 María R. Muciño-Reyes1,2
María R. Muciño-Reyes1,2 Edlin Guerra-Castro3,4
Edlin Guerra-Castro3,4 Raúl E. Castillo-Cupul5
Raúl E. Castillo-Cupul5 Carlos E. Paz-Ríos6
Carlos E. Paz-Ríos6 Nuno Simões2,4,7*
Nuno Simões2,4,7*- 1Posgrado en Ciencias Biológicas, Universidad Nacional Autónoma de México, Unidad de Posgrado, Mexico City, Mexico
- 2Unidad Multidisciplinaria de Docencia e Investigación Sisal, Facultad de Ciencias, Universidad Nacional Autónoma de México, Hunucmá, Mexico
- 3Escuela Nacional de Estudios Superiores Unidad Mérida, Universidad Nacional Autónoma de México, Mérida, Mexico
- 4Laboratorio Nacional de Resiliencia Costera, Laboratorios Nacionales, Consejo Nacional de Humanidades, Ciencias y Tecnologías (CONAHCYT), Ciudad de México, Mexico
- 5Kalanbio A.C. Research for Conservation, Mérida, Mexico
- 6Instituto de Ecología, Pesquerías y Oceanografía de la Universidad Autónoma de Campeche (EPOMEX-UAC), Campeche, Mexico
- 7International Chair for Coastal and Marine Studies in Mexico, Harte Research Institute for Gulf of Mexico Studies, Texas A&M University-Corpus Christi, Corpus Christi, TX, United States
Despite the ecological importance of sandy beaches, the diversity and distribution of macrocrustacean communities in the eastern Mexican coast (Mexican coast of the Gulf of Mexico and Caribbean Sea) remain understudied. As the first large-scale sampling effort along the eastern Mexican coast, this study aimed to estimate macrocrustacean diversity and evaluate sampling methodologies, while providing historical context for macrocrustacean research in the region. Two sampling designs using sediment cores (4 versus 6 cores per transect) were implemented across 15 sandy beaches. A total of 3,352 organisms were collected, representing 22 species from 17 genera, spanning 14 families, 7 suborders, 5 orders, and 2 superorders. Amphipoda and Isopoda exhibited the highest species richness and abundance. The study yielded seven new geographical records (Haustorius jayneae, Rhepoxynius epistomus, Americorchestia salomani, Heterodina mosaica, Cassidinidea ovalis, Exosphaeroma diminutum, and Sphaeroma walkeri). Results demonstrated that the 6-core sampling design provided better diversity representation. An updated checklist comprising 77 species/taxa for the eastern Mexican coast was compiled, integrating historical and new data. This comprehensive assessment enhances our understanding of these vulnerable ecosystems and emphasizes the need for broader temporal and spatial scale studies to inform effective conservation strategies.
Introduction
Sandy beaches are important ecosystems that provide vital ecosystem services such as shoreline protection, sediment transport and storage, pollutant decomposition, water filtration, nutrient recycling, and biodiversity maintenance (Defeo et al., 2009). Additionally, they are tourist destinations of great economic importance, such as the Mexican Caribbean beaches that received 38 million international tourists in 2022 (SECTUR and DATATUR, 2022). Sandy beaches + are dynamic ecosystems that are inhabited by numerous macrocrustacean species, many with particular adaptations (including small size, tidal rhythms, orientational responses, and behavioral flexibility) that result in their not being found in any other environment (Chelazzi and Vannini, 1988; Brown, 1996; Scapini, 2006). The role of macrocrustaceans in sandy beaches is of great importance. Within the trophic chain of this environment, they serve as primary consumers by feeding on detritus, and in turn, they are part of the diet of polychaetes, mollusks, birds, and fishes, some of those being commercially important (Bocher et al., 2001; Bergamino et al., 2013). Additionally, the abundance of crustaceans serves as an indicator of good beach health (Cardoso et al., 2016). Moreover, these organisms play a key role in ecosystem processes by contributing to the mineralization of organic matter and the recycling of nutrients (Defeo et al., 2009). Some macrocrustacean species, mainly the talitrid amphipods and hippid crabs, have been used as bioindicators of environmental variability produced by freshwater discharges, human impact, human use, and ecological quality conditions in beach and dune environments (Lercari and Defeo, 1999; Fanini et al., 2005; Barca-Bravo et al., 2008; Noriega et al., 2012; Gonçalves et al., 2013; Bessa et al., 2014; Nourisson et al., 2014; Cardoso et al., 2016).
However, many phenomena threaten beach ecosystems and their associated fauna with serious consequences. For example, pollution and habitat modification due to population growth (more than 40% of the world’s population lives near the coast), recreational seashore activities, beach modification through engineering works, accelerated rates of erosion, and the potential rise in the sea levels due to effects of global climate change can all negatively impact these ecosystems and fauna (Martínez et al., 2007; Schlacher et al., 2008; Defeo et al., 2009). In addition, oil industry accidents have had serious environmental consequences in the coastal zone, mostly represented by sandy beaches (Jernelöv and Lindén, 2010; Sun et al., 2015; Joye, 2016). And the recent massive arrivals and decomposition of Sargassum on sandy beaches, especially in the Caribbean, has had a significant effect on the mortality of communities associated with this environment (Rodríguez-Martínez et al., 2019). Despite the importance of sandy beaches and the anthropogenic threats affecting them, there remains a knowledge gap regarding the diversity and distribution patterns of the macrocrustacean communities inhabiting the extensive sandy beaches of the eastern Mexican coast (Gulf of Mexico and the Mexican Caribbean).
On the eastern Mexican coast, previous studies on benthic crustaceans have focused heavily on coral reefs (Oliva-Rivera, 2003; Winfield and Escobar-Briones, 2007; Winfield et al., 2007, 2010, 2013, 2020; Ortíz et al., 2013; Paz-Ríos et al., 2013b, 2019), shallow waters and coastal lagoons (Winfield and Ortiz, 1996; Raz-Guzmán and Soto, 2017). Knowledge about macrobenthic crustaceans on beaches is still limited even though the eastern Mexican coastline extends for 3,294 km, with sandy beaches being its most widely distributed habitat (92.46%) (Silva-Casarín et al., 2011; INEGI, 2022). The first effort to describe the crustacean fauna associated with eastern Mexican beaches was undertaken by Dexter (1976). Since then, nine additional studies have been conducted up to the present date, most of them at a local scale (Méndez-Ubach et al., 1985; Pacheco-Ríos, 2010; Miranda, 2012; Martínez, 2013; Paz-Ríos et al., 2013a; Wildish and Lecroy, 2014; Rocha-Ramírez et al., 2016; González et al., 2017; Guerra-Castro et al., 2020). Only one study has covered a large spatial scale (29 beaches, primarily in Veracruz state) but with limited sampling effort per beach (1 replicate) (Méndez-Ubach et al., 1985); and only one has focused on insular sandy beach habitats within the Gulf of Mexico (Isla Pérez, Alacranes Reef National Park) (Paz-Ríos et al., 2013a). As a result of these studies, a total of 45 species/taxa have been described, with Veracruz state being the best represented at 30 species/taxa, followed by Yucatán (19), Quintana Roo (3), Campeche (6), and Tamaulipas (4). It is noteworthy that the state of Tabasco lacks any information regarding the macrocrustaceans inhabiting its beaches, making this the first work addressing this subject for that state.
The present study provides quantitative and qualitative estimates about macrocrustacean diversity on 15 beaches along the eastern Mexican coast, a sampling effort unprecedented in the western Atlantic, as previous extensive sampling efforts have been limited to local or state-level scales, or when covering larger regions, were restricted to single beaches per state. In addition to the regional scale addressed by this work, we tested two sampling designs at the local (beach) scale to measure their effectiveness. Moreover, this information, together with information found in theses and published articles, has been used to create an updated checklist of sandy beach macrocrustaceans. This compilation is intended to serve as baseline data for forthcoming studies examining the multifaceted impacts of pollution, erosion, beach modification, as well as the influences stemming from tourism development, global climate change, oil exploitation, and the unusual arrival of Sargassum.
Materials and methods
Study area
The study area includes 15 beaches distributed along the southern part of the Gulf of Mexico and the northern Caribbean Sea (eastern Mexican coast) (Figure 1), spanning low (18° N) and mid-latitude (22°N) and experiencing a diversity of climates conditions along its wide geographic range. In the northern zone (Tamaulipas state), the climate is semi-arid with an average annual rainfall of 800 mm. In the other states, the climate is warm-humid to sub-humid, with the highest levels of precipitation in Veracruz, Tabasco, and Campeche (1,500-3,500 mm) and to a lesser extent in Yucatán and Quintana Roo (600-1,400 mm) (de la Lanza-Espino et al., 2013). The beaches of the states of Tamaulipas, Veracruz and Tabasco are characterized by their terrigenous origin (sediments supplied by rivers), their location in front of a narrow continental shelf and are influenced by the warm current of El Lazo (Silva-Casarín et al., 2011). The sandy coasts that border the Yucatan Peninsula are the product of biogenic genesis, situated on an emergent karst platform composed predominantly of calcium carbonate sediments. Devoid of surface fluvial inputs, these coastal landscapes are intricately governed by the dynamic interaction of the Yucatan and Caribbean currents.
