A. Activated human platelets express Fas-L and induce apoptosis in Fas-positive tumor cells. J Leukoc Biol. (2001) 69:123–8. doi: 10.1189/jlb.69.1.123
106. Crist, SA, Elzey, BD, Ludwig, AT, Griffith, TS, Staack, JB, Lentz, SR, et al. Expression of TNF-related apoptosis-inducing ligand (TRAIL) in megakaryocytes and platelets. Exp Hematol. (2004) 32:1073–81. doi: 10.1016/j.exphem.2004.07.022
107. Elzey, BD, Sprague, DL, and Ratliff, TL. The emerging role of platelets in adaptive immunity. Cell Immunol. (2005) 238:1–9. doi: 10.1016/j.cellimm.2005.12.005
108. Elzey, BD, Schmidt, NW, Crist, SA, Kresowik, TP, Harty, JT, Nieswandt, B, et al. Platelet-derived CD154 enables T-cell priming and protection against listeria monocytogenes challenge. Blood. (2008) 111:3684–91. doi: 10.1182/blood-2007-05-091728
109. Iannacone, M, Sitia, G, Isogawa, M, Whitmire, JK, Marchese, P, Chisari, FV, et al. Platelets prevent IFN-alpha/beta-induced lethal hemorrhage promoting CTL-dependent clearance of lymphocytic choriomeningitis virus. Proc Natl Acad Sci U S A. (2008) 105:629–34. doi: 10.1073/pnas.0711200105
110. Chapman, LM, Aggrey, AA, Field, DJ, Srivastava, K, Ture, S, Yui, K, et al. Platelets present antigen in the context of MHC class I. J Immunol. (2012) 189:916–23. doi: 10.4049/jimmunol.1200580
111. Lang, PA, Contaldo, C, Georgiev, P, el-Badry, AM, Recher, M, Kurrer, M, et al. Aggravation of viral hepatitis by platelet-derived serotonin. Nat Med. (2008) 14:756–61. doi: 10.1038/nm1780
112. Guo, L, Shen, S, Rowley, JW, Tolley, ND, Jia, W, Manne, BK, et al. Platelet MHC class I mediates CD8+ T-cell suppression during sepsis. Blood. (2021) 138:401–16. doi: 10.1182/blood.2020008958
113. Wu, Y, Zeng, Z, Guo, Y, Song, L, Weatherhead, JE, Huang, X, et al. Candida albicans elicits protective allergic responses via platelet mediated T helper 2 and T helper 17 cell polarization. Immunity. (2021) 54:2595–2610.e7. doi: 10.1016/j.immuni.2021.08.009
114. la Manna, MP, Orlando, V, Badami, GD, Tamburini, B, Shekarkar Azgomi, M, Lo Presti, E, et al. Platelets accumulate in lung lesions of tuberculosis patients and inhibit T-cell responses and mycobacterium tuberculosis replication in macrophages. Eur J Immunol. (2022) 52:784–99. doi: 10.1002/eji.202149549
115. Schuhmann, MK, Stoll, G, Bieber, M, Vögtle, T, Hofmann, S, Klaus, V, et al. CD84 links T cell and platelet activity in cerebral Thrombo-inflammation in acute stroke. Circ Res. (2020) 127:1023–35. doi: 10.1161/CIRCRESAHA.120.316655
116. Scherlinger, M, Guillotin, V, Douchet, I, Vacher, P, Boizard-Moracchini, A, Guegan, J-P, et al. Erratum for the research article: Selectins impair regulatory T cell function and contribute to systemic lupus erythematosus pathogenesis. Sci Transl Med. (2021) 13:eabl7656. doi: 10.1126/scitranslmed.abl7656
117. Rachidi, S, Metelli, A, Riesenberg, B, Wu, BX, Nelson, MH, Wallace, C, et al. Platelets subvert T cell immunity against cancer via GARP-TGFbeta axis. Sci Immunol. (2017) 2:eaai7911. doi: 10.1126/sciimmunol.aai7911
118. Hinterleitner, C, Strähle, J, Malenke, E, Hinterleitner, M, Henning, M, Seehawer, M, et al. Platelet PD-L1 reflects collective intratumoral PD-L1 expression and predicts immunotherapy response in non-small cell lung cancer. Nat Commun. (2021) 12:7005. doi: 10.1038/s41467-021-27303-7
