
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci., 12 March 2025
Sec. Marine Conservation and Sustainability
Volume 12 - 2025 | https://doi.org/10.3389/fmars.2025.1516792
 Thibaut de Bettignies1,2*†
Thibaut de Bettignies1,2*† Lucie Vanalderweireldt2,3,4†
Lucie Vanalderweireldt2,3,4† Marie Launay5,6
Marie Launay5,6 Grégoire Moutardier7,8Vanina Pasqualini5Éric Durieux5
Grégoire Moutardier7,8Vanina Pasqualini5Éric Durieux5 Line Le Gall7
Line Le Gall7The Mediterranean Sea is a “sea under siege”, facing numerous anthropogenic pressures leading to the spread of invasive species and the degradation of reefs, particularly the reduction of habitat-forming macroalgae to turf algae or sea urchin barrens. The coastal waters of Corsica, often considered as a “reference state” due to their lower human impact, have not been thoroughly studied with regard to infralittoral reef communities. This study aimed to characterize these communities and identify their biotic and abiotic drivers through an innovative monitoring protocol (VIT-Reef). From 2019 to 2021, we monitored infralittoral reef communities at 59 sites between 15 and 35 m depth within and adjacent to three MPAs, using innovative rapid assessment dive surveys for 74 benthic taxa. Our results highlighted vertical zonation in reef communities, in particular the depth extent of photophilic algae and the emergence of pre-coralligenous communities. Important ecological implications of taxa covariation were discussed, such as the negative relationship between canopy-forming macroalgae and the invasive algae, Caulerpa cylindracea. Notably, geographical contrasts were observed along northern, southern and western MPAs, primarily influenced by algal community changes. Cystoseira (sensu lato) forests were identified as the climax state on western reefs, while northern and southern reefs showed significant cover of Dictyota spp. and crustose corallines respectively, with concerning levels of Caulerpa cylindracea in deeper southern reefs. Our results underscored the structuring role of Cystoseira spp. forests and the effects of light, nutrients and Posidonia oceanica meadows extending into the reefs. This study highlighted the role of algae in structuring reef communities and will contribute to establish a baseline for conservation planning and predicting future changes in these high priority infralittoral communities.
Coastal marine environments provide vital ecosystem functions, resulting in some of the highest ecosystem goods and services on the planet (Costanza et al., 1997; 2017). At the same time, as a land-sea interface with increasing population densities and economic activities, the coastal zone concentrates most of the local anthropogenic pressures that threaten the stability of these ecosystems (Halpern et al., 2015; Hewitt et al., 2016). Coastal urban development, nutrient loading, chemical pollution, increased turbidity, sedimentation, over-exploitation and destructive fishing, along with the spread of invasive species, diseases, and climate change stressors have already led to the degradation of marine habitats and in particular to the decline of habitat-forming species (such as macroalgae, seagrass and corals) (Pandolfi et al., 2003; Airoldi and Beck, 2007; Waycott et al., 2009; de Bettignies et al., 2021). Temperate reefs are particularly affected by these stressors and severe changes in their ecosystem dynamics can lead to abrupt transitions in ecological communities, known as regime shifts (Ling et al., 2015; Wernberg et al., 2016; Filbee-Dexter and Wernberg, 2018). These shifts typically involve changes from macroalgal forests (i.e., kelp and fucoid species) to turf algal seascapes (Filbee-Dexter and Wernberg, 2018) or to urchin barrens (Filbee-Dexter and Scheibling, 2014; Ling et al., 2015). The proliferation of turf algae and sea urchins typically leads to habitat homogenization with the loss of engineering and foundation species, inhibiting the re-establishment of native canopy-forming algae.
All of these effects have a greater impact in the Mediterranean, as the semi-enclosed nature of the basin and the concentration of anthropogenic activities make Mediterranean temperate reefs highly sensitive to local human pressures and climate change (Garrabou et al., 2009; Bulleri et al., 2010; Durrieu de Madron et al., 2011; Coll et al., 2012). This level of threat supports the expression “the Mediterranean is a sea under siege”, which is often quoted in the scientific literature to alert the general public and motivate conservation action (Galil, 2000; Lejeusne et al., 2010; Coll et al., 2012). Several authors have further argued for a “tropicalisation” of the Mediterranean Sea (Bianchi and Morri, 2003; Peleg et al., 2020). This process is the result of two main phenomena: first, the seawater temperature of the entire basin is warming rapidly; second, the widespread introduction of non-indigenous species with tropical affinities (Bianchi and Morri, 2003; Lejeusne et al., 2010; Raitsos et al., 2010; Durrieu de Madron et al., 2011). These “tropical invaders” are either directly introduced by human activities (e.g., aquaculture, aquariology, maritime transport) or originate from Lessepsian immigration (Bianchi and Morri, 2003; Giangrande et al., 2020). They can affect community dynamics by altering patterns of competition between native species or even transform the current ecosystems into new ones (Lejeusne et al., 2010). Among the most threatening invaders is Caulerpa cylindracea Sonder, 1845, a green alga, that has spread across vast areas of temperate Mediterranean reefs (Klein and Verlaque, 2008; Piazzi et al., 2016). It has become one of the dominant species along entire coastlines, leading to large-scale homogenization of temperate reefs and a reduction in the associated diversity (Piazzi et al., 2005; Klein and Verlaque, 2008; Bulleri et al., 2010).
To address such conservation issues in the Mediterranean, monitoring techniques have been developed to assess the ecological status of reefs and detect critical changes beyond their natural variability, such as the Cartography Littoral Method (CARLIT) (Blanfuné et al., 2017), the Reef-Ecosystem-Based Quality Index (reef-EBQI) (Thibaut et al., 2017), and the Coralligenous Bioconstructions Quality Index (CBQI) (Ferrigno et al., 2017). In addition, new technological methods, such as environmental DNA, photogrammetry and underwater hyperspectral imaging (e.g., Díaz-Ferguson and Moyer, 2014; Dumke et al., 2018; Marre et al., 2019) are being developed to complement and overcome some limitations of traditional survey methods, mainly time, logistics and impact (Thomsen et al., 2012). Despite extensive efforts to develop, calibrate and deploy multi-integrated indicators, there is still a lack of large scale monitoring and description of reference conditions for temperate reefs (Hill et al., 2014). Building on this statement, we propose here a complementary and operational approach, derived from the terrestrial method known as Rapid Assessment Survey (RAS; Ward and Larivière, 2004) and based on the benthic community (composition and abundance) for a closed list of indicative species of algae and fauna. This Visual Inventory in limited Time-space for Mediterranean temperate Reefs (VIT-Reef) is an easy-to-use semi-quantitative inventory that can be carried out by two divers in less than 30 minutes to cover 150 m². It has the potential to provide a rapid, standardized and repeatable measure of the benthic infralittoral rock community, providing practical and operational tools for greater spatial replication of reef monitoring, but is not yet linked to reef ecological assessment per se (e.g., CARLIT, reef-EBQI, CBQI). Although Mediterranean benthic infralittoral communities play an important ecological role, they are still poorly represented in the literature.
Here, we applied this innovative reef monitoring method to the Corsican coast, (France, NW Mediterranean), in and around three marine protected areas (MPAs), including the Scandola Nature Reserve, the Bonifacio Straits Nature Reserve, and the Marine Natural Park of Cap Corse and Agriate (Office de l’Environnement de la Corse, 2011). Corsica is the fourth largest island in the Mediterranean Sea and is often considered a marine biodiversity hotspot (Coll et al., 2012), as well as a “reference state” due to the low impact of human activities along the island coastline (Holon et al., 2015; Vanalderweireldt et al., 2022). Down to 30 – 35 m depth, the Corsican rocky substrate is mainly covered by canopy habitats (e.g., Cystoseira sensu lato), where it forms a highly productive marine forest providing critical habitats for shelter and food for higher trophic levels (Hoffmann et al., 1992; Sales and Ballesteros, 2010). Nevertheless, canopy-forming species are threatened by the spread of the green macroalga Caulerpa cylindracea. This species is a major concern due to its severe impact on biodiversity, its reduction of substrate complexity, and its promotion of turf-forming algae (Klein and Verlaque, 2008; Piazzi et al., 2016). To better understand the structure of infralittoral reef communities and the potential impact of C. cylindracea, we aimed to characterize benthic rocky infralittoral communities and specifically assess the spread of Caulerpa cylindracea using an innovative monitoring protocol. To this end, we used multivariate analysis to characterize, for the first time, infralittoral reef communities between 15 and 35 m depth and to identify which species and environmental variables, including an index of anthropogenic pressure, drive variation in benthic community structure. This community analysis provided an unprecedented opportunity to jointly investigate reef community structure and assess the extent of Caulerpa cylindracea invasion.
The Corsican coast forms a unique spatial unit in the Mediterranean, consisting of extensive meadows of the seagrass P. oceanica, mostly spreading on sandy bottoms (Pasqualini et al., 1998; Telesca et al., 2015), and highly productive Cystoseira (sensu lato) forests inhabit rocky reefs down to 30 – 35 m depth (Sales and Ballesteros, 2010; Blanfuné et al., 2016). From 2019 to 2021, subtidal reefs were surveyed along the Corsican coast in and around three MPAs: the Marine Natural Park of Cap Corse and Agriate (MNP CCA) in the north, the Scandola Nature Reserve (SNR) in the west and the Bonifacio Straits Nature Reserve (BSNR) in the south (Figure 1). In total, benthic communities were assessed at 59 sites during three research campaigns “Our planet reviewed” led by the French Muséum National d’Histoire Naturelle (8 – 23 May 2019, 13 – 30 October 2020, 14 – 27 May 2021). All sampling was conducted in late spring or fall, thus avoiding potential, biomass variation (algae), mortality, or recruitment peaks often observed in mid‐summer or mid‐winter. A relative balanced number of sites were surveyed within the three regions divided into shallow sites from 15 – 25 m depth and deeper sites from 25 – 35 m depth (Figure 1, Table 1). These depth ranges are characteristic of reefs dominated by infralittoral photophilic algae, reflecting the influence of light penetration in the water column. Shallower reefs, from 1 to 15 m depth, have been more extensively explored historically and were therefore the least explored during the 2019 – 2021 biodiversity surveys, thus limiting the depth range of the present study.
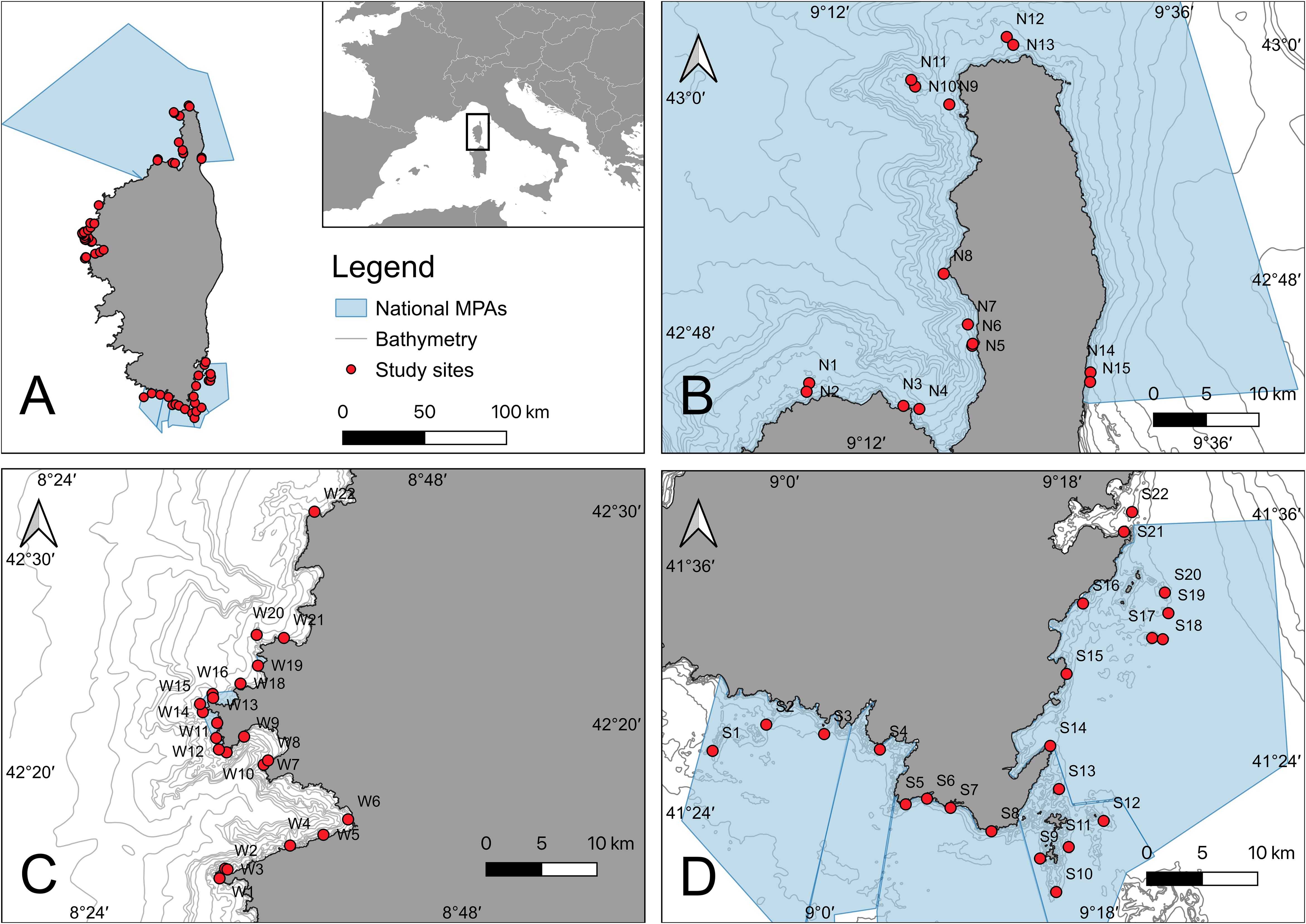
Figure 1. (A) Map of the study areas with the three MPAs investigated around the island of Corsica, (B) Marine Natural Park of Cap Corse and Agriate (MNPCCA) (n=15 sites, N1–N15), (C) the Scandola Nature Reserve (SNR) (n=22 sites, W1–W22), and (D) the Bonifacio Straits Nature Reserve (BNSR) (n=22 sites, S1–S22).