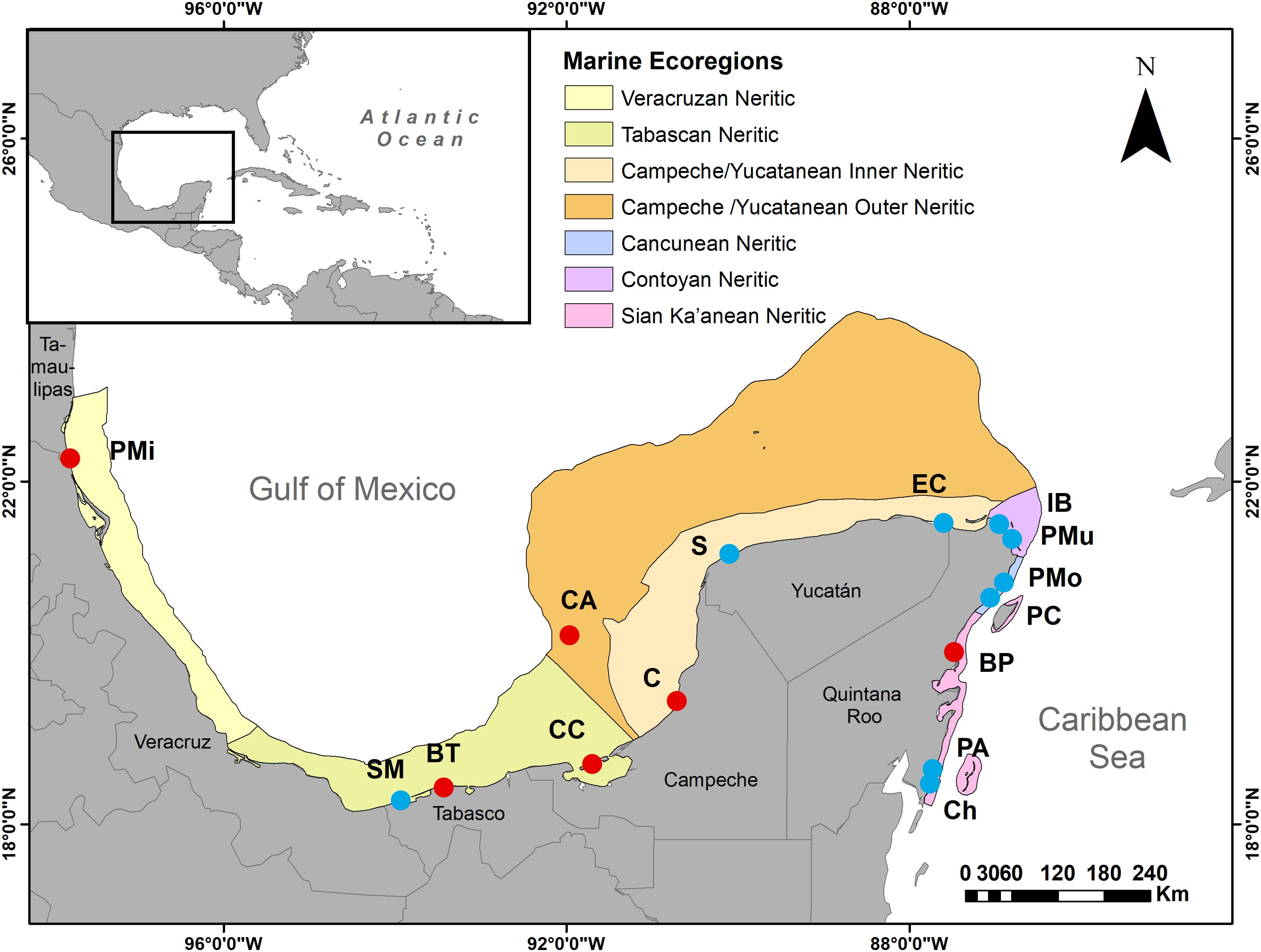
Figure 1. Sand beaches studied on the eastern Mexican coast. Marine Ecoregions of North America (Wilkinson et al., 2009: level 3 shallow sub-regions) are also shown to provide a geographical context, along the six Mexican coastal states. Dots represent beach localities: PMi, Playa Miramar; SM, Sánchez Magallanes; BT, Barra de Tupilco; CC, Ciudad del Carmen; CA, Cayo Arcas; C, Champotón; S, Sisal; EC, El Cuyo; IB, Isla Blanca; PMu, Playa Mujeres; PMo, Puerto Morelos; PC, Playa del Carmen; BP=Boca Paila; PA, Puerto Ángel; Ch, Chahuayxol. Dot´s colors represent two different sampling protocols (further described in the materials and methods section, Figure 2). Red: sample design A, Blue: sample design B.
Sampling protocol
Sampling was conducted within a one-year time frame (March, April, and June 2019, and in June 2020) at 15 localities (beaches) (Figure 1; Table 1) representing all states along the eastern Mexican coast except Veracruz, where sampling was precluded by conditions imposed during the SARS CoV-2 pandemic. The sampling design was hierarchical and nested across five spatial scales: regions (kilometers), subregions (kilometers), localities (hundreds of meters), sites (tens of meters), and cores (meters). None of the 15 localities were within 2 km of a river mouth, coastal lagoon, lake, or urban communities. At each locality, three sites separated by 50 m were selected and at each site one of three different sampling designs (A or B) was used (Table 1; Figure 2). In design A, at each of the 3 locality sites, two transects were drawn parallel to the beach, the first in the infratidal zone and the second in the intertidal zone. Along each transect, 3 sediment samples separated by 1 m (replicates) were obtained. In design B, the sampling effort was doubled on the replicate scale, i.e., 6 sediment samples per transect (infratidal and intertidal) were collected at each site (1, 2, and 3) for each locality.
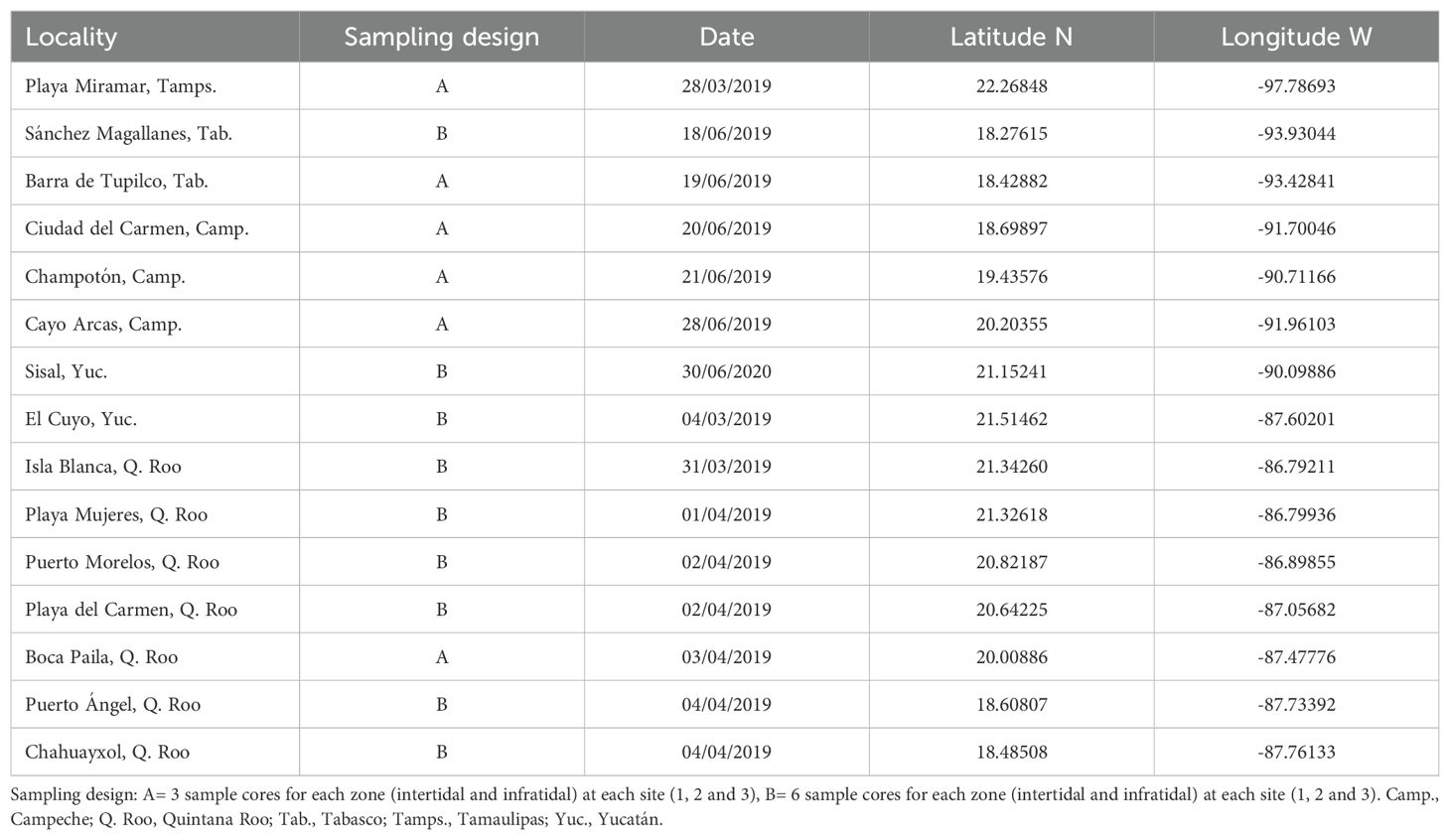
Table 1. Sampled localities, sampling design type, date, and geographical coordinates.
