
95% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
SYSTEMATIC REVIEW article
Front. Mar. Sci. , 14 March 2025
Sec. Marine Fisheries, Aquaculture and Living Resources
Volume 12 - 2025 | https://doi.org/10.3389/fmars.2025.1539725
This article is part of the Research Topic Alternative Feed Ingredients and their Functional Properties in Aquaculture View all 10 articles
 Paolo Guttuso1,2*
Paolo Guttuso1,2* Natacha Nogueira3
Natacha Nogueira3 Sonia K. M. Gueroun3,4
Sonia K. M. Gueroun3,4 Jamileh Javidpour5
Jamileh Javidpour5 João Canning-Clode3,4,6
João Canning-Clode3,4,6 Carlos A. P. Andrade3,4
Carlos A. P. Andrade3,4Jellyfish’s potential for feed production remains largely unexplored and research on their nutritional benefits in aquafeeds is still limited. This systematic review analyzed the nutritional composition of jellyfish and its potential as a sustainable aquaculture feed ingredient, evaluating advantages and limitations. Data from 65 studies were categorized into proximate composition, amino acids, fatty acids, and mineral content. Good proportion of methionine and lysine, high amount of collagen-derived amino acids (glycine, proline, hydroxyproline), the presence of taurine and beneficial long-chain fatty acids (mainly ARA), as well as richness in minerals such as Na, K, Cl, Mg, and Zn, constitute attractive key characteristics for feed application. However, challenges remain, including high moisture and ash content, elevated aluminum levels from present processing methods, and compositional variability. Improved processing methods may enhance their use, but further research is needed to address digestibility, optimize processing, and assess long-term sustainability. This study positions jellyfish as a valuable, sustainable supplement for aquaculture feed, though comprehensive evaluations are necessary to unlock their full potential and ensure consistent quality in commercial applications.
Jellyfish, belong to the Medusozoa classes Hydrozoa, Scyphozoa, and Cubozoa (Boero, 2013), and hold a unique position as one of the oldest metazoan animal groups on Earth (Cartwright et al., 2007), populating oceans worldwide from surface to bottom (Graham et al., 2014). In general, they present a bipartite life cycle with an asexual reproductive sessile stage (polyp, hydroid) and a sexual reproductive pelagic stage (medusa) (Jarms and Morandini, 2019).
Jellyfish (herein referring only to the Class Scyphozoa) have a rich cultural history in China, where they have been esteemed as a food source for centuries and recognized for their medicinal properties (Hsieh and Rudloe, 1994). This tradition extends to other Asian countries (e.g., Japan, Malaysia, Korea), where there is substantial market demand (Kingsford et al., 2000; Hsieh et al., 2001; Omori and Nakano, 2001; Raposo et al., 2022) leading to the establishment of an important fisheries sector, particularly in Southeast Asia. About 40 jellyfish species are commercially fished for food purposes, focusing on species like Rhopilema esculentum and Nemopilema nomurai, mainly in China (Brotz, 2016). Additionally, there has been a notable expansion of jellyfish fisheries in Western countries like the USA and Mexico, driven by Eastern market demands and the exploitation of new species such as Stomolophus meleagris (Brotz et al., 2017).
While jellyfish have traditionally been consumed primarily in Asian cuisines, recent European regulations have highlighted their potential as novel food sources, emphasizing the biochemical characterization and bioactive properties of Mediterranean jellyfish species (Regulation (EU) 2283/2015). Beyond food, jellyfish are being explored for various applications such as agriculture fertilizer (Hussein and Saleh, 2014), cosmetics (Zhuang et al., 2009) and biomedical application (Addad et al., 2011), driven by research efforts into their bioactive (Leone et al., 2015, Leone et al., 2019; Upata et al., 2022) and functional properties, particularly collagen (Barzideh et al., 2013).
In recent decades, global jellyfish populations have surged, attributed to anthropogenic factors like climate change (Purcell, 2005), overfishing (Roux et al., 2013) and coastal eutrophication (Purcell et al., 1999). This led to significant blooms impacting various human activities such as tourism (Ruiz-Frau, 2023), coastal industries, fisheries, and aquaculture (Purcell et al., 2007; Dong et al., 2010; Bosch-belmar et al., 2021). Despite these challenges, jellyfish roles in marine ecosystems are being re-evaluated, recognizing their contributions to regulating, supporting, and provisioning ecosystem service as well as economic and social benefits (Doyle et al., 2014).
In this context, expanding jellyfish exploitation may present an exciting opportunity within the Blue Economy framework. This includes advancements in harvesting and processing techniques as well as the potential to tap into currently under-exploited species (Edelist et al., 2021). By utilizing these gelatinous organisms as a valuable resource, we can reshape their perception and unlock their potential.
Furthermore, aquaculture is an emerging industry as a key catalyst in harnessing jellyfish biomass to address the growing demand for sustainable and cost-effective fish feeds. Concerns regarding traditional fish meal and fish oil sources (Hua et al., 2019; Naylor et al., 2021) have fueled the search for alternatives, with jellyfish offering a compelling solution (Duarte et al., 2022; Eroldoğan et al., 2023). Promising results exist from experiments using live jellyfish or fresh portions in marine fish and crustaceans’ diets (Table 1). The scientific literature contains a wealth of valuable data on jellyfish chemical composition, which has been collected and analyzed in reviews focused on biotechnological application (Merquiol et al., 2019; D’Ambra and Merquiol, 2022) and ecological studies (Ikeda, 2014; Hubot et al., 2022).
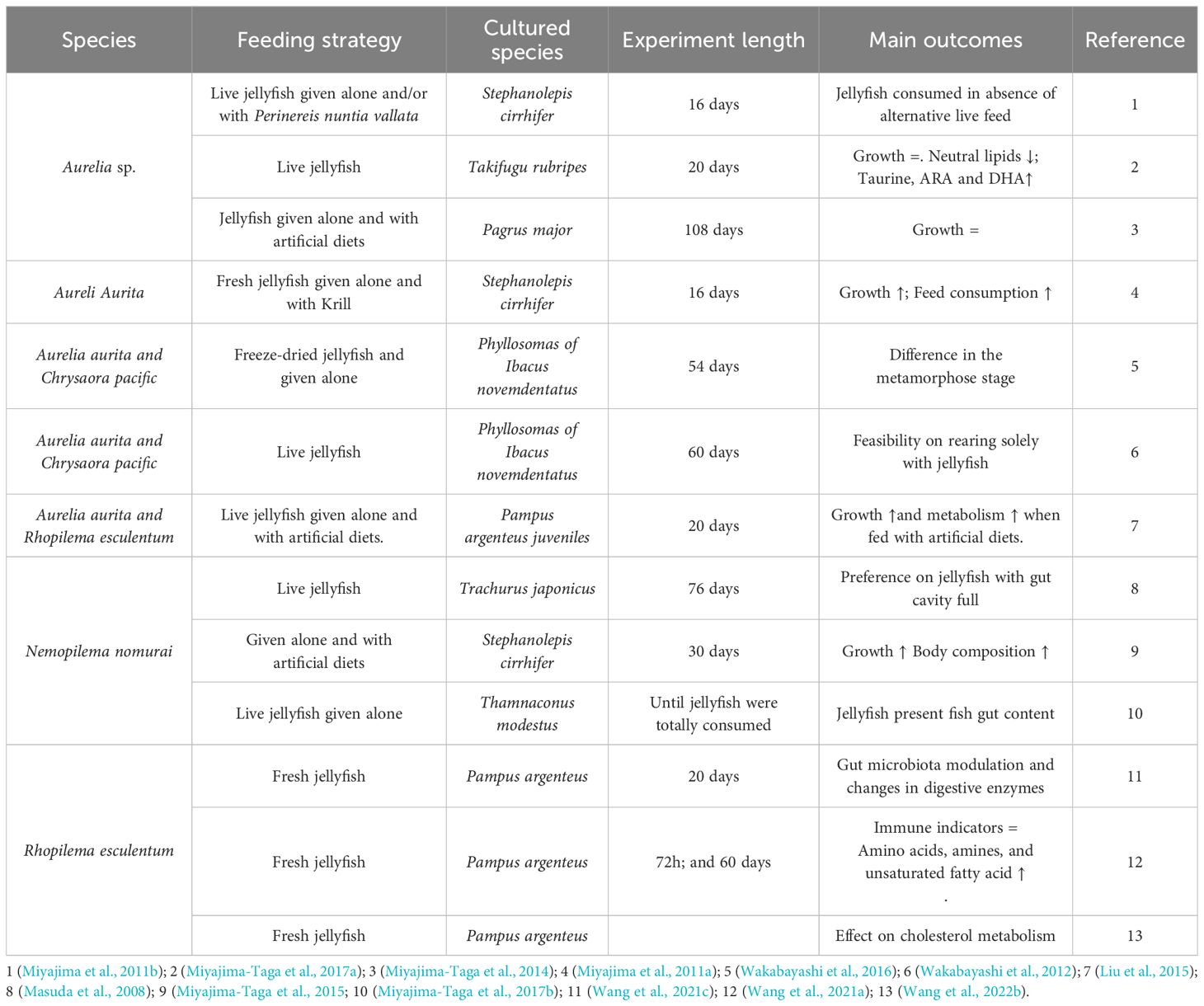
Table 1. Jellyfish used to feed aquatic species.
To fully evaluate jellyfish potential as a novel feed ingredient, a crucial first step is to gather and standardize all available data on species, origin, processing methods and chemical composition (Glencross et al., 2020). In this context, this study aims to standardize and evaluate all the available data on jellyfish nutritional composition through a systematic review method and assessing their nutritional value. Furthermore, this evaluation considers the specific requirements of aquafeed production and ultimately contributes to the exploration of jellyfish as a viable and sustainable source for aquaculture feed.
To ensure a robust and reliable systematic review, we followed the guidelines recommended by the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA), with certain adjustments to better suit our study’s objectives (Liberati et al., 2009; Moher et al., 2009).
A thorough screening of publication lists obtained from three bibliographic databases (i.e. PubMed, Web of Science, and Google Scholar) was conducted. The Google Scholar database was processed with the use of the platform provided by Harzing (2007) (Harzing, 2007) to overstep limitation in Boolean search tools and importable items. The last search date for Google Scholar was October 2020, while for the other databases, it was March 2023.
The search queries in each database utilized the same keywords (“jellyfish” “scyphozoa”, “schyphomedusa, “nutrient” “microelement” “macroelement”, “vitamin”, “protein”, “lipid*”, “ash”, “organic matter”, “carbohydrate”, “amino acid”, “fatty acid”, “elemental”, “biochemical”, “nutritional”, “gross” “proximate”, composition”, “compound”) and were expressed in the appropriate language of the specific database (Supplementary File 1).
To manage the large number of items, a three-step eligibility criteria process was implemented (Figure 1). Firstly, titles were screened for relevance to the paper’s objective or the potential to report nutritional data. Secondly, structured eligibility criteria were applied to define the nutritional composition of jellyfish, excluding publications reporting data on specific substrates like collagen or gelatin and considering only papers focusing on the medusa stage of the Scyphozoa class. Thirdly, only scientific publications with available data in English were considered. Briefly, only data related to wild organisms, whole body, oral arms, bell, and raw material were included, while entirely reared and processed organisms, other body parts such as mesoglea and gonads, and extraction substrates such as collagen and gelatin were excluded. However, aspects influencing nutritional composition and potential use as feed components were discussed in the text.
