 Charles Raine
Charles Raine Ian Giles
Ian Giles- Centre for Rheumatology, Department of Inflammation, Division of Medicine, University College London, London, United Kingdom
Rheumatoid arthritis (RA) is the most common inflammatory rheumatic disease and has a female predominance of around 3:1. The relationship between sex hormones and RA has been of great interest to researchers ever since Philip Hench's observations in the 1930's regarding spontaneous disease amelioration in pregnancy. Extensive basic scientific work has demonstrated the immunomodulatory actions of sex hormones but this therapeutic potential has not to date resulted in successful clinical trials in RA. Epidemiological data regarding both endogenous and exogenous hormonal factors are inconsistent, but declining estrogen and/or progesterone levels in the menopause and post-partum appear to increase the risk and severity of RA. This review assimilates basic scientific, epidemiological and clinical trial data to provide an overview of the current understanding of the relationship between sex hormones and RA, focusing on estrogen, progesterone and androgens.
Introduction
Rheumatoid arthritis (RA) is a chronic multisystem inflammatory disease which causes a destructive symmetrical polyarthritis. It is characterized by production of the autoantibodies rheumatoid factor (RF) and anti-citrullinated protein antibodies (ACPA). Common to many autoimmune disorders, there is a female predominance, with a female:male distribution of around 3:1 (1), although this gender disparity is less marked than in other inflammatory rheumatic conditions such as systemic lupus erythematosus. Other observations implicating the importance of sex hormones in the pathogenesis of RA include its peak incidence at menopause (2), reduced disease activity in pregnancy and flare in the postpartum period (3). Such phenomena would suggest that declining female sex hormones, principally oestrogren and progesterone, increase the risk of RA in menopause and post-partum, while increased levels in pregnancy are protective. However, despite extensive study, the relationship between sex hormones and RA pathogenesis remains complex.
This review will synthesize lines of evidence regarding the impact of sex hormones on the pathogenesis of RA, focusing on estrogen, progesterone and androgens. We present an overview of the data from basic laboratory research through to a comprehensive evaluation of epidemiological studies and clinical trials.
Strategy
We searched PubMed for original articles, systematic reviews and meta-analyses in English, pubished between 1 January 1990 and 31 December 2021. Reference lists in retrieved articles were also reviewed to identify further articles of relevance. Search terms used comprised combinations of the following: “Rheumatoid arthritis”, “sex hormones/steroids”, “(o)estrogen”, “progesterone”, “androgens”, “risk factors”, “disease activity”, “menarche”, “pregnancy”, “menopause”, “contraceptive”, “breastfeeding” and “hormone replacement therapy”.
Rheumatoid arthritis pathogenesis
A detailed discussion of the pathogenesis of rheumatoid arthritis it outside the scope of this article, and the reader is directed to other recent excellent reviews (4–7). In brief, the etiology of RA is recognized to depend on complex interactions between genes and environment, with a resultant breakdown of immune tolerance leading to inflammation in synovial joints. The strongest genetic component is found in the human leukocyte antigen (HLA) class II molecule HLA-DRB1, with over 80 % of patients carrying the shared epitope of the HLA-DRB1*04 cluster (8), which is also associated with severity of disease (9). RA pathogenesis is thought to be initiated years before the development of symptoms and likely involves the induction by environmental factors, the most established of which is smoking (10), of post-translational modifications. These processes lead to the activation by antigen-presenting cells of an adaptive immune response, with production of the hallmark autoantibodies RF (targeting immunoglobulin G) and ACPA (6). Interactions of immune cells, including T and B cells, plasma cells and monocytes, and pro-inflammatory cytokines, most importantly tumor necrosis factor-alpha (TNF-α) and interleukin (IL)-6, lead to an infux and local activation of inflammatory cells in the synovium. Macrophage-like synoviocytes proliferate and produce TNF-α, IL-6 and IL-1, among other pro-inflammatory mediators. Infiltrating immune cells include CD4+ T cells and mature B cells producing RF and ACPAs. The synovial membrane expands and is filled with new blood vessels. Within the synovium, abundant osteoclasts are the primary mediators of bone erosion, while fibroblast-like synoviocytes secrete matrix metalloproteinases into the synovial fluid leading to cartilage degradation.
Sex hormone signaling
The sex hormones estrogen, progesterone and androgens all bind to nuclear receptors which belong to the type 1 family of nuclear hormone transcription factors (to which the glucocorticoid receptor also belongs) (11–13). In brief, binding of these hormones to their receptors in the cytosol leads to dissociation of heat shock proteins, homodimerization, translocation into the nucleus and subsequent binding to hormone response elements within regulatory regions of target genes, resulting in modification of gene transcription. Classically, oestrogens bind the cytosolic receptors estrogen receptor (ER)- α and/or ERβ to mediate genomic effects; in addition, oestrogens may also bind to membrane receptors such as G protein-coupled estrogen receptor 1 (GPER1) to trigger downstream signaling cascades that non-directly influence gene expression. The progesterone receptor (PR) and androgen receptor (AR), which are closely related, mediate classical genomic effects in a similar fashion to oestrogens, with hormone-receptor complexes binding to progresterone- and androgen-response elements respectively.
Sex hormones and genetic risk of rheumatoid arthritis
Estrogen and progesterone
Genome-wide and targeted gene analysis studies have revealed single nucleotide polymorphisms (SNPs) in sex steroid-associated genes that influence the risk and course of RA. These include SNPs in ESR2 which encodes estrogen receptor- β (ERβ) conferring a reduced risk of erosive arthritis (14) and an improved chance of responding to anti-tumor necrosis factor (TNF) therapy (15); other SNPs associated with a reduced risk of erosive disease include those in cytochrome p450 enzymes CYP1B1 and CYP2C9, which convert oestrogens to anti-inflammatory hydroxy-oestrogens (14). We are not aware of any studies evaluating genes involved in progesterone signaling and the risk or severity of RA.
Androgens
SNPs in the cytochrome B5-encoding gene CYB5A have been associated with a reduced susceptibility to RA in women (16). Such polymorphisms lead to an increased production by the cytochrome B5 enzyme of androgens from precursor hormones, suggesting that higher androgen levels protect against the development of RA.
Sex hormone signaling in the immune system
There is a significant body of evidence of the immunomodulatory effects mediated by sex hormones in autoimmune disease [extensively reviewed in Hughes (17), Wilder and Elenkov (18), Hughes and Choubey (19), Kanik and Wilder (20), Cutolo and Straub (21) and Moulton (22)].
Estrogen
Oestrogens have complex interactions with the immune system which may be pro- or anti-inflammatory depending on the cell type and concentrations involved (21, 22). With relevance to B cell function, at physiological concentrations the estrogen-ERα complex binds to the promoter of the AICDA gene which in turn stimulates expression of the enzyme activation-induced cytidine deaminase (AID). This enzyme is the master regulator of somatic hypermutation and class switch recombination. Therefore, oestrogens support immunoglobulin class switching in B cells and would logically be expected to have a deleterious effect on autoimmune diseases characterized by autoantibody production (23).
T cell ontogeny comprises initial development as haematopoetic precusors in the bone-marrow which then migrate to the thymus. Here, a process of positive and negative selection occurs, such that T cells which react strongly with antigen presented by major histocompatibility complex survive, but those that react with self-antigen are eliminated (except a proportion of CD4+ cells that survive as regulatory T cells (Treg), see below). The process of negative selection is regulated by the transcription factor AIRE, which serves to promote self-tolerance. This protein has emerged as a key factor in sexual dimorphism in autoimmunity, with females expressing lower levels of AIRE post-puberty compared to males (24). Male castration reduces expression of AIRE, while ERα deficient mice showed no difference in expression between sexes. Moreover, estrogen treatment was found to decrease AIRE expression in human thymic tissue via methylation of CpG sites in the AIRE promoter (24).
