 Pengfei Liu
Pengfei Liu Kuan Jiang1
Kuan Jiang1 Fajin Chen
Fajin Chen Renming Jia
Renming Jia Chao Wang
Chao Wang- 1College of Oceanology and Meteorology, Guangdong Ocean University, Zhanjiang, China
- 2Guangxi Key Laboratory of Beibu Gulf Marine Resources, Environment and Sustainable Development, Fourth Institute of Oceanography, Ministry of Natural Resources, Beihai, China
- 3Key Laboratory of Space Marine Remote Sensing and Application, Ministry of Natural Resources, Beijing, China
- 4Laboratory for Coastal Ocean Variation and Disaster Prediction, College of Oceanology and Meteorology, Guangdong Ocean University, Zhanjiang, China
The export and transformation of riverine dissolved organic matter (DOM) at the river-sea interface are critical to understanding carbon budgets in tropical regions, yet remain underexplored in small watersheds. This study analyzed dissolved organic carbon (DOC), chromophoric (CDOM), and fluorescent DOM (FDOM) over five cruises from summer 2021 to summer 2022 in the Suixi Estuary-Zhanjiang Bay continuum, a eutrophic tropical transition zone in the northwestern South China Sea. Seasonal variations were pronounced for DOM in the continuum and two endmembers. Freshwater DOM, dominated by terrestrial organic materials, was closely tied to soil leaching and erosion, correlating with precipitation patterns. Conversely, seawater DOM, enriched in protein-like FDOM, varied with the West-Guangdong Coastal Current and its associated primary production. Linear decreases in DOM with increasing salinity indicate that conservative mixing largely shapes DOM distributions in the continuum, especially for humic-like FDOM. Non-conservative processes, including flocculation, phytoplankton production, and microbial transformation, variably alter DOM components across seasons. The Suixi River in summer contributed considerably but disproportionately to the annual DOM flux (47-59%), due to intense soil leaching processes that mobilize refractory terrigenous DOM. The deviation observed in the Suixi River’s DOC-CDOM relationship from that of global rivers, coupled with a distinctive single-peak pattern between the carbon-specific absorption coefficient and water yield in global rivers, highlights the critical role of watershed nature in influencing river-exported DOM composition. The notably high DOC and CDOM yields for the Suixi River further emphasize the importance of small, tropical rivers in shaping the estuarine and coastal carbon budget.
1 Introduction
Estuarine and coastal areas play a crucial role in regulating the carbon cycle by controlling the nature and magnitude of terrigenous carbon entering the ocean (Bauer et al., 2013; García-Martín et al., 2021). Dissolved organic matter (DOM), a major reduced carbon pool in these regions, is critical for ecological and biogeochemical functions (Bauer and Bianchi, 2011; Dai et al., 2022). DOM mainly originates from allochthonous sources such as river runoff (Spencer and Raymond, 2024), groundwater discharge (McDonough et al., 2022), atmospheric deposition (Safieddine and Heald, 2017), and anthropogenic activities (Guo et al., 2014), as well as autochthonous production through in situ biological activities (Carlson et al., 2024). Given the heterogeneous nature of climate, geography, hydrodynamics, biogeochemical backgrounds, and human pressures, the source, sink, transport, and transformation of DOM in estuarine and coastal areas vary significantly across both temporal and spatial dimensions (Guo et al., 2007, 2014; García-Martín et al., 2021). This variability introduces substantial uncertainties in quantifying the carbon budgets on regional and global scales (Bauer et al., 2013).
River runoff is a major source of allochthonous DOM to the ocean, contributing approximately 260 to 300 Tg C/yr globally (Spencer and Raymond, 2024; Liu et al., 2024). Small rivers, though often overlooked, typically exhibit higher concentrations of dissolved organic carbon (DOC) and contribute significantly to the global flux, especially in tropical regions (Raymond and Spencer, 2015). Tropical rivers have displayed an increasing trend in DOC output over recent decades driven by rising terrestrial organic carbon inputs, even as the global riverine DOC flux generally declines (Li et al., 2019a). Despite their importance, the dynamics of DOC transport by tropical rivers—particularly smaller ones—and their impact on estuaries and coastal bays are not well-studied. This gap is significant considering the vast number of these rivers, their marked seasonality, and their high sensitivity to climate change and human activities (Liu et al., 2024; Spencer and Raymond, 2024).
Riverine DOM, originating from soil and vegetation (i.e., terrigenous DOM), aquatic biological production, and anthropogenic inputs, exhibits considerable diversity in quality, quantity, and biogeochemical reactivities (Li et al., 2019a). Terrigenous DOM, rich in highly unsaturated compounds like lignin phenols and humic acid (Lehmann and Kleber, 2015; Osterholz et al., 2016), predominates in riverine transport to estuaries and coastal bays (Wen et al., 2021). Biological activities (e.g., phytoplankton production, microbial metabolism) could contribute both simple bio-labile molecules (e.g., amino acids, carbohydrate, Rochelle-Newall and Fisher, 2002; Medeiros et al., 2015) and complex bio-refractory molecules (humic substances, Romera-Castillo et al., 2010; Jiao et al., 2024), accounting for ~20% of riverine DOM flux in Chinese rivers (Wen et al., 2021). Anthropogenic inputs could contribute substantial bioavailable DOM to estuarine and coastal systems, especially in regions experiencing increased human activities (Guo et al., 2014; Tzortziou et al., 2015; Wen et al., 2021). Optical analyses results have shown that terrigenous and microbial-derived DOM in abundant in chromophore (i.e., chromophoric DOM, CDOM) and humic-like fluorophores (i.e., fluorescent DOM, FDOM) (Guo et al., 2011, 2014), while the bio-labile DOM components from primary production and anthropogenic inputs are more enriched in protein-like fluorophores (i.e., FDOMP, Guo et al., 2014; Romera-Castillo et al., 2010; Tzortziou et al., 2015). Qualitive optical parameters, such as spectral slope, carbon-normalized absorption coefficient, humification degree, and freshness, provide insights into the source, nature, fate of DOM from land to sea (Helms et al., 2008; Hansen et al., 2016). The optical information has demonstrated a high potential for photochemical and microbial degradations of riverine DOM, thereby influencing its biogeochemical transformation in estuaries and coastal bays (Fichot and Benner, 2014), as well as the actual flux of DOM to the sea (Liu et al., 2024).