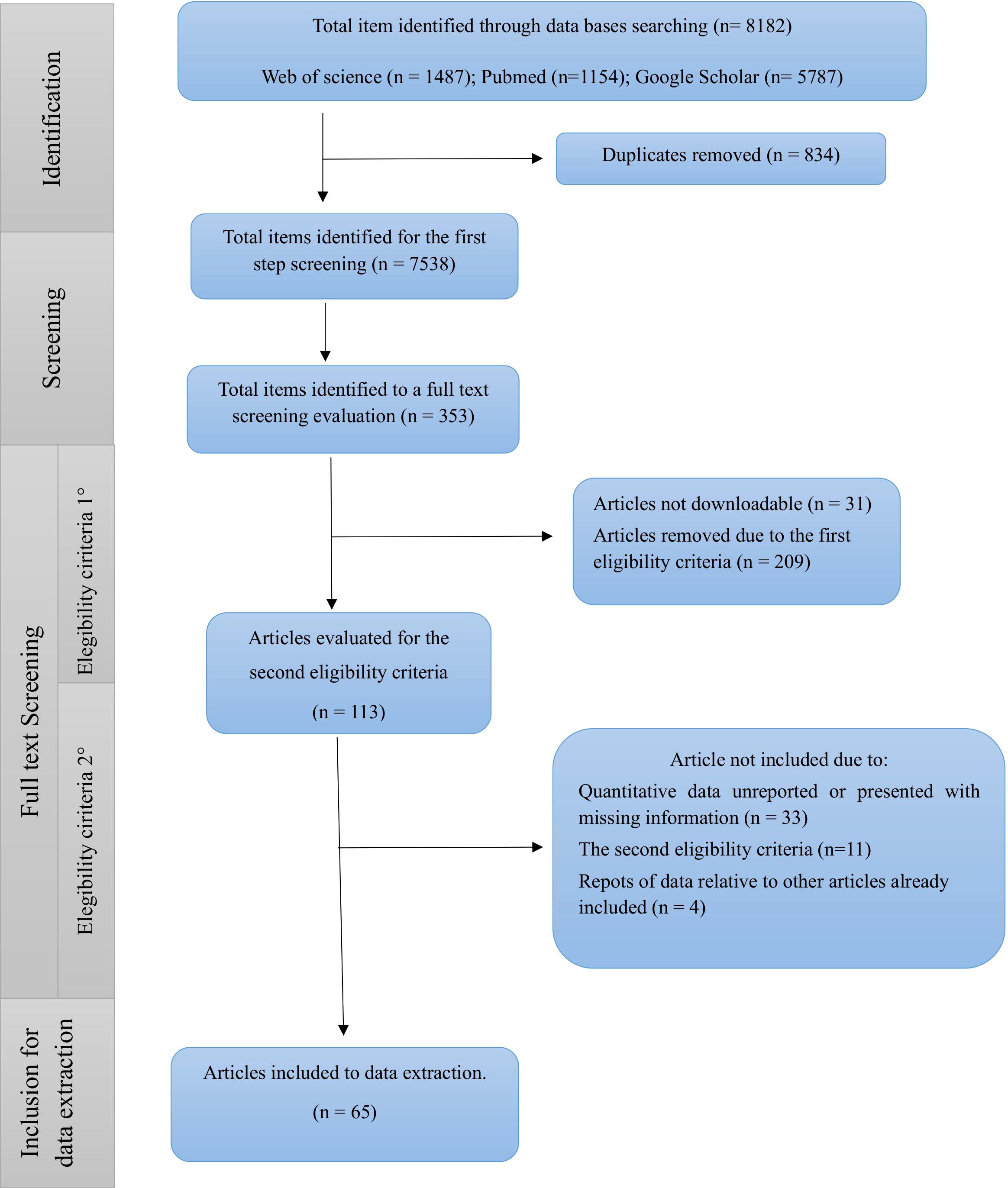
Figure 1. Flow diagram of the steps followed for the identification, eligibility and selection of studies.
Qualitative parameters included in the final tables were family, species, and body parts. Data related to factors not evaluated in the eligibility criteria process, such as site, size, and sex, were extracted as a range or mean if directly available in the reference. Nutritional composition data, if only graphically presented were extracted with an online software WebPlotDigitizer (Rohatgi, 2017).
Data were categorized into four main groups: proximate composition (Pc), amino acids (AA), fatty acids (FA), and minerals (Mi). Each nutritional compound was converted into a unique unit of measurement, applying appropriate unit conversions (Supplementary File 1). Water content data, if missing, was back-calculated from the respective specular dry weight values reported in the references.
Furthermore, nutritional composition data for the most used aquafeed ingredients and supplements were extracted from the NRC (Nutrient Requirements of Fish and Shrimp, 2011) (NRC, 2011) and other sources as needed. Data were unit-converted for direct comparison with the jellyfish nutritional profile.
The systematic search across three electronic databases initially yielded 7538 items, from which 353 publications were identified as potentially highly relevant. Through the multi-step eligibility criteria process, 64 articles were ultimately selected for the extraction of main nutritional compounds as indicated in the flow diagram in Figure 1.
An overview of data set reveals that the Mediterranean Sea, the Northwest Pacific, and the Northeast Atlantic are the primary geographical regions where the nutritional composition of jellyfish has been evaluated (Figure 2A). Additionally, Figure 2A highlights the relative proportion of jellyfish species analyzed for their nutritional composition, categorized by the journal fields in which the data were published. Notably, the field of Aquaculture Nutrition exhibited limited interest in the evaluation of jellyfish nutritional composition
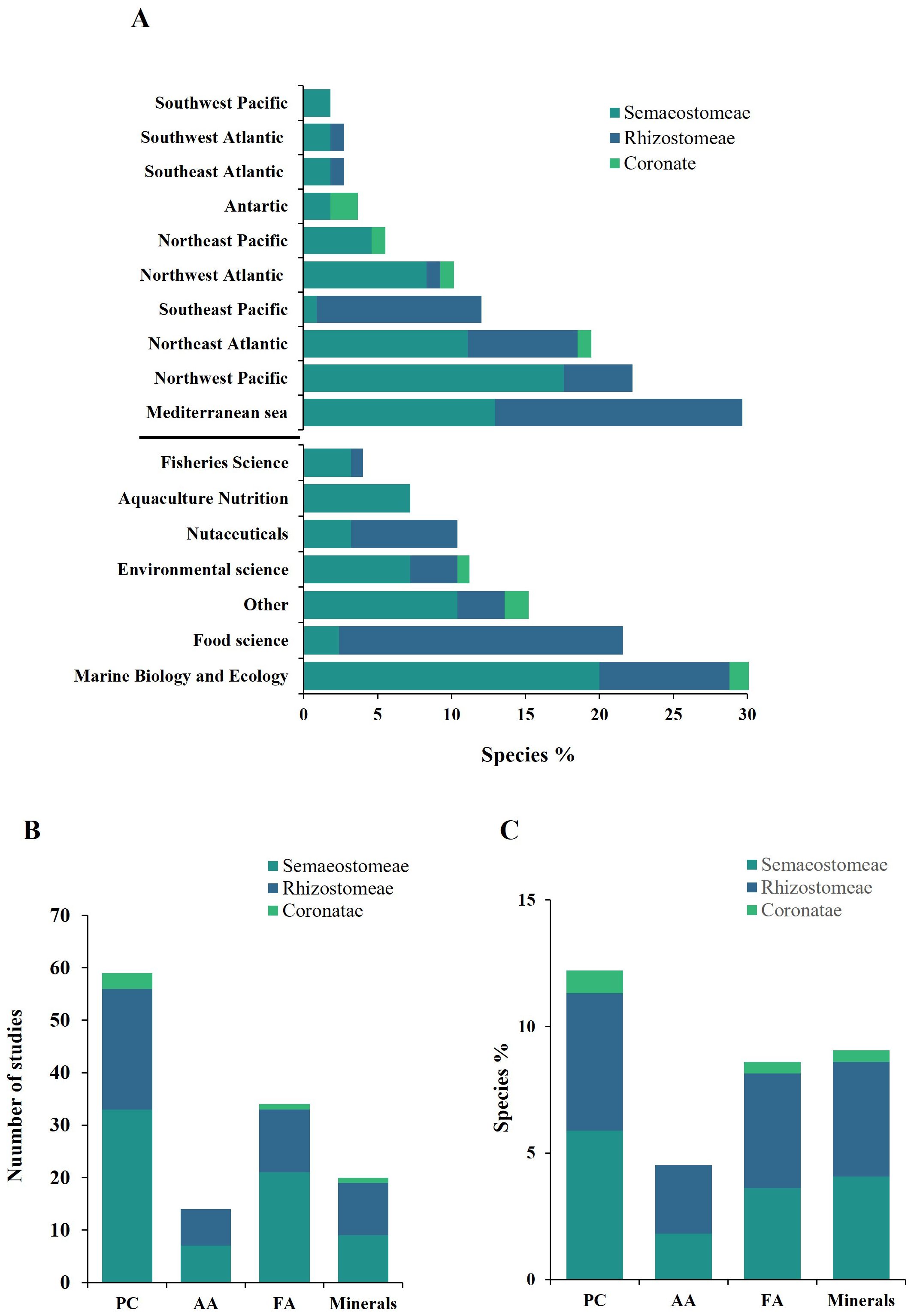
Figure 2. Representativeness of jellyfish species evaluated for nutritional composition. (A) Percentage of jellyfish species out of the total included in this study accounting the geographic location of species collection and the field of interest of the journal that published the nutritional assessment. (B) Number of studies that evaluated nutritional composition of jellyfish species categorized by each nutritional category. (C) Percentage of species evaluated in this study out of the total known species by each nutritional category. Total species: number of accepted species (Jarms and Morandini, 2019); PC (proximate composition); AA (amino acids); FA (fatty acids).
The nutritional variables characterizing the dataset are summarized in Figure 2B. Semaeostomeae, Rhizostomeae, and Coronatae orders accounted for 58%, 36.9%, and 5.1% of the total references, respectively. Aurelia was the most frequently mentioned genus (27.6% of the total references), followed by Rhizostoma (11.8% of the total references). Most of the works reported proximate composition values, but body part data varied. 40% percent of the works did not indicate a specific body part, while oral arms (13%), bell (15%), and whole body (32%) were reported in the remaining works.
Semaeostomeae, representing 51.8% of Scyphozoa diversity (Jarms and Morandini, 2019), are the most studied group within this class, with 12.2% (protein content - Pc), 4.5% (amino acid - AA), 8.6% (fatty acid - FA), and 9.0% (mineral - Mi) of species having been analyzed for nutritional composition (Figure 2C). While the Rhizostoma order constitutes 22% of Scyphozoa diversity, it has been more extensively studied, the Coronate order, the second most diverse within Scyphozoa, has been minimally investigated, with less than 1% of species analyzed for most nutritional components, and no amino acid composition data available (Figure 2C).
Across Scyphozoa species, Pc varied as follows: water content ranged from 91.1 to 98% wet weight (ww), ash from 15.4 to 85.6% dry weight (dw), proteins from 0.2 to 76.8% DW, lipids from 0.17 to 12.3% DW, and carbohydrates from 0.1 to 22.71% DW (Supplementary File 2). Ash content predominated, with consistent ranges observed across species. Protein content exhibited its highest values in Rhopilema esculentum (38.12-53.87% DW) and Stomolophus meleagris (76.8% DW). Lipids were the least abundant fraction, with exceptions such as Cotylorhiza tuberculata. (12.3% DW). The protein and lipid contents varied depending on the body part analyzed. Overall, oral arms exhibited higher protein (13.4-53.87% DW) and lipid concentrations 0.2-2.2% DW) than bell (protein: 6.6-38.12% DW; lipid: 0.17–1% DW), while the whole body showing intermediate values (protein: 1.1-34.2% DW; lipid: 0.3-5.8% DW). Carbohydrate data found in the literature were limited, typically falling within a range of 0.06% DW (for Eupilema inexpectata) to 22.71% DW for (Chrysaora pacifica).
The observed variability in jellyfish chemical composition likely stems from several factors that will be discussed further ahead as well as their potentialities and constraints for aquafeed production.
Jellyfish, characterized by their high moisture and consistent ash content, generally exhibit lower protein content (DW) compared to established feed ingredients like fish meal and soybean meal (Figure 3). However, their protein content is comparable to seaweed (14.4 ± 6.6% DW) and to other plant-based sources such as corn (10.6 ± 1.3% DW), and wheat (15.6 ± 2.5% DW). Certain species, such as Rhopilema esculentum and Rhopilema hispidum (oral arms) display protein content within the range of soya bean meal (Figure 3).