In contrast, oestrogens have been found to have a range of direct anti-inflammatory actions on T cells. Silibinin, a natural agonist of ERβ was shown to reduce the in-vitro expression of pro-inflammatory interleukin (IL)-17 and TNFα in T cells from healthy donors and patients with active RA (25). This effect was mediated via down modulation of the expression of the epigenetic modifier microRNA-155. Silibinin has also been shown to stimulate apoptosis of human RA synoviocytes in vitro (26) and reduce the production of inflammatory cytokines in rats with collagen-induced arthritis (26).
Oestrogens generally exert inhibitory effects on pro-inflammatory TH1 cells, while they may inhibit pro-inflammatory TH17 cells via ERα or have the opposite effect via ERβ. At high concentrations such as seen in pregnancy, oestrogens induce the secretion of IL-10 and suppress TNFα production in T cells, supporting an anti-inflammatory cytokine milieu (27).
Forkhead box (FOXP3) expressing Treg cells are highly suppressive regulators of the immune response through secretion of anti-inflammatory cytokines such as IL-10 as well as cell-cell contact mechanisms. The FOXP3 locus possesses sex steroid response elements enabling direct binding of hormones and subsequent modulation of FOXP3 activation. Endogenous complexes of estradiol and ERβ were shown to bind directly and activate the FOXP3 promoter in Treg cells from both human cervical cancer specimens and healthy controls (28).
It has been shown that estrogen treatment before immunization with collagen can retard disease development in collagen-induced arthritis (CIA), a mouse model of RA (29). In CIA, exogenous estrogen administration ameliorated postpartum flare (30), while joint inflammation improved with treatment of non-pregnant arthritis-prone mice with estradiol or progesterone at pregnancy-like levels (31), or by treatment with high-potency estrogen alone (32). Estradiol has been linked to expansion of anti-inflammatory Treg cells in pregnant mice with autoimmune encephalitis (33) and pregnancy-induced amelioration of joint inflammation has been achieved by treatment of non-pregnant SKG mice (a model of human RA) with either estradiol or progesterone at pregnancy-like levels (31).
Progesterone
In contrast to the dichotomous effects of oestrogens, progesterone exhibits broad anti-inflammatory actions (17, 21), including: inhibition of AID (in opposition to the effect of oestrogens); inhibition of the TH1 and TH17 response; and inhibition of NK cells, neutrophils and macrophages.
Progesterone promotes the TH2 response by enhancing IL-4 and IL-10 production in human T cells (27). During pregnancy, lymphocytes express progesterone receptors and release a protein named progesterone-induced blocking factor, which has strong anti-natural killer (NK) cell activity, and also secrete IL-10 (34). Progesterone has been shown to induce FOXP3+ Tregs from naive murine CD4+ T cells via suppression of mammalian target of rapamycin (mTOR) signaling (35); furthermore, progesterone promotes differentiation of human fetal cord blood T cells into Tregs and suppresses their differentiation to TH17 cells (36). In multiple sclerosis, Tregs express high levels of estrogen and progesterone receptors, and each hormone enhances the suppressive function of Tregs in vitro (37).
Recently, progesterone was shown to suppress activation (as measured by CD69 and CD25 expression) of ex vivo CD4+ T cells from healthy human females in a dose-dependent fashion (using doses similar to those found in the placenta) (38); furthermore, RNA sequencing analysis showed significant transcriptomic changes involving downregulation of immune-related genes and pathways important in RA, such as signal transducer and activator of transcription (STAT)-1 and STAT3.
Androgens
Androgens such as testosterone have a range of anti-inflammatory effects in vivo, reducing secretion of inflammatory cytokines such as TNF, IL-1 and IL-6 by monocytes and inhibiting B cell lymphopoiesis and antibody production (39). In inflammatory rheumatic diseases serum androgen levels are often reduced (40) owing to the stimulation by inflammatory cytokines such as TNF, IL-1 and IL-6 of the aromatase enzyme in immune cells and fibroblasts. Androgens are also known to bind and upregulate tyrosine-protein phosphatase non-receptor type 1 (PTPN1), which has a broad range of functions in cell growth and immune function. PTPN1 inhibits janus kinase (JAK)-2 and tyrosine kinase 2, part of the JAK-STAT pathway which is integral to TH1 cell-mediated immune responses and the production of IL-12 and IFNγ. This signaling pathway is of especial interest in RA given the established therapeutic options targeting the JAK-STAT pathway. It is not yet known whether response to these treatments differs between men and women with RA.
Conversely to oestrogens, androgens promote AIRE transcription by recruiting androgen receptors to AIRE promoter regions, leading to higher expression in mice and human thymus in males compared to females (41). Male sex and androgen treatment were protective in a mouse model of multiple sclerosis, but to date this effect has not been investigated in models of rheumatic diseases including RA (41).
Similarly to oestrogens, androgens can bind to the FOXP3 androgen response element, leading to acetylation of histone H4 and activation of FOXP3; there is a strong androgen-dependent increase of FOXP3 expression in T cells from women in the ovulatory phase of the menstrual cycle but not from men (42).
Sex hormones and pathogenesis of RA: Epidemiology
There is extensive epidemiological data implicating the importance of sex hormones in the pathophysiology of RA, but results are conflicting. Large modern cohort studies and meta-analyses have often contradicted the findings of early smaller retrospective studies. We summarize the results of large modern epidemiological studies (case-control, cohort studies and meta-analyses) concerning sex hormones and RA in Tables 1–3 (43–64). Table 1 shows studies which found an association between endogenous hormonal factors and risk or course of RA, Table 2 lists the reports which found no such association, and Table 3 shows studies concerning exogenous hormonal factors and RA (Table 3) (nb, some studies are listed more than once where appropriate).

Table 1. Epidemiological studies showing an association between endogenous sex hormones and rheumatoid arthritis pathogenesis.

Table 2. Epidemiological studies showing no association between endogenous sex hormones and RA pathogenesis.