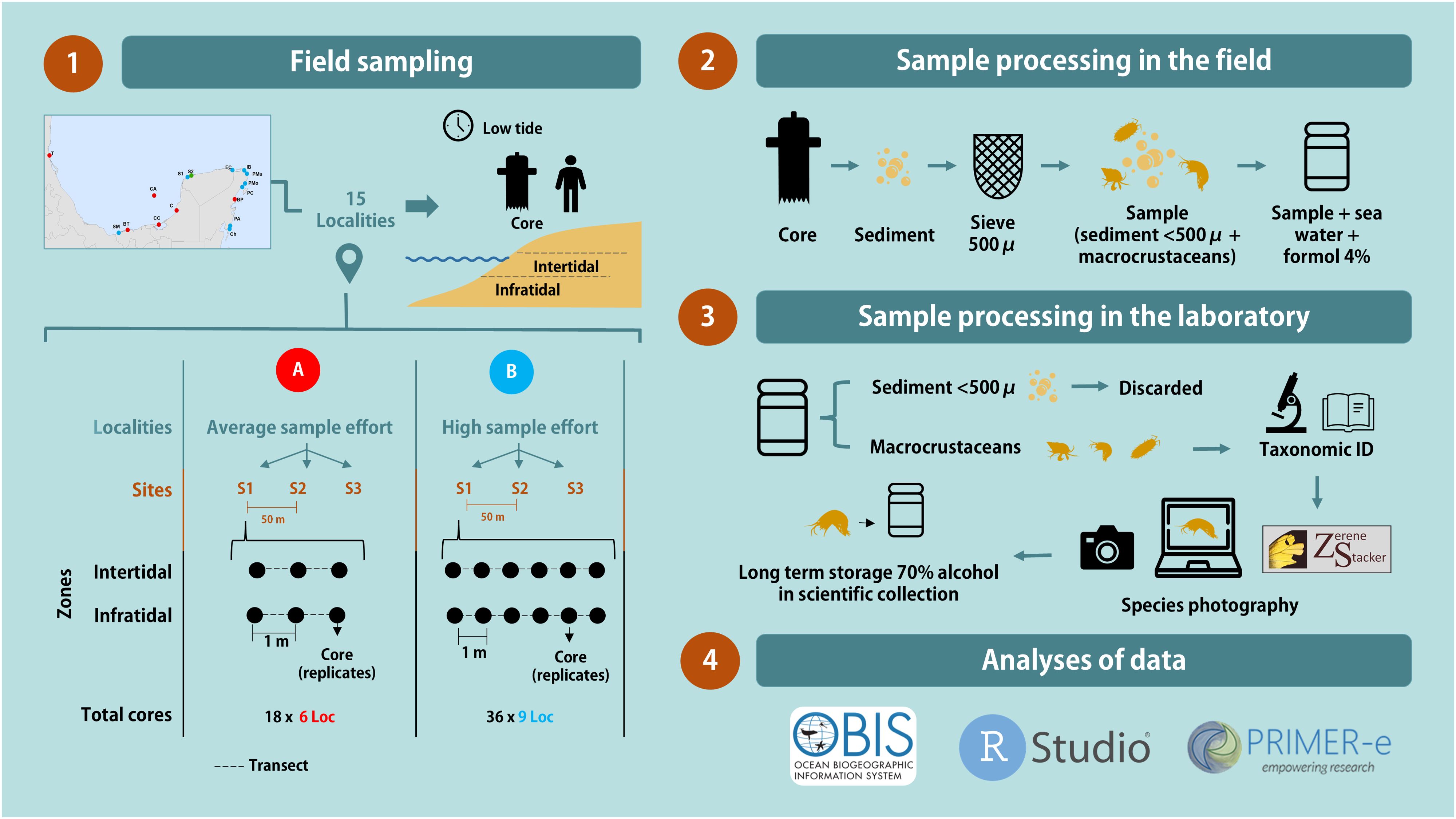
Figure 2. Flowchart infographic of the used methodology sequence. Sampling designs: (A) Increase beach sample size and geographic representation while reducing field and lab work time. (B) Enhance local sampling for comparisons across spatial scales, with increased field and lab work time.
The objectives of using three sampling designs were as follows: design A, to increase the number of sampled beaches and enhance geographic representativeness while reducing the time invested in field and laboratory work; design B, to increase the local sampling effort to enable comparisons across different spatial scales, ranging from meters to hundreds of thousands of kilometers, albeit with an increased time requirement for field sampling and laboratory work.
The samples were collected using a PVC sediment corer (diameter 15 cm and length 30 cm; equivalent area 0.0177 m2, volume of 0.005 m3). Each sediment core was sieved in the field through a mesh bag (mesh size 500 µm), which retained the macrofauna and sediment particles larger than 500 µm, then sorted organisms were fixed with 4% formaldehyde.
Laboratory Protocols
In the laboratory, organisms were separated from the sediment, counted, and identified to the lowest possible taxonomic category using taxonomic keys for each group (Abele and Kim, 1986; LeCroy, 2000, 2002, 2004, 2007, 2011; Hsueh, 2015). The specimens were then preserved in 70% alcohol at the Colección de Crustáceos de Yucatán (YUC-CC) from Unidad Multidisciplinaria de Docencia e Investigación, Sisal, Facultad de Ciencias, UNAM.
Analyses of data
Based on the species/taxa identified in the present study, a faunistic list was compiled following the taxonomic classifications proposed by Ahyong et al. (2011) and Lowry and Myers (2017). In addition, the number of collected organisms was counted, and alpha diversity (species richness) per locality was calculated.
The distribution and abundance patterns of species/taxa along the coast were identified with an ordered shade plot. The abundance values were transformed into natural logarithms (plus 1) to downweigh the effect of highly abundant species. The species/taxa were ordered based on their similarity (Bray-Curtis) in their spatial distribution with respect to the locality, using a hierarchical cluster analysis using the PRIMER7 software (Clarke et al., 2014).
To evaluate the two sampling designs (A and B), three extrapolation and rarefaction curves based on sample size, coverage, and completeness were derived using the incidence data of each species per locality, and the incidence data per region (Gulf of Mexico and Caribbean Sea) with the iNEXT package (Chao et al., 2014; Hsieh et al., 2016) in R Studio. These measures allowed us to infer the representativeness of the sampling effort at each locality and each region. The biological data collected were organized into a Darwin Core dataset that is available in the OBIS website (https://doi.org/10.15468/zfgt5w) (Muciño-Reyes et al., 2024).
Results
Among the 3,352 organisms collected from the 432 sediment cores, 28 species/taxa were recognized (Supplementary 1, 2; Figures 3, 4), representing 22 species, 17 genera, 14 families, 7 suborders, 5 orders, and 2 superorders (Eucarida and Peracarida). The superorder Peracarida had higher species/taxa richness (22) and relative abundance (99.8%) than Eucarida (5 species/taxa and 0.2%). Of the 5 orders identified, Isopoda was represented by 13 species, followed by Amphipoda with 7 species, Decapoda with 6 species/taxa, and Tanaidacea and Cumacea with 1 species each. Of the total abundance, Amphipoda accounted for 69.8%, Isopoda 29.9%, and the rest of the orders 0.3%. The amphipod Mexorchestia carpenteri raduloviciae Wildish and Lecroy, 2014 was the most abundant, with 66.41% of the total abundance, followed by the isopods Excirolana braziliensis Richardson (1912) with 16.74%, and Tylos marcuzzi Giordani Soika (1954) with 9.05%, then the peracarids Excirolana mayana (Ives, 1891) with 3.22% and Haustorius jayneae Foster and Lecroy (1991) with 2.78%, and all other species/taxa with <1% each.

Figure 3. Amphipods and decapods found in beaches of the eastern Mexican coast: (A) Americorchestia salomani, (B) Cerapus benthophilus, (C) Haustorius jayneae, (D) Melita planaterga, (E) Mexorchestia carpenteri raduloviciae, (F) Parhyalella whelpleyi, (G) Rhepoxynius epistomus, (H) Emerita talpoida.

Figure 4. Isopods found in beaches of the eastern Mexican coast: (A) Ancinus sp 1., (B) Cassidinidea ovalis, (C) Excirolana braziliensis, (D) Excirolana mayana, (E) Excorallana berbicencis, (F) Excorallana sp. 1, (G) Excorallana cf. productatelson, (H) Exosphaeroma diminutum, (I) Exosphaeroma sp.1, (J) Heterodina mosaica, (K) Sphaeroma walkeri, (L) Tylos marcuzzi.
Species richness was highest on the beaches of Champotón (7 species, sample coverage= 1), Sánchez Magallanes (6, s. c. = 0.89) and Puerto Morelos (6, s. c. = 0.29), and the lowest at Cayo Arcas and Chahuayxol with 1 species each (s. c. = 0.73 and 1, respectively) (Figure 5; Table 2). Abundance was greatest at Chahuayxol and Isla Blanca (together, 73% of the total), and the lowest at Barra de Tupilco (5 individuals, 0.15%), Boca Paila (4 individuals, 0.12%) and Ciudad del Carmen (3 individuals, 0.09%) (Figure 5). No macrocrustaceans were found at the Playa del Carmen (Figure 5). Most of the species had a restricted distribution, being observed in only one locality. In contrast, Excirolana brasiliensis was distributed in 9 of the 15 sampled localities and E. mayana was found in 7 localities (Figure 6).