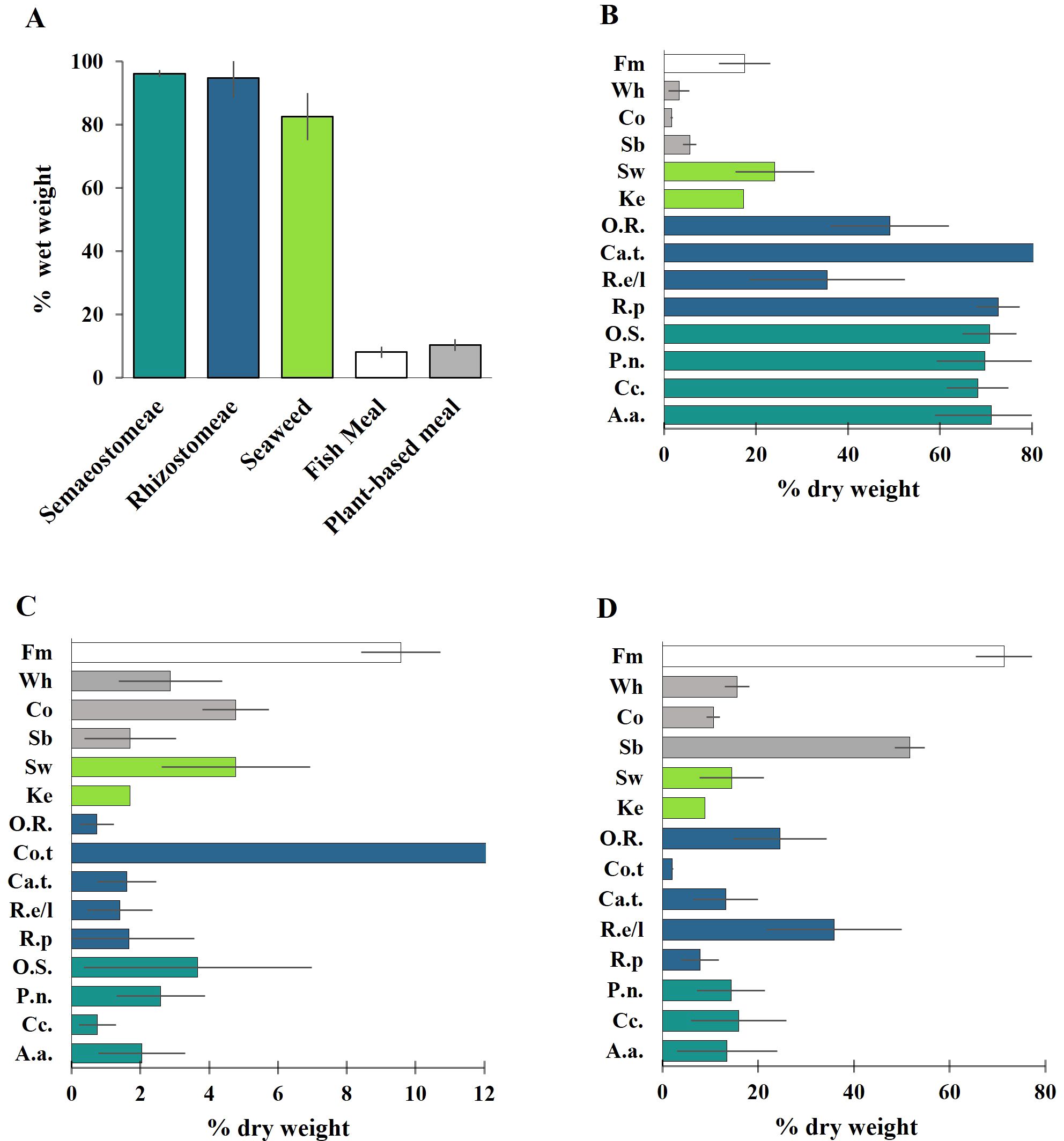
Figure 3. Proximate composition of (A) Moisture (% of ww); (B) Ash (% DW); (C) Lipid (% DW); (D) Protein (% DW) in jellyfish and reference ingredient for aqua-feed production. Note: A.a. (Aurelia aurita), C.c. (Cyanea capillata), P.n (Pelagia noctulica), O.S. (Other Sematostomae), R.p.(Rhizostoma pulmo), R.e/l (Rophilema esculentum and Rhopilema hispidum), Ca.t. (Catostylus tagi); Co.t. (Cotylorhiza tuberculata); O.R. (Other Rizhostoma); Sb (Soyabean); Co (Corn); Wh (Wheat); Fm (Fish meal); Ke (Kelp); Sw (Seaweed). Value are reported as mean and standard deviation of all the values extracted; Jellyfish (This study), other ingredients (NRC, 2011).
Considering the quality of proteins in terms of essential amino acids (EAA), jellyfish present a heterogeneous profile (Supplementary File 2), with lower relative amounts of EAA compared to traditional protein sources like fish meal and soyabean meal (Figure 4). Moreover, jellyfish demonstrate a variable quantitative EAA profile across species (Semaestomae generally displaying lower amino acid levels compared to Rhizostomae) and body parts, with oral arms consistently exhibiting higher EAA values compared to bell segments (Supplementary File 2). Notably, certain species like Acromitus hardenbergi (0.57-14.8% DW), Rhopilema esculentum (0.29-7.96% DW), and Rhopilema hispidum (0.06-9.8% DW) display particularly high net quantitative EAA profiles. Along with non-EAAs as glycine, proline, hydroxyproline, aspartic acid, glutamic acid and alanine also arginine resulted consistent in jellyfish (Supplementary File 2).
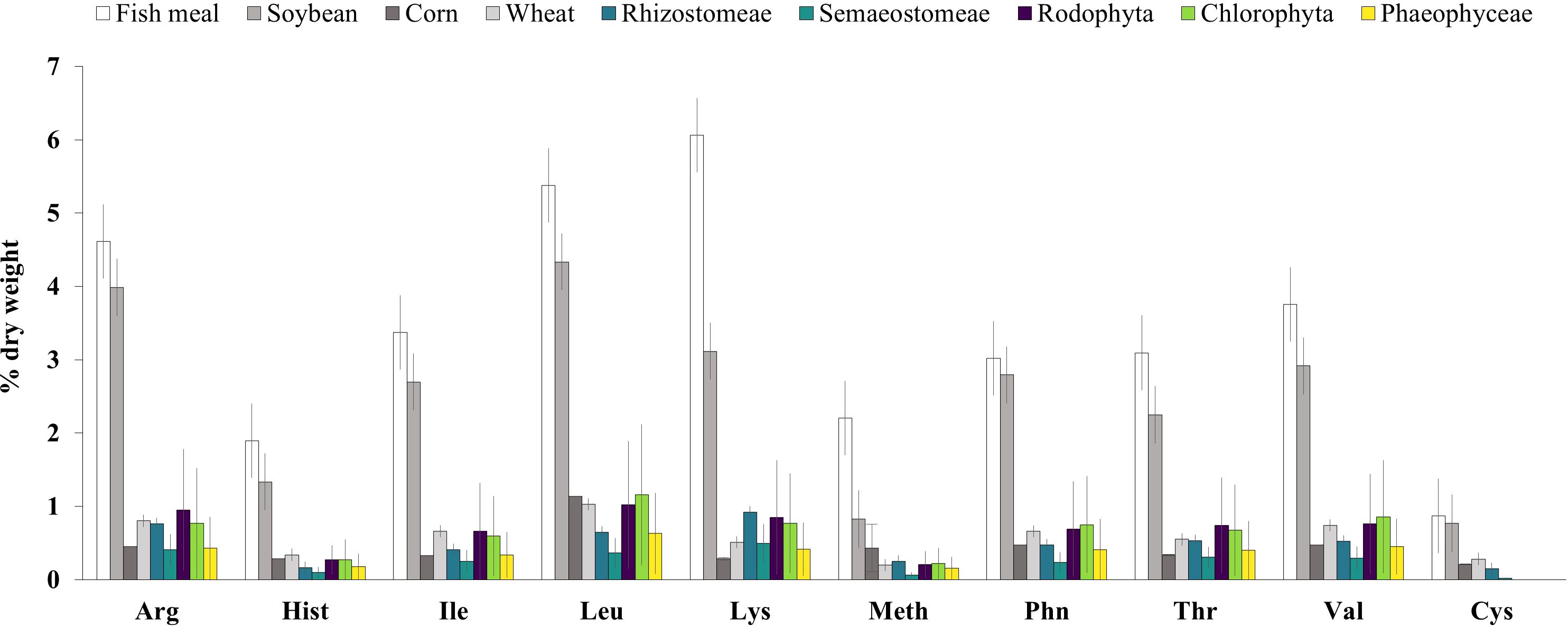
Figure 4. Essential amino acid content (% dry weight) in jellyfish and reference ingredient for aqua-feed production. Note: Value are reported as mean and standard deviation of all the values extracted; Jellyfish (this study), fish meal and plant-based ingredients (NRC, 2011), seaweeds (Angell et al., 2016). Arg, arginine; His, histidine; Ile, isoleucine; Leu, leucine; Lys, lysine; Met, methionine; Phe, phenylalanine; Thr, threonine; Trp, tryptophan; Val, valine, Cys, cysteine.
Amino acid composition, expressed as % of total amino acid (%TAA), is relatively consistent among species (Tables 2, 3). Under this perspective, jellyfish have similar or higher proportions of EAA (42.6- 48.1%TAA in Semaeostomeae and 49.2-54.1%TAA in Rhizostomeae) compared to fishmeal (43.4%TAA), soybean meal (46.0%TAA) or seaweed (45.7%TAA) (Angell et al., 2016). Moreover, the proportion of limiting amino acids in jellyfish demonstrates comparability to commonly used and alternative protein sources such as fish meal, soybean meal, and seaweed. Methionine levels in jellyfish (1.5- 1.9% TAA) fall within the range observed for seaweed (1.25% TAA) and fish meal (2.8% TAA) and the proportion of lysine in jellyfish (6.4-12.2% TAA) also appears to be higher or comparable to the one observed in soybean meal (6.66% TAA), seaweed (5.88% TAA), and fish meal (7.4% TAA) (Angell et al., 2016).
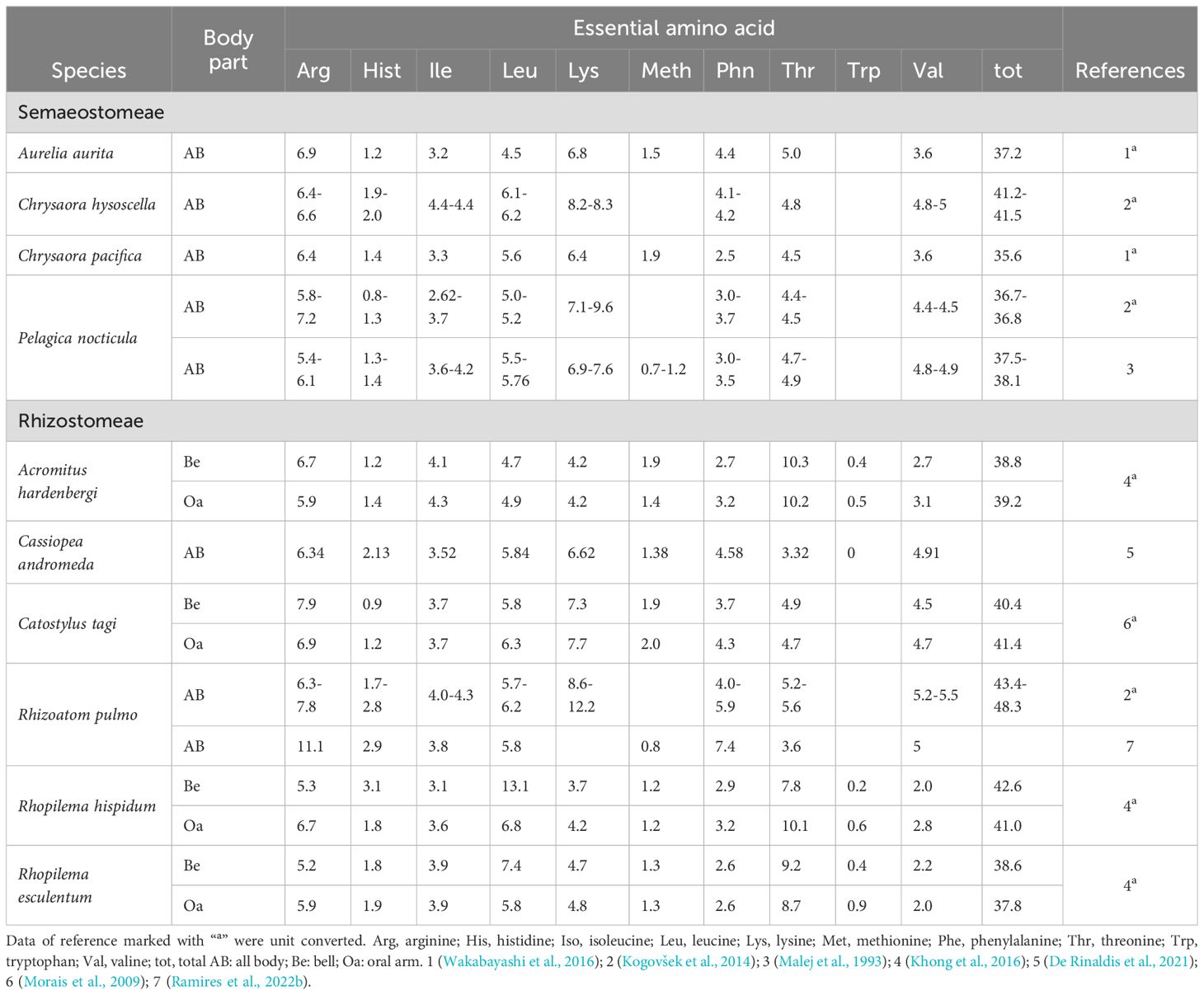
Table 2. Total essential amino acid composition (% of total amino acid) in jellyfish reported in this study.