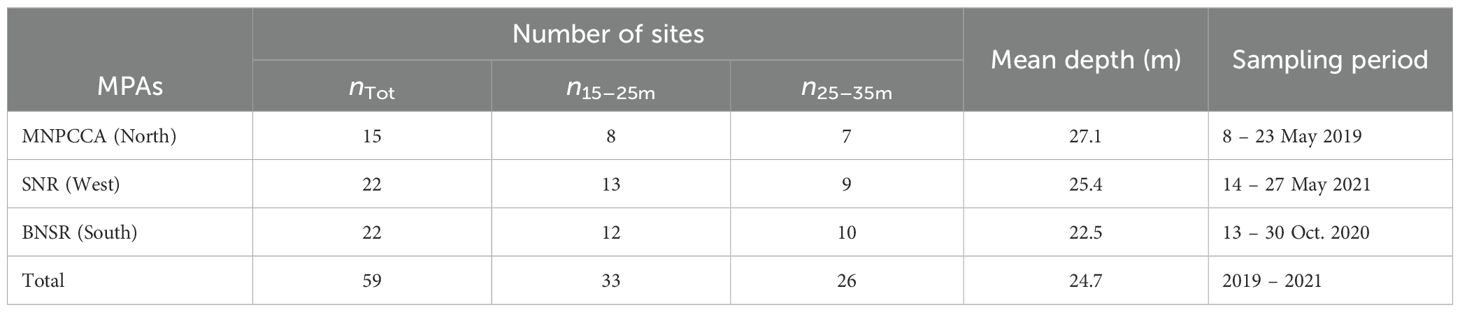
Table 1. Summary of the surveys carried out within the three MPAs: the Marine Natural Park of Cap Corse and Agriate (MNPCCA), the Scandola Nature Reserve (SNR) and the Bonifacio Straits Nature Reserve (BNSR) from May 2019–2021.
The filtering of reef taxa prior to field deployment followed a three-step selection process. First, a preliminary list of known marine benthic species was compiled based on the French National Inventory of Natural Heritage in the French Mediterranean (INPN) database (n~1000 sp.). The list was then cross-checked and reduced based on the characteristic species and determinant species for reef habitats listed in the French classification of Mediterranean benthic habitats for reefs (Michez et al., 2014; La Rivière et al., 2021) and the French ZNIEFF (Natural Zone of Interest for Ecology, Flora and Fauna; Lepareur et al., 2022) inventory species list, respectively (n~300 sp.). Finally, the list was streamlined to the finest taxonomic resolution possible according to species characteristics, expert feedback and field requirements (n=74 taxa). Indeed, taxa were selected on the basis of habitat association and depth range (reef-associated species for the 10 – 40 m depth range), species longevity and phenology (long-lived taxa with minimal seasonal variation), mobility (mostly sessile species), size (generally > 10 cm), ease of in situ identification (during dives), and ecological/structural importance as indicated by the scientific literature and from expert judgment (de Bettignies et al., 2020). The final list contains 74 taxa (61 species levels, 9 genus levels and 4 morphological groups) (Supplementary Table S1 in the Supplementary Material).
The VIT-Reef protocol was developed based on the Rapid Assessment Surveys (RAS; Ward and Larivière, 2004) protocol using a semi-quantitative approach adapted from the SACFOR scale (Mieszkowska et al., 2006; Lévêque et al., 2017), considering standard operating procedures for safety. For each site, a 30 m transect was deployed along a selected (relatively) homogeneous topographic reef and along bathymetric contours to maintain constant depth, avoiding extensive reef walls, overhangs and depth variations greater than 5 m. During transect deployment, the first diver deployed the tape while the second diver videotaped the topographic features 1.5 m above the measuring tape for future estimation of mean reef slope (i.e., Reef index). Once the transect was deployed, the two divers separately initiated a 25 min visual census of all listed taxa (Supplementary Table S1 in the Supplementary Material) along a 150 m2 belt transect (30 m long × 5 m wide). While typical 0.25 m2 photographic quadrats are often preferred for circalittoral coralligenous assemblages (Deter et al., 2012; Çinar et al., 2020), they can be adapted for low complexity infralittoral communities such as turf or encrusting algae-dominated barrens, as in the eastern Mediterranean (Tsirintanis et al., 2018; Nikolaou et al., 2023) but are not suitable for diverse multilayered communities such as those found throughout Corsica. Given the complexity and diversity of the Corsican reefs, the infralittoral communities are then better captured by a large belt transect with a comprehensive list of indicative species to be identified in situ, without the need for extensive image post-processing (reef coverage equivalent to 600 photographic quadrats of 0.25 m2). In addition, the area of the belt transect was chosen to balance the practical constraints of diving (air consumption and safety) with the need for a comprehensive reef coverage. To standardize the transect width (5 m), the length (m) was converted to the average number of fin kicks (per diver) along the transect line during practical dives, to allow for standardized exploration distance away from the transect line (2.5 m from each side). Additional practical taxa identification dives were conducted at the beginning of each research campaign prior to VIT-Reef deployment to ensure rapid, accurate, and consistent taxa identification among divers.
During VIT-Reef deployment, taxa abundance was assessed using a semi-quantitative scale (0 to 4), allowing a rapid in situ assessment of (CL) large countable taxa (> 10 cm), (CS) small countable taxa (< 10 cm), (SM) space-occupying taxa which are considered as meadow/crustose species and (ST) turf/understorey taxa (Mieszkowska et al., 2006, Table 2). Given the relative complexity of reef habitat in these regions, less common sciaphilous habitats (depending on photophilic conditions of transect deployment) such as walls, overhangs, and small fissures were still searched for listed taxa when encountered along the transect. However, abundance values for these taxa were minimal as horizontal and more photophilous conditions were preferred. After each dive, the two divers compared and merged their scores to minimize sampling biases between operators. In total, the abundance values of 49 countable taxa (dominated by faunal species, except for Codium bursa) and the coverage values of 25 space-occupying taxa (dominated by algal species, except for the encrusting sponges Crambe crambe and Phorbas tenacior) were estimated at 59 sites, including 33 sites at 15 – 25 m depth and 26 sites at 25 – 35 m depth (Figure 1; Table 1). Special attention was given to the estimation of the cover of Caulerpa cylindracea, considering the seasonal variation of fronds and ramets and the different sampling times. Priority was then given to the percentage cover of the stolons and rhizoids network (more perennial structure). Prior to analysis, VIT-Reef scores were converted to semi-quantitative abundances as class range means. Two Hellinger transformed matrix species were then constructed for countable taxa (CL–CS, mostly fauna) and space-occupying taxa (SM–ST, mostly algae). Hellinger pretransformation of species data is required to reduce skewness and to avoid the use of double zeros (Borcard et al., 2018).
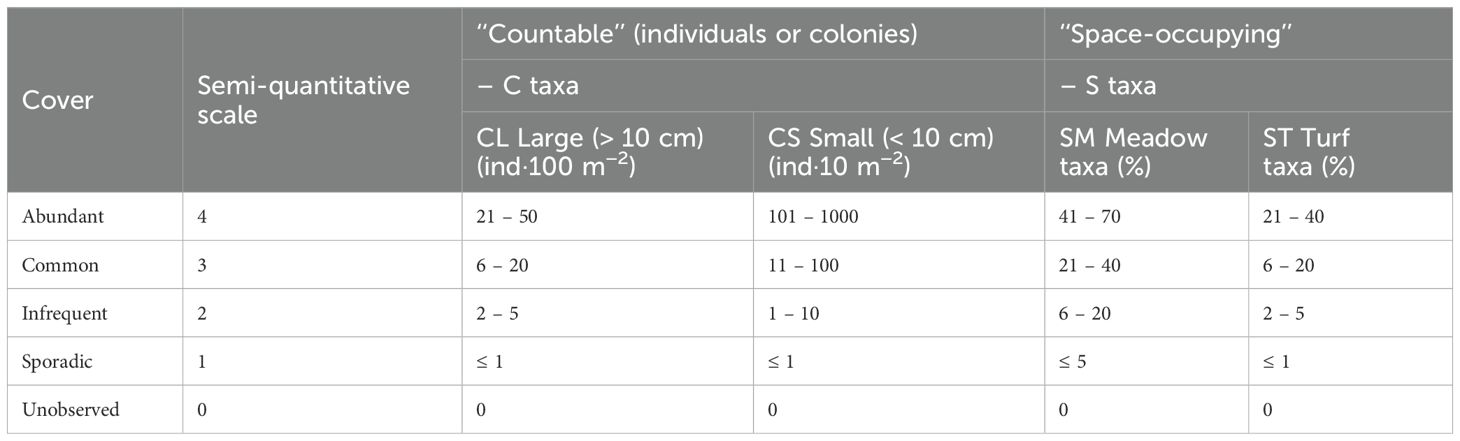
Table 2. VIT-Reef survey and semi-quantitative scale adapted for: “countable” taxa of (CL) large size (> 10 cm) and of (CS) small size (< 10 cm), ‘‘space-occupying’’ taxa considered as (SM) meadow/crustose taxa and as (ST) turf/understorey taxa.
In this study, we collected 17 explanatory variables to identify the main environmental drivers of subtidal reef communities. From field surveys, we estimated the maximum depth of the sampling site (m), the semi-quantitative abundances of Cystoseira (sensu lato) (% of cover) and the nearby Posidonia oceanica reef meadows (% of cover), and the index of reef slope (Reef index; unitless). The Reef index was estimated from the field videos recorded on each VIT-Reef transect. The degree of reef slope was used to classify VIT-Reef transects into four slope categories (1: flat < 10°, 2: gentle slope 10° – 40°, 3: steep slope 40° – 80° and 4: wall > 80°). The reef slope index was then calculated as:
where score represents the slope category i (1 – 4) and cover, the corresponding percentage of transect cover of the four slope categories (%). Reef index ranges from 0.25 to 1.
Twelve physico-chemical variables were extracted from modelling rasters obtained from Bio-Oracle (Tyberghein et al., 2012; Assis et al., 2017) and Ocle (De la Hoz et al., 2018). Eleven physico-chemical variables were collected from the Bio-Oracle satellite data (surface layers only): minimum temperature (Tmin; °C), maximum temperature (Tmax; °C), temperature range (Tr; °C), minimum salinity (Sal; PSU), maximum nitrate concentration (NO3; mol·m−3), phosphate concentration maximum (PO4; mol·m−³), maximum dissolved oxygen concentration (DO; mol·m−³), maximum photosynthetically available radiation at the surface (PAR; Einstein·m−2·day−1), maximum chlorophyll-a concentration (Chl-a; mg·m−³), maximum primary productivity (PP; g·m−3·day−1), and maximum current velocity (Veloc; m−1). These data were compiled from monthly in situ measurements recorded between 2000 and 2014, previously assigned to a grid resolution of 5 arcmin (Tyberghein et al., 2012; Assis et al., 2017). The significant wave height variable (SWH; m) was collected from the Ocle satellite data (De la Hoz et al., 2018). SWH corresponds to the average height of the highest wave measured across Europe during the winter period of 2000–2015. The SWH data were transformed to match the same resolution as the Bio-Oracle variables (5 arcmin and monthly measurements). We supplemented the dataset of explanatory variables with an estimated index of the anthropogenic pressure (Impact; unitless) extracted from the IMPACT model of Andromède (Holon et al., 2015; 2018). The IMPACT index combined global anthropogenic pressures from coastal engineering, anchoring, urban runoff, land cover, coastal population and fisheries and assigned a score in each location (Holon et al., 2015, 2018). The IMPACT model considered both the distance from the sources of disturbance and the bathymetry. Our use of historical Bio-Oracle and Ocle data was driven by their ease of access and consistent spatial coverage, which minimizes the noise of short-term variability and ensures comparability across sites. Given that Mediterranean benthic communities respond primarily to long-term environmental conditions, we assumed that multi-annual climatologies would be well suited to identify the main spatial drivers of community structure.