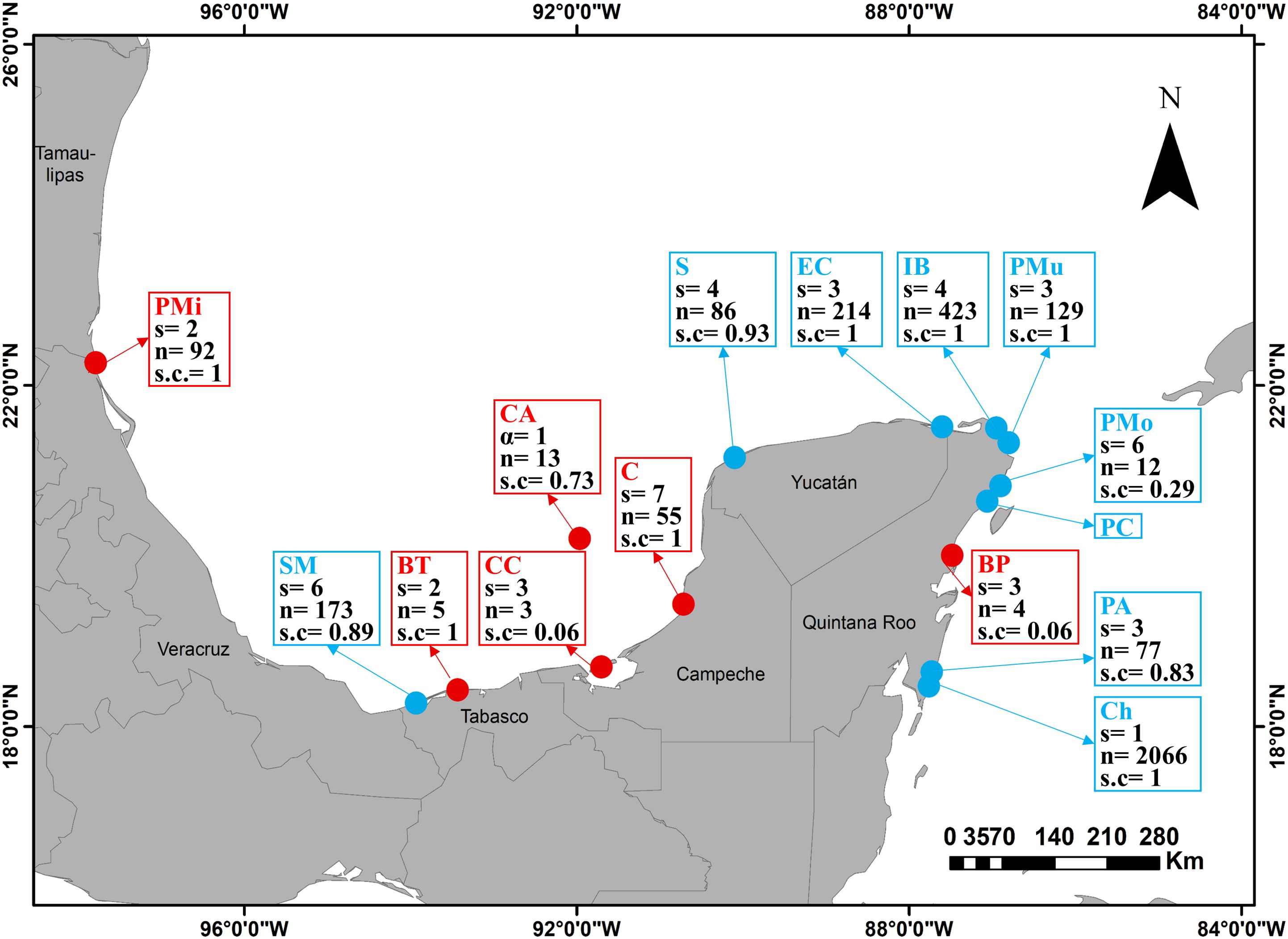
Figure 5. Species richness (s), abundance (n), and sample coverage (s. c.) of macrocrustaceans collected from 15 beaches on the eastern Mexican coast. Beaches localities acronyms can be read on Figure 1. Dot´s colors represent two different sampling protocols (further described in the materials and methods section, Figure 2).

Figure 6. Abundance and distribution of sampled macrocrustaceans by locality in the eastern Mexican coast. The color intensity of the rectangles represents the transformed abundance values (Log(X+1)) of each species/taxon per locality. The species are grouped based on a similarity analysis (Bray-Curtis) of their abundances. GM, Gulf of Mexico; CS, Caribbean Sea. The acronyms of the localities are in the caption of Figure 1.
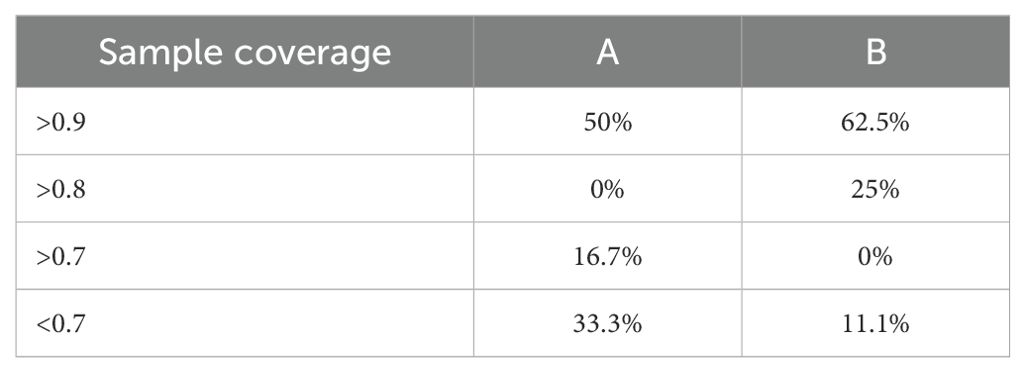
Table 2. Comparison of sampling designs A and B: percentage of localities by sampling coverage categories.
Sampling design B is more effective than sampling design A based on sampling coverage values (Table 2; Figure 7). The observed richness values were more similar to those extrapolated in most of the localities with sampling design B (36 sampling units) than in the localities with sampling design A (18 sampling units) (Figure 7A). Sample completeness exceeded 0.75 at 13 of the 16 localities, regardless of sampling effort (Figure 7B), i.e., a deficit in species detection of <25%. Among the three localities with sample completeness of <0.7, two represented sampling design A, and one represented design B; in these, species diversity could be much higher than that observed in the present study. Over 87% of localities with sampling design B had a sampling coverage greater than 0.8, while only half of the localities with sampling design A achieved a similar sampling coverage. The percentage of sites with lower sampling coverage (<0.7) is higher in sampling design A (Figure 7C). In contrast, the extrapolation curves of richness values and sampling coverage at the regional level do not reach the asymptote, suggesting that a greater number of beaches should be sampled at the regional scale (Figures 7D–F).
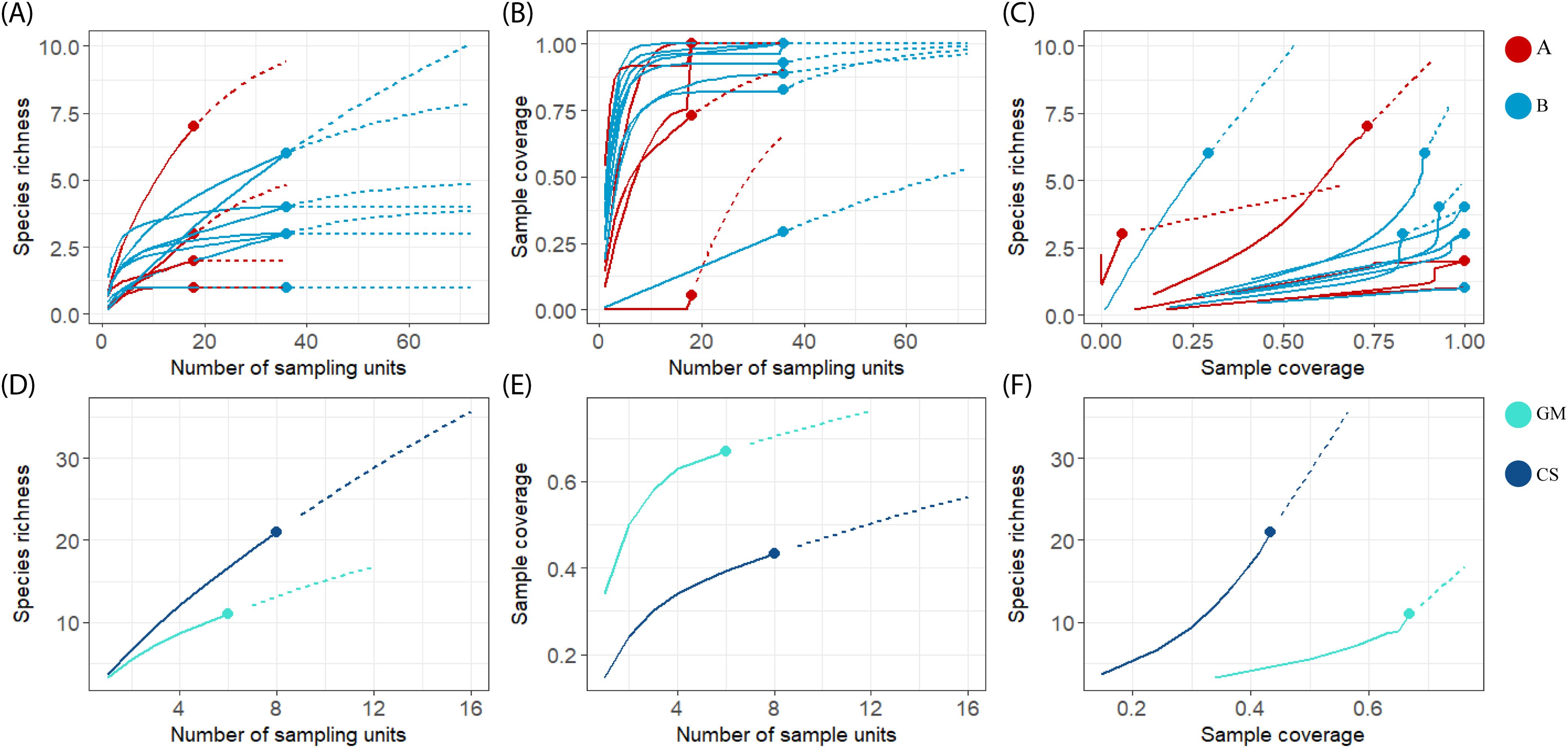
Figure 7. Interpolation and extrapolation of richness values and sample coverage (iNEXT Package from RStudio) of 15 localities from eastern Mexican coast with two different sampling designs (A and B, Figure 2), and two regions: GM, Gulf of Mexico and CS, Caribbean Sea. (A, D) Species accumulation curves based on the sample size and the incidence of species for each locality, (B, E) Sample-coverage accumulation curves based on incidence for each locality, (C, F) Sampled completeness curves.
Discussion
This study marks a significant milestone as it presents the first large-scale survey of macrobenthic crustaceans on sandy beaches along the eastern Mexican coast. Our findings, covering 15 beaches along almost 2400km of coastline, have expanded the documented richness to 70 species. This includes the discovery of eight species recorded for the first time in the region, with Heterodina mosaica (Kensley and Schotte, 1987), being a notable addition, extending its known range from Belize to the Gulf of Mexico (Kensley and Schotte, 1987). Other taxa, such as Haustorius jayneae Foster and Lecroy, 1991, Rhepoxynius epistomus (Shoemaker, 1938), Cassidinidea ovalis (Say, 1818), Exosphaeroma diminutum Menzies and Frankenberg, 1966, Sphaeroma walkeri Stebbing, 1905 and Americorchestia salomani Bousfield, 1991 are new records for the eastern Mexican coast and further underscore the rich diversity of these ecosystems. The inclusion of four additional species or taxa—Cerapus benthophilus Thomas and Heard, 1979, Parhyalella whelpleyi (Shoemaker, 1933), Excorallana berbicensis Boone, 1918, and an isopod of the family Munnidae—further enriches our understanding of sandy beach ecosystems in the eastern Mexican coast (Table 3). This compilation of historical and novel data establishes a crucial baseline for monitoring coastal biodiversity and assessing anthropogenic impacts, such as erosion, pollution, and the massive arrival of Sargassum. The observed increase in species diversity is particularly significant, given that previous studies had documented only 57 taxa across decades of intermittent sampling.