Previous studies have shown diverse mixing behaviors of DOM in estuaries and coastal bays across different compounds, spatially and temporally (Guo et al., 2011; Osterholz et al., 2016; Massicotte et al., 2017). Allochthonous CDOM and humic-like FDOM (FDOMH) often exhibit conservative mixing behaviors in various estuaries and coastal bays, as seen in the Mississippi and Atchafalaya River plume regions (Chen and Gardner, 2004), Yangtze River estuary (Guo et al., 2007; 2014), Pearl River estuary (Li et al., 2019b), Jiulong River estuary (Guo et al., 2011), among others (Yamashita et al., 2008; Markager et al., 2011; Zhao et al., 2021). Despite their resistance to microbial utilization (Jiao et al., 2024), these components are susceptible to photodegradation in estuaries and coastal environments (Moran et al., 2000), and may rapidly diminish under favorable conditions of residence time and sunlight exposure (Fichot and Benner, 2014). The priming effect may also accelerate the biodegradation of terrigenous refractory DOM in these regions (Letscher et al., 2011; Bianchi, 2011; Zhuang et al., 2021; Spencer and Raymond, 2024). Additionally, abiotic processes (e.g., sorption-desorption, flocculation) can significantly alter DOM level and compositions in estuaries and coastal environments (McCallister et al., 2006; Asmala et al., 2014). Hence, allochthonous inputs, along with biotic and abiotic transformations, substantially shape the composition, flux, and fate of riverine DOM as it travels from land to sea. These dynamics have been extensively studied in major estuaries like those of Yangtze River estuary and Pearl River estuary (Guo et al., 2014; Li et al., 2019b). However, there remains a significant gap in research across the small river-estuary-coastal bay continuum, especially in tropical regions (Spencer and Raymond, 2024).
In this study, DOC, CDOM and FDOM were measured in the Suixi Estuary-Zhanjiang Bay continuum, a typical tropical estuary and semi-closed bay in the northwestern South China Sea, from summer 2021 to summer 2022. Together with the hydrological and environmental parameters, we aim to (1) investigate seasonal variations of DOM in the freshwater and seawater endmembers; (2) elucidate the distributions and mixing behaviors of DOM using a two-endmember mixing model; and (3) compare the composition, sources, and fluxes of riverine DOM with those from other global rivers.
2 Materials and methods
2.1 Study region
Suixi Estuary-Zhanjiang Bay continuum, situated in the northwest South China Sea (SCS), stands as a tropical estuarine and coastal bay heavily influenced by human activities. Geographically, the continuum can be divided into two distinct areas: the upper Suixi Estuary and the lower Zhanjiang Bay (Figure 1). The upper estuary experiences a substantial impact from freshwater discharge, predominantly from the Suixi River, with an average annual runoff of 10.4 × 108 m3. This discharge plays a significant role in shaping the upper continuum’s hydrology. In addition, the upper continuum serves as a central hub for oyster breeding activities and is burdened with significant pollution (Lao et al., 2022). Conversely, the lower Zhanjiang Bay is strongly influenced by seawater intrusion from the northwestern South China Sea (SCS), through a narrow channel (~2 km wide), consistently affected by the West-Guangdong Coastal Current (WGCC) persists outside Zhanjiang Bay year-round. During the summer months, this intrusion may intensify pollutant retention within the bay (Lao et al., 2023a). In recent years, there has been a noticeable surge in the discharge of industrial sewage, aquaculture wastewater, and ship navigation wastewater into Zhanjiang Bay, contributing significantly to its pollution load (Zhang et al., 2021). Additionally, the bay’s slow water exchange rate promotes nutrient accumulation, exacerbating eutrophication and further degrading water quality (Liang et al., 2024). Consequently, the complex interaction of seasonal variations in river discharge, intrusion of the SCS waters, and human activities results in significant fluctuations in the hydrology, environment, and ecology of Zhanjiang Bay.

Figure 1. Map of sampling stations in the Suixi Estuary-Zhanjiang Bay continuum. WGCC represents the West-Guangdong Coastal Current.
2.2 Field observation and sample collection
Five seasonal cruises were conducted in Zhanjiang Bay, located in the northwestern SCS, on the following dates: June 26-30, 2021 (Summer 2021); September 29-October 3, 2021 (Autumn 2021); December 7-10, 2021 (Winter 2021); March 19-21, 2022 (Spring 2022); and June 23-25, 2022 (Summer 2022). Temperature and salinity were measured at twenty-three stations using a calibrated SBE 37 CTD unit (Seabird, USA). Surface water samples (0.5m depth) were collected at all stations using a Niskin bottle, from the upper Suixi River to the offshore SCS. Bottom water samples were obtained at the depth of 1m above the seafloor. Water samples were first collected for dissolved oxygen (DO) analysis. Samples for inorganic nutrient analyses were filtered through MF-Millipore™ filters (0.45 μm) and stored frozen until analysis. Samples for chlorophyll a (Chl-a) and DOM analyses were filtered immediately through pre-combusted (500°C, 5 h) GF/F filters (nominal pore size ∼0.7 μm) under a gentle vacuum. Filter membranes for Chl-a samples were stored frozen until analysis. Filtrates were transferred into pre-combusted glass vials with Teflon-lined caps and stored frozen for analyses of DOC, CDOM and FDOM. Analyses of DO and ammonium were completed within 8 hours, and other analyses were conducted within 24 hours of sampling.
2.3 Measurements of environmental parameters
The DO concentrations were determined using the Winkler titration method (Carpenter, 1965), achieving a precision of 0.07 mg/L. Chl-a was extracted from samples using 90% aqueous acetone, shielded from light, and incubated at 4°C for 24 hours. The Chl-a concentrations were measured with a precision of 0.3 μg/L using a Turner Trilogy fluorometer (Welschmeyer, 1994). The concentrations of nitrate (, nitrite , and soluble reactive phosphate (SRP) were determined using a San++ continuous flow analyzer (Skalar, Netherlands). Ammonium (NH4+-N) concentration was measured via spectrophotometry. Dissolved inorganic nitrogen (DIN) was calculated by summing the concentrations of , , and NH4+-N. The nitrogen-to-phosphorus ratio (N/P, mol/mol) was computed as the ratio of DIN to SRP, providing additional insights into the nutrient dynamics. The detection limits for DIN and SRP were 0.05 μmol/L with an analytical precision was better than ± 2%.
2.4 Measurements of DOC
DOC concentrations were determined using a TOC-LCPH analyzer operating in high-temperature catalytic oxidation mode (Shimadzu, Japan). A five-point standard curve was established using potassium hydrogen phthalate standards. DOC concentrations were calculated by first subtracting the running blank, determined as the average peak area of Milli-Q water acidified with HCl, from the average peak area of the samples (injected 2–3 times) and then dividing by the slope of the standard curve. The precision of the DOC analysis was maintained at ±1.0 μmol/L, as validated by DOC Consensus Reference Material (CRM) provided by D.A. Hansell from the University of Miami (https://hansell-lab.earth.miami.edu/consensus-reference-material/index.html). The coefficient of variation for DOC analysis, based on replicate measurements, was approximately 2%.