Prior to further analyses, we performed permutational analyses of variance (PERMANOVA) to test the effect of depth ranges (two levels: 15 – 25 m depth vs. 25 – 35 m depth) on reef communities (Table 1) separately for each Hellinger-transformed species matrix (CL–CS and SM–ST). PERMANOVA analyses were based on Euclidean distances and performed with 9999 permutations. Homogeneity of dispersion was checked before each PERMANOVA according to Anderson (2001). We then performed a centered PCA on the Hellinger-transformed species abundances on the two species matrices (CL–CS; SM–ST) to illustrate the structuring of the species communities according to depth ranges. PCA is an ordination technique that allows to summarize a multivariate dataset (Borcard et al., 2018). It allows to describe the similarities between sites (according to their taxa composition) and how the abundances of the different taxa covary. We then performed a similarity percentages breakdown (SIMPER) procedure to assess the percentage contribution of the most contributing taxa (Clarke, 1993), which was then displayed as a vector superimposed on the 2D PCA plots. PERMANOVA, SIMPER and PCA were performed using the R software v.4.1.2 running with the vegan package (Oksanen et al., 2020). We then divided the species matrices into depth ranges (15 – 25 m and 25 – 35 m depth) to explore the concordance between CL–CS vs. SM–ST (PCIA), and the structure of the subtidal communities for each depth range (PCA).
Two Procrustean co-inertia analyses (PCIA, Dray et al., 2003) were performed to assess the concordance between CL–CS (mostly fauna) and SM–ST (mostly algae) site compositions in the two bathymetric ranges. PCIA is a multivariate technique that measures the similarity between site compositions and identifies which taxa of the two datasets covary. In short, PCIA finds linear combinations of two sets (CL-CS and SM–ST taxa) with maximum covariance. The significance of the overall relationship was assessed using a permutation procedure (PROTEST) with 999 permutations. The results were presented graphically in two bi-plots (15 – 25 m and 25 – 35 m depth sites) where sites are represented by arrows according to their CL–CS and SM–ST composition. The length of a line segment with arrow is therefore inversely proportional to the concordance between CL–CS and SM–ST compositions and can be interpreted as a residual of the covariation model of the PCIA. The PCIA was performed using the R software v.4.1.2 with the ade4 package (Thioulouse et al., 2018).
We performed PCA analysis on the two species matrices (CL–CS; SM–ST), divided into bathymetric subsets (15 – 25 and 25 – 35 m depth locations) to characterize the structuring of the species communities. Centered PCA was performed on Hellinger-transformed species abundances and using the R software v.4.1.2 with the vegan package (Oksanen et al., 2020). Only taxa contributing more than 5% to the first two principal components (PC1–2) were considered significant and included in further analysis.
We used redundancy analysis (RDA) to test for the most important environmental variable (matrix of explanatory variables) that influences the composition of subtidal communities (matrix of response data, CL–CS or SM–ST data). RDA is a canonical ordination technique that combines regression and PCA. In RDA, each of the canonical axis is a linear combination of all explanatory variables – and their statistical significance is tested by permutations (999 permutations). We measured the fit of the RDA models by computing an R2 as the ratio of the explained variation divided by the total variation of CL–CS or SM–ST composition. Then, we calculated the adjusted R2 (R2adj) using the Ezekiel’s formula (Ezekiel, 1930 cited in Borcard et al., 2018) to account for the number of explanatory variables included in RDA models. We first selected explanatory variables to reduce their number using the ordiR2step function from the vegan package, followed by checking for collinearity with Variance Inflation Factors (VIF). This procedure allows the selection of a significant subset of environmental variables that maximizes the R2adj of the global model (Blanchet et al., 2008). RDA were performed using the R software v.4.1.2 with the vegan package (Oksanen et al., 2020).
To identify species associated with Caulerpa cylindracea, we applied two LASSO regression models. First, we performed a binary LASSO model (logistic regression) to identify species predicting the presence/absence of C. cylindracea. The response variable was binary (presence = 1, absence = 0), and all countable (CL–CS) and space-occupying (SM–ST) taxa were included as predictors. Fivefold cross-validation was used to optimize the lambda (λ), and the 1-SE lambda (λ = 0.10) was chosen for model simplicity. A final logistic regression was then refitted using the selected predictors to assess their significance. Model performance was assessed using classification accuracy, AIC, and residual deviance. In addition, a continuous LASSO model was run to examine the species associated with C. cylindracea abundance. Thus, the response variable was continuous, with the same set of predictors as in the binary model. We used fivefold cross-validation, selecting the 1-SE lambda (λ = 4.91) for regularization. A final linear regression was refitted using the retained predictors, with model performance evaluated using AIC, residual deviance, and Mean Squared Error (MSE). LASSO models were run using the R software v.4.1.2 with the dply (Wickham et al., 2023) and the glmnet (Friedman et al., 2010) packages.
One-way PERMANOVAs significantly confirmed the effect of depth on reef communities (CL–CS: F1,58 = 4.755, p < 0.001; SM–ST: F1,58= 3.328, p = 0.002), and PCA biplots highlighted this effect along the x-axis for the countable taxa and along the y-axis for the space-occupying taxa (Figure 2). The proportion of variance accounted for by the first two PCA components was 23.3% and 39.6% for countable taxa (i.e., fauna matrix CL–CS) and space-occupying taxa (i.e., algal matrix SM–ST), respectively. The taxa contributing most to these differences in community structure − specifically, those contributing more than 5% to the first two principal components (PC1–2) − were grey-ball sponges (most likely Ircinia variabilis), Balanophyllia europaea, Codium bursa, Dictyota spp. and Halopteris spp. at 15 – 25 m sites, and Eunicella cavolini, Axinella verrucosa, Caulerpa cylindracea and crustose corallines at 25 – 35 m sites (Figures 2A, B).
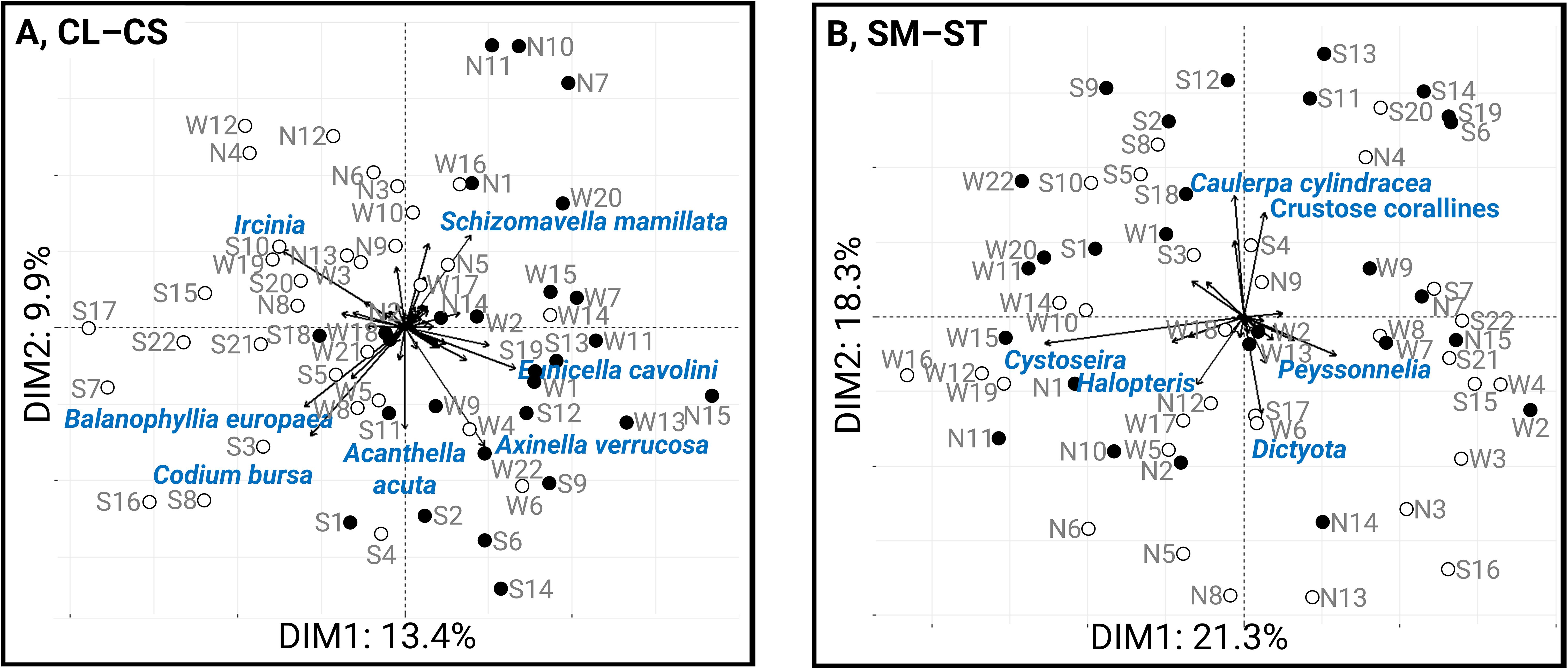
Figure 2. PCA biplots for (A) countable species (mostly fauna) and (B) space-occupying species (mostly algal taxa) performed for all sites (not divided into depth ranges). White circles indicate shallow sites (15 – 25 m) and black circles indicate deep sites (25 – 35 m). The arrows represent species contributing more than 5% of the variance along the first two principal components. The direction and length of each arrow indicate the influence of each species on the ordination space, with longer arrows representing greater influence. Site names (e.g., S1, W3) are shown in the plot.
The PCIA analysis evaluated the concordance between countable taxa (i.e., fauna matrix CL–CS) and space-occupying taxa (i.e., algal matrix SM–ST) (Figure 3) and identified a significant association (PROTEST: 15 – 25 m, m2 = 0.50, p < 0.0001; 25 – 35 m, m2 = 0.46, p < 0.001; Figures 3A, B). Residuals values were highest (> 0.05) for S16, S10, N3, S15, S8 at 15 – 25 m depth, and N14, S13, S1, W11, W15 at 25 – 35 m depth (Figures 3A, B). The PCIA plots also showed a spatial segregation of sites between the northern, western and southern communities.
