
94% of researchers rate our articles as excellent or good
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.
Find out more
ORIGINAL RESEARCH article
Front. Mar. Sci., 05 March 2025
Sec. Marine Megafauna
Volume 12 - 2025 | https://doi.org/10.3389/fmars.2025.1474460
 Martin J. Baptist1,2*
Martin J. Baptist1,2* Mardik F. Leopold1
Mardik F. Leopold1 Marinka E. B. van Puijenbroek1
Marinka E. B. van Puijenbroek1 Hans Verdaat1
Hans Verdaat1 Nicole Janinhoff-Verdaat1Dennis R. Lammertsma3
Nicole Janinhoff-Verdaat1Dennis R. Lammertsma3 Wim J. Dimmers3Paul F. A. M. Römkens3
Wim J. Dimmers3Paul F. A. M. Römkens3 Jan Burgers3
Jan Burgers3Due to the benefits to the ecosystem there is a growing incentive to limit management interventions of stranded whales. This study was conducted to test the impact of a stranded whale carcass decomposing in a natural dune ecosystem and to gain experience in managing a beached whale carcass. A 4.70 m male juvenile minke whale Balaenoptera acutorostrata carcass washed up on the western side of the uninhabited island Rottumerplaat, the Netherlands. The carcass was moved the same night into the dunes to prevent it from washing back into the sea. Regular field visits were undertaken and wildlife trail cameras were placed to document the decomposition stages and the attraction of avifauna. Pitfall traps were placed to monitor insect fauna next to the carcass. Vegetation plots were surveyed to document changes to the dune vegetation and soil cores were taken to determine the increase in metal, carbon and nutrient contents. Decomposition of the carcass until the bare skeleton within the temperate dune ecosystem took about two years. The carcass was highly beneficial for beetle biodiversity, attracting different beetle species at various stages of decomposition. In total, 129 species of beetles were found near the carcass, of which eight species are specific for carcasses. For scavenging birds such as gulls, magpies and carrion crows the skin of the minke whale was too tough to break open. While the initial nutrient leakage from the carcass was toxic to dune plants immediately around the carcass, it eventually promoted lush vegetation growth due to increased fertility. The bioavailable metal contents in the soil for potassium (K) and nickel (Ni) showed a significant increase due to the presence of the whale carcass up to six months. Significantly increased levels of arsenic (As), cobalt (Co) and vanadium (V) were found up to one and a half years later and sodium (Na) and manganese (Mn) persisted in elevated levels up to two years later. Recommendations were given on management interventions when leaving a whale carcass on site.
Mammal carcasses play a significant ecological role in ecosystems by serving as nutrient-rich resources that support diverse communities of scavengers and decomposers (Barton et al., 2013; Olea et al., 2019; Benbow et al., 2020; van Klink et al., 2020). Upon the death of a mammal, its carcass undergoes a series of ecological processes that not only recycle nutrients but also shape the dynamics of local ecosystems. Initially, scavengers such as carnivores, birds of prey, and scavenging invertebrates are attracted to the carcass and feed on the nutrient-rich tissues. This serves as a pathway for energy transfer within the various parts of the food-web of an ecosystem (Wilson and Wolkovich, 2011). Carcass size, location and season influence the richness and composition of vertebrate scavenger communities (Turner et al., 2017). As the carcass decomposes, microbial communities colonize the remains, breaking down the organic matter. This microbial activity further accelerates the decomposition process, releasing additional nutrients into the environment. Consequently, carcass decomposition locally enriches the soil with essential nutrients such as nitrogen, phosphorus, and carbon, which are essential for the growth of primary producers and the functioning of ecosystems (Quaggiotto et al., 2019).
In marine ecosystems, whales represent a particularly significant source of carrion due to their large body sizes. When whales die, their carcasses usually sink to the ocean floor, where they become known as whale falls (Smith and Baco, 2003; Li et al., 2022). These whale falls create localized hotspots of biodiversity and productivity, supporting unique communities of organisms adapted to extreme conditions of pressure, temperature, and darkness in the deep sea. However, not all whale carcasses sink into the deep sea, some dead cetaceans decompose in shallow waters (Glover et al., 2013) or strand ashore. Decomposition of stranded carcasses may then add to ecosystem services of the beach and dune ecosystem (Quaggiotto et al., 2022). Particularly Coleoptera (beetles) form an important part of the carrion fauna (Olea et al., 2019). Certain beetles, like carrion beetles (Silphidae) and skin beetles (Dermestidae), rely on decomposing flesh as a primary food source. These species help break down the carcass and recycle nutrients into the ecosystem. Some beetle species lay their eggs in carcasses, providing their larvae with immediate access to food. For example, burying beetles (Nicrophorus spp.) not only consume carrion but also prepare it as a food source for their young (Milne and Milne, 1976). Different beetle species arrive at different stages of decomposition leading to multi-stage succession. Initially, species like rove beetles (Staphylinidae) and carrion beetles are attracted to fresh carcasses. As the decomposition progresses, skin beetles and bone beetles complete recycling of organic matter (Olea et al., 2019). By consuming carcasses, beetles contribute to nutrient cycling, and support other scavenger species linking carcasses to broader food web dynamics (DeVault et al., 2003).
Based on studies along US coasts, Geraci and Lounsbury (1993) distinguish seven management options after stranding: leave it, bury it, move it, return it to the sea, bring it to a rendering plant, blow it up with explosives or burn it. Tucker et al. (2018) distinguish four feasible options after stranding: leaving the carcass to decompose on site, beach burial, towing the carcass back to sea, or disposal to a rendering plant. Leaving a whale carcass on site is considered to be the most cost-effective and the most natural method, but raises issues of unpleasant appearance and unpleasant smell as well as changes in soil and groundwater chemistry. In the Netherlands large stranded whales are considered chemical waste that require formal disposal because they may have stored high levels of cadmium (MinVenW, 2007). Another issue can be contamination of the environment with euthanasia agents, if applied (Tucker et al., 2018). Because stranded whales are a food source for scavengers and provide nutrients for coastal sediments (Quaggiotto et al., 2022), leaving them for natural decomposition is encouraged, provided that legal and safety requirements can be met (Tavares et al., 2021; Quaggiotto et al., 2022). A better assessment of the ecological impact of the various stranding management techniques, and the comparison of their costs and benefits, is essential for management decisions. Only through a deeper scientific understanding of the socio-ecological context characteristic of stranded whale carcasses will we be able to delineate the most efficient management in increasingly densely populated coastal areas (Quaggiotto et al., 2022).
Cetacean strandings occur regularly along the coast of the Netherlands. Harbour porpoises Phocoena phocoena strand most frequently, with about 450 individuals annually between 2000-2019 (Keijl et al., 2021). Large whales strand irregularly, with annual averages of 0.9 minke whale Balaenoptera acutorostrata, 0.8 fin whale Balaenoptera physalus and 0.8 sperm whale Physeter macrocephalus between 2000-2019 (Keijl et al., 2021). Large stranded whales in the Netherlands are removed from the beach because of concerns about an exploding carcass, zoonotic diseases, odor nuisance and soil contamination with heavy metals.
A carcass of a 4.70 m male juvenile minke whale washed up on the uninhabited island Rottumerplaat, the Netherlands, on 25 November 2020. No necropsy was carried out to determine the cause of death of the animal, or whether it had eaten until death (Castrillon and Bengtson Nash, 2020). The carcass was moved the same night into the dunes at a height of 4.5 m above mean sea level, to prevent it from washing back into the sea (Figure 1). The aim of securing the carcass was to leave it for natural decomposition, to study the related ecological processes and to gain management experience in leaving a whale carcass in place. The objectives of this study were to assess (1) the carcass’ decomposition rate, (2) the attractiveness to avifauna, (3) the attractiveness to insect fauna, particularly beetles, (4) the effects on dune vegetation, and (5) the contamination with nutrients and metals to the soil, particularly cadmium. Further, we discuss recommendations on management interventions when leaving a carcass of a beached whale on site.

Figure 1. (a) Europe (light yellow) and the Netherlands (yellow), (b) The Wadden Sea (brown), (c) Wadden islands Schiermonnikoog (left) and Rottumerplaat (right) in the Dutch Wadden Sea, (d) Satellite view of Rottumerplaat, (e) Aerial UAV image of the minke whale carcass located inside the red circle. Satellite images by Google Earth, UAV image by The Fieldwork Company.
Rottumerplaat is a small beach barrier island of 7 km2 in the Dutch part of the Wadden Sea, consisting of dunes, salt marshes, sand flats and mud flats (Reise et al., 2010). At 53° N, the area contains typical temperate zone coastal dunes with a dominant vegetation of Ammophila arenaria (Hesp, 2004; Wiedemann and Pickart, 2004; De Groot et al., 2017). Winter temperatures in the Netherlands are generally mild, with average daytime highs ranging from 3°C to 6°C. Frost and snowfall occur occasionally but are typically short-lived. Summers are warm but not excessively hot, with average daytime highs between 20°C and 25°C. Heatwaves with temperatures exceeding 30°C are rare but can occur. Rainfall is frequent but usually light, with no distinct dry season. On average, the Netherlands receives 700–900 mm of precipitation annually.
This study was conducted opportunistically, field protocols and equipment were prepared in two days’ time. The minke whale carcass was first visited three days after stranding, on 28-11-2020 (DD-MM-YYYY), followed by monthly field visits on 16-12-2020, 27-01-2021, 25-02-2021, 30-03-2021, 28-04-2021, 28-05-2021 and 09-07-2021. In the following year, when the changes in the carcass were less prevalent, the carcass was visited less frequently on 06-05-2022, 10-08-2022 and 12-12-2022. The island Rottumerplaat is only accessible with a small vessel at high tide, leaving a time window of three hours for field work activities during each site visit.
During each field visit photos were taken of the carcass to document the state of decomposition. This was classified into one of five categories, known as Decomposition Condition Categories (DCCs), based on external characteristics following IJsseldijk et al. (2019). DCC1 - Extremely fresh carcass, just dead: no bloating or sloughing of skin; fresh smell; clear, glassy eyes. DCC2 - Fresh carcass: fresh smell, minimal drying and wrinkling of skin, eyes and mucous membranes; carcass not bloated, tongue and penis not protruded. DCC3 - Moderate decomposition: bloating evident, with tongue and penis often distended; skin cracked and started sloughing; characteristic (mild) odor can be expected; mucous membranes dry, eyes sunken. DCC4 - Advanced decomposition: carcass may be intact, but collapsed; skin sloughing; epidermis may be largely missing; strong odor. DCC5 - Mummified or skeletal remains: skin may be draped over skeletal remains; any remaining tissues are desiccated. The odour of the carcass could not objectively and quantitatively be determined without specialized equipment to determine volatile organic compounds (Verheggen et al., 2017). Therefore, a qualitative description of the odour was made during each field visit. This was done consistently by the same person for continuity.