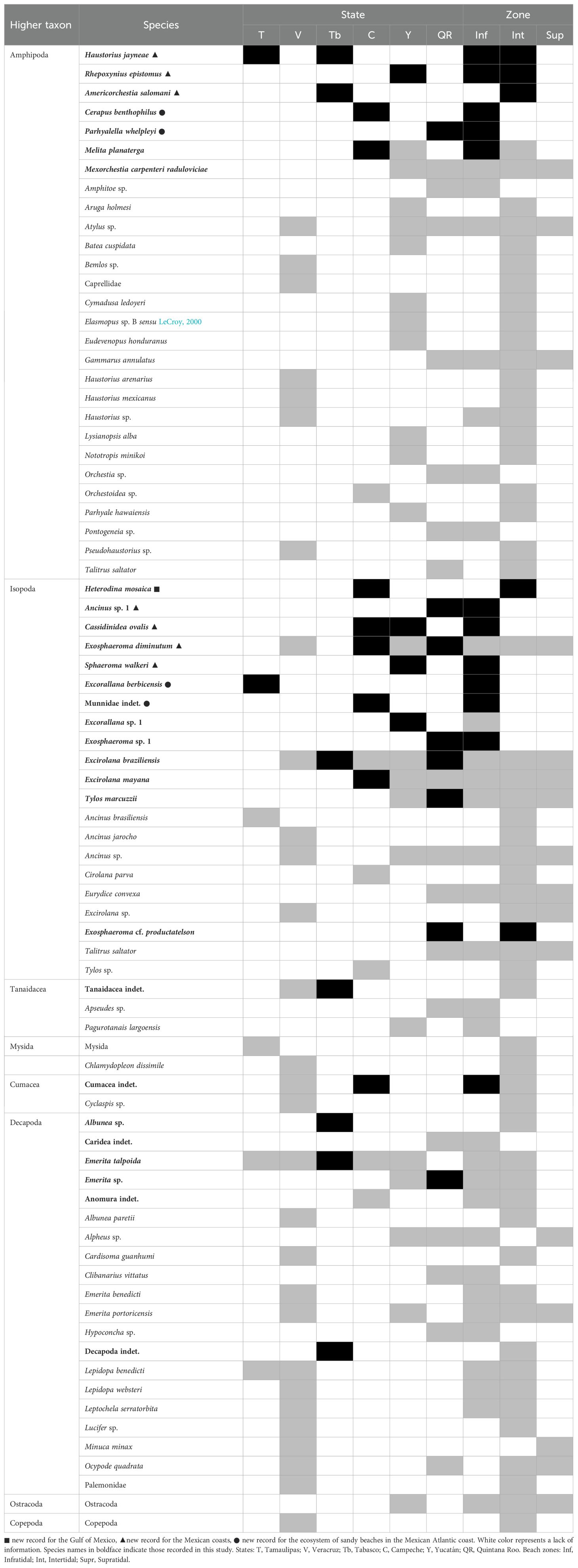
Table 3. Historically documented species/taxa (dark grey), including the new information provided in this study (black), on the sandy beaches from the eastern Mexican coast.
Ecological and biogeographical insights
In general, Excirolana braziliensis exhibited the widest distribution across beaches from Tabasco to Quintana Roo. However, the restricted distribution of other taxa, with 19 out of 24 species recorded at a single locality, underscores the influence of local environmental conditions and habitat specificity. For example, the amphipod Mexorchestia carpenteri raduloviciae was the most abundant species, accounting for 66.41% of individuals and primarly associated with Sargassum deposits on Quintana Roo’s beaches (Figure 6). The association of this talitrid amphipod with macroalgal wrack underscores the critical role of stranded algae in providing food and refuge, enhancing habitat complexity, and supporting high macrofaunal abundance (Parker et al., 2001; Tanaka and Leite, 2003; Ali et al., 2018).
From a regional perspective, the environmental transition between regions such as Campeche and Yucatán, characterized by a mix of calcareous sediments and diverse water masses, emerged as a macrocrustacean biodiversity hotspot (Escobar-Briones and Jiménez-Guadarrama, 2010). This finding aligns with biogeographical theories that highlight the importance of transition zones in fostering species richness by combining elements of neighboring ecoregions and hosting endemic species (Smith et al., 2001). However, besides the potential effect of this transitional region, the division of sampling into beach zones (supratidal, intertidal, infratidal) further expanded the ecological distribution records of species such as Melita planaterga Kunkel, 1910 and the order Cumacea, illustrating the relevance of different spatial scales in determining the distribution of species. Particularly, Melita planaterga and other Cumacea species, previously known for inhabiting intertidal zones (Paz-Ríos et al., 2013a; Hidalgo, 2017), were now observed in infratidal areas.
In addition, this study revealed potential cryptic diversity within some taxa. Specifically, morphological differences in the isopods Exosphaeroma sp., Excorallana sp., Exosphaeroma cf. productatelson, and Ancinus sp. compared to known species, suggest the need for molecular analyses to confirm their taxonomic status.
First, the isopod Exosphaeroma sp. found in this work in P. Morelos is very similar to E. antillensis. Both species have two broadly subconical protrusions on the pleotelson. However, Exosphaeroma sp. has a pleotelson with an entire posterior margin while E. antillensis has a pleotelson with an emarginate posterior margin. Furthermore, in the former the margins of the uropodal exopods and endopods are smooth, whereas in the latter these margins are crenulated (Kensley and Schotte, 1987). Second, the isopod belonging to the genus Excorallana sp. collected in Sisal is very similar to E. subtilis, previously documented in Florida (Boyko et al., 2019). However, Excorallana sp. has two subconical protuberances on the pleotelson that E. subtilis does not have. Third, Exosphaeroma cf. productatelson Menzies and Glynn, 1968 is distinguished by a fully crenulated outer margin of the uropodal exopod, and a frontal lamina that is equally long and wide. Fourth, two specimens (females) of the genus Ancinus found on the beach of Boca Paila (Caribbean Sea) were very similar to A. depressus (Say, 1818), a species reported from the northern Gulf of Mexico (Schotte et al., 2009). However, in Ancinus sp. the dorsal integument granules are much smaller and the two pairs of antennae are also smaller than in A. depressus: antenna 1 does not extend beyond the fourth thoracic segment and antenna 2 does not extend beyond the third thoracic segment.
Such taxonomic identification efforts of potential new species from sandy beaches would enhance our understanding of species richness and evolutionary processes in these underexplored ecosystems (Kensley and Schotte, 1987; Boyko et al., 2019).
Methodological insights and conservation implications
The state-level disparities in faunal knowledge highlight the importance of localized conservation efforts informed by robust baseline data (Figure 8A). For instance, the increased species richness documented in Campeche and Quintana Roo emphasizes the need for targeted protection of these biodiversity hotspots. Conversely, the limited data available for Tabasco and Tamaulipas call for prioritizing these regions in future research and conservation planning. Much of the knowledge about macrocrustaceans on Mexican beaches along the eastern coast has been generated in recent years, and the present study represents a significant contribution (Figure 8B). However, there is still a considerable information gap for many Mexican beaches. This is also the case for beaches on the western Mexican coast (Pacific) where only three studies have documented macrocrustaceans of sandy beaches (Dexter, 1976; Torres and Lowry, 2011; Torres-Alfaro et al., 2012). Given the regional ecological and economic importance of sandy beaches, integrating this biodiversity data into state and national coastal management strategies is essential. The deposition of specimens in public collections and the open availability of data through the Ocean Biodiversity Information System (OBIS) ensure that this study’s findings can support evidence-based decision-making and foster collaboration among stakeholders.
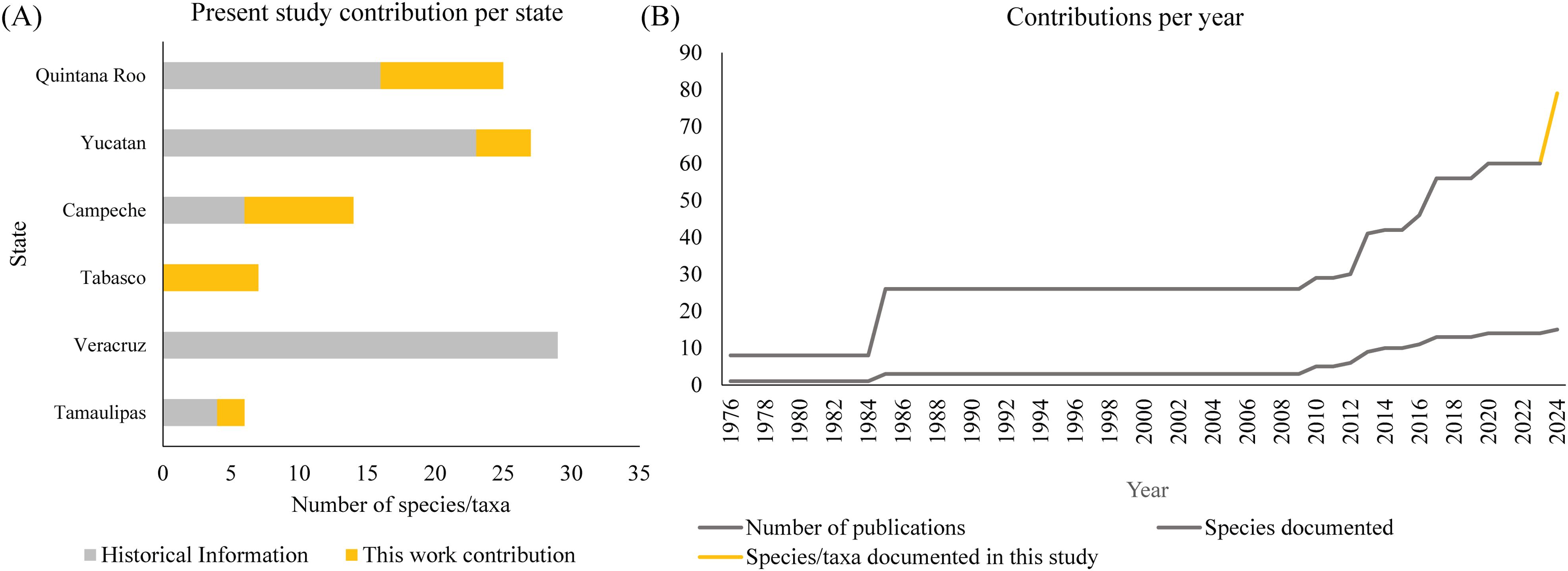
Figure 8. (A) State of knowledge per year of macrocrustaceans species/taxa associated with the sandy beaches from the easter Mexican coast. (B) Contribution of the macrocrustacean species/taxa documented in this study on the sandy beaches of the eastern Mexican coast. Data obtained from Dexter, 1976; Arriaga-Becerra, 1985; Méndez-Ubach et al., 1985; Rocha-Ramírez et al., 2010, 2016; Pacheco-Ríos, 2010; Miranda, 2012; Martínez, 2013; Paz-Ríos et al., 2013a; Wildish and Lecroy, 2014; González et al., 2017; Hidalgo, 2017; Guerra-Castro et al., 2020.
