 Hamzeh Izadi
Hamzeh Izadi Homa Asadi1
Homa Asadi1 Marjan Bemani
Marjan Bemani
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
REVIEW article
Front. Mater., 04 March 2025
Sec. Biomaterials and Bio-Inspired Materials
Volume 12 - 2025 | https://doi.org/10.3389/fmats.2025.1537067
Chitin is the second most abundant polysaccharide on Earth, after cellulose, and it is mainly obtained from the shells of crustaceans. While chitin is currently derived from shrimp shells, there is a growing interest in commercializing chitin sourced from insects. This review compares chitin from various sources, including crustaceans, insects, fungi, and mollusks, based on several factors: purity, molecular weight, crystallinity, cost, and sustainability. Although crustaceans yield higher amounts of chitin and exhibit better crystallinity, insects present significant advantages in terms of sustainability, ease of extraction, and lower impurity levels. Chitin derived from insects is emerging as a sustainable alternative due to its simpler extraction processes and reduced environmental impact. This work highlights the increasing potential of insects as a reliable source for sustainable chitin production.
Natural polymers like chitin, starch, cellulose, hemicellulose, and alginate are widely used in our daily lives to perform various essential functions (Silva et al., 2021; Muthukumaran et al., 2022). Polysaccharides and their derivatives, especially chitin and starch, attract considerable attention due to their extraordinary chemical properties. Starch and chitin are byproducts classified as modified polymers, derived biopolymers, and polysaccharide-containing materials (Crini, 2005). Chitin has the chemical formula (C8H13O5N)n. The structure of chitin is most similar to that of cellulose, while its function is most similar to that of keratin. There are minor structural differences between cellulose and chitin, but they are chemically identical. Chitin has an acetamide group at the C2 position instead of a hydroxyl group within the glucose unit (Hamed et al., 2016).
Chitin is a naturally occurring polysaccharide found abundantly in nature. Microfibrils of chitin are structural components found in the extracellular matrix of various invertebrates, including sponge skeletal fibers, mollusk shells, nematodes eggshell and pharynx, crustacean shells, arthropod exoskeletons, and fungi cell walls, forming natural biocomposites (Roberts, 1992; Ehrlich et al., 2007; Chen and Peng, 2019; Liu et al., 2019; Yu et al., 2024). Chitin in fungi is primarily sourced from the mycelium of species like Mucor rouxii, Aspergillus niger, and Lentinus edodes (Huq et al., 2022). Chitin makes up the exoskeleton, the wings, the antennae, and the trachea (Mei et al., 2024).
Chitosan and chitin are polysaccharides composed of N-acetyl-D-glucosamine and D-glucosamine units. Chitosan is derived from chitin through several methods, primarily involving chemical and enzymatic processes. Chemical methods are the predominant approach that involves the deproteination, demineralization, decolorization, and deacetylation of chitin (Gandhi et al., 2014; Ibitoye et al., 2018). Deacetylation is a process that eliminates acetyl groups from chitin’s molecular chain, yielding chitosan. The degree of deacetylation (DD) determines the extent of chitin conversion to chitosan (Kumar et al., 2020). DD varies based on factors such as NaOH concentration, reaction temperature, and duration. Chitosan’s DD can span from 56% to 99%, influenced by production techniques and species. Chitosan represents a partially deacetylated form of chitin, where a portion of N-acetyl-D-glucosamine transforms into D-glucosamine (Kumari and Kishor, 2020). Enzymatic methods utilize chitinases and chitosanases to transform chitin into chitosan. This method is increasingly attracting attention due to its ability to produce chitosan with a well-defined structure and a lower environmental impact compared to chemical methods (Kaczmarek et al., 2019; Pratiwi et al., 2023). In microbial methods, certain fungi, such as Pleurotus spp., can extract chitosan through fermentation processes. This approach is regarded as eco-friendly and can yield chitosan with notable antimicrobial properties (Johney et al., 2016). The efficiency, cost, and environmental impact of these methods differ significantly, with enzymatic and microbial approaches offering more sustainable alternatives to conventional chemical extraction. Chitin and chitosan are not present in mammals but can be degraded in the body by several proteases such as lysozyme, papain, and pepsin (Pangburn et al., 1982). Chitin and its derivatives, chitosan, are utilized in various fields such as water treatment, cosmetics, healthcare, and agrochemicals. Their biodegradable nature and non-toxic properties make them valuable for sustainable practices. They are employed in producing organic fertilizers that have been shown to enhance crop productivity. Chitin promotes plant growth by fostering beneficial microbial activity in the soil, enhancing nutrient availability and absorption by plants. This results in higher crop yields and improved plant health (Sharp, 2013; Malerba and Cerana, 2020a). Chitin and its derivatives enhance plants’ ability to withstand both biotic and abiotic stresses by activating their defense mechanisms. This protection helps safeguard crops against pathogens and environmental challenges, ultimately leading to increased productivity (Malerba and Cerana, 2020b; Shahrajabian et al., 2021). Chitin functions as a bioadsorbent, facilitating the remediation of contaminated soils by extracting heavy metals and enhancing soil characteristics. This process not only boosts soil fertility but also promotes sustainable agricultural practices (Malerba and Cerana, 2020b; Anedo et al., 2024). A recent study has shown that fertilizers enriched with chitin are effective in controlling nematode populations, which pose a threat to crops. This approach decreases the reliance on chemical nematicides, providing an environmentally friendly alternative (Kisaakye et al., 2024). Additionally, chitin is utilized in various industrial processes, including creating edible films and as a thickening and stabilizing agent in foods and food emulsions. Insoluble fibers, such as chitin, are indigestible in the human gastrointestinal tract. They are essential for increasing stool bulk and softness, which helps promote regular bowel movements and reduce intestinal transit time. Moreover, chitin serves as a dietary fiber that enhances gut microbiota, playing a crucial role in gut health. This enrichment can lead to improved gut function and may help prevent conditions such as constipation and hemorrhoids (Rodriguez et al., 2020; Kipkoech, 2023). However, insects offer an efficient method for chitin production, requiring significantly less land, water, and feed, which reduces their environmental impact. Their shorter life cycles and rapid growth allow for frequent harvesting, enhancing production efficiency. Additionally, insect farming is generally more cost-effective due to these lower resource demands and quicker growth rates, further decreasing overall production costs. Insect farming often uses organic waste as feed, promoting waste reduction and resource recycling in line with circular economy principles (Zainol Abidin et al., 2020a; Triunfo et al., 2022; Rehman et al., 2023; Saenz-Mendoza et al., 2023). These environmental and economic benefits make insect-based chitin a promising alternative to traditional sources. This review aims to qualitatively and quantitatively compare the primary sources of chitin production, focusing on their chemical properties, production methods, environmental impacts, and applications across various fields, including agriculture, healthcare, and industrial processes. It seeks to highlight the advantages and limitations of traditional and alternative sources, such as insect farming and evaluates sustainable and eco-friendly methods of chitin extraction.
Chitin is widely found in animal and plant kingdoms and is a crucial constituent of the exoskeletons of insects, crustaceans, and other arthropods. Its primary function is to provide a protective barrier for the inner soft tissues. Chitin acts as a watertight barrier, preventing delicate tissues from drying out. This helps to ensure the survival of these organisms by providing hydration protection to their inner tissues (Roberts, 1992; Moussian, 2019). The cuticles of insects, crustaceans, and spiders are mostly composed of sclerotized chitin, which can have diverse appearances. Chitin is also found in the iridophores of the eyes, the epidermis of cephalopods and arthropods, as well as the cuticles of some vertebrates (Zhang et al., 2021). It is also present in the cell walls of fungi and some algae. The exoskeleton of oysters and spiders, particularly Theraphosidae also contains chitin (Machałowski et al., 2020). Insects usually have 30%–45% protein, 25%–40% lipids, and 10%–15% chitin in their exoskeletons. In contrast, crustacean shells consist of 20%–40% protein, 20%–50% calcium carbonate, and 15%–40% chitin. Nevertheless, certain insects may contain more than 40% chitin in their exoskeletons (Spranghers et al., 2017; Mohan et al., 2020; Basawa et al., 2023).
Chitin is a major component of the insect’s external body, providing the necessary support and protection against various external factors, such as physical damage, predators, and environmental stresses. Therefore, chitin plays a significant role in insects’ survival, growth, and interaction with their surroundings. Insects heavily rely on chitin to form their cuticles and peritrophic membranes. However, the importance of chitin in insects can be summarized as follows: (Figure 1).
• Chitin provides structural support to insects’ exoskeletons, protecting internal organs and safeguarding them from physical damage, providing and maintaining rigidity and shape to their bodies.
•The chitinous exoskeleton provides physical, pathogenic, and predatory protection, aiding insects in surviving their environment.
•Chitin acts as a watertight barrier against dehydration, preventing excessive water loss from the body.
•Insects undergo molting, a process by which they shed their exoskeleton to grow. Chitin, a flexible yet durable material, plays a crucial role in this process and can be shed and regenerated during each molt.
•Chitin provides the support and structure for insects with wings for efficient flight
•Chitin is present in sensory structures such as antennae and sensory hairs, where it aids in detecting environmental cues and communicating with the insect’s surroundings.
•Chitin protects midgut epithelial cells and lines the foregut and hindgut, providing them with protection and support.
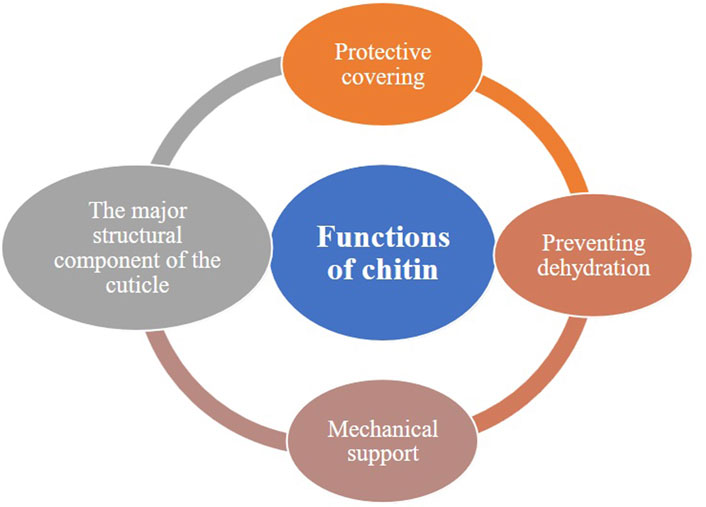
Figure 1. The primary functions of chitin.
Overall, the unique properties of chitin make it an essential component in various biological contexts, and its metabolism is tightly regulated during growth and morphogenesis (Merzendorfer and Zimoch, 2003; Yu et al., 2024).
Crustaceans are a diverse group of arthropods that include shrimp, lobsters, crabs, and crayfish, among others. One of the defining features of crustaceans is their exoskeleton, which is composed primarily of chitin, a polysaccharide that provides structural support and protection. In crustaceans, chitin is integrated within a matrix of proteins and mainly carbonate minerals, which provide structural integrity and rigidity to the exoskeleton. The main sources of chitin are the shells of shrimp, crabs, and lobsters, which constitute a significant portion of seafood processing waste. These shells typically contain 6.0%–25.0% chitin by weight, making them a rich extraction source (Tables 1, 2). Utilizing crustacean waste for chitin extraction helps reduce environmental pollution and supports sustainable practices by transforming waste into valuable products. However, crustaceans are a vital source of chitin due to their structural makeup, biosynthetic processes, and the potential for sustainable extraction methods (Younes and Rinaudo, 2015; Zhang et al., 2021; Amiri et al., 2022; Verardi et al., 2023).

Table 1. A comparison between animal and non-animal sources of chitin.

Table 2. Sources and properties of chitin.
Chitin is a long-chain polymer of N-acetylglucosamine, derived from glucose, and serves as a key structural component in the exoskeletons of arthropods, including insects. Insects are a significant natural source of chitin, constituting about 1.2%–60.0% of the dry weight of their cuticles, which vary by species and developmental stage (Merzendorfer and Zimoch, 2003; Hahn et al., 2020). Chitin forms a robust outer layer of the exoskeleton and is essential to the peritrophic matrix that lines the gut, acting as a barrier against pathogens and aiding digestion (Tables 1, 2). This polysaccharide provides structural integrity and rigidity to the insect exoskeleton while serving as an attachment matrix for cuticular proteins. Insects, such as beetles, crickets, flies, locusts, and mealworms, are commonly harvested for chitin extraction. Compared to crustaceans like shrimp and crabs, insects are easier to farm and process, making them an environmentally sustainable and attractive source of chitin, particularly in sustainable agriculture and waste reduction contexts (Merzendorfer and Zimoch, 2003; Hahn et al., 2020; Muthukrishnan et al., 2020).
Chitin is a versatile biopolymer found in various sources, each with distinct properties and applications (Figure 2) (Abo Elsoud and El Kady, 2019). Fungi are becoming an important source of chitin, with values reported of 8%–16% (w/w) chitin (plus chitosan) on a dry matter basis, mainly present in their cell walls. Notably, Zygomycetes fungi contain more chitin than other fungal groups. This chitin is covalently bonded to glucans, complicating extraction while imparting unique properties for diverse applications (Cord-Landwehr and Moerschbacher, 2021; Huq et al., 2022). Fungal sources of chitin and chitosan have several advantages over other sources, such as unique molecular weight homogeneity, charge and viscosity distribution, and the absence of heavy metals (Bastiaens et al., 2019). Moreover, fungal chitin, sourced from food and biotechnology waste streams, provides a sustainable option with consistent quality, unlike crustaceans (Cord-Landwehr and Moerschbacher, 2021). Fungal chitin is often preferred in biomedical applications due to its high purity and consistent quality. It's also non-toxic and biodegradable, making it suitable for various industries. Therefore, fungal chitin is preferred for vegan and cosmetic products due to its non-animal origin, appealing to consumers seeking cruelty-free options. Moreover, its availability is stable year-round, making it a reliable resource for industrial use (Cord-Landwehr and Moerschbacher, 2021; Huq et al., 2022). However, extracting chitin from fungal cell walls poses challenges, primarily due to the complex structure resulting from covalent bonds with glucans, which can lead to lower purity levels (Cord-Landwehr and Moerschbacher, 2021) Moreover, compared to other sources like crustacean shells, chitin from fungal sources, such as Ganoderma lucidum, may have lower thermal stability but can still be a viable alternative (Philibert et al., 2017). However, fungi are a promising source of chitin, providing opportunities for sustainable production and various applications across multiple industries (Tables 1, 2). Some studies have also explored the concurrent production of chitin from both shrimp shells and fungal sources, where the fungi can help deproteinate and demineralize the shrimp shells, providing a nitrogen source for their growth (Abo Elsoud and El-Kady, 2019).