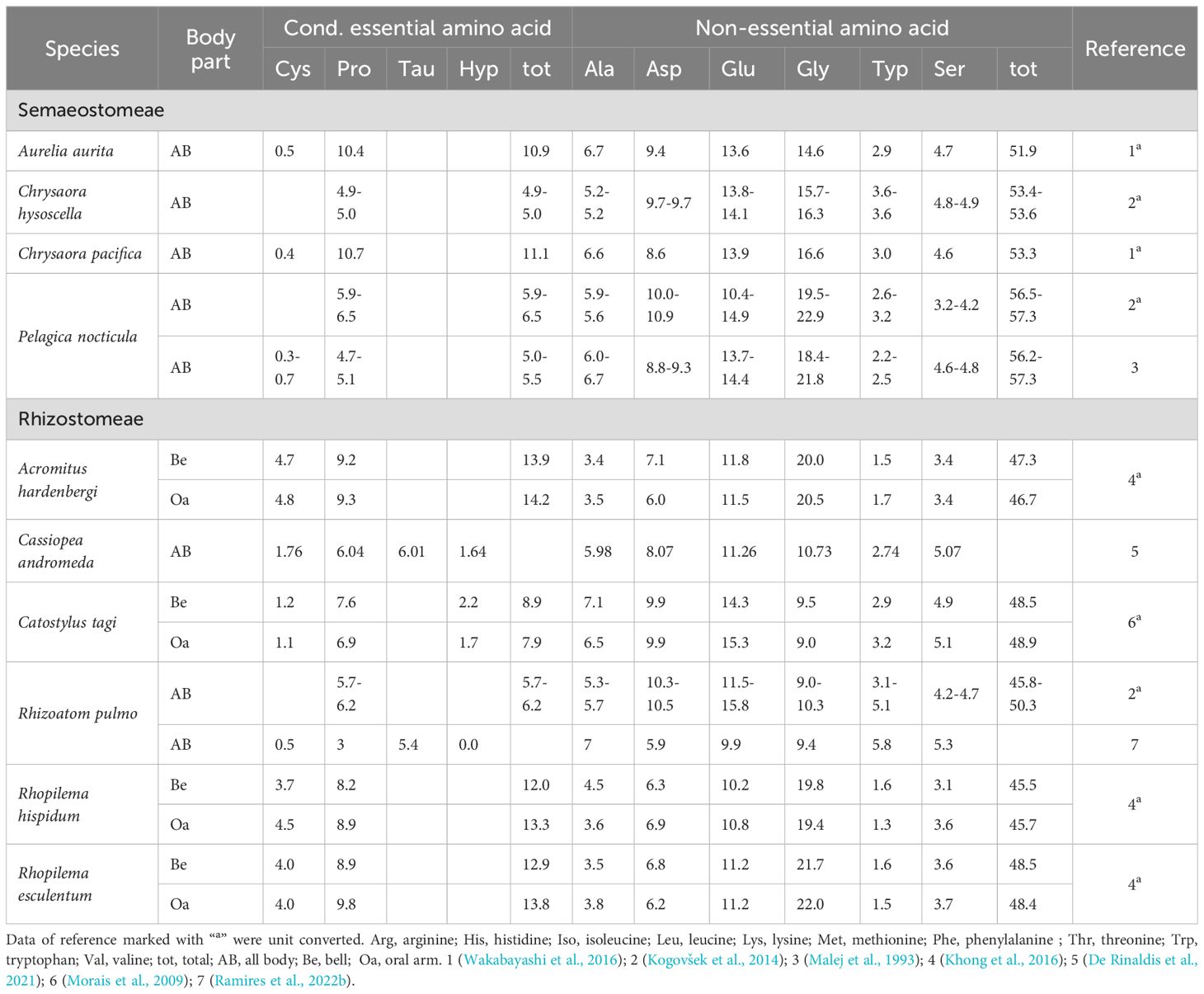
Table 3. Total conditional and non-essential amino acid composition (% of total amino acid) in jellyfish reported in this study.
The overall lipid content (% DW) of jellyfish is low, varying between 0.2 and 5.8, and comparable to some plant-based ingredients (Figure 3). Nevertheless, some species such as Cotylorhiza tuberculata, (12.3% DW) Cyanea nozakii, (8.1% DW) and Stygiomedusa gigantea (10.2% DW) display lipid values similar or even higher than those found in fish meal (9.58 ± 1.16) (Figure 3).
In terms of fatty acid composition, saturated fatty acids (SFA) and polyunsaturated fatty acids (PUFA), particularly arachidonic acid (ARA) (2.8-23.7), eicosapentaenoic acid (EPA) (1.23-25.9), and docosahexaenoic acid (DHA) (0.8-25.9), were more abundant than monounsaturated fatty acids (MUFA) (Supplementary File 2). However, high variability was observed among species, with some species like Aurelia aurita and Pelagia noctiluca, presenting consistently low levels of these compounds (Supplementary File 2).
Despite the similarities between jellyfish and fish oil fatty acid profiles, the ω3/ω6 ratio is higher in fish oil (12.4- 24,1% DW) (NRC, 2011) due to the greater amount of ARA in jellyfish (Figure 5).
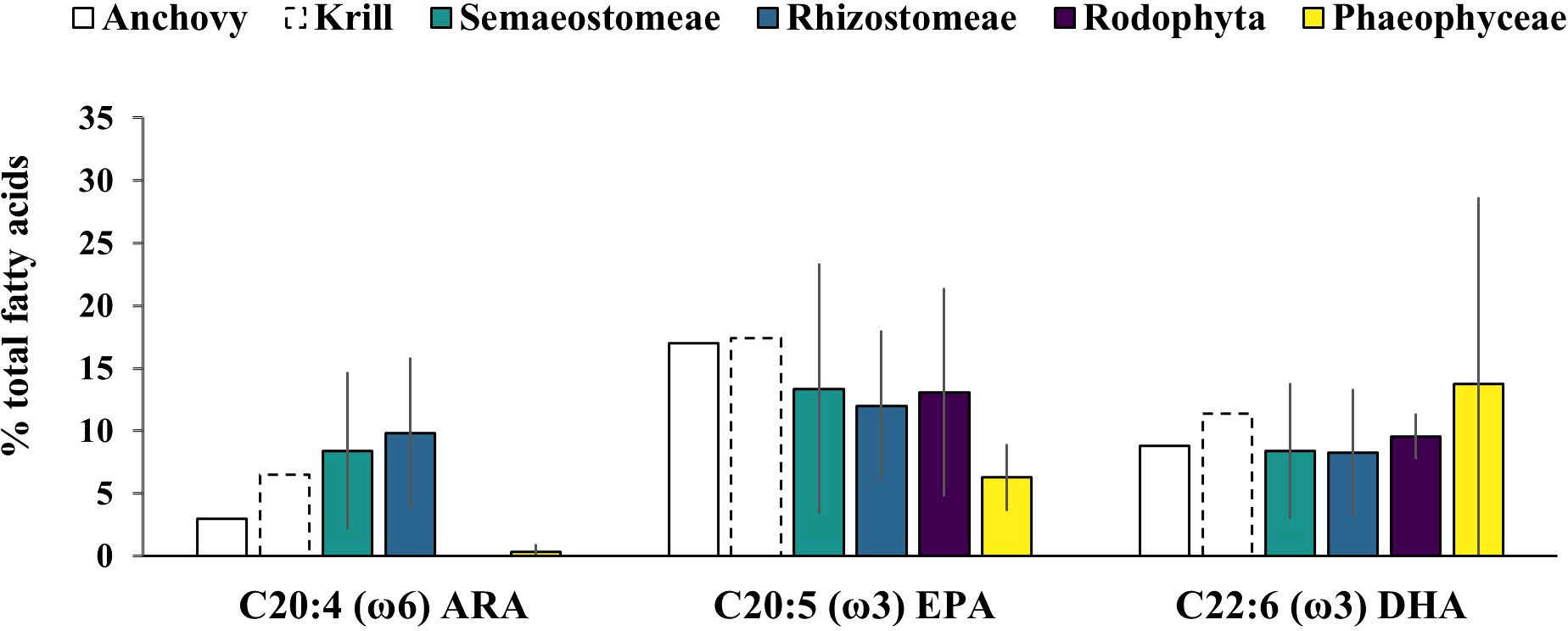
Figure 5. Most representative PUFA (% of total fatty acids) in jellyfish and reference ingredient for aqua-feed production. Note: Value are reported as mean and standard deviation of all the values extracted; Value are reported as mean and standard deviation of all the values extracted; Jellyfish (this study), anchovy and krill (NRC, 2011), seaweed (Rocha et al., 2021).
Macroelements, including sodium (Na) (37.4-80.79 g/kg DW), potassium (K) (1.26 – 2.29 g/kg DW), calcium (Ca) (1.33 -2.36 g/kg DW), magnesium (Mg) (4.27 – 6.92 g/kg DW), phosphorus (P) (0.046 – 59.55 g/kg DW), and chlorine (Cl) (326-587.6 g/kg DW), are abundant in jellyfish, with variations observed among species and body parts and lower concentrations reported for Pellagia noctiluca (Supplementary File 2). Rizhostoma species generally exhibit higher concentrations of K (5.57-126.70 g/kg DW) compared to Sematostoma species (1.6-19.66 g/kg DW). Compared to fish meal, jellyfish exhibit lower Ca content, but comparable levels to seaweed (Supplementary File 1). Regarding P, its content in jellyfish is variable, but generally comparable to other ingredients used in aquafeeds (Supplementary File 1).
Microelements in jellyfish display more variability, with variable concentrations for iron (Fe) (0.59- 252 mg/kg DW), copper (Cu) (0.11 – 49.82 mg/kg DW), zinc (Zn) (3.61 – 400 mg/kg DW), manganese (Mn) (0.11 – 18.66 mg/kg DW) and selenium (Se) (0.31-5.49 mg/kg DW) content (Supplementary File 2). No data on iodine (I) were encountered. Zn concentrations, especially in species associated with symbiotic dinoflagellates, namely - Cotylorhiza tuberculata (Furla et al., 2011; Enrique-Navarro et al., 2022) and Cassiopea sp (Templeman and Kingsford, 2010; Templeman et al., 2021), - show potential for fish nutrition.
Collagen is a prominent protein constituent of jellyfish that in some species accounts for about 50% of its total protein content (Merquiol et al., 2019). Given the high content of glycine, proline, and hydroxyproline in collagen, it was not surprising to find these amino acids abundant in jellyfish (Supplementary File 2). Proline and hydroxyproline are conditionally EAA whose dietary intake might become necessary under specific conditions like rapid growth, illness, or deficiencies in other nutrients (Li and Wu, 2018) and consequently represent an attractive characteristic of jellyfish for aquatic animal nutrition. Glycine, as one of the major non-EAA in jellyfish may play a role in gut health and immune function in fish (Hoseini et al., 2022; Aidee et al., 2023). In terms of amino acid composition, jellyfish contain high proportions of specific amino acids comparable to or higher than those found in commonly used protein sources. The consistent proportion of lysine and arginine (Table 2) unravel appealing characteristics accounting that their requirements for fish and crustaceans are among the highest in the comparison with other EAA (Xing et al., 2024). Therefore, limitations associated with plant-based feedstuffs, particularly regarding AA balance deficiencies (Cai et al., 2022), could be effectively supplemented by incorporating conditionally EAA and/or EAA from selected jellyfish species/parts, namely R. esculentum and R. hispidum arms, and A. hardenbergi. However, accounting the net amount of protein, jellyfish emerge as a more suitable feed ingredient to fulfil the protein requirements of freshwater fish, as marine species typically require higher levels of dietary proteins (40–55%) compared to most freshwater fish (25–40%) (Velasco Santamaría and Corredor Santamaría, 2011; Bowyer et al., 2013). Moreover, despite jellyfish possessing promising protein content, processing methods can significantly impact the bioavailability and digestibility of jellyfish protein. Studies on mammals have shown that hydrolyzed collagen (gelatin) can decrease food efficiency and protein bioavailability (Bordin and Naves, 2015). This finding highlights the importance of investigating processing techniques specific for jellyfish to optimize their nutritional value as an aquafeed ingredient. Beyond its nutritional merits, jellyfish collagen, peptides and free amino acids exhibit a plethora of bioactive functionalities to be dealt with further ahead.
ARA represents the jellyfish characteristic fatty acid. Studies suggest that ARA may play a significant role in growth performance, reproduction, survival, and stress resistance in marine organisms (Xu et al., 2010; Torrecillas et al., 2017, 2018; Ding et al., 2018). However, excessive ARA intake can lead to issues such as reduced performance in Litopenaeus vannamei (Araújo et al., 2020) and problems as at metamorphosis in flatfish (NRC, 2011). Jellyfish could serve as a natural source of ARA to enrich diets according to experimental evidence. Takifugu rubripes, tiger puffer, fed with Aurelia sp. showed increased proportions of polar lipids, ω −3 and ω −6 highly unsaturated fatty acids, especially ARA and DHA (Miyajima-Taga et al., 2017a). Similarly, threadsail filefish (Stephanolepis cirrhifer) fed artificial diet supplemented with N. nomurai presented high levels of ARA (Miyajima-Taga et al., 2015, Miyajima-Taga et al., 2017a). Jellyfish could be used to supplement terrestrial plant oils as lipid source, mitigating their commonly reported deficiencies in long chain PUFA (Zhang et al., 2024). Despite the generally low lipid content in jellyfish, certain species with higher lipid levels—such as Cotylorhiza tuberculata, Cyanea nozakii, and Stygiomedusa gigantea—could serve as a notable lipid source with an attractive fatty acid profile. Additionally, reared jellyfish have been reported to exhibit significantly higher lipid content (De Domenico et al., 2025) and elevated levels of SFA and PUFA, alongside lower MUFA levels (Wang et al., 2021b), compared to their wild counterparts. These findings suggest the potential for jellyfish to be utilized as a lipid source, though further research is required to optimize such practices.
Mineral nutrition in fish, though less studied than other nutrients (Lall and Kaushik, 2021), is essential for their growth and health (NRC, 2011). In jellyfish differences in macro element concentrations between body parts suggest influence from osmotic balance and floating capacity (Costa et al., 2019).