In addition to the field work carried out in this study, this work is also the first effort to compile faunal data on macrocrustaceans in the sandy beaches of the Mexican coast of the Gulf of Mexico and the Caribbean Sea and their distribution in three zones of the beach (infratidal, intertidal and supratidal) (Table 3). Of 14 publications traced on this topic, nine are scientific articles, four are undergraduate theses and one is a doctoral thesis. The first published record on the macrofauna of sandy beaches along these coasts (Dexter, 1976) reported eight benthic macrocrustaceans from Tamaulipas, Veracruz and Campeche. A further 18 species were added (Arriaga-Becerra, 1985; Méndez-Ubach et al., 1985) followed by hiatus until information regarding macrocrustaceans was contributed by studies in Veracruz (Pacheco-Ríos, 2010; Rocha-Ramírez et al., 2010, 2016; Miranda, 2012; Martínez, 2013; Hidalgo, 2017), Yucatán (Paz-Ríos et al., 2013a; Guerra-Castro et al., 2020) and Quintana Roo (Wildish and Lecroy, 2014; González et al., 2017): this gradually increased the species/taxa total to 59, to which the present findings have added a further 19 species/taxa (Table 3; Figure 8).
From a methodological perspective, using a core sampling method and comparing two designs at the local scale provided valuable insights into sampling efficiency (Figure 7). Design B, involving 36 cores per locality, achieved a balance between observed and extrapolated richness, making it the preferred approach for regional-scale studies. However, including additional techniques, such as baited traps and manual collections, would likely enhance the detection of elusive or cryptic species. For example, Exosphaeroma sp., collected in P. Morelos, and Excorallana sp., found in Sisal, presented morphological traits distinct from those in existing taxonomic keys, underscoring the potential for discovering cryptic or undescribed species (Kensley and Schotte, 1987; Munilla et al., 1998; Beyst et al., 2001; Boyko et al., 2019).
Temporal variability remains an essential but unexplored aspect of this study. Seasonal changes in beach morphology and associated fauna have significantly influenced diversity and abundance patterns (McLachlan and Defeo, 2018). Addressing this limitation would require long-term monitoring, which, while resource-intensive, is essential for understanding the temporal dynamics of sandy beach ecosystems. Future studies could adopt a hierarchical approach, balancing spatial and temporal scales to identify the key processes shaping biodiversity patterns. The geographic extension of this study allowed for the identification of significant spatial variations in species richness and composition. This spatial pattern highlights the need for future studies to harmonize sampling efforts across all regions of the eastern Mexican coast.
While this study provides a foundational understanding of macrobenthic crustacean diversity, several avenues for future research emerge. Expanding the geographic scope to include historically well-sampled regions like Veracruz and extending temporal coverage to capture seasonal dynamics are critical next steps. Incorporating environmental variables, such as sediment grain size, organic content, and hydrodynamic conditions, would enhance the explanatory power of biodiversity patterns observed in this study.
Additionally, advancing molecular techniques could resolve taxonomic ambiguities and uncover hidden diversity within cryptic taxa. Such approaches would complement traditional morphology-based identifications and provide a more comprehensive picture of biodiversity. Collaborative efforts across disciplines, integrating ecological, genetic, and oceanographic data, hold promise for addressing the complex challenges facing sandy beach ecosystems.
This study marks a significant milestone in understanding the diversity of macrobenthic crustaceans along the Mexican Atlantic coast. Integrating historical and novel data establishes a comprehensive baseline for future research and conservation. The insights gained from this sampling effort provide a roadmap for optimizing biodiversity assessments in sandy beach ecosystems in Mexico. As coastal environments face increasing human activities and climate change pressures, this work underscores the urgency of protecting these vulnerable yet invaluable habitats. We take a crucial step toward sustainable management and preservation by fostering a deeper appreciation for their biodiversity and ecological roles.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://doi.org/10.15468/zfgt5w.
Ethics statement
The manuscript presents research on animals that do not require ethical approval for their study.
Author contributions
MM: Data curation, Formal analysis, Investigation, Project administration, Visualization, Writing – original draft, Writing – review & editing. EG: Conceptualization, Formal analysis, Funding acquisition, Methodology, Resources, Validation, Writing – review & editing. RC: Investigation, Writing – review & editing. CP: Conceptualization, Methodology, Validation, Writing – review & editing. NS: Conceptualization, Funding acquisition, Methodology, Resources, Supervision, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. Financing was granted by a Consejo Nacional de Humanidades, Ciencia y Tecnología graduate scholarship (CVU: 630719), Harte Research Institute, the Harte Charitable Foundation, Texas A&M University and the PAPIIT IV 300123 project “Resilience of Coastal Socie-ecosystems Associated with Tourism in the Face of COVID-19 and Climate Change” conducted by the Laboratorio Nacional de Resiliencia Costera.
Acknowledgments
This paper is part of the requirements for obtaining a Doctoral degree at the Posgrado en Ciencias Biológicas, Universidad Nacional Autónoma de México. We are very thankful to M. en C. Johnny Omar Valdez Iuit who helped during field trips and data collection. We acknowledge the help of Dra. Patricia Guadarrama Chávez and Lic. Camilo Rendón Valdez (UMDI-Facultad de Ciencias and LIPC-UNAM) who provided technical support. We especially thank our doctoral thesis advisors, Dra. Elva Guadalupe Escobar Briones and Dr. Adolfo Gracia Gasca, for their guidance and support throughout this research.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2025.1514137/full#supplementary-material
References
Abele L. G., Kim W. (1986). An Illustrated Guide to the Marine Decapod Crustaceans of Florida (Florida: Technical Series of the Satate of Florida Department of Environmental Regulation).
Ahyong S. T., Lowry J. K., Alonso M., Bamber R. N., Boxshall G. A., Castro P., et al. (2011). “Subphylum crustacea Brünnich, 1772,” in Animal biodiversity: an outline of higher-level classification and survey of taxonomic richness. Ed. Z. Z. Q. vol. 3148, 165–191. doi: 10.11646/zootaxa.3148.1.33
Ali M., Al-Ghunaim A., Subrahmanyam M. N. V., Al-Enezi Y., Al-Said T., Al-Zakri W., et al. (2018). On the diversity of amphipods inhabiting sargassum as well as clear areas in Kuwait coastal waters, with an assessment of the effect of turbidity and notes on their abundance, composition, and distribution: A preliminary study. Crustaceana 91, 767–819. doi: 10.1163/15685403-00003799
Arriaga-Becerra R. E. (1985). Estudio preliminar de la macrofuana de los invertebrados de las playas arenosas de Quintana Roo y Yucatán (México) y su relación con el sedimento. (thesis). Mexico City (MEX): Universidad Nacional Autónoma de México.
Barca-Bravo S., Servia M. J., Cobo F., Gonzalez M. A. (2008). The effect of human use of sandy beaches on developmental stability of Talitrus saltator (Montagu 1808) (Crustacea, Amphipoda). A study on fluctuating asymmetry. Mar. Ecol. 29, 91–98. doi: 10.1111/j.1439-0485.2007.00208.x
Bergamino L., Gómez J., Barboza F. R., Lercari D. (2013). Major food web properties of two sandy beaches with contrasting morphodynamics, and effects on the stability. Aquat. Ecol. 47, 253–261. doi: 10.1007/s10452-013-9440-5
Bessa F., Gonçalves S. C., Franco J. N., André J. N., Cunha P. P., Marques J. C. (2014). Temporal changes in macrofauna as response indicator to potential human pressures on sandy beaches. Ecol. Indic. 41, 49–57. doi: 10.1016/j.ecolind.2014.01.023
Beyst B., Buysse D., Dewicke A., Mees J. (2001). Surf zone hyperbenthos of Belgian sandy beaches: Seasonal patterns. Estuar. Coast. Shelf Sci. 53, 877–895. doi: 10.1006/ecss.2001.0808
Bocher P., Cherel Y., Labat J. P., Mayzaud P., Razouls S., Jouventin P. (2001). Amphipod-based food web: Themisto gaudichaudii caught in nets and by seabirds in Kerguelen waters, southern Indian Ocean. Mar. Ecol. Prog. Ser. 223, 261–276. doi: 10.3354/meps223261
Bousfield E. L. (1991). New sandhoppers (Crustacea: Amphipoda) from the gulf coast of the united states. Gulf Res. Rep. 8, 271–283.
Boyko C., Bruce N. L., Hadfield K. A., Merrin K. L., Ota Y., Poore G. C. B., et al. (2019). WoRMS Isopoda: World Marine, Freshwater and Terrestrial Isopod Crustaceans database (version 2019-03-05). Available online at: http://marinespecies.org/isopoda/aphia.php?p=taxdetails&id=260377 (Accessed April 11, 2023).