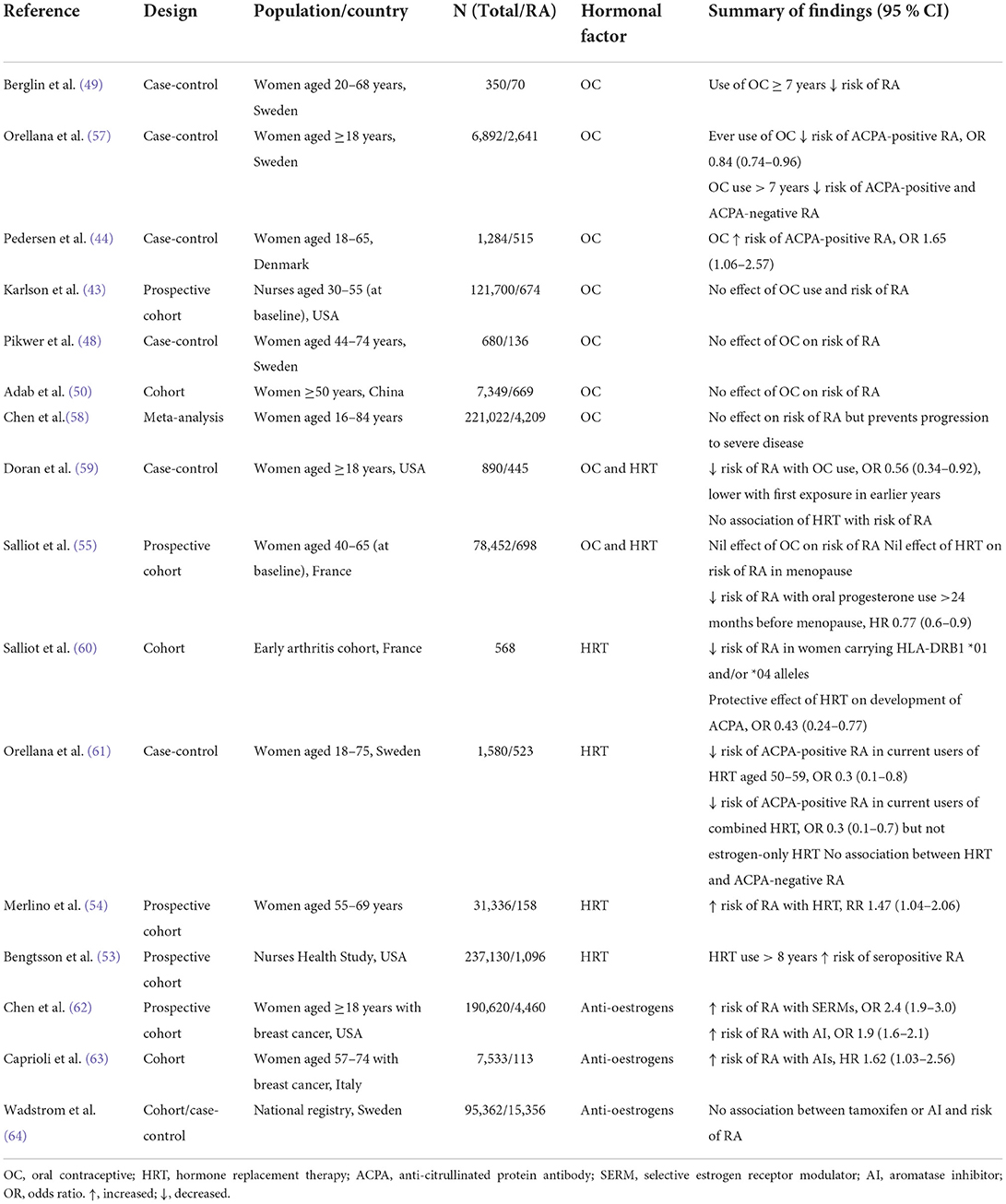
Table 3. Epidemiological studies of exogenous sex hormones and RA pathogenesis.
Endogenous sex hormones
Menarche
An abnormally early menarche was found to be associated with an increased risk of seropositive RA in a large prospective cohort study [relative risk (RR) 1.6 [95% confidence interval (CI) 1.1–2.4] for menarche <10 years] (43). This finding was supported by a more recent cohort study, although the association was weaker (HR) 1.20 for menarche <13 years (95% CI 0.9 - 1.5) (55). The data on menarche is conflicting, however, as a case-control study found an increased risk of anti-citrullinated protein antibody (ACPA)-positive RA with menarche at ≥15 vs. ≤ 12 years (44).
Pregnancy and post-partum
Hormonal changes in pregnancy
Pregnancy is associated with considerable increases in both estrogen (around four to six fold) and progesterone (around three to eight fold) (65). These hormones rise progressively through pregnancy, peaking at the third trimester and falling within weeks post-partum.
Pregnancy, parity and risk of RA
In an initial small case-control study, pregnancy was associated with a reduced risk of RA onset which did not reach statistical significance [odds ratio (OR) 0.3, 95% CI 0.04–2.6], while there was an increased incidence of RA during the first 3 months post-partum (OR 5.6, 95% CI 1.8–17.6) (66). Similar results were found in another early case-control study (67). The risk of RA onset has been shown to persist for the first year (OR 3.8, 95% CI 1.45–9.93) (68) and up to 24 months post-delivery (incidence rate ratio 1.7, 95% CI 1.11–2.70) (69).
In early studies, nulliparity was associated with a roughly two-fold risk of RA onset (70), but modern studies have shown conflicting findings regarding parity and risk of RA. Some studies have found that having more than one pregnancy increased the risk of RA (46), particularly with young age at first pregnancy (45, 46), while others have found a protective effect of parity (45, 47, 71). One retrospective cohort study found a dose-response relationship between the protective effect of parity and risk of RA (45), but this was not replicated in a later meta-analysis (47). The most recent systematic review found no association between gravidity and parity and the risk of developing RA (56).
Pregnancy and RA disease activity
RA was first observed to spontaneously remit in pregnancy in the 1930s (72). Historical studies reported that up to 90% of patients improved during pregnancy, but this effect is less apparent with modern treatment regimens (3, 73). As safety data regarding the use of traditional and biologic disease modifying drugs in pregnancy accumulates (74, 75), increasing numbers of patients maintain low disease activity or remission through pregnancy with a treat-to-target strategy (76).
Recent systematic reviews of the potential mechanisms of RA disease improvement in pregnancy have not identified any human studies that have specifically investigated the impact of rises in estrogen or progesterone on alterations of RA disease activity in pregnancy (77, 78).
Breastfeeding
In an early study, breastfeeding was associated with post-partum flare in inflammatory polyarthritis; this finding led the authors to suggest that prolactin may be responsible for this phenomenon (79). Since then, duration of breastfeeding has been most consistently associated with a decreased risk of RA (48, 50, 51). Other case-control (57) and large prospective cohort (43, 55) studies have found no significant association between breastfeeding and risk of RA, while a single study identified an increased risk (49). The data regarding the role of prolactin in human RA is limited and conflicting (80), but in a murine model of collagen-induced arthritis, treatment with bromocriptine, an inhibitor of prolactin secretion, suppressed post-partum flare (81).
Menopause
A consistent finding is the increased risk of RA in early menopause (52, 82, 83). In the large Nurses Health Study cohort, menopause at <44 years increased the risk of seronegative RA [hazard ratio (HR) 2.4, 95% CI 1.5–4.0] (53). The menopause has also been associated with the development of ACPA in first degree relatives of patients with RA (84).
Androgens
In men with RA, low serum levels of testosterone were found to be strongly predictive of seronegative disease (OR 0.31, 95% CI 0.12–0.85) but not significantly predictive of seropositive disease (85). Men with untreated hypogonadism have been found to be at increased risk of a range of autoimmune diseases, including RA (HR 1.31, 95% CI 1.22–1.44) (86), as are men with Klinefelter syndrome (RR 3.3, 95% CI 2.0–5.2)(87).
Exogenous sex hormones
Oral contraceptives
There are conflicting reports on the effect of oral contraceptives (OC) on the risk of RA.
Early reports suggested a beneficial effect, with a case-control study (n = 115) showing lower current use of OC in new cases of inflammatory polyarthritis (OR 0.22, 95% CI 0.06–0.85) (88). A further case control study found that OC use roughly halved the risk of RA, with stronger protection from earlier OC preparations (59). A matched case-control study found, conversely, an increased risk of ACPA-positive RA with OC use (44). Most recent reports, however, show no such effect (43, 50, 58, 89), although a meta-analysis has suggested that OC may reduce the risk of progression to severe disease in established RA (58). Two studies have suggested that extended OC use (>7 years) may protect against RA (49, 57). The differing findings regarding OC use and risk of RA have been suggested to be due to a lowering of their estrogen content over time (59, 90).