Figure 2. Approximate chitin content from various sources (the numbers represent the maximum amounts).
Chitin is primarily known from crustaceans but is also present in mollusks, which include squids, cuttlefish, snails, clams, and chitons (Weiss, 2012; Chan et al., 2018). In mollusks, chitin is mainly found in shells, pens, and structural components, though in lower quantities than crustaceans. Molluscan chitin has a high degree of polymerization and crystallinity, providing excellent mechanical strength and biocompatibility (Tables 1, 2). It often exists alongside proteins and minerals, contributing to structural integrity, particularly in cephalopods like squids, where chitin forms specific associations with various proteins (Weiss, 2012; Younes and Rinaudo, 2015; Chan et al., 2018).
The physicochemical characteristics of chitin found in insects and other organisms can vary slightly, which affects properties like mechanical strength and stiffness (Figure 3 and Table 3). Some traits that may exhibit differences include crystallinity, molecular weight, acetylation level, fiber structure, and cross-linking with other molecules (Wang et al., 2013; Zargar et al., 2015; Moussian, 2019; Khajavian et al., 2022). Chitin is a linear polymer of N-acetylglucosamine units linked by β-1,4 glycosidic bonds. It forms a robust and flexible matrix in insects and other organisms, offering structural support and protection. Chitin imparts mechanical strength and toughness to insect and crustacean exoskeletons, with properties influenced by crystallinity, molecular weight, and acetylation degree. Despite a similar basic structure, variations in chitin’s physicochemical properties contribute to diverse functions and adaptations in these organisms (Merzendorfer and Zimoch, 2003; Erko et al., 2013; Moussian, 2013; Liu et al., 2019; Machałowski et al., 2020; Mohan et al., 2020; Zhang et al., 2021). However, the physicochemical, morphological, and structural characteristics of chitin and chitosan are influenced by the characteristics of the species, such as life stage, metamorphosis type, and genus (Mohan et al., 2020; Mohan et al., 2022). The main characteristics of chitin are (Figure 2).

Figure 3. The primary physicochemical properties of chitin and the grouping are based on the main sources (Amiri et al., 2022).

Table 3. The physicochemical properties of chitin derived from various sources.
Chitin is recognized for existing in multiple allomorphic forms, each characterized by varying degrees of crystallinity resulting from the hydrogen bonding interactions between its polymer chains. The degree of crystallinity (CrI) in chitin can fluctuate considerably, influenced by its source and the methods employed in its preparation (Table 4). Typically, CrI values range from 0.57 to 0.93, depending on the specific calculation method utilized (Casadidio et al., 2019). The crystallinity of chitin is a measure of the degree to which the polymer exhibits a regular, ordered structure (Soon et al., 2018). Crystalline regions within chitin provide strength and rigidity to structures like the exoskeleton, while amorphous regions contribute flexibility (Ogawa et al., 2011; Wang et al., 2013). Comparing the crystallinity of chitin from different sources involves examining the structural order and organization of its polymer chains across various biological origins. Several factors, including the source of the chitin, extraction methods, and processing conditions, influence its crystallinity. Chitin fibers associate with one another in three possible crystalline arrangements: α-chitin, β-chitin, or γ-chitin. These arrangements further contribute to chitin’s structural and functional diversity, with each allomorph exhibiting unique properties and degrees of crystallinity (Figures 4–6) (Lv et al., 2023). Two-dimensional sheets of α-chitin crystals form laminae. These sheets consist of antiparallel chitin fibers associated with β-folded proteins. α-Chitin is a primary constituent of the arthropod cuticle, crustaceans’ shell, and the fibrous extracellular matrix in sponges (Wang et al., 2013; Kumari and Kishor, 2020 January 1). β-Chitin found in hard biomaterials like the squid beak, β-chitin forms a tri-dimensional composite material with proteins. γ-Chitin is less well-studied and contributes to the physical barrier found in insect cocoons (Moussian, 2019). Chitin is a structural polymer that produces strong fibers both inside and outside cells. The fibers bond weakly with one another, increasing overall structural strength (Moussian, 2019; Hou et al., 2021). Moreover, chitin can be cross-linked with other molecules to enhance its strength, stability, and resistance to degradation (Jiao et al., 2023). For example, in fungal cell walls (e.g., Candida albicans), chitin forms strong bonds with glucans. Among arthropods and other invertebrates, chitin links with proteins either covalently or non-covalently (Arroyo et al., 2016; Wegrzynowska-Drzymalska et al., 2020; Wang et al., 2021). However, based on crystallinity, different sources of chitin can be categorized as: High crystallinity: Chitins derived from crustaceans such as crabs and shrimp typically demonstrate higher crystallinity. This is due to the highly ordered arrangement of chitin chains in their shells, which provides the necessary rigidity and strength. The crystallinity in crustaceans is influenced by the tight association with calcium carbonate and proteins, forming a complex matrix that enhances structural order (Wang et al., 2013; Pădurețu et al., 2019; Poerio et al., 2020; Hou et al., 2021; Jędrzejczak et al., 2024). Moderate to high crystallinity: Insect chitin also exhibits a relatively high degree of crystallinity, although it may be slightly lower than that of crustaceans. This is because insect cuticles are composed of chitin fibers embedded in a matrix of proteins, which may not be as densely packed as in crustaceans. The crystallinity can vary among different insect species and developmental stages. For example, some insects have softer, less crystalline exoskeletons in their larval stages compared to the highly crystalline cuticles in adults (Zhang et al., 2000; Sáenz-Mendoza et al., 2018; Henriques et al., 2020; Kumar et al., 2020; Poerio et al., 2020; Soetemans et al., 2020; Triunfo et al., 2022). Low crystallinity: Chitin extracted from fungi typically exhibits lower crystallinity compared to that from crustaceans and insects. In fungi, chitin is combined with other polysaccharides such as glucans and proteins, forming a less ordered matrix. Significant amorphous regions and a more heterogeneous composition in fungal cell walls contribute to the lower overall crystallinity (Blumenthal and Roseman, 1957; Velásquez and Pirela, 2016; Poerio et al., 2020).
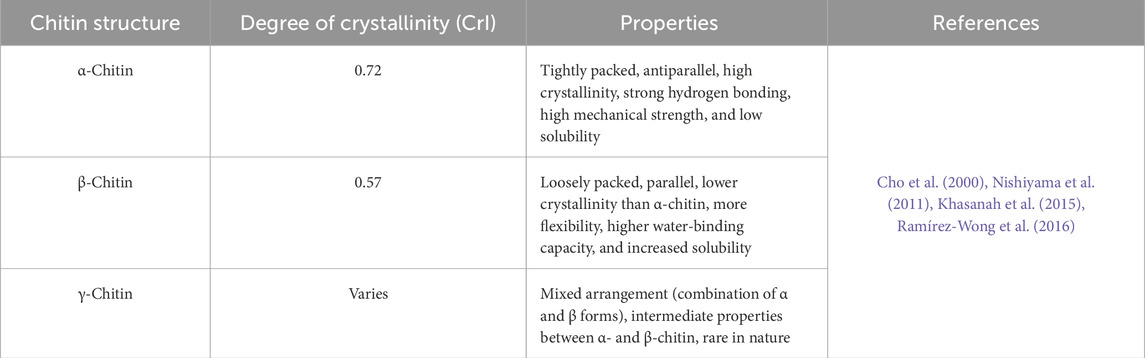
Table 4. The crystallinity properties of different chitin structures.
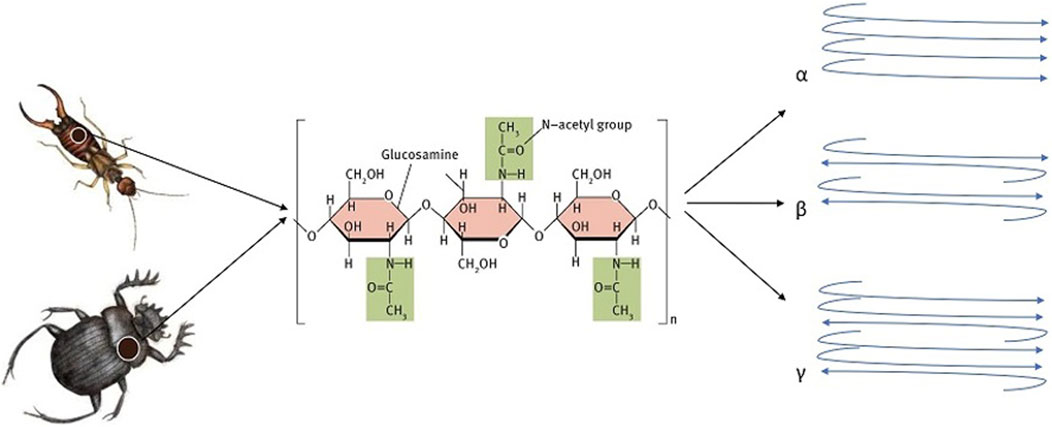
Figure 4. Schematic shapes of three forms of chitin (a, ẞ, and Y).

Figure 5. The X-ray diffraction (XRD) spectrum for (A) Chitin 1, (B) Chitin 2, and (C) Chitin 3 was obtained from the darkling beetle, Zophobas morio larvae, over a scanning angle range of 5°–45° at 30 kV and 30 mA. This data is reprinted with permission (5950520533931) from (Soon et al., 2018).

Figure 6. FTIR spectrograms illustrate the differences between (A) chitin and (B) chitosan extracted from the darkling beetle, Zophobas morio larvae, highlighting the effects of varying sodium hydroxide concentrations. This data is reprinted with permission (5950520533931) from (Soon et al., 2018).
Chitin’s crystalline arrangements and its association with proteins define the mechanical properties of tissues and organisms, making it a versatile extracellular polysaccharide. The variation in crystallinity reflects differences in the biological roles and structural requirements of chitin in these organisms. Crustaceans need a highly ordered structure for their protective shells, insects require moderately crystalline exoskeletons for flexibility and protection, while fungi have less crystalline chitin due to their distinct structural and functional requirements (Kabalak et al., 2020; Iber et al., 2022).
Comparing the molecular weight (MW) of chitin from different sources involves analyzing the polymer length of chitin chains. The molecular weight of chitin varies depending on the source, which affects its properties and applications. Chitin is a biopolymer made up of over 5,000 sugar units, with varying molecular weights (MW) depending on the number of monomeric units in the biopolymer, the source, and the deacetylation treatment (Yeul and Rayalu, 2013; Hahn et al., 2020). Here’s how these sources compare based on molecular weight: High molecular weight: Chitin from crustaceans generally has a high molecular weight. This is because crustaceans produce long, continuous chitin chains, which contribute to the mechanical strength and rigidity of their exoskeletons (Reddy and Yang, 2015; El Knidri et al., 2018; Zhang et al., 2018; Kumari and Kishor, 2020; Hou et al., 2021). Moderate to high molecular weight: Insect chitin usually has a slightly lower molecular weight compared to crustacean chitin. However, it still tends to be relatively high due to the need for durable and flexible exoskeletons. The molecular weight of insect chitin can vary widely among different insect species and life stages (Sáenz-Mendoza et al., 2018; Kumar et al., 2020; Kumari and Kishor, 2020; Hou et al., 2021; Pedrazzani et al., 2024). Low molecular weight: Chitin from fungal sources often has a lower molecular weight than that from crustaceans and insects. Fungal chitin is synthesized in shorter chains, which are adequate for the structural roles they play within the cell walls. The molecular weight of fungal chitin generally ranges from about 50,000 to 300,000 Da (Hou et al., 2021).
Chitin from crustaceans generally has the highest molecular weight, followed by chitin from insects, with fungal chitin having the lowest molecular weight. This variation reflects the different biological requirements and structural roles of chitin in these organisms. Crustaceans require long, high molecular weight chitin chains for strong and rigid exoskeletons, insects need moderately high molecular weight chitin for flexible yet protective exoskeletons, and fungi have shorter chitin chains suitable for their cell wall structures.
Comparing the purity of chitin from different sources involves assessing the level of contaminants or impurities present in the chitin extract. Purity can significantly impact the quality and suitability of chitin for various applications. Here’s how these sources compare based on purity: High purity: Chitin extracted from crustacean shells can achieve high levels of purity with proper processing techniques. However, achieving absolute purity can be challenging due to the presence of other biomolecules, such as proteins, lipids, and minerals, which are often tightly associated with chitin in the exoskeleton. Common contaminants in crustacean chitin include proteins like chitin-binding proteins and residual pigments from the shells (Zhang et al., 2018; Panchal and Desai, 2022). Moderate purity: Chitin from insect exoskeletons can also be purified to relatively high levels, but it may contain more impurities compared to crustacean chitin. Similar to crustaceans, insect chitin is associated with proteins and other biomolecules. Insect chitin may contain proteins, pigments, and other components present in the cuticle matrix (Panchal and Desai, 2022). Low purity: Chitin extracted from fungal cell walls generally has lower purity compared to crustacean and insect chitin. Fungal cell walls contain a complex mixture of polysaccharides, proteins, and other cell wall components, which can be challenging to separate from chitin. Common contaminants in fungal chitin include glucans, proteins, lipids, and other cell wall constituents (Panchal and Desai, 2022).
Achieving high-purity chitin requires careful extraction and purification processes to minimize contaminants and impurities. Crustacean chitin can be purified to relatively high levels due to the availability of well-established extraction methods, while insect chitin may contain slightly more impurities. Fungal chitin, on the other hand, is often associated with a more complex mixture of cell wall components, making it more challenging to obtain pure chitin extracts (Panchal and Desai, 2022).