2.5 CDOM and FDOM analyses
The absorption spectrum of CDOM was measured at room temperature using a UV-1780 ultraviolet-visible spectrophotometer (Shimadzu, Japan) equipped with a pair of 10-cm quartz cuvettes (L = 0.1 m). Fresh Milli-Q water was served as a blank, with one Milli-Q water sample analyzed after every 5 samples to monitor any potential shifts in the blank. The spectrophotometer scanned from 800 nm to 240 nm at intervals of 0.5 nm. Absorbance values (Aλ) were corrected by subtracting the Milli-Q water blank and were then converted to Napierian absorption coefficients, denoted as aλ (m-1), using the following formula:
In this study, we report the Napierian absorption coefficients at 350 nm (a350) to quantify CDOM in the Suixi Estuary-Zhanjiang Bay continuum. The choice of a350 is due to its frequent use in studies across the land-sea interface (Massicotte et al., 2017). Additionally, the spectral slope between 275–295 nm (S275-295, μm-1) was determined through linear regression of the natural log-transformed absorption spectra (Helms et al., 2008). For broader comparisons, CDOM absorption coefficients at other commonly used wavelengths (e.g., a254, a280, a300, a325, a355, and a412), along with the spectral slope over the range of 350–400 nm (S350-400), are included in Supplementary Table S1. Furthermore, specific ultraviolet absorbance at 254 nm (SUVA254, m2/g C) was calculated by dividing the decadal absorption coefficient (i.e., A254/0.1) by the dissolved organic carbon (DOC) concentration, expressed in mg/L (Weishaar et al., 2003).
Fluorescence excitation-emission matrices (EEMs) of FDOM were measured using an F-7100 fluorophotometer with a 1-cm quartz cuvette at room temperature (Hitachi, Japan). Emission (em) scans were conducted from 280 to 600 nm at 2-nm intervals, paired with excitation (ex) wavelengths ranging from 240 to 450 nm at 5-nm intervals. Both the em and ex slit widths were set to 5 nm. The inner filter effects were corrected following an absorbance-based method (Kothawala et al., 2013). EEMs were then corrected for blank values and normalized using freshly prepared Milli-Q water. Fluorescence intensity was expressed in Raman unit (RU, Lawaetz and Stedmon, 2009).
A total of 203 calibrated EEMs were decomposed using parallel factor analysis (PARAFAC) on MATLAB 2020b with the DOM Fluor toolbox (Stedmon et al., 2003). The model was constrained to nonnegative values, and robustness was confirmed via split-half analysis. Model validation extended to comparing results with published fluorescent components from the Open Fluor database (https://openfluor.lablicate.com/, accessed August 8, 2024; Murphy et al., 2014), using the Tucker Congruence Coefficient (TCC) (Wang et al., 2022). Four qualitative fluorescence parameters derived from EEMs were employed to characterize DOM regarding sources, composition, and reactivity. The humification index (HIX), calculated as the ratio of the integrated emission spectra from 435-480 nm to that from 300-345 nm with excitation at 254 nm, indicating the degree of DOM humification (Zsolnay et al., 1999). The biological index (BIX), computed as the ratio of fluorescence intensity emitted at 380 nm to that emitted at 430 nm upon excitation at 310 nm, providing insights into the presence of fresh biological material in DOM (Huguet et al., 2009). The fluorescence index (FI), calculated as the ratio of emission intensity at 470 nm to that at 520 nm under an excitation of 370 nm, was used to infer DOM source (Cory and McKnight, 2005). Furthermore, the ratio of humic-like fluorescence to protein-like fluorescence (H/P) was calculated, serving as an indicator of the relative abundance of these components within the DOM pool (Hansen et al., 2016).
2.6 River discharge and precipitation data
River discharge data for the Suixi River was source from Wang et al. (2022). Daily precipitation data for the study region was obtained from the China Meteorological Data Service Centre (https://data.cma.cn/). For this study, the cumulative daily precipitation for the 15 days preceding each of the five cruise was calculated.
2.7 Statistical analyses
To assess the significance of differences in environmental and DOM parameters between surface and bottom waters, as well as across different seasons, a two-tailed t-test was applied (α = 0.05), using IBM SPSS Statistics 23. Additionally, Pearson correlation analyses were conducted to explore relationships between DOM variables and salinity, as well as between non-conservative DOM and environmental variables. These analyses were performed using OriginPro 2024b, with significance levels (p-value) determined by a two-tailed test.
3 Results
3.1 Distributions of hydrological and environmental parameters
The Suixi Estuary-Zhanjiang Bay continuum exhibits significant seasonal variations in water temperature, ranging from 20.4°C to 33.4°C throughout the year (Figures 2A–E; Supplementary Figures S1A–E). The highest temperatures were recorded during the summer and autumn, contrasting with the cooler temperatures observed in winter and spring (Table 1). Surface water temperatures were slightly higher than those at the bottom of the continuum (t-test, p< 0.05), except for summer 2021 (t-test, p > 0.05). Salinity also showed significant seasonal fluctuations, ranging from 0.4 to 33.1, with lower values typically in summer and autumn and higher in winter and spring. (Table 1; Figures 2F–J; Supplementary Figures S1F–J). A significant difference in salinity between surface and bottom waters is evident across all seasons (t-test, p< 0.05). This variation was more pronounced in the upper estuary compared to the lower bay (Figures 2F–J; Supplementary Figures S1F–J).
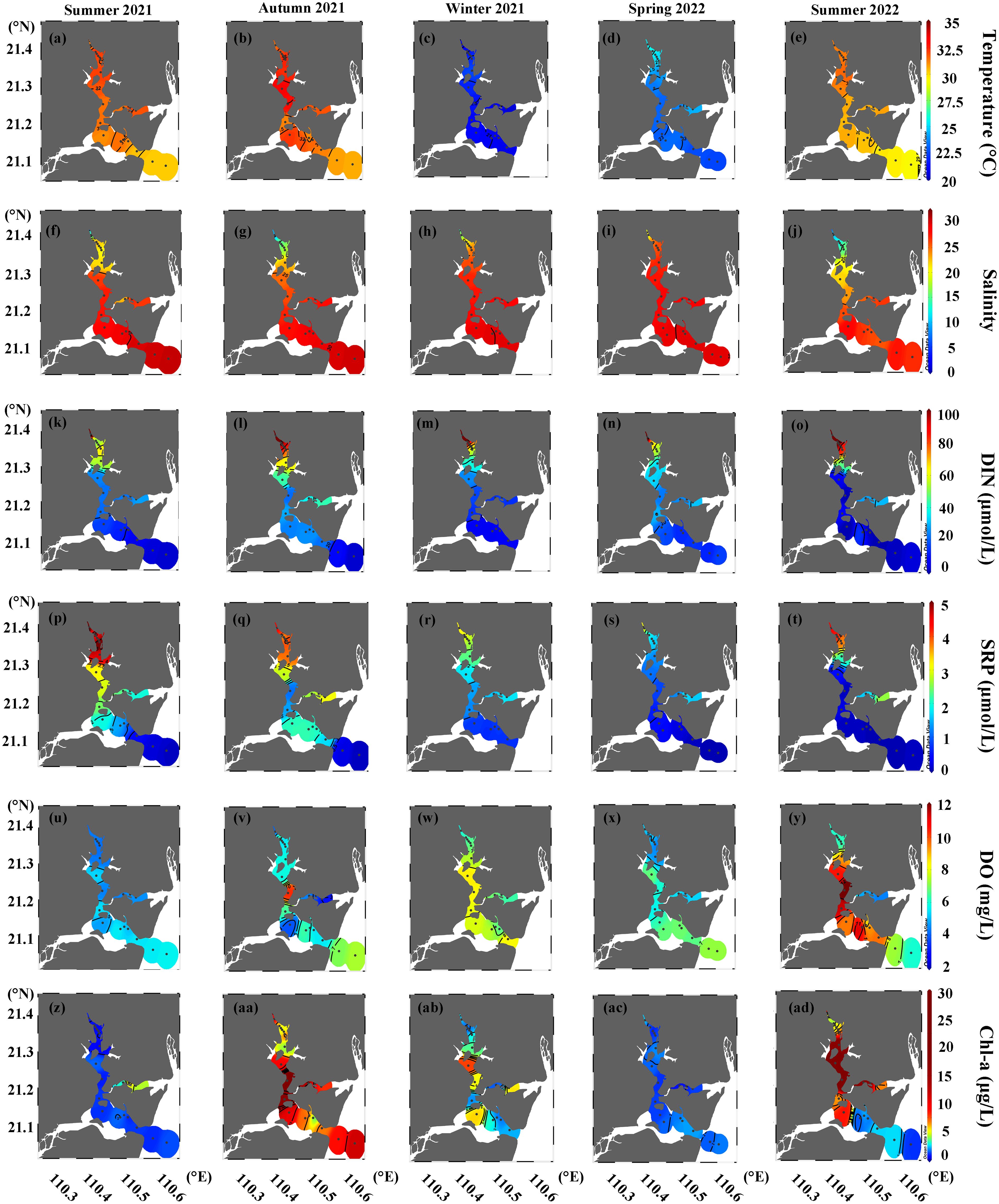
Figure 2. Seasonal distributions of (A–E) temperature, (F–J) salinity, (K–O) DIN, (P–T) SRP, (U–Y) DO, and (Z–AD) Chl-a in surface waters of the Suixi Estuary-Zhanjiang Bay continuum.