Hormone replacement therapy
Use of hormone replacement therapy (HRT) was associated with a significantly elevated risk of RA in a large prospective study of postmenopausal women (RR 1.47, 95% CI 1.04–2.06) (54). Conversely, a case-control study found a reduced risk of ACPA-positive RA in women aged 50–70 years who were current users of combined (estrogen plus progestogen) HRT (OR 0.3, 95% CI 0.1–0.7) (61). This finding supported that of an earlier cohort study which found a protective effect of HRT on the development of ACPA in early arthritis (60). Other studies have found no association between HRT use and risk of RA (59).
Anti-estrogen agents
Selective estrogen receptor modulators (SERMs) and aromatase inhibitors (AI) are used for the treatment of breast cancer. SERMs competitively inhibit estrogen binding to its receptor and may have agonistic or antagonistic properties in different target tissues, while AI reduce endogenous production of estrogen. A large national database study found an increased incidence of RA with use of either of these agents (OR 2.4 for SERMs (95% CI 1.9–3.0) and 1.9 (95% CI 1.6–2.1) for AI, each with >12 months treatment) (62). In another large population-based study, the use of aromatase inhibitors to treat breast cancer was associated with an increased risk of RA (adjusted HR 1.62, 95% CI 1.03–2.56) (63). In contrast, a recent large national registry study found no increased risk of RA with either tamoxifen (a SERM) or AI (64).
Clinical trials of sex hormones in RA
Estrogen and progesterone
Investigations into the therapeutic potential of sex hormones in RA have been disappointing. A randomized placebo-controlled trial of adjuvant estrogen therapy at physiological doses compatible with pregnancy in postmenopausal female RA patients was negative (91). Randomized controlled trials of ERα (92) and ERβ (93) agonists in RA were both negative. Large randomized control trials of HRT (either estrogen or estrogen and progesterone combined) in postmenopausal women with RA failed to demonstrate any benefit on symptom severity over placebo (94).
Interestingly, however, the data emerging from the field of multiple sclerosis, another disease that improves in pregnancy, is more encouraging regarding the potential therapeutic benefit of estrogen: a placebo-controlled phase 2 trial of estriol met its endpoint of a significant reduction in relapse rate (95). Progesterone or its synthetic derivatives have not been studied in clinical trials as a treatment option in RA.
Androgens
Well-designed trials of supplemental androgen therapy in patients with RA are lacking. Two preliminary studies of treatment with testosterone in the 1990s suggested positive results in male (n = 7) (96) and postmenopausal female (97) patients (n = 107). Meanwhile, two trials of dehydroepiandrosterone (DHEA) have been published. A small open-label trial of DHEA treatment in elderly RA patients (six post-menopausal female and five male) found no benefit on disease activity (98), while a randomized placebo-controlled trial of DHEA in pre-menopausal RA patients (n = 46) found improvements in quality of life but not disease activity scores (99).
Discussion
Despite extensive work in both basic laboratory and clinical studies, the exact impact of sex hormones on RA pathogenesis remains controversial. The epidemiological data described above is conflicting, with both high and low estrogen/progesterone states being found to be protective or risk factors, or to have no effect on development or progression of RA, in different studies. Namely, age of menarche, age of menopause, parity status, breastfeeding, use of the oral contraceptive, and HRT have all been found by different studies to have opposing effects on the risk of RA. Meta-analyses pertaining to parity and risk of RA found only a borderline, or zero, association (47, 56), as did that regarding the oral contraceptive (58). In contrast, the studies relating to breastfeeding have been more consistent, in that only one case-control study found an increased risk of RA with breastfeeding (49), while several others have found a protective effect, including four reporting a dose-response relationship (43, 48, 50, 51). These discrepancies are likely explained to an extent by heterogeneity in study design, variations in study populations and definitions of reproductive variables (e.g., early menarche), and inherent limitations in case-control studies such as recall bias and lack of consideration of confounding variables. On a biological level, many environmental and genetic factors may influence sex steroid signaling via their intracellular receptors, which may be another reason for the conflicting data from epidemiological studies.
Despite discrepancies in the published literature, several patterns do emerge. The most consistent findings are the increased risk of RA at early menopause and post-partum, and decreased disease activity in pregnancy. Therefore, declining estrogen and/or progesterone levels (in post-partum and menopause) are consistently linked to the onset of RA, while high levels of these hormones are protective during pregnancy (although many other factors may be relevant to reduced disease activity in pregnancy, as we and others have previously noted).
While there is an abundance of epidemiological data regarding reproductive factors and risk of RA, one possible avenue of research which, to our knowledge, has not been explored is the study of individuals with gender dysphoria [except a single case report (100)]. For instance, it would be intriguing to study whether the risk of RA in transgender individuals is modified by gender affirming therapy, or whether disease activity of pre-existing RA is altered by such treatments. This topic is discussed in detail elsewhere in this chapter.
There is a wealth of basic scientific data demonstrating the immunomodulatory actions of oestrogens, progestogens and androgens. The therapeutic potential of these hormones for treating RA suggested by results from animal models has to date not translated into successful clinical trials. In humans it is likely that there is a much more complex interaction between sex hormones and a multitude of genetic and environmental risk factors (e.g., smoking, obesity, alcohol consumption) for RA. Progestogens and androgens both exhibit more broadly anti-inflammatory actions than those of oestrogens and it is interesting to note the recent finding that perimenopausal oral progestogen use reduced the risk of RA (55) and that combined HRT, but not estrogen alone, strongly reduced the risk of ACPA-positive disease (61). However, neither of these hormones have been evaluated in large well-designed trials of RA, perhaps due to concern regarding the potential side effects of systemic administration.
It is important to point that there are other sex-related factors which have been proposed as being of potential importance in RA pathogenesis that we have not considered in the present review, including microchimerism (101), sex chromosomes (102) and sex differences in gut microbiota (103). However, none of these other factors have to date shown such direct links with RA disease onset and/or progression.
In terms of future study, the rapidly expanding field of high throughput multiomics technologies (e.g., genomics, transcriptomics, proteomics and metabolomics), particularly at the single cell level, is starting to dissect the pathobiological basis of clinical heterogeneity in human disease, including in RA (104). With both epidemiological and in-vitro studies proving ultimately insufficient to unravel the relationship between sex hormones and RA pathogenesis, the application of these novel techniques are a tantalizing proposition for investigators in this area.
Conclusion
Sex hormones are immunomodulatory with pleiotropic effects on the immune system. There are conflicting reports regarding endogenous and exogenous sex hormones and RA pathogenesis, but declining estrogen levels in the menopause and post-partum are consistently associated with an increased risk and severity of RA. These findings, however, have not translated into improved therapies in RA, although progesterone and androgens warrant further evaluation as potential therapeutic agents in clinical trials.
Author contributions
CR and IG conceived of the article. CR performed the literature search, reviewed the source papers, and drafted the manuscript. IG independently performed the search, contributed to inclusion/exclusion of source literature, and edited the final manuscript. All authors contributed to the article and approved the submitted version.