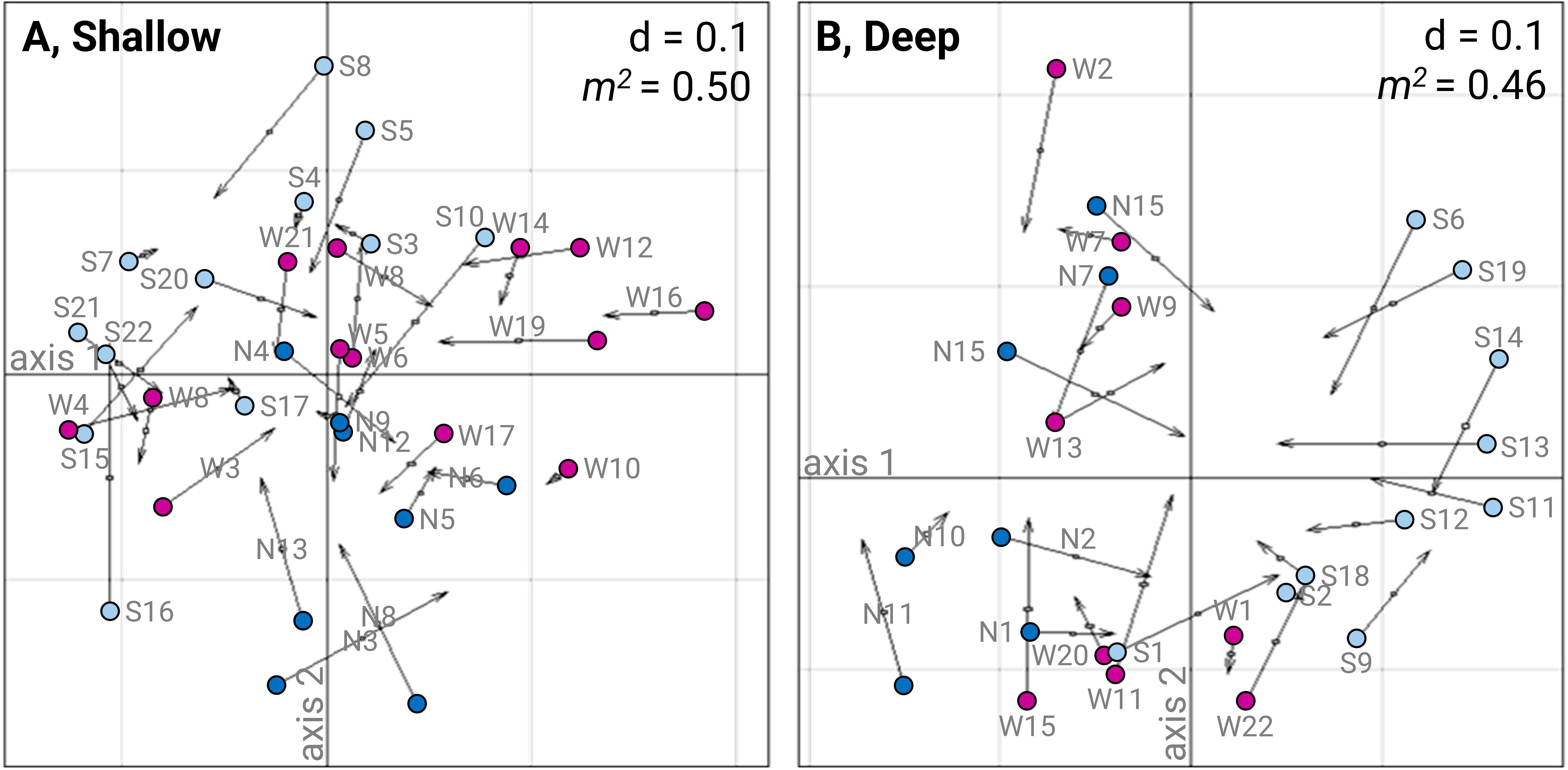
Figure 3. Graphical results of the PCIA analysis performed between the countable species matrix (CL–CS) and the space-occupying species matrix (SM–ST). (A) Sites from 15 – 25 m depth (n=33). (B) Sites from 25 – 35 m depth (n=26). Sites are represented in dark blue diamonds for northern sites (MNPCCA, N1–N15), in magenta triangles for western sites (SNR, W1–W22), and in light blue circles for southern sites (BNSR, S1–S22). Each site is plotted twice: once for the fauna (CL–CS) and once for the algae (SM–ST), connected by arrows. Arrow lengths indicate the degree of concordance between ordinations (shorter arrows represent stronger concordance), while their directions reflect shift between position. The value of d indicates the size of grid squares, and the m12 statistics are given.
When analyzing variation in community structure using PCA, the plots showed a similar segregation of sites between the north, west and south coasts (Figure 4). For countable fauna taxa, the first two components accounted for 24.8% of the variance at 15 – 25 m depth, and 28.0% at 25 – 35 m depth (Figures 4A, B). Below 25 m depth, the taxa that contributed most to variation in community structure were Axinella damicornis, Balanophyllia europaea, Codium bursa, orange-cushion sponges (Dictyonella incisa or Corticium candelabrum), and Petrosia ficiformis (> 5% of contribution, Figure 4A). PC1 (13.9%) was mainly driven by A. damicornis, B. europaea, and C. bursa while PC2 (10.9%) was mainly driven by B. europaea, orange-cushion sponges, and P. ficiformis (> 5% of contribution). In Figure 4A, A. damicornis (upper left panel) was opposed to orange-cushion sponges (lower left panel), B. europaea (upper right panel), and C. bursa coupled with P. ficiformis (lower right panel). Above 25 m depth, the taxa contributing most to variation in community structure were Acanthella acuta, Axinella verrucosa, Codium bursa, Eunicella cavolini, Halocynthia papillosa, Myriapora truncata, Paramuricea clavata, and Schizomavella mamillata (> 5% of contribution, Figure 4B). PC1 (16.7%) was mainly driven by A. verrucosa, A. acuta, and S. mamillata, while PC2 (11.3%) was mainly driven by C. bursa, E. cavolini, H. papillosa, and P. clavata (> 10% of contribution). In Figure 4B, the PCA showed six main groups of taxa: C. bursa (upper left panel), distinguished from A. acuta and A. verrucose (middle left panels), from E. cavolini and P. clavata (lower panel), from S. mamillata (right panel) and from H. papillosa (upper right panel).
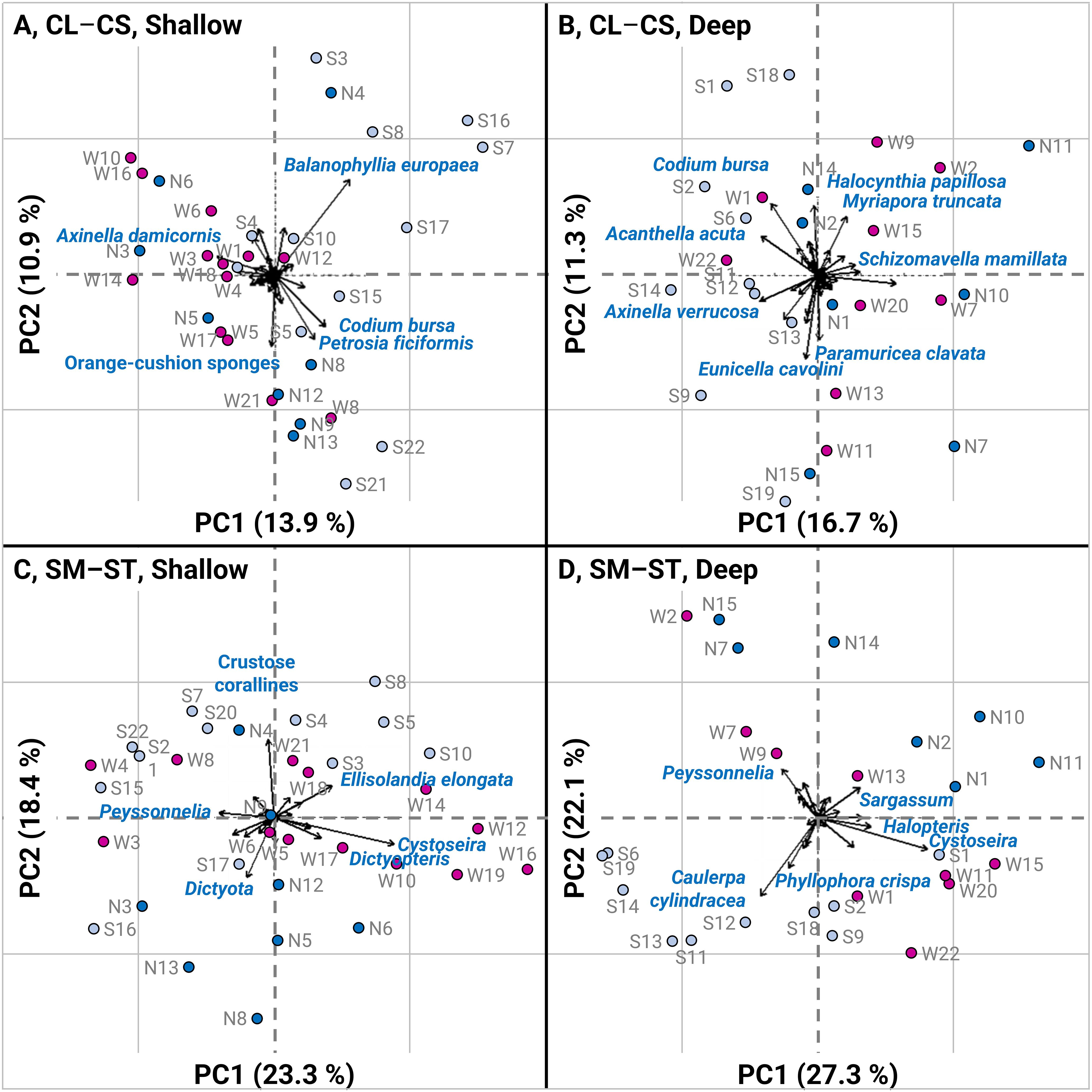
Figure 4. PCA analyses performed on: (A) CL–CS community from shallow sites (15 – 25 m); (B) CL–CS community from deep sites (25 – 35 m); (C) SM–ST community from shallow sites (15 – 25 m); (D) SM–ST community from deep sites (25 – 35 m). Symbols represent the sites surveyed in the northern (MNPCCA, dark blue circles), in the western (SNR, magenta circles) and in the southern (BNSR, light blue circles) coasts of Corsica. The arrows represent the subtidal species, and only taxa contributing more than 5% to PC1–2 are shown on the graphs.
For space-occupying algal taxa, the first two components accounted for 41.7% of the variance at 15 – 25 m depth, and 49.4% at 25 – 35 m depth (Figures 4C, D). Below 25 m depth, the taxa contributing most to variation in community structure were crustose corallines, Cystoseira s.l. spp., Dictyopteris spp., Dictyota spp., Ellisolandia elongata, and Peyssonnelia spp. (> 5% of the contribution, Figure 4C). PC1 (23.3%) was mainly driven by Cystoseira s.l. spp. and Peyssonnelia spp., while PC2 (18.4%) was mainly driven by crustose corallines and Dictyota spp. (> 10% of the contribution). In Figure 4C, Peyssonnelia spp. (left panel) were contrasted with Dictoyta spp. (lower panel), to Cystoseira spp. and Dictyopteris spp. (lower right panel), to E. elongata (upper right panel), and to crustose corallines (upper panel). Above 25 m depth, the taxa contributing most to variation in community structure were Caulerpa cylindracea, Cystoseira s.l. spp., Halopteris spp., Peyssonnelia spp., Phyllophora crispa, and Sargassum spp. (> 5% of the contribution, Figure 4D). PC1 (27.3%) was mainly driven by C. cylindracea, and Cystoseira s.l. spp., while PC2 (22.1%) was mainly driven by C. cylindracea, Peyssonnelia spp., and P. crispa, (> 10% of the contribution). In Figure 4D, Peyssonnelia spp. (upper left panel), differed from C. cylindracea and P. crispa (lower left panel) and from Cystoseira s.l. spp., Halopteris spp. and Sargassum spp. (right panels). The spatial segregation of sites between the northern, western and southern regions was strongly explained by algal communities at both depth ranges, as shown by the PCA plots (Figures 4C, D). To better illustrate this spatial structure and the differences between regions, the percentage cover of the algal taxa contributing the most (Cystoseira s.l. spp., Peyssonnelia spp., crustose corallines, Caulerpa cylindracea, Phyllophora crispa, Dictyota spp.) was plotted on maps (Figures 5A–F).
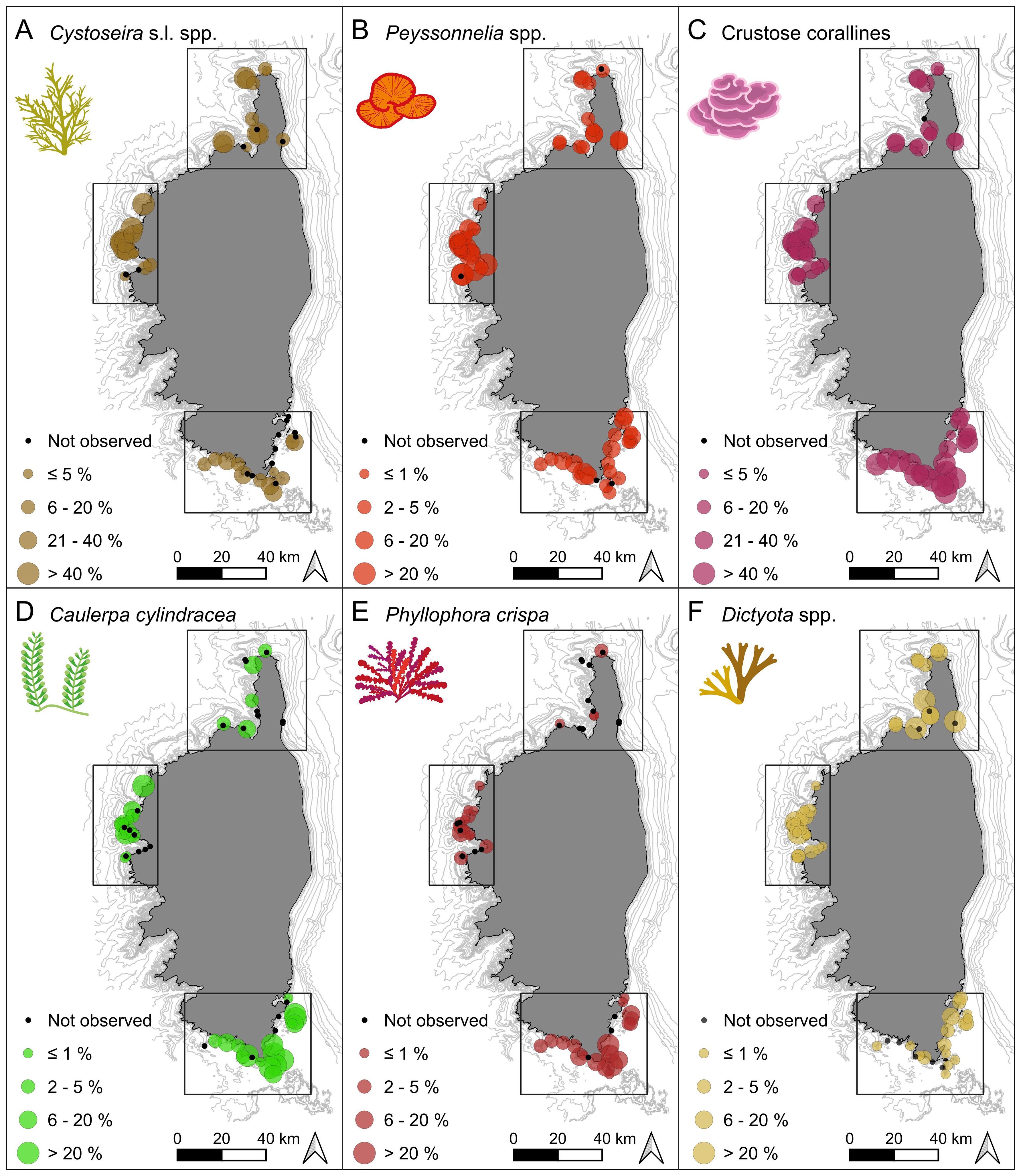
Figure 5. Abundance maps of the algal taxa contributing most to the spatial structure: (A) Cystoseira (sensu lato) spp., (B) Peyssonnelia spp., (C) Crustose corallines, (D) Caulerpa cylindracea, (E) Phyllophora crispa and (F) Dictyota spp.