119. Tan, S, Li, S, Min, Y, Gisterå, A, Moruzzi, N, Zhang, J, et al. Platelet factor 4 enhances CD4(+) T effector memory cell responses via Akt-PGC1alpha-TFAM signaling-mediated mitochondrial biogenesis. J Thromb Haemost. (2020) 18:2685–700. doi: 10.1111/jth.15005
120. Min, Y, Hao, L, Liu, X, Tan, S, Song, H, Ni, H, et al. Platelets fine-tune effector responses of naive CD4(+) T cells via platelet factor 4-regulated transforming growth factor beta signaling. Cell Mol Life Sci. (2022) 79:247. doi: 10.1007/s00018-022-04279-1
121. Tan, S, Zhang, J, Sun, Y, Gisterå, A, Sheng, Z, Malmström, RE, et al. Platelets enhance CD4+ central memory T cell responses via platelet factor 4-dependent mitochondrial biogenesis and cell proliferation. Platelets. (2022) 33:360–70. doi: 10.1080/09537104.2021.1936479
122. Yu, H, Hu, W, Song, X, and Zhao, Y. Immune modulation of platelet-derived mitochondria on memory CD4(+) T cells in humans. Int J Mol Sci. (2020) 21:6295. doi: 10.3390/ijms21176295
123. Mulet, M, Zamora, C, Porcel, JM, Nieto, JC, Pajares, V, Muñoz-Fernandez, AM, et al. Platelet factor 4 regulates T cell effector functions in malignant pleural effusions. Cancer Lett. (2020) 491:78–86. doi: 10.1016/j.canlet.2020.06.014
124. Chen, X, Xu, Y, Chen, Q, Zhang, H, Zeng, Y, Geng, Y, et al. The phosphatase PTEN links platelets with immune regulatory functions of mouse T follicular helper cells. Nat Commun. (2022) 13:2762. doi: 10.1038/s41467-022-30444-y
125. Polasky, C, Wendt, F, Pries, R, and Wollenberg, B. Platelet induced functional alteration of CD4(+) and CD8(+) T cells in HNSCC. Int J Mol Sci. (2020) 21:7507. doi: 10.3390/ijms21207507
126. Zhang, X, Liu, H, Tang, N, Zhou, S, Yu, J, Li, B, et al. Engineered PD-L1-expressing platelets reverse new-onset type 1 diabetes. Adv Mater. (2020) 32:e1907692. doi: 10.1002/adma.202006930
127. Paletta, A, di Diego García, F, Varese, A, Erra Diaz, F, García, J, Cisneros, JC, et al. Platelets modulate CD4(+) T-cell function in COVID-19 through a PD-L1 dependent mechanism. Br J Haematol. (2022) 197:283–92. doi: 10.1111/bjh.18062
128. Luo, L, Zhang, J, Lee, J, and Tao, A. Platelets, not an insignificant player in development of allergic asthma. Cells. (2021) 10. doi: 10.3390/cells10082038
129. Dai, XP, Wu, FY, Cui, C, Liao, XJ, Jiao, YM, Zhang, C, et al. Increased platelet-CD4(+) T cell aggregates are correlated with HIV-1 permissiveness and CD4(+) T cell loss. Front Immunol. (2021) 12:799124. doi: 10.3389/fimmu.2021.799124
130. Zhu, A, Real, F, Zhu, J, Greffe, S, de Truchis, P, Rouveix, E, et al. HIV-sheltering platelets from immunological non-responders induce a dysfunctional glycolytic CD4(+) T-cell profile. Front Immunol. (2021) 12:781923. doi: 10.3389/fimmu.2021.781923
131. Elzey, BD, Tian, J, Jensen, RJ, Swanson, AK, Lees, JR, Lentz, SR, et al. Platelet-mediated modulation of adaptive immunity: a communication link between innate and adaptive immune compartments. Immunity. (2003) 19:9–19. doi: 10.1016/S1074-7613(03)00177-8
132. Elzey, BD, Grant, JF, Sinn, HW, Nieswandt, B, Waldschmidt, TJ, and Ratliff, TL. Cooperation between platelet-derived CD154 and CD4(+) T cells for enhanced germinal center formation. J Leukoc Biol. (2005) 78:80–4. doi: 10.1189/jlb.1104669
133. Cognasse, F, Hamzeh-Cognasse, H, Lafarge, S, Chavarin, P, Cogné, M, Richard, Y, et al. Human platelets can activate peripheral blood B cells and increase production of immunoglobulins. Exp Hematol. (2007) 35:1376–87. doi: 10.1016/j.exphem.2007.05.021
134. Kasuya, A, Tokura, Y, and Honda, T. CD40L from platelet, endothelial cell, and smooth muscle cell may contribute to the proliferation of tumor cell in intravascular large B cell lymphoma. J Dermatol. (2021) 48:e180–1. doi: 10.1111/1346-8138.15786
135. Field, DJ, Aggrey-Amable, AA, Blick, SK, Ture, SK, Johanson, A, Cameron, SJ, et al. Platelet factor 4 increases bone marrow B cell development and differentiation. Immunol Res. (2017) 65:1089–94. doi: 10.1007/s12026-017-8951-x
136. Pavord, S, Scully, M, Hunt, BJ, Lester, W, Bagot, C, Craven, B, et al. Clinical features of vaccine-induced immune thrombocytopenia and thrombosis. N Engl J Med. (2021) 385:1680–9. doi: 10.1056/NEJMoa2109908
137. Klok, FA, Pai, M, Huisman, MV, and Makris, M. Vaccine-induced immune thrombotic thrombocytopenia. Lancet Haematol. (2022) 9:e73–80. doi: 10.1016/S2352-3026(21)00306-9
138. Greinacher, A, Selleng, K, Palankar, R, Wesche, J, Handtke, S, Wolff, M, et al. Insights in ChAdOx1 nCoV-19 vaccine-induced immune thrombotic thrombocytopenia. Blood. (2021) 138:2256–68. doi: 10.1182/blood.2021013231
139. Greinacher, A., Platelet fc receptors for IgG and IgE. in The immunoglobulin receptors and their physiological and pathological roles in immunity, Winkel, J.G.J.Van De, and Hogarth, P.M., Editors. (1998), Springer Netherlands: Dordrecht. p. 119–134.
140. Beutier, H, Hechler, B, Godon, O, Wang, Y, Gillis, CM, de Chaisemartin, L, et al. Platelets expressing IgG receptor FcgammaRIIA/CD32A determine the severity of experimental anaphylaxis. Sci Immunol. (2018) 3:eaan5997. doi: 10.1126/sciimmunol.aan5997
141. Joseph, M, Capron, A, Ameisen, JC, Capron, M, Vorng, H, Pancré, V, et al. The receptor for IgE on blood platelets. Eur J Immunol. (1986) 16:306–12. doi: 10.1002/eji.1830160318
142. Joseph, M, Gounni, AS, Kusnierz, JP, Vorng, H, Sarfati, M, Kinet, JP, et al. Expression and functions of the high-affinity IgE receptor on human platelets and megakaryocyte precursors. Eur J Immunol. (1997) 27:2212–8. doi: 10.1002/eji.1830270914
143. Saris, A, Steuten, J, Schrijver, DP, van Schijndel, G, Zwaginga, JJ, van Ham, SM, et al. Inhibition of dendritic cell activation and modulation of T cell polarization by the platelet Secretome. Front Immunol. (2021) 12:631285. doi: 10.3389/fimmu.2021.631285
144. Ki, KK, Faddy, HM, Flower, RL, and Dean, MM. Platelet concentrates modulate myeloid dendritic cell immune responses. Platelets. (2018) 29:373–82. doi: 10.1080/09537104.2017.1306045
145. Ki, KK, Johnson, L, Faddy, HM, Flower, RL, Marks, DC, and Dean, MM. Immunomodulatory effect of cryopreserved platelets: altered BDCA3(+) dendritic cell maturation and activation in vitro. Transfusion. (2017) 57:2878–87. doi: 10.1111/trf.14320
146. Singh, MV, Suwunnakorn, S, Simpson, SR, Weber, EA, Singh, VB, Kalinski, P, et al. Monocytes complexed to platelets differentiate into functionally deficient dendritic cells. J Leukoc Biol. (2021) 109:807–20. doi: 10.1002/JLB.3A0620-460RR
147. Nishat, S, Wuescher, LM, and Worth, RG. Platelets enhance dendritic cell responses against Staphylococcus aureus through CD40-CD40L. Infect Immun. (2018) 86:e00186. doi: 10.1128/IAI.00186-18
148. Nakanishi, T, Inaba, M, Inagaki-Katashiba, N, Tanaka, A, Vien, PTX, Kibata, K, et al. Platelet-derived RANK ligand enhances CCL17 secretion from dendritic cells mediated by thymic stromal lymphopoietin. Platelets. (2015) 26:425–31. doi: 10.3109/09537104.2014.920081
149. Amison, RT, Cleary, SJ, Riffo-Vasquez, Y, Bajwa, M, Page, CP, and Pitchford, SC. Platelets play a central role in sensitization to allergen. Am J Respir Cell Mol Biol. (2018) 59:96–103. doi: 10.1165/rcmb.2017-0401OC
150. Han, P, Han, D, Arshad, N, Lee, JS, Tatsuno, K, Robinson, E, et al. Platelet P-selectin initiates cross-presentation and dendritic cell differentiation in blood monocytes. Sci Adv. (2021) 7:eaaz1580. doi: 10.1126/sciadv.abl8894
151. Nording, H, Sauter, M, Lin, C, Steubing, R, Geisler, S, Sun, Y, et al. Activated platelets Upregulate beta2 integrin mac-1 (CD11b/CD18) on dendritic cells, which mediates heterotypic cell-cell interaction. J Immunol. (2022) 208:1729–41. doi: 10.4049/jimmunol.2100557
152. Martinson, JA, Bae, J, Klingemann, HG, and Tam, YK. Activated platelets rapidly up-regulate CD40L expression and can effectively mature and activate autologous ex vivo differentiated DC. Cytotherapy. (2004) 6:487–97. doi: 10.1080/14653240410005249-1
153. Czapiga, M, Kirk, AD, and Lekstrom-Himes, J. Platelets deliver costimulatory signals to antigen-presenting cells: a potential bridge between injury and immune activation. Exp Hematol. (2004) 32:135–9. doi: 10.1016/j.exphem.2003.11.004
154. Duffau, P, Seneschal, J, Nicco, C, Richez, C, Lazaro, E, Douchet, I, et al. Platelet CD154 potentiates interferon-alpha secretion by Plasmacytoid dendritic cells in systemic lupus erythematosus. Sci Transl Med. (2010) 2:47ra63. doi: 10.1126/scitranslmed.3001001
155. Katoh, N, Soga, F, Nara, T, Tamagawa-Mineoka, R, Nin, M, Kotani, H, et al. Effect of serotonin on the differentiation of human monocytes into dendritic cells. Clin Exp Immunol. (2006) 146:354–61. doi: 10.1111/j.1365-2249.2006.03197.x
156. Xia, CQ, and Kao, KJ. Effect of CXC chemokine platelet factor 4 on differentiation and function of monocyte-derived dendritic cells. Int Immunol. (2003) 15:1007–15. doi: 10.1093/intimm/dxg100
Keywords: platelet, monocyte, macrophage, T cell, B cell, dendritic cell
Citation: Yan C, Wu H, Fang X, He J and Zhu F (2023) Platelet, a key regulator of innate and adaptive immunity. Front. Med. 10:1074878. doi: 10.3389/fmed.2023.1074878
Edited by:
Xiao He, The University of Utah, United StatesReviewed by:
Demba Sarr, University of Georgia, United StatesCopyright © 2023 Yan, Wu, Fang, He and Zhu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Feng Zhu, OTI0MjQzMTQ2QHFxLmNvbQ==
†These authors have contributed equally to this work
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.