Funding
CR received support from the British Society of Rheumatology (BSR CID-2146992), the Rosetrees Trust (M707), University College London, and an Investigator Initiated Studies grant from UCB Biopharma SRL (UCB: IIS-2019-134708). IG has received speaker's fees and travel fees from UCB. The funders were not involved in the study design, collection, analysis, interpretation of data, the writing of this article or the decision to submit it for publication.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher's note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Cross M, Smith E, Hoy D, Carmona L, Wolfe F, Vos T, et al. The global burden of rheumatoid arthritis: estimates from the global burden of disease 2010 study. Ann Rheum Dis. (2014) 73:1316–22. doi: 10.1136/annrheumdis-2013-204627
2. Goemaere S, Ackerman C, Goethals K, De Keyser F, Van der Straeten C, Verbruggen G, et al. Onset of symptoms of rheumatoid arthritis in relation to age, sex and menopausal transition. J Rheumatol. (1990) 17:1620–2.
3. Jethwa H, Lam S, Smith C, Giles I. Does rheumatoid arthritis really improve during pregnancy? A systematic review and metaanalysis. J Rheumatol. (2019) 46:245–50. doi: 10.3899/jrheum.180226
4. Smolen JS, Aletaha D, McInnes IB. Rheumatoid arthritis. Lancet. (2016) 388:2023–38. doi: 10.1016/S0140-6736(16)30173-8
5. Smolen JS, Aletaha D, Barton A, Burmester GR, Emery P, Firestein GS, et al. Rheumatoid arthritis. Nat Rev Dis Primers. (2018) 4:18001. doi: 10.1038/nrdp.2018.1
6. Choy E. Understanding the dynamics: pathways involved in the pathogenesis of rheumatoid arthritis. Rheumatology (Oxford). (2012) 51:v3–11. doi: 10.1093/rheumatology/kes113
7. Deane KD, Holers VM. Rheumatoid arthritis pathogenesis, prediction, and prevention: an emerging paradigm shift. Arthritis Rheumatol. (2021) 73:181–93. doi: 10.1002/art.41417
8. Smolen JS, Aletaha D, Koeller M, Weisman MH, Emery P. New therapies for treatment of rheumatoid arthritis. Lancet. (2007) 370:1861–74. doi: 10.1016/S0140-6736(07)60784-3
9. Weyand CM, Hicok KC, Conn DL, Goronzy JJ. The influence of HLA-DRB1 genes on disease severity in rheumatoid arthritis. Ann Intern Med. (1992) 117:801–6. doi: 10.7326/0003-4819-117-10-801
10. Klareskog L, Padyukov L, Alfredsson L. Smoking as a trigger for inflammatory rheumatic diseases. Curr Opin Rheumatol. (2007) 19:49–54. doi: 10.1097/BOR.0b013e32801127c8
11. Fuentes N, Silveyra P. Estrogen receptor signaling mechanisms. Adv Protein Chem Struct Biol. (2019) 116:135–70. doi: 10.1016/bs.apcsb.2019.01.001
12. Michmerhuizen AR, Spratt DE, Pierce LJ, Speers CW. ARe we there yet? Understanding androgen receptor signaling in breast cancer. NPJ Breast Cancer. (2020) 6:47. doi: 10.1038/s41523-020-00190-9
13. Grimm SL, Hartig SM, Edwards DP. Progesterone receptor signaling mechanisms. J Mol Biol. (2016) 428:3831–49. doi: 10.1016/j.jmb.2016.06.020
14. Sanchez-Maldonado JM, Caliz R, Canet L, Horst RT, Bakker O, den Broeder AA, et al. Steroid hormone-related polymorphisms associate with the development of bone erosions in rheumatoid arthritis and help to predict disease progression: results from the REPAIR consortium. Sci Rep. (2019) 9:14812. doi: 10.1038/s41598-019-51255-0
15. Canet LM, Sanchez-Maldonado JM, Caliz R, Rodriguez-Ramos A, Lupianez CB, Canhao H, et al. Polymorphisms at phase I-metabolizing enzyme and hormone receptor loci influence the response to anti-TNF therapy in rheumatoid arthritis patients. Pharmacogenomics J. (2019) 19:83–96. doi: 10.1038/s41397-018-0057-x
16. Stark K, Straub RH, Rovensky J, Blazickova S, Eiselt G, Schmidt M. CYB5A polymorphism increases androgens and reduces risk of rheumatoid arthritis in women. Arthritis Res Ther. (2015) 17:56. doi: 10.1186/s13075-015-0574-9
17. Hughes GC. Progesterone and autoimmune disease. Autoimmun Rev. (2012) 11:A502–14. doi: 10.1016/j.autrev.2011.12.003
18. Wilder RL, Elenkov IJ. Hormonal regulation of tumor necrosis factor-alpha, interleukin-12 and interleukin-10 production by activated macrophages. A disease-modifying mechanism in rheumatoid arthritis and systemic lupus erythematosus? Ann N Y Acad Sci. (1999) 876:14–31. doi: 10.1111/j.1749-6632.1999.tb07619.x
19. Hughes GC, Choubey D. Modulation of autoimmune rheumatic diseases by oestrogen and progesterone. Nat Rev Rheumatol. (2014) 10:740–51. doi: 10.1038/nrrheum.2014.144
20. Kanik KS, Wilder RL. Hormonal alterations in rheumatoid arthritis, including the effects of pregnancy. Rheum Dis Clin North Am. (2000) 26:805–23. doi: 10.1016/S0889-857X(05)70170-8
21. Cutolo M, Straub RH. Sex steroids and autoimmune rheumatic diseases: state of the art. Nat Rev Rheumatol. (2020) 16:628–44. doi: 10.1038/s41584-020-0503-4
22. Moulton VR. Sex Hormones in acquired immunity and autoimmune disease. Front Immunol. (2018) 9:2279. doi: 10.3389/fimmu.2018.02279
23. Pauklin S, Sernandez IV, Bachmann G, Ramiro AR, Petersen-Mahrt SK. Estrogen directly activates AID transcription and function. J Exp Med. (2009) 206:99–111. doi: 10.1084/jem.20080521
24. Dragin N, Bismuth J, Cizeron-Clairac G, Biferi MG, Berthault C, Serraf A, et al. Estrogen-mediated downregulation of AIRE influences sexual dimorphism in autoimmune diseases. J Clin Invest. (2016) 126:1525–37. doi: 10.1172/JCI81894
25. Dupuis ML, Conti F, Maselli A, Pagano MT, Ruggieri A, Anticoli S, et al. The natural agonist of estrogen receptor beta silibinin plays an immunosuppressive role representing a potential therapeutic tool in rheumatoid arthritis. Front Immunol. (2018) 9:1903. doi: 10.3389/fimmu.2018.01903
26. Tong WW, Zhang C, Hong T, Liu DH, Wang C, Li J, et al. Silibinin alleviates inflammation and induces apoptosis in human rheumatoid arthritis fibroblast-like synoviocytes and has a therapeutic effect on arthritis in rats. Sci Rep. (2018) 8:3241. doi: 10.1038/s41598-018-21674-6
27. Correale J, Arias M, Gilmore W. Steroid hormone regulation of cytokine secretion by proteolipid protein-specific CD4+ T cell clones isolated from multiple sclerosis patients and normal control subjects. J Immunol. (1998) 161:3365–74.