Microelements are typically more limited than macro element in compound feeds and dietary supplementation of trace minerals is commonly employed to ensure optimal growth and health of cultured species (Watanabe et al., 1997). Jellyfish could serve as a potential future source of minerals for dietary supplementation. However, attention must be paid to possible pollution by toxic trace elements (Muñoz-Vera et al., 2015, Muñoz-Vera et al., 2016; Templeman et al., 2021), particularly aluminium (Al) previously detected in the jellyfish, Catostylus tagi (Morais et al., 2009). Monitoring of toxic trace element levels in jellyfish is essential to ensure their safety for use in feed and food. Nevertheless, despite the potential anthropogenic impacts, jellyfish generally exhibit low levels of toxic elements, suggesting their suitability for consumption within regulatory limits. For instance, toxic elements (arsenic (As), cadmium (Cd), lead (Pb), mercury (Hg)) concentrations in Rhizostoma pulmo were found below the limit levels for human consumption allowed by Australian, USA, and EU Food Regulations (Basso et al., 2021).
The composition of jellyfish biomass can vary due to several factors beyond body parts and taxonomy. For instance, size and water content (Malej et al., 1993; Lucas, 1994) affect protein content. In Aurelia aurita (Lucas, 1994) and Pelagia noctiluca (Malej et al., 1993), larger organisms show decreased protein content, likely due to egg transfer and organic content loss. Contrastingly, according to Schaub et al., 2023, Aurelia labiata shows increased protein with bell diameter, though larger diameter ranges indicate an opposite trend (Luskow et al., 2022). These variations suggest complex interactions beyond size, such as diet and trophic factors (Schaub et al., 2023). Seasonal shifts in diet and life cycle are the primary sources of nutritional variability. Although, size-based dietary shifts significantly influence lipid and fatty acid composition in Aurelia labiata (Schaub et al., 2023), the absence of size-based influence on lipid and fatty acid composition in Nemopilema nomurai confirmed the need to account for species-specific trophic habits (Wang et al., 2022a). Notable trophic transitions include shifts from grazing to detritus (Fukuda and Naganuma, 2001), from microzooplankton to microplankton and resuspended particles (Javidpour et al., 2016), and from seston to zooplankton diets (Wang et al., 2020). Pelagia noctiluca’s generalist trophic habits (Milisenda et al., 2018) further underscore the plasticity of jellyfish trophic interactions. Lipid composition varies with life stages, especially reproductive tissues. Gonads maintain consistent nutritional content, affecting overall organism composition during reproduction as indicated by energetic measurements (Doyle et al., 2007), direct lipid (Milisenda et al., 2014) and fatty acid content (Milisenda et al., 2018; Stenvers et al., 2020).
Although, gonadal lipid content has a crucial role as reserve during sexual maturation (Stenvers et al., 2020), food sources, nutritional status, and energy costs were further underlined to be the source of variation in FA profiles between wild and farmed Rhopilema esculentum. Specifically, farmed specimens were characterized by high level of SFA and PUFA and the lowest level of MUFA probably mirroring the adequate supply of specific diet in farmed specimens (Wang et al., 2021b). Macroelement composition in jellyfish reflects seawater composition (De Barba et al., 2016) but varies with species, body parts, and ecological factors (Costa et al., 2019). Microelement distribution has been studied in Cotylorhiza tuberculata (Muñoz-Vera et al., 2015) Rhizostoma pulmo (Muñoz-Vera et al., 2016) and Cassiopea sp (Templeman and Kingsford, 2010) suggesting that the variability in concentrations has a close relationship with water quality at the collection site. Zn and As concentration related with animal size in Cotylorhiza tuberculate and species-specific Zn level were mainly associated to symbiotic dinoflagellates living in Cotylorhiza tuberculata (Furla et al., 2011; Enrique-Navarro et al., 2022) and Cassiopea sp (Templeman and Kingsford, 2010; Templeman et al., 2021). Furthermore, for those jellyfish that harbor symbiotic photosynthetic dinoflagellates (zooxanthellate jellyfish), the strength of this association can be influenced by the life stage and geographic location of the jellyfish species (Djeghri et al., 2019). This variation in the symbiotic relationship ultimately affects the overall chemical composition of the jellyfish. Also, methodological factors like different drying methods may affect nutritional assessment (Siddiqui et al., 2024), impacting in the overall chemical composition (Fukushi et al., 2005; Emadodin et al., 2020), including amino acid content (Kogovšek et al., 2014; Yuferova, 2015; Leone et al., 2019).
Research and innovation in aquafeed formulation continue to explore diverse sources of bioactive compounds to optimize feed efficiency, promote animal health, and minimize environmental impact in aquaculture operations. Presently, the most promising sources of functioning feed additives are plant based, yeasts, mushrooms, seaweed, and their derivatives (Van Doan et al., 2019; Agboola et al., 2021; Firmino et al., 2021), besides the traditional sources such as fish discards and processing byproducts (Ozogul et al., 2021), and synthetic and semi-synthetic compounds (Wang and Hui, 2021).
However, multiple studies have demonstrated that extracts, collagens, and hydrolysates derived from various jellyfish species exhibit a range of potentially beneficial properties for aquaculture. Research suggests these products, particularly from species like Catostylus tagi (Morais et al., 2009) and Rhopilema nomadica (Leone et al., 2015, 2019), possess antioxidant activity, potentially helping to reduce oxidative stress and improve the overall health of organisms, and additionally prolonging shelf life of feeds. Extracts from Rhopilema tetrapilema (Esparza-Espinoza et al., 2023) have also shown antimutagenic properties, which could be beneficial for cell health. Other studies on Rhopilema esculentum suggest potential anti-fatigue effects (Ding et al., 2011), which could benefit aquaculture animals by reducing stress and improving their resilience. Additionally, extracts from Stomolophus nomurai demonstrate immunostimulant activity (Sugahara et al., 2006), with potential to boost the immune system of organisms.
Moreover, the presence of free amino acids in jellyfish (Tables 4, 5), including taurine, hydroxyproline, glycine, arginine, glutamic acid, and alanine, holds significant potential for enhancing the nutritional performance of farmed animals by stimulating feeding in various marine and freshwater fish (Kasumyan and Doving, 2003) and in shrimps (Tantikitti, 2014). Taurine has been shown to enhance growth performance and feed efficiency when supplemented in low-fish meal diets (Magalhães et al., 2019; Sampath et al., 2020). Therefore, jellyfish, rich in taurine, a prominent free amino acid in Aurelia sp., Aurelia aurita, and Nemopilema nomurai, present a promising source for this amino acid supplementation in aquafeeds.

Table 4. Total free essential amino acid (% of total amino acid) composition in jellyfish reported in this study.
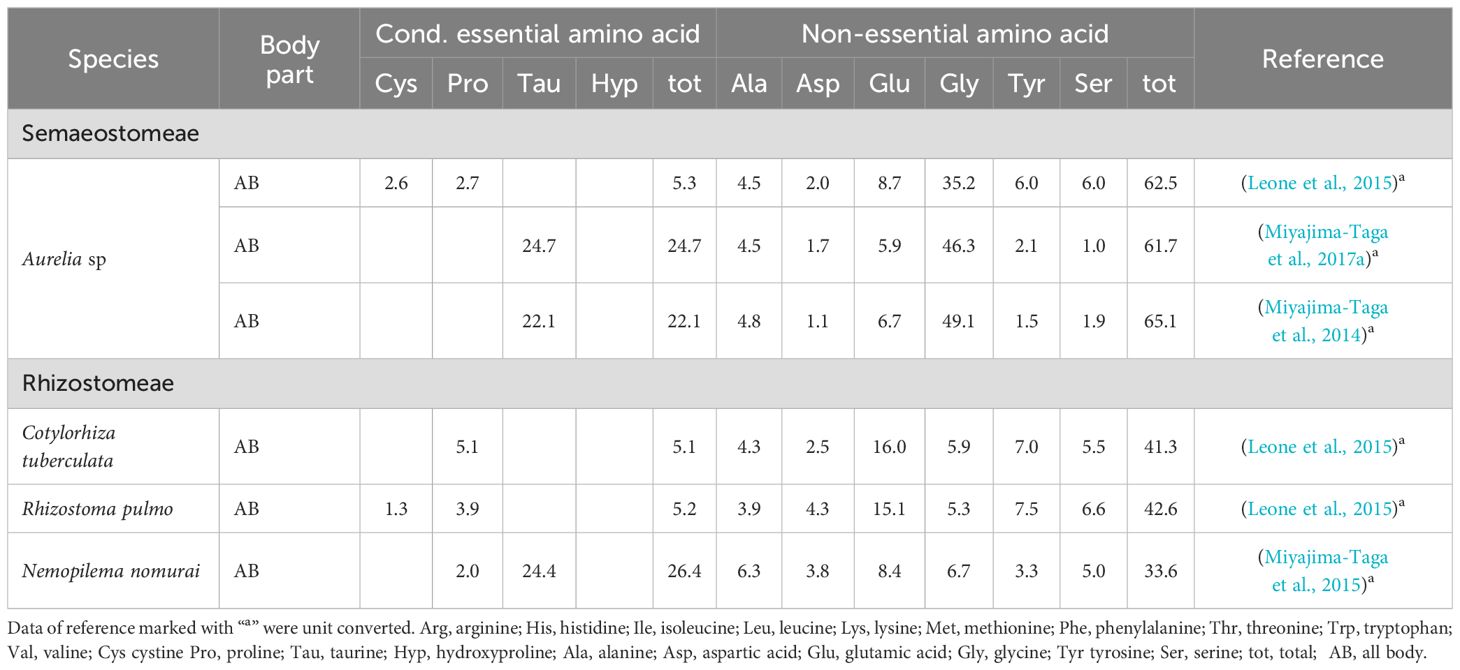
Table 5. Total free conditional and non-essential amino acid (% of total amino acid) composition in jellyfish reported in this study.
Observations of several fish species attracted to jellyfish carcasses as bait (Sweetman et al., 2014; Dunlop et al., 2018) and to jellyfish portions in controlled feeding experiments, particularly with A. aurita (Miyajima et al., 2011b) and N. nomurai (Miyajima-Taga et al., 2015), also suggest that jellyfish possess unique characteristics that could make them valuable attractants for aquaculture feeds. This approach presents a sustainable alternative for enhancing the palatability of current feed trends that incorporate reduced levels of fish meal (Yue et al., 2022) as opposed to traditional fish derived attractants (He et al., 2022).
Fishery by-catches, including jellyfish may cause problems of waste management and disposal, and are costly (Coppola et al., 2021; D’Ambra and Merquiol, 2022). The multi-valorization of marine discarded wastes into high value-added materials approach helps to overcome these major issues, subsequently contributing to the reduction of marine environmental polluting discard accumulation in coastal areas (Govindharaj et al., 2019).
To conduct a meaningful evaluation of a new ingredient for feed purposes, we have not only characterized its nutritional aspects but also assessed the variability in composition, source, and species of origin, which constitutes a crucial element and essential step (Glencross et al., 2007).
According to the biochemical characterization in this review, jellyfish represent a valuable but complex resource for feed applications, offering nutritional variability and bioactive functionalities that can be harnessed to improve aquaculture sustainability and performance. The most relevant jellyfish properties, previously discussed, that are crucial for effective feed formulation and maximizing their potential as underutilized marine resources are as follows:
- Crude protein content (DW) of the oral arms (R. esculentum and R. hispidum) makes them potential feed ingredients for low-trophic level or low-protein-demanding aquaculture species.
- Good proportion of limiting EAA such as methionine and lysine and richness in collagen-derived amino acids, lacking in plant ingredients (Li et al., 2011), may grant their use as feed supplements in feeds. Glycine, proline, and hydroxyproline can improve feed conversion ratios, leading to faster growth and reduced production costs (Li and Wu, 2018) and stimulate gut health and immune function in fish (Hoseini et al., 2022; Aidee et al., 2023).
- Despite low lipid (DW), the presence of PUFA, particularly ARA may serve as a natural source of long-chain PUFA to enrich diets.
- Low levels of insoluble carbohydrates and fiber, as these components negatively affect the fish growth and feed conversion ratio (Nagappan et al., 2021)
- High concentrations of mineral ions Na, K, Cl, Mg (particularly in the Rizhostoma group) and Zn could be used to be usually part of minerals in mixes added in feed formulations.
- Jellyfish species possess unique characteristics that could make them valuable feed attractants that however needs further investigations.
On the other hand, there are still challenges that need to be addressed if jellyfish are to be used as an aquafeed ingredient:
- Though differences in moisture content between jellyfish and other potential feed ingredients like seaweed may not be as substantial as initially perceived (Figure 3), high water content in jellyfish poses logistical challenges for transport and biomass preservation, demanding costly drying methods. In situ valorization of biomass would facilitate logistics associated with storing and transport, eliminating most associated economic and environmental costs (Lopes et al., 2015)
- High ash content in jellyfish presents another potential limitation, affecting energy levels and feed quality.
- Processing techniques originate elevated aluminum levels, posing safety concerns (Bleve et al., 2021). Recent advances, including alum-free treatment methods (Bleve et al., 2021) and innovative processing techniques like thermal processing (Leone et al., 2019) and fermentation (Ramires et al., 2022a), show promise in improving nutritional features and safety standards.