Orthophotos of the carcass were taken during each on-site visit. Eleven ground control points were placed on and next to the carcass. Their geopositions in xyz-dimension were measured with 2 cm accuracy using a Trimble R6 Model 3 GNSS. Three Canon 850D DSLR cameras equipped with an Irix 11 mm f/4 wide-angle lens were mounted on a four meter long aluminum pole that was carried between two persons at a height of about one meter above the carcass, walking from snout to tail and reverse. The remotely controlled cameras synchronously took overlapping orthophotos. The photos were stitched to mosaics using Agisoft photogrammetry software to obtain a 3D georeferenced high resolution image. The horizontal resolution of a composite photo was 0.3 - 0.4 mm per pixel. Based on the altitude data, a digital elevation model of the carcass was created with a resolution of 0.6 - 0.8 mm per pixel.
Seven wildlife cameras on four positions were operational at distances of 4.0 m, 5.8 m, 8.7 m and 11.4 m. Three cameras were Reconyx HF2X and four were Snyper 4G with a mobile data connection. Images were taken every hour day and night and, additionally also triggered by motion. Every image was followed by a 5 s video. Images and videos were stored locally on an SD card, and a lower resolution image was sent to the whale’s e-mail address by the 4G cameras. During each field visit, the images and videos were retrieved from the SD cards, stored on a laptop and subsequently transferred to the Agouti repository for wildlife camera data by Wageningen University and Research (www.agouti.eu). The photos up to and including July 2021, in total 41,280 photos, have been stored in the Agouti repository. All photos were screened for the presence of birds to identify the species. A subset of 5,900 photos from all four camera positions has been annotated in Agouti for species behaviour. Photos were recorded up to 27-06-2023 after which the cameras were dismantled.
In December 2020 three pitfall traps for insect fauna were buried near the whale carcass. A pitfall trap consists of a 1 L plastic bucket with a roof to prevent rainwater falling in. The buckets contained a preservative consisting of 100 ml propylene glycol and 200 ml of water. When insects, such as beetles, walk around the carcass and fall into the trap, they are preserved (Greenslade, 1964; Hohbein and Conway, 2018). Three traps were placed at different distances to the carcass. Trap 1 was placed at 0.3 m, next to the head, trap 2 at 0.5 m, next to the lower abdomen, and trap 3 at 3 m, behind the tail fluke as a control sample. The contents of the pitfall traps were collected monthly. In May 2022 two extra pitfall traps were added close to the carcass. Trap 4 was placed between the whale’s jawbones at a distance of 0 m of the carcass, and trap 5 was placed against the vertebrae of the spine at the caudal peduncle at a distance of 0.05 m. All five traps were emptied monthly until August 2022. In December 2022, only traps 1, 3, and 4 could be retrieved. Only the lid of pitfall trap 2 was found, and there was no trace of trap 5. All captured insects were determined to species level where possible and counted. Sufficient data for statistical analysis was collected on the number of beetles (Coleoptera). To investigate the effects of time and distance on the total number of beetles, we applied a linear model using log-transformed beetle counts as the response variable. Specifically, we modeled the natural logarithm of species counts as a function of two predictors, time and distance:
where β0 represents the intercept, β1 and β2 are regression coefficients and є denotes the residual error. Log-transformation was applied to normalize its distribution, stabilize variance and make the residuals more homogeneously scattered. Time in months and distance to the carcass in metres were used as continuous predictor variables. The model was fitted using the ordinary least squares method. We assessed the significance of predictors through t-tests on the estimated coefficients, while the overall fit of the model was evaluated using the R2 statistic and residual diagnostics. Diagnostic plots for residual vs. fitted values, quantile-quantile residuals, Scale-Location and Cook’s distance were used to validate the adequacy of the model. All analyses were performed using R Statistical Software (R Core Team, 2024).
Soil samples were taken to monitor nutrients and minerals in the soil. Samples were taken using a soil auger with a diameter of 5 cm with a sampling depth of 10 cm. Soil samples were collected simultaneously with the vegetation surveys, except for July 2021, so in November 2020 (T0), May 2021 (T1), May 2022 (T2) and December 2022 (T3). Samples were taken next to the head (S1), next to the abdomen (S2), at a distance of 1.3 m from the abdomen (S3), and a control sample more than 4 m away from the tail fluke (S4). The T0 soil samples are assumed not to have been disturbed by the presence of the carcass, as it had only recently been placed there, and these served as control as well. Soil samples were analyzed by the Soil Chemical Biological Laboratory at Wageningen University & Research. Analyses were done by extraction with 0.001 M CaCl2, which provides a measure of the nutrients and minerals that are biologically available to plants. NH4, NO3+NO2, total N (Nts), P-PO4, pH and C were analyzed by means of extraction and flow-through analysis CFA. Al, Cu, Fe, K, Mg, Mn, Na, P, S and Zn were analyzed by means of extraction and mass spectrometry ICP-AES. As, Cd, Co, Cr, Cu, Ni, Pb and V were analyzed by means of extraction and mass spectrometry ICP-MS. For all analyses the detection limit of the analytical equipment is provided, denoting the lowest concentration of the component in the sample whose presence can be reliably determined. To evaluate the effect of the carcass on soil contents of metals and non-metals we applied a linear model using the natural logarithm of biologically available soil content as response variable and treatment as a categorical predictor. The model was expressed as:
where β0 is the intercept corresponding to the baseline (control) treatment, βk represent the estimated effect of a treatment group Tk relative to the control treatment, and є denotes the residual error. The response variable was log-transformed to normalize its distribution, stabilize variance and make the residuals more homogeneously scattered. The values for pH were not transformed because of their logarithmic scale. The predictor variable is ‘whale treatment’. Treatment is zero for the samples collected in November 2020 (T0) and the control samples that were taken more than 4 m away from the tail fluke (S4). Treatment is one for all other samples. Three linear models were fitted to analyze the duration of the effect on soil contents, i.e. for changes between control treatment and T1 (month 6), for changes between control treatment and T2 (month 18) and for changes between control treatment and T3 (month 25). Only constituents measured in levels well above the detection limit were analyzed. The models were fitted using the ordinary least squares method. We assessed the significance of predictors through t-tests on the estimated coefficients, while the fit of each model was evaluated using the R2 statistic and residual diagnostics. Diagnostic plots for residual vs. fitted values, quantile-quantile residuals, Scale-Location and Cook’s distance were used to validate the adequacy of the models. All analyses were performed using R Statistical Software (R Core Team, 2024).
The dune in which the carcass was placed consisted of disturbed soil and was already a nutrient-rich site with many rabbit droppings. The vegetation composition was mapped near the carcass using three permanent quadrats (PQs) of 1 m2 each. PQ1 was located on the higher eastern side of the carcass near the head, PQ2 at the lower western side of the carcass near the abdomen, and PQ3 at 15 m northwest of the carcass as a control. The permanent quadrats were marked in the field using buried magnets. Vegetation surveys were conducted in November 2020, March 2021, July 2021, May 2022 and December 2022. All vascular plants and mosses were recorded applying the modified Londo scale (Londo, 1976) and the average vegetation height was estimated. Small samples of mosses were collected for later identification under a microscope. Lichens were not found in the permanent quadrats.
The decomposition of the carcass is presented by orthophotos made during each site visit (Figure 2) and with photos from the wildlife cameras (Supplementary Material). The change in volume of the decomposing minke whale carcass was determined for the first seven months since stranding (Figure 3). Each site visit, observations were made on the status of the decomposing whale (Table 1).

Figure 2. Orthophotos of the minke whale carcass. Due to growing vegetation, the photogrammetry software sometimes partly failed to stitch the orthophotos together.
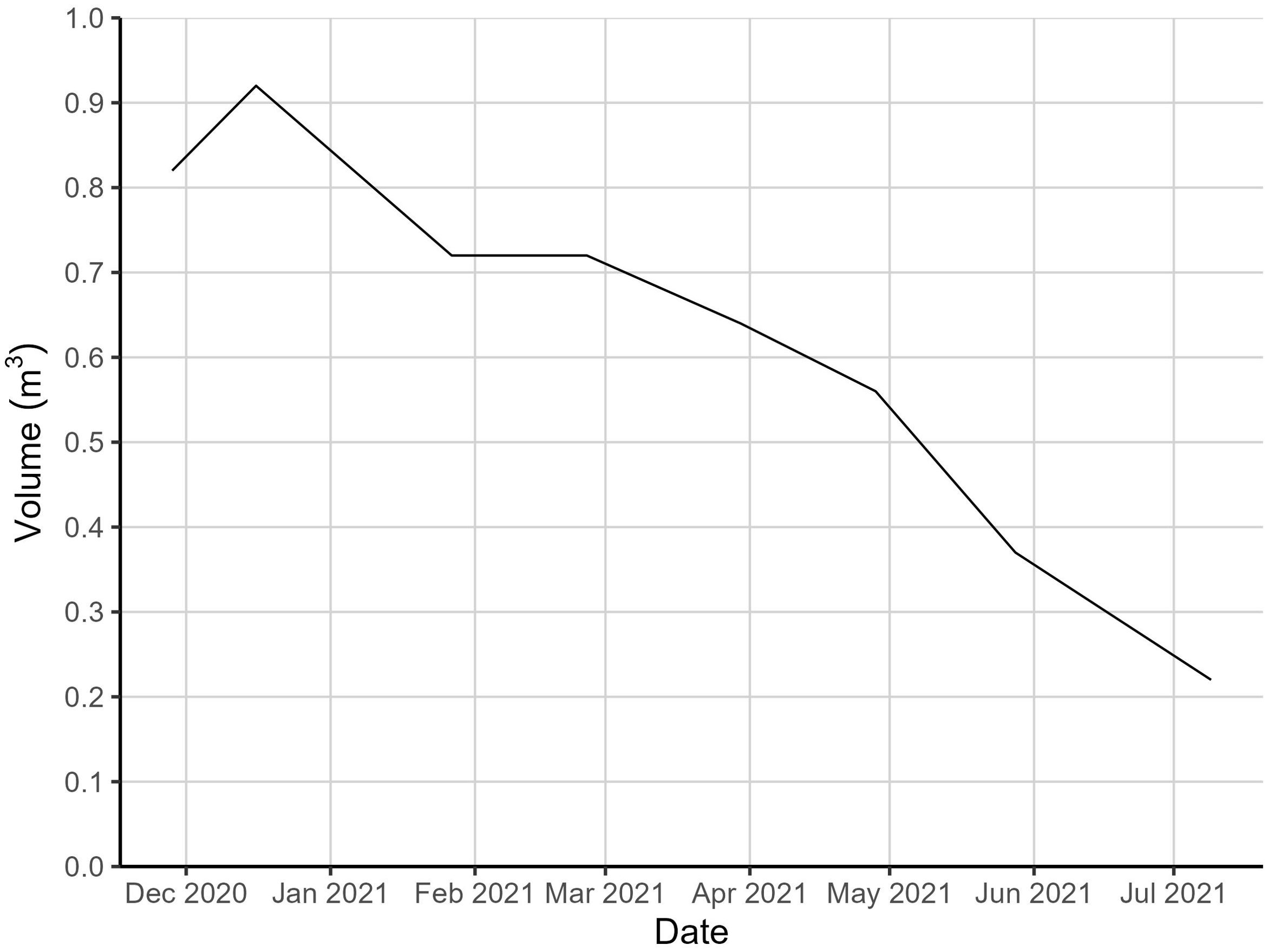
Figure 3. Development of the volume of the minke whale carcass over time.