Brown A. C. (1996). Behavioural plasticity as a key factor in the survival and evolution of the macrofauna on exposed sandy beaches. Rev. Chil. Hist. Nat. 69, 469–474.
Cardoso R. S., Barboza C. A. M., Skinner V. B., Cabrini T. M. B. (2016). Crustaceans as ecological indicators of metropolitan sandy beaches health. Ecol. Indic. 62, 154–162. doi: 10.1016/j.ecolind.2015.11.039
Chao A., Gotelli N. J., Hsieh T. C., Sander E. L., Ma K. H., Colwell R. K. (2014). Rarefaction and extrapolation with Hill numbers: A framework for sampling and estimation in species diversity studies. Ecol. Monogr. 84, 45–67. doi: 10.1890/13-0133.1
Clarke K. R., Tweedley J. R., Valesini F. J. (2014). Simple shade plots aid better long-term choices of data pre-treatment in multivariate assemblage studies. J. Mar. Biol. Assoc. United Kingdom 94, 1–16. doi: 10.1017/S0025315413001227
Defeo O., McLachlan A., Schoeman D. S., Schlacher T. A., Dugan J., Jones A., et al. (2009). Threats to sandy beach ecosystems: A review. Estuar. Coast. Shelf Sci. 81, 1–12. doi: 10.1016/j.ecss.2008.09.022
de la Lanza-Espino G., Ortiz-Pérez M. A., Pérez-Carbajal J. L. (2013). Diferenciación hidrogeomorfológica de los ambientes costeros del Pacífico, del Golfo de México y del Mar Caribe. Investig. Geogr. 81, 33–50. doi: 10.14350/rig.33375
Escobar-Briones E. G., Jiménez-Guadarrama E. L. (2010). Macrocrustáceos (Peracarida, Decapoda) de fondos carbonatados del sector occidental del Banco de Campeche en el sur del Golfo de México. Rev. Mex. Biodivers. 81, S63–S72. doi: 10.22201/ib.20078706e.2010.0.226
Fanini L., Cantarino C. M., Scapini F. (2005). Relationships between the dynamics of two Talitrus saltator populations and the impacts of activities linked to tourism. Oceanologia 47, 93–112.
Foster J. M., Lecroy S. E. (1991). Haustorius jayneae, a new species of haustoriid amphipod from the northern gulf of mexico, with notes on its ecology at panama city beach, florida U.S.A. Gulf Res. Rep. 8, 259–270.
Giordani Soika A. (1954). Studi di ecologia e biogeografia 12. ecologia, sistematica, biogeografia ed evoluzione del tylos latreillei auct. (Isop. tylidae). Boll. del Mus. Civ. di Stor. Nat. di Venezia 7, 63–83.
Gonçalves S. C., Anastácio P. M., Marques J. C. (2013). Talitrid and Tylid crustaceans bioecology as a tool to monitor and assess sandy beaches’ ecological quality condition. Ecol. Indic. 29, 549–557. doi: 10.1016/j.ecolind.2013.01.035
González A., Torruco D., Torruco Á.D. (2017). The sandy beaches of Quintana Roo, Mexico: The intertidal fauna diversity. Rev. Biol. Mar. Oceanogr. 52, 361–373. doi: 10.4067/s0718-19572017000200015
Guerra-Castro E., Hidalgo G., Castillo-Cupul R. E., Muciño-Reyes M., Noreña-Barroso E., Quiroz-Deaquino J., et al. (2020). Sandy beach macrofauna of yucatán state (Mexico) and oil industry development in the gulf of Mexico: first approach for detecting environmental impacts. Front. Mar. Sci. 7. doi: 10.3389/fmars.2020.589656
Hidalgo G. (2017). Cominidades intermareales de la macrofauna en playas arenosas del litoral central de Veracruz (Golfo de México: Un enfoque de integridad ecológica. Universidad Veracruzana). Available at: https://cdigital.uv.mx/bitstream/handle/123456789/46427/QuirozCortesMCarmen.pdf?sequence=2&isAllowed=y (Accessed November 9, 2020).
Hsieh T. C., Ma K. H., Chao A. (2016). iNEXT: an R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods Ecol. Evol. 7, 1451–1456. doi: 10.1111/2041-210X.12613
Hsueh P. W. (2015). A new species of Emerita (Decapoda, Anomura, Hippidae) from Taiwan, with a key to species of the genus. Crustaceana 88, 247–258. doi: 10.1163/15685403-00003413
INEGI (2022). Anuario Estadístico y Geográfico de los Estados Unidos Mexicanos 2021. Available online at: https://www.inegi.org.mx/contenidos/productos/prod_serv/contenidos/espanol/bvinegi/productos/nueva_estruc/889463904359.pdf (Accessed July 4, 2023).
Ives J. E. (1891). Crustacea from the northern coast of yucatan, the harbor of vera cruz, the west coast of florida and the bermuda islands. Proc. Acad. Nat. Sci. Philadelphia 43, 176–207.
Jernelöv A., Lindén O. (2010). Ixtoc I: A Case Study of the World ‘ s Largest Oil Spill. Ambio 10, 299–306. Available at: http://www.jstor.org/stable/4312725 (Accessed January 3, 2019).
Joye S. B. (2016). The Gulf of Mexico ecosystem - Before, during and after the Deepwater Horizon oil well blowout. Deep. Res. Part II Top. Stud. Oceanogr. 129, 1. doi: 10.1016/j.dsr2.2016.04.022
Kensley B., Schotte M. (1987). New records of isopod crustacea from the Caribbean, the Florida Keys, and the Bahamas. Proc. Biol. Soc Washingt. 100, 216–247.
LeCroy S. E. (2000). An Illustrated Guide to the Nearshore Marine and Estuarine Gammaridean Amphipoda of Florida. Volume 1. Families Gammaridae, Hadziidae, Isaeidae, Melitidae and Oedicerotidae (Tallahassee, Florida: Florida Department of Environmental Protection).
LeCroy S. E. (2002). An Illustrated Identification Guide to the Nearshore Marine and Estuarine Gammaridean Amphipoda of Florida. Volume 2. Families Ampeliscidae, Amphilochidae, Ampithoidae, Aoridae, Argissidae and Haustoriidae. (Tallahassee, Florida: Florida Department of Environmental Protection).
LeCroy S. E. (2004). An Illustrated Identification Guide to the Nearshore Marine and Estuarine Gmmaridean Amphipoda of Florida. Volume 3. Families Bateidae, Biancolinidae, Cheluridae, Colomastigidae, Corophiidae, Cyproideidae and Dexaminidae. (Tallahassee, Florida: Florida Department of Environmental Protection).
LeCroy S. E. (2007). An Illustrated Identification Guide To the Nearshore Marine and Estuarine Gammaridean Amphipoda of Florida. Volume 4. Families Anamixidae, Eusiridae, Hyalellidae, Hyalidae, Iphimediidae, Ischyroceridae, Lysianassidae, Megaluropidae And Melphidippidae. (Tallahassee, Florida: Florida Department of Environmental Protection).
LeCroy S. E. (2011). An Illustrated Identification Guide To the Nearshore Marine and Estuarine Gammaridean Amphipoda of Florida (Tallahassee, Florida: Florida Department of Environmental Protection).
Lercari D., Defeo O. (1999). Effects of freshwater discharge in sandy beach populations: The mole crab Emerita brasiliensis in Uruguay. Estuar. Coast. Shelf Sci. 49, 457–468. doi: 10.1006/ecss.1999.0521
Lowry J. K., Myers A. A. (2017). A phylogeny and classification of the amphipoda with the establishment of the new order Ingolfiellida (Crustacea: Peracarida). Zootaxa 4265, 1–89. doi: 10.11646/zootaxa.4265.1.1
Martínez M. G. (2013). Morfodinámica y variación espacio-temporal de la fauna macrobentónica de cuatro playas arenosas de Veracruz, México (thesis). (Tlalnepantla (MEX): Universidad Nacional Autónoma de México).
Martínez M. L., Intralawan A., Vázquez G., Pérez-Maqueo O., Sutton P., Landgrave R. (2007). The coasts of our world: Ecological, economic and social importance. Ecol. Econ. 63, 254–272. doi: 10.1016/j.ecolecon.2006.10.022
Méndez-Ubach M. N., Solis-Weiss V., Carranza-Edwards A. (1985). La importancia de la ganulometría en la distribución de organismos bentónicos. Estudio de playas del estado de Veracruz, México. An. del Inst. Cienc. del Mar. y Limnol. 13, 45–56.
Menzies R. J., Frankenberg D. (1966). Handbook on the common marine isopod crustacea of georgia (Athens, Georgia: University of Georgia Press).
Menzies R. J., Glynn P. W. (1968). The common marine isopod crustacea of puerto rico: A handbook for marine biologists. Stud. Fauna Curacao other Caribb. Islands 27, 1–133.
Miranda M. (2012). Estructura de las comunidades de macroinvertebrados intermareales en las playas del centro de Veracruz. (Thesis). Estado de México (MEX): Universidad Nacional Autónoma de México.
Muciño-Reyes M. D. R., Simoes N., Guerra-Castro E., Castillo-Cupul R. E., Paz-Ríos C. E. (2024). Data from: Community of macrocrustaceans from the Mexican sandy beaches of the Gulf of México and Caribbean Sea collected by the National Autonomous University of Mexico (UNAM). Version 1.5. Caribb. OBIS Node. Occur. dataset. doi: 10.15468/zfgt5w
Munilla T., Corrales M. J., San Vicente C. (1998). Suprabenthic assemblages from Catalan beaches: zoological groups. Orsis 13, 67–78. Available at: http://ddd.uab.es/pub/orsis/02134039v13p67.pdf%5Cnhttp://ddd.uab.cat/pub/orsis/02134039v13p67.pdf%5Cnhttp://www.raco.cat/index.php/Orsis/article/viewFile/24411/24245.
Noriega R., Schlacher T. A., Smeuninx B. (2012). Reductions in ghost crab populations reflect urbanization of beaches and dunes. J. Coast. Res. 28, 123–131. doi: 10.2112/JCOASTRES-D-09-00173.1
Nourisson D. H., Bessa F., Scapini F., Marques J. C. (2014). Macrofaunal community abundance and diversity and talitrid orientation as potential indicators of ecological long-term effects of a sand-dune recovery intervention. Ecol. Indic. 36, 356–366. doi: 10.1016/j.ecolind.2013.08.005
Oliva-Rivera J. J. (2003). The amphipod fauna of Banco Chinchorro, Quintana Roo, Mexico with ecological notes. Bull. Mar. Sci. 73, 77–89.
Ortíz M., Winfield I., Cházaro-Olvera S., López-Del Río B., Lozano-Aburto M.Á. (2013). Isópodos (Crustacea: Peracarida) del Área Natural Protegida Arrecife Tuxpan-Lobos, Veracruz, México: lista de especies y registros nuevos. Novit. Caribaea. 6, 63–75. doi: 10.33800/nc.v0i6.108
Pacheco-Ríos S. (2010). Patrones de distribución de la macroinfauna en cuatro playas arenosas del Estado de Veracruz, México. (thesis). (Tlalnepantla (MEX): Universidad Nacional Autónoma de México).
Parker J. D., Duffy J. E., Orth R. J. (2001). Plant species diversity and composition: Experimental effects on marine epifaunal assemblages. Mar. Ecol. Prog. Ser. 224, 55–67. doi: 10.3354/meps224055
Paz-Ríos C. E., Simões N., Ardisson P. (2013a). Intertidal and shallow water amphipods (Amphipoda : Gammaridea and Corophiidea) from Isla Pérez, Alacranes Reef, southern Gulf of Mexico. Nauplius 21, 179–194. doi: 10.1590/S0104-64972013000200005
Paz-Ríos C. E., Simões N., Ardisson P. (2013b). Records and observations of amphipods (Amphipoda : Gammaridea and Corophiidea) from fouling assemblages in the Alacranes Reef, southern Gulf of Mexico. Mar. Biodivers. Rec. 6, 1–16. doi: 10.1017/S175526721300064X
Paz-Ríos C. E., Simões N., Pech D. (2019). Species richness and spatial distribution of benthic amphipods (Crustacea: Peracarida) in the Alacranes Reef National Park, Gulf of Mexico. Mar. Biodivers. 49, 673–682. doi: 10.1007/s12526-017-0843-8
Raz-Guzmán A., Soto L. A. (2017). Updated checklist and zoogeographic remarks of benthic amphipods (Crustacea: Peracarida: Amphipoda) of two coastal lagoons in the western Gulf of Mexico. Rev. Mex. Biodivers. 88, 715–734. doi: 10.1016/j.rmb.2017.07.002
Richardson H. (1912). Descriptions of a new genus of isopod crustaceans, and of two new species from south america. Proc. United States Natl. Museum 43, 201–204.
Rocha-Ramírez A., Chávez-López R., Antillón-Zaragoza I., Fuentes-Mendoza F. A. (2016). Variación nictemeral de los ensamblajes de macrocrustáceos en una playa arenosa del centro-norte de Veracruz, México. Rev. Mex. Biodivers. 87, 92–100. doi: 10.1016/j.rmb.2016.01.025
Rocha-Ramírez A., Chávez-López R., Peláez-Rodríguez E. (2010). Ancinus jarocho (Isopoda: Sphaeromatidea: Ancinidae), a new species from the central Gulf of Mexico, Mexico. Zootaxa. 2397, 61–68. doi: 10.11646/zootaxa.2397.1.6
Rodríguez-Martínez R. E., Medina-Valmaseda A. E., Blanchon P., Monroy-Velázquez L. V., Almazán-Becerril A., Delgado-Pech B., et al. (2019). Faunal mortality associated with massive beaching and decomposition of pelagic Sargassum. Mar. pollut. Bull. 146, 201–205. doi: 10.1016/j.marpolbul.2019.06.015
Say T. (1818). An account of the crustacea of the united states. J. Acad. Nat. Sci. Philadelphia 1, 423–441.
Scapini F. (2006). Keynote papers on sandhopper orientation and navigation. Mar. Freshw. Behav. Physiol. 39, 73–85. doi: 10.1080/10236240600563412
Schlacher T. A., Schoeman D. S., Dugan J., Lastra M., Jones A., Scapini F., et al. (2008). Sandy beach ecosystems: Key features, sampling issues, management challenges and climate change impacts. Mar. Ecol. 29, 70–90. doi: 10.1111/j.1439-0485.2007.00204.x
Schotte M., Markham J. C., Wilson G. D. F. (2009). “Isopoda (Crustacea) of the Gulf of Mexico,” in Gulf of Mexico-Origins, Waters, and Biota. Eds. Felder D. L., Camp D. K. (Texas A&M Press, College Station, Texas), 973–986.
Silva-Casarín R., Lacouture M., Durón F., Pedroza-Paez D., Pérez M. A., Baldwin E., et al. (2011). Caracterización de la zona costera y planteamiento de elementos técnicos para la elaboración de criterios de regulación y manejo sustentable. México: UNAM-SEMARNAT.
Shoemaker C. R. (1938). Two new species of amphipod crustaceans from the east coast of the united states. J. Washingt. Acad. Sci. 28, 326–332.
Smith T. B., Kark S., Schneider C. J., Wayne R. K., Moritz C. (2001). Biodiversity hotspots and beyond: The need for preserving environmental transitions [1. Trends Ecol. Evol. 16, 431. doi: 10.1016/S0169-5347(01)02201-7
Stebbing T. R. R. (1905). Report on the isopoda collected by professor herdman at ceylon, in 1902. Rep. to Gov. Ceylon Pearl Oyster Fish. Gulf Manaar Suppl. Rep. 4, 1–12.
Sun S., Hu C., Tunnell J. W. (2015). Surface oil footprint and trajectory of the Ixtoc-I oil spill determined from Landsat/MSS and CZCS observations. Mar. pollut. Bull. 101, 632–641. doi: 10.1016/j.marpolbul.2015.10.036
Tanaka M. O., Leite F. P. P. (2003). Spatial scaling in the distribution of macrofauna associated with Sargassum stenophyllum (Mertens) Martius: Analyses of faunal groups, gammarid life habits, and assemblage structure. J. Exp. Mar. Bio. Ecol. 293, 1–22. doi: 10.1016/S0022-0981(03)00233-8
Thomas J. D., Heard R. (1979). A new species of cerapus sa (Crustacea:Amphipoda) from the northern gulf of mexico with notes on its ecology.
Torres G., Lowry J. (2011). “Peracarids from three low-energy fine-sand beaches of Mexico: western coast of Gulf of California,” in Crustaceana Monographs. (Brill Academic Publishers). 15, 311–324. doi: 10.1163/ej.9789004174252.i-354.207 (Accessed January 03, 2019).
Torres-Alfaro G. M., López-Olmos J. R., De Cruz-Agüero G. (2012). Crustáceos peracáridos de las playas arenosas de la Bahía de la Paz, Baja California Sur, México, durante 2001 a 2012. Cienc. y Mar. XVI, 37–42.
Wildish D. J., Lecroy S. E. (2014). Mexorchestia: A new genus of talitrid amphipod (Crustacea, Amphipoda, Talitridae) from the Gulf of Mexico and Caribbean Sea, with the description of a new species and two new subspecies. Zootaxa 3856, 555–577. doi: 10.11646/zootaxa.3856.4.5
Wilkinson T., Wiken E., Creel J., Hourigan T., Agardy T., Herrmann H., et al. (2009). Marine ecoregions of north america (Montreal, Canada: Commission for Environmental Cooperation).
Winfield I., Abarca-Arenas L., Cházaro-Olvera S. (2007). Crustacean macrofoulers in the Veracruz coral reef system, SW Gulf of Mexico: checklist, spatial distribution and diversity. Cah. Biol. Mar. 48, 287–295.
Winfield I., Abarca-ávila M., Ortiz M., Lozano-Aburto M.Á. (2013). Tanaidaceans (Peracarida: Tanaidacea) from the National Park Veracruzan Coral Reef System: Checklist and new records. Rev. Mex. Biodivers. 84, 471–479. doi: 10.7550/rmb.32499
Winfield I., Cházaro-Olvera S., Horta-Puga G., Lozano-Aburto M.Á., Arenas-Fuentes V. (2010). Macrocrustáceos incrustantes en el Parque Nacional Sistema Arrecifal Veracruzano: Biodiversidad, abundancia y distribución. Rev. Mex. Biodivers. 81, 165–175. doi: 10.22201/ib.20078706e.2010.0.219
Winfield I., Escobar-Briones E. (2007). Anfípodos (Crustacea : Gammaridea) del sector norte del Mar Caribe : listado faunístico, registros nuevos y distribución espacial. Rev. Mex. Biodivers. 78, 51–61. doi: 10.22201/ib.20078706e.2007.001.411
Winfield I., Ortiz M. (1996). Riqueza específica de crustáceos peracáridos en el Complejo Lagunar de Alvarado, Veracruz. Rev. Zool. 2, 57–68.
Keywords: benthic fauna, crustaceans, Caribbean Sea, Gulf of Mexico, intertidal, infratidal
Citation: Muciño-Reyes MR, Guerra-Castro E, Castillo-Cupul RE, Paz-Ríos CE and Simões N (2025) Unveiling the diversity of macrobenthic crustaceans on sandy beaches of the eastern Mexican coast: new records and an updated checklist. Front. Mar. Sci. 12:1514137. doi: 10.3389/fmars.2025.1514137
Received: 20 October 2024; Accepted: 13 January 2025;
Published: 07 February 2025.
Edited by:
Jin Sun, Ocean University of China, ChinaReviewed by:
Aisha Belal, National Institute of Oceanography and Fisheries, EgyptEmanuel Pereira, University of Buenos Aires, Argentina
Luz Verónica Monroy Velázquez, National Autonomous University of Mexico, Mexico
Copyright © 2025 Muciño-Reyes, Guerra-Castro, Castillo-Cupul, Paz-Ríos and Simões. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Nuno Simões, bnNAY2llbmNpYXMudW5hbS5teA==