To understand the main environmental drivers shaping the community composition of CL–CS and SM–ST, results from RDA and LASSO models were examined.
Redundancy analysis (RDA) performed on the CL–CS matrix of 49 countable taxa (mostly fauna species, except for Codium bursa) indicated that maximum depth, Posidonia oceanica reef meadow cover, salinity, maximum photosynthetically available radiation at the surface (PAR), significant wave height (SWH), Chl-a concentration, and anthropogenic index impact were the most important predictors of the community structure (Figure 6A). The adjusted R2 of the RDA model, which refers to the amount of variance explained, reached 19.0% with RDA axes 1–4 accounting for 25.0% of the explained variation. The global test of the RDA model and its associated first four canonical axes were significant (RDA model: F8,59 = 2.910, p = 0.001; RDA1–4: p = 0.001; 0.001; 0.001; 0.010). In Figure 6A, RDA1 was mainly driven by depth (left panel) and reef meadow cover of P. oceanica (right panel, proportion explaining 9.9%). On the other hand, salinity, Cystoseira s.l. spp., Chl-a (lower panel), and meadow cover (upper panel) were the most important variables related to RDA2 (proportion explained of 6.9%) (Figure 6A).
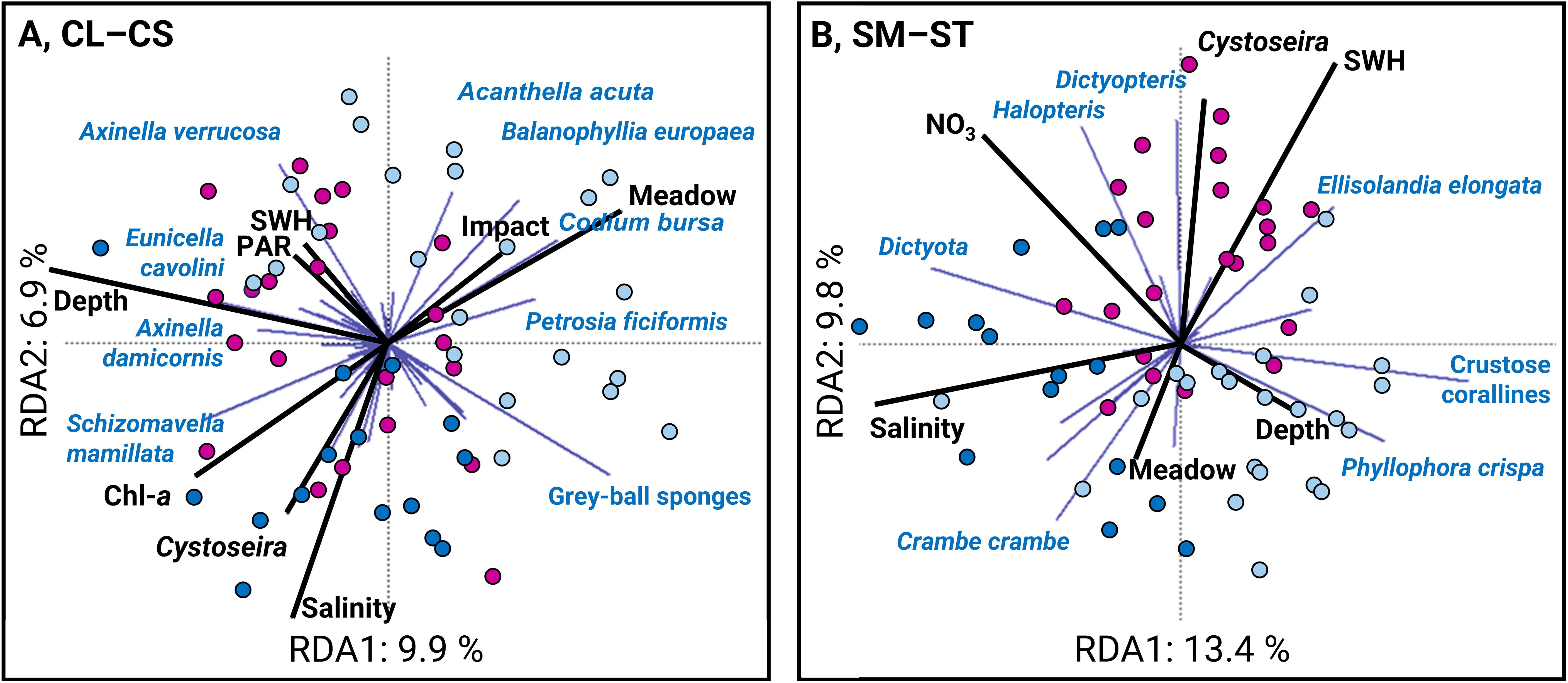
Figure 6. Projections of fauna and algal communities and environmental variables in redundancy analysis (RDA). Community matrices (A) CL–CS and (B) SM–ST are Hellinger-transformed and multidimensional scaling is based on Mahalanobis distance. The points represent the 59 sites surveyed in the North (MNPCCA, n=15 dark blue points), in the West (SNR, n=22 magenta points) and in the South (BNSR, n=22 light blue points) of Corsica. Depth, the maximum depth; Meadow, Posidonia oceanica meadow cover; Cystoseira, the cover of Cystoseira spp.; PAR, the maximum photosynthetically available radiation at the surface; SWH, the significant wave height; Impact, the anthropogenic impact index.
The RDA performed on the SM–ST matrix, composed of 25 space-occupying taxa (mostly algae, except for the encrusting sponges Crambe crambe and Phorbas tenacior), indicated that the maximum depth, the cover of Posidonia oceanica (reef) meadows (% of cover), the salinity, the NO3 concentration and the SWH (significant wave height), were the most important drivers of the SM–ST community structure (Figure 6B). The adjusted R2 of the RDA model, referring to the amount of variance explained, reached 31.0%. RDA axes 1–4 accounted for 35.0% of the explained variation. The global test of the RDA model and its associated first four canonical axes were significant (RDA model: F6,59 = 5.297, p < 0.001; RDA1–4: p = 0.001; 0.001; 0.001; 0.001). On the one hand, the first axis was mainly driven by the salinity and NO3 (left panel), in contrast to SWH and depth (right panel; proportion explained 13.4%) (Figure 6B). On the other hand, the second axis was mainly driven by P. oceanica meadows (lower panel), in contrast to NO3 concentration and SWH (upper panel; proportion explained 9.8%) (Figure 6B).
In the binary LASSO model, ten species were identified as the strongest predictors of Caulerpa cylindracea presence, including species from countable taxa (CL–CS): Anemonia viridis (coefficient: 0.14), Axinella damicornis (coef.: 0.08), Chondrosia reniformis (coef.: 0.09), Cribrinopsis crassa (coef.: 0.11), Paracentrotus lividus (coef.: 0.05), Smittina cervicornis (coef.: – 0.26), and from space-occupying taxa (SM–ST): Cutleria multifida (coef.: – 0.01), Halimeda tuna (coef.: – 0.23), Phyllophora crispa (coef.: 0.28), Sphaerococcus coronopifolius (coef.: – 0.10). When these predictors were included in the final logistic regression model, none remained statistically significant (all p > 0.05), although P. crispa had the lowest p-value (0.08). Model performance was relatively strong, with a classification accuracy of 91.5%, AIC = 43.31, and a residual deviance of 21.31. In addition, the second LASSO continuous regression model retained only one species, P. crispa, as a predictor of C. cylindracea abundance (coefficient: 1.01). The refitted linear model confirmed a significant positive relationship (p < 0.001), with P. crispa having a coefficient of 5.96 ± 1.18 (SE). The overall model fit resulted in a residual deviance of 4607.4, an AIC of 430.55 and a Mean Squared Error (MSE) of 78.09.
The VIT-Reef protocol has provided ecological insights around Corsica, which are discussed in detail below. The protocol could be adapted to monitor reefs in the western Mediterranean, but the closed list of indicative taxa may need to be relevant to other Mediterranean regions. After training in transect deployment and taxa identification, VIT-Reef has the potential to be implemented by different actors (citizen science programs, MPA managers, NGOs, environmental engineering consultants, or research units) for infralittoral reef conservation issues, given its limited logistical and technical constraints. Indeed, such a belt transect approach can be implemented by two divers in less than 30 minutes to cover 150 m², but site level transect replication may still be required depending on the objectives. In addition, the semi-quantitative classes could be revised to score countable and space-occupying taxa based on percent cover only, in order to simplify in situ taxa scoring and the subsequent statistical analysis. However, we believe that this approach is better suited for multilayered infralittoral reef communities and allows for the detection of more taxa than classical quadrats, without the need for extensive image post-processing in the case of photography.
Preliminary analyses of the two depth ranges (15 – 25 m vs. 25 – 35 m) provided insights into the vertical zonation patterns for both algal and fauna taxa, in particular the depth extent of infralittoral communities dominated by photophilic algae and the emergence of pre-coralligenous communities, indicating a shift towards circalittoral communities. Such a pronounced vertical zonation has already been demonstrated for the Mediterranean Sea, despite its reduced tidal range (Ballesteros, 1992), and has rather been explained by the interactive effects of light, hydrodynamics and nutrients (Zabala and Ballesteros, 1989; Ballesteros, 1993), but the literature on this topic remained scarce. For the Corsican reefs studied, the most structuring species driving this vertical zonation (and hence differences between sites) were species or groups of species well known for their light affinities. In summary, shallow sites (15 – 25 m) were characterized by a high cover of turf-forming algal species (Halopteris spp. and Dictyota spp.) and a high number of long-lived and photophilic species such as the green balloon-like macroalga Codium bursa (life span > 15 years; Vidondo and Duarte, 1998), a solitary Mediterranean coral Balanophyllia europaea (life span > 20 years; Goffredo et al., 2004) and grey-ball sponges (Ircinia variabilis most likely). Cystoseira (sensu lato) forest did not explain such depth zonation (given its taxonomic resolution in this study), as it was found across the entire depth range. On the other hand, deep sites (25 – 35 m) attested the intrusion of typical coralligenous species (Ballesteros, 2006) into the infralittoral communities, with Eunicella cavolini being the most abundant gorgonian species, Axinella verrucosa a circalittoral-like erect sponge and an increasing cover of encrusting corallines. While the vertical zonation pattern was evident here, the transition to coralligenous communities appeared gradual as evidenced by a mix of infralittoral and circalittoral species in the 25 – 35 m depth range. This transitional area can thus be referred as pre-coralligenous communities. In the western Mediterranean, the 25 – 35 m depth range is considered shallow for coralligenous reefs, where they begin to develop mostly on vertical walls and overhangs and are referred to as “cliff morphology” (~ 20 – 50 m) (Bracchi et al., 2017; Piazzi et al., 2022). Further down (usually below 50 m), the “platform morphology” of coralligenous reefs develops horizontally (Ballesteros, 2006; Piazzi et al., 2022). Further analyses were then replicated at each depth range to avoid the confounding effect of depth ranges when discussing spatial structure, given the transition in reef communities.
Interestingly, most deep sites (25 – 35 m) were heavily colonized by Caulerpa cylindracea, a non-indigenous species endemic to southwestern Australia with temperate warm to pan-tropical affinities (Verlaque et al., 2003) and reported as the most serious invader in the Mediterranean (Piazzi et al., 2005; Montefalcone et al., 2015). Since its first report in Libya in 1990 (Nizamuddin, 1991) and according to its spread kinetics (Verlaque et al., 2000; Piazzi et al., 2005), C. cylindracea has probably affected all the Mediterranean coasts on all types of substrates and communities from 0 to at least 50 m depth, indicating a wide ecological niche (Verlaque et al., 2000; Piazzi et al., 2005). While algal turf has been proposed as a facilitating factor for the establishment of C. cylindracea on Mediterranean reefs (Piazzi et al., 2003; Bulleri and Benedetti-Cecchi, 2008), here the opposite pattern was observed with the spread of C. cylindracea and its co-occurrence with encrusting corallines in deep sites (25 – 35 m), opposite to algal turf depths.
We hypothesized that the facilitation provided by algal turf was overcome by biotic inhibitors to the establishment and/or spread of C. cylindracea in the 15 – 25 m depth range (infralittoral). In particular, the presence of a dense Cystoseira (sensu lato) canopy has been proposed as the main limiting factor for C. cylindracea invasion (Bulleri et al., 2010, 2011), but the exact mechanisms remain unresolved (Piazzi et al., 2016). Extensive C. cylindracea mats were repeatedly observed in the coralligenous habitat (between 40 and 65 m depth), often covering more than 50% (Le Gall L., pers. com.). This suggested that coralligenous communities may be more vulnerable to C. cylindracea invasion than infralittoral communities, which are protected by a dense and more resistant macroalgal canopy. Moreover, the circalittoral habitat may serve as an important source of vegetative dispersal for C. cylindracea to nearby infralittoral communities. If a shift from macroalgal canopy to algal turf occurs (as observed in disturbed Mediterranean reefs), the recipient habitat may become more susceptible to successful establishment of C. cylindracea. In this context, C. cylindracea can be considered as a “passenger” rather than a “driver” of habitat degradation (Bulleri et al., 2010). However, once established, it has the potential to become a “driver” of change, preventing the recovery of native macroalgal canopy species (once disturbed) by facilitating turf growth, which in turn increases sediment deposition (Bulleri et al., 2010).
Covariations (in the same or opposite direction) among structuring taxa provided relevant ecological information for each depth range. At shallow depths (15 – 25 m), Codium bursa and Petrosia ficormis co-varied and were predominantly found in lighted conditions. Both species are known to reach large sizes only under sufficient light exposure. Indeed, the photosynthetic capacity of C. bursa decreases with increasing size (Geertz-Hansen et al., 1994) and the trophic source from the cyanobacterial symbiotic association (symbiocortex) of P. ficiformis contributes to the increase in size with light conditions (Bavestrello et al., 1994). At deeper depths (25 – 35 m), the two gorgonian species Eunicella cavolini and Paramuricea clavata covaried, as expected, as we usually observe them in sympatric populations (Ponti et al., 2014; Valisano et al., 2019). Both species are characteristic of coralligenous communities in the northwestern Mediterranean where they increase habitat complexity (Harmelin and Marinopoulos, 1994; Linares et al., 2008).
Most surprisingly, the alien green alga Caulerpa cylindracea and the fleshy red alga Phyllophora crispa were both found in high abundance at deeper sites. The regression analysis further supported a strong association between these two species, with P. crispa emerging as the only significant predictor of C. cylindracea abundance in the continuous model. The refitted linear regression confirmed this positive relationship, suggesting that P. crispa may play a facilitating role in the establishment of C. cylindracea. Dense mats of P. crispa may create structural conditions that facilitate the colonization and persistence of C. cylindracea, as has been observed in other species (Bonifazi et al., 2017). The architectural complexity of P. crispa mats (Bonifazi et al., 2017) increases microhabitat heterogeneity, potentially providing more anchoring points for C. cylindracea and promoting sediment accumulation. This, in turn, could facilitate stolon elongation and the lateral spread of C. cylindracea (Bulleri et al., 2010; Piazzi et al., 2016). While our binary LASSO model initially identified additional potential species associations, none remained statistically significant after refitting the logistic regression, underscoring the particularly strong association between P. crispa and C. cylindracea abundance rather than mere presence. Across both depth ranges (15 – 25 m vs. 25 – 35 m), Cystoseira (sensu lato) forests showed opposite trends compared to the shrubs of the encrusting red algae Peyssonnelia spp. The later thrive only in low light conditions, typically found on overhangs, vertical walls, crevices and caves in the infralittoral zone; and is one of the most representative genera of coralligenous algae in the circalittoral (Barberá et al., 2012). On the contrary, extensive Cystoseira (sensu lato) forest are usually found in high light conditions on low-sloping reefs. Such changes in community structure were indicative of differences in both light conditions and/or reef substrate slope for all sites, regardless of depth range.
Overall, photophilic and sciaphilic species drove variation in spatial structure at shallow and deep depths ranges (15 – 25 m vs. 25 – 35 m) respectively, which the algal community being a key explanatory factor. Co-inertia analyses showed a strong correlation between the countable fauna and space-occupying algae matrices (m2 > 0.46). These results strongly suggested a bottom-up control of benthic primary producers towards higher trophic levels, probably not through trophic dependence, as most of fauna taxa listed in this study are sessile filter-feeders, but rather through competition for space and/or habitat provisioning. Further down in the circalittoral zone, the competition release with algal forests allows the development of complex and well-structured faunal communities (mainly composed of sponges, gorgonians, corals, and bryozoans), also called “animal forests”, which will then play a key role as ecosystem engineers (Rossi, 2013).
Given the different sampling times, a potential confounding effect of time (or season) may have been introduced into our study, weakening further comparisons of spatial patterns between regions. However, we believe that our results are not significantly biased by time. In fact, the VIT-Reef protocol minimizes such temporal bias by prioritizing long-lived and sessile taxa with minimal seasonal variation, avoiding extreme seasons (i.e., mortality or recruitment peaks observed in mid-summer or mid-winter), paying special attention to estimating Caulerpa cylindracea cover from stolon/rhizoid rather than frond/ramet, and buffering moderate fluctuations in taxa abundance and cover thanks to the coarse categorical approach of the semi-quantitative method.
Reef communities showed geographical contrasts along the northern, southern and western Corsican MPA regions, but these patterns varied with both depth ranges and communities studied (fauna vs. algae). The northern and western MPA regions showed similar fauna communities across both depth ranges but contrasted with the southern region. In particular, most southern sites showed higher abundance of specific structuring species, i.e., the scleractinian Balanophyllia europaea in shallow sites and the two sponge species Acanthella acuta and Axinella verrucosa in deeper sites. In southern shallow sites, the shift from erect to crustose coralline algae may in turn have favored the photophilic solitary scleractinian, Balanophyllia europaea (Goffredo et al., 2004), probably by releasing competition for space and/or improving substrate under light exposure. In addition, extensive development of Posidonia oceanica on southern coastal reefs (over 20% coverage in 60% of sites) resulted in limited areas (mostly rocky outcrops of large boulders and overhangs tops) for large erect algae, dominated instead by coralline, probably due to higher disturbance regime. The combination of these biotic and abiotic conditions appeared to be particularly favorable for the colonization of B. europaea, as evidenced by its high numbers in these reef areas. On the contrary, many southern deep sites displayed significant structural difference compared to northern and western sites, due to the high abundance of the two sciaphilic sponges, A. acuta, A. verrucosa (Santín et al., 2019). Low light conditions may then be more prevalent for southern deep reef communities, given the complexity of deep reefs with numerous small walls, overhangs, crevices and caves (mean reef slope index > 0.75), which physically reduce incident light on reefs and favor sciaphilic species. Indeed, such abiotic (depth, as a proxy for incident light reduction) and biotic (cover of Cystoseira s. l. and Posidonia oceanica on reefs) factors best explained the regional partitioning of fauna communities. Within this depth range (15 – 35 m) in the Mediterranean, while the downstream effect of Cystoseira (sensu lato) canopy on surrounding communities (recruitment and development) is well established (Benedetti-Cecchi et al., 2001; Mangialajo et al., 2008), the effect of seagrass meadows thriving on reefs has rarely been documented. It is interesting to observe the opposite trend between extensive cover of Posidonia oceanica and Cystoseira (sensu lato) which might indicate a relative competitive exclusion of these two engineer species under certain conditions, particularly in the southern region of Corsica, where extensive cover of P. oceanica on reefs was documented alongside minimal cover of Cystoseira (sensu lato).
Algal communities better explained the geographical contrasts between the three MPA regions, as indicated by the higher proportion of variance explained by the multivariate analyses, especially in the 25 – 35 m depth range. In summary, at shallow depths (15 – 25 m), the western region had the highest cover of Cystoseira (sensu lato) together with Dictyopteris spp. thriving in low slope reefs characterized by abundant light, while Dictyota spp. and crustose corallines thrived in the northern and southern regions, respectively. As expected, Peyssonnelia spp. increased in shallow southern and western reefs with more vertical features (low light conditions). At deeper depths (25 – 35 m), the southern region exhibited a higher abundance of Caulerpa cylindracea and Phyllophora crispa than the northern region, while the western region had more Cystoseira (sensu lato) and/or Peyssonnelia spp. depending of reef slopes. As for the fauna communities, the presence of dense stands of Cystoseira (sensu lato) has been proposed to structure the associated algal communities (Benedetti-Cecchi et al., 2001; Mangialajo et al., 2008), together with higher water movement and nutrient concentration, probably responsible for the extensive development of Cystoseira (sensu lato) forests themselves along the Corsican coast. To a lesser extent, and as mentioned above, the extent of Posidonia oceanica on the reefs is another crucial determinant of the algal communities, restricting reef areas for large erect algae development.
This study of the spatial structure of infralittoral communities on Corsican reefs provided important insights into depth zonation (between 15 and 35 m depth), species covariation, regional variations in community structure, environmental drivers and potential future disturbances mediated by Caulerpa cylindracea. Not surprisingly, algal communities explained most of the variation in spatial structure and Cystoseira (sensu lato) was found to be a driving factor together with light (depth), nutrient and hydrodynamic conditions. More surprisingly, Posidonia oceanica, which also develops on reefs, acted as another influential driver, its extensive coverage confining reef communities to specific areas and thereby structuring the observed fauna and algal communities. Well-developed Cystoseira (sensu lato) forests, which represent the climax and reference state of Mediterranean infralittoral communities, were found on most western reefs (especially in the Scandola Nature Reserve) and on some northern reefs away from the Gulf of Saint-Florent (near the Agriates desert and the Centuri shoals). On the contrary, most of the northern and southern reefs were characterized respectively by a high cover of Dictyota spp. and crustose corallines, with a worrying cover of Caulerpa cylindracea in the southern deeper reefs.
These findings raised concerns, as they did not align with the characteristics of undisturbed infralittoral reefs in the region. Further comparisons with historical data and appropriate calibrations against known pressure gradients will be required to propose ecological assessments of sites from the VIT-Reef protocol. The few reef-EBQI sites in Corsica have been classified as high or good ecological status (Thibaut et al., 2017). The reef-EBQI is based on a broad set of functional compartments from a conceptual ecosystem model and allows the assessment of the quality of the ecosystem functioning, but therefore requires more measurements and extensive diving (i.e., at least 2 days for 2 divers) than VIT-Reef. While the depth range is different between the current deployment of the reef-EBQI (1 – 10 m) and VIT-Reef (15 – 35 m) protocols, it is important to test the VIT-Reef protocol in other reef-EBQI ecological status frameworks (moderate, poor, and bad) (Thibaut et al., 2017). Deployment in known stressed environments will allow disentangling the relationship between reef community variation and pressure, and to relate VIT-Reef to reef-EBQI scores.
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
TB: Conceptualization, Data curation, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. LV: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Supervision, Validation, Visualization, Writing – original draft, Writing – review & editing. ML: Formal Analysis, Investigation, Writing – original draft, Writing – review & editing. GM: Formal Analysis, Investigation, Writing – review & editing. VP: Funding acquisition, Supervision, Writing – review & editing. ÉD: Funding acquisition, Writing – review & editing. LG: Investigation, Resources, Supervision, Visualization, Writing – review & editing.
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This research was carried out within as part of the “Our planet reviewed” expeditions of the French Muséum National d’Histoire Naturelle. This work was partly supported by the research programme of the University of Corsica “BiodivCorse -Explorer la biodiversité de la Corse” (Collectivité de Corse -État) (grant number CPER project (N°40137)).
We thank Dr Aurélien Boyé (Laboratoire d'Ecologie Benthique Côtière, Ifremer) and Dr Stéphane Dray (UMR 5558, CNRS, Université Claude Bernard Lyon 1) for providing their advice on multivariate analysis. We kindly thank Andromède for providing an index of the anthropogenic pressure extracted from their IMPACT model. We are sincerely grateful to the editor, Stanislao Bevilacqua, and two reviewers, Julian Evans and Federico Pinna, for their valuable insights. Their comments have significantly improved our manuscript.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declare that no Generative AI was used in the creation of this manuscript.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2025.1516792/full#supplementary-material
Airoldi L., Beck M. (2007). “Loss, status and trends for coastal marine habitats of Europe,” in Oceanography and Marine Biology: Annual Review, 45 (London, United Kingdom: Taylor & Francis Group), 345–405.
Anderson M. J. (2001). A new method for non-parametric multivariate analysis of variances. Austral Ecol. 26, 32–46. doi: 10.1111/j.1442-9993.2001.01070.pp.x
Assis J., Tyberghein L., Bosh S., Verbruggen H., Serrão E., De Clerck O. (2017). Bio-ORACLE v2.0: extending marine data layers for bioclimatic modelling. Global Ecol. Biogeography 27, 277–284. doi: 10.1111/geb.12693
Ballesteros E. (1992). “Els vegetals i la zonacio litoral: especies, comunitats i factors que influeixen en la seva distribucio,” in Arxius de la Secció de Ciències, vol. 101. (Barcelona, Spain: Institut d’Estudis Catalans), 1–616.
Ballesteros E. (1993). Algues bentoniques i fanerogames marines. Monografies la Societat d’Historia Natural les Balears 2, 503–530.
Ballesteros E. (2006). “Mediterranean coralligenous assemblages: a synthesis of present knowledge,” in Oceanography and Marine Biology: Annual Review, 44 (London, United Kingdom: Taylor & Francis Group), 123–195.
Barberá C., Moranta J., Ordines F., Ramón M., De Mesa A., Díaz-Valdés M., et al. (2012). Biodiversity and habitat mapping of Menorca Channel (western Mediterranean): implications for conservation. Biodiversity Conserv. 21, 701–728. doi: 10.1007/s10531-011-0210-1
Bavestrello G., Pansini M., Sarà M. (1994). “The variability and taxonomic status of different Petrosia-like sponges in the Mediterranean Sea,” in Sponges in time and space: biology, chemistry, paleontology. Eds. van Soest R. W. M., van Kempen T. M. G., Braekman J. C. (Balkema, Rotterdam), 83–92.
Benedetti-Cecchi L., Pannacciulli F., Bulleri F., Moschella P. S., Airoldi L., Relini G., et al. (2001). Predicting the consequences of anthropogenic disturbance: large-scale effects of loss of canopy algae on rocky shores. Mar. Ecol. Prog. Ser. 214, 137–150. doi: 10.3354/meps214137
Bianchi C. N., Morri C. (2003). Global sea warming and “tropicalization” of the Mediterranean Sea: biogeographic and ecological aspects. Biogeographia 24, 319–327. doi: 10.21426/B6110129
Blanchet F. G., Legendre P., Borcard D. (2008). Forward selection of explanatory variables. Ecology 89, 2623–2632. doi: 10.1890/07-0986.1
Blanfuné A., Boudouresque C. F., Verlaque M., Thibaut T. (2016). The fate of Cystoseira crinita, a forest-forming Fucale (Phaeophyceae, Stramenopiles), in France (North Western Mediterranean Sea). Estuarine Coast. Shelf Sci. 181, 196–208. doi: 10.1016/j.ecss.2016.08.049
Blanfuné A., Thibaut T., Boudouresque C. F., Mačić V., Markovic L., Palomba L., et al. (2017). The CARLIT method for the assessment of the ecological quality of European Mediterranean waters: relevance, robustness and possible improvements. Ecol. Indic. 72, 249–259. doi: 10.1016/j.ecolind.2016.07.049
Bonifazi A., Ventura D., Gravina M. F., Lasinio G. J., Belluscio A., Ardizzone G. D. (2017). Unusual algal turfs associated with the rhodophyta Phyllophora crispa: benthic assemblages along a depth gradient in the Central Mediterranean Sea. Estuarine Coast. Shelf Sci. 185, 77–93. doi: 10.1016/j.ecss.2016.12.013
Bracchi V. A., Basso D., Marchese F., Corselli C., Savini A. (2017). Coralligenous morphotypes on subhorizontal substrate: a new categorization. Continental Shelf Res. 144, 10–20. doi: 10.1016/j.csr.2017.06.005
Bulleri F., Alestra T., Ceccherelli G., Tamburello L., Pinna S., Sechi N., et al. (2011). Determinants of Caulerpa racemosa distribution in the north-western Mediterranean. Mar. Ecol. Prog. Ser. 431, 55–67. doi: 10.3354/meps09137
Bulleri F., Balata D., Bertocci I., Tamburello L., Benedetti-Cecchi L. (2010). The seaweed Caulerpa racemosa on Mediterranean rocky reefs: from passenger to driver of ecological change. Ecology 91, 2205–2212. doi: 10.1890/09-1857.1
Bulleri F., Benedetti-Cecchi L. (2008). Facilitation of the introduced green alga Caulerpa racemosa by resident algal turfs: experimental evaluation of underlying mechanisms. Mar. Ecol. Prog. Ser. 364, 77–86. doi: 10.3354/meps07484
Çinar M. E., Féral J.-P., Arvanitidis C., David R., Taşkin E., Sini M., et al. (2020). Coralligenous assemblages along their geographical distribution: testing of concepts and implications for management. Aquat. Conservation: Mar. Freshw. Ecosyst. 30, 1578–1594. doi: 10.1002/aqc.3365
Clarke K. R. (1993). Non-parametric multivariate analyses of changes in community structure. Aust. J. Ecol. 18, 117–143. doi: 10.1111/j.1442-9993.1993.tb00438.x
Coll M., Piroddi C., Albouy C., Ben Rais Lasram F., Cheung W. W., Christensen V., et al. (2012). The Mediterranean Sea under siege: spatial overlap between marine biodiversity, cumulative threats and marine reserves. Global Ecol. Biogeography 21, 465–480. doi: 10.1111/j.1466-8238.2011.00697.x
Costanza R., d’Arge R., De Groot R., Farber S., Grasso M., Hannon, et al. (1997). The value of the world’s ecosystem services and natural capital. Nature 387, 253–260. doi: 10.1038/387253a0
Costanza R., De Groot R., Braat L., Kubiszewski I., Fioramonti L., Sutton P., et al. (2017). Twenty years of ecosystem services: How far have we come and how far do we still need to go? Ecosystem Serv. 28, 1–16. doi: 10.1016/j.ecoser.2017.09.008
de Bettignies T., Grimond J., Le Gall L., Sartoretto S. (2020). La Planète Revisité au Parc Naturel Marin du Cap Corse et de l’Agriate: conservation des récifs infralittoraux (dominés par les algues photophiles, entre 15 et 35m). Preliminary report UMS PatriNat OFB/CNRS/MNHN. (Paris, France: PatriNat (OFB-CNRS-MNHN)). 27 pp.
de Bettignies T., Hébert C., Assis J., Bartsch I., Bekkby T., Christie H., et al. (2021). Case Report for kelp forests habitat. OSPAR 787/2021. London, UK: OSPAR Commission. 39 pp.
De la Hoz C., Ramos E., Acevedo A., Puente A., Losada Í., Juanes J. (2018). OCLE: A European open access database on climate change effects on littoral and oceanic ecosystems. Prog. Oceanography 168, 222–231. doi: 10.1016/j.pocean.2018.09.021
Deter J., Descamp P., Boissery P., Ballesta L., Holon F. (2012). A rapid photographic method detects depth gradient in coralligenous assemblages. J. Exp. Mar. Biol. Ecol. 418, 75–82. doi: 10.1016/j.jembe.2012.03.006
Díaz-Ferguson E. E., Moyer G. R. (2014). “History, applications, methodological issues and perspectives for the use of environmental DNA (eDNA) in marine and freshwater environments,” in Revista de Biología Tropical, 62 (San Pedro de Montes de Oca, Costa Rica: Universidad de Costa Rica), 1273–1284.
Dray S., Chessel D., Thioulouse J. (2003). Procrustean co-inertia analysis for the linking of multivariate datasets. Écoscience 10, 110–119. doi: 10.1080/11956860.2003.11682757
Dumke I., Nornes S. M., Purser A., Marcon Y., Ludvigsen M., Ellefmo S. L., et al. (2018). First hyperspectral imaging survey of the deep seafloor: high-resolution mapping of manganese nodules. Remote Sens. Environ. 209, 19–30. doi: 10.1016/j.rse.2018.02.024
Durrieu de Madron X. D., Guieu C., Sempéré R., Conan P., Cossa D., D’Ortenzio F., et al. (2011). Marine ecosystems’ responses to climatic and anthropogenic forcings in the Mediterranean. Prog. Oceanography 91, 97–166. doi: 10.1016/j.pocean.2011.02.003
Ferrigno F., Russo G. F., Sandulli R. (2017). Coralligenous Bioconstructions Quality Index (CBQI): a synthetic indicator to assess the status of different types of coralligenous habitats. Ecol. Indic. 82, 271–279. doi: 10.1016/j.ecolind.2017.07.020
Filbee-Dexter K., Scheibling R. E. (2014). Sea urchin barrens as alternative stable states of collapsed kelp ecosystems. Mar. Ecol. Prog. Ser. 495, 1–25. doi: 10.3354/meps10573
Filbee-Dexter K., Wernberg T. (2018). Rise of turfs: a new battlefront for globally declining kelp forests. BioScience 68, 64–76. doi: 10.1093/biosci/bix147
Friedman J., Tibshirani R., Hastie T. (2010). Regularization paths for Generalized Linear Models via coordinate descent. J. Stat. Software 33, 1–22. doi: 10.18637/jss.v033.i01
Galil B. S. (2000). A sea under siege – Alien species in the Mediterranean. Biol. Invasions 2, 177–186. doi: 10.1023/A:1010057010476
Garrabou J., Coma R., Bensoussan N., Bally M., Chevaldonné P., Cigliano M., et al. (2009). Mass mortality in Northwestern Mediterranean rocky benthic communities: effects of the 2003 heat wave. Global Change Biol. 15, 1090–1103. doi: 10.1111/j.1365-2486.2008.01823.x
Geertz-Hansen O., Enríquez S., Duarte C. M., Agustí S., Vaqué D., Vidondo B. (1994). Functional implications of the form of Codium bursa, a balloon-like Mediterranean macroalga. Mar. Ecol. Prog. Ser. 108, 153–160. doi: 10.3354/meps108153
Giangrande A., Pierri C., Del Pasqua M., Gravili C., Gambi M. C., Gravina M. F. (2020). The Mediterranean in check: biological invasions in a changing sea. Mar. Ecol. 41, e12583. doi: 10.1111/maec.12583
Goffredo S., Mattioli G., Zaccanti F. (2004). Growth and population dynamics model of the Mediterranean solitary coral Balanophyllia europaea (Scleractinia, Dendrophylliidae). Coral Reefs 23, 433–443. doi: 10.1007/s00338-004-0395-9
Halpern B. S., Frazier M., Potapenko J., Casey K. S., Koenig K., Longo C., et al (2015). Spatial and temporal changes in cumulative human impacts on the world’s ocean. Nat. Commun. 6 (1), 1–7. doi: 10.1038/ncomms8615
Harmelin J. G., Marinopoulos J. (1994). Population structure and partial mortality of the gorgonian Paramuricea clavata (Risso) in the north-western Mediterranean (France, Port-Cros Island). Mar. Life 4, 5–13.
Hewitt J. E., Ellis J. I., Thrush S. F. (2016). Multiple stressors, nonlinear effects and the implications of climate change impacts on marine coastal ecosystems. Global Change Biol. 22, 2665–2675. doi: 10.1111/gcb.13176
Hill N. A., Lucieer V., Barrett N. S., Anderson T. J., Williams S. B. (2014). Filling the gaps: predicting the distribution of temperate reef biota using high-resolution biological and acoustic data. Estuarine Coast. Shelf Sci. 147, 137–147. doi: 10.1016/j.ecss.2014.05.019
Hoffmann L., Renard R., Demoulin V. (1992). Phenology, growth, and biomass of Cystoseira balearica in Calvi (Corsica). Mar. Ecol. Prog. Ser. 80, 249–254. doi: 10.3354/meps080249
Holon F., Marre G., Parravicini V., Mouquet N., Bockel T., Descamp P., et al. (2018). A predictive model based on multiple coastal anthropogenic pressures explains the degradation status of a marine ecosystem: implications for management and conservation. Biol. Conserv. 222, 125–135. doi: 10.1016/j.biocon.2018.04.006
Holon F., Mouquet N., Boissery P., Bouchoucha M., Delaruelle G., Tribot A. S., et al. (2015). Fine-scale cartography of human impacts along French Mediterranean coasts: a relevant map for the management of marine ecosystems. PloS One 10, e0135473. doi: 10.1371/journal.pone.0135473
Klein J., Verlaque M. (2008). The Caulerpa racemosa invasion: a critical review. Mar. pollut. Bull. 56, 205–225. doi: 10.1016/j.marpolbul.2007.09.043
La Rivière M., Michez N., Delavenne J., Andres S., Fréjefond C., Janson A.-L., et al. (2021). Fiches descriptives des biocénoses benthiques de Méditerranée (Paris: UMS PatriNat (OFB-CNRS-MNHN), 660 pp.
Lejeusne C., Chevaldonné P., Pergent-Martini C., Boudouresque C. F., Pérez T. (2010). Climate change effects on a miniature ocean: the highly diverse, highly impacted Mediterranean Sea. Trends Ecol. Evol. 25, 250–260. doi: 10.1016/j.tree.2009.10.009
Lepareur F., Manceau M., Reyjol Y., Touroult J., Robert S., Vest F., et al. (2022). The nationwide ‘ZNIEFF’ inventory in France: an open dataset of more than one million species data in zones of high ecological value. Biodiversity Data J. 10, e83369. doi: 10.3897/BDJ.10.e83369
Lévêque L., Broudin C., Camusat M., Fontana Y., Houbin C., Leclerc J.-C., et al. (2017). Inventaire et caractérisation des communautés benthiques du Plateau de la Méloine. Research Report (Roscoff, France: Station Biologique de Roscoff), 132 pp.
Linares C., Coma R., Garrabou J., Díaz D., Zabala M. (2008). Size distribution, density and disturbance in two Mediterranean gorgonians: Paramuricea clavata and Eunicella singularis. J. Appl. Ecol. 45, 688–699. doi: 10.1111/j.1365-2664.2007.01419.x
Ling S. D., Scheibling R. E., Rassweiler A., Johnson C. R., Shears N., Connell S. D., et al. (2015). Global regime shift dynamics of catastrophic sea urchin overgrazing. Philos. Trans. R. Soc. B: Biol. Sci. 370, 20130269. doi: 10.1098/rstb.2013.0269
Mangialajo L., Chiantore M., Cattaneo-Vietti R. (2008). Loss of fucoid algae along a gradient of urbanisation, and structure of benthic assemblages. Mar. Ecol. Prog. Ser. 358, 63–74. doi: 10.3354/meps07400
Marre G., Holon F., Luque S., Boissery P., Deter J. (2019). Monitoring marine habitats with photogrammetry: a cost-effective, accurate, precise and high-resolution reconstruction method. Front. Mar. Sci. 6. doi: 10.3389/fmars.2019.00276
Michez N., Fourt M., Aish A., Bellan G., Bellan-Santini D., Chevaldonné P., et al. (2014). Typologie des biocénoses benthiques de Méditerranée, Version 2. Report SPN 2014 – MNHN. Paris, France: UMS PatriNat, Muséum national d'Histoire naturelle. 33 pp.
Mieszkowska N., Kendall M. A., Hawkins S. J., Leaper R., Williamson P., Hardman-Mountford N. J., et al. (2006). “Changes in the range of some common rocky shore species in Britain − a response to climate change?,” in Marine biodiversity: patterns and processes, assessment, threats, management and conservation. Eds. Hawkins S. J., Hutchinson N., Jensen A. C., Williams J. A. (The Marine Biological Association of the United Kingdom, Plymouth), 241–251.
Montefalcone M., Morri C., Parravicini V., Bianchi C. N. (2015). A tale of two invaders: divergent spreading kinetics of the alien green algae Caulerpa taxifolia and Caulerpa cylindracea. Biol. Invasions 17, 2717–2728. doi: 10.1007/s10530-015-0908-1
Nikolaou A., Tsirintanis K., Rilov G., Katsanevakis S. (2023). Invasive fish and sea urchins drive the status of canopy forming macroalgae in the Eastern Mediterranean. Biology 12, 763. doi: 10.3390/biology12060763
Office de l’Environnement de la Corse (2011). Analyse stratégique régionale de la Corse: enjeux et propositions de création d’aires marines protégées. OEC Report (Corte, OEC: Office de l'Environnement de la Corse).
Oksanen J., Blanchet F. G., Friendly M., Kindt R., Legendre P., McGlinn D., et al. (2020). Vegan: community ecology package (version 2.5-7). Available online at: https://CRAN.R-project.org/package=vegan (Accessed February 5, 2024).
Pandolfi J. M., Bradbury R. H., Sala E., Hughes T. P., Bjorndal K. A., Cooke R. G., et al. (2003). Global trajectories of the long-term decline of coral reef ecosystems. Science 301, 955–958. doi: 10.1126/science.1085706
Pasqualini V., Pergent-Martini C., Clabaut P., Pergent G. (1998). Mapping of Posidonia oceanica using aerial photographs and side scan sonar: application off the island of Corsica (France). Estuarine Coast. Shelf Sci. 47, 359–367. doi: 10.1006/ecss.1998.0361
Peleg O., Guy-Haim T., Yeruham E., Silverman J., Rilov G. (2020). Tropicalization may invert trophic state and carbon budget of shallow temperate rocky reefs. J. Ecol. 108, 844–854. doi: 10.1111/1365-2745.13316
Piazzi L., Balata D., Bulleri F., Gennaro P., Ceccherelli G. (2016). The invasion of Caulerpa cylindracea in the Mediterranean: the known, the unknown, and the knowable. Mar. Biol. 163, 161. doi: 10.1007/s00227-016-2910-1
Piazzi L., Balata D., Ceccherelli G., Cinelli F. (2005). Interactive effect of sedimentation and Caulerpa racemosa var. cylindracea invasion on macroalgal assemblages in the Mediterranean Sea. Estuarine Coast. Shelf Sci. 64, 467–474. doi: 10.1016/j.ecss.2005.03.010
Piazzi L., Ceccherelli G., Balata D., Cinelli F. (2003). Early patterns of Caulerpa racemosa recovery in the Mediterranean Sea: the influence of algal turfs. J. Mar. Biol. Assoc. United Kingdom 83, 27–29. doi: 10.1017/S0025315403006751h
Piazzi L., Ferrigno F., Guala I., Cinti M. F., Conforti A., De Falco G., et al. (2022). Inconsistency in community structure and ecological quality between platform and cliff coralligenous assemblages. Ecol. Indic. 136, 108657. doi: 10.1016/j.ecolind.2022.108657
Ponti M., Perlini R. A., Ventra V., Grech D., Abbiati M., Cerrano C. (2014). Ecological shifts in Mediterranean coralligenous assemblages related to gorgonian forest loss. PloS One 9, e102782. doi: 10.1371/journal.pone.0102782
Raitsos D. E., Beaugrand G., Georgopoulos D., Zenetos A., Pancucci-Papadopoulou A. M., Theocharis A., et al. (2010). Global climate change amplifies the entry of tropical species into the Eastern Mediterranean Sea. Limnology Oceanography 55, 1478–1484. doi: 10.4319/lo.2010.55.4.1478
Rossi S. (2013). The destruction of the ‘animal forests’ in the oceans: towards an over-simplification of the benthic ecosystems. Ocean Coast. Manage. 84, 77–85. doi: 10.1016/j.ocecoaman.2013.07.004
Sales M., Ballesteros E. (2010). Long-term comparison of algal assemblages dominated by Cystoseira crinita (Fucales, Heterokontophyta) from Cap Corse (Corsica, North Western Mediterranean). Eur. J. Phycology 45, 404–412. doi: 10.1080/09670262.2010.498585
Santín A., Grinyó J., Ambroso S., Uriz M. J., Dominguez-Carrió C., Gili J. M. (2019). Distribution patterns and demographic trends of demosponges at the Menorca Channel (Northwestern Mediterranean Sea). Prog. Oceanography 173, 9–25. doi: 10.1016/j.pocean.2019.02.002
Telesca L., Belluscio A., Criscoli A., Ardizzone G., Apostolaki E. T., Fraschetti S., et al. (2015). Seagrass meadows (Posidonia oceanica) distribution and trajectories of change. Sci. Rep. 5, 12505. doi: 10.1038/srep12505
Thibaut T., Blanfuné A., Boudouresque C. F., Personnic S., Ruitton S., Ballesteros E., et al. (2017). An ecosystem-based approach to assess the status of Mediterranean algae-dominated shallow rocky reefs. Mar. pollut. Bull. 117, 311–329. doi: 10.1016/j.marpolbul.2017.01.029
Thioulouse J., Dray S., Dufour A. B., Siberchicot A., Jombart T., Pavoine S. (2018). Multivariate analysis of ecological data with ade4 (New York: Springer). doi: 10.1007/978-1-4939-8850-1
Thomsen P. F., Kielgast J., Iversen L. L., Wiuf C., Rasmussen M., Gilbert M. T. P., et al. (2012). Monitoring endangered freshwater biodiversity using environmental DNA. Mol. Ecol. 21, 2565–2573. doi: 10.1111/j.1365-294X.2011.05418.x
Tsirintanis K., Sini M., Doumas O., Trygonis V., Katsanevakis S. (2018). Assessment of grazing effects on phytobenthic community structure at shallow rocky reefs: an experimental field study in the North Aegean Sea. J. Exp. Mar. Biol. Ecol. 503, 31–40. doi: 10.1016/j.jembe.2018.01.008
Tyberghein L., Verbruggen H., Pauly K., Troupin C., Mineur F., De Clerck O. (2012). Bio-ORACLE: a global environmental dataset for marine species distribution modelling. Global Ecol. Biogeography 21, 272–281. doi: 10.1111/j.1466-8238.2011.00656.x
Valisano L., Palma M., Pantaleo U., Calcinai B., Cerrano C. (2019). Characterization of North–Western Mediterranean coralligenous assemblages by video surveys and evaluation of their structural complexity. Mar. pollut. Bull. 148, 134–148. doi: 10.1016/j.marpolbul.2019.07.012
Vanalderweireldt L., Albouy C., Le Loc’h F., Millot R., Blestel C., Patrissi M., et al. (2022). Ecosystem modelling of the Eastern Corsican Coast (ECC): case study of one of the least trawled shelves of the Mediterranean Sea. J. Mar. Syst. 235, 103798. doi: 10.1016/j.jmarsys.2022.103798
Verlaque M., Boudouresque C. F., Meinesz A., Gravez V. (2000). The Caulerpa racemosa complex (Caulerpales, Ulvophyceae) in the Mediterranean Sea. Botanica Marina 43, 49–68. doi: 10.1515/BOT.2000.005
Verlaque M., Durand C., Huisman J. M., Boudouresque C. F., Le Parco Y. (2003). On the identity and origin of the Mediterranean invasive Caulerpa racemosa (Caulerpales, Chlorophyta). Eur. J. Phycology 38, 325–339. doi: 10.1080/09670260310001612592
Vidondo B., Duarte C. M. (1998). Population structure, dynamics, and production of the Mediterranean macroalga Codium bursa (Chlorophyceae). J. Phycology 34, 918–924. doi: 10.1046/j.1529-8817.1998.340918.x
Ward D. F., Larivière M. C. (2004). Terrestrial invertebrate surveys and rapid biodiversity assessment in New Zealand: lessons from Australia. New Z. J. Ecol. 28, 151–159. Availavle online at: https://www.jstor.org/stable/24058222
Waycott M., Duarte C. M., Carruthers T. J., Orth R. J., Dennison W. C., Olyarnik S., et al. (2009). Accelerating loss of seagrasses across the globe threatens coastal ecosystems. Proc. Natl. Acad. Sci. 106, 12377–12381. doi: 10.1073/pnas.0905620106
Wernberg T., Bennett S., Babcock R. C., de Bettignies T., Cure K., Depczynski M., et al. (2016). Climate-driven regime shift of a temperate marine ecosystem. Science 353, 169–172. doi: 10.1126/science.aad8745
Wickham H., François R., Henry L., Müller K., Vaughan D. (2023). dplyr: a grammar of data manipulation. R package version 1.1.4. Available online at: https://CRAN.R-project.org/package=dplyr (Accessed February 5, 2024).
Keywords: canopy-forming seaweed, Caulerpa cylindracea, Cystoseira (sensu lato) forest, rapid assessment survey, temperate reefs
Citation: de Bettignies T, Vanalderweireldt L, Launay M, Moutardier G, Pasqualini V, Durieux É and Le Gall L (2025) The role of algae in structuring reef communities: innovative monitoring and ecological insights within a Mediterranean conservation priority area. Front. Mar. Sci. 12:1516792. doi: 10.3389/fmars.2025.1516792
Received: 24 October 2024; Accepted: 20 February 2025;
Published: 12 March 2025.
Edited by:
Stanislao Bevilacqua, University of Trieste, ItalyReviewed by:
Julian Evans, University of Malta, MaltaCopyright © 2025 de Bettignies, Vanalderweireldt, Launay, Moutardier, Pasqualini, Durieux and Le Gall. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Thibaut de Bettignies, dGhpYmF1dC5kZS1iZXR0aWduaWVzQG1uaG4uZnI=
†These authors have contributed equally to this work and share first authorship
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.