The ratio of N-acetyl-D-glucosamine to β-linked D-glucosamine units determines the molecular weight and deacetylation degree (DD0 of chitosan. Higher DD generally leads to increased water solubility and improved biological activities (Amor et al., 2023). Chitin molecules can be partially or fully acetylated, impacting the polymer’s properties. Chitin is usually more acetylated in insects than in other arthropods, affecting solubility and enzymatic degradation. While insoluble in most solvents, chitin can dissolve in strong acids or bases. Its solubility in insects and arthropods depends on acetylation degree and exoskeleton components. Chitin is composed of repeating N-acetylglucosamine (GlcNAc) units. The degree of acetylation (DA) refers to the proportion of acetyl groups attached to the GlcNAc units. In chitin, the DA is typically around 0.90, indicating the presence of certain amine groups (though some deacetylation can occur during extraction, resulting in about 5%–15% amine groups in chitin). Acetylation level is a crucial factor that determines the properties of chitin. The degree of acetylation (DA) measures the proportion of acetyl groups in chitin and affects its physicochemical properties, such as solubility, chain conformation, and reactivity (Mouyna et al., 2020). Chitins may have different levels of acetylation depending on their sources such as fungi, insects, crustaceans, or mollusks, which may affect their potential uses (Brigode et al., 2020). Low deacetylation degree: Chitin from crustacean shells typically has a low DD, ranging from 10%–15%, when deacetylated using conventional chemical methods like treatment with concentrated alkali solutions. The highly crystalline structure of crustacean chitin makes it resistant to deacetylation (Zhao et al., 2010; Poerio et al., 2020; Novikov et al., 2023). Moderate deacetylation degree: Chitin from insect exoskeletons, like those of silkworms or bees, can reach a DD of around 50%–70% through chemical methods. The degree of deacetylation depends on the specific insect source and the deacetylation conditions. For example, chitosan extracted from Blaps lethifera, Pimelia fernandezlopezi, and Musca domestica showed different DD values (Zhao et al., 2010; Poerio et al., 2020; Amor et al., 2023). High deacetylation degree: Chitin derived from fungal cell walls, such as those from M. rouxii or A. niger, can achieve a higher DD of around 70%–90% through chemical deacetylation. Fungal chitin has a less crystalline structure compared to crustacean chitin, making it more susceptible to deacetylation (Zhao et al., 2010).
As the degree of deacetylation increases, chitin becomes more soluble and transforms into chitosan, which exhibits greater solubility in aqueous solutions, particularly under acidic conditions. The solubility of chitosan can be tailored by controlling the degree of deacetylation during its production. Fungal chitin generally achieves the highest DD through chemical deacetylation, followed by insect cuticles and crustacean shells. Enzymatic deacetylation offers the potential for higher and more controlled DD, but its efficiency varies based on the chitin source and process parameters (Zhao et al., 2010; Poerio et al., 2020; Novikov et al., 2023).
Besides chitin and proteins, the arthropod cuticles and crustacean shells also contain various minerals that make them strong and rigid. These minerals include calcium carbonate, calcium phosphate, and magnesium carbonate. (Erko et al., 2013; Moussian, 2013). The minerals found in arthropod cuticles and crustacean shells are crucial for their mechanical properties and structure. They add strength and rigidity to the cuticle, which acts as a protective shield for the animal’s body against any damage. Some animals have heavily mineralized cuticles, which makes them appear as if they have a hard, shell-like exterior. This is commonly observed in crustaceans like crabs and lobsters (Plotnick, 1990; Erko et al., 2013). The main sources of chitin differ in their demineralization requirements during the extraction process. The demineralization step is crucial for crustacean shells due to their high mineral content, while it is less critical for insect cuticles and not required for fungal chitin. The use of dilute HCl is the most common method for demineralization across all sources, but the specific conditions, such as acid concentration, temperature, and duration, may vary depending on the source (Younes and Rinaudo, 2015; Ibram et al., 2019; Abidin et al., 2020; Pellis et al., 2022).
Here’s a comparison of the common chitin sources based on the demineralization step (Zhao et al., 2010; Ibram et al., 2019; Vicente et al., 2021; Pellis et al., 2022; Novikov et al., 2023): Crustacean shells: Chitin from crustacean shells requires a demineralization step to remove the high mineral content, primarily calcium carbonate. Insect cuticles: Insects have fewer minerals than the shells of crustaceans (Khayrova et al., 2021). However, chitin from insect exoskeletons also requires a demineralization step, but the mineral content is generally lower than in crustacean shells. As a result, chitin and chitosan extracted from insects may have different physicochemical attributes than those extracted from crabs and shrimp (Saenz-Mendoza et al., 2023). Fungal chitin: Chitin derived from fungal cell walls does not require a demineralization step, as fungi generally have a lower mineral content than crustacean shells. Unlike other invertebrates (such as sponges, mollusks, or crustaceans), the cuticle of spiders lacks minerals such as CaCO3, which allows for the elimination of the demineralization step during chitin separation (Machałowski et al., 2020). The origin of chitin affects not only its crystallinity and purity but also its polymer chain arrangement and thus its properties (Rinaudo, 2006; Kumari and Kishor, 2020). The dosage of the main mineral elements present in the exoskeleton of different chitin sources shows that the content varies with the species and thus the processing conditions (Tolaimate et al., 2003). Two separate studies have shown differences in the structure of chitin obtained from different species of insects. In the first study (Waśko et al., 2016), chitin isolated from adults of Hermetia illucens was found to have a relatively smooth surface and was composed of parallel distributed fibers. In comparison, the larval chitin had a more complex structure, with a convex shape composed of repeated honeycomb-like units. In the second study (Erdogan and Kaya, 2016a), chitin obtained from adults and nymphs of Dociostaurus maroccanus was found to be composed of long nanofibers with large nanopores. High similarity was found in the properties of chitin and chitosan from nymphs and adults of the species. In experiments conducted on the potato beetles, Leptinotarsa decemlineata, it was found that chitin extracted from the adults and larvae of the species had a similar structure to nanofibers but differed in the number of pores. A study investigated chitin structure in four grasshopper species using various analytical techniques. Differences in chitin quantity and surface structure were observed between male and female grasshoppers. Despite gender variations, elemental analysis, thermal properties, and crystalline index values for chitin were similar. Enzymatic digestion showed no significant differences in digestion rates between chitins from both sexes and commercial chitin (Kaya et al., 2015e).
The deproteinization process for chitin extraction involves the removal of protein content from the raw material. The deproteinization method should be used to extract chitin from different sources. Deproteinization of chitin involves the use of alkaline solutions, primarily NaOH, to eliminate the protein content present in the raw material (Zainol Abidin et al., 2020b).
There are three different crystalline allomorphs of chitin found in nature which include α-chitin, β-chitin, and γ-chitin (Erdogan and Kaya, 2016a; Kaya et al., 2017a; Hou et al., 2021). α-chitin is the predominant chitin in insects and other arthropods (Figure 4). It has a highly ordered crystalline structure, with chitin molecules arranged in parallel chains. α-chitin is recognized for its high tensile strength and rigidity (Hou et al., 2021). It is the most consistent polymorphic form of chitin in nature and is present in crustaceans such as shrimps, lobsters, and crabs, as well as in the chitinous cuticles of insects. It can also be found in fungal cell walls (Ifuku et al., 2009; Hassainia et al., 2018). In a study, the structure of α-chitin has been determined through X-ray diffraction, revealing an orthorhombic unit cell with specific dimensions and a space group of P212121. The chains form hydrogen-bonded sheets and have a statistical mixture of side-chain orientations. The structure contains two types of amide groups with different hydrogen bonding, explaining the lack of swelling in water due to extensive intermolecular hydrogen bonding (Minke and Blackwell, 1978).
β-chitin is a less common form of chitin found in certain species, such as some crustaceans and mollusks. It has a different crystal structure compared to α-chitin, with the chains of chitin molecules arranged in an antiparallel fashion. This form of chitin is less rigid than α-chitin. It is characterized by loosely packed parallel chains with weak intermolecular interactions, making it more soluble and droplet-like than the α form. Additionally, the β form of chitin has been derived from squids (Pillai et al., 2009; Hamed et al., 2016; Hossin et al., 2021; Hou et al., 2021). γ-chitin is a type of chitin that is found in certain types of algae and fungi. It has a unique crystal structure that distinguishes it from the more well-known α-chitin and β-chitin. This is a less common form of chitin that has a disordered structure with random orientations of N-acetylglucosamine units. However, further research is required to fully comprehend its characteristics and prevalence in insects and other arthropods (Hou et al., 2021). A schematic shape of three forms is depicted in Figure 4. There is a mix of α and β forms in γ-chitin. The presence of γ-chitin is lower than that of α and β forms, and it has been examined in only a few studies. It is noteworthy that α chitin can convert to the β form, but the reverse is not true (Velásquez and Pirela, 2016; Hou et al., 2021).
Both α and β chitins keep a powerful network dominated by intrachain hydrogen bonds between the groups of C═O⋯NH and C═O⋯OH within an interval of 0.47 nm. In α form conformation, supernumerary inter-chain hydrogen bonds connect the hydroxymethyl groups while this type of interaction is not observed in the β conformation (Roy et al., 2017). These three types differ mainly in the degree of hydration, the size of the unit cell, and the number of chitin chains in each cell (Merzendorfer and Zimoch, 2003; Yu et al., 2024). On the other hand, the properties of pure chitin depend on their molecular weight, degree of acetylation, purity, and dispersion index. For example, in a study, the chitin of crab shells and mealworms was determined by X-ray diffraction and FTIR analysis. In this research, α form of chitin was observed in four crystalline reflections, shown at 9.4°, 19.3°, 20.8°, and 23.3° in the chitin of crab shell, and at 9.44°, 19.3°, 20.7°, and 23.3° in the chitin of mealworm by crystalline structure. α-form showed doublet at amide I band in crab shell and mealworm chitin (Kaur and Dhillon, 2015). In another study, chitin was extracted from various marine crustacean shells using chemical methods, showing a chitin content of 17.50%–23.75% on a dry weight basis. The chitin had low molecular weight and was similar to commercial chitin based on various analyses. The extracted chitin exhibited α-form characteristics, with a crystalline index value ranging from 80.3% to 80.8%. Analysis showed nanofiber and nanopore structures (Mohan et al., 2021). Chitin polymorphs α, β, and γ were isolated and analyzed for their crystalline structures and characteristics using various spectroscopic and analytical techniques. Molecular weights, spectral features, X-ray diffraction patterns, and thermal properties were determined for each chitin type. Differences in crystalline reflections, thermal decomposition temperatures, and activation energies were observed, highlighting the distinct properties of α-chitin, β-chitin, and γ-chitin. The study also demonstrated the temperature sensitivity differences among the chitin polymorphs based on their activation energies (Jang et al., 2004). Chitin can have different crystalline organizations and interact with chitin-binding proteins to create higher-order structures. This combination defines the mechanical properties of tissues and organisms (Moussian, 2019). Analysis through techniques like ATR-FTIR, NMR spectroscopy, and WAXS confirmed Bombyx mori as a promising source of α-chitin (Jędrzejczak et al., 2024). Overall, chitin, an abundant biopolymer, exhibits diverse structural conformations through natural processes like biopolymerization and self-assembly, leading to unique physical effects and mechanical properties (Hou et al., 2021).
Chitin is an amino polysaccharide found in yeast cells, arthropod cuticles, fungal cell walls, and crustacean shells (Rehman et al., 2023; Sanjanwala et al., 2024). When comparing these sources of chitin, several factors come into play: Purity, as the purity of chitin extracted from different sources can vary. Some sources may require more extensive purification processes to remove impurities such as proteins and minerals; Yield: The quantity of chitin extracted from a specific source varies depending on the species, size, and processing methods employed. Some sources may contain higher amounts of chitin compared to others. The quality of chitin can also vary depending on factors such as the degree of deacetylation (which affects its solubility and other properties) and the presence of contaminants; Cost and availability: The cost and availability of chitin from various sources can vary depending on factors such as geographical location, seasonal variability, and demand.
The choice of chitin source depends on factors such as intended applications, availability, cost considerations, and sustainability concerns. Researchers and industries are exploring various sources and methods for chitin extraction to meet the increasing demand for this versatile biopolymer. If we are ranking sources based solely on the amount of chitin that can be obtained, it generally follows this order (Merzendorfer and Zimoch, 2003; Ma et al., 2020; Khayrova et al., 2021; Iber et al., 2022; Lv et al., 2023; Thakur et al., 2023): Crustacean shells: Crustaceans have relatively large exoskeletons compared to insects and fungi. Therefore, they typically yield the highest amount of chitin per individual organism; Insect exoskeletons: Although insects have smaller bodies than crustaceans, they often exist in larger populations and have a higher overall biomass. Therefore, insect exoskeletons can still yield a significant amount of chitin, especially if sourced from species that are abundant or cultivated in large numbers; Fungal cell walls: Fungi generally have smaller biomass than insects and crustaceans. Additionally, not all fungi produce chitin in significant quantities. Therefore, while fungal cell walls can be a source of chitin, the overall yield may be lower compared to insect and crustacean sources. So, based solely on the amount of chitin that can be obtained from each source, the ranking would be crustacean shells˃ insect exoskeletons˃ fungal cell walls.
One way to compare the chitin content of various organisms is through chemical analysis of the chitin found in their exoskeletons or cell walls. This involves isolating the chitin from the tissues and quantifying it using techniques like Fourier Transform Infrared Spectroscopy (FTIR) or High-Performance Liquid Chromatography (HPLC) (Qin et al., 2019; Ghosh and Dhepe, 2021; Tsurkan et al., 2021). Another approach is to assess the thickness or density of chitinous structures in different organisms, as these traits can indicate the amount of chitin present. For instance, insects with thicker exoskeletons may have higher chitin content than those with thinner exoskeletons. Furthermore, genetic studies can help compare chitin content across organisms by examining the genes related to chitin synthesis and degradation. This allows researchers to understand how chitin production and turnover are regulated in various species. Combining chemical, morphological, and genetic analyses can comprehensively compare chitin content in different organisms, providing insights into the role of chitin in their biology (Kabalak et al., 2020; Khayrova et al., 2021; Lv et al., 2023). The chitin content in the shells of commercial organisms like shrimps, crabs, and crayfish is approximately 20%. In contrast, certain insect species have a chitin content of around 15%. Additionally, the chitin content in some aquatic insect species ranges from 10% to 20% (Table 5) (Susana Cortizo et al., 2008; Sagheer et al., 2009; Wang et al., 2013).
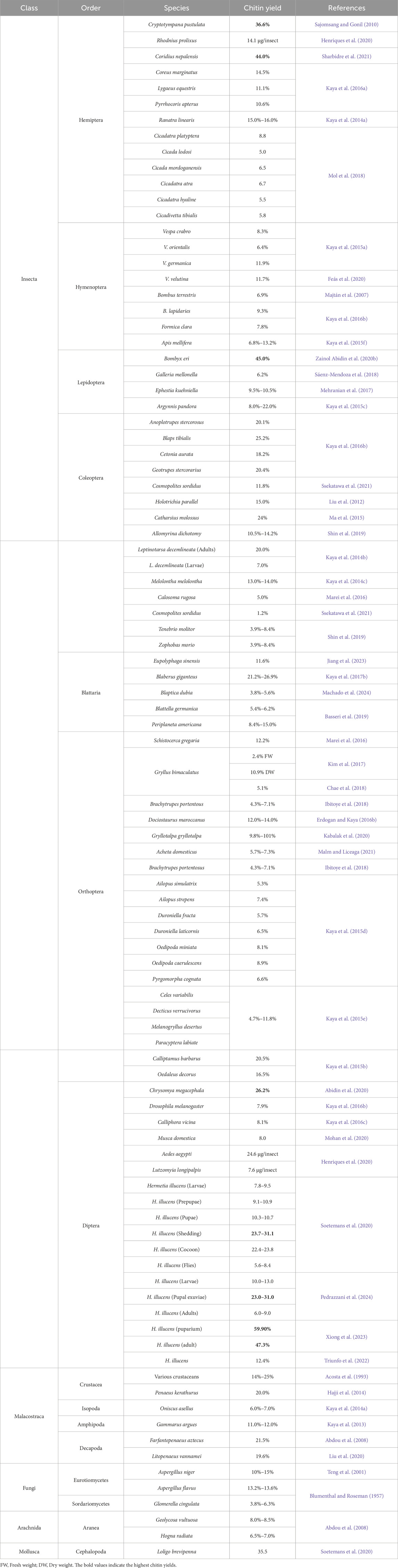
Table 5. The amount of chitin in different species of insects and other organisms.
Chitin content, purity, and structural properties differ among species, developmental stages, sexes, and body parts. Additionally, abiotic factors like processing methods and environmental and cultivation conditions influence the quality and quantity of chitin (Weiss, 2012; Chan et al., 2018; Hahn et al., 2020). Table 5 presents chitin yields across various insect species and other organisms. The data, organized by species, order, class, and yield, facilitates comparisons of chitin levels. This information is valuable for applications that utilize chitin due to its versatility in pharmaceuticals, biotechnology, agriculture, and materials science. Certain insects, such as B. eri (45%) and H. illucens puparium (59.9%), exhibit notably high chitin yields. These species may be prioritized for chitin extraction due to their superior yields, making them more efficient sources.
Animal and non-animal sources of chitin offer distinct content ranges, purity levels, and extraction requirements, impacting their applications and sustainability. Animal-derived chitin, present at higher concentrations (8%–64%) in shell structures, undergoes rigorous demineralization and deproteinization processes to remove non-chitin substances (Table 1). This high-purity chitin is widely applied in biomedical fields, such as drug delivery systems and biodegradable materials, where quality and functionality are paramount. In contrast, non-animal sources, such as fungi, yield lower chitin content (8%–16%) and produce chitosan, a soluble chitin derivative with unique chemical properties. Fungal chitin extraction is less intensive, focusing primarily on residue removal, making it an eco-friendly alternative to animal chitin. The selection of a chitin source depends on specific application needs, sustainability considerations, and resource availability. While animal-derived chitin offers higher yields, concerns about environmental impact, such as overfishing, may limit its use. Fungal chitin, though yielding less chitin per source, is a more sustainable option, frequently employed in food and pharmaceutical industries due to its distinct biochemical profile. Thus, each chitin source has advantages and limitations, influencing its suitability across diverse industries. This revision provides improved clarity, consistency, and balanced comparison of the two chitin sources, emphasizing their unique benefits and environmental considerations.
The dataset spans species across multiple taxonomic classes, including insects, fungi, crustaceans, and mollusks, allowing for a comprehensive cross-taxonomic comparison of chitin yields. Such diversity provides insights into environmentally and economically viable choices, as some species may be more sustainable or cost-effective to cultivate and harvest. On a broad taxonomic scale, insects (Class Insecta) exhibit notable variability in chitin yield percentages, distinguishing them from other groups like fungi and crustaceans. Crustacean species such as Farfantepenaeus aztecus (21.5% chitin content) and specific life stages of the black soldier fly (H. illucens) show high chitin levels, making them strong candidates for high-yield chitin extraction. Among insects, the orders Lepidoptera and Diptera are particularly promising, with species like B. eri (Lepidoptera), C. megacephala, and H. illucens (Diptera) achieving chitin yields as high as 59.9% in certain developmental stages, such as the puparium of H. illucens. This trend suggests that insects within these orders may serve as highly efficient sources for chitin harvesting. In addition, substantial variability exists within certain insect orders, such as Orthoptera, where chitin yields range from 1.2% in Cosmopolites sordidus to approximately 26.2% in Calliptamus barbarus, highlighting the extensive yield variation within a single order. Furthermore, the physicochemical properties of chitin differ significantly across taxa and are partially specific to each order. These differences manifest in surface morphology, which can vary even among species within the same order, suggesting that taxonomic and developmental factors influence chitin properties. This comprehensive taxonomic analysis underscores the potential for selecting species not only based on yield but also considering the physicochemical attributes of chitin, which may impact its functionality across different applications (Kaya et al., 2016a). However, findings indicate a correlation between chitin content, crystalline index, and taxonomical relationships. In this line, chitin was isolated from 16 arthropod species (13 insects and 3 arachnids) in a study. Among Insecta, chitin content was highest in Coleoptera (18.2%–25.2%), followed by Hemiptera, Odonata, Hymenoptera, Diptera, and Blattodea. Additionally, Coleoptera demonstrated higher crystalline index values compared to other orders (Kaya et al., 2016b). Among crustaceans, the composition of shrimp shells varies according to species, seasonal changes, and geographical locations. The components of shrimp shell waste consist of 10%–25% chitin, 13%–50% protein, 15%–70% minerals (Babu et al., 2008), and low-fat value (Cira et al., 2002). A study characterized the functional properties and microstructure of chitin extracted from the house cricket, Acheta domesticus, field cricket, Gryllus bimaculatus, and black soldier fly, H. illucens cocoons compared to commercial shrimp chitin. Various properties like solubility, emulsion capacity, and purity were determined. G. bimaculatus chitin showed high fat absorption and emulsion capacity, while H. illucens chitin had the highest deacetylation level. The highest values for purity were found in commercial chitin and G. bimaculatus, followed by H. illucens. Overall, insect chitin exhibited similar characteristics to commercial chitin, suggesting its suitability for industrial applications (Ndiritu et al., 2023).
Insects vs. Crustaceans: The chitin yield in crustaceans, such as F. aztecus (21.5%) and Litopenaeus vannamei (19.6%), is relatively high and comparable to that of many high-yield insect species. Chitin structures from two European species, the common cockchafer, Melolontha melolontha, and Crustacea, Oniscus asellus, were isolated and compared. Both underwent similar isolation procedures using HCl and NaOH. Physicochemical properties of the chitins were analyzed, showing higher chitin content, purer composition, and more pores in M. melolontha compared to O. asellus (Kaya et al., 2014a). Various analyses confirmed similarities between the chitins, but differences in nanofiber adherence and pore structures were observed. M. melolontha was deemed a more appealing chitin source due to its purity (Kaya et al., 2014b). A study demonstrated that Holotrichia parallel (Coleoptera: Scarabaeidae) chitin was compared with commercial chitin from shrimp using various analytical techniques such as infrared spectroscopy, X-ray diffraction, scanning electron microscopy, and elemental analysis. Chitin from both sources exhibited comparable chemical structures and physicochemical properties (Liu et al., 2012). These indicate that crustaceans could serve as a viable alternative for chitin extraction, particularly in regions where they are abundant.
Mollusks and Fungi: The cephalopod Loligo brevipenna exhibits a fresh weight yield of 35.5%, which is significant; however, it is not directly comparable to the dry weight percentages reported for other organisms. In fungi, A. niger contains a chitin content of approximately 10%–15%, suggesting that fungi could serve as an alternative, although they may require more specialized processing methods.
Chitin extracted from various body parts of insects shows differences in content, surface morphologies, and degradation temperatures. The study investigated the correlation between chitin properties and honeybee body parts—head, thorax, abdomen, legs, and wings. The highest chitin content was found in the legs, while the lowest was in the thorax. Different surface morphologies were observed across body parts, with the abdomen showing the most variation. Maximum degradation temperatures varied among body parts, with the head chitin having the lowest value (Kaya et al., 2015f). A study investigates the physicochemical properties of chitin extracted from the butterfly species’ wings and other body parts, Argynnis pandora. The findings revealed that the chitin content in the wings (22.0%) is significantly higher than that in other body parts (8.0%). Additionally, the wings exhibited a single surface morphology, while four distinct morphologies were observed in the other body parts. (Kaya et al., 2015c). However, the findings suggest a strong connection between the surface morphology of chitin and the specific body part from which it is isolated.
In certain species, such as the black soldier fly, H. illucens, chitin content varies significantly across different life stages (Larvae: 10.0%–13.0%, Prepupae: 9.1%–10.9%, Pupae: 10.3%–10.7%, Puparium: 59.9%, Adult: 47.3%) (Triunfo et al., 2022) (Henriques et al., 2020). The cocoon and puparium stages exhibit higher chitin percentages compared to adults. This variation suggests the potential for optimizing extraction processes by targeting specific life stages. Harvesting chitin at specific developmental stages, particularly from the puparium, could yield higher extraction rates, enhancing efficiency for large-scale chitin production. Chitin was extracted from different life stages of the mealworm beetle, Tenebrio molitor, and Oryctes rhinoceros with varying yields (Shin et al., 2019). Chitin and chitosan extracted from Colorado potato beetle, L. decemlineata were analyzed, revealing significant differences between larvae and adults. Adult beetles had a chitin content of 20% based on dry weight, while larvae had only 7.0%. Additionally, adults produced higher yields of chitosan, with 72.0% compared to 67.0% in larvae (Kaya et al., 2014c). Chitin was extracted from different life stages of the mealworm beetles (T. molitor, Zophobas morio) and the rhinoceros beetle (Allomyrina dichotoma) through processes of deproteinization and demineralization. The yields were as follows: 4.6%, 8.4%, and 3.9% from the larvae, adult, and superworm of the mealworm beetle, respectively; and 10.5%, 12.7%, and 14.2% from the larvae, pupa, and adult stages of the rhinoceros beetle (Shin et al., 2019). The chitin content of adult (14.0%) and nymph (12.0%) D. maroccanus, measured using the same method, was similar. The crystalline index values of chitin from adult and nymph grasshoppers were 71% and 74%, respectively (Erdogan and Kaya, 2016b). A study investigated the changes in the physicochemical properties of chitin in the insect Vespa crabro throughout its developmental stages: larva, pupa, and adult. The chitin content increased progressively with growth, reaching levels of 2.1%, 6.2%, and 10.3% at each stage, with a significant threefold increase occurring between the larva and pupa stages. Notably, the surface characteristics of chitin changed considerably as the insect matured (Kaya et al., 2016c)
In four grasshopper species, Celes variabilis, Decticus verrucivorus, Melanogryllus desertus, and Paracyptera labiate, males had higher chitin content than females, with the dry weight of chitin varying between species from 4.7% to 11.8% (Kaya et al., 2015e). A recent study examined how gender affects the physicochemical properties of chitin isolates sourced from various body parts of the coleopteran species, Melolontha sp. The results revealed significant differences in chitin content, crystallinity, thermal stability, and surface morphology between samples extracted from male and female specimens across eight body parts, including antennae, head, and legs. Furthermore, interactions between the chitin samples and bovine serum albumin indicated potential differences in their applications based on the gender of the specimens (Kaya et al., 2016d). Testing a new method utilizing calcofluor fluorescent staining followed by microplate readings on disease vector species like Lutzomyia longipalpis, Aedes aegypti, and Rhodnius prolixus revealed significant variations in chitin levels, influenced by species, sex, and instar. For instance, in L. longipalpis females, the chitin content is approximately 12.0 μg per insect, while individual males have about 8.0 μg of chitin (Henriques et al., 2020).
It should be noted that the chitin content of different sources may vary depending on the extraction and bleaching techniques employed. The chitin extraction process can significantly influence the final chitin content from a particular source. Extraction of chitin can utilize a range of methods, such as biological, chemical, and physical techniques. The primary methods employed in industry and medical fields are biological and chemical. However, these methods differ in their ability to target and eliminate unwanted components like proteins and minerals with varying degrees of effectiveness. Harsh chemicals may degrade some chitin along with impurities, resulting in reduced yield. Conversely, milder techniques could leave behind more proteins or minerals, thereby diluting the chitin content. Additionally, certain methods, particularly those involving high temperatures or strong chemicals, have the potential to degrade the chitin itself, leading to a diminished yield of usable chitin. Using different extraction methods revealed variations in chitin levels across developmental stages of the black soldier fly. Additionally, bleaching led to a notable decrease in chitin content at various developmental stages of the black soldier flies (Soetemans et al., 2020; Hahn et al., 2022).
The mineral content is a crucial factor influencing the efficiency and yield of chitin extraction from different natural sources. The mineral content in chitin-containing materials varies significantly depending on the source. For example, crab shells typically contain 20%–30% chitin, 30%–40% protein, and 30%–50% minerals (mainly calcium carbonate). However, this composition can differ based on factors such as species, season, and even the body part of the organism. Calcium carbonate is a common mineral found in chitin sources, but other minerals like sodium, potassium, and magnesium might also be present in smaller amounts. Insect cuticles have lower levels of inorganic material than crustacean shells, making their demineralization treatment more convenient and economical with milder chemicals. Animals with lower mineral content, such as insects, tend to have higher chitin yields and require milder extraction conditions compared to highly mineralized crustacean sources. The mineral content in insects can vary, with some sources like cicadas having around 10% minerals (Berezina, 2016; Abidin et al., 2020; Poerio et al., 2020; Terkula Iber et al., 2022; Triunfo et al., 2022).
The amount of chitin in different species is presented in Table 5. Certain insects have less chitin than shrimp exoskeletons, but this is not universally true. For instance, B. eri, Coridius nepalensis, and P. cicadae have notably higher chitin levels at 45.0%, 44.0%, and 36.6% respectively (Figure 7). These amounts surpass the chitin content of shrimp, a significant source of commercial chitin and chitosan. It should be noted that in a fermentation process, the chitin yield of H. illucens puparium and adults was substantially higher (Xiong et al., 2023). Edible insects boast rich biodiversity and significant chitin content, positioning them as an underutilized asset (Ndiritu et al., 2023). Of the studied insects, the chitin content was highest in B. eri, followed by two hemipteran species (C. nepalensis, P. cicadae), and H. illucens. However, the black soldier fly, H. illucens, is an edible insect that can be easily raised under various conditions and is considered a potent source of chitin, even at the commercial level (Eggink and Dalsgaard, 2023; Pedrazzani et al., 2024). In this line, a study explored the potential of the black soldier fly (BSF) as a chitin source compared to shrimp shells. Different extraction methods were tested, and BSF chitin was found more challenging to extract and purify due to strong protein binding. The purity of BSF chitin ranged from 47.6% to 79.9%, lower than shrimp chitin at 88.3% (Pedrazzani et al., 2024). In another study, chitin was extracted from different developmental stages of the BSF, with varying chitin content. The physicochemical evaluation revealed that BSF chitin was α-chitin, similar to shrimp chitin (Soetemans et al., 2020). Research explores using black soldier fly puparium and adults with a continuous fermentation method employing specific bacteria for chitin extraction. Results show successful extraction of α-chitin with varying rates and yields based on the developmental stage of the flies. The study suggests that simultaneous continuous fermentation could offer a novel biological approach for chitin extraction from black soldier flies (Xiong et al., 2023). Comparing this research’s results with others indicates the significant effect of the extraction method on chitin yield. However, the increasing demand for sustainable alternatives to crustaceans as a source of chitin and chitosan highlights bioconverter insects like H. illucens as potential sources. A study (Triunfo et al., 2022) emphasized the suitability of pupal exuviae for high chitin yields. Analysis shows that insect-derived polymers are similar to commercial ones, making them suitable for industrial and biomedical applications. The chitin from H. illucens is noted for its fibrillary nature, conducive to producing fibrous materials, while chitosan exhibits properties that could support antimicrobial applications (Triunfo et al., 2022). Researchers have found that the larvae of B. eri are a promising source of chitin (Huet et al., 2020). A study compared the chitin extracted from B. eri larvae to shrimp chitin. While both exhibited similar characteristics in terms of crystallographic structures and thermal stability, they differed in crystallinity and morphological structures. Furthermore, insect chitins treated with room-temperature ionic liquid showed higher digestibility when subjected to enzymatic degradation, making them a more promising source for N-acetyl-D-glucosamine production (Huet et al., 2020). Another study focused on extracting chitin from the edible insect, C. nepalensis and compared it with commercial shrimp chitin. Results showed a chitin yield of 44.0% from C. nepalensis with a degree of acetylation of 57. 7%. The extracted chitin displays a mixture of chitin and chitosan, differing in crystalline structure and surface morphology from shrimp chitin. This research highlights the potential of C. nepalensis as a novel biomaterial source (Sharbidre et al., 2021). Research on the impact of various food wastes on H. illucens larvae revealed no significant differences in chitin and chitosan composition based on feeding substrates. The study underscores the importance of understanding how feeding substrates relate to valuable by-products, such as chitosan, in industrial insect production (Soetemans et al., 2020; Eggink and Dalsgaard, 2023; Xiong et al., 2023; Adamaki-Sotiraki et al., 2024; Pedrazzani et al., 2024). However, when choosing the best sources of chitin for extraction, several factors such as the amount of chitin, the purity, and the availability of its sources need to be considered. Although some insects can provide the highest amount of chitin, crustacean shells (such as those of crabs, shrimp, and krill) have been the traditional commercial sources due to their moderate but consistent yield of chitin. These shells are usually readily available from seafood processing industries. However, insects offer a potentially more sustainable option with a consistent supply, making them a promising source of high yield and purity of chitin. That being said, crustacean shells remain a good option due to their established processing techniques and easily available supply.
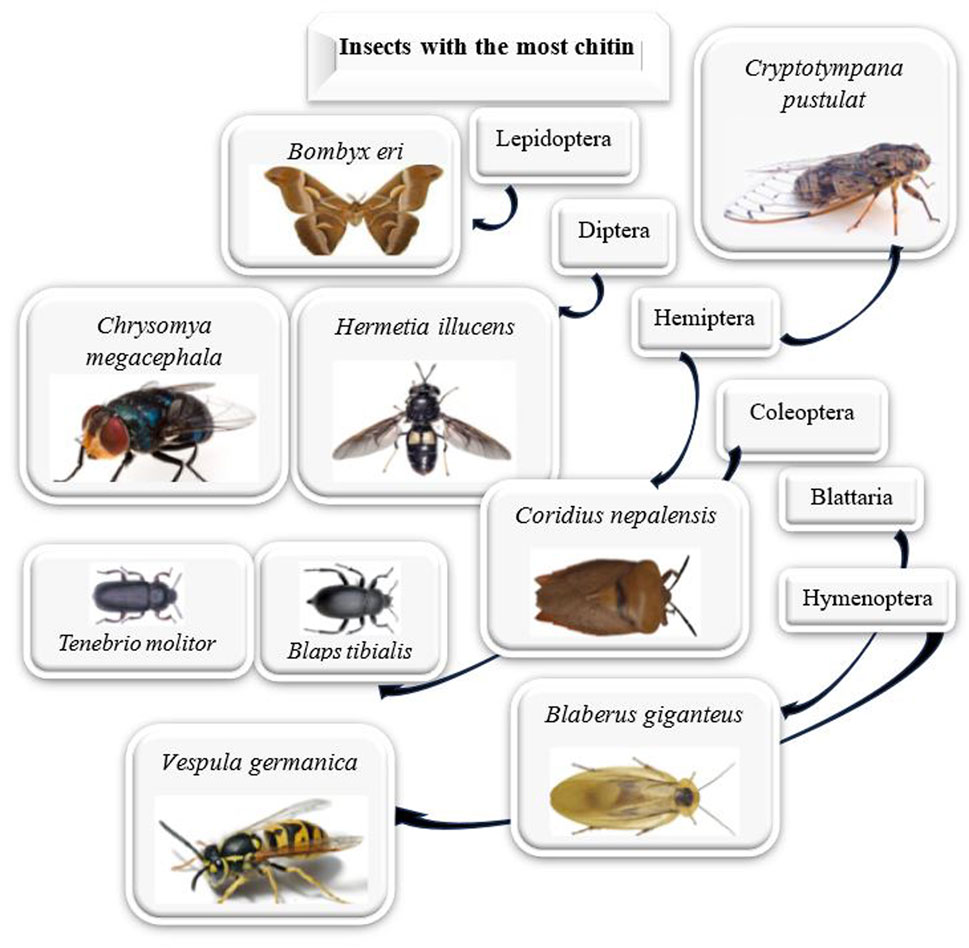
Figure 7. The highest chitin content among various insect orders.
Insect chitin degrades more easily than shrimp chitin when treated with 6 N HCl and the enzyme chitinase. N-deacetylation of insect chitin is also simpler than that of crustaceous chitin. In a study, the enzymatic reaction of beetle and shrimp chitins was conducted heterogeneously in an acetate buffer solution. The hydrolysis rates of beetle and shrimp chitins by chitinase vary, with beetle chitin being hydrolyzed faster than shrimp chitin, suggesting that cockroach chitin has a higher affinity for chitinase than shrimp chitin (Zhang et al., 2000). The catechol compounds in insect cuticles inhibit chitin crystallinity, making the amorphous structure of insect chitin more easily destructible. N-deacetylation of insect chitin is simpler compared to crustaceous chitin, with approximately 94.0% of N-acetyl groups removed in a single treatment using 40% NaOH for 4 h at 110°C. Silkworm chitin, when treated with 2 N HCl under similar conditions, had 55% of its N-acetyl groups removed. Beetle chitin exhibited greater chitinase affinity than shrimp chitin (Zhang et al., 2000). The deacetylation process results in a copolymer of N-acetyl-glucosamines and glucosamines. When the resulting copolymer contains more than 50% N-acetyl-glucosamine units, it is typically referred to as chitin. For chitin polymers, the percentage of N-acetyl-glucosamine units is termed the degree of acetylation (DA) and can vary from 50% to 100% (Sivashankari and Prabaharan, 2017). A higher degree of acetylation reduces the solubility of chitin. Shrimp shells usually contain chitin with a degree of acetylation exceeding 50% (Sivashankari and Prabaharan, 2017; Yanat et al., 2023). This high acetylation level contributes to its lower solubility. Insects, however, offer chitin with superior characteristics. Studies indicate that chitin sourced from insects has a more favorable balance of deacetylation and acetylation than aquatic crustaceans (Kumar et al., 2020). While crustacean shells, such as those from shrimp, have long been the traditional and reliable source of chitin due to established processing methods and consistent yields, insects—particularly species like H. illucens, B. eri, and C. nepalensis—are emerging as promising alternative sources. Insects offer several advantages, including higher chitin content in certain species, greater biodegradability, and potentially more efficient extraction methods, making them a sustainable and underutilized resource for chitin production. Furthermore, the distinct characteristics of insect chitin, such as its more favorable balance of deacetylation, suggest it may provide unique benefits in industrial and biomedical applications. However, factors such as purity, yield, and the availability of these insect sources must be considered when selecting the optimal chitin provider, as crustaceans still maintain an advantage in specific commercial contexts. Nevertheless, the growing demand for eco-friendly and sustainable alternatives positions insects as a viable future chitin and chitosan production source.
Chitin, the second most abundant polysaccharide on Earth, is a crucial resource with diverse applications in agriculture, medicine, and industry. Its sources, crustaceans, insects, fungi, and mollusks, each have distinct advantages and limitations. Crustaceans provide high chitin yield and superior crystallinity but present environmental and economic challenges due to their mineral content and seasonal availability. In contrast, insects serve as a sustainable alternative, offering high-purity chitin with lower environmental impact and minimal resource requirements, while also contributing to organic waste recycling. Fungal chitin, despite its lower purity and yield, holds promise for biomedical applications due to its consistent availability and cruelty-free sourcing. Mollusks provide unique mechanical and biocompatible properties, though in limited quantities. The physicochemical properties such as crystallinity, molecular weight, and degree of deacetylation vary among these sources, affecting their functionality for various applications. These insights emphasize the need for eco-friendly, cost-effective extraction methods tailored to each source to enhance chitin’s utility. Additionally, the potential of insect-based chitin aligns with circular economy principles, advocating for its wider adoption. Future research should aim to refine extraction processes, improve chitin yield and purity, and investigate novel applications to fully utilize this biopolymer in a resource-constrained environment.
HI: Conceptualization, Data curation, Investigation, Methodology, Resources, Software, Validation, Visualization, Writing–original draft, Writing–review and editing. HA: Investigation, Resources, Writing–original draft. MB: Writing–review and editing.
The author(s) declare that no financial support was received for the research, authorship, and/or publication of this article.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmats.2025.1537067/full#supplementary-material
Abdou, E. S., Nagy, K. S. A., and Elsabee, M. Z. (2008). Extraction and characterization of chitin and chitosan from local sources. Bioresour. Technol. 99, 1359–1367. doi:10.1016/j.biortech.2007.01.051
Abidin, N. A. Z., Kormin, F., Abidin, N. A. Z., Anuar, N. A. F. M., and Bakar, M. F. A. (2020). The potential of insects as alternative sources of chitin: an overview on the chemical method of extraction from various sources. Int. J. Mol. Sci. 21, 1–25. doi:10.3390/ijms21144978
Abo Elsoud, M., Elmansy, A., and Abdelhamid, S. A. (2022). Economic and non-seasonal source for production of chitin and chitosan. J. Chem. Rev. 4, 222–240. doi:10.22034/jcr.2022.342454.1173
Abo Elsoud, M. M., and El Kady, E. M. (2019). Current trends in fungal biosynthesis of chitin and chitosan. Bull. Natl. Res. Cent. 43, 59. doi:10.1186/s42269-019-0105-y
Acosta, N., Jiménez, C., Borau, V., and Heras, A. (1993). Extraction and characterization of chitin from crustaceans. Biomass Bioenergy 5, 145–153. doi:10.1016/0961-9534(93)90096-M
Adamaki-Sotiraki, C., Abenaim, L., Mannucci, A., Rumbos, C. I., Bedini, S., Castagna, A., et al. (2024). Performance of Hermetia illucens (Diptera: stratiomyidae) larvae reared on organic waste diets and pupal chitin and chitosan yield. Environ. Sci. Poll. Res. 31, 37366–37375. doi:10.1007/s11356-024-33545-x
Amiri, H., Aghbashlo, M., Sharma, M., Gaffey, J., Manning, L., Moosavi Basri, S. M., et al. (2022). Chitin and chitosan derived from crustacean waste valorization streams can support food systems and the UN sustainable development goals. Nat. Food 3, 822–828. doi:10.1038/s43016-022-00591-y
Amor, I. B., Hemmami, H., Laouini, S. E., Abdelaziz, A. G., and Barhoum, A. (2023). Influence of chitosan source and degree of deacetylation on antibacterial activity and adsorption of AZO dye from water. Biomass Convers. Biorefin 14, 16245–16255. doi:10.1007/s13399-023-03741-9
Anedo, E. O., Beesigamukama, D., Mochoge, B., Korir, N. K., Haukeland, S., Cheseto, X., et al. (2024). Evolving dynamics of insect frass fertilizer for sustainable nematode management and potato production. Front. Plant Sci. 15, 1343038. doi:10.3389/fpls.2024.1343038
Arroyo, J., Farkaš, V., Sanz, A. B., and Cabib, E. (2016). Strengthening the fungal cell wall through chitin-glucan cross-links: effects on morphogenesis and cell integrity. Cell. Microbiol. 18, 1239–1250. doi:10.1111/cmi.12615
Babu, C. M., Chakrabarti, R., and Surya Sambasivarao, K. R. (2008). Enzymatic isolation of carotenoid-protein complex from shrimp head waste and its use as a source of carotenoids. LWT - Food Sci. Technol. 41, 227–235. doi:10.1016/j.lwt.2007.03.006
Basawa, R., Kabra, S., Khile, D. A., Faruk Abbu, R. U., Parekkadan, S. J., Thomas, N. A., et al. (2023). Repurposing chitin-rich seafood waste for warm-water fish farming. Heliyon 9, e18197. doi:10.1016/j.heliyon.2023.e18197
Basseri, H., Bakhtiyari, R., Hashemi, S. J., Baniardelani, M., Shahraki, H., and Hosainpour, L. (2019). Antibacterial/antifungal activity of extracted chitosan from American cockroach (Dictyoptera: blattidae) and German cockroach (Blattodea: blattellidae). J. Med. Entomol. 56, 1208–1214. doi:10.1093/jme/tjz082
Bastiaens, L., Soetemans, L., D’Hondt, E., and Elst, K. (2019). “Sources of chitin and chitosan and their isolation,” in Chitin and chitosan Wiley, 1–34. doi:10.1002/9781119450467.ch1
Berezina, N. (2016). Production and application of chitin. Physic Sci. Rev. 1. doi:10.1515/psr-2016-0048
Blumenthal, H. J., and Roseman, S. (1957). Quantitative estimation of chitin in fungi. J. Bacteriol. 74, 222–224. doi:10.1128/jb.74.2.222-224.1957
Brigode, C., Hobbi, P., Jafari, H., Verwilghen, F., Baeten, E., and Shavandi, A. (2020). Isolation and physicochemical properties of chitin polymer from insect farm side stream as a new source of renewable biopolymer. J. Clean. Prod. 275, 122924. doi:10.1016/j.jclepro.2020.122924
Casadidio, C., Peregrina, D. V., Gigliobianco, M. R., Deng, S., Censi, R., and Di Martino, P. (2019). Chitin and chitosans: characteristics, eco-friendly processes, and applications in cosmetic science. Mar. Drugs 17, 369. doi:10.3390/md17060369
Chae, K. S., Shin, C. S., and Shin, W. S. (2018). Characteristics of cricket (Gryllus bimaculatus) chitosan and chitosan-based nanoparticles. Food Sci. Biotechnol. 27, 631–639. doi:10.1007/s10068-018-0314-4
Chan, V. B. S., Johnstone, M. B., Wheeler, A. P., and Mount, A. S. (2018). Chitin facilitated mineralization in the eastern oyster. Front. Mar. Sci. 5. doi:10.3389/fmars.2018.00347
Chen, Q., and Peng, D. (2019). Nematode chitin and application. Adv. Exp. Med. Biol. 1142, 209–219. doi:10.1007/978-981-13-7318-3_10
Cho, Y.-W., Jang, J., Park, C. R., and Ko, S.-W. (2000). Preparation and solubility in acid and water of partially deacetylated chitins. Biomacromolecules 1, 609–614. doi:10.1021/bm000036j
Cira, L. A., Huerta, S., Hall, G. M., and Shirai, K. (2002). Pilot scale lactic acid fermentation of shrimp wastes for chitin recovery. Process Biochem. 37, 1359–1366. doi:10.1016/S0032-9592(02)00008-0
Cord-Landwehr, S., and Moerschbacher, B. M. (2021). Deciphering the ChitoCode: fungal chitins and chitosans as functional biopolymers. Fungal Biol. Biotechnol. 8, 19. doi:10.1186/s40694-021-00127-2
Crini, G. (2005). Recent developments in polysaccharide-based materials used as adsorbents in wastewater treatment. Prog. Polym. Sci. 30, 38–70. doi:10.1016/j.progpolymsci.2004.11.002
Eggink, K. M., and Dalsgaard, J. (2023). Chitin contents in different black soldier fly (Hermetia illucens) life stages. J. Insects Food Feed 9, 855–864. doi:10.3920/JIFF2022.0142
Ehrlich, H., Krautter, M., Hanke, T., Simon, P., Knieb, C., Heinemann, S., et al. (2007). First evidence of the presence of chitin in skeletons of marine sponges. Part II. Glass sponges (Hexactinellida: Porifera). J. Exp. Zool. B Mol. Dev. Evol. 308B, 473–483. doi:10.1002/jez.b.21174
El Knidri, H., Belaabed, R., Addaou, A., Laajeb, A., and Lahsini, A. (2018). Extraction, chemical modification and characterization of chitin and chitosan. Int. J. Biol. Macromol. 120, 1181–1189. doi:10.1016/j.ijbiomac.2018.08.139
Erdogan, S., and Kaya, M. (2016a). High similarity in physicochemical properties of chitin and chitosan from nymphs and adults of a grasshopper. Int. J. Biol. Macromol. 89, 118–126. doi:10.1016/j.ijbiomac.2016.04.059
Erdogan, S., and Kaya, M. (2016b). High similarity in physicochemical properties of chitin and chitosan from nymphs and adults of a grasshopper. Int. J. Biol. Macromol. 89, 118–126. doi:10.1016/j.ijbiomac.2016.04.059
Erko, M., Hartmann, M. A., Zlotnikov, I., Valverde Serrano, C., Fratzl, P., and Politi, Y. (2013). Structural and mechanical properties of the arthropod cuticle: comparison between the fang of the spider Cupiennius salei and the carapace of American lobster Homarus americanus. J. Struct. Biol. 183, 172–179. doi:10.1016/j.jsb.2013.06.001
Feás, X., Vázquez-Tato, M. P., Seijas, J. A., Pratima, G., Nikalje, A., and Fraga-López, F. (2020). Extraction and physicochemical characterization of chitin derived from the Asian hornet, Vespa velutina Lepeletier 1836 (Hym.: vespidae). Molecules 25, 384. doi:10.3390/molecules25020384
Gandhi, M., Shanavas, J., Rebello, S., and S, J. M. (2014). A Simple and effective method for extraction of high purity chitosan from shrimp shell waste. Proc. Intl. Conf. Adv. Appl. Sci. Environ. Engineering-ASEE. doi:10.15224/978-1-63248-004-0-93
Ghosh, N., and Dhepe, P. L. (2021). HPLC method development for chitin and chitosan valorisation chemistry. Carbohydr. Polym. Technol. Appl. 2, 100139. doi:10.1016/j.carpta.2021.100139
Hahn, T., Tafi, E., Paul, A., Salvia, R., Falabella, P., and Zibek, S. (2020). Current state of chitin purification and chitosan production from insects. J. Chem. Technol. Biotechnol. 95, 2775–2795. doi:10.1002/jctb.6533
Hahn, T., Tafi, E., von Seggern, N., Falabella, P., Salvia, R., Thomä, J., et al. (2022). Purification of Chitin from pupal exuviae of the black soldier fly. Waste Biomass Valorization 13, 1993–2008. doi:10.1007/s12649-021-01645-1
Hajji, S., Younes, I., Ghorbel-Bellaaj, O., Hajji, R., Rinaudo, M., Nasri, M., et al. (2014). Structural differences between chitin and chitosan extracted from three different marine sources. Int. J. Biol. Macromol. 65, 298–306. doi:10.1016/j.ijbiomac.2014.01.045
Hamed, I., Özogul, F., and Regenstein, J. M. (2016). Industrial applications of crustacean by-products (chitin, chitosan, and chitooligosaccharides): a review. Trends Food Sci. Technol. 48, 40–50. doi:10.1016/j.tifs.2015.11.007
Hassainia, A., Satha, H., and Boufi, S. (2018). Chitin from agaricus bisporus: extraction and characterization. Int. J. Biol. Macromol. 117, 1334–1342. doi:10.1016/J.IJBIOMAC.2017.11.172
Henriques, B. S., Garcia, E. S., Azambuja, P., and Genta, F. A. (2020). Determination of chitin content in insects: an alternate method based on calcofluor staining. Front. Physiol. 11, 117. doi:10.3389/fphys.2020.00117
Hossin, M. A., Al Shaqsi, N. H. K., Al Touby, S. S. J., and Al Sibani, M. A. (2021). A review of polymeric chitin extraction, characterization, and applications. Arab. J. Geosci. 14, 1870. doi:10.1007/S12517-021-08239-0
Hou, J., Aydemir, B. E., and Dumanli, A. G. (2021). Understanding the structural diversity of chitins as a versatile biomaterial. Philos. Trans. A Math. Phys. Eng. Sci. 379, 20200331. doi:10.1098/rsta.2020.0331
Huet, G., Hadad, C., Husson, E., Laclef, S., Lambertyn, V., Araya Farias, M., et al. (2020). Straightforward extraction and selective bioconversion of high purity chitin from Bombyx eri larva: toward an integrated insect biorefinery. Carbohydr. Polym. 228, 115382. doi:10.1016/J.CARBPOL.2019.115382
Huq, T., Khan, A., Brown, D., Dhayagude, N., He, Z., and Ni, Y. (2022). Sources, production and commercial applications of fungal chitosan: a review. J. Bioresour. Bioprod. 7, 85–98. doi:10.1016/j.jobab.2022.01.002
Iber, B. T., Kasan, N. A., Torsabo, D., and Omuwa, J. W. (2022). A review of various sources of chitin and chitosan in nature. J. Renew. Mater 10, 1097–1123. doi:10.32604/JRM.2022.018142
Ibitoye, E. B., Lokman, I. H., Hezmee, M. N. M., Goh, Y. M., Zuki, A. B. Z., and Jimoh, A. A. (2018). Extraction and physicochemical characterization of chitin and chitosan isolated from house cricket. Biomed. Mat. 13, 025009. doi:10.1088/1748-605X/aa9dde
Ibram, A., Ionescu, A.-M., and Cadar, E. (2019). Comparison of extraction methods of chitin and chitosan from different sources. Eur. J. Med. Nat. Sci. 2, 23. doi:10.26417/688wvv48e
Ifuku, S., Nogi, M., Abe, K., Yoshioka, M., Morimoto, M., Saimoto, H., et al. (2009). Preparation of chitin nanofibers with a uniform width as α-chitin from crab shells. Biomacromolecules 10, 1584–1588. doi:10.1021/BM900163D
Jang, M., Kong, B., Jeong, Y., Lee, C. H., and Nah, J. (2004). Physicochemical characterization of α-chitin, β-chitin, and γ-chitin separated from natural resources. J. Polym. Sci. A Polym. Chem. 42, 3423–3432. doi:10.1002/pola.20176
Jędrzejczak, E., Frąckowiak, P., Sibillano, T., Brendler, E., Giannini, C., Jesionowski, T., et al. (2024). Isolation and structure analysis of chitin obtained from different developmental stages of the mulberry silkworm (Bombyx mori). Molecules 29, 1914. doi:10.3390/molecules29091914
Jiang, F., Li, X., Duan, Y., Li, Q., Qu, Y., Zhong, G., et al. (2023). Extraction and characterization of chitosan from Eupolyphaga sinensis Walker and its application in the preparation of electrospinning nanofiber membranes. Colloids Surf. B Biointerfaces 222, 113030. doi:10.1016/j.colsurfb.2022.113030
Jiao, D., Li, Z., Hu, J. Y., Zhang, X. N., Guo, J., Zheng, Q., et al. (2023). Supramolecular cross-linking affords chitin nanofibril nanocomposites with high strength and water resistance. Compos Sci. Technol. 244, 110295. doi:10.1016/J.COMPSCITECH.2023.110295
Johney, J., Eagappan, K., and Ragunathan, R. R. (2016). Microbial extraction of chitin and chitosan from Pleurotus spp, its characterization and antimicrobial activity. Int. J. Curr. Pharm. Res. 9, 88. doi:10.22159/ijcpr.2017v9i1.16623
Kabalak, M., Aracagök, D., and Torun, M. (2020). Extraction, characterization and comparison of chitins from large bodied four Coleoptera and Orthoptera species. Int. J. Biol. Macromol. 145, 402–409. doi:10.1016/J.IJBIOMAC.2019.12.194
Kaczmarek, M. B., Struszczyk-Swita, K., Li, X., Szczęsna-Antczak, M., and Daroch, M. (2019). Enzymatic Modifications of chitin, chitosan, and chitooligosaccharides. Front. Bioeng. Biotechnol. 7, 243. doi:10.3389/fbioe.2019.00243
Kaur, S., and Dhillon, G. S. (2015). Recent trends in biological extraction of chitin from marine shell wastes: a review. Crit. Rev. Biotechnol. 35, 44–61. doi:10.3109/07388551.2013.798256
Kaya, M., Akyuz, B., Bulut, E., Sargin, I., Eroglu, F., and Tan, G. (2016a). Chitosan nanofiber production from Drosophila by electrospinning. Int. J. Biol. Macromol. 92, 49–55.
Kaya, M., Bağrıaçık, N., Seyyar, O., and Baran, T. (2015a). Comparison of chitin structures derived from three common wasp species (Vespa crabro Linnaeus, 1758, Vespa orientalis Linnaeus, 1771 and Vespula germanica (Fabricius, 1793)). Arch. Insect Biochem. Physiol. 89, 204–217. doi:10.1002/arch.21237
Kaya, M., Baran, T., Asan-Ozusaglam, M., Cakmak, Y. S., Tozak, K. O., Mol, A., et al. (2015b). Extraction and characterization of chitin and chitosan with antimicrobial and antioxidant activities from cosmopolitan Orthoptera species (Insecta). Biotechnol. Bioprocess Eng. 20, 168–179. doi:10.1007/s12257-014-0391-z
Kaya, M., Baran, T., Erdoğan, S., Menteş, A., Aşan Özüsağlam, M., and Çakmak, Y. S. (2014a). Physicochemical comparison of chitin and chitosan obtained from larvae and adult Colorado potato beetle (Leptinotarsa decemlineata). Mater Sc. Eng. C 45, 72–81. doi:10.1016/j.msec.2014.09.004
Kaya, M., Baran, T., Mentes, A., Asaroglu, M., Sezen, G., and Tozak, K. O. (2014b). Extraction and characterization of α-chitin and chitosan from six different aquatic invertebrates. Food Biophys. 9, 145–157. doi:10.1007/s11483-013-9327-y
Kaya, M., Baublys, V., Can, E., Šatkauskienė, I., Bitim, B., Tubelytė, V., et al. (2014c). Comparison of physicochemical properties of chitins isolated from an insect (Melolontha melolontha) and a crustacean species (Oniscus asellus). Zoomorphology 133, 285–293. doi:10.1007/s00435-014-0227-6
Kaya, M., Baublys, V., Sargin, I., Šatkauskienė, I., Paulauskas, A., Akyuz, B., et al. (2016b). How taxonomic relations affect the physicochemical properties of chitin. Food Biophys. 11, 10–19. doi:10.1007/s11483-015-9404-5
Kaya, M., Bitim, B., Mujtaba, M., and Koyuncu, T. (2015c). Surface morphology of chitin highly related with the isolated body part of butterfly (Argynnis pandora). Int. J. Biol. Macromol. 81, 443–449. doi:10.1016/j.ijbiomac.2015.08.021
Kaya, M., Bulut, E., Mujtaba, M., Sivickis, K., Sargin, I., Akyuz, B., et al. (2016c). Gender influences differentiation of chitin among body parts. Arch. Insect Biochem. Physiol. 93, 96–109. doi:10.1002/arch.21344
Kaya, M., Erdogan, S., Mol, A., and Baran, T. (2015d). Comparison of chitin structures isolated from seven Orthoptera species. Int. J. Biol. Macromol. 72, 797–805. doi:10.1016/j.ijbiomac.2014.09.034
Kaya, M., Lelešius, E., Nagrockaitė, R., Sargin, I., Arslan, G., Mol, A., et al. (2015e). Differentiations of chitin content and surface morphologies of chitins extracted from male and female grasshopper species. PLoS One 10, e0115531. doi:10.1371/journal.pone.0115531
Kaya, M., Mujtaba, M., Bulut, E., Akyuz, B., Zelencova, L., and Sofi, K. (2015f). Fluctuation in physicochemical properties of chitins extracted from different body parts of honeybee. Carbohydr. Polym. 132, 9–16. doi:10.1016/j.carbpol.2015.06.008
Kaya, M., Mujtaba, M., Ehrlich, H., Salaberria, A. M., Baran, T., Amemiya, C. T., et al. (2017a). On chemistry of γ-chitin. Carbohydr. Polym. 176, 177–186. doi:10.1016/j.carbpol.2017.08.076
Kaya, M., Özcan Tozak, K., Baran, T., Sezen, G., and Sargın, İ. (2013). Natural porous and nano fiber chitin structure from Gammarus argaeus (Gammaridae: Crustacea). EXCLI J.; 12:503–510. doi:10.1016/j.ijbiomac.2016.07.021
Kaya, M., Sargin, I., Sabeckis, I., Noreikaite, D., Erdonmez, D., Salaberria, A. M., et al. (2017b). Biological, mechanical, optical, and physicochemical properties of natural chitin films obtained from the dorsal pronotum and the wing of cockroach. Carbohydr. Polym. 163, 162–169. doi:10.1016/j.carbpol.2017.01.022
Kaya, M., Sofi, K., Sargin, I., and Mujtaba, M. (2016d). Changes in physicochemical properties of chitin at developmental stages (larvae, pupa and adult) of Vespa crabro (wasp). Carbohydr. Polym. 145, 64–70. doi:10.1016/j.carbpol.2016.03.010
Khajavian, M., Vatanpour, V., Castro-Muñoz, R., and Boczkaj, G. (2022). Chitin and derivative chitosan-based structures — preparation strategies aided by deep eutectic solvents: a review. Carbohydr. Polym. 275, 118702. doi:10.1016/J.CARBPOL.2021.118702
Khasanah, K. R., Sato, H., Takahashi, I., and Ozaki, Y. (2015). Intermolecular hydrogen bondings in the poly(3-hydroxybutyrate) and chitin blends: their effects on the crystallization behavior and crystal structure of poly (3-hydroxybutyrate). Polym. Guildf. 75, 141–150. doi:10.1016/j.polymer.2015.08.011
Khayrova, A., Lopatin, S., and Varlamov, V. (2021). Obtaining chitin, chitosan and their melanin complexes from insects. Int. J. Biol. Macromol. 167, 1319–1328. doi:10.1016/j.ijbiomac.2020.11.086
Kim, M., Song, Y., Han, Y. S., Jo, Y. H., Choi, M. H., Park, Y., et al. (2017). Production of chitin and chitosan from the exoskeleton of adult two-spotted field crickets (Gryllus bimaculatus). Entomol. Res. 47, 279–285. doi:10.1111/1748-5967.12239
Kipkoech, C. (2023). Beyond proteins—edible insects as a source of dietary fiber. Polysaccharides 4, 116–128. doi:10.3390/polysaccharides4020009
Kisaakye, J., Beesigamukama, D., Haukeland, S., Subramanian, S., Thiongo, P. K., Kelemu, S., et al. (2024). Chitin-enriched insect frass fertilizer as a biorational alternative for root-knot nematode (Meloidogyne incognita) management. Front. Plant Sci. 15, 1361739. doi:10.3389/fpls.2024.1361739
Kumar, M., Vivekanand, V., and Pareek, N. (2020). “Insect chitin and chitosan: structure, properties, production, and implementation prospective,” in Natural materials and products from insects: chemistry and applications (Cham: Springer International Publishing), 51–66. doi:10.1007/978-3-030-36610-0_4
Kumari, S., and Kishor, R. (2020). Chitin and chitosan: origin, properties, and applications. Handb. Chitin Chitosan: Volume 1, 1–33. doi:10.1016/B978-0-12-817970-3.00001-8
Liu, S., Sun, J., Yu, L., Zhang, C., Bi, J., Zhu, F., et al. (2012). Extraction and characterization of chitin from the beetle Holotrichia parallela Motschulsky. Molecules 17, 4604–4611. doi:10.3390/molecules17044604
Liu, X., Zhang, J., and Zhu, K. Y. (2019). Chitin in arthropods: biosynthesis, modification, and metabolism. Adv. Exp. Med. Biol. 1142, 169–207. doi:10.1007/978-981-13-7318-3_9
Liu, Y., Xing, R., Yang, H., Liu, S., Qin, Y., Li, K., et al. (2020). Chitin extraction from shrimp (Litopenaeus vannamei) shells by successive two-step fermentation with Lactobacillus rhamnoides and Bacillus amyloliquefaciens. Int. J. Biol. Macromol. 148, 424–433. doi:10.1016/J.IJBIOMAC.2020.01.124
Lopez-Santamarina, A., Mondragon, A. del C., Lamas, A., Miranda, J. M., Franco, C. M., and Cepeda, A. (2020). Animal-origin prebiotics based on chitin: an alternative for the future? a critical review. Foods 9, 782. doi:10.3390/foods9060782
Lv, J., Lv, X., Ma, M., Oh, D. H., Jiang, Z., and Fu, X. (2023). Chitin and chitin-based biomaterials: a review of advances in processing and food applications. Carbohydr. Polym. 299, 120142. doi:10.1016/J.CARBPOL.2022.120142
Ma, J., Xin, C., and Tan, C. (2015). Preparation, physicochemical and pharmaceutical characterization of chitosan from Catharsius molossus residue. Int. J. Biol. Macromol. 80, 547–556. doi:10.1016/j.ijbiomac.2015.07.027
Ma, X., Gözaydın, G., Yang, H., Ning, W., Han, X., Poon, N. Y., et al. (2020). Upcycling chitin-containing waste into organonitrogen chemicals via an integrated process. Proc. Natl. Acad. Sci. U. S. A. 117, 7719–7728. doi:10.1073/PNAS.1919862117
Machado, S. S. N., Silva, J. B. A. da, Nascimento, R. Q., Lemos, P. V. F., Assis, D. de J., Marcelino, H. R., et al. (2024). Insect residues as an alternative and promising source for the extraction of chitin and chitosan. Int. J. Biol. Macromol. 254, 127773. doi:10.1016/j.ijbiomac.2023.127773
Machałowski, T., Amemiya, C., and Jesionowski, T. (2020). Chitin of araneae origin: structural features and biomimetic applications: a review. Appl. Phys. A 126, 678. doi:10.1007/s00339-020-03867-x
Majtán, J., Bíliková, K., Markovič, O., Gróf, J., Kogan, G., and Šimúth, J. (2007). Isolation and characterization of chitin from bumblebee (Bombus terrestris). Int. J. Biol. Macromol. 40, 237–241. doi:10.1016/j.ijbiomac.2006.07.010
Malerba, M., and Cerana, R. (2020a). Chitin- and chitosan-based derivatives in plant protection against biotic and abiotic stresses and in recovery of contaminated soil and water. Polysaccharides 1, 21–30. doi:10.3390/polysaccharides1010003
Malerba, M., and Cerana, R. (2020b). Chitin- and chitosan-based derivatives in plant protection against biotic and abiotic stresses and in recovery of contaminated soil and water. Polysaccharides 1, 21–30. doi:10.3390/polysaccharides1010003
Malm, M., and Liceaga, A. M. (2021). Physicochemical properties of chitosan from two commonly reared edible cricket species, and its application as a hypolipidemic and antimicrobial agent. Polysaccharides 2, 339–353. doi:10.3390/polysaccharides2020022
Marei, N. H., El-Samie, E. A., Salah, T., Saad, G. R., and Elwahy, A. H. M. (2016). Isolation and characterization of chitosan from different local insects in Egypt. Int. J. Biol. Macromol. 82, 871–877. doi:10.1016/j.ijbiomac.2015.10.024
Mehranian, M., Pourabad, R. F., Bashir, N. S., and Taieban, S. (2017). Physicochemical characterization of chitin from the Mediterranean flour moth, Ephestia kuehniella Zeller (Lepidoptera: Pyralidae). J. Macromol. Sci. A 54, 720–726. doi:10.1080/10601325.2017.1332461
Mei, Z., Kuzhir, P., and Godeau, G. (2024). Update on chitin and chitosan from insects: sources, production, characterization, and biomedical applications. Biomimetics 9, 297. doi:10.3390/biomimetics9050297
Merzendorfer, H., and Zimoch, L. (2003). Chitin metabolism in insects: structure, function and regulation of chitin synthases and chitinases. J. Exp. Biol. 206, 4393–4412. doi:10.1242/jeb.00709
Minke, R., and Blackwell, J. (1978). The structure of α-chitin. J. Mol. Biol. 120, 167–181. doi:10.1016/0022-2836(78)90063-3
Mohan, K., Ganesan, A. R., Ezhilarasi, P. N., Kondamareddy, K. K., Rajan, D. K., Sathishkumar, P., et al. (2022). Green and eco-friendly approaches for the extraction of chitin and chitosan: a review. Carbohydr. Polym. 287, 119349. doi:10.1016/J.CARBPOL.2022.119349
Mohan, K., Ganesan, A. R., Muralisankar, T., Jayakumar, R., Sathishkumar, P., Uthayakumar, V., et al. (2020). Recent insights into the extraction, characterization, and bioactivities of chitin and chitosan from insects. Trends Food Sci. Technol. 105, 17–42. doi:10.1016/j.tifs.2020.08.016
Mohan, K., Muralisankar, T., Jayakumar, R., and Rajeevgandhi, C. (2021). A study on structural comparisons of α-chitin extracted from marine crustacean shell waste. Carbohydr. Polym. Technol. Appl. 2, 100037. doi:10.1016/j.carpta.2021.100037
Mol, A., Kaya, M., Mujtaba, M., and Akyuz, B. (2018). Extraction of high thermally stable and nanofibrous chitin from Cicada (Cicadoidea). Entomol. Res. 48, 480–489. doi:10.1111/1748-5967.12299
Moussian, B. (2013). “The arthropod cuticle,” in Arthropod biology and evolution (Berlin, Heidelberg: Springer Berlin Heidelberg), 171–196. doi:10.1007/978-3-642-36160-9_8
Moussian, B. (2019). Chitin: structure, chemistry and biology. Adv. Exp. Med. Biol. 1142, 5–18. doi:10.1007/978-981-13-7318-3_2
Mouyna, I., Dellière, S., Beauvais, A., Gravelat, F., Snarr, B., Lehoux, M., et al. (2020). What are the functions of chitin deacetylases in Aspergillus fumigatus? Front. Cell. Infect. Microbiol. 10, 28. doi:10.3389/fcimb.2020.00028
Muthukrishnan, S., Mun, S., Noh, M. Y., Geisbrecht, E. R., and Arakane, Y. (2020). Insect cuticular chitin contributes to form and function. Curr. Pharm. Des. 26, 3530–3545. doi:10.2174/1381612826666200523175409
Muthukumaran, P., Suresh Babu, P., Shyamalagowri, S., Aravind, J., Kamaraj, M., and Govarthanan, M. (2022). Polymeric biomolecules-based nanomaterials: production strategies and pollutant mitigation as an emerging tool for environmental application. Chemosphere 307, 136008. doi:10.1016/j.chemosphere.2022.136008
Ndiritu, A., Kinyuru, J., Onyango, A., and Kipkoech, C. (2023). Functional and microstructural characteristics of chitin extracted from field cricket, house cricket, and black soldier fly cocoons. J. Food Meas. Charact. 17, 5903–5912. doi:10.1007/s11694-023-02086-1
Nishiyama, Y., Noishiki, Y., and Wada, M. (2011). X-Ray structure of anhydrous β-chitin at 1 Å resolution. Macromolecules 44, 950–957. doi:10.1021/ma102240r
Novikov, V.Yu., Derkach, S. R., Konovalova, I. N., Dolgopyatova, N. V., and Kuchina, Y. A. (2023). Mechanism of heterogeneous alkaline deacetylation of chitin: a review. Polymers 15, 1729. doi:10.3390/polym15071729
Ogawa, Y., Hori, R., Kim, U. J., and Wada, M. (2011). Elastic modulus in the crystalline region and the thermal expansion coefficients of α-chitin determined using synchrotron radiated X-ray diffraction. Carbohydr. Polym. 83, 1213–1217. doi:10.1016/j.carbpol.2010.09.025
Pădurețu, C. C., Isopescu, R. D., Gîjiu, C. L., Rău, I., Apetroaei, M. R., and Schröder, V. (2019). Optimization of chitin extraction procedure from shrimp waste using Taguchi method and chitosan characterization. Mol. Cry Liq. Cry 695, 19–28. doi:10.1080/15421406.2020.1723902
Panchal, K., and Desai, M. (2022). “Chitin: production, purification, and characterization,” in Biotic elicitors. Springer protocols handbooks. Editors D. Amin, N. Amaresan, and S. Ray (New York, NY: Humana). doi:10.1007/978-1-0716-2601-6_11
Pangburn, S. H., Trescony, P. V., and Heller, J. (1982). Lysozyme degradation of partially deacetylated chitin, its films and hydrogels. Biomaterials 3, 105–108. doi:10.1016/0142-9612(82)90043-6
Pedrazzani, C., Righi, L., Vescovi, F., Maistrello, L., and Caligiani, A. (2024). Black soldier fly as a new chitin source: extraction, purification and molecular/structural characterization. LWT 191, 115618. doi:10.1016/j.lwt.2023.115618
Pellis, A., Guebitz, G. M., and Nyanhongo, G. S. (2022). Chitosan: sources, processing and modification techniques. Gels 8, 393. doi:10.3390/gels8070393
Philibert, T., Lee, B. H., and Fabien, N. (2017). Current status and new perspectives on chitin and chitosan as functional biopolymers. Appl. Biochem. Biotechnol. 181, 1314–1337. doi:10.1007/s12010-016-2286-2
Pillai, C. K. S., Paul, W., and Sharma, C. P. (2009). Chitin and chitosan polymers: chemistry, solubility and fiber formation. Prog. Polym. Sci. Oxf. 34, 641–678. doi:10.1016/j.progpolymsci.2009.04.001
Plotnick, R. E. (1990). Paleobiology of the arthropod cuticle. Short Courses Paleontology 3, 177–196. doi:10.1017/S2475263000001793
Poerio, A., Petit, C., Jehl, J. P., Arab-Tehrany, E., Mano, J. F., and Cleymand, F. (2020). Extraction and physicochemical characterization of chitin from Cicada orni sloughs of the south-eastern French Mediterranean basin. Molecules 25, 2543. doi:10.3390/MOLECULES25112543
Pratiwi, R. D., El Muttaqien, S., Gustini, N., Difa, N. S., Syahputra, G., and Rosyidah, A. (2023). Eco-friendly synthesis of chitosan and its medical application: from chitin extraction to nanoparticle preparation. ADMET DMPK 11, 435–455. doi:10.5599/admet.1999
Qin, Z., Hou, Y., Ahamed, W., Li, Y., and Zhao, L. (2019). “Detection and separation of chito/chitin oligosaccharides,” in Oligosaccharides of chitin and chitosan (Singapore: Springer Singapore), 83–105. doi:10.1007/978-981-13-9402-7_5
Ramírez-Wong, D. G., Ramírez-Cardona, M., Sánchez-Leija, R. J., Rugerio, A., Mauricio-Sánchez, R. A., Hernández-Landaverde, M. A., et al. (2016). Sustainable-solvent-induced polymorphism in chitin films. Green Chem. 18, 4303–4311. doi:10.1039/C6GC00628K
Reddy, N., and Yang, Y. (2015). “Introduction to chitin, chitosan, and alginate fibers,” in Innovative biofibers from renewable resources (Berlin, Heidelberg: Springer Berlin Heidelberg), 93–94. doi:10.1007/978-3-662-45136-6_23
Rehman, K., Hollah, C., Wiesotzki, K., Heinz, V., Aganovic, K., Rehman, R., et al. (2023). Insect-derived chitin and chitosan: a still unexploited resource for the edible insect sector. Sustainability 15, 4864. doi:10.3390/su15064864
Rinaudo, M. (2006). Chitin and chitosan: properties and applications. Prog. Polym. Sci. 31, 603–632. doi:10.1016/j.progpolymsci.2006.06.001
Roberts, G. A. F. (1992). Chitin chemistry. London: Macmillan Education UK. doi:10.1007/978-1-349-11545-7
Rodriguez, J., Neyrinck, A. M., Zhang, Z., Seethaler, B., Nazare, J.-A., Robles Sánchez, C., et al. (2020). Metabolite profiling reveals the interaction of chitin-glucan with the gut microbiota. Gut Microbes 12, 1810530. doi:10.1080/19490976.2020.1810530
Roy, J. C., Salaün, F., Giraud, S., Ferri, A., Chen, G., and Guan, J. (2017). “Solubility of chitin: solvents, solution behaviors and their related mechanisms,” in Solubility of polysaccharides London, United Kingdom: IntechOpen Limited. doi:10.5772/intechopen.71385
Saenz-Mendoza, A. I., Zamudio-Flores, P. B., García-Anaya, M. C., Velasco, C. R., Acosta-Muñiz, C. H., Espino-Díaz, M., et al. (2023). Insects as a potential source of chitin and chitosan: physicochemical, morphological and structural characterization. -A review. Emir J. Food Agric. doi:10.9755/ejfa.2023.v35.i5.3095
Sáenz-Mendoza, A. I., Zamudio-Flores, P. B., Palomino-Artalejo, G. A., Tirado-Gallegos, J. M., García-Cano, V. G., Ornelas-Paz, J. J., et al. (2018). Physicochemical, morphological and structural characterization of the chitin and chitosan of Tenebrio molitor and Galleria mellonella insects. Rev. Mex. Ing. Quim 19, 39–56. doi:10.24275/uam/izt/dcbi/revmexingquim/2019v18n1/Saenz
Sagheer, F. A.Al, Al-Sughayer, M. A., Muslim, S., and Elsabee, M. Z. (2009). Extraction and characterization of chitin and chitosan from marine sources in Arabian Gulf. Carbohydr. Polym. 77, 410–419. doi:10.1016/j.carbpol.2009.01.032
Sajomsang, W., and Gonil, P. (2010). Preparation and characterization of α-chitin from cicada sloughs. Mat. Sci. Eng. C 30, 357–363. doi:10.1016/j.msec.2009.11.014
Sanjanwala, D., Londhe, V., Trivedi, R., Bonde, S., Sawarkar, S., Kale, V., et al. (2024). Polysaccharide-based hydrogels for medical devices, implants and tissue engineering: a review. Int. J. Biol. Macromol. 256, 128488. doi:10.1016/j.ijbiomac.2023.128488
Shahrajabian, M. H., Chaski, C., Polyzos, N., Tzortzakis, N., and Petropoulos, S. A. (2021). Sustainable agriculture systems in vegetable production using chitin and chitosan as plant biostimulants. Biomolecules 11, 819. doi:10.3390/biom11060819
Sharbidre, A., Sargar, S., Gogoi, H., and Patil, R. (2021). Characterization of chitin content extracted from edible insect, Coridius nepalensis (Westwood, 1837) (Hemiptera: dinidoridae). Int. J. Trop. Insect Sci. 41, 1893–1900. doi:10.1007/s42690-020-00386-3
Sharp, R. (2013). A review of the applications of chitin and its derivatives in agriculture to modify plant-microbial interactions and improve crop yields. Agronomy 3, 757–793. doi:10.3390/agronomy3040757
Shin, C. S., Kim, D. Y., and Shin, W. S. (2019). Characterization of chitosan extracted from mealworm beetle (Tenebrio molitor, Zophobas morio) and rhinoceros beetle (Allomyrina dichotoma) and their antibacterial activities. Int. J. Biol. Macromol. 125, 72–77. doi:10.1016/j.ijbiomac.2018.11.242
Silva, A. C. Q., Silvestre, A. J. D., Vilela, C., and Freire, C. S. R. (2021). Natural polymers-based materials: a contribution to a greener future. Molecules 27, 94. doi:10.3390/molecules27010094
Sivashankari, P. R., and Prabaharan, M. (2017). “Deacetylation modification techniques of chitin and chitosan,” Chitosan Based Biomater., 1. 117–133. doi:10.1016/B978-0-08-100230-8.00005-4
Soetemans, L., Uyttebroek, M., and Bastiaens, L. (2020). Characteristics of chitin extracted from black soldier fly in different life stages. Int. J. Biol. Macromol. 165, 3206–3214. doi:10.1016/j.ijbiomac.2020.11.041
Soon, C. Y., Tee, Y. B., Tan, C. H., Rosnita, A. T., and Khalina, A. (2018). Extraction and physicochemical characterization of chitin and chitosan from Zophobas morio larvae in varying sodium hydroxide concentration. Int. J. Biol. Macromol. 108, 135–142. doi:10.1016/j.ijbiomac.2017.11.138
Spranghers, T., Ottoboni, M., Klootwijk, C., Ovyn, A., Deboosere, S., De Meulenaer, B., et al. (2017). Nutritional composition of black soldier fly (Hermetia illucens) prepupae reared on different organic waste substrates. J. Sci. Food Agric. 97, 2594–2600. doi:10.1002/jsfa.8081
Ssekatawa, K., Byarugaba, D. K., Wampande, E. M., Moja, T. N., Nxumalo, E., Maaza, M., et al. (2021). Isolation and characterization of chitosan from Ugandan edible mushrooms, Nile perch scales and banana weevils for biomedical applications. Sci. Rep. 11, 4116. doi:10.1038/s41598-021-81880-7
Susana Cortizo, M., Berghoff, C. F., and Alessandrini, J. L. (2008). Characterization of chitin from Illex argentinus squid pen. Carbohydr. Polym. 74, 10–15. doi:10.1016/j.carbpol.2008.01.004
Teng, W. L., Khor, E., Tan, T. K., Lim, L. Y., and Tan, S. C. (2001). Concurrent production of chitin from shrimp shells and fungi. Carbohydr. Res. 332, 305–316. doi:10.1016/S0008-6215(01)00084-2
Terkula Iber, B., Azman Kasan, N., Torsabo, D., and Wese Omuwa, J. (2022). A review of various sources of chitin and chitosan in nature. J. Renew. Mater 10, 1097–1123. doi:10.32604/jrm.2022.018142
Thakur, D., Bairwa, A., Dipta, B., Jhilta, P., and Chauhan, A. (2023). An overview of fungal chitinases and their potential applications. Protoplasma 260, 1031–1046. doi:10.1007/s00709-023-01839-5
Tolaimate, A., Desbrieres, J., Rhazi, M., and Alagui, A. (2003). Contribution to the preparation of chitins and chitosans with controlled physico-chemical properties. Polym. Guildf. 44, 7939–7952. doi:10.1016/j.polymer.2003.10.025
Triunfo, M., Tafi, E., Guarnieri, A., Salvia, R., Scieuzo, C., Hahn, T., et al. (2022). Characterization of chitin and chitosan derived from Hermetia illucens, a further step in a circular economy process. Sci. Rep. 12, 6613. doi:10.1038/s41598-022-10423-5
Tsurkan, M. V., Voronkina, A., Khrunyk, Y., Wysokowski, M., Petrenko, I., and Ehrlich, H. (2021). Progress in chitin analytics. Carbohydr. Polym. 252, 117204. doi:10.1016/j.carbpol.2020.117204
Velásquez, C. L., and Pirela, M. R. (2016). “Biochemical aspects of the chitin fungicidal activity in agricultural uses,” in Chitosan in the preservation of agricultural commodities (Elsevier), 279–298. doi:10.1016/B978-0-12-802735-6.00010-0
Verardi, A., Sangiorgio, P., Moliterni, S., Errico, S., Spagnoletta, A., and Dimatteo, S. (2023). Advanced technologies for chitin recovery from crustacean waste. Clean. Technol. Recycl 3, 4–43. doi:10.3934/ctr.2023002
Vicente, F. A., Huš, M., Likozar, B., and Novak, U. (2021). Chitin deacetylation using deep eutectic solvents: ab initio -supported process optimization. ACS Sustain Chem. Eng. 9, 3874–3886. doi:10.1021/acssuschemeng.0c08976
Wang, J., Chen, Z., and Naguib, H. E. (2021). Preparation of a novel double crosslinked chitin aerogel via etherification with high strength. Carbohydr. Polym. 265, 118014. doi:10.1016/j.carbpol.2021.118014
Wang, Y., Chang, Y., Yu, L., Zhang, C., Xu, X., Xue, Y., et al. (2013). Crystalline structure and thermal property characterization of chitin from Antarctic krill Euphausia superba. Euphausia Superba. Carbohydr. Polym. 92, 90–97. doi:10.1016/j.carbpol.2012.09.084
Waśko, A., Bulak, P., Polak-Berecka, M., Nowak, K., Polakowski, C., and Bieganowski, A. (2016). The first report of the physicochemical structure of chitin isolated from Hermetia illucens. Int. J. Biol. Macromol. 92, 316–320. doi:10.1016/j.ijbiomac.2016.07.038
Wegrzynowska-Drzymalska, K., Grebicka, P., Mlynarczyk, D. T., Chelminiak-Dudkiewicz, D., Kaczmarek, H., Goslinski, T., et al. (2020). Crosslinking of chitosan with dialdehyde chitosan as a new approach for biomedical applications. Materials 13, 3413. doi:10.3390/ma13153413
Weiss, I. (2012). Species-specific shells: chitin synthases and cell mechanics in molluscs. Z Krist. Cryst. Mater 227, 723–738. doi:10.1524/zkri.2012.1530
Xiong, A., Ruan, L., Ye, K., Huang, Z., and Yu, C. (2023). Extraction of chitin from black soldier fly (Hermetia illucens) and its puparium by using biological treatment. Life 13, 1424. doi:10.3390/life13071424
Yanat, M., Colijn, I., de Boer, K., and Schroën, K. (2023). Comparison of the degree of acetylation of chitin nanocrystals measured by various analysis methods. Polym. (Basel) 15, 294. doi:10.3390/polym15020294
Yeul, V. S., and Rayalu, S. S. (2013). Unprecedented chitin and chitosan: a chemical overview. J. Polym. Environ. 21, 606–614. doi:10.1007/s10924-012-0458-x
Younes, I., and Rinaudo, M. (2015). Chitin and chitosan preparation from marine sources. Structure, properties and applications. Mar. Drugs 13, 1133–1174. doi:10.3390/md13031133
Yu, A., Beck, M., Merzendorfer, H., and Yang, Q. (2024). Advances in understanding insect chitin biosynthesis. Insect Biochem. Mol. Biol. 164, 104058. doi:10.1016/j.ibmb.2023.104058
Zainol Abidin, N. A., Kormin, F., Zainol Abidin, N. A., Mohamed Anuar, N. A. F., and Abu Bakar, M. F. (2020a). The potential of insects as alternative sources of chitin: an overview on the chemical method of extraction from various sources. Int. J. Mol. Sci. 21, 4978. doi:10.3390/ijms21144978
Zainol Abidin, N. A., Kormin, F., Zainol Abidin, N. A., Mohamed Anuar, N. A. F., and Abu Bakar, M. F. (2020b). The potential of insects as alternative sources of chitin: an overview on the chemical method of extraction from various sources. Int. J. Mol. Sci. 21, 4978. doi:10.3390/ijms21144978
Zargar, V., Asghari, M., and Dashti, A. (2015). A review on chitin and chitosan polymers: structure, chemistry, solubility, derivatives, and applications. ChemBioEng Rev. 2, 204–226. doi:10.1002/CBEN.201400025
Zhang, J., Feng, M., Lu, X., Shi, C., Li, X., Xin, J., et al. (2018). Base-free preparation of low molecular weight chitin from crab shell. Carbohydr. Polym. 190, 148–155. doi:10.1016/j.carbpol.2018.02.019
Zhang, M., Haga, A., Sekiguchi, H., and Hirano, S. (2000). Structure of insect chitin isolated from beetle larva cuticle and silkworm (Bombyx mori) pupa exuvia. Int. J. Biol. Macromol. 27, 99–105. doi:10.1016/S0141-8130(99)00123-3
Zhang, X., Yuan, J., Li, F., and Xiang, J. (2021). Chitin synthesis and degradation in crustaceans: a genomic view and application. Mar. Drugs 19, 153. doi:10.3390/md19030153
Keywords: insects, chitin, chitosan, crustacean, arthropod, demineralization
Citation: Izadi H, Asadi H and Bemani M (2025) Chitin: a comparison between its main sources. Front. Mater. 12:1537067. doi: 10.3389/fmats.2025.1537067
Received: 29 November 2024; Accepted: 07 February 2025;
Published: 04 March 2025.
Edited by:
Luiz Fernando Romanholo Ferreira, Catholic University of Brasilia (UCB), BrazilReviewed by:
Nazrin Asmawi, Universiti Putra Malaysia Bintulu Sarawak Campus, MalaysiaCopyright © 2025 Izadi, Asadi and Bemani. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Hamzeh Izadi, aXphZGlAdnJ1LmFjLmly
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.