- Lack of consistent composition of an ingredient would affect the nutritional value of the feed and feed efficiency for farmed animals (with implications for growth performance and health), hindering feed formulation and requiring constant adjustments (Sørensen, 2012; Fabà et al., 2018). Therefore, the need for developing standardized processing methods to ensure consistent quality and minimize variability in jellyfish-based feed ingredients.
- Significant knowledge gap exists regarding the digestibility of different jellyfish species and their various body parts by fish.
Despite the promising findings, further research and innovation are necessary to overcome limitations. Moreover, the safe utilization of jellyfish in both feed and food applications requires a thorough risk assessment, as outlined by Bonaccorsi et al. (2020), and strict adherence to established safety parameters, like any new aquafeed ingredient (Bleve et al., 2019).
Finally, assessing the long-term sustainability of utilizing jellyfish as an aquafeed ingredient is crucial. While this study positioned jellyfish as a potential alternative to currently limited feed ingredients, a dedicated sustainability assessment is needed. This analysis should consider factors such as harvesting practices and potential ecological impacts in the case of sustainable fishery exploitation (as recommended by Edelist et al., 2021) or take the shape of a comparative environmental assessment between a valorization process to produce a feed ingredient and different waste management options such as, composting, incineration and landfilling for a waste disposal scenario (Lopes et al., 2015) following jellyfish blooms.
The nutritional variability and bioactive functionalities of jellyfish underscore their potential as a feed supplement for aquaculture feeds rather than a novel major feed ingredient. Conditionally essential amino acids (glycine, proline, glutamic acid and taurine) proportion of EEA (lysine and methionine), the long chain fatty acid ARA and selected minerals such as Na, K, Cl, Mg, and Zn were highlighted in this study as a most promising source of supplementing ingredients.
Continued research and development efforts are needed to elucidate the full potential of jellyfish in feed applications, optimize processing techniques, and evaluate their impact on animal growth, health, and product quality. By associating jellyfish species from specific geographic regions to their applications in situ by the feed industry, this study may additionally contribute to the development and sustainability of jellyfish fisheries.
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
PG: Conceptualization, Data curation, Methodology, Writing – original draft, Writing – review & editing. NN: Writing – review & editing. SG: Methodology, Writing – review & editing. JJ: Writing – review & editing. JC-C: Funding acquisition, Writing – review & editing. CA: Conceptualization, Funding acquisition, Writing – original draft, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This research was funded by the project GoJelly - A gelatinous solution to plastic pollution, funding from the European Union’s Horizon 2020 research and innovation program (grant agreement No. 774499), and by the project ACUICONECTA (1/MAC/1/1.1/0123) from the European Union´s Cooperation Program INTERREG VI-D MAC 2021-2027. Additionally, this study had the support of FCT through the strategic project UIDB/04292/2020 awarded to MARE and through project LA/P/0069/2020 granted to the Associate Laboratory ARNET.
PG was funded by the project GoJelly and is currently under the scholarship (https://doi.org/10.54499/2021.05635.BD). Finally, JC-C is funded by national funds through FCT, under the Scientific Employment Stimulus Institutional Call (CEECINST/00098/2018).
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2025.1539725/full#supplementary-material
Addad S., Exposito J. Y., Faye C., Ricard-Blum S., Lethias C. (2011). Isolation, characterization and biological evaluation of jellyfish collagen for use in biomedical applications. Mar. Drugs 9, 967–983. doi: 10.3390/md9060967
Agboola J. O., Øverland M., Skrede A., Hansen J.Ø. (2021). Yeast as major protein-rich ingredient in aquafeeds: a review of the implications for aquaculture production. Rev. Aquac. 13, 949–970. doi: 10.1111/raq.12507
Aidee K., Fausto S., Acevedo-villavicencio L. N., Flores-zarate A. V., Huang F., Giacoman-martinez A., et al. (2023). Glycine : The smallest anti-inflammatory micronutrient. Int. J. Mol. Sci. 24, 11236. doi: 10.3390/ijms241411236
Angell A. R., Angell S. F., de Nys R., Paul N. A. (2016). Seaweed as a protein source for mono-gastric livestock. Trends Food Sci. Technol. 54, 74–84. doi: 10.1016/j.tifs.2016.05.014
Araújo B. C., Flores-Galvez K., Honji R. M., Barbosa V. M., Viana M. T., Tinajero A., et al. (2020). Arachidonic acid effects on the overall performance, fatty acid profile, hepatopancreas morphology and lipid-relevant genes in Litopenaeus vannamei juveniles. Aquaculture 523, 735207. doi: 10.1016/j.aquaculture.2020.735207
Barzideh Z., Latiff A. A., Gan C. Y., Benjakul S., Karim A. A. (2013). Isolation and characterisation of collagen from the ribbon jellyfish (Chrysaora sp.). Int. J. Food Sci. Technol. 49, 1490–1499. doi: 10.1111/ijfs.12464
Basso L., Papadia P., Rizzo L., Migoni D., Fanizzi F. P., Piraino S. (2021). Trace metals do not accumulate over time in the edible mediterranean jellyfish Rhizostoma pulmo (Cnidaria, Scyphozoa) from urban coastal waters. Water 13, 1410. doi: 10.3390/w13101410
Bleve G., Ramires F. A., De Domenico S., Leone A. (2021). An alum-free jellyfish treatment for food applications. Front. Nutr. 8. doi: 10.3389/fnut.2021.718798
Bleve G., Ramires F. A., Gallo A., Leone A. (2019). ). Identification of safety and quality parameters for preparation of jellyfish based novel food products. Foods Nutr. 8. doi: 10.3389/fnut.2021.718798
Boero F. (2013). “Review of jellyfish blooms in the mediterranean and black sea,” in Studies and reviews, vol. 92. (General Fisheries Commission for the Mediterranean, Rome, FAO), 1–53. doi: 10.1080/17451000.2014.880790
Bonaccorsi G., Garamell G., Cavallo G., Lorini C. (2020). A Systematic review of risk assessment associated with jellyfish consumption as a potential novel food. Foods 9, 1–15. doi: 10.3390/foods9070935
Bordin C. C. D., Naves M. M. V. (2015). Hydrolyzed collagen (gelatin) decreases food efficiency and the bioavailability of high-quality protein in rats. Rev. Nutr. 28, 421–430. doi: 10.1590/1415-52732015000400008
Bosch-belmar M., Milisenda G., Basso L., Doyle T. K., Leone A., Piraino S., et al. (2021). Jellyfish impacts on marine aquaculture and fisheries. Rev. Fish. Sci. Aquac. 0, 1–18. doi: 10.1080/23308249.2020.1806201
Bowyer J. N., Qin J. G., Stone D. A. J. (2013). Protein, lipid and energy requirements of cultured marine fish in cold, temperate and warm water. Rev. Aquac. 5, 10–32. doi: 10.1111/j.1753-5131.2012.01078.x
Brotz L. (2016). Jellyfish fisheries of the world (T). (University of British Columbia). Available online at: https://open.library.ubc.ca/collections/ubctheses/24/items/1.0340647
Brotz L., Schiariti A., López-Martínez J., Álvarez-Tello J., Peggy Hsieh Y. H., Jones R. P., et al. (2017). Jellyfish fisheries in the Americas: origin, state of the art, and perspectives on new fishing grounds. Rev. Fish Biol. Fish. 27, 1–29. doi: 10.1007/s11160-016-9445-y
Cai W., Liu H., Han D., Zhu X., Jin J., Yang Y., et al. (2022). Complete replacement of fishmeal with plant protein ingredients in gibel carp (Carassius auratus gibelio) diets by supplementation with essential amino acids without negative impact on growth performance and muscle growth-related biomarkers. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.759086
Cartwright P., Halgedahl S. L., Hendricks J. R., Jarrard R. D., Marques A. C., Collins A. G., et al. (2007). Exceptionally preserved jellyfishes from the Middle Cambria. PloS One 2, 1–7. doi: 10.1371/journal.pone.0001121
Coppola D., Lauritano C., Esposito F. P., Riccio G., Rizzo C., de Pascale D. (2021). Fish waste: From problem to valuable resource. Mar. Drugs 19, 1–39. doi: 10.3390/MD19020116
Costa R., Capillo G., Albergamo A., Volsi R. L., Bartolomeo G., Bua G., et al. (2019). A multi-screening evaluation of the nutritional and nutraceutical potential of the mediterranean jellyfish Pelagia noctiluca. Mar. Drugs 17, 1–21. doi: 10.3390/md17030172
D’Ambra I., Merquiol L. (2022). Jellyfish from fisheries by-catches as a sustainable source of high-value compounds with biotechnological applications. Mar. Drugs 20, 266. doi: 10.3390/md20040266
De Barba F. F. M., Bazi C. C., Pessatti M. L., Resgalla C. (2016). Macromedusae of southern Brazil: temporal variation, population structure and biochemical composition. Braz. J. Oceanogr. 64, 127–136. doi: 10.1590/S1679-87592016101806402
De Domenico S., Toso A., De Rinaldis G., Mammone M., Fumarola L. M., Piraino S., et al. (2025). Wild or reared? Cassiopea andromeda jellyfish as a potential biofactory. Mar. Drugs 23, 19. doi: 10.3390/md23010019
De Rinaldis G., Leone A., De Domenico S., Bosch-Belmar M., Slizyte R., Milisenda G., et al. (2021). Biochemical characterization of Cassiopea andromeda, another Red Sea jellyfish in the western Mediterranean Sea. Mar. Drugs 19, 498. doi: 10.3390/md19090498
Ding J. F., Li Y. Y., Xu J. J., Su X. R., Gao X., Yue F. P. (2011). Study on effect of jellyfish collagen hydrolysate on anti-fatigue and anti-oxidation. Food Hydrocoll. 25, 1350–1353. doi: 10.1016/j.foodhyd.2010.12.013
Ding Z., Zhou J., Kong Y., Zhang Y., Cao F., Luo N., et al. (2018). Dietary arachidonic acid promotes growth, improves immunity, and regulates the expression of immune-related signaling molecules in Macrobrachium nipponense (De Haan). Aquaculture 484, 112–119. doi: 10.1016/j.aquaculture.2017.11.010
Djeghri N., Pondaven P., Stibor H., Dawson M. N. (2019). Review of the diversity, traits, and ecology of zooxanthellate jellyfishes. Mar. Biol. 166, 1–19. doi: 10.1007/s00227-019-3581-6
Dong Z., Liu D., Keesing J. K. (2010). Jellyfish blooms in China: dominant species, causes and consequences. Mar. pollut. Bull. 60, 954–963. doi: 10.1016/j.marpolbul.2010.04.022
Doyle T. K., Hays G. C., Harrod C., Houghton D. R. (2014). “Ecological and societal benefits of jellyfish,” in Jellyfish blooms (Springer, Dordrecht). doi: 10.1007/978-94-007-7015-7_5
Doyle T. K., Houghton J. D. R., McDevitt R., Davenport J., Hays G. C. (2007). The energy density of jellyfish: estimates from bomb-calorimetry and proximate-composition. J. Exp. Mar. Bio. Ecol. 343, 239–252. doi: 10.1016/j.jembe.2006.12.010
Duarte I. M., Marques S. C., Leandro S. M., Calado R. (2022). An overview of jellyfish aquaculture: for food, feed, pharma and fun. Rev. Aquac. 14, 265–287. doi: 10.1111/raq.12597
Dunlop K. M., Jones D. O. B., Sweetman A. K. (2018). Scavenging processes on jellyfish carcasses across a fjord depth gradient. Limnol. Oceanogr. 63, 1146–1155. doi: 10.1002/lno.10760
Edelist D., Angel D. L., Canning-clode J., Gueroun S. K. M., Aberle N., Javidpour J., et al. (2021). Jellyfishing in europe: current status, knowledge gaps, and future directions towards a sustainable practice. Sustainibility 13, 12445. doi: 10.3390/su132212445
Emadodin I., Reinsch T., Ockens R.-R., Taube F. (2020). Assessing the potential of jellyfish as an organic soil amendment to enhance seed germination and seedling establishment in sand dune restoration. Agronomy 10, 863. doi: 10.3390/agronomy10060863
Enrique-Navarro A., Huertas E., Flander-Putrle V., Bartual A., Navarro G., Ruiz J., et al. (2022). Living inside a jellyfish: the symbiosis case study of host-specialized dinoflagellates, “zooxanthellae”, and the Scyphozoan Cotylorhiza tuberculata. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.817312
Eroldoğan O. T., Glencross B., Novoveska L., Gaudêncio S. P., Rinkevich B., Varese G. C., et al. (2023). From the sea to aquafeed: a perspective overview. Rev. Aquac. 15, 1028–1057. doi: 10.1111/raq.12740
Esparza-Espinoza D. M., del-Carmen-Santacruz-Ortega ,. H., Plascencia-Jatomea M., Aubourg S. P., Salazar-Leyva J. A., Rodríguez-Felix F., et al. (2023). Chemical-structural identification of crude gelatin from jellyfish (Stomolophus meleagris) and evaluation of Its potential biological activity. Fishes 8, 1–15. doi: 10.3390/fishes8050246
Fabà L., Solà-Oriol D., Balfagon A., Coma J., Gasa J. (2018). Assessing the effect of ingredients variability on the composition of the final complete feed for swine. Can. J. Anim. Sci. 99, 7–14. doi: 10.1139/cjas-2017-0157
Firmino J. P., Galindo-Villegas J., Reyes-López F. E., Gisbert E. (2021). Phytogenic bioactive compounds shape fish mucosal immunity. Front. Immunol. 12. doi: 10.3389/fimmu.2021.695973
Fukuda Y., Naganuma T. (2001). Potential dietary effects on the fatty acid composition of the common jellyfish Aurelia aurita. Mar. Biol. 138, 1029–1035. doi: 10.1007/s002270000512
Fukushi K., Tsujimoto J. I., Yokota K. (2005). Determination of inorganic constituents in jellyfish. Bunseki Kagaku 54, 175–178. doi: 10.2116/bunsekikagaku.54.175
Furla P., Richier S., Allemand D. (2011). “Physiological adaptation to symbiosis in cnidarians,” in Coral reefs: an ecosystem in transition (Springer Netherlands, Dordrecht), 187–195. doi: 10.1007/978-94-007-0114-4_12
Glencross B. D., Baily J., Berntssen M. H. G., Hardy R., MacKenzie S., Tocher D. R. (2020). Risk assessment of the use of alternative animal and plant raw material resources in aquaculture feeds. Rev. Aquac. 12, 703–758. doi: 10.1111/raq.12347
Glencross B. D., Booth M., Allan G. L. (2007). A feed is only as good as its ingredients - A review of ingredient evaluation strategies for aquaculture feeds. Aquac. Nutr. 13, 17–34. doi: 10.1111/j.1365-2095.2007.00450.x
Govindharaj M., Roopavath U. K., Rath S. N. (2019). Valorization of discarded marine eel fish skin for collagen extraction as a 3D printabl blue biomaterial for tissue engineering. J. Clean. Prod. 230, 412–419. doi: 10.1016/j.jclepro.2019.05.082
Graham W. M., Gelcich S., Robinson K. L., Duarte C. M., Brotz L., Purcell J. E., et al. (2014). Linking human well-being and jellyfish: ecosystem services, impacts, and societal responses. Front. Ecol. Environ. 12, 515–523. doi: 10.1890/130298
Harzing A. (2007). Publish or perish. Available online at: https://harzing.com/resources/publish-or-perish (Accessed October 25, 2020).