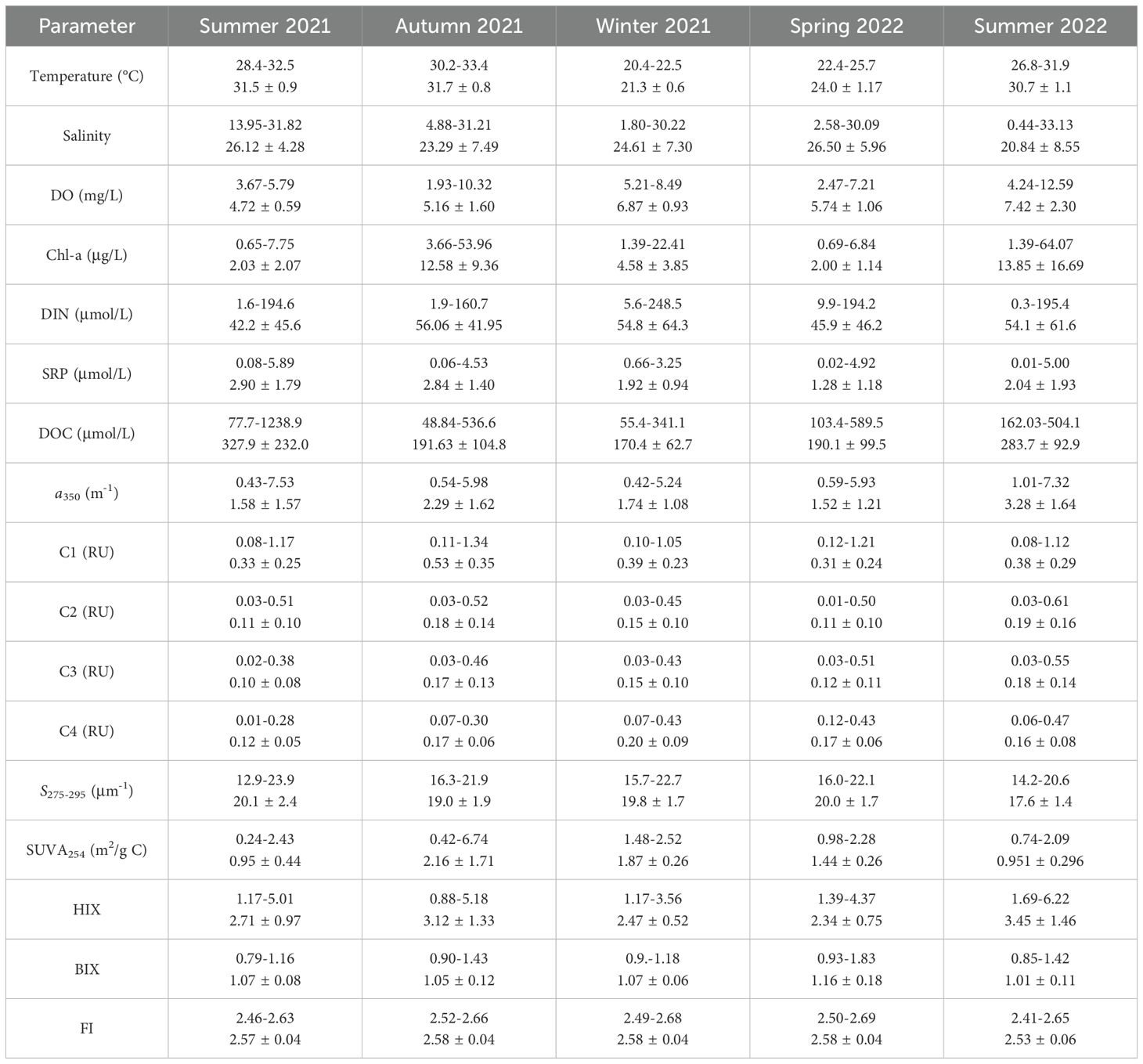
Table 1. Ranges and mean values (± SD) of hydrological, environmental, and DOM parameters across fiver seasons in the Suixi Estuary-Zhanjiang Bay continuum.
DIN concentrations in the continuum varied widely from 0.3 to 248.5 μmol/L, peaking in autumn 2021 and reaching the lowest in summer 2021 (Table 1; Figures 2K–O; Supplementary Figures S1K–O). SRP concentrations fluctuated between 0.01 and 5.89 μmol/L, with the highest average during summer 2021 and the lowest during spring 2022 (Table 1). Notable differences between surface and bottom DIN levels were observed during winter 2021 and spring 2022 (t-test, p< 0.05), whereas SRP concentrations showed no significant variation between depths across any season (p > 0.05). Both DIN and SRP concentrations generally peaked near the Suixi River in the upper estuary and decrease towards the seaward direction across all seasons (Figures 2P–T; Supplementary Figures S1P–T).
DO concentrations exhibited seasonal variations ranging from 1.93 mg/L to 12.59 mg/L (Table 1), with peaks in the center of Zhanjiang Bay during autumn 2021 and summer 2022 (Figures 2U–Y; Supplementary Figures S1U-Y). In these seasons, surface water DO concentrations were significantly higher than those in bottom water (t-test, p = 0.008 and p = 0.01, respectively), while no significant differences were noted in other seasons (p > 0.05). Chl-a concentrations range from 0.65 μg/L to 64.07 μg/L, peaking during autumn 2021 and summer 2022, with lower concentrations noted during summer 2021 and spring 2022 (Table 1). Significant disparities in Chl-a concentrations between surface and bottom waters were noted in summer 2021 (t-test, p< 0.05). Corresponding to the observed high DO concentrations, elevated Chl-a levels (> 20 μg/L) were observed in the central continuum during both autumn 2021 and summer 2022 in both surface and bottom waters (Figures 2Z–AD; Supplementary Figures S1Z-AD).