28. Adurthi S, Kumar MM, Vinodkumar HS, Mukherjee G, Krishnamurthy H, Acharya KK, et al. Oestrogen Receptor-alpha binds the FOXP3 promoter and modulates regulatory T-cell function in human cervical cancer. Sci Rep. (2017) 7:17289. doi: 10.1038/s41598-017-17102-w
29. Ganesan K, Balachandran C, Manohar BM, Puvanakrishnan R. Comparative studies on the interplay of testosterone, estrogen and progesterone in collagen induced arthritis in rats. Bone. (2008) 43:758–65. doi: 10.1016/j.bone.2008.05.025
30. Mattsson R, Mattsson A, Holmdahl R, Whyte A, Rook GA. Maintained pregnancy levels of oestrogen afford complete protection from post-partum exacerbation of collagen-induced arthritis. Clin Exp Immunol. (1991) 85:41–7. doi: 10.1111/j.1365-2249.1991.tb05679.x
31. Inoue K, Inoue E, Imai Y. Female sex hormones ameliorate arthritis in SKG mice. Biochem Biophys Res Commun. (2013) 434:740–5. doi: 10.1016/j.bbrc.2013.03.111
32. Subramanian S, Tovey M, Afentoulis M, Krogstad A, Vandenbark AA, Offner H. Ethinyl estradiol treats collagen-induced arthritis in DBA/1LacJ mice by inhibiting the production of TNF-alpha and IL-1beta. Clin Immunol. (2005) 115:162–72. doi: 10.1016/j.clim.2005.01.006
33. Polanczyk MJ, Hopke C, Huan J, Vandenbark AA, Offner H. Enhanced FoxP3 expression and Treg cell function in pregnant and estrogen-treated mice. J Neuroimmunol. (2005) 170:85–92. doi: 10.1016/j.jneuroim.2005.08.023
34. Szereday L, Varga P, Szekeres-Bartho J. Cytokine production by lymphocytes in pregnancy. Am J Reprod Immunol. (1997) 38:418–22. doi: 10.1111/j.1600-0897.1997.tb00321.x
35. Lee JH, Lydon JP, Kim CH. Progesterone suppresses the mTOR pathway and promotes generation of induced regulatory T cells with increased stability. Eur J Immunol. (2012) 42:2683–96. doi: 10.1002/eji.201142317
36. Lee JH, Ulrich B, Cho J, Park J, Kim CH. Progesterone promotes differentiation of human cord blood fetal T cells into T regulatory cells but suppresses their differentiation into Th17 cells. J Immunol. (2011) 187:1778–87. doi: 10.4049/jimmunol.1003919
37. Aristimuno C, Teijeiro R, Valor L, Alonso B, Tejera-Alhambra M, de Andres C, et al. Sex-hormone receptors pattern on regulatory T-cells: clinical implications for multiple sclerosis. Clin Exp Med. (2012) 12:247–55. doi: 10.1007/s10238-011-0172-3
38. Hellberg S, Raffetseder J, Rundquist O, Magnusson R, Papapavlou G, Jenmalm MC, et al. Progesterone dampens immune responses in in vitro activated CD4(+) T cells and affects genes associated with autoimmune diseases that improve during pregnancy. Front Immunol. (2021) 12:672168. doi: 10.3389/fimmu.2021.672168
39. Gubbels Bupp MR, Jorgensen TN. Androgen-Induced Immunosuppression. Front Immunol. (2018) 9:794. doi: 10.3389/fimmu.2018.00794
40. Cutolo M, Wilder RL. Different roles for androgens and estrogens in the susceptibility to autoimmune rheumatic diseases. Rheum Dis Clin North Am. (2000) 26:825–39. doi: 10.1016/S0889-857X(05)70171-X
41. Zhu ML, Bakhru P, Conley B, Nelson JS, Free M, Martin A, et al. Sex bias in CNS autoimmune disease mediated by androgen control of autoimmune regulator. Nat Commun. (2016) 7:11350. doi: 10.1038/ncomms11350
42. Walecki M, Eisel F, Klug J, Baal N, Paradowska-Dogan A, Wahle E, et al. Androgen receptor modulates Foxp3 expression in CD4+CD25+Foxp3+ regulatory T-cells. Mol Biol Cell. (2015) 26:2845–57. doi: 10.1091/mbc.E14-08-1323
43. Karlson EW, Mandl LA, Hankinson SE, Grodstein F. Do breast-feeding and other reproductive factors influence future risk of rheumatoid arthritis? Results from the nurses' health study. Arthritis Rheum. (2004) 50:3458–67. doi: 10.1002/art.20621
44. Pedersen M, Jacobsen S, Klarlund M, Pedersen BV, Wiik A, Wohlfahrt J, et al. Environmental risk factors differ between rheumatoid arthritis with and without auto-antibodies against cyclic citrullinated peptides. Arthritis Res Ther. (2006) 8:R133. doi: 10.1186/ar2022
45. Jorgensen KT, Pedersen BV, Jacobsen S, Biggar RJ, Frisch M. National cohort study of reproductive risk factors for rheumatoid arthritis in Denmark: a role for hyperemesis, gestational hypertension and pre-eclampsia? Ann Rheum Dis. (2010) 69:358–63. doi: 10.1136/ard.2008.099945
46. Orellana C, Wedren S, Kallberg H, Holmqvist M, Karlson EW, Alfredsson L, et al. Parity and the risk of developing rheumatoid arthritis: results from the Swedish Epidemiological Investigation of Rheumatoid Arthritis study. Ann Rheum Dis. (2014) 73:752–5. doi: 10.1136/annrheumdis-2013-203567
47. Ren L, Guo P, Sun QM, Liu H, Chen Y, Huang Y, et al. Number of parity and the risk of rheumatoid arthritis in women: a dose-response meta-analysis of observational studies. J Obstet Gynaecol Res. (2017) 43:1428–40. doi: 10.1111/jog.13370
48. Pikwer M, Bergstrom U, Nilsson JA, Jacobsson L, Berglund G, Turesson C. Breast feeding, but not use of oral contraceptives, is associated with a reduced risk of rheumatoid arthritis. Ann Rheum Dis. (2009) 68:526–30. doi: 10.1136/ard.2007.084707
49. Berglin E, Kokkonen H, Einarsdottir E, Agren A, Rantapaa Dahlqvist S. Influence of female hormonal factors, in relation to autoantibodies and genetic markers, on the development of rheumatoid arthritis in northern Sweden: a case-control study. Scand J Rheumatol. (2010) 39:454–60. doi: 10.3109/03009741003742763
50. Adab P, Jiang CQ, Rankin E, Tsang YW, Lam TH, Barlow J, et al. Breastfeeding practice, oral contraceptive use and risk of rheumatoid arthritis among Chinese women: the Guangzhou Biobank Cohort Study. Rheumatology (Oxford). (2014) 53:860–6. doi: 10.1093/rheumatology/ket456
51. Chen H, Wang J, Zhou W, Yin H, Wang M. Breastfeeding and risk of rheumatoid arthritis: a systematic review and metaanalysis. J Rheumatol. (2015) 42:1563–9. doi: 10.3899/jrheum.150195
52. Beydoun HA. el-Amin R, McNeal M, Perry C, Archer DF. Reproductive history and postmenopausal rheumatoid arthritis among women 60 years or older: Third National Health and Nutrition Examination Survey. Menopause. (2013) 20:930–5. doi: 10.1097/GME.0b013e3182a14372
53. Bengtsson C, Malspeis S, Orellana C, Sparks JA, Costenbader KH, Karlson EW. Association between menopausal factors and the risk of seronegative and seropositive rheumatoid arthritis: results from the nurses' health studies. Arthritis Care Res (Hoboken). (2017) 69:1676–84. doi: 10.1002/acr.23194
54. Merlino LA, Cerhan JR, Criswell LA, Mikuls TR, Saag KG. Estrogen and other female reproductive risk factors are not strongly associated with the development of rheumatoid arthritis in elderly women. Semin Arthritis Rheum. (2003) 33:72–82. doi: 10.1016/S0049-0172(03)00084-2
55. Salliot C, Nguyen Y, Gusto G, Gelot A, Gambaretti J, Mariette X, et al. Female hormonal exposures and risk of rheumatoid arthritis in the French E3N-EPIC cohort study. Rheumatology (Oxford). (2021) 60:4790–800. doi: 10.1093/rheumatology/keab101
56. Chen WMY, Subesinghe S, Muller S, Hider SL, Mallen CD, Scott IC. The association between gravidity, parity and the risk of developing rheumatoid arthritis: a systematic review and meta-analysis. Semin Arthritis Rheum. (2020) 50:252–60. doi: 10.1093/rheumatology/kez107.028
57. Orellana C, Saevarsdottir S, Klareskog L, Karlson EW, Alfredsson L, Bengtsson C. Oral contraceptives, breastfeeding and the risk of developing rheumatoid arthritis: results from the Swedish EIRA study. Ann Rheum Dis. (2017) 76:1845–52. doi: 10.1136/annrheumdis-2017-211620
58. Chen Q, Jin Z, Xiang C, Cai Q, Shi W, He J. Absence of protective effect of oral contraceptive use on the development of rheumatoid arthritis: a meta-analysis of observational studies. Int J Rheum Dis. (2014) 17:725–37. doi: 10.1111/1756-185X.12413
59. Doran MF, Crowson CS, O'Fallon WM, Gabriel SE. The effect of oral contraceptives and estrogen replacement therapy on the risk of rheumatoid arthritis: a population based study. J Rheumatol. (2004) 31:207–13.
60. Salliot C, Bombardier C, Saraux A, Combe B, Dougados M. Hormonal replacement therapy may reduce the risk for RA in women with early arthritis who carry HLA-DRB1 *01 and/or *04 alleles by protecting against the production of anti-CCP: results from the ESPOIR cohort. Ann Rheum Dis. (2010) 69:1683–6. doi: 10.1136/ard.2009.111179
61. Orellana C, Saevarsdottir S, Klareskog L, Karlson EW, Alfredsson L, Bengtsson C. Postmenopausal hormone therapy and the risk of rheumatoid arthritis: results from the Swedish EIRA population-based case-control study. Eur J Epidemiol. (2015) 30:449–57. doi: 10.1007/s10654-015-0004-y
62. Chen JY, Ballou SP. The effect of antiestrogen agents on risk of autoimmune disorders in patients with breast cancer. J Rheumatol. (2015) 42:55–9. doi: 10.3899/jrheum.140367
63. Caprioli M, Carrara G, Sakellariou G, Silvagni E, Scire CA. Influence of aromatase inhibitors therapy on the occurrence of rheumatoid arthritis in women with breast cancer: results from a large population-based study of the Italian Society for Rheumatology. RMD Open. (2017) 3:e000523. doi: 10.1136/rmdopen-2017-000523
64. Wadstrom H, Pettersson A, Smedby KE, Askling J. Risk of breast cancer before and after rheumatoid arthritis, and the impact of hormonal factors. Ann Rheum Dis. (2020) 79:581–6. doi: 10.1136/annrheumdis-2019-216756
65. Amin S, Peterson EJ, Reed AM, Mueller DL. Pregnancy and rheumatoid arthritis: insights into the immunology of fetal tolerance and control of autoimmunity. Curr Rheumatol Rep. (2011) 13:449–55. doi: 10.1007/s11926-011-0199-1
66. Silman A, Kay A, Brennan P. Timing of pregnancy in relation to the onset of rheumatoid arthritis. Arthritis Rheum. (1992) 35:152–5. doi: 10.1002/art.1780350205
67. Lansink M, de Boer A, Dijkmans BA, Vandenbroucke JP, Hazes JM. The onset of rheumatoid arthritis in relation to pregnancy and childbirth. Clin Exp Rheumatol. (1993) 11:171–4.
68. Peschken CA, Robinson DB, Hitchon CA, Smolik I, Hart D, Bernstein CN, et al. Pregnancy and the risk of rheumatoid arthritis in a highly predisposed North American Native population. J Rheumatol. (2012) 39:2253–60. doi: 10.3899/jrheum.120269
69. Wallenius M, Skomsvoll JF, Irgens LM, Salvesen KA, Koldingsnes W, Mikkelsen K, et al. Postpartum onset of rheumatoid arthritis and other chronic arthritides: results from a patient register linked to a medical birth registry. Ann Rheum Dis. (2010) 69:332–6. doi: 10.1136/ard.2009.115964
70. Silman AJ. Parity status and the development of rheumatoid arthritis. Am J Reprod Immunol. (1992) 28:228–30. doi: 10.1111/j.1600-0897.1992.tb00799.x
71. Guthrie KA, Dugowson CE, Voigt LF, Koepsell TD, Nelson JL. Does pregnancy provide vaccine-like protection against rheumatoid arthritis? Arthritis Rheum. (2010) 62:1842–8. doi: 10.1002/art.27459
72. Hench P editor editor The ameliorating effect of pregnancy on chronic atropic infections rheumatoid arthritis fibrosititis and intermittent hydrarthrosis. Boca Raton, FL: CRC Press (1938).
73. Hazes JM, Coulie PG, Geenen V, Vermeire S, Carbonnel F, Louis E, et al. Rheumatoid arthritis and pregnancy: evolution of disease activity and pathophysiological considerations for drug use. Rheumatology (Oxford). (2011) 50:1955–68. doi: 10.1093/rheumatology/ker302
74. Flint J, Panchal S, Hurrell A, van de Venne M, Gayed M, Schreiber K, et al. BSR and BHPR guideline on prescribing drugs in pregnancy and breastfeeding-Part I: standard and biologic disease modifying anti-rheumatic drugs and corticosteroids. Rheumatology (Oxford). (2016) 55:1693–7. doi: 10.1093/rheumatology/kev404
75. Gotestam Skorpen C, Hoeltzenbein M, Tincani A, Fischer-Betz R, Elefant E, Chambers C, et al. The EULAR points to consider for use of antirheumatic drugs before pregnancy, and during pregnancy and lactation. Ann Rheum Dis. (2016) 75:795–810. doi: 10.1136/annrheumdis-2015-208840
76. Smeele HT, Roder E, Wintjes HM. Kranenburg-van Koppen LJ, Hazes JM, Dolhain RJ. Modern treatment approach results in low disease activity in 90% of pregnant rheumatoid arthritis patients: the PreCARA study. Ann Rheum Dis. (2021) 80:859–64. doi: 10.1136/annrheumdis-2020-219547
77. Raine C, Austin K, Giles I. Mechanisms determining the amelioration of rheumatoid arthritis in pregnancy: a systematic review. Semin Arthritis Rheum. (2020) 50:1357–69. doi: 10.1016/j.semarthrit.2020.03.006
78. Forger F, Villiger PM. Immunological adaptations in pregnancy that modulate rheumatoid arthritis disease activity. Nat Rev Rheumatol. (2020) 16:113–22. doi: 10.1038/s41584-019-0351-2
79. Barrett JH, Brennan P, Fiddler M, Silman A. Breast-feeding and postpartum relapse in women with rheumatoid and inflammatory arthritis. Arthritis Rheum. (2000) 43:1010–5. doi: 10.1002/1529-0131(200005)43:5<1010::AID-ANR8>3.0.CO;2-O
80. Tang MW, Garcia S, Gerlag DM, Tak PP, Reedquist KA. Insight into the endocrine system and the immune system: a review of the inflammatory role of prolactin in rheumatoid arthritis and psoriatic arthritis. Front Immunol. (2017) 8:720. doi: 10.3389/fimmu.2017.00720
81. Whyte A, Williams RO. Bromocriptine suppresses postpartum exacerbation of collagen-induced arthritis. Arthritis Rheum. (1988) 31:927–8. doi: 10.1002/art.1780310717
82. Pikwer M, Bergstrom U, Nilsson JA, Jacobsson L, Turesson C. Early menopause is an independent predictor of rheumatoid arthritis. Ann Rheum Dis. (2012) 71:378–81. doi: 10.1136/ard.2011.200059
83. Wong LE, Huang WT, Pope JE, Haraoui B, Boire G, Thorne JC, et al. Effect of age at menopause on disease presentation in early rheumatoid arthritis: results from the Canadian Early Arthritis Cohort. Arthritis Care Res (Hoboken). (2015) 67:616–23. doi: 10.1002/acr.22494
84. Alpizar-Rodriguez D, Mueller RB, Moller B, Dudler J, Ciurea A, Zufferey P, et al. Female hormonal factors and the development of anti-citrullinated protein antibodies in women at risk of rheumatoid arthritis. Rheumatology (Oxford). (2017) 56:1579–85. doi: 10.1093/rheumatology/kex239
85. Pikwer M, Giwercman A, Bergstrom U, Nilsson JA, Jacobsson LT, Turesson C. Association between testosterone levels and risk of future rheumatoid arthritis in men: a population-based case-control study. Ann Rheum Dis. (2014) 73:573–9. doi: 10.1136/annrheumdis-2012-202781
86. Baillargeon J, Al Snih S, Raji MA, Urban RJ, Sharma G, Sheffield-Moore M, et al. Hypogonadism and the risk of rheumatic autoimmune disease. Clin Rheumatol. (2016) 35:2983–7. doi: 10.1007/s10067-016-3330-x
87. Seminog OO, Seminog AB, Yeates D, Goldacre MJ. Associations between Klinefelter's syndrome and autoimmune diseases: English national record linkage studies. Autoimmunity. (2015) 48:125–8. doi: 10.3109/08916934.2014.968918
88. Brennan P, Bankhead C, Silman A, Symmons D. Oral contraceptives and rheumatoid arthritis: results from a primary care-based incident case-control study. Semin Arthritis Rheum. (1997) 26:817–23. doi: 10.1016/S0049-0172(97)80025-X
89. Liao KP, Alfredsson L, Karlson EW. Environmental influences on risk for rheumatoid arthritis. Curr Opin Rheumatol. (2009) 21:279–83. doi: 10.1097/BOR.0b013e32832a2e16
90. Benagiano G, Benagiano M, Bianchi P, D'Elios MM, Brosens I. Contraception in autoimmune diseases. Best Pract Res Clin Obstet Gynaecol. (2019) 60:111–23. doi: 10.1016/j.bpobgyn.2019.05.003
91. van den Brink HR, van Everdingen AA, van Wijk MJ, Jacobs JW, Bijlsma JW. Adjuvant oestrogen therapy does not improve disease activity in postmenopausal patients with rheumatoid arthritis. Ann Rheum Dis. (1993) 52:862–5. doi: 10.1136/ard.52.12.862
92. van Vollenhoven RF, Houbiers JG, Buttgereit F, In 't Hout J, Boers M, Leij S, et al. The selective estrogen receptor alpha agonist Org 37663 induces estrogenic effects but lacks antirheumatic activity: a phase IIa trial investigating efficacy and safety of Org 37663 in postmenopausal female rheumatoid arthritis patients receiving stable background methotrexate or sulfasalazine. Arthritis Rheum. (2010) 62:351–8. doi: 10.1002/art.27196
93. Roman-Blas JA, Castaneda S, Cutolo M, Herrero-Beaumont G. Efficacy and safety of a selective estrogen receptor beta agonist, ERB-041, in patients with rheumatoid arthritis: a 12-week, randomized, placebo-controlled, phase II study. Arthritis Care Res (Hoboken). (2010) 62:1588–93. doi: 10.1002/acr.20275
94. Walitt B, Pettinger M, Weinstein A, Katz J, Torner J, Wasko MC, et al. Effects of postmenopausal hormone therapy on rheumatoid arthritis: the women's health initiative randomized controlled trials. Arthritis Rheum. (2008) 59:302–10. doi: 10.1002/art.23325
95. Voskuhl RR, Wang H, Wu TC, Sicotte NL, Nakamura K, Kurth F, et al. Estriol combined with glatiramer acetate for women with relapsing-remitting multiple sclerosis: a randomised, placebo-controlled, phase 2 trial. Lancet Neurol. (2016) 15:35–46. doi: 10.1016/S1474-4422(15)00322-1
96. Cutolo M, Balleari E, Giusti M, Intra E, Accardo S. Androgen replacement therapy in male patients with rheumatoid arthritis. Arthritis Rheum. (1991) 34:1–5. doi: 10.1002/art.1780340102
97. Booji A, Biewenga-Booji CM, Huber-Bruning O, Cornelis C, Jacobs JW, Bijlsma JW. Androgens as adjuvant treatment in postmenopausal female patients with rheumatoid arthritis. Ann Rheum Dis. (1996) 55:811–5. doi: 10.1136/ard.55.11.811
98. Giltay EJ, van Schaardenburg D, Gooren LJ, von Blomberg BM, Fonk JC, Touw DJ, et al. Effects of dehydroepiandrosterone administration on disease activity in patients with rheumatoid arthritis. Br J Rheumatol. (1998) 37:705–6. doi: 10.1093/rheumatology/37.6.705
99. Sandoughi M, Kaykhaei MA, Langarizadeh E, Dashipour A. Effects of dehydroepiandrosterone on quality of life in premenopausal women with rheumatoid arthritis: A preliminary randomized clinical trial. Int J Rheum Dis. (2020) 23:1692–7. doi: 10.1111/1756-185X.13975
100. Troum OM, Quismorio FP. Jr. Rheumatoid arthritis in a male transsexual. J Rheumatol. (1985) 12:640–1.
101. Yan Z, Aydelotte T, Gadi VK, Guthrie KA, Nelson JL. Acquisition of the rheumatoid arthritis HLA shared epitope through microchimerism. Arthritis Rheum. (2011) 63:640–4. doi: 10.1002/art.30160
102. Lambert NC. Nonendocrine mechanisms of sex bias in rheumatic diseases. Nat Rev Rheumatol. (2019) 15:673–86. doi: 10.1038/s41584-019-0307-6
103. Gomez A, Luckey D, Taneja V. The gut microbiome in autoimmunity: Sex matters. Clin Immunol. (2015) 159:154–62. doi: 10.1016/j.clim.2015.04.016
Keywords: rheumatoid arthritis, estrogen, progesterone, androgens, pathogenesis, pregnancy
Citation: Raine C and Giles I (2022) What is the impact of sex hormones on the pathogenesis of rheumatoid arthritis? Front. Med. 9:909879. doi: 10.3389/fmed.2022.909879
Received: 31 March 2022; Accepted: 05 July 2022;
Published: 22 July 2022.
Edited by:
Veena Taneja, Mayo Clinic, United StatesReviewed by:
Maria Francisca Moraes Fontes, Fundação Champalimaud, PortugalMarco Krasselt, University Hospital Leipzig, Germany
Copyright © 2022 Raine and Giles. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Charles Raine, Yy5yYWluZUB1Y2wuYWMudWs=