He G., Chen X., Zeng Q., Zhu W., Chen Z., Tan B., et al. (2022). Effects of compound feed attractants on growth performance, feed utilization, intestinal histology, protein synthesis, and immune response of white shrimp (Litopenaeus Vannamei). Animals 12, 2550. doi: 10.3390/ani12192550
Hoseini S. M., Moghaddam A. A., Ghelichpour M., Pagheh E., Haghpanah A., Gharavi B., et al. (2022). Dietary glycine supplementation modulates antioxidant and immune responses of beluga, Huso huso, juveniles. Aquac. Rep. 23, 101026. doi: 10.1016/j.aqrep.2022.101026
Hsieh Y. H. P., Leong F. M., Rudloe J. (2001). Jellyfish as food. Hydrobiologia 451, 11–17. doi: 10.1023/A:1011875720415
Hsieh Y. P., Rudloe J. (1994). Potential of utilizing jellyfish as food in Western countrie. Trends Food Sci. Technol. 5, 225–229. doi: 10.1016/0924-2244(94)90253-4
Hua K., Cobcroft J. M., Cole A., Condon K., Jerry D. R., Mangott A., et al. (2019). The future of aquatic protein: implications for protein sources in aquaculture diets. One Earth 1, 316–329. doi: 10.1016/j.oneear.2019.10.018
Hubot N., Giering S. L. C., Lucas C. H. (2022). Similarities between the biochemical composition of jellyfish body and mucus. J. Plankton Res. 00, 1–8. doi: 10.1093/plankt/fbab091
Hussein O. S., Saleh O. I. (2014). Effect of soaking in jellyfish on some parameters of wheat and lentil seedlings. IOSR J. Environ. Sci. Toxicol. Food Technol. 8, 32–39. doi: 10.9790/2402-08423239
Ikeda T. (2014). Synthesis toward a global model of metabolism and chemical composition of medusae and ctenophores. J. Exp. Mar. Bio. Ecol. 456, 50–64. doi: 10.1016/j.jembe.2014.03.006
Javidpour J., Cipriano A. N., Agnes M., Jan M. (2016). Temporal dietary shift in jellyfish revealed by stabl isotope analysis. Mar. Biol. 163, 1–9. doi: 10.1007/s00227-016-2892-0
Kasumyan A. O., Doving K. B. (2003). Taste preferences in fishes. Fish Fish. 4, 289–347. doi: 10.1046/j.1467-2979.2003.00121.x
Khong N. M. H., Yusoff F. M., Jamilah B., Basri M., Maznah I., Chan K. W., et al. (2016). Nutritional composition and total collagen content of three commercially important edible jellyfish. Food Chem. 196, 953–960. doi: 10.1016/j.foodchem.2015.09.094
Kingsford M. J., Pitt K. A., Gillanders B. M., Brotz L., Cheung W. W. L., Kleisner K., et al. (2000). Management of jellyfish fisheries, with special reference to the order Rhizostomeae. Hydrobiologia 103, 1–179. doi: 10.1007/s10750-012-1039-7
Kogovšek T., Tinta T., Klun K., Malej A. (2014). Jellyfish biochemical composition: Importance of standardised sample processing. Mar. Ecol. Prog. Ser. 510, 275–288. doi: 10.3354/meps10959
Lall S. P., Kaushik S. J. (2021). Nutrition and metabolism of minerals in fish. Animals 11, 1–41. doi: 10.3390/ani11092711
Leone A., Lecci R. M., Durante M., Meli F., Piraino S. (2015). The bright side of gelatinous blooms: Nutraceutical value and antioxidant properties of three Mediterranean jellyfish (Scyphozoa). Mar. Drugs 13, 4654–4681. doi: 10.3390/md13084654
Leone A., Lecci R. M., Milisenda G., Piraino S. (2019). Mediterranean jellyfish as novel food: effects of thermal processing on antioxidant, phenolic, and protein contents. Eur. Food Res. Technol. 245, 1611–1627. doi: 10.1007/s00217-019-03248-6
Li X., Rezaei R., Li P., Wu G. (2011). Composition of amino acids in feed ingredients for animal diets. Amino Acids 40, 1159–1168. doi: 10.1007/s00726-010-0740-y
Li P., Wu G. (2018). Roles of dietary glycine, proline, and hydroxyproline in collagen synthesis and animal growth. Amino Acids 50, 29–38. doi: 10.1007/s00726-017-2490-6
Liberati A., Altman D. G., Tetzlaff J., Mulrow C., Gøtzsche P. C., Ioannidis J. P. A., et al. (2009). The PRISMA statement for reporting systematic reviews and meta-analyses of studies that evaluate health care interventions: explanation and elaboration. J. Clin. Epidemiol. 62, 1–34. doi: 10.1016/j.jclinepi.2009.06.006
Liu C., Chen S.Q., Zhuang Z.M., Yan J.P., Liu C.L., Cui H.T. (2015). Potential of utilizing jellyfish as food in culturing Pampus argenteus juveniles. Hydrobiologia 754, 189–200. doi: 10.1007/s10750-014-1869-6
Lopes C., Antelo L. T., Franco-Uría A., Alonso A. A., Pérez-Martín R. (2015). Valorisation of fish by-products against waste management treatments – Comparison of environmental impacts. Waste Manage. 46, 103–112. doi: 10.1016/j.wasman.2015.08.017
Lucas C. H. (1994). Biochemical composition of Aurelia aurita in relation to age and sexual maturity. J. Exp. Mar. Bio. Ecol. 183, 179–192. doi: 10.1016/0022-0981(94)90086-8
Luskow F., Galbraith M. D., Hunt B. P. V., Perry R. I., Boersma M., Pakhomov E. A. (2022). Gelatinous and soft-bodied zooplankton in the Northeast Pacific Ocean: phosphorus content and potential resilience to phosphorus limitation. Hydrobiologia 849, 1543–1557. doi: 10.1007/s10750-021-04758-9
Magalhães R., Martins N., Martins S., Lopes T., Diáz-Rosales P., Pousão-Ferreira P., et al. (2019). Is dietary taurine required for white seabream (Diplodus sargus) juveniles? Aquaculture 502, 296–302. doi: 10.1016/j.aquaculture.2018.12.019
Malej A., Fagnelli J., Pezdi J. (1993). Stabl isotope and biochemical fraction in the marine pelagic food chain: the jellyfish Pelagia noctiluca and net zooplankton. Mar. Biol. 116, 265–270. doi: 10.1007/BF00355475
Masuda R., Yamashita Y., Matsuyama M. (2008). Jack mackerel Trachurus japonicus juveniles use jellyfish for predator avoidance and as a prey collector. Fish. Sci. 74, 276–284. doi: 10.1111/j.1444-2906.2008.01522.x
Merquiol L., Romano G., Ianora A., D’Ambra I. (2019). Biotechnological applications of scyphomedusae. Mar. Drugs 17, 1–26. doi: 10.3390/md17110604
Milisenda G., Rosa S., Fuentes V. L., Boero F., Guglielmo L., Purcell J. E., et al. (2014). Jellyfish as prey: Frequency of predation and selective foraging of Boops boops (Vertebrata, Actinopterygii) on the mauve stinger Pelagia noctiluca (Cnidaria, Scyphozoa). PloS One 9, 1–10. doi: 10.1371/journal.pone.0094600
Milisenda G., Rossi S., Vizzini S., Fuentes V. L., Purcell J. E., Tilves U., et al. (2018). Seasonal variability of diet and trophic level of the gelatinous predator Pelagia noctiluca (Scyphozoa). Sci. Rep. 8, 1–13. doi: 10.1038/s41598-018-30474-x
Miyajima Y., Masuda R., Kurihara A., Kamata R., Yamashita Y., Takeuchi T. (2011a). Juveniles of threadsail filefish, Stephanolepis cirrhifer, can survive and grow by feeding on moon jellyfish Aurelia Aurita. Fish. Sci. 77, 41–48. doi: 10.1007/s12562-010-0305-8
Miyajima Y., Masuda R., Yamashita Y. (2011b). Feeding preference of threadsail filefish Stephanolepis cirrhifer on moon jellyfish and lobworm in the laboratory. Plankt. Benthos Res. 6, 12–17. doi: 10.3800/pbr.6.12
Miyajima-Taga Y., Masuda R., Kurihara A., Komi R., Yamashita Y., Takeuchi T. (2017a). Efficacy of feeding tiger puffer Takifugu rubripes on moon jellyfish with respect to nutritional composition and behavioural traits. Aquac. Nutr. 24, 504–514. doi: 10.1111/anu.12583
Miyajima-Taga Y., Masuda R., Kurihara A., Yamashita Y., Takeuchi T. (2014). Feeding moon jellyfish improves the tilting behavior of hatchery-reared red sea bream juveniles. Nippon Suisan Gakkaishi (Japanese Ed.) 80, 934–945. doi: 10.2331/suisan.80.934
Miyajima-Taga Y., Masuda R., Kurihara A., Yamashita Y., Takeuchi T. (2015). Effect of giant jellyfish Nemopilema nomurai as supplemental feed on threadsail filefish Stephanolepis cirrhifer. Nippon Suisan Gakkaishi (Japanese Ed). 81, 701–714. doi: 10.2331/suisan.81.701
Miyajima-Taga Y., Masuda R., Yamashita Y. (2017b). Feeding capability of black scraper Thamnaconus modestus on giant jellyfish Nemopilema nomurai evaluated through field observations and tank experiments. Environ. Biol. Fishes 100, 1237–1249. doi: 10.1007/s10641-017-0639-5
Moher D., Liberati A., Tetzlaff J., Altman D. G. (2009). Preferred reporting items for systematic reviews and meta-analyses: the PRISMA statement. J. Clin. Epidemiol. 62, 1006–1012. doi: 10.1016/j.jclinepi.2009.06.005
Morais Z. B., Pintao A. M., Costa I. M., Calejo M. T., Bandarra N. M., Abreu P. (2009). Composition and in vitro antioxidant effects of jellyfish Catostylus tagi from sado estuary (SW Portugal). J. Aquat. Food Prod. Technol. 18, 90–107. doi: 10.1080/10498850802581799
Muñoz-Vera A., García G., García-Sánchez A. (2015). Metal bioaccumulation pattern by Cotylorhiza tuberculata (Cnidaria, Scyphozoa) in the Mar Menor coastal lagoon (SE Spain). Environ. Sci. pollut. Res. 22, 19157–19169. doi: 10.1007/s11356-015-5119-x
Muñoz-Vera A., Peñas Castejón J. M., García G. (2016). Patterns of trace element bioaccumulation in jellyfish Rhizostoma pulmo (Cnidaria, Scyphozoa) in a Mediterranean coastal lagoon from SE Spain. Mar. pollut. Bull. 110, 143–154. doi: 10.1016/j.marpolbul.2016.06.069
Nagappan S., Das P., AbdulQuadir M., Thaher M., Khan S., Mahata C., et al. (2021). Potential of microalgae as a sustainable feed ingredient for aquaculture. J. Biotechnol. 341, 1–20. doi: 10.1016/j.jbiotec.2021.09.003
Naylor R. L., Hardy R. W., Buschmann A. H., Bush S. R., Cao L., Klinger D. H., et al. (2021). A 20-year retrospective review of global aquaculture. Nat. 591, 551–563. doi: 10.1038/s41586-021-03308-6
NRC (2011). Nutrient requirements of fish and shrimp. (Washington, DC: The National Academies Press). doi: 10.17226/13039
Omori M., Nakano E. (2001). Jellyfish fisheries in southeast Asia. Hydrobiologia 451, 19–26. doi: 10.1023/A:1011879821323
Ozogul F., Cagalj M., Šimat V., Ozogul Y., Tkaczewska J., Hassoun A., et al. (2021). Recent developments in valorisation of bioactive ingredients in discard/seafood processing by-products. Trends Food Sci. Technol. 116, 559–582. doi: 10.1016/j.tifs.2021.08.007
Purcell J. E. (2005). Climate effects on formation of jellyfish and ctenophore blooms: a review. J. Mar. Biol. Assoc. United Kingdom 85, 461–476. doi: 10.1017/S0025315405011409
Purcell J. E., Malej A., Benović A. (1999). Potential links of jellyfish to eutrophication and fisheries. In Ecosystems at the Land-Sea Margin: Drainage Basin to Coastal Sea (eds Malone T. C., Malej A., Harding L. W., Smodlaka N., Turner R. E.). doi: 10.1029/ce055p0241
Purcell J. E., Uye S. I., Lo W. T. (2007). Anthropogenic causes of jellyfish blooms and their direct consequences for humans: A review. Mar. Ecol. Prog. Ser. 350, 153–174. doi: 10.3354/meps07093
Ramires F. A., Bleve G., De Domenico S., Leone A. (2022a). Combination of solid state and submerged fermentation strategies to produce a new jellyfish-based food. Foods 11, 3974. doi: 10.3390/foods11243974
Ramires F. A., De Domenico S., Migoni D., Fanizzi F. P., Angel D. L., Slizyte R., et al. (2022b). Optimization of a calcium-based treatment method for jellyfish to design food for the future. Foods 11, 2697. doi: 10.3390/foods11172697
Raposo A., Alasqah I., Alfheeaid H. A., Alsharari Z. D., Alturki H. A., Raheem D. (2022). Jellyfish as food: A narrative review. Foods 11, 1–20. doi: 10.3390/foods11182773
Rocha C. P., Pacheco D., Cotas J., Marques J. C., Pereira L., Gonçalves A. M. M. (2021). Seaweeds as valuable sources of essential fatty acids for human nutrition. Int. J. Environ. Res. Public Health 18, 4968. doi: 10.3390/ijerph18094968
Rohatgi A. (2017). WebPlotDigitizer Version 3.12 (Austin, TX, USA). Available online at: http://arohatgi.info/WebPlotDigitizer.
Roux J. P., van der Lingen C. D., Gibbons M. J., Moroff N. E., Shannon L. J., Smith A. D. M., et al. (2013). Jellyfication of marine ecosystems as a likely consequence of overfishing small pelagic fishes: lessons from the Benguela. Bull. Mar. Sci. 89, 249–284. doi: 10.5343/bms.2011.1145
Ruiz-Frau A. (2023). Impacts of jellyfish presence on tourists ‘ holiday destination choices and their willingness to pay for mitigation measures. J. Environ. Plan. Manage. 66, 2107–2125. doi: 10.1080/09640568.2022.2061926
Sampath W. W. H. A., Zhang Y., Liu J., Yang M., Zhang W., Mai K. (2020). Dietary taurine improves muscle growth and texture characteristics in juvenile turbot (Scophthalmus maximus). Aquac. Rep. 17, 0–6. doi: 10.1016/j.aqrep.2020.100305
Schaub J., McLaskey A. K., Forster I., Hunt B. P. V. (2023). Size-based changes in trophic ecology and nutritional quality of moon jellyfish (Aurelia labiata). Ecosphere 14, 1–17. doi: 10.1002/ecs2.4430
Siddiqui S. A., Ucak İ., Jain S., Elsheikh W., Ali Redha A., Kurt A., et al. (2024). Impact of drying on techno-functional and nutritional properties of food proteins and carbohydrates - A comprehensive review. Dry. Technol. 42, 592–611. doi: 10.1080/07373937.2024.2303580
Sørensen M. (2012). A review of the effects of ingredient composition and processing conditions on the physical qualities of extruded high-energy fish feed as measured by prevailing methods. Aquac. Nutr. 18, 233–248. doi: 10.1111/j.1365-2095.2011.00924.x
Stenvers V., Chi X., Javidpour J. (2020). Seasonal variability of the fatty acid composition in Aurelia aurita (Cnidaria: Scyphozoa): implications for gelativore food web studies. J. Plankton Res. 42, 440–452. doi: 10.1093/plankt/fbaa026
Sugahara T., Ueno M., Goto Y., Shiraishi R., Doi M., Akiyama K., et al. (2006). Immunostimulation effect of jellyfish collagen. Biosci. Biotechnol. Biochem. 70, 2131–2137. doi: 10.1271/bbb.60076
Sweetman A. K., Smith C. R., Dale T., Jones D. O. B. (2014). Rapid scavenging of jellyfish carcasses reveals the importance of gelatinous material to deep-sea food webs. Proc. Biol. Sci. 281, 20142210. doi: 10.1098/rspb.2014.2210
Tantikitti C. (2014). Feed palatability and the alternative protein sources in shrimp feed. Songklanakarin J. Sci. Technol. 36, 51–55.
Templeman M. A., Kingsford M. J. (2010). Trace element accumulation in Cassiopea sp. (Scyphozoa) from urban marine environments in Australia. Mar. Environ. Res. 69, 63–72. doi: 10.1016/j.marenvres.2009.08.001
Templeman M. A., McKenzie M. R., Kingsford M. J. (2021). The utility of jellyfish as marine biomonitors. Mar. pollut. Bull. 173, 113056. doi: 10.1016/j.marpolbul.2021.113056
Torrecillas S., Román L., Rivero-Ramírez F., Caballero M. J., Pascual C., Robaina L., et al. (2017). Supplementation of arachidonic acid rich oil in European sea bass juveniles ( Dicentrarchus labrax ) diets: Effects on leucocytes and plasma fatty acid profiles, selected immune parameters and circulating prostaglandins levels. Fish Shellfish Immunol. 64, 437–445. doi: 10.1016/j.fsi.2017.03.041
Torrecillas S., Betancor M. B., Caballero M. J., Rivero F., Robaina L., Izquierdo M., et al. (2018). Supplementation of arachidonic acid rich oil in European sea bass juveniles (Dicentrarchus labrax) diets: effects on growth performance, tissue fatty acid profile and lipid metabolism. Fish Physiol. Biochem. 44, 283–300. doi: 10.1007/s10695-017-0433-5
Upata M., Siriwoharn T., Makkhun S., Yarnpakdee S., Regenstein J. M., Wangtueai S. (2022). Tyrosinase inhibitory and antioxidant activity of enzymatic protein hydrolysate from jellyfish (Lobonema smithii). Foods 11, 615. doi: 10.3390/foods11040615
Van Doan H., Hoseinifar S. H., Esteban M.Á., Dadar M., Thu T. T. N. (2019). “Chapter 2 - mushrooms, seaweed, and their derivatives as functional feed additives for aquaculture: an updated view,” in Studies in natural product chemistry;, vol. 62. (Susan Dennis, Amsterdam: Atta-ur-Rahman, Elsevier), 41–90. doi: 10.1016/B978-0-444-64185-4.00002-2
Velasco Santamaría Y., Corredor Santamaría W. (2011). Nutritional requirements of freshwater ornamental fish: a review. Rev. MVZ Córdoba 16, 2458–2469. doi: 10.21897/rmvz.283
Wakabayashi K., Sato R., Ishii H., Akiba T., Nogata Y., Tanaka Y. (2012). Culture of phyllosomas of Ibacus novemdentatus (Decapoda: Scyllaridae) in a closed recirculating system using jellyfish as food. Aquaculture 330–333, 162–166. doi: 10.1016/j.aquaculture.2011.12.005
Wakabayashi K., Sato H., Yoshie-Stark Y., Ogushi M., Tanaka Y. (2016). Differences in the biochemical compositions of two dietary jellyfish species and their effects on the growth and survival of Ibacus novemdentatus phyllosomas. Aquac. Nutr. 22, 25–33. doi: 10.1111/anu.12228
Wang Y., Gong Y., Zhang J., Tang Y., Shi X., Shi J. (2021b). Intra- and inter-specific variation in edible jellyfish biomarkers and implications for origin traceability and authentication. Front. Mar. Sci. 8, 738689. doi: 10.3389/fmars.2021.755048
Wang Z., Hui C. (2021). Contemporary advancements in the semi-synthesis of bioactive terpenoids and steroids. Org. Biomol. Chem. 19, 3791–3812. doi: 10.1039/d1ob00448d
Wang J., Li C., Yang G., Tao Z., Wang Y., Xian H. (2022a). Dietary preferences and potential ecological impact on the zooplankton community of Nemopilema nomurai based on stabl isotope and fatty acid analyses. J. Oceanol. Limnol. 40, 1085–1096. doi: 10.1007/s00343-021-1024-y
Wang Y., Wang Q., Lian S., Yue Y., Gao Q., Peng S. (2021c). Dietary jellyfish affect digestive enzyme activities and gut microbiota of Pampus argenteus. Comp. Biochem. Physiol. - Part D Genomics Proteomics 40, 100923. doi: 10.1016/j.cbd.2021.100923
Wang Y. B., Wang Q., Lian S. Y., Yue Y. F., Gao Q. X., Peng S. M. (2022b). Proteomic and transcriptomic analysis of the effects of dietary jellyfish on the midgut of Pampus argenteus. Aquaculture 561. doi: 10.1016/j.aquaculture.2022.738689
Wang Q., Zeng J., Wang Y., Zhao J., Ma L., Shi Z., et al. (2021a). Alternations in the liver metabolome, skin and serum antioxidant function of silver pomfret (Pampus Argenteus) is induced by jellyfish feeding. 3 Biotech. 11, 1–10. doi: 10.1007/s13205-021-02702-1
Wang P., Zhang F., Liu M., Sun S., Xian H. (2020). Isotopic evidence for size-based dietary shifts in the jellyfish Cyanea nozakii in the northern East China Sea. J. Plankton Res. 42, 689–701. doi: 10.1093/plankt/fbaa042
Watanabe T., Kiron V., Satoh S. (1997). Trace minerals in fish nutrition. Aquaculture 151, 185–207. doi: 10.1016/S0044-8486(96)01503-7
Xing S., Liang X., Zhang X., Oliva-Teles A., Peres H., Li M., et al. (2024). Essential amino acid requirements of fish and crustaceans, a meta-analysis. Rev. Aquac. 16, 1069–1086. doi: 10.1111/raq.12886
Xu H., Ai Q., Mai K., Xu W., Wang J., Ma H., et al. (2010). Effects of dietary arachidonic acid on growth performance, survival, immune response and tissue fatty acid composition of juvenile Japanese seabass, Lateolabrax japonicus. Aquaculture 307, 75–82. doi: 10.1016/j.aquaculture.2010.07.001
Yue Y., Chen M., Bao X., Yu Y., Shi W., Kumkhong S., et al. (2022). Effects of three feed attractants on the growth performance and meat quality of the largemouth bass (Micropterus salmoides). Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.1029969
Yuferova A. A. (2015). The impact of different drying modes of scyphozoan jellyfish Rhopilema Esculentum and Aurelia Aurita on the protein and carbohydrate components in their composition and the possibility of their use as dried prepared food. J. Food Process Eng. 40. doi: 10.1111/jfpe.12326
Zhang Z., Miar Y., Huyben D., Colombo S. M. (2024). Omega-3 long-chain polyunsaturated fatty acids in Atlantic salmon: functions, requirements, sources, de novo biosynthesis and selective breeding strategies. Rev. Aquac. 16, 1030–1041. doi: 10.1111/raq.12882
Keywords: jellyfish, aquafeed, nutritional composition, bioactive, amino acids, fatty acids, minerals
Citation: Guttuso P, Nogueira N, Gueroun SKM, Javidpour J, Canning-Clode J and Andrade CAP (2025) Is jellyfish a suitable ingredient for aquafeed? A comprehensive review of nutritional potential and limitation. Front. Mar. Sci. 12:1539725. doi: 10.3389/fmars.2025.1539725
Received: 04 December 2024; Accepted: 07 February 2025;
Published: 14 March 2025.
Edited by:
Gladstone Sagada, Victory Farms, KenyaReviewed by:
Laura Braga Ribeiro, Portuguese Institute for Sea and Atmosphere (IPMA), PortugalCopyright © 2025 Guttuso, Nogueira, Gueroun, Javidpour, Canning-Clode and Andrade. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Paolo Guttuso, Z3V0dHVzb3Bhb2xvQGdtYWlsLmNvbQ==
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.