3.2 PARAFAC results
PARAFAC analysis identified four major fluorescent components (Supplementary Figure S2), including three humic-like components (C1, C2, C3) and one protein-like component (C4). C1 exhibited excitation/emission maxima at 270/460 nm, aligning with the typical humic-like peak A (Coble, 2007). C2 displayed two excitation maxima at 270 and 355 nm and one emission maximum at 448 nm, spanning the region of both peak A and peak C, characteristic of UVC and UVA humic fluorophores (Coble, 2007). These components, primarily C1 and C2, are initially considered terrestrial-derived humic-like fluorophores (Coble, 2007), widely distributed in land-ocean interface, strongly associated with soil organic materials and microbial activities (Guo et al., 2014; Li et al., 2019b). C3, with excitation/emission maxima at 305/392 nm, originally identified as a marine-derived humic-like component, is also prevalent across various natural aquatic ecosystems. Biological activities (e.g., microbial activities, phytoplankton production) have been demonstrated to the primary autochthonous sources of C3 in seawater (Yamashita et al., 2008; Romera-Castillo et al., 2010). C4, displaying excitation/emission maxima at 275/340 nm, represents a protein-like component consistent with tryptophan-like peak T (Coble, 2007). It is indicative of autochthonous DOM production and anthropogenic input (Yamashita and Tanoue, 2003; Guo et al., 2014; Yamashita et al., 2015). Comparisons with the OpenFluor database confirmed that four FDOM components in this study were consistent with PARAFAC-derived components from previous studies (Murphy et al., 2014). C1, C2 and C3 have 10, 17 and 28 pairs of match-ups, respectively, with a TCC of >0.95 for both excitation and emission (TCCex,em), suggesting a consistent presence across diverse environments (e.g., Li et al., 2015; Dainard et al., 2019; Gamrani et al., 2023). Protein-like C4, with TCCex,em values exceeding 0.95, was matched in 103 previous studies, including various natural aquatic ecosystems (Yamashita et al., 2010; Gao and Gueguen, 2017), plankton community culture experiments (Stedmon and Markager, 2005; Bittar et al., 2015), and wastewater (Cohen et al., 2014; Huang et al., 2024).
3.3 Distributions of DOC and quantitative optical parameters
DOC concentrations in the Suixi Estuary-Zhanjiang Bay continuum displayed significant variability, ranging from 55.4 to 1235.9 μmol/L. The summer of 2021 recorded the highest mean concentration at 327.9 ± 232.0 μmol/L, while the lowest was in the spring of 2022 at 170.4 ± 62.7 μmol/L (Table 1). A statistically significant difference in DOC concentrations between surface and bottom waters was observed only in the summer of 2022 (t-test, p = 0.01), with higher averages in surface waters. The upper Suixi Estuary consistently showed the highest DOC concentrations, which decreased progressively towards the sea across all seasons. Notably high DOC levels were documented in the central region of Zhanjiang Bay during the summers of 2021 and 2022 (Figures 3A–E; Supplementary Figures S3A–E).
Strong positive correlations were found among CDOM absorption coefficients at various wavelengths (i.e., a254, a280, a300, a325, a350, a355, and a412) in the continuum (r > 0.97, p< 0.0001, n = 195). Thus, for simplicity, this study focuses on the a350 results, which ranged from 0.42 m-1 to 7.32 m-1 (Table 1). The average a350 peaked in summer 2022 (3.28 ± 1.64 m-1) and reached its minimum in spring 2022 (1.52 ± 1.21 m-1), with significant difference noted in summer 2022 between surface and bottom waters (t-test, p = 0.003). The highest a350 values were consistently observed in the upper Suixi estuary, diminishing towards the sea throughout the year. (Figures 3F–J; Supplementary Figures S3F–J).
The intensities of three humic-like components (C1, C2 and C3) demonstrated strong correlations (r > 0.97, p< 0.0001, n = 195), ranging from 0.08-1.17 RU, 0.01-0.61 RU, and 0.02-0.55 RU, respectively. The peak mean intensity of C1 was recorded in autumn 2021 (0.53 ± 0.35 RU), with the lowest occurring in spring 2022 (0.31 ± 0.24 RU). Conversely, the highest mean intensities for C2 and C3 were observed in the summer of 2022, whereas their lowest values were noted in the summer of 2021 (Table 1). The protein-like C4 intensity ranged from 0.01 to 0.47 RU, peaking in winter 2021 (0.20 ± 0.09 RU) and showing the lowest value in summer 2021 (0.12 ± 0.05 RU). The significant differences between surface and bottom waters were detected only in summer 2022 for all components (t-test, p values ranging from 0.002 to 0.006). The intensities of three humic-like components (C1, C2, and C3) were strongly correlated with the a350 (r > 0.83, p< 0.0001, n = 195), exhibiting similar distribution patterns throughout the Suixi Estuary-Zhanjiang Bay continuum (Figures 3K–Y; Supplementary Figures S3K–Y). However, the intensities of protein-like C4 showed a positive correlation with DOC concentrations at each season (r: 0.58-0.95, p< 0.0001) and followed a similar distribution pattern across the continuum (Figures 3Z–AD; Supplementary Figures S3Z–AD).
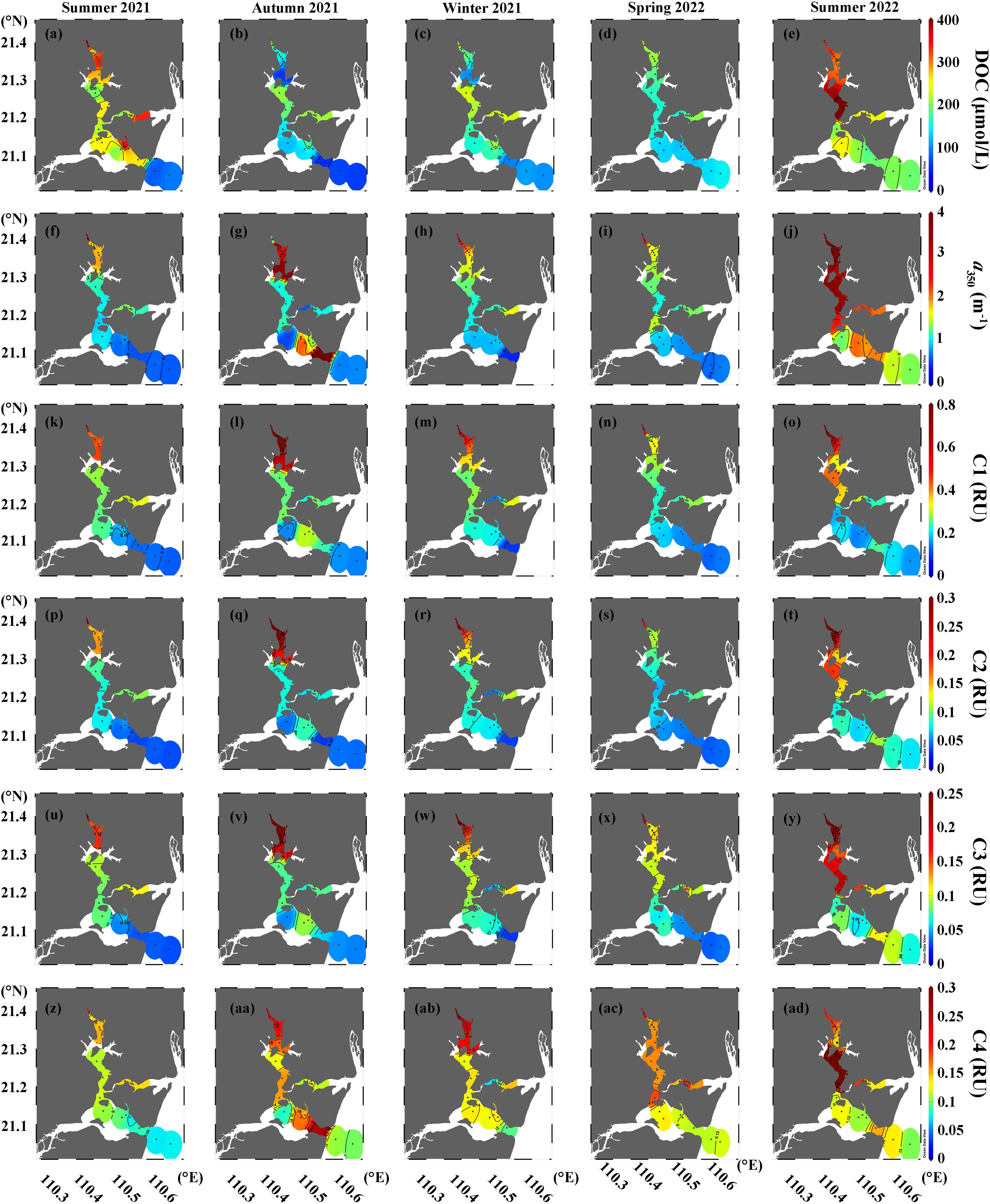
Figure 3. Seasonal distributions of (A–E) DOC, (F–J) a350, and (K–AD) intensities of four FDOM components in surface waters of the Suixi Estuary-Zhanjiang Bay continuum.
3.4 Distributions of qualitative optical parameters
The spectral slope S275-295 ranged from 12.9 to 22.7 μm-1, showing a contrasting spatial distribution pattern compared to a350 (Figures 4A–E; Supplementary Figures S4A–E). The highest mean S275-295 value occurred in summer 2021, while the lowest was noted in summer 2022. SUVA254 values showed a broad range from 0.24 to 6.74 m2/g C, peaking in autumn 2021, with the lowest values occurring in summers 2021 and 2022 (Table 1). Generally, SUVA254 decreased towards the sea, except in autumn when significantly lower values were recorded in the central continuum (Figures 4F–J; Supplementary Figures S4F–J). HIX varied from 0.88 to 6.22, decreasing from the upper estuary to lower bay, with the highest mean value in summer 2022 (Figures 4K-O; Supplementary Figures S4K–O). Conversely, BIX values ranged from 0.79 to 1.83, gradually increasing towards the sea and recording the lowest mean value in summer 2022 (Table 1; Figures 4P-T; Supplementary Figures S4P–T). FI values, varying from 2.41 to 2.61, showing the minimal spatial and temporal variation (Figures 4U–Y; Supplementary Figures S4U–Y). No significant differences were found between surface and bottom water for all DOM qualitative parameters (p > 0.05).
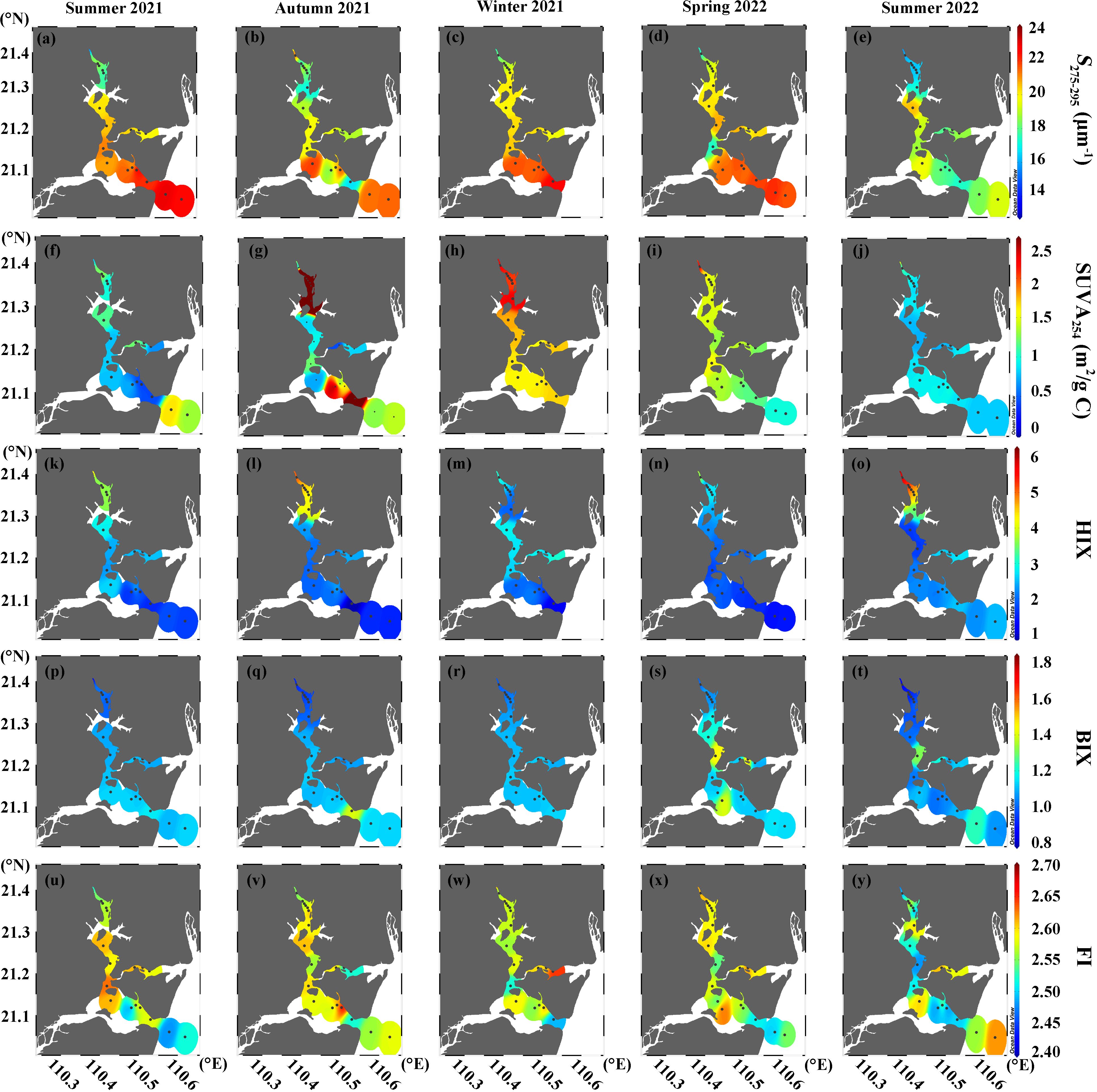
Figure 4. Seasonal distributions of (A–E) S275-295, (F–J) SUVA254, (K–O) HIX, (P–T) BIX, and (U–Y) FI in surface waters of the Suixi Estuary-Zhanjiang Bay continuum.
4 Discussion
4.1 Seasonal variability of freshwater and seawater DOM endmembers
Seasonal variations in riverine DOM endmember are well-documented across different climate regions (Guo et al., 2011; Li et al., 2019b; Yang et al., 2019). These variations affect the dynamics of riverine DOM in estuarine and coastal environments (Fichot and Benner, 2014) and introduce uncertainty (~30%) in quantifying global riverine DOM fluxes (Liu et al., 2024). In the Suixi estuary-Zhanjiang Bay continuum, freshwater DOC endmember values ranged from 355 to 708 μmol/L, peaking in autumn 2021 and reaching their lowest in winter 2021 (Table 2). A strong positive correlation was found between DOC and humic-like C1 in freshwater endmembers (r = 0.88, p = 0.05, n = 5). Freshwater endmembers with high levels of DOC and C1 typically exhibited low S275-295 and BIX, alongside high HIX (Table 2), suggesting a predominance of terrestrial organic materials with high humification degrees, low freshness, and high molecular weights in the Suixi River DOM pool (Hansen et al., 2016; Huguet et al., 2009; Helms et al., 2008). Protein-like components constituted only 10-18% of the total fluorescence intensity in freshwater DOM pool (Table 2), underscoring the dominance of natural terrestrial inputs. This is consistent with the high-productivity tropical forests and wetlands surrounding the Suixi River watershed (Zhong et al., 2022). Soil leaching and erosion, driven by high annual precipitation (~1800 mm, Chen et al., 2021), are likely the main contributors to the river’s high terrigenous DOM content. This is supported by a strong correlation between the freshwater DOC and C1 endmember values and 15-d cumulative precipitation prior to each cruise (r > 0.96, p< 0.05, n = 4), except for spring 2022. A short residence time of DOM in soil environments, driven by high precipitation, may limit its mineralization, thereby facilitating rapid transport to the river and maintaining high DOC levels in the riverine endmember.
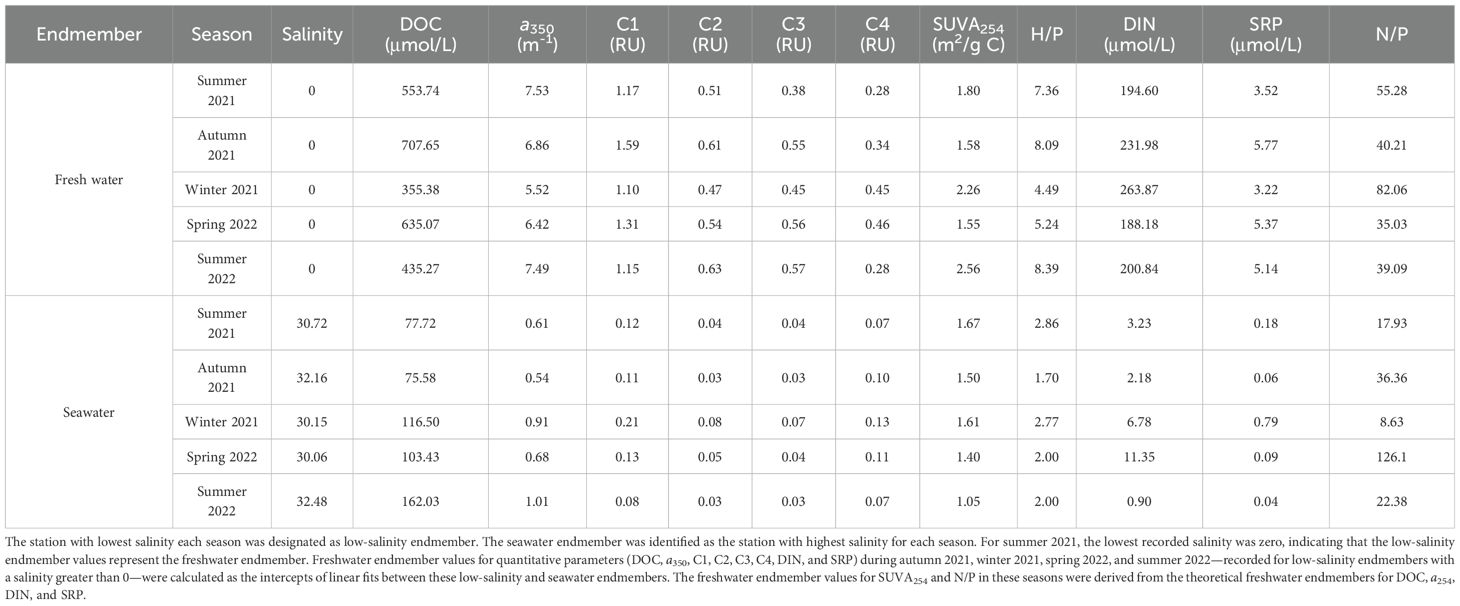
Table 2. Seasonal variations in DOM and nutrient endmember values for freshwater and seawater in the Suixi Estuary-Zhanjiang Bay continuum.
In seawater endmembers, DOM levels were consistently lower (74-91%) than in freshwater endmembers. Differences in CDOM and FDOMH (89-91%) being more pronounced compared to DOC (78 ± 12%) and FDOMP (74 ± 2.5%). This suggests decreased aromaticity and increased bioavailability, as indicated by lower SUVA254 and higher H/P values (Table 2). The seawater endmembers were mainly influenced by the West-Guangdong Coastal Current (WGCC) and tidal activities (Ding et al., 2017; Huang et al., 2021; Lao et al., 2022), contributing to modest variations in seawater salinity endmember values in this study (30.06-32.48, Table 2). In comparison, the variation in DOM within seawater endmembers was pronounced, contrasting with other systems, such as Pearl River and Jiulong River estuaries, which showed minimal variations in their seawater endmembers (Guo et al., 2007, 2011; Li et al., 2019b). In summer 2021, an intensified WGCC transported terrigenous materials from rivers in western Guangdong (e.g., Peral River, Moyang River, and Jian River), which contributed to the relatively low salinity but high H/P and SUVA254 values in seawater endmember (Table 2). Conversely, summer 2022 saw the highest DOC and lowest SUVA254 values in seawater endmembers, indicative of significant contributions from primary production, as evidenced by depleted levels of DIN and SRP (Table 1). The lack of correlation between salinity and DOM within seawater endmembers (p > 0.05) further highlights the importance of in situ biogeochemical transformations in shaping DOM within seawater endmembers. Seasonal variability in both freshwater and seawater DOM endmembers, driven by natural and anthropogenic factors, delineates the complex dynamics of DOM in the Suixi Estuary-Zhanjiang Bay continuum. With reduced runoff and enhanced dredging activity, salinity in the Suixi Estuary-Zhanjiang Bay continuum has risen by 18% over the past two decades (Lao et al., 2022), undoubtedly altering the DOM budget and transformation in the continuum and emphasizing the need for continued research.
4.2 Mixing behaviors of DOM in the Suixi Estuary-Zhanjiang Bay continuum
The transport of dissolved organic matter (DOM) from rivers to the ocean, especially through estuaries and coastal bays, involves complex physicochemical and biogeochemical processes that significantly alter the composition, reactivity, flux, and fate of riverine DOM as it reaches the ocean shelf (Raymond and Spencer, 2015; Spencer and Raymond, 2024).These transformations can be assessed through the relationships between DOM and salinity gradients (Huguet et al., 2009; Fellman et al., 2011; Guo et al., 2011; Vidal et al., 2023). Our findings indicate significant linear decreases in DOC, a350 and FDOM intensities with increasing salinity (p< 0.05), suggesting that conservative mixing of freshwater and seawater is the primary factor shaping the distribution of DOM in the Suixi Estuary-Zhanjiang Bay continuum (Figure 5). However, the R2 values for these linear fits varied substantially across different DOM parameters and seasons, highlighting the complexity of the interactions. The linear fittings between FDOMH and salinity demonstrated the highest R2 values (0.85-0.97 for C1, 0.86-0.98 for C2, and 0.83-0.96 for C3), followed by those between a350 and salinity (0.46-0.99), between FDOMP and salinity (0.22-0.89), and between DOC and salinity (0.25-0.92). Salinity explained over 83% of the variation in FDOMH, indicating that these components were dominated by the conservative mixing of freshwater and seawater endmembers. Non-conservative processes such as flocculation (Guo et al., 2007; Asmala et al., 2014), photobleaching (Moran et al., 2000; Catalá et al., 2013), and microbial activities (Klinkhammer et al., 2000; Yang et al., 2019) appeared to have a relatively small impact on their distributions across all seasons. Conservative mixing accounted for most of the variability in CDOM distribution during winter 2021 and in spring and summer 2022 (R2 ≥ 0.86) but explained only half of the variability in CDOM in autumn 2021 (R2 = 0.46). DOC and protein-like C4, proxies for labile DOM produced by primary production and degraded by microbes but relatively resistant to photobleaching (Yang et al., 2019), exhibited the expected weakest correlations with salinity in the Suixi Estuary-Zhanjiang Bay continuum (Figure 5). This was particularly noticeable in autumn 2021 and summer 2022 (0.22 ≤ R2 ≤ 0.63), periods characterized by higher Chl-a concentrations (Figure 3; Supplementary Figure S3). These diverse mixing behaviors among different DOM components and across seasons demonstrate the substantial modification riverine DOM undergoes when passing through estuarine environments (Spencer and Raymond, 2024), influencing the flux and fate of allochthonous DOM entering the northern SCS.
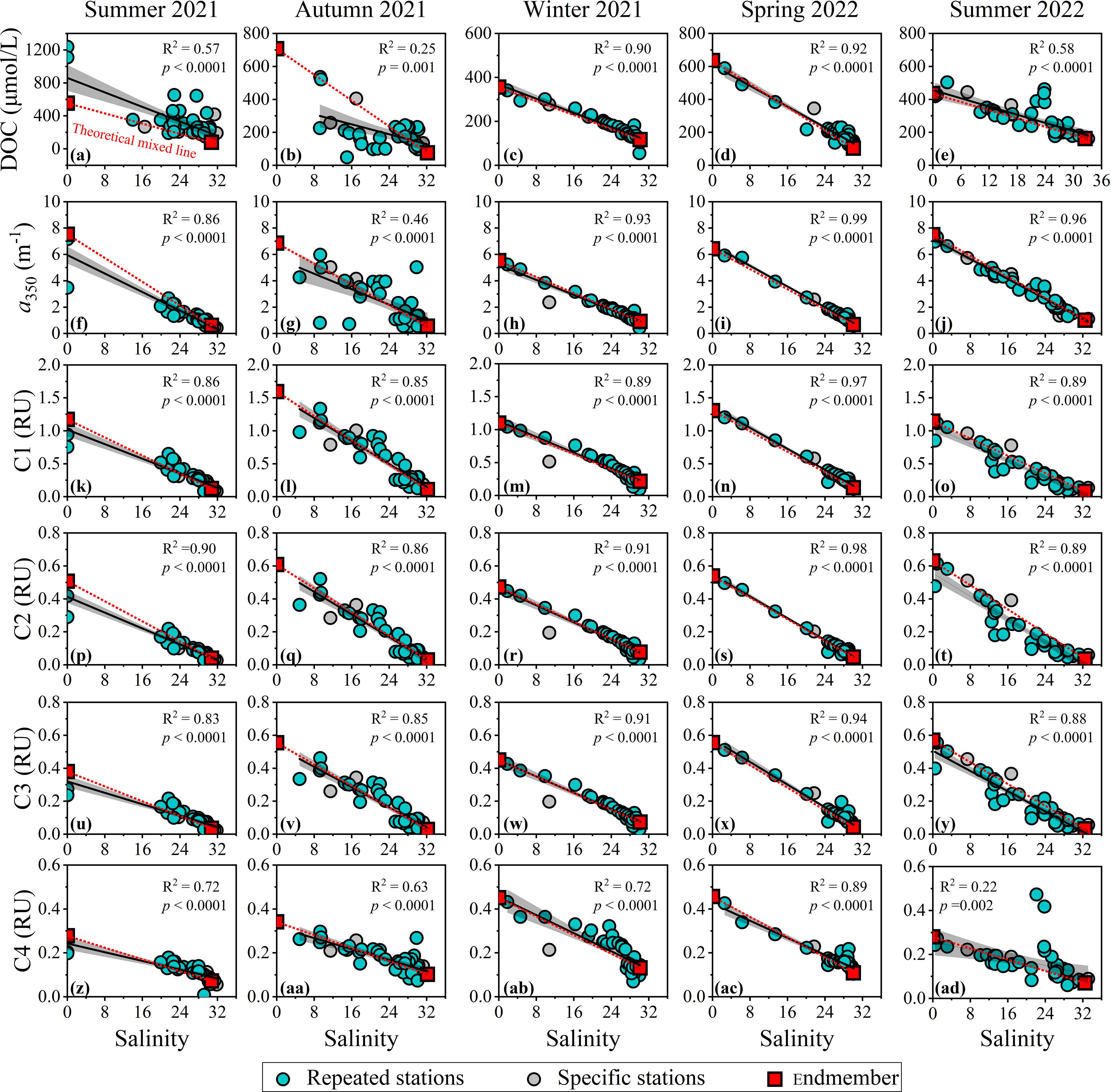
Figure 5. Seasonal variations in the relationships between (A–E) DOC, (F–J) a350, and (K–AD) intensities of four FDOM components with salinity across the Suixi Estuary-Zhanjiang Bay continuