
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Med. , 18 October 2022
Sec. Nephrology
Volume 9 - 2022 | https://doi.org/10.3389/fmed.2022.988080
This article is part of the Research Topic Recent Advances on Omics and Biomarkers Research in Renal Transplantation View all 6 articles
 Dalia Raïch-Regué1,2
Dalia Raïch-Regué1,2 Javier Gimeno1,3
Javier Gimeno1,3 Laura Llinàs-Mallol1,2Silvia Menéndez1,3David Benito1,2
Laura Llinàs-Mallol1,2Silvia Menéndez1,3David Benito1,2 Dolores Redondo1,2
Dolores Redondo1,2 M. José Pérez-Sáez1,2
M. José Pérez-Sáez1,2 Marta Riera1,2
Marta Riera1,2 Elaine F. Reed4
Elaine F. Reed4 Julio Pascual1,2*
Julio Pascual1,2* Marta Crespo1,2*
Marta Crespo1,2*Antibody-mediated rejection (ABMR) caused by donor-specific HLA-antibodies (DSA) is a mediator of allograft loss after kidney transplantation (KT). DSA can activate microvascular endothelium damage through the mTOR pathway. In this study we assessed the mTOR pathway activation by DSA in KT with ABMR (ABMR + DSA+) compared to controls (ABMR−DSA−), biopsies with ABMR changes without DSA (ABMR + DSA−) and DSA without ABMR changes (ABMR−DSA+), and the potential modulation by mTOR inhibitors (mTORi). We evaluated 97 biopsies: 31 ABMR + DSA+, 33 controls ABMR-DSA−, 16 ABMR + DSA−, and 17 ABMR-DSA+ cases. Regarding immunosuppression of full ABMR + DSA+ and controls, 21 biopsies were performed under mTORi treatment (11 of them ABMR + DSA+ cases) and 43 without mTORi (20 of them ABMR + DSA+) so as to explore its effect on the mTOR pathway. Biopsies were stained for C4d, Ki67, and phosphorylated (p) S6RP, ERK, and mTOR by immunohistochemistry. Labeling was graded according to peritubular capillary staining. ABMR biopsies showed significantly higher C4d, p-S6RP, and Ki67 staining in peritubular capillaries (PTC) compared to controls, and light differences in p-ERK or p-mTOR. mTORi treatment did not modify p-S6RP, p-mTOR, and p-ERK staining. Diffuse p-S6RP in PTC in the biopsies significantly associated with circulating HLA-DSA independently of graft rejection, and with worse death-censored graft survival. These findings suggest that activation of endothelium through the mTOR pathway evidence different mechanisms of damage in ABMR + DSA+ and ABMR + DSA− despite similar histological injury.
Antibody-mediated rejection (ABMR) is one of the leading causes of renal allograft loss (1). Antibodies against human leukocyte antigens (HLA) of the donor (DSA) are associated with poor allograft survival, frequently preceding this type of graft rejection (2, 3). ABMR is characterized by microvascular lesions in the form of inflammation and/or tissue remodeling in the presence of HLA-DSA (1, 4). Circulating HLA-DSA bind to graft endothelial cells and exert multiple effector functions that may produce damage, such as immune cell recruitment, complement activation or transduction of intracellular signals, leading to proliferation of graft vasculature (5). In fact, subsequent chronic transplant glomerulopathy and peritubular capillary basement membrane multilayering are diagnostic criteria for chronic ABMR. Understanding and documentation of these mechanisms of damage is paramount to diagnose, prevent and treat injury produced in ABMR.
Activation of complement in the form of C4d deposits in peritubular capillaries (PTC) was originally proposed as ABMR marker in renal transplant biopsies (6). Initially, C4d positivity was incorporated in the definition of ABMR in the Banff Classification (7), showing strong correlation with the presence of DSA (8). However, not all ABMR cases are C4d positive, as recognized by the Banff classification since 2013 (9), and there are also C4d-positive biopsies without evidence of rejection (10). Therefore, the implementation of new molecular diagnostic markers for ABMR may improve diagnosis and management of ABMR.
Vascular injury in ABMR is accompanied by endothelial cell activation (11). Microvascular endothelial cells express HLA constitutively (12), and expression of HLA class-II increases after transplantation (13). HLA class-II DSA are more strongly associated with microvascular injury and chronic endothelial lesions than HLA class-I DSA (14). Binding of anti-HLA class-I and –II antibodies to microvascular endothelial cells allows the recruitment of Integrin-β4 and other not well defined molecules, which transduce cytoplasmatic signals through the mammalian target of rapamycin (mTOR) pathway (15–17). mTOR is a protein kinase that forms two molecular complexes, 1 and 2, with distinct functional capacities. mTOR also integrates the input from other extracellular signals, such as insulin, growth factors, amino acids, and oxygen (18), and is a key regulator of cell growth, cell proliferation and survival, protein synthesis and autophagy (19). Indeed, mTOR was described to play important roles in different tissues, such as liver or brain, and to be dysregulated in human diseases like cancer or diabetes (20). In this line, up-regulated mTOR activity was found in active injury in both native and transplanted human kidneys (21). Furthermore, phosphorylation of the mTOR pathway proteins S6RP and 70S6K has been proposed as new ABMR markers on transplanted hearts (22, 23). Inhibitors of mTOR (mTORi; rapamycin or everolimus) have been used for two decades to prevent organ transplant rejection. In vitro, everolimus has shown to inhibit anti-HLA antibody-mediated signaling, migration and proliferation of endothelial cells (15, 24).
We hypothesized that detection of phosphorylated (p)-mTOR and its downstream signals S6RP and ERK in kidney allografts may be useful as diagnostic biomarkers of ABMR in the presence of circulating HLA-DSA, and the use of mTORi may modulate the pathway in vivo. Therefore, we evaluated the activation signals of the mTOR pathway: p-mTOR, p-S6RP and p-ERK, as well as microvascular endothelial proliferation in kidney allografts with histological criteria of ABMR with and without HLA-DSA, DSA without ABMR, and with normal samples. We also assessed the potential modulation of the mTOR pathway by mTORi immunosuppression in vivo.
We included 97 kidney transplant (KT) recipients with graft-biopsies performed between 2011 and 2015, around 24 months post-transplantation [IQR 14–56]. First, the study group comprised 31 patients diagnosed of ABMR with HLA-DSA (ABMR+DSA+), and 33 age-matched recipients with normal biopsies (ABMR–DSA–) according to (25). Of them, 21 patients received treatment with mTORi (20 Everolimus, 1 Rapamycin) at the time of biopsy (11 ABMR + DSA+) and 43 did not (20 ABMR + DSA+). Second, we included patients with an ABMR histological phenotype who lacked HLA-DSA (ABMR + DSA−, n = 16) and patients with circulating HLA-DSA without ABMR histological diagnosis (ABMR−DSA+, n = 17). Patient characteristics and clinical data are shown in Tables 1–4 and Supplementary Table 1. The study was approved by the CEIC Parc de Salut Mar Ethical Research Board (2018/7873I) and all patients signed informed consents. The clinical and research activities being reported are consistent with the Principles of the Declaration of Istanbul and the Declaration of Helsinki.
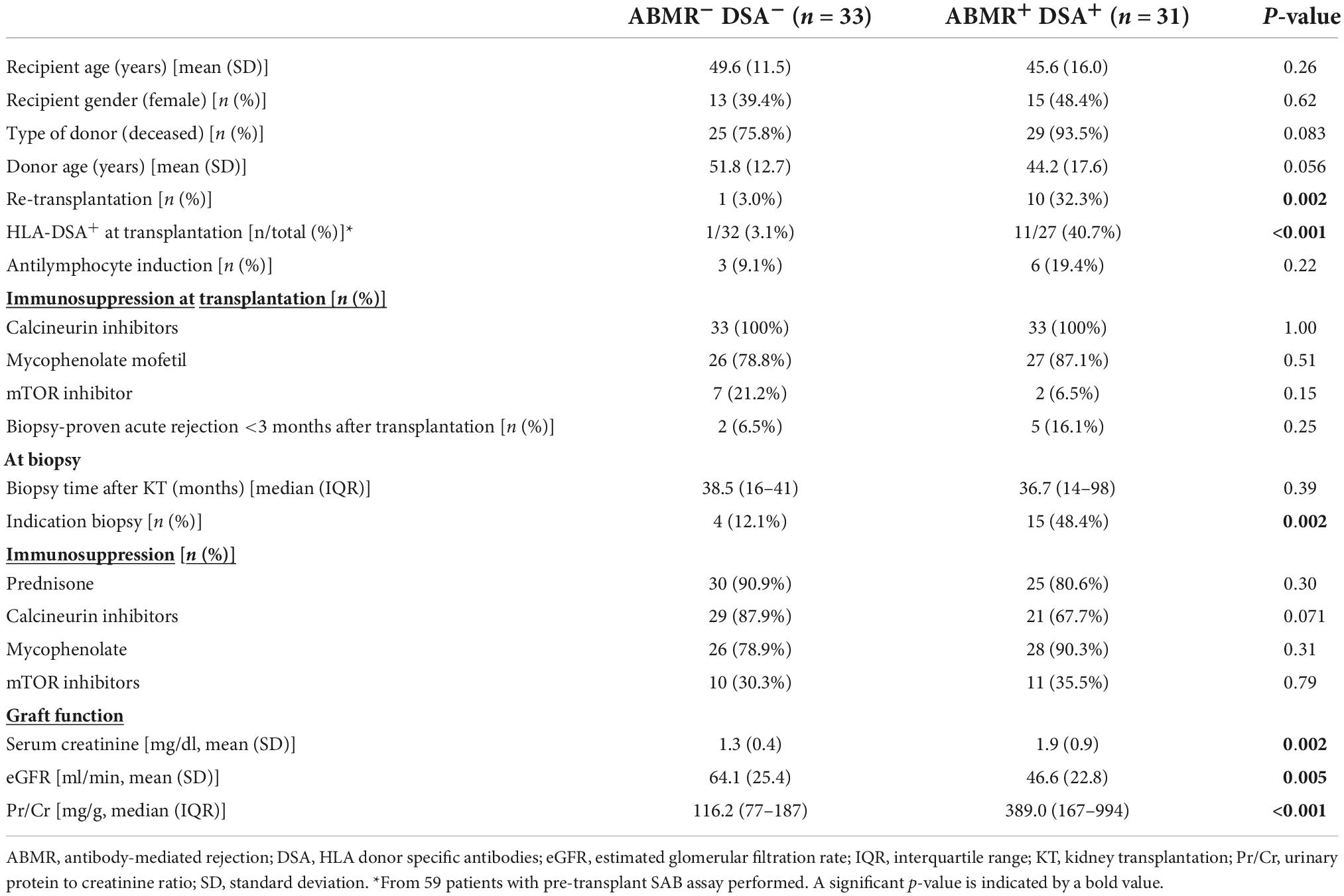
Table 1. Main demographic and clinical characteristics of patients with ABMR and HLA-DSA (ABMR+DSA+) and without ABMR and HLA-DSA (ABMR– DSA–).
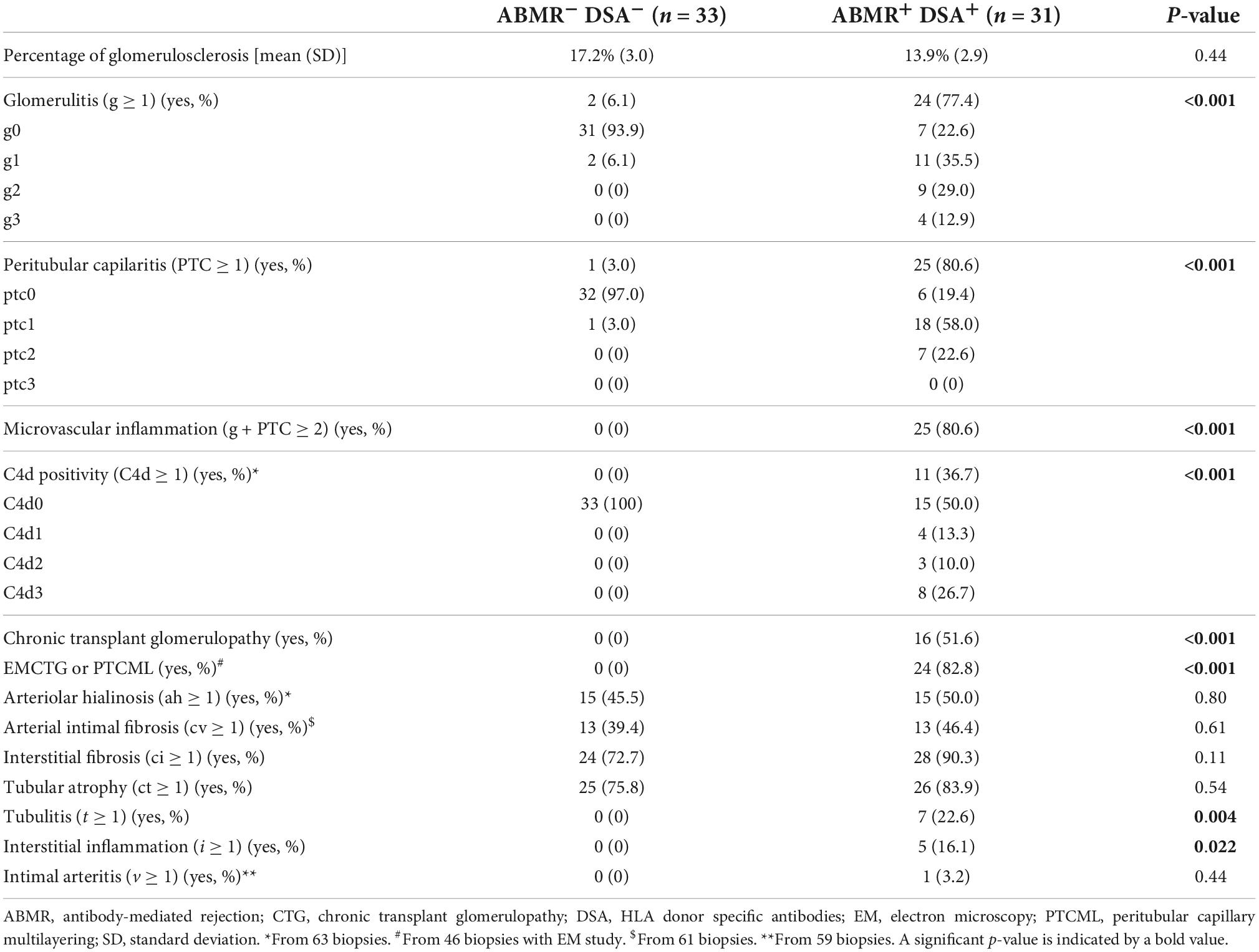
Table 2. Comparison of the histopathologic lesions in the biopsy of patients with ABMR and HLA-DSA (ABMR+DSA+) and without ABMR and HLA-DSA (ABMR– DSA–).
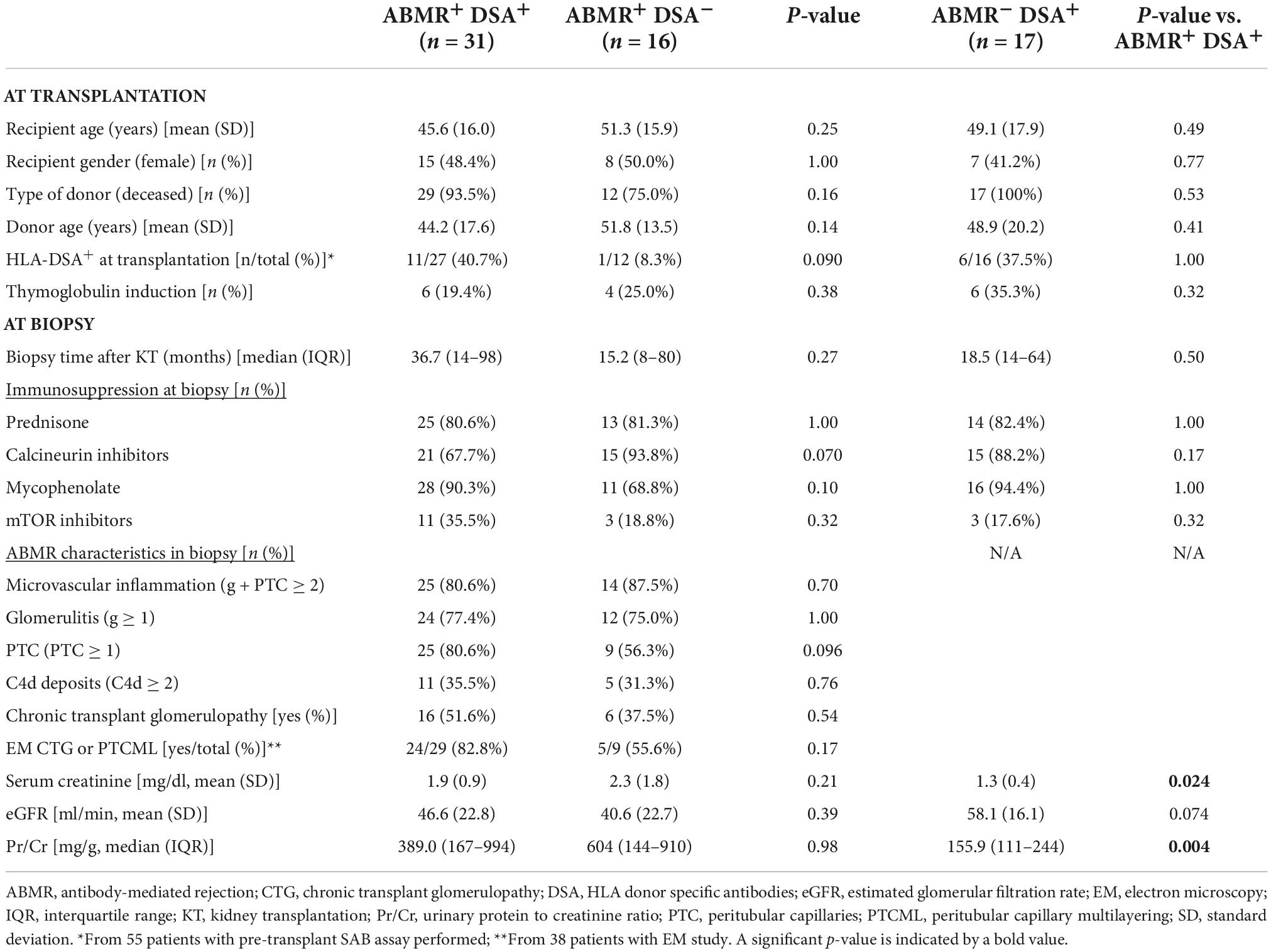
Table 3. Main demographic and clinical characteristics of patients with ABMR and DSA compared to ABMR without HLA-DSA (left) and to non-ABMR with HLA-DSA (right).
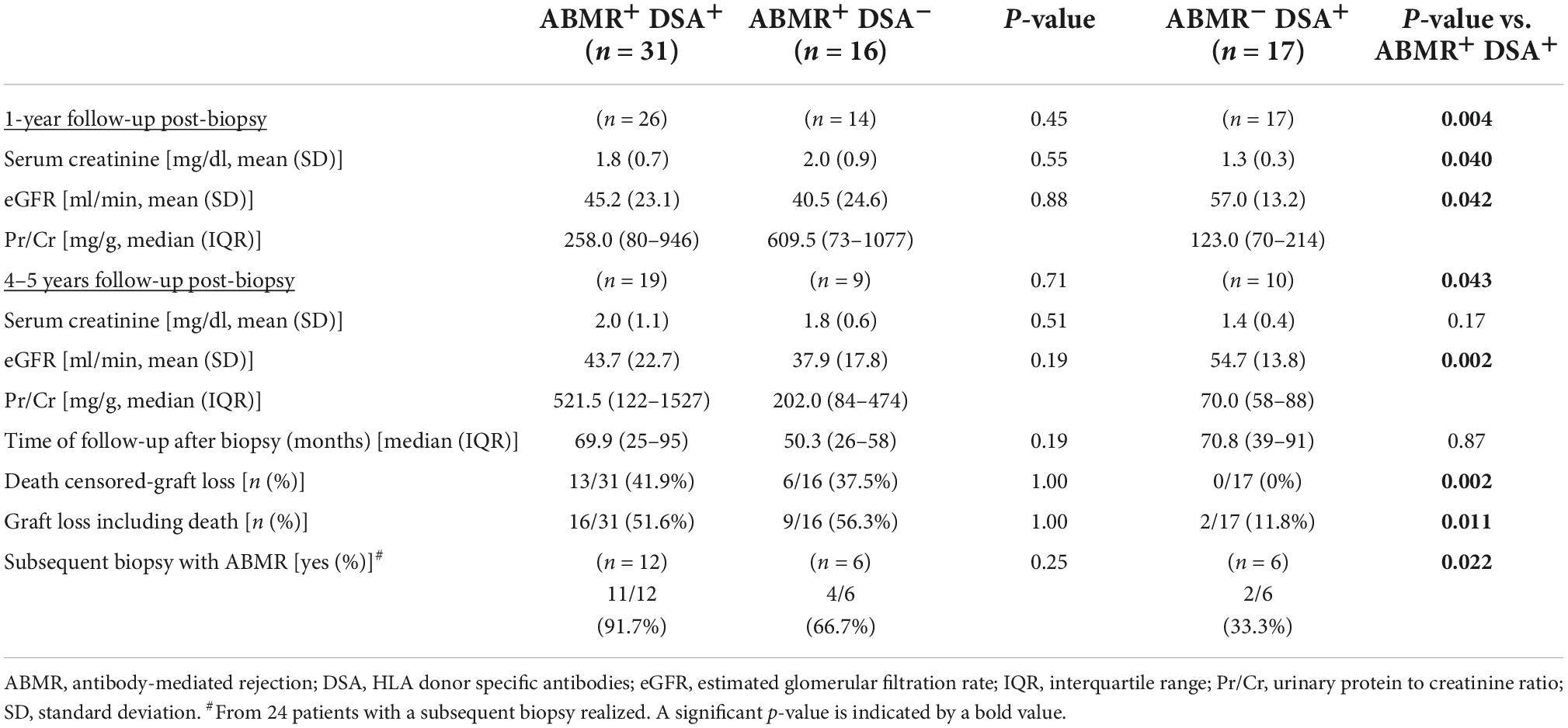
Table 4. Clinical follow-up of patients with ABMR and DSA compared to ABMR without HLA-DSA (left) and to non-ABMR with HLA-DSA (right).
Immunohistochemistry was performed on 3-μm paraffin allograft-biopsy sections, placed on plus-charged glass slides. After deparaffinization, heat antigen retrieval was performed in a pH = 9 EDTA-buffered solution (Dako, CA, United States). Endogenous peroxidase was quenched. Primary antibodies were incubated for 1 h at room temperature: anti-phospho-mTOR at Ser2448 (clone 49F9) rabbit mAb at 1:100, anti-phospho-S6 ribosomal protein at Ser235/236 (clone 91B2) rabbit mAb at 1:80, and anti-phospho-ERK1/2 (clone 20G11) rabbit mAb at 1:150, all from Cell Signaling Technology (Netherlands), and anti-Ki67 (clone MIB-1; Agilent, Dako) mAb at 1:100. Antigen–antibody reaction was detected by an anti-mouse/rabbit Ig–dextran-polymer coupled with peroxidase (Flex+, Dako). Sections were visualized with 3,3’-diaminobenzidine and counterstained with hematoxylin. Staining was performed on a Dako Autostainer platform. Protein expression was blindly assessed by one pathologist (J.G.).
Peritubular capillaries labeling was graded according to the staining intensity scale: 0 = none; 1 = rare; 2 = focal; 3 = multifocal/diffuse. A score ≥ 1 was considered positive for p-mTOR, and a score = 3 was considered positive for p-ERK and p-S6RP, according to the staining sensitivity of each marker. Due to limitations on the amount of sample, staining of p-S6RP was performed in 95 samples, p-ERK in 61 samples, p-mTOR in 61 samples, and Ki67 in 40 samples from the initial cohort.
C4d immunostaining was performed on 3-μm −80°C frozen biopsy sections. Antigen retrieval was performed with Cell Conditioning 1 (Roche, United States) for 8min. Primary anti-C4d polyclonal antibody (Cell Marque, CA, United States) was incubated for 28 min at room temperature and performed on a Benchmark XT platform (Roche). Sections were visualized using the Roche Optiview Kit. C4d staining was classified as minimally, focally or diffusely positive according to Banff classification.
Sera samples were collected contemporaneously to the biopsy (0, [−15, 34.5] days) and stored at −80°C until their analysis. Antibody testing was performed using Luminex® HLA Single Antigen Bead assays (LABScreen, One Lambda, CA, United States) with a normalized mean fluorescence intensity (MFI) of 1000 as a cutoff for identification of a relevant HLA antibody. Donor HLA antibody specificity was ascribed considering donor HLA A, B, DRB1 and some C and DQB typing. Antibodies against some C or DQ antigens were assigned considering linkage disequilibrium.
Comparisons between normally distributed variables were carried out using Student’s t-test, and non-parametric variables were analyzed with U Mann-Whitney test or with Kruskal–Wallis test. Normal distribution of continuous variables was tested with Kolgorov-Smirnoff and Shapiro-Wilk tests. Chi-squared or Fisher’s exact tests were used for dichotomous variables. AUC (Area under the curve) values for “C4d,” “p-S6RP,” and “C4d and p-S6RP” were compared using the DeLong test for correlated AUC, in order to determine the best test in terms of diagnostic performance. A p < 0.05 was considered statistically significant. Statistical analysis was performed using SPSS® v.22.0 (IBM Corp., New York, United States).
The data that support the findings of this study are available from the corresponding author upon reasonable request.
For this study, first we evaluated 64 patients, 31 had a biopsy with ABMR diagnosis and circulating HLA-DSA (ABMR+DSA+), and 33 with normal biopsies (ABMR–DSA–) were selected as controls. Baseline characteristics were similar in both groups (Table 1). Almost half of the ABMR+DSA+ biopsies were performed for indication, significantly more than ABMR–DSA–, and ABMR+DSA+ showed worse serum creatinine and glomerular filtration rate (eGFR), and higher protein-creatinine ratio (Pr/Cr) at biopsy than ABMR–DSA– (Table 1). As expected, ABMR+DSA+ biopsies presented worse histopathologic lesions compared to ABMR–DSA– biopsies (Table 2). Clinical characteristics of patients treated or not with mTORi were similar, except for the proportion of patients receiving calcineurin inhibitors and/or mycophenolate (Supplementary Table 1).
Then, we included a group of patients with ABMR changes without HLA-DSA (ABMR+DSA–, n = 16) and a group with HLA-DSA but no ABMR changes (ABMR–DSA+, n = 17) in the biopsies. At baseline, there were no significant differences among groups (Table 3). At biopsy, patients with ABMR–DSA+ showed a significantly lower serum creatinine and lower protein-creatinine ratio (Pr/Cr) compared to ABMR+DSA+.
Patient follow-up at 1 and 4/5 years is shown in Table 4. Interestingly, we observed that ABMR–DSA+ patients had better graft function and lower proteinuria, less frequently a subsequent biopsy with ABMR, and less graft failure in comparison with 41.9% censored-graft loss in ABMR+DSA+ group. No significant differences were found when comparing ABMR+DSA+ with ABMR+DSA– during the follow-up. Data on death-censored graft survival is consistent with these results, showing a significant difference on graft survival between ABMR- and ABMR+ cases regardless of DSA detection (Supplementary Figure 1).
Considering all ABMR cases, with and without DSA (n = 47), we identified 2 mixed rejections, both of them corresponding to TCMR grade IIA (v1 lesions), and 8 cases with borderline changes concomitant to the ABMR histology.
Detection of p-S6RP, p-ERK, and p-mTOR in PTC (Figure 1) was scored as previously described (23). P-S6RP and P-ERK positive cases showed a granular cytoplasmic staining in endothelial cells, with some cases displaying a similar staining pattern in glomerular endothelial cells and epithelial tubular cells. P-mTOR staining was also granular and cytoplasmic in PTC endothelium, less intense than the other markers, and was found in some biopsies in epithelial tubular cells with strong apical and membranous staining. Due to limitations on the amount of sample, staining of p-S6RP was performed in 95 samples, p-ERK in 61 samples, p-mTOR in 61 samples, and Ki67 in 40 samples.
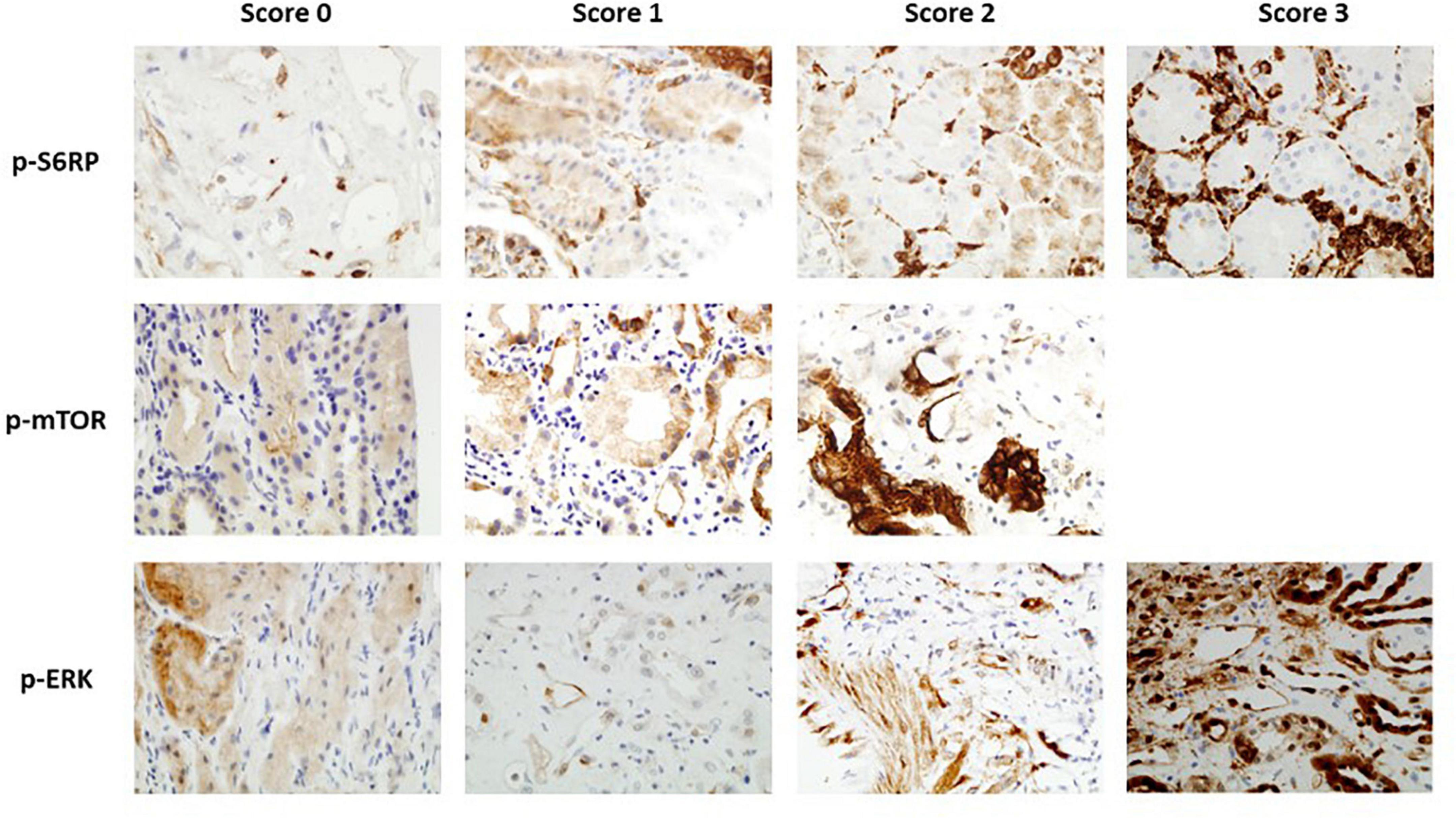
Figure 1. Immunohistochemistry staining of p-S6RP, p-ERK, and p-mTOR in kidney transplant biopsies (40×). Representative images of different staining scores for each protein are shown.
We analyzed whether endothelial positive staining of p-S6RP, p-ERK or p-mTOR and C4d in PTC of renal allografts associated with ABMR+DSA+ compared to ABMR–DSA–. As expected, C4d was significantly increased in biopsies with ABMR compared to control biopsies (p < 0.0001; Figure 2A). Interestingly, staining of p-S6RP was significantly increased in ABMR+DSA+ biopsies compared to ABMR–DSA– biopsies (p = 0.0003; Figure 2A). There was a slight difference between groups regarding the staining of p-ERK (p = 0.023) and no difference with p-mTOR (p = 0.072) (Figure 2A).
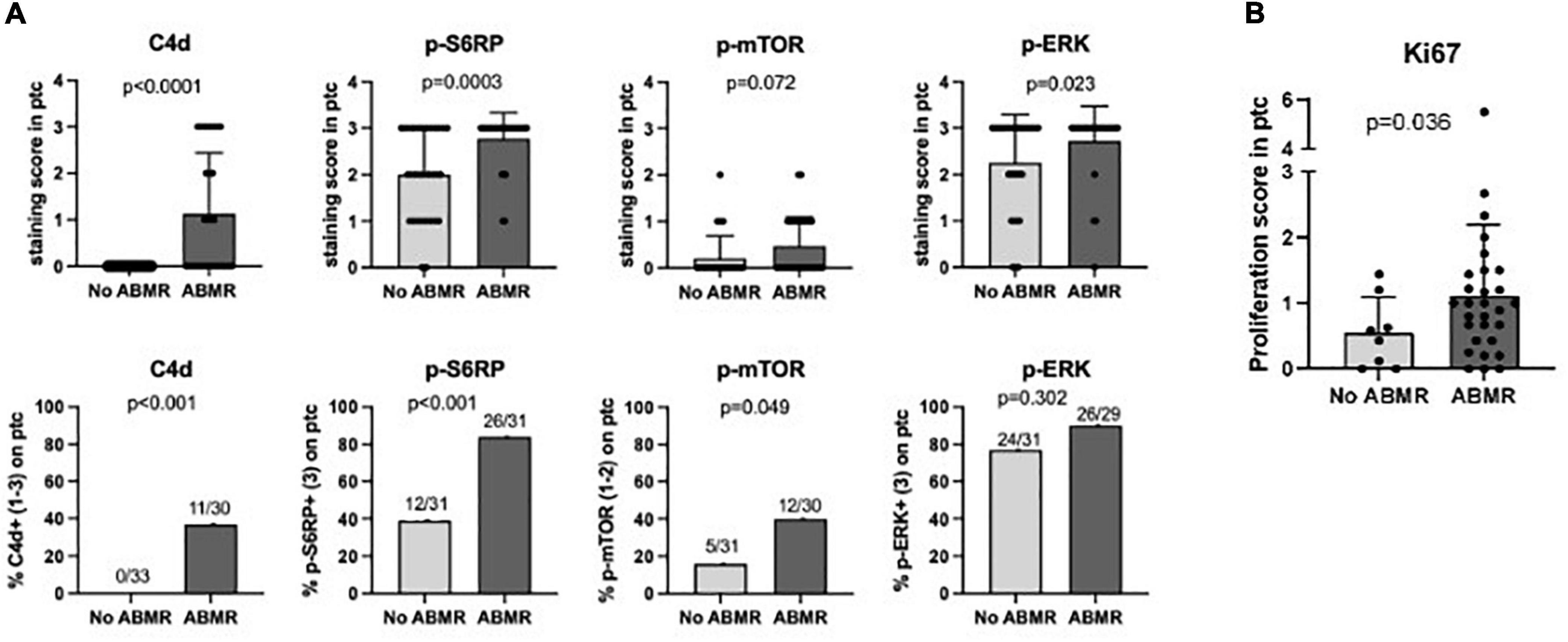
Figure 2. Staining of p-S6RP in peritubular capillaries is increased in ABMR biopsies. (A) Staining score (upper graphs) and percentage of positive staining (lower graphs) of C4d, p-S6RP, p-ERK, and p-mTOR in PTC of renal allografts biopsies with ABMR + DSA+ (n = 31) compared to ABMR–DSA– (n = 33). Mann Whitney and Fisher exact tests were used to assess significant differences between these two groups. (B) Staining of the proliferation marker Ki67 was analyzed in biopsies with and without ABMR. Plots show the means (bars), standard deviation (error bars), and individual values (dots) for each sample. Mann Whitney and Fisher exact tests were used to assess significant differences between groups.
Overall, these results suggest that positive staining of p-S6RP in PTC of renal biopsies is associated with the interaction of DSA with endothelium. Given that HLA-DSA have been reported to activate proliferation of endothelial cells, staining with the proliferation marker Ki67 was analyzed in biopsies with and without ABMR. Accordingly, ABMR+DSA+ biopsies showed a significantly increased Ki67 staining score in endothelium of PTC compared with ABMR–DSA– biopsies (p = 0.036, Figure 2B).
We could not find any significant correlation between p-S6RP PTC staining and peritubular capillaritis, PTC multilayering or chronic transplant glomerulopathy (Supplementary Table 2). Interestingly, death-censored graft survival was worse in patients with positive staining of p-S6RP in PTC compared with those with negative staining of p-S6RP (p = 0.028, Figure 3A). Nevertheless, considering the impact that ABMR and DSA detection may have in graft survival, we further analyzed graft survival stratifying patients according to the presence or absence of ABMR or DSA, and positive or negative p-S6RP staining (Figure 3B). Our results reflect that worse graft survival is observed in patients with ABMR and positive p-S6RP staining, compared to those with ABMR and negative p-S6RP staining, or without ABMR regardless of p-S6RP staining (p < 0.001, Figure 3B). Inferior allograft survival was also observed in cases with DSA and positive p-S6RP staining compared with DSA and negative p-S6RP staining, or non-DSA detection and positive or negative p-S6RP staining cases (p = 0.042, Figure 3B).
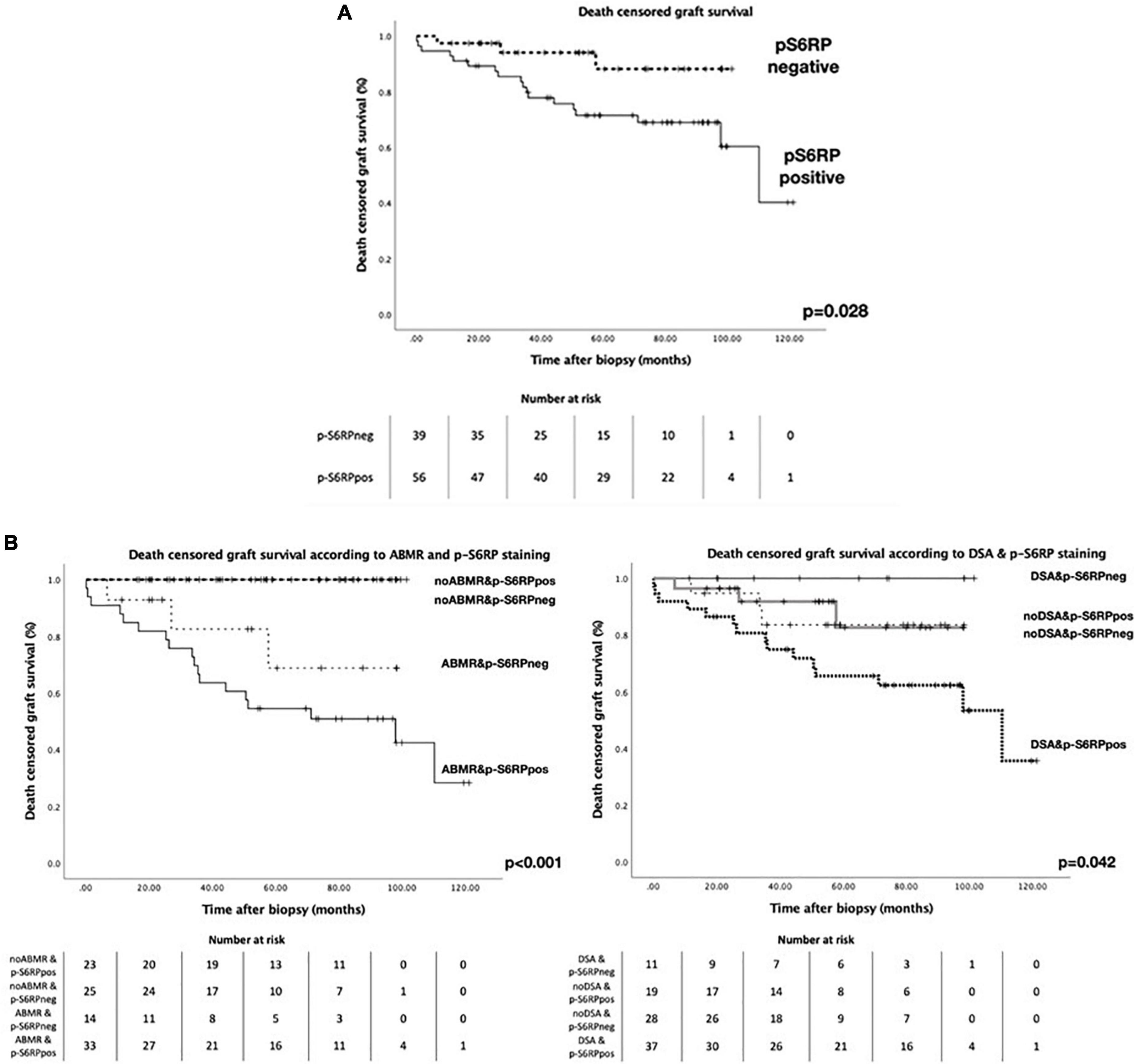
Figure 3. Death censored graft survival in patients with biopsies with p-S6RP positive and negative staining. (A) Kaplan-Meier survival curve representing death censored graft survival in patients with positive or negative p-S6RP staining in PTC. (B Left), Kaplan-Meier survival curve representing death censored graft survival in patients with and without ABMR diagnosis according to the p-S6RP staining in the biopsy. P-value between ABMR+ cases with positive or negative p-S6RP staining is p = 0.21. (B Right), Kaplan-Meier survival curve representing death censored graft survival in patients with and without DSA diagnosis according to the p-S6RP staining in the biopsy. P-value between DSA+ cases with positive or negative p-S6RP staining is p = 0.043.
We evaluated the potential modulation of the mTOR pathway by mTORi immunosuppression. Results showed that treatment with mTORi did not significantly modify C4d staining or phosphorylation of S6RP, ERK or mTOR (Figures 4A,B). Excluding from the analysis four patients treated with mTORi combined with tacrolimus, as their mTORi trough levels were lower, we observed similar results (data not shown). Analysis of mTORi treatment on active (n = 14) vs. chronic (n = 32) ABMR showed no apparent impact of mTORi on the studied phosphorylated proteins (Figure 4C). These results may suggest that mTORi treatment in our cohort do not modulate p-mTOR, p-S6RP, p-ERK or C4d staining neither in chronic nor in active ABMR, although they need to be confirmed with a higher number of samples.
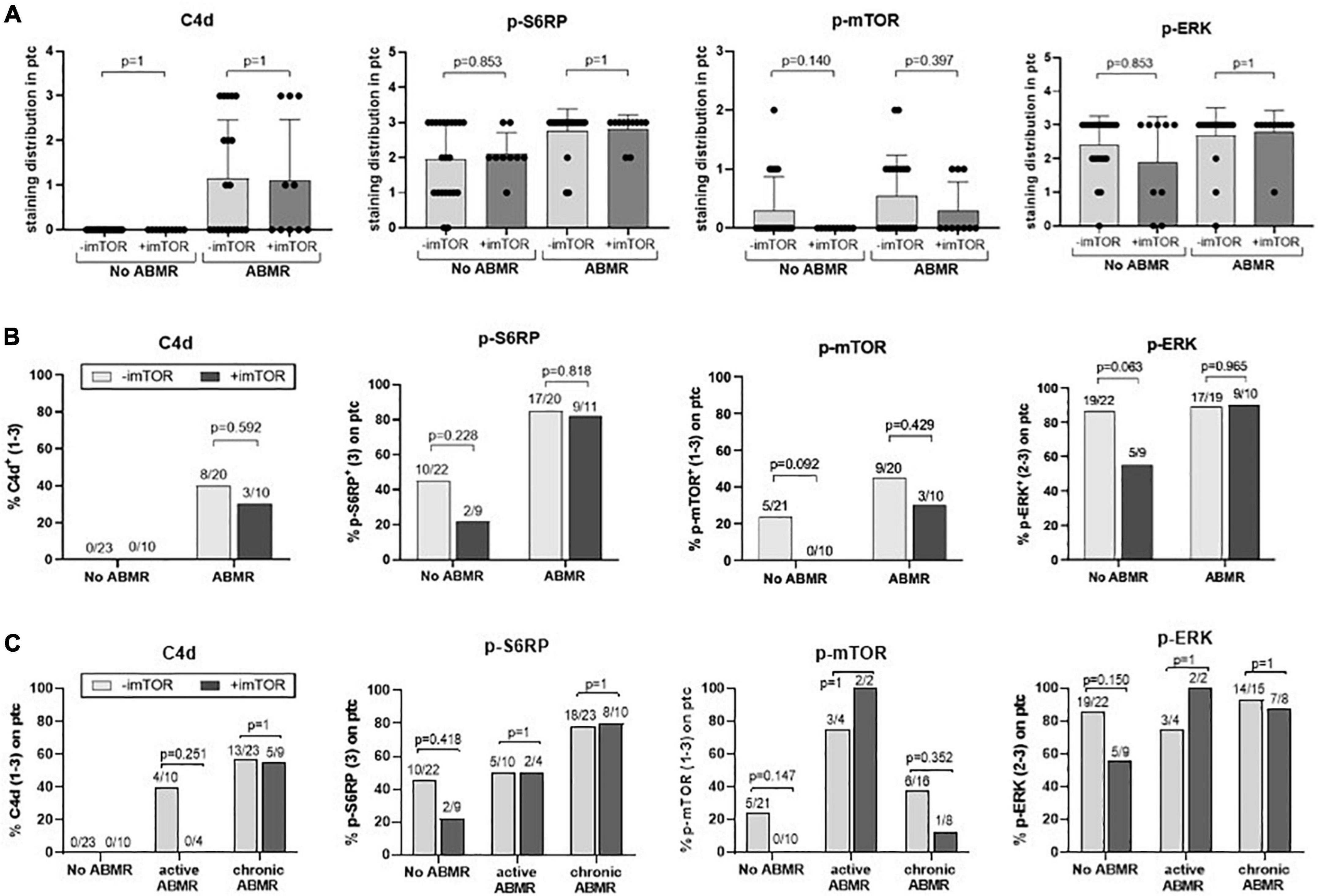
Figure 4. Potential modulation of C4d, p-S6RP, p-mTOR, and p-ERK staining by mTORi immunosuppression. Staining score (A) and percentage of positive staining (B) of C4d, p-S6RP, p-ERK, and p-mTOR in PTC of renal allografts biopsies with ABMR + DSA+ treated (n = 11) or not (n = 20) with mTORi, and ABMR–DSA– treated (n = 4) or not (n = 7) with mTORi. Plots of staining score show the means (bars), standard deviation (error bars), and individual values (dots) for each sample. Kruskal-Wallis test was used to assess significant differences among groups for staining score and Fisher exact test for proportions of staining. (C) Percentage of positive staining for C4d, p-S6RP, p-ERK, and p-mTOR in PTC of renal allografts biopsies with active and chronic ABMR, compared to no ABMR, receiving or not mTORi immunosuppression. Fisher exact test was used to assess significant differences among groups.
Then we assessed whether the positive staining of p-S6RP in ABMR depended upon the presence of circulating HLA-DSA at the time of biopsy. We incorporated a group of patients (n = 16) with histological damage in biopsies compatible with ABMR who had no detectable HLA-DSA, and found that p-S6RP was significantly higher in ABMR+DSA+ compared to ABMR+DSA– (p = 0.0242, Figure 5A). Next we evaluated whether p-S6RP positivity only occurred in patients with circulating HLA-DSA who had ABMR on their biopsies, and a new group of patients (n = 17) with DSA+ABMR– was included. There was no significant difference on p-S6RP staining between ABMR+DSA+ and ABMR–DSA+ biopsies (p = 1, Figure 5B). These results support an association of positive p-S6RP staining with HLA-DSA independently of histological damage.
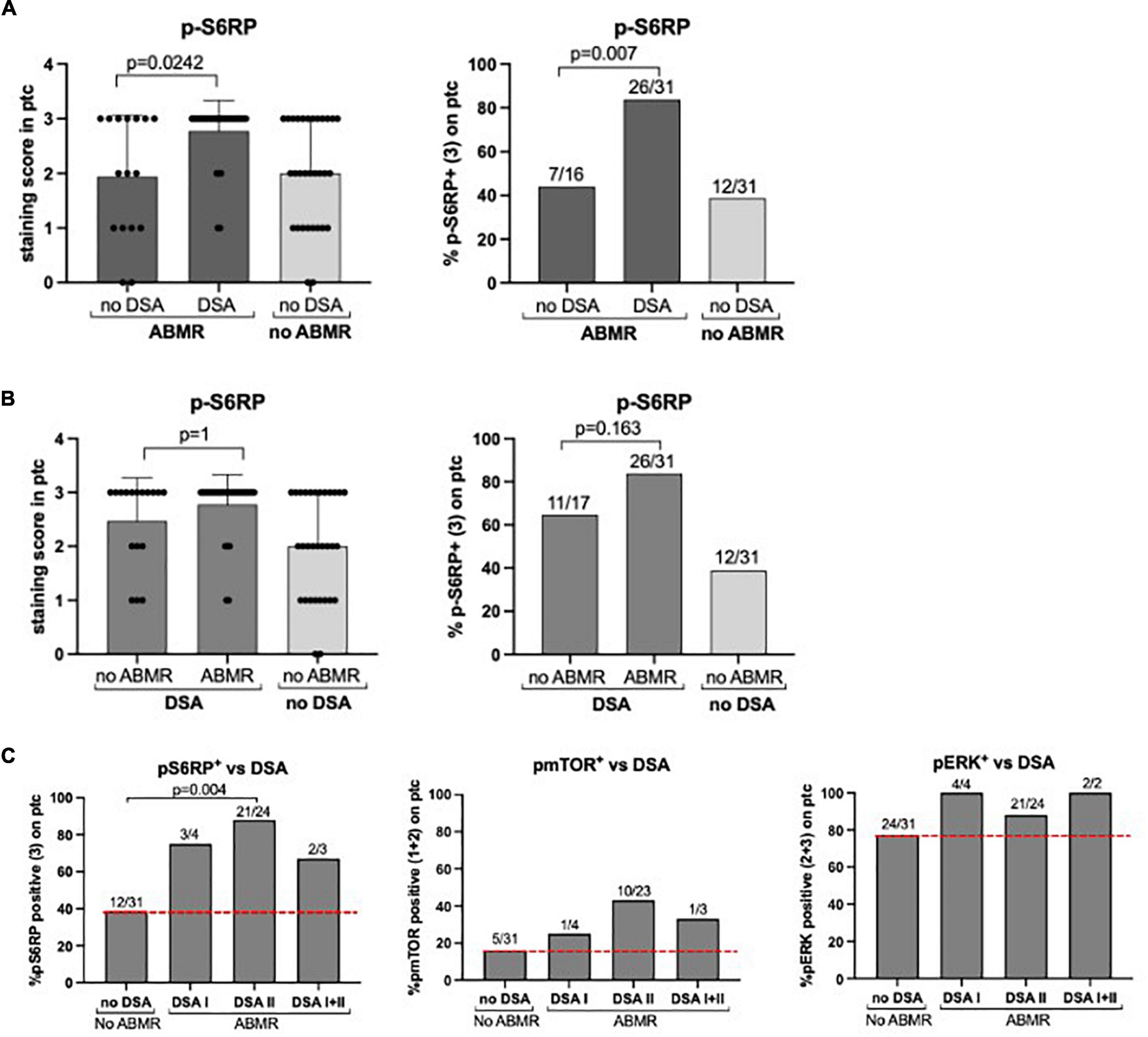
Figure 5. P-S6RP in peritubular capillaries is associated with HLA-DSA. (A) Comparison of p-S6RP staining score in biopsies compatible with ABMR with DSA (n = 31) and with no detectable HLA-DSA (n = 16). (B) Comparison of p-S6RP staining score in biopsies with DSA and ABMR (n = 31) with biopsies with HLA-DSA but not ABMR (n = 18). Plots show the means (bars), standard deviation (error bars), and individual values (dots) for each sample. Mann Whitney test was used to assess significant differences between groups. (C) Percentage of positive staining (bars) for p-S6RP, p-mTOR, and p-ERK on biopsies with DSA class I, II, or I&II compared to biopsies with no DSA and no rejection (red dotted line).
We also assessed potential differences in p-S6RP staining depending on the type or amount of circulating HLA-DSA. A higher percentage of diffuse p-S6RP staining was observed in ABMR biopsies with DSA class I, II, or I&II compared to no rejection biopsies without DSA (Figure 4C). Considering all four groups of patients, positive p-S6RP staining was associated with total DSA (p < 0.001) and with HLA class-II DSA (p = 0.002), but not with DSA HLA class-I (p = 0.52) (Table 5). The amount of circulating DSA, employing as surrogate the sum of the mean fluorescence intensity of all circulating DSA, did not correlate with p-S6RP staining intensity (Supplementary Figure 2). There was a higher proportion of positive p-mTOR staining biopsies with class-II DSA compared to biopsies with no DSA (Figure 5C) and an association of p-mTOR staining with class-II DSA (p = 0.014, Table 5). We found no differences in p-ERK staining comparing different types of DSA with no DSA samples (Figure 5C).
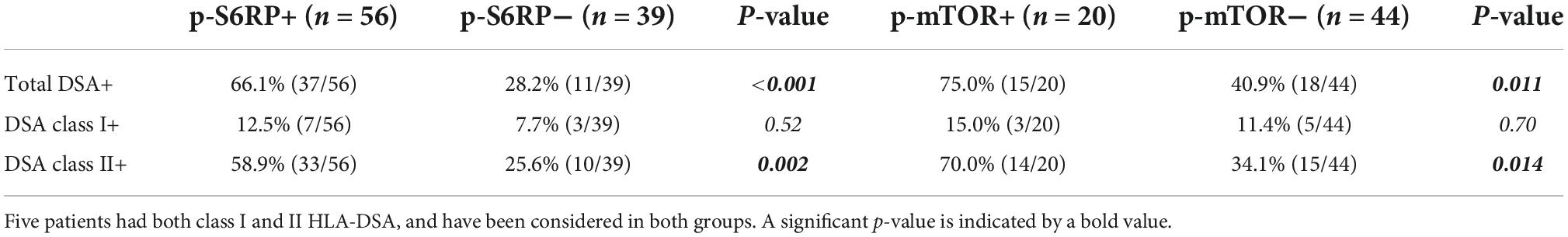
Table 5. Percentage of positive and negative p-S6RP and p-mTOR staining in peritubular capillaries in biopsy cases with class I, class II, and total DSA.
Staining of p-S6RP and C4d did not correlate (p = 0.37, Table 6). However, we analyzed whether p-S6RP staining could improve the C4d diagnostic capacity for those cases with ABMR without DSA. Accuracy diagnostics of “C4d,” “p-S6RP,” and “C4d and p-S6RP” for ABMR+DSA+ and ABMR+DSA– are shown through sensitivity and specificity in Table 7, and AUC values for each method were compared in order to determine the best test in terms of diagnostic performance. We found that p-S6RP improves the sensitivity compared to C4d, and that both C4d and p-S6RP together significantly improve the accuracy diagnostics vs. C4d alone, when comparing ABMR with and without HLA-DSA (p = 0.017; Table 7). These results indicate that positive p-S6RP staining complements the diagnostic capacity of C4d, which could be helpful for cases with ABMR+ DSA–.

Table 6. Correlation table between p-S6RP and C4d staining.
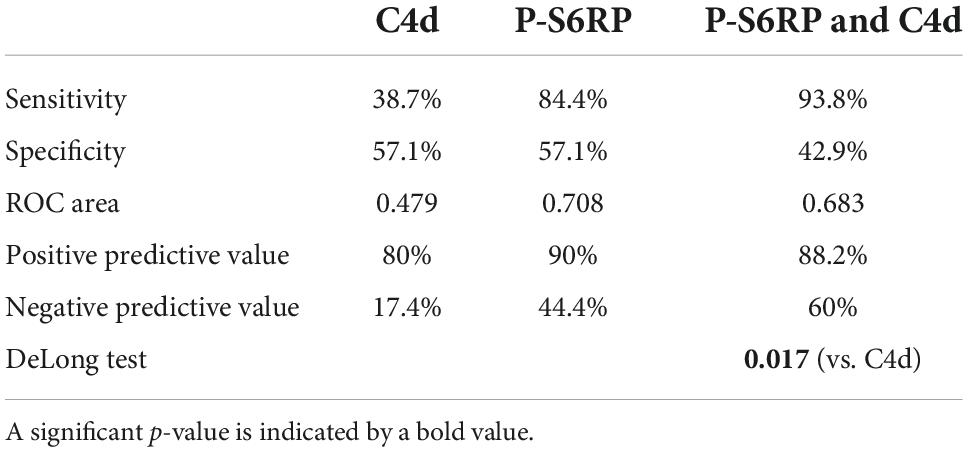
Table 7. Diagnostic capability of C4d, p-S6RP, and p-S6RP&C4d to distinguish DSA+ from DSA– in cases with ABMR+.
The diagnostic approach to transplant rejection based on molecular biomarkers may help in the selection of appropriate treatment strategies. We hypothesized that staining of phosphorylated mTOR and its downstream signals S6RP and ERK in kidney allografts may be useful as diagnostic biomarkers and therapeutic targets in ABMR. To test this, we used phosphorylation-specific antibodies to elucidate the effects of anti-HLA antibody-induced signal transduction in PTC endothelium in vivo. Our findings support that p-S6RP in peritubular capillary endothelium of kidney graft biopsies is associated with circulating HLA-DSA in the presence or absence of ABMR. In contrast, we could not find a clear significant difference on the phosphorylation of mTOR or ERK comparing ABMR+DSA+ with ABMR-DSA- biopsies. This could be due to the redundancy of the biologic system in which growth factors and cytokines can also activate PI3K, which in turn phosphorylates mTOR at Ser2448. Furthermore, the phosphorylation of intracellular ERK in vitro is described to depend only on mTORC2 activation in the case of HLA class-I DSA (26), and independent of mTOR in the case of HLA class-II DSA (17).
To our knowledge, this is the first study to evaluate the phosphorylation of mTOR proteins in kidney allografts in the context of ABMR. The association between ABMR and the capillary staining of phosphorylated S6K and S6RP was previously reported in heart allograft biopsies (22, 23). Furthermore, class I or class II ligation on in vitro endothelial cells results in a time and dose-dependent increase in Ser235/236 phosphorylation of S6RP (22). We were expecting to see a correlation between p-S6RP intensity staining and renal damage, however we could not find this correlation, and both active and chronic ABMR presented the same p-S6RP intensity. Instead, we found that high p-S6RP staining was associated with HLA-DSA independently of histological damage.
As novelty, we also had the opportunity to analyze the value of p-S6RP staining in biopsies of KT patients with ABMR without DSA, and in patients with DSA without ABMR. We found that positive staining of p-S6RP was associated with HLA-DSA, specifically with HLA class-II, despite the absence of histological damage compatible with ABMR. Previous work in heart allografts found an association of p-S6RP staining with circulating anti-HLA antibodies, especially also with class II antibodies (22), in patients with cardiac rejection. Interestingly, plasma-derived HLA-DQ antibodies phosphorylate Akt, S6K, and S6RP in microvascular endothelial cells in vitro (27). In a clinical setting, we have found that p-S6RP staining evidences the activation of intracellular pathways in the presence of HLA-DSA and could serve as a biomarker of their existence, contributing to ABMR diagnosis. Of note, ABMR without evidence of HLA-DSA showed less frequently p-S6RP in PTC, reflecting that in those cases other triggers could be involved and they may produce damage through other mechanisms. Furthermore, a few cases of ABMR+DSA– cases exhibited p-S6RP in PTC potentially due to a low level of HLA DSA undetected with SAB at the time of biopsy (28). In our experience, four ABMR+DSA– with diffuse p-S6RP showed detectable DSA later on as shown in Tables 3, 4.
Based on the knowledge of the mTOR pathway and previous in vitro studies, we hypothesized that treatment with mTORi in KT patients could modulate the phosphorylation of mTOR and its downstream signals, interfering damage in cases with ABMR. No significant differences in p-S6RP, p-mTOR, p-ERK or C4d staining neither in chronic nor in active ABMR under different immunosuppression were found. These results could suggest that clinical dose and exposure to mTORi may not have been enough in these cases to inhibit the intracellular signaling. The low number of ABMR cases treated with mTORi is one of the limitations of our study, and a higher number of cases would be necessary to confirm these results. Previous in vitro studies reported that everolimus effectively inhibited the mTOR activation pathway in anti-HLA class-I antibody-activated endothelial cells (24), pointing to a potential beneficial effect of everolimus in the prevention of chronic ABMR. Of interest, due to the relevance of the mTOR pathway in regulating essential aspects of cell biology, other proteins may have a compensatory activity that allows restoring some of the long-term inhibited mTOR functions. Actually, Ser235/236 phosphorylation depends on both p-70S6K and p-90RSK. Therefore, it is conceivable that Ser235/236 p-S6RP was not affected by mTORi because it was phosphorylated by p90RSK, an enzyme downstream of ERK not altered by rapalogs. The lack of inhibition of p-ERK by mTORi may be due to compensatory over-activation of ERK in response to antibody-mediated ligation of class II molecules in EC, as previously described (17). Furthermore, the mTOR activity in the kidney has been identified in many different cell types (29) and has been related to ischemia-reperfusion injury (30), interstitial fibrosis (31), and renal carcinoma (32). Therefore, many factors modulate and may impact the mTOR pathway activation in the kidney.
Overall our findings support that p-S6RP in capillary endothelium of kidney graft biopsies is associated with circulating HLA-DSA, independently of the existence of damage in the form of ABMR. These results suggest that p-S6RP staining in kidney allografts evidences the activation of intracellular pathways due to HLA-DSA, and therefore it could serve as a sensitive surrogate biomarker of HLA-DSA for ABMR diagnosis in cases where DSA are not available or not detected yet. p-S6RP emerges as a potential useful tool for prediction of ABMR in the presence of HLA DSA. The lack of correlation of p-S6RP and C4d staining in PTC, despite both being highly associated with circulating HLA DSA and/or ABMR, illustrate two different mechanisms of damage activated by HLA DSA, which deserve different treatment approaches. Larger studies are needed to see if pS6RP could be of interest to assess the value of treatment efficacy in late ABMR in clinical trials.
The raw data supporting the conclusion of this article will be made available by the authors, without undue reservation.
DR-R participated in the design and performance of the research, the data analysis, the figure preparation, and wrote the manuscript. JG participated in the design, performance of the research, and the figure preparation. LL-M participated in acquisition and analysis of the data and the figure preparation. SM, DB, and MR participated in the performance of the research. DR and MP-S participated in acquisition of the data. ER and JP participated in the interpretation of the data. MC participated in the research design, data acquisition, interpretation of results, and wrote the manuscript. All authors contributed to the article and approved the submitted version.
This study was performed with funding from projects PI10/01370, PI13/00598, PI16/00617, and PI PI20/00090, funded by Instituto de Salud Carlos III (ISCIII) and co-funded by the European Union, RD12/0021/0024 (Spanish Ministry of Health ISCIII FIS-FEDER), RD12/0021/0024 and RD16/0009/0013 (ISCIII FEDER RedinRen) and 201822-10 (Fundació la Marató de TV3). DR-R was supported by a Sara Borrell Fellowship from Instituto de Salud Carlos III (CD17/00025). SM was supported by Departament de Salut, Generalitat de Catalunya (PERIS SLT006/17/00040).
We acknowledge the technical support received by the Cancer Molecular Therapy group (IMIM) directed by Dr. Albanell, and we thank the statistical assessment of Xavier Duran Jordà (IMIM). We are also indebted to Anna Faura, Sara Alvarez, and Rosa Causadias for their technical support. JG did this study as part of his doctoral thesis program at the Department of Medicine from the Universitat Autonoma of Barcelona (UAB).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmed.2022.988080/full#supplementary-material
Supplementary Figure 1 | Death censored graft survival in ABMR + DSA+, ABMR-DSA−, ABMR + DSA−, and ABMR-DSA+ patients. Kaplan-Meier survival curve representing death censored graft survival in the four groups of patients included in the study.
Supplementary Figure 2 | Correlation between p-S6RP staining intensity and the MFI of detected DSA.
ABMR, antibody mediated rejection; CTG, chronic transplant glomerulopathy; DSA, donor-specific HLA antibodies; eGFR, estimated glomerular filtration rate; HLA, human leukocyte antigens; KT, kidney transplantation; mTOR, mammalian or mechanistic target of rapamycin; mTORi, mTOR inhibitor; p-S6RP, phospho-S6 ribosomal protein at Ser235/236; p-ERK, phospho-ERK1/2 (p44/42 MAPK) at Thr202/Tyr204; p-mTOR, phospho-mTOR at Ser2448; Pr/Cr, urinary protein to creatinine ratio; PTC, peritubular capillaries; TG, transplant glomerulopathy.
1. Einecke G, Sis B, Reeve J, Mengel M, Campbell PM, Hidalgo LG, et al. Antibody-mediated microcirculation injury is the major cause of late kidney transplant failure. Am J Transplant. (2009) 9:2520–31. doi: 10.1111/j.1600-6143.2009.02799.x
2. Lee P-C, Terasaki PI, Takemoto SK, Lee P-H, Hung C-J, Chen Y-L, et al. All chronic rejection failures of kidney transplants were preceded by the development of HLA antibodies. Transplantation. (2002) 74:1192–4. doi: 10.1097/00007890-200210270-00025
3. Redondo-Pachón D, Pérez-Sáez MJ, Mir M, Gimeno J, Llinás L, García C, et al. Impact of persistent and cleared preformed HLA DSA on kidney transplant outcomes. Hum Immunol. (2018) 79:424–31. doi: 10.1016/j.humimm.2018.02.014
4. Gimeno J, Redondo D, Pérez-Sáez MJ, Naranjo-Hans D, Pascual J, Crespo M. Impact of the Banff 2013 classification on the diagnosis of suspicious versus conclusive late antibody-mediated rejection in allografts without acute dysfunction. Nephrol Dial Transplant. (2016) 31:1938–46. doi: 10.1093/ndt/gfw223
5. Hickey MJ, Valenzuela NM, Reed EF. Alloantibody generation and effector function following sensitization to human leukocyte antigen. Front Immunol. (2016) 7:30. doi: 10.3389/fimmu.2016.00030
6. Mauiyyedi S, Crespo M, Collins AB, Schneeberger EE, Pascual MA, Saidman SL, et al. Acute humoral rejection in kidney transplantation: II. Morphology, immunopathology, and pathologic classification. J Am Soc Nephrol. (2002) 13:779–87. doi: 10.1681/ASN.V133779
7. Racusen LC, Halloran PF, Solez K. Banff 2003 meeting report: new diagnostic insights and standards. Am J Transplant. (2004) 4:1562–6. doi: 10.1111/j.1600-6143.2004.00585.x
8. Haririan A, Kiangkitiwan B, Kukuruga D, Cooper M, Hurley H, Drachenberg C, et al. The impact of c4d pattern and donor-specific antibody on graft survival in recipients requiring indication renal allograft biopsy. Am J Transplant. (2009) 9:2758–67. doi: 10.1111/j.1600-6143.2009.02836.x
9. Haas M, Sis B, Racusen LC, Solez K, Glotz D, Colvin RB, et al. Banff 2013 meeting report: inclusion of C4d-negative antibody-mediated rejection and antibody-associated arterial lesions. Am J Transplant. (2014) 14:272–83. doi: 10.1111/ajt.12590
10. Dominy KM, Willicombe M, Al Johani T, Beckwith H, Goodall D, Brookes P, et al. Molecular assessment of C4d-positive renal transplant biopsies without evidence of rejection. Kidney Int Rep. (2019) 4:148–58. doi: 10.1016/j.ekir.2018.09.005
11. Hayde N, Broin PO, Bao Y, de Boccardo G, Lubetzky M, Ajaimy M, et al. Increased intragraft rejection-associated gene transcripts in patients with donor-specific antibodies and normal biopsies. Kidney Int. (2014) 86:600–9. doi: 10.1038/ki.2014.75
12. Muczynski KA, Cotner T, Anderson SK. Unusual expression of human lymphocyte antigen class II in normal renal microvascular endothelium. Kidney Int. (2001) 59:488–97. doi: 10.1046/j.1523-1755.2001.059002488.x
13. Hall BM, Bishop GA, Duggin GG, Horvath JS, Philips J, Tiller DJ. Increased expression of HLA-DR antigens on renal tubular cells in renal transplants: relevance to the rejection response. Lancet (London, England). (1984) 2:247–51. doi: 10.1016/s0140-6736(84)90297-6
14. Issa N, Cosio FG, Gloor JM, Sethi S, Dean PG, Moore SB, et al. Transplant glomerulopathy: risk and prognosis related to anti-human leukocyte antigen class II antibody levels. Transplantation. (2008) 86:681–5. doi: 10.1097/TP.0b013e3181837626
15. Jindra PT, Jin Y-P, Rozengurt E, Reed EF. HLA class I antibody-mediated endothelial cell proliferation via the mTOR pathway. J Immunol. (2008) 180:2357–66. doi: 10.4049/jimmunol.180.4.2357
16. Zhang X, Rozengurt E, Reed EF. HLA class I molecules partner with integrin β4 to stimulate endothelial cell proliferation and migration. Sci Signal. (2010) 3:ra85. doi: 10.1126/scisignal.2001158
17. Jin Y-P, Valenzuela NM, Zhang X, Rozengurt E, Reed EF. HLA class II-triggered signaling cascades cause endothelial cell proliferation and migration: relevance to antibody-mediated transplant rejection. J Immunol. (2018) 200:2372–90. doi: 10.4049/jimmunol.1701259
18. Hay N, Sonenberg N. Upstream and downstream of mTOR. Genes Dev. (2004) 18:1926–45. doi: 10.1101/gad.1212704
19. Kim D-H, Sarbassov DD, Ali SM, King JE, Latek RR, Erdjument-Bromage H, et al. mTOR interacts with raptor to form a nutrient-sensitive complex that signals to the cell growth machinery. Cell. (2002) 110:163–75. doi: 10.1016/s0092-8674(02)00808-5
20. Zoncu R, Efeyan A, Sabatini DM. mTOR: from growth signal integration to cancer, diabetes and ageing. Nat Rev Mol Cell Biol. (2011) 12:21–35. doi: 10.1038/nrm3025
21. Li W, Zhang PL. Up-regulated mTOR pathway indicates active disease in both human native and transplant kidneys. Ann Clin Lab Sci. (2013) 43:378–88.
22. Lepin EJ, Zhang Q, Zhang X, Jindra PT, Hong LS, Ayele P, et al. Phosphorylated S6 ribosomal protein: a novel biomarker of antibody-mediated rejection in heart allografts. Am J Transplant. (2006) 6:1560–71. doi: 10.1111/j.1600-6143.2006.01355.x
23. Li F, Wei J, Valenzuela NM, Lai C, Zhang Q, Gjertson D, et al. Phosphorylated S6 kinase and S6 ribosomal protein are diagnostic markers of antibody-mediated rejection in heart allografts. J Heart Lung Transplant. (2015) 34:580–7. doi: 10.1016/j.healun.2014.09.047
24. Jin YP, Valenzuela NM, Ziegler ME, Rozengurt E, Reed EF. Everolimus inhibits anti-HLA i antibody-mediated endothelial cell signaling, migration and proliferation more potently than sirolimus. Am J Transplant. (2014) 14:806–19. doi: 10.1111/ajt.12669
25. Haas M, Loupy A, Lefaucheur C, Roufosse C, Glotz D, Seron D, et al. The Banff 2017 Kidney Meeting Report: Revised diagnostic criteria for chronic active T cell-mediated rejection, antibody-mediated rejection, and prospects for integrative endpoints for next-generation clinical trials. Am J Transplant. (2018) 18:293–307. doi: 10.1111/ajt.14625
26. Jindra PT, Jin Y-P, Jacamo R, Rozengurt E, Reed EF. MHC class I and integrin ligation induce ERK activation via an mTORC2-dependent pathway. Biochem Biophys Res Commun. (2008) 369:781–7. doi: 10.1016/j.bbrc.2008.02.093
27. Cross AR, Lion J, Poussin K, Assayag M, Taupin J-L, Glotz D, et al. HLA-DQ alloantibodies directly activate the endothelium and compromise differentiation of FoxP3high regulatory T lymphocytes. Kidney Int. (2019) 96:689–98. doi: 10.1016/j.kint.2019.04.023
28. Luque S, Lúcia M, Melilli E, Lefaucheur C, Crespo M, Loupy A, et al. Value of monitoring circulating donor-reactive memory B cells to characterize antibody-mediated rejection after kidney transplantation. Am J Transplant. (2019) 19:368–80. doi: 10.1111/ajt.15055
29. Fantus D, Rogers NM, Grahammer F, Huber TB, Thomson AW. Roles of mTOR complexes in the kidney: implications for renal disease and transplantation. Nat Rev Nephrol. (2016) 12:587–609. doi: 10.1038/nrneph.2016.108
30. Alshaman R, Truong L, Oyekan A. Role of mechanistic target of rapamycin (mTOR) in renal function and ischaemia-reperfusion induced kidney injury. Clin Exp Pharmacol Physiol. (2016) 43:1087–96. doi: 10.1111/1440-1681.12648
31. Ogawa S, Ishimura T, Miyake H, Fujisawa M. Expression profile of mammalian target of rapamycin-related proteins in graft biopsy specimens: significance for predicting interstitial fibrosis after kidney transplantation. Int J Urol. (2017) 24:223–9. doi: 10.1111/iju.13287
Keywords: transplantation—kidney, donor specific antibodies, mammalian target of rapamycin (mTOR), phosphorylation, peritubular capillaries, ribosomal protein S6 (S6RP), antibody mediated allograft rejection
Citation: Raïch-Regué D, Gimeno J, Llinàs-Mallol L, Menéndez S, Benito D, Redondo D, Pérez-Sáez MJ, Riera M, Reed EF, Pascual J and Crespo M (2022) Phosphorylation of S6RP in peritubular capillaries of kidney grafts and circulating HLA donor-specific antibodies. Front. Med. 9:988080. doi: 10.3389/fmed.2022.988080
Received: 06 July 2022; Accepted: 26 September 2022;
Published: 18 October 2022.
Edited by:
Xiaopeng Hu, Capital Medical University, ChinaReviewed by:
Petra Hruba, Institute for Clinical and Experimental Medicine (IKEM), CzechiaCopyright © 2022 Raïch-Regué, Gimeno, Llinàs-Mallol, Menéndez, Benito, Redondo, Pérez-Sáez, Riera, Reed, Pascual and Crespo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Julio Pascual, anVscGFzY3VhbEBnbWFpbC5jb20=; Marta Crespo, bWNyZXNwb0Bwc21hci5jYXQ=
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.