Table 1. Decomposition Condition Category (DCC), observations and volume (m3) of the decomposing minke whale carcass over time.
Based on the annotated wildlife trail cameras’ footage the most frequently observed birds near the whale carcass were Eurasian oystercatcher Haematopus ostralegus (40%), Eurasian magpie Pica pica (32%) and carrion crow Corvus corone (21%) (Figure 4). Two pairs of oystercatcher nested near the carcass and they often foraged next to it. The oystercatcher chicks also stayed near and on top of the carcass for foraging for days, despite the presence of magpies. Apparently, the carcass and the immediate vicinity of the carcass provided a good food source. Bird species recorded as foraging on the carcass itself were carrion crow, oystercatcher, magpie, herring gull Larus argentatus, great black-backed gull Larus marinus, black redstart Phoenicurus ochruros and fieldfare Turdus pilaris. Of these, carrion crow was seen most often (54%), followed by magpie (28%) and great black-backed gull (11%). These bird species foraged on the carcass meat. Foraging oystercatcher, black redstart and fieldfare foraged on the insect imagos and maggots found on or in the carcass. Other species of insect-eating birds were also observed foraging in the vegetation next to the carcass. A total of 26 bird species were seen, including four scavengers, thirteen insectivores, two mixed scavenger-insectivores, and the remaining seven species were passers-by that were more or less accidentally visiting the carcass (Table 2).

Figure 4. Wildlife camera image showing the three most often observed bird species in one shot: Eurasian oystercatcher, Eurasian magpie and carrion crow. Note that a cover of one of the pitfall traps is visible on the right.
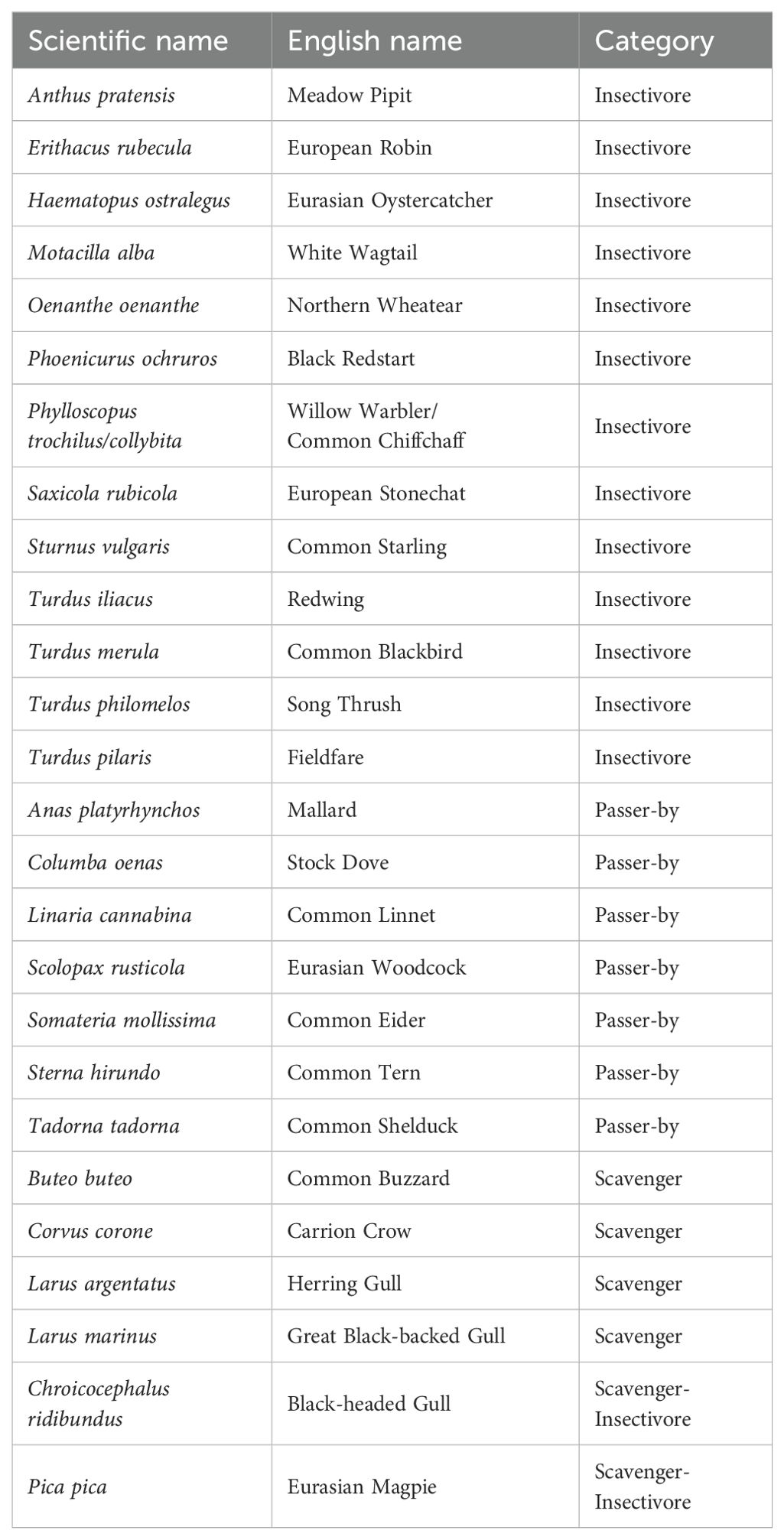
Table 2. Bird species observed on or near the minke whale carcass up to and including eight months since stranding, November 2020 – July 2021.
A large number of beetles were found in the pitfall traps, a total of 1373 specimens from 57 species in 2021 and a total of 4667 specimens from 104 species in 2022 (Supplementary Material). Over both years, 129 species of beetles were found near the carcass, of which eight species are specific for carcasses (Table 3). Three species of carrion beetles (Silphidae) were found in 2021. These (larger) beetles have not been observed on Rottumerplaat before. The burying beetle Nicrophorus vespillo, shore sexton beetle Necrodes littoralis and sinuate burying beetle Thanatophilus sinuatus are true carrion-eaters that feed on flesh and maggots of flesh flies. New observations were also made of the flesh- and maggot-eating hairy rove beetle Creophilus maxillosus and the black scavenger beetle Saprinus semistriatus. Large numbers of the smooth carrion beetle Saprinus planiusculus showed that these may also have been attracted to the carrion. The three large carrion beetle species and the hairy rove beetle were observed only in 2021 when the carcass was still ‘fresh’. The carnivorous smooth carrion beetle Saprinus planiusculus was also found in much higher numbers (390 ind.) in 2021 than in 2022 (6 ind.). Large numbers of the skin and dried animal remains-eater black larder beetle Dermestes frischii were found in 2021 (159 ind.) compared to 2022 (16 ind.). In contrast, species that specialise in eating bones such as the common furniture beetle Omosita colon were found in much higher numbers (3664 ind.) in 2022 than in 2021 (305 ind.), showing multi-stage succession of beetle species.
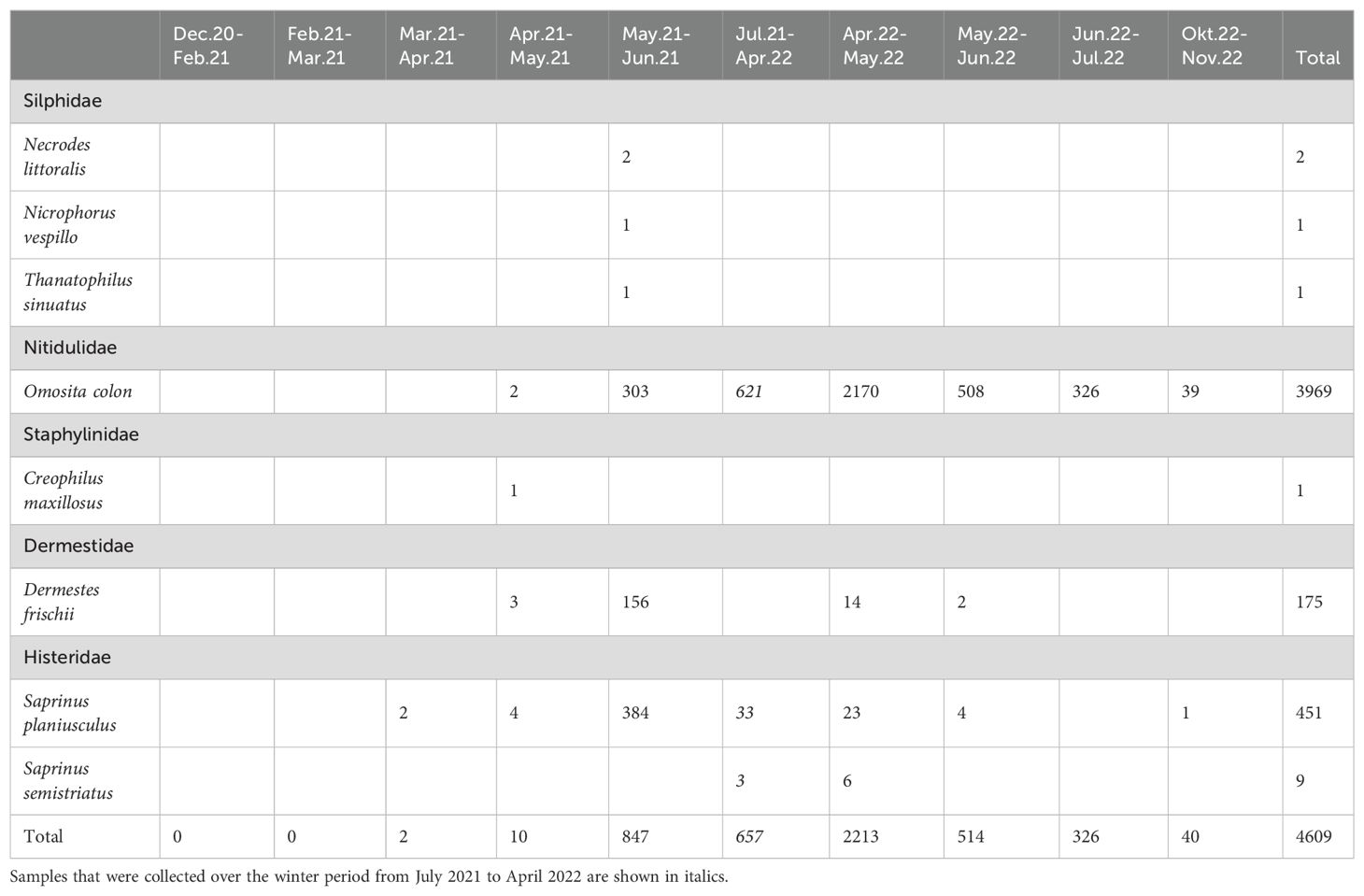
Table 3. Numbers of beetles specific for carcasses found at the minke whale over time.
Over the five month period from February 2021 to June 2021 the effect of time was statistically significant and positive (t(12) = 4.48, p <.001), whereas the effect of distance was statistically non-significant and negative (t(12) = -0.86, p = 0.408). This shows a seasonal trend in increasing beetle numbers from winter to summer. From May 2022 to November 2022 the effect of time was statistically non-significant and negative (t(15) = -0.26, p = 0.797), whereas the effect of distance was statistically significant and negative (t(15) = -2.91, p = 0.011). The two additional pitfall traps that were placed at 0 and 0.05 m to the carcass in May 2022 showed significantly higher numbers of beetles, indicating that beetles stay very close to the carcass.
All biologically available metal contents showed significant changes for whale treatment in one or more of the comparison periods. Potassium (K) and nickel (Ni) showed significantly enhanced levels 6 months after placement of the carcass, followed by a levelling to background soil contents. For arsenic (As), cobalt (Co) and vanadium (V) the significant increase in soil contents lasted for 18 months. Sodium (Na) showed significantly increased levels for all compared time periods and together with manganese (Mn) persisted in significantly higher concentrations 25 months after placement of the carcass. Magnesium (Mg) did not show significantly elevated contents at 6 months, but did at 18 and 25 months (Table 4; Figure 5). The other cationic (heavy) metals cadmium (Cd), copper (Cu), iron (Fe), molybdenum (Mo), lead (Pb) and zinc (Zn) remained below the detection limit during the measuring period.
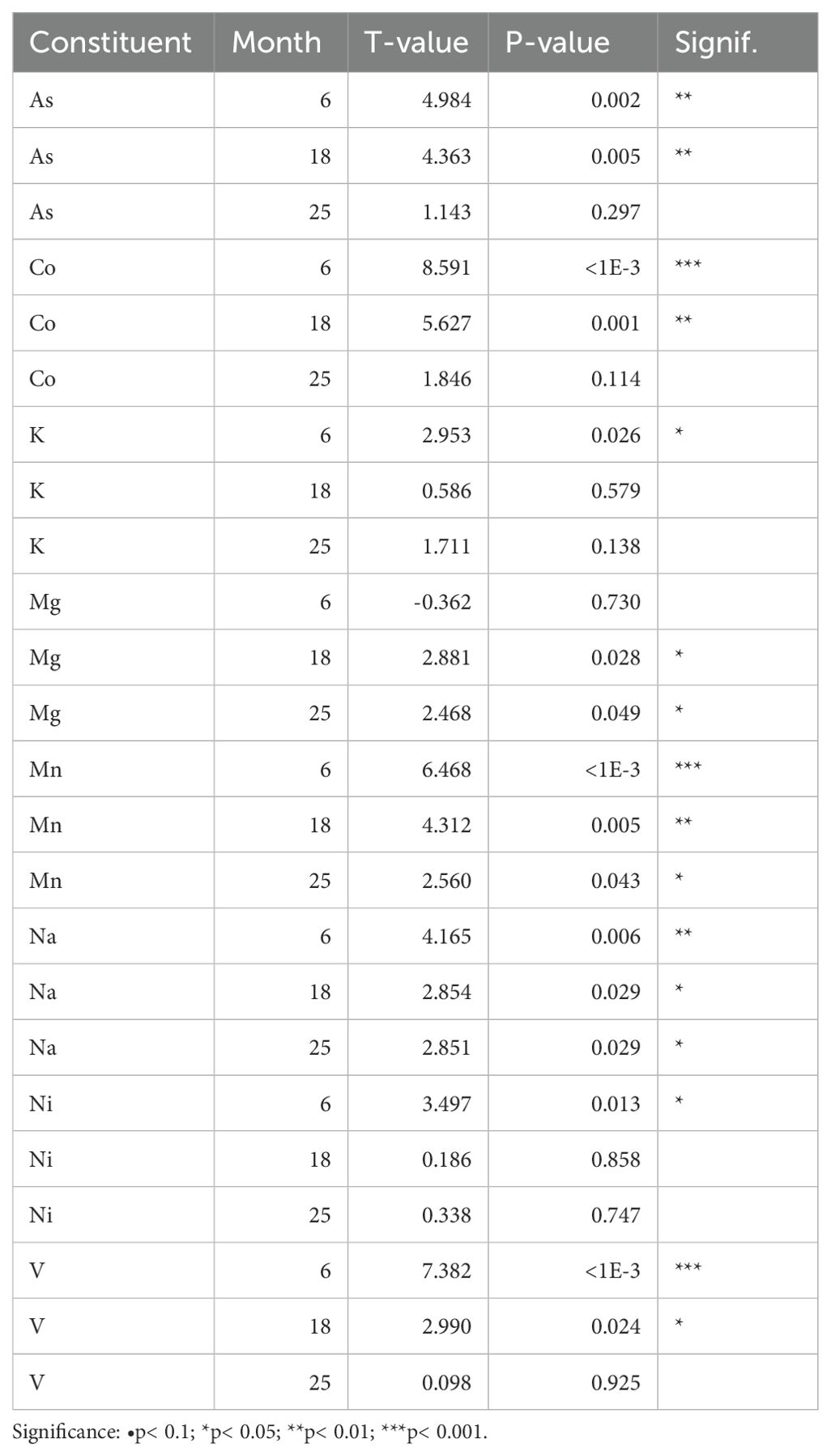
Table 4. Effect of ‘whale treatment’ on soil metal contents after 6, 18 or 25 months.
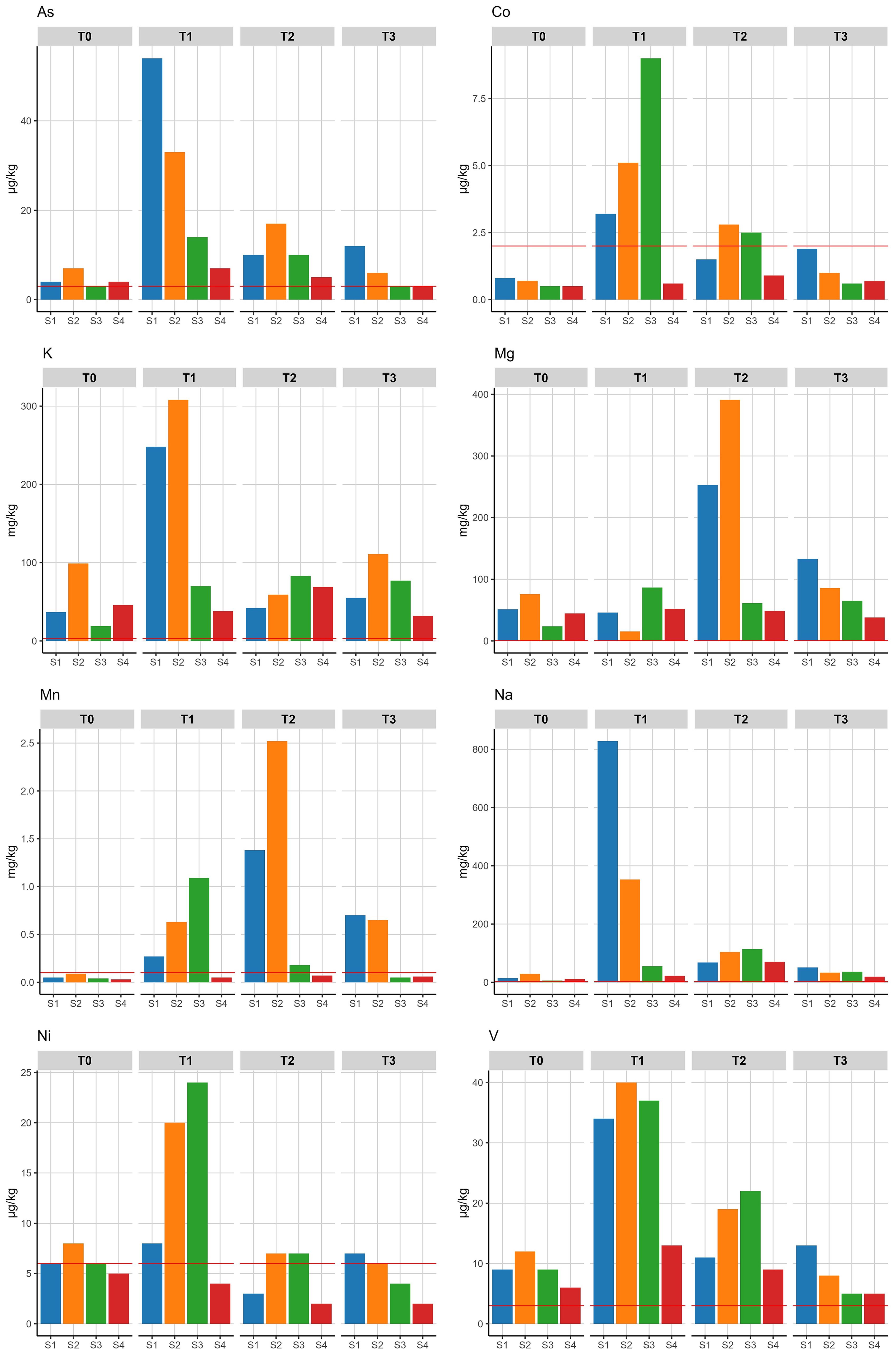
Figure 5. Biologically available metal contents in soil samples measured above the detection limit of the analytical equipment (red line). S1 and S2 at 0.15 m, S3 at 1.3 m, S4 at 4.5 m. T0 = month 0, T1 = month 6, T2 = month 18, T3 = month 25.
For non-metals, a significant effect of the presence of the whale carcass showed for all constituents, with long-lasting duration. Carbon (C) and sulphur (S) contents showed significantly elevated levels at 6 months and 18 months, as well as elevated levels at 25 months in close proximity to the carcass. Total nitrogen (Nts) showed significantly elevated levels at 6 months and 18 months with a significant increase in ammonium (N-NH4) at six months and a significant increase in nitrate+nitrite ((N-(NO3+NO2)) at 25 months. Significantly increased levels for phosphorus (P), mainly in the form of orthophosphate (P-PO4), showed throughout the measurement period. The pH decreased 25 months after placement of the carcass, although with low significance (Table 5; Figure 6).
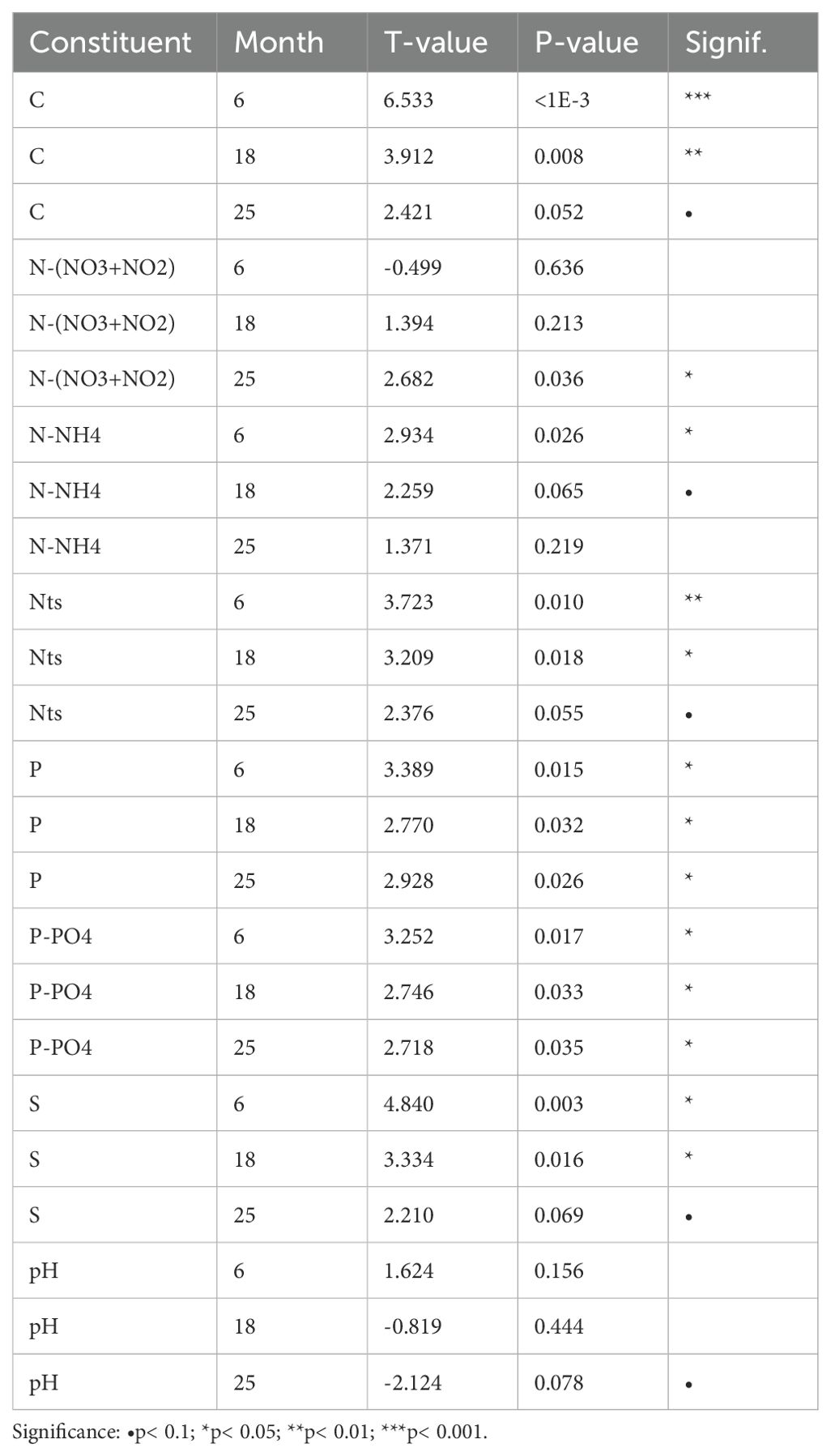
Table 5. Effect of ‘whale treatment’ on soil non-metal contents after 6, 18 or 25 months.
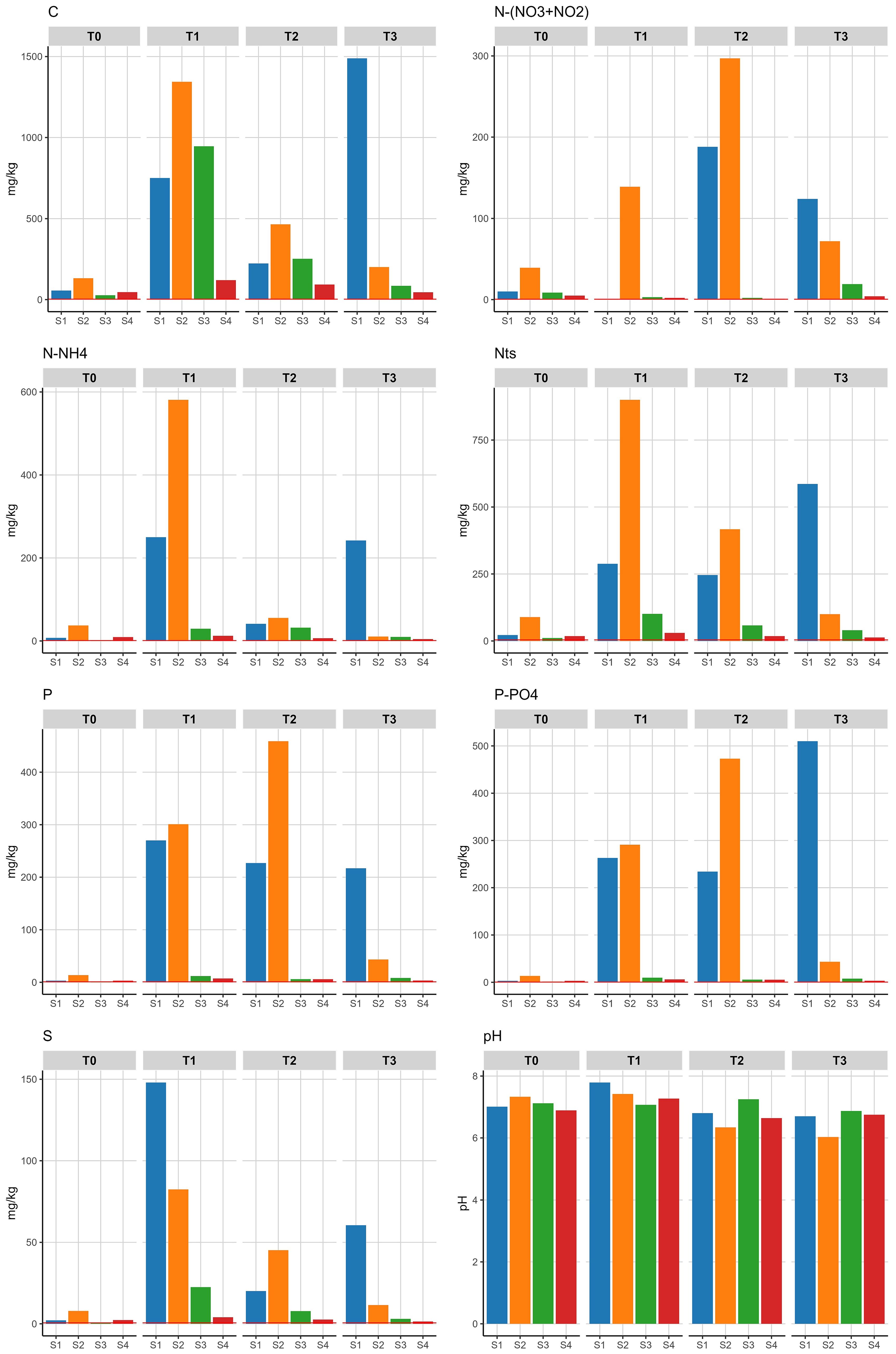
Figure 6. Biologically available non-metal contents in soil samples measured above the detection limit of the analytical equipment (red line). S1 and S2 at 0.15 m, S3 at 1.3 m, S4 at 4.5 m. T0 = month 0, T1 = month 6, T2 = month 18, T3 = month 25.
No effect of the carcass on the vegetation composition in the PQs could be established. Vegetation mortality occurred in a zone up to 23 cm around the carcass in the spring and summer following the stranding. Immediately outside the zone of dead vegetation, accelerated growth and high vigor of spring flowering plants was observed, which can be attributed to the increased nutrient supply. The dead zone had almost completely disappeared by the following spring in May 2022. Stinging nettle Urtica dioica spread around the carcass and the increase in common bent Agrostis capillaris and creeping thistle Cirsium arvense also indicated an increase in available nutrients. After two years, the dead zone had completely disappeared and vegetation grew through the skeleton, especially common nettle was lush green. There were no developments in the morphology of the dune as there was no shifting sand.
As far as we know this is the first decomposing whale carcass located in a temperate dune to have been closely monitored. Whale falls in the ocean have been studied more often (Li et al., 2022), but beached dead whales are usually disposed of, or when stranded at remote locations, left without scientific monitoring. A similar carcass of an adult minke whale that stranded in the dunes of the German island Juist on 24 September 2012 was left on site and the decomposition process was documented photographically: two years since stranding the skull was exposed, but not all the body tissues had decomposed. Experiments on decomposition and associated biodiversity benefits in dune environments have been described for fish carcasses that were deliberately left in Australian dunes monitored with wildlife cameras (Huijbers et al., 2013). This attracted birds of prey, foxes, cats, dogs and ghost crabs (Schlacher et al., 2013).
The whale carcass at Rottumerplaat showed a discoloration of the skin to light brown two months after washing ashore, and the skin darkened progressively to reddish-brown and dark-brown over the next months. This was probably caused by chemical reactions occurring by the breakdown of fats, fatty acids and amino acids (Zamora and Hidalgo, 2005; Domínguez et al., 2019). Notable was the occurrence of many spots of white color and fungal-like structures after four months of decomposition. This was most likely ‘efflorescence’ from saltpeter (potassium nitrate) which is known from white deposits on walls, by potassium and ammonium leaching from the skin. Two months later there were no longer any fungus-like, white, fluffy structures on the skin, but there was still a flat white crust. The disappearance of salt efflorescence can be explained by increased precipitation, as potassium nitrate is a highly soluble salt.
The decomposing whale carcass did not become a feeding frenzy for birds, although there were plenty of birds around. Carcasses of seals and porpoises on Dutch beaches are opened and eaten by birds, such as gulls and crows. However, the skin of the minke whale seemed too tough to break open. Only minor traces of bird pecking, mainly on extremities such as the mandibles, fins or tail fluke were observed. Birds that would probably be physically capable of opening the carcass, such as the white-tailed eagle Haliaeetus albicilla do occur in the region, but no eagle was seen foraging on the carcass. Rats, wild boar or foxes that are able to open the carcass do not occur on the island of Rottumerplaat, which may result in a significant reduction in the amount of carrion consumed by other scavengers (Tobajas et al., 2021). At times, rabbits seemed to be scavenging on the carcass, which is not unlikely (Clauss et al., 2016).
Many beetle species were observed near the whale carcass, in total 129 different species. Of these, as many as 74 species were new to Rottumerplaat, that is, not yet described in Zekhuis and De Vries (2012), the standard work on the fauna of the Rottum islands. Probably the area has been poorly surveyed for beetles before. Carcasses are already available on Rottumerplaat, e.g., of stranded harbour porpoises and seals, or of birds and rabbits, but the associated beetle fauna has not been previously studied in detail. Compared to smaller carcasses such as those of birds or land mammals, this large whale carcass attracted high numbers of beetles. And because it’s decomposition took a long time, several generations of beetles could be raised. Moreover, skin and bone remains will remain for an even longer time, which can attract other specialized beetle species. A decomposing whale carcass therefore has high additional value for beetle biodiversity.
A whale carcass buried below the groundwater table (in the saturated zone) can bring high loads of carbon and ammonium to the groundwater (Heiss, 2020). Via groundwater, this can reach the sea, leading to nutrient enrichment and even attracting sharks (Heiss, 2020). An experimental study by Tucker et al. (2019) showed that transport of constituents from whale carcasses buried above the groundwater table (in the unsaturated zone) is low. They consider it unlikely that a whale carcass buried above the water table at a distance of more than 25 m from the tide line would leak constituents to groundwater. The whale carcass at Rottumerplaat was laid down well above the water table.
The metal contents in the soil for potassium (K) and nickel (Ni) showed a short term increase due to the presence of the whale carcass. The measured available concentrations for Ni (up to 25 μg/kg) are however well below typical observed available concentrations in sandy soils that range from 50 to 100 μg/kg (Mol et al., 2012). Increased levels of arsenic (As), cobalt (Co) and vanadium (V) were found up to one and a half years later and sodium (Na) and manganese (Mn) persisted in elevated levels up to two years. The temporal increase in the levels of anionic species of As and V can be explained partly by the rather high dissolved concentrations of P since both As en V can be mobilized at high concentrations of P in soil (Sun et al., 2019; Wu et al., 2022). In addition, the small increase in pH in locations S1 and S2 also contributes to the release of both elements. The total amounts of As and V released are small compared to natural background levels for both elements (Mol et al., 2012) and the added inputs to soils will have a limited or no effect on the ecosystem. Other cationic (heavy) metals could not be detected or remained below the detection limit during the measuring period. The low availability of cationic elements is likely due to the neutral pH that prevails in all locations. In fact the pH in locations S1 and S2 tends to increase temporarily due to the presence of the carcass which further reduces the mobility of these cationic metals. Based on these observations, the impact of the (heavy) metals from this whale carcass is expected to have little influence on the environment.
One of the reasons why a beached whale is considered chemical waste that should be disposed of in the Netherlands is that these animals may have stored high levels of cadmium (MinVenW, 2007). Whales absorb heavy metals and chlorinated hydrocarbons from their food during their lifetime and this bioaccumulation increases with age, especially in the kidney and liver that act as a sink of such contaminants in the animal body (Wagemann and Muir, 1984; Hansen et al., 1990; UNEP, 1991; Bellante et al., 2012). The levels can be so high that eating whale meat should be discouraged (Caurant and Amiard-Triquet, 1995; Endo et al., 2002, 2003). However, the actual scale of this problem, in the case of a cetacean stranded on the Dutch coast, is far from clear (Evans, 2013). Available information indicates that relatively high levels of hazardous components are found in squid eating toothed whales (such as pilot whales Globicephalus sp. and sperm whales Physeter macrocephalus), with the highest concentrations found in the liver and kidneys (Law et al., 1996, 1997; Holsbeek et al., 1999; Gajdosechova et al., 2016; Lischka et al., 2021). Prey species from deeper and colder waters have lower growth rates and accumulate more cadmium (Ciesielski et al., 2006). Cetaceans that eat more fish than squid accumulate, as far as we know, lower amounts of heavy metals (Hansen et al., 2016). Baleen whales in particular, which eat lower trophic level prey (krill), have lower levels of contaminants (about a factor of 10 lower for metals) in their tissues, but this general ‘rule’ does not necessarily apply to every individual: between different individuals of the same species, levels can differ by more than a factor 10 (Bowles, 1999). Moreover, most species of baleen whales that beach on North Sea coast are fish-eaters rather than krill eaters, and the larger baleen whales that do eat mainly krill may contain many pollutants in absolute terms. In the soil samples near the minke whale carcass at Rottumerplaat no bioavailable cadmium was found above the detection limit. This means that the cadmium levels were very low, indicating that this young animal had not accumulated much cadmium during its short lifetime. In view of potentially high levels of organochlorines in stranded whales (Holsbeek et al., 1999), we recommend to pay more attention to soil contamination with PCBs and DDT instead of heavy metals in further studies on decomposing whales.
No effect of the carcass on the vegetation composition in the PQs could be established. The changes in vegetation composition and cover observed in the PQs can be attributed to seasonal influences and grazing by rabbits. However, as organic material from the whale’s tissues seeped into the soil, the soil was enriched with the nutrients phosphorus, sulfur and nitrogen in close proximity to the carcass. Phosphorus and nitrogen showed increased soil contents over a period of two years and nitrogen was mainly present in the form of ammonium (Figure 6). Vegetation died in a zone up to 23 cm around the carcass. A zone of dead vegetation is a normal phenomenon around large carcasses, e.g., near bison Bison bison carcasses in the North American prairie (Knapp et al., 1999). Although the nutrients initially released occur in concentrations toxic to vegetation, these spots eventually become zones of high fertility (Wenting et al., 2023). This was indeed also observed by lush vegetation dominated by eutrophic species growing in and among the whale skeletal remains 1.5 year since stranding.
Stranded whales are a fascinating subject for the general public (Sousa and Brito, 2012). However, leaving a carcass in place is not a viable option for sites located near residential areas, or on beaches with a lot of public traffic, because of the smell and unpleasant sight of decomposition (Tucker et al., 2018). Offshore disposal by towing a floating carcass out to sea is therefore advocated as a viable option (Meynecke et al., 2024), but we consider the risks for navigation and other user functions too high in crowded seas. Such a carcass might strand again, in a more populated area, or pose a hazard to shipping. We recommend using a suitable net that is securely anchored to fix a whale carcass on a beach. Another option, as was chosen at Rottumerplaat, is to move the carcass further up the shore to a flood-free level. This requires heavy equipment and a whale that is not too heavy. There is also a risk of a carcass exploding. This can occur with freshly stranded animals due to the accumulation of decomposition gases (Reisdorf et al., 2012). Gas buildup depends on temperature, the structural integrity of the carcass, as well as biotic (bacterial type and activity) factors (Moore et al., 2020). The buildup of pressure inside the carcass often leads to the hissing escape of gases from openings such as the mouth, blowhole, or anus. Explosions are only known to occur when someone pierces the whale with a sharp object or when a carcass is moved or transported (Reisdorf et al., 2012). We recommend to not transport a fully bloated whale carcass.
The smell of the minke whale carcass at Rottumerplaat was not strong shortly after stranding. While the smell became stronger over time, it never became an overwhelming stench. Downwind of the carcass, it was easily detectable from a distance of about 10-30 m. Bird watchers who had their shelter 45 m away from the carcass reported that on some days with a mild wind from the ‘right’ direction, they could smell the carcass from near their shelter. The smell was never intense but at times unpleasant. However, not every large cetacean that washes ashore can be left to decompose at any location. Size matters, because a big carcass takes longer to decompose and probably emits a heavier and longer-lasting odor. Only limited information could be obtained on distances for human perception of the odor from a whale carcass. The greatest distance at which the stench of a dead whale was noticeable is 5 km, according to online newspaper articles. Without quantitative information on the odor emission from a decomposing whale carcass and its relationship with size, season, wind conditions and humidity there is no well-founded advice to be given on a minimum distance from habituation. However, if a stinking carcass leads to odor nuisance, it remains possible to remove and dispose of the carcass, or to bury it.
There is a risk of illegal removal of specific parts of a carcass dependent on the species. With smaller species, this may involve the (entire) skull; teeth or baleen. Sperm whale teeth are highly sought-after and tradable collectibles (although this is illegal), and ambergris from the intestines of a sperm whale (1 in 100 individuals; (Clarke, 2006)) is also highly valuable. Sperm whale teeth should be removed to prevent looting. If the decision were made to leave a baleen whale carcass on site, consideration should be given to pre-emptively removing the baleen, both for research purposes and to prevent looting. Apart from the risk of illegal removal of specific parts of a carcass by humans, birds or small mammals could also move small bones away, so there is no guarantee for a pristine skeleton. When a carcass is left to decompose on an easily accessible and publicly available location we recommend to have conservation stewards nearby to reduce unwanted behavior from the public. Stewards can provide information about the carcass and its value for biodiversity. We recommend to station a mobile hut for stewards near the carcass. However, considering the potential odor nuisance, it should not be too close.
A whale carcass can have value for museums or for science. Allowing an animal to decay may be incompatible with this, although it is not excluded beforehand that skeletal parts could still be salvaged and preserved. However, the quality of such material would be lower after a lengthy decaying process. Universities and research institutes collaborate worldwide on research into the pathology, cause of death and diet of stranded cetaceans. If an animal remains in place, such research cannot take place, unless a mode is found involving a combination of pathological/biological research followed by leaving the animal. This could have other advantages as well, because chopping carcasses up into smaller pieces facilitates scavengers and accelerates decomposition (Quaggiotto et al., 2022). Moreover, during a necropsy, samples can be taken from organs suspected to contain high concentrations of toxic constituents and the size or weight of these organs can be determined to gain more insight into the presence of contaminants in various tissues, and in environmental contamination resulting from leaving the carcass. Organs suspected to contain high concentrations of toxic constituents, such as the kidneys (metals) or blubber (organic micro-pollutants), can be removed, reducing the environmental impact of leaving the carcass. The remains of a carcass after a necropsy are easier to move than a complete carcass. This is especially relevant for very large carcasses, such as those of adult fin whales or sperm whales and if at any point later in time it becomes necessary to remove the carcass, due to odour problems, enforcement issues, or potential soil contamination.
Last, but not least, there is the risk for zoonotic or epizootic infections (MinLNV, 2020). The risk of contracting a transmissible disease from cetaceans to humans is considered low (Dr. Lonneke IJsseldijk, Utrecht University, personal communication). Although various pathogens, carried by cetaceans, are potentially dangerous to humans, they are generally treatable and not fatal: brucella, erysipelothrix, salmonella, toxoplasmosis, campylobacter, and mycoplasma. Risks of zoonotic infections mainly arise from contact with sharp teeth that carry oral flora. Dolphins have sharper teeth than larger toothed whales (sperm whales), so one should be particularly careful with dolphins. In addition, risks of zoonoses are high with contact with open wounds or via aerosols from exhalation when an animal is still alive, or for example during sawing or cleaning operations on dead animals (IJsseldijk, pers. comm). To minimize risks of zoonotic infections it is recommended to leave whales for decomposition in areas with no or limited public access, exclude animals with visible, suspect skin defects and remove the teeth of toothed whales.
Recent infections among marine mammals with highly pathogenic avian influenza have raised attention to the sensitivity of areas with many seals. Seals and other pinnipeds (Gamarra-Toledo et al., 2023) can become infected by avian influenza (Zohari et al., 2014; Bodewes et al., 2015b, a; Shin et al., 2019; Mirolo et al., 2023; Puryear et al., 2023). Infections have been found in multiple species of cetaceans, including harbour porpoise, bottlenose dolphin Tursiops truncatus and white-sided dolphin Lagenorhynchus acutus (Adlhoch et al., 2023). Until recently, it was assumed that marine mammals could only be infected by birds, but avian influenza viruses constantly mutate and are now better adapted to infecting and replicating in mammals (Bordes et al., 2023; Gamarra-Toledo et al., 2023). Epizootic spread of avian influenza from a stranded dead cetacean to seals with high conservation value should be prevented by not leaving a whale carcass near their resting locations. We recommend sampling a stranded whale for avian influenza, using swabs from the blowhole. Because several days could elapse between sampling and the test results being available, precautions should be taken to remove a stranded animal if it turns out to be infected.
This study examined the decomposition of a juvenile minke whale carcass over two years in the Wadden Sea’s dune ecosystem, emphasizing its ecological impact. The carcass significantly contributed to biodiversity, particularly among beetle species, which exhibited a multi-stage succession. Nutrients from the whale enriched the soil, initially causing vegetation die-off but later promoting lush plant growth. The natural dune vegetation showed no long-term adverse effects. Bioavailable metal levels in the soil changed temporarily, with potassium (K) and nickel (Ni) increasing shortly, while arsenic (As), cobalt (Co), and vanadium (V) remained elevated for up to 1.5 years, and Sodium (Na) and manganese (Mn) persisted for two years. No bioavailable cadmium (Cd) was found above the detection limit and metal concentrations posed no lasting environmental risk to soil quality.
To prevent displacement by waves and ensure controlled decomposition, securing beached whales with nets or relocating them to flood-free zones is recommended. Removing valuable parts, such as sperm whale teeth and baleens, can prevent looting. In publicly accessible areas, mobile conservation stewards can provide supervision and communication. A partial necropsy may facilitate sample collection, reduce environmental impact, and ease later transport if necessary. For safety, decomposition should occur in areas with restricted public access, and carcasses with visible abnormalities should be excluded. Toothed whale teeth removal is advised. Stranded whales should be tested for avian influenza, and placement near seal haul-out sites should be avoided. These management interventions ensure ecological benefits while mitigating risks.
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author/s.
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because the animal was already dead when research started.
MB: Conceptualization, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Visualization, Writing – original draft. ML: Conceptualization, Methodology, Writing – original draft. Mv: Formal analysis, Visualization, Writing – review & editing. HV: Investigation, Writing – review & editing. NJ: Investigation, Writing – review & editing. DL: Investigation, Resources, Writing – review & editing. WD: Investigation, Resources, Writing – review & editing. PR: Writing – review & editing. JB: Investigation, Writing – review & editing.
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was commissioned by Rijkswaterstaat, Ministry of Infrastructure and Water Management, the Netherlands, under contracts 31167916 and 31186666.
We are thankful to Rick Hoeksema and Sophia Bats (RWS-NN), Jan Kostwinner and Freek Jan de Wal (crew of the MS Harder, Wadden Unit, Ministry of Agriculture, Nature and Food Quality), Jaap Kloosterhuis (Staatsbosbeheer), Jannes Heusinkveld (The Fieldwork Company), Peter Nobels (Soil Chemistry Laboratory, WUR), Theodoor Heijerman (retired Biosystematics Group, WUR), Patrick Jansen, Elke Wenting and Yorick Liefting (Wildlife Ecology and Conservation Group, WUR), Oliver Katenhusen and Annika Kruse (NLWKN), Jos Houbraken (Westerdijk Fungal Biodiversity Institute), Guido Keijl (Naturalis), Jan Willem Zwart (national coordinator cetacean strandings, Waddenunit), Lonneke IJsseldijk (University Utrecht) and last but not least the bird watchers of Rottumerplaat season 2021 Addo van der Eijk & Frank Oosterhoff and season 2022 Marc van der Aa & Holmer Vonk.
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2025.1474460/full#supplementary-material
Adlhoch C., Fusaro A., Gonzales J. L., Kuiken T., Marangon S., Niqueux É., et al. (2023). Avian influenza overview September – December 2022. EFSA J. 21. doi: 10.2903/j.efsa.2023.7786
Barton P. S., Cunningham S. A., Lindenmayer D. B., Manning A. D. (2013). The role of carrion in maintaining biodiversity and ecological processes in terrestrial ecosystems. Oecologia 171, 761–772. doi: 10.1007/s00442-012-2460-3
Bellante A., Sprovieri M., Buscaino G., Buffa G., Di Stefano V., Manta D. S., et al. (2012). Distribution of Cd and As in organs and tissues of four marine mammal species stranded along the Italian coasts. J. Environ. Monit. 14, 2382–2391. doi: 10.1039/c2em30161j
Benbow M. E., Receveur J. P., Lamberti G. A. (2020). Death and decomposition in aquatic ecosystems. Front. Ecol. Evol. 8. doi: 10.3389/fevo.2020.00017
Bodewes R., Bestebroer T. M., van der Vries E., Verhagen J. H., Herfst S., Koopmans M. P., et al. (2015a). Avian influenza A(H10N7) virus-associated mass deaths among harbor seals. Emerg. Infect. Dis. 21, 720–722. doi: 10.3201/eid2104.141675
Bodewes R., Rubio García A., Brasseur S. M., Sanchez Conteras G. J., van de Bildt M. W. G., Koopmans M. P. G., et al. (2015b). Seroprevalence of antibodies against seal influenza A(H10N7) virus in harbor seals and gray seals from the Netherlands. PLoS One 10, e0144899. doi: 10.1371/journal.pone.0144899
Bordes L., Vreman S., Heutink R., Roose M., Venema S., Pritz-Verschuren S. B. E., et al. (2023). Highly pathogenic avian influenza H5N1 virus infections in wild red foxes (Vulpes vulpes) show neurotropism and adaptive virus mutations. Microbiol. Spectr. 11. doi: 10.1128/spectrum.02867-22
Bowles D. (1999). An overview of the concentrations and effects of metals in cetacean species. J. Cetacean Res. Manage. Special Is, 125–148. doi: 10.47536/jcrm.v1i1.267
Castrillon J., Bengtson Nash S. (2020). Evaluating cetacean body condition; a review of traditional approaches and new developments. Ecol. Evol. 10, 6144–6162. doi: 10.1002/ece3.v10.12
Caurant F., Amiard-Triquet C. (1995). Cadmium contamination in pilot whales Globicephala melas: source and potential hazard to the species. Mar. pollut. Bull. 30, 207–210. doi: 10.1016/0025-326X(94)00126-T
Ciesielski T., Szefer P., Bertenyi Z., Kuklik I., Skóra K., Namiesnik J., et al. (2006). Interspecific distribution and co-associations of chemical elements in the liver tissue of marine mammals from the Polish Economical Exclusive Zone, Baltic Sea. Environ. Int. 32, 524–532. doi: 10.1016/j.envint.2005.12.004
Clarke R. (2006). The origin of ambergris. Latin Am. J. Aquat. Mammals 5, 7–21. doi: 10.5597/lajam00087
Clauss M., Lischke A., Botha H., Hatt J.-M. (2016). Carcass consumption by domestic rabbits (Oryctolagus cuniculus). Eur. J. Wildl Res. 62, 143–145. doi: 10.1007/s10344-015-0980-y
De Groot A. V., Janssen G. M., Isermann M., Stock M., Glahn M., Elschot K., et al. (2017). “Beaches and dunes,” in Wadden Sea Quality Status Report. Ed. Kloepper S., et al (Common Wadden Sea Secretariat, Wilhelmshaven, Germany).
DeVault T. L., Rhodes O. E., Shivik J. A. (2003). Scavenging by vertebrates: Behavioral, ecological, and evolutionary perspectives on an important energy transfer pathway in terrestrial ecosystems. Oikos 102, 225–234. doi: 10.1034/j.1600-0706.2003.12378.x
Domínguez R., Pateiro M., Gagaoua M., Barba F. J., Zhang W., Lorenzo J. M. (2019). A comprehensive review on lipid oxidation in meat and meat products. Antioxidants 8, 429. doi: 10.3390/antiox8100429
Endo T., Haraguchi K., Sakata M. (2002). Mercury and selenium concentrations in the internal organs of toothed whales and dolphins marketed for human consumption in Japan. Sci. Total Environ. 300, 15–22. doi: 10.1016/S0048-9697(02)00137-7
Endo T., Hotta Y., Haraguchi K., Sakata M. (2003). Mercury contamination in the red meat of whales and dolphins marketed for human consumption in Japan. Environ. Sci. Technol. 37, 2681–2685. doi: 10.1021/es034055n
Evans P. G. H. (2013). Proceedings of the ECS/ASCOBANS/ACCOBAMS Joint Workshop on chemical pollution and marine mammals. Ed. Evans P. G. H. (Cádiz, Spain: European Cetacean Society).
Gajdosechova Z., Lawan M. M., Urgast D. S., Raab A., Scheckel K. G., Lombi E., et al. (2016). In vivo formation of natural HgSe nanoparticles in the liver and brain of pilot whales. Sci. Rep. 6, 34361. doi: 10.1038/srep34361
Gamarra-Toledo V., Plaza P. I., Gutiérrez R., Inga-Diaz G., Saravia-Guevara P., Pereyra-Meza O., et al. (2023). Mass mortality of sea lions caused by highly pathogenic avian influenza A(H5N1) virus. Emerg. Infect. Dis. 29, 2553–2556. doi: 10.3201/eid2912.230192
Geraci J. R., Lounsbury V. J. (1993). Marine mammals ashore: a field guide for strandings (College Station, Texas: Publication TAMU-SG-93-601: Texas A&M University Sea Grant College Program).
Glover A. G., Higgs N. D., Bagley P. M., Carlsson R., Davies A. J., Kemp K. M., et al. (2013). Bone-eating worms from the Antarctic: the contrasting fate of whale and wood remains on the Southern Ocean seafloor. Proc. R. Soc. B: Biol. Sci. 280, 20131390. doi: 10.1098/rspb.2013.1390
Greenslade P. J. M. (1964). Pitfall trapping as a method for studying populations of Carabidae (Coleoptera). J. Animal Ecol. 33 (2), 301-310
Hansen A. M., Bryan C. E., West K., Jensen B. A. (2016). Trace element concentrations in liver of 16 species of cetaceans stranded on Pacific Islands from 1997 through 2013. Arch. Environ. Contam Toxicol. 70, 75–95. doi: 10.1007/s00244-015-0204-1
Hansen C. T., Nielsen C. O., Dietz R., Hansen M. M. (1990). Zinc, cadmium, mercury and selenium in minke whales, belugas and narwhals from West Greenland. Polar Biol. 10, 529–539. doi: 10.1007/BF00233702
Heiss J. W. (2020). Whale burial and organic matter impacts on biogeochemical cycling in beach aquifers and leachate fluxes to the nearshore zone. J. Contam Hydrol 233, 103656. doi: 10.1016/j.jconhyd.2020.103656
Hesp P. A. (2004). “Coastal dunes in the tropics and temperate regions: location, formation, morphology and vegetation processes,” in Coastal Dunes, Ecology and Conservation. Eds. Martínez M. L., Psuty N. P. (Springer Verlag, Berlin Heidelberg).
Hohbein R. R., Conway C. J. (2018). Pitfall traps: A review of methods for estimating arthropod abundance. Wildl Soc. Bull. 42, 597–606. doi: 10.1002/wsb.928
Holsbeek L., Joiris C. R., Debacker V., Ali I. B., Roose P., Nellissen J.-P., et al. (1999). Heavy metals, organochlorines and polycyclic aromatic hydrocarbons in sperm whales stranded in the southern North Sea during the 1994/1995 winter. Mar. pollut. Bull. 38, 304–313. doi: 10.1016/S0025-326X(98)00150-7
Huijbers C. M., Schlacher T. A., Schoeman D. S., Weston M. A., Connolly R. M. (2013). Urbanisation alters processing of marine carrion on sandy beaches. Landsc Urban Plan 119, 1–8. doi: 10.1016/j.landurbplan.2013.06.004
IJsseldijk L. L., Brownlow A. C., Mazzariol S. (2019). European best practice on cetacean postmortem investigation and tissue sampling. doi: 10.31219/osf.io/zh4ra
Keijl G. O., Bakker Paiva M. F., IJsseldijk L. L., Kamminga P. (2021). Cetaceans stranded in the Netherlands in 2015-2019. Lutra 64, 19–44.
Knapp A., Blair J. M., Briggs J. M., Collins S. L., Hartnett D. C., Johnson L. C., et al. (1999). The keystone role of bison in North American tallgrass prairie: Bison increase habitat heterogeneity and alter a broad array of plant, community, and ecosystem processes. Bioscience 49, 39–50. doi: 10.2307/1313492
Law R. J., Morris R. J., Allchin C. R., Jones B. R. (1997). Metals and chlorobiphenyls in tissues of sperm whales (Physeter macrocephalus) and other cetacean species exploiting similar diets. Bull. Kon. Bel. Inst. Natuurwet. 67, 79–89.
Law R. J., Stringer R. L., Allchin C. R., Jones B. R. (1996). Metals and organochlorines in sperm whales (Physeter macrocephalus) stranded around the North Sea during the 1994/1995 winter. Mar. pollut. Bull. 32, 72–77. doi: 10.1016/0025-326X(95)00182-M
Li Q., Liu Y., Li G., Wang Z., Zheng Z., Sun Y., et al. (2022). Review of the impact of Whale Fall on biodiversity in deep-sea ecosystems. Front. Ecol. Evol. 10, 885572. doi: 10.3389/fevo.2022.885572
Lischka A., Betty E. L., Braid H. E., Pook C. J., Gaw S., Bolstad K. S. R. (2021). Trace element concentrations, including Cd and Hg, in long-finned pilot whales (Globicephala melas edwardii) mass stranded on the New Zealand coast. Mar. pollut. Bull. 165, 112084. doi: 10.1016/j.marpolbul.2021.112084
Londo G. (1976). The decimal scale for releves of permanent quadrats. Vegetatio 33, 61–64. doi: 10.1007/BF00055300
Meynecke J.-O., Zigic S., Perez L., Dunn R. J. K., Benfer N., Gustafson J., et al. (2024). Dead on the beach? Predicting the drift of whale remains improves management for offshore disposal. J. Mar. Sci. Eng. 12, 1156. doi: 10.3390/jmse12071156
Milne L. J., Milne M. (1976). The social behavior of burying beetles. Sci. Am. 235, 84–89. doi: 10.2307/24950419
MinLNV (2020). “Leidraad stranding levende grote walvisachtigen,” in Gravenhage: Ministerie van Landbouw, Natuur en Voedselkwaliteit (LNV), DG Natuur, Visserij en Landelijk Gebied.
MinVenW (2007). Samenwerkingsregeling Bestrijding Kustverontreiniging RWS-diensten (Rijswijk: Ministerie van Verkeer en Waterstaat, Rijkswaterstaat Noordzee).
Mirolo M., Pohlmann A., Ahrens A. K., Kühl B., Rubio-Garcìa A., Kramer K., et al. (2023). Highly pathogenic avian influenza A virus (HPAIV) H5N1 infection in two European grey seals (Halichoerus grypus) with encephalitis. Emerg. Microbes Infect. 12, e2257810. doi: 10.1080/22221751.2023.2257810
Mol G., Spijker J. H., van Gaans P., Römkens P. F. A. M. (2012). Geochemische bodematlas van Nederland (Wageningen: Wageningen Academic Publishers).
Moore M. J., Mitchell G. H., Rowles T. K., Early G. (2020). Dead cetacean? Beach, bloat, float, sink. Front. Mar. Sci. 7, 333. doi: 10.3389/fmars.2020.00333
Olea P. P., Mateo-Tomás P., Sánchez-Zapata J. A. (2019). Carrion Ecology and Management. Eds. Olea P. P., Mateo-Tomás P., Sánchez-Zapata J. A. (Cham, Switzerland: Springer).
Puryear W., Sawatzki K., Hill N., Foss A., Stone J. J., Doughty L., et al. (2023). Highly pathogenic avian influenza A(H5N1) virus outbreak in new England seals, United States. Emerg. Infect. Dis. 29, 786–791. doi: 10.3201/eid2904.221538
Quaggiotto M. M., Evans M. J., Higgins A., Strong C., Barton P. S.. (2019). Dynamic soil nutrient and moisture changes under decomposing vertebrate carcasses. Biogeochemistry 146, 71–82. doi: 10.1007/s10533-019-00611-3
Quaggiotto M. M., Sánchez-Zapata J. A., Bailey D. M., Payo-Payo A., Navarro J., Brownlow A., et al. (2022). Past, present and future of the ecosystem services provided by cetacean carcasses. Ecosyst. Serv. 54, 101406. doi: 10.1016/j.ecoser.2022.101406
R Core Team (2024). R: A Language and Environment for Statistical Computing (Vienna, Austria: R Foundation for Statistical Computing). Available at: https://www.R-project.org.
Reisdorf A. G., Bux R., Wyler D., Benecke M., Klug C., Maisch M. W., et al. (2012). Float, explode or sink: postmortem fate of lung-breathing marine vertebrates. Paleobiodivers Paleoenviron 92, 67–81. doi: 10.1007/s12549-011-0067-z
Reise K., Baptist M. J., Burbridge P., Dankers N., Fischer L., Flemming B., et al. (2010). The Wadden Sea - a universally outstanding tidal wetland (Wilhelmshaven, Germany: Common Wadden Sea Secretariat, Wadden Sea Ecosystem No. 29).
Schlacher T. A., Strydom S., Connolly R. M. (2013). Multiple scavengers respond rapidly to pulsed carrion resources at the land–ocean interface. Acta Oecologica 48, 7–12. doi: 10.1016/j.actao.2013.01.007
Shin D.-L., Siebert U., Lakemeyer J., Grilo M., Pawliczka I., Wu N.-H., et al. (2019). Highly pathogenic avian influenza A(H5N8) virus in Gray Seals, Baltic Sea. Emerg. Infect. Dis. 25, 2295–2299. doi: 10.3201/eid2512.181472
Smith C. R., Baco A. R. (2003). Ecology of whale falls at the deep-sea floor. Oceanography Mar. Biology: an Annu. Rev. 41, 311–354.
Sousa A., Brito C. (2012). Historical strandings of cetaceans on the Portuguese coast: anecdotes, people and naturalists. Mar. Biodivers Rec 5. doi: 10.1017/S1755267211000959
Sun W., Li X., Padilla J., Elbana T. A., Selim H. M. (2019). The influence of phosphate on the adsorption–desorption kinetics of vanadium in an acidic soil. J. Environ. Qual 48, 686–693. doi: 10.2134/JEQ2018.08.0316
Tavares M., Alvares D. J., Borges-Martins M. (2021). Spatiotemporal patterns of carrion biomass of marine tetrapods at the ocean–land interface on the southern Brazilian coastline. Mar. Ecol. Prog. Ser. 672, 57–72. doi: 10.3354/MEPS13812
Tobajas J., Descalzo E., Ferreras P., Mateo R., Margalida A. (2021). Effects on carrion consumption in a mammalian scavenger community when dominant species are excluded. Mamm. Biol. 101, 851–859. doi: 10.1007/s42991-021-00163-w
Tucker J. P., Santos I. R., Crocetti S., Butcher P. (2018). Whale carcass strandings on beaches: Management challenges, research needs, and examples from Australia. Ocean Coast. Manag 163, 323–336. doi: 10.1016/j.ocecoaman.2018.07.006
Tucker J. P., Santos I. R., Davis K. L., Butcher P. A. (2019). Whale carcass leachate plumes in beach groundwater: A potential shark attractant to the surf? Mar. pollut. Bull. 140, 219–226. doi: 10.1016/j.marpolbul.2019.01.043
Turner K. L., Abernethy E. F., Conner L. M., Rhodes O. E., Beasley J. C. (2017). Abiotic and biotic factors modulate carrion fate and vertebrate scavenging communities. Ecology 98, 2413–2424. doi: 10.1002/ecy.2017.98.issue-9
UNEP (1991). ICES/JOC/UNEP Review of contaminant in marine mammals (Nairobi: United Nations Environment Programme).
van Klink R., van-Laar-Wiersma J., Vorst O., Smit C. (2020). Rewilding with large herbivores: Positive direct and delayed effects of carrion on plant and arthropod communities. PLoS One 15, e0226946. doi: 10.1371/journal.pone.0226946
Verheggen F., Perrault K. A., Megido R. C., Dubois L. M., Francis F., Haubruge E., et al. (2017). The odor of death: an overview of current knowledge on characterization and applications. Bioscience 67, 600–613. doi: 10.1093/biosci/bix046
Wagemann R., Muir D. C. G. (1984). Concentrations of heavy metals and organochlorines in marine mammals of Northern waters: Overview and evaluation. Can. Tech Rep. Fish Aquat Sci. 1279, 97.
Wenting E., Jansen P. A., Laugeman M. J. B., van Langevelde F. (2023). Leakage of nutrients into the soil due to carrion decomposition can enhance plant growth. J. Soil Sci. Plant Nutr. 23, 6874–6879. doi: 10.1007/S42729-023-01430-0/FIGURES/2
Wiedemann A. M., Pickart A. J. (2004). “Temperate zone coastal dunes,” in Coastal Dunes, Ecology and Conservation. Eds. Martínez M. L., Psuty N. P. (Springer-Verlag, Berlin Heidelberg).
Wilson E. E., Wolkovich E. M. (2011). Scavenging: How carnivores and carrion structure communities. Trends Ecol. Evol. 26, 129–135. doi: 10.1016/j.tree.2010.12.011
Wu J., Liang J., Björn L. O., Li J., Shu W., Wang Y. (2022). Phosphorus-arsenic interaction in the ‘soil-plant-microbe’ system and its influence on arsenic pollution. Sci. Total Environ. 802, 149796. doi: 10.1016/J.SCITOTENV.2021.149796
Zamora R., Hidalgo F. J. (2005). Coordinate contribution of lipid oxidation and Maillard reaction to the nonenzymatic food browning. Crit. Rev. Food Sci. Nutr. 45, 49–59. doi: 10.1080/10408690590900117
Keywords: biodiversity, cetaceans, decomposition, management, North Sea, stranding, whale carcass
Citation: Baptist MJ, Leopold MF, van Puijenbroek MEB, Verdaat H, Janinhoff-Verdaat N, Lammertsma DR, Dimmers WJ, Römkens PFAM and Burgers J (2025) Decomposition of a minke whale carcass in a temperate dune ecosystem. Front. Mar. Sci. 12:1474460. doi: 10.3389/fmars.2025.1474460
Received: 01 August 2024; Accepted: 17 February 2025;
Published: 05 March 2025.
Edited by:
Mark E Bond, Florida International University, United StatesReviewed by:
Brian Kot, City University of Hong Kong, Hong Kong SAR, ChinaCopyright © 2025 Baptist, Leopold, van Puijenbroek, Verdaat, Janinhoff-Verdaat, Lammertsma, Dimmers, Römkens and Burgers. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Martin J. Baptist, bWFydGluLmJhcHRpc3RAd3VyLm5s
Disclaimer: All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article or claim that may be made by its manufacturer is not guaranteed or endorsed by the publisher.
Research integrity at Frontiers
Learn more about the work of our research integrity team to safeguard the quality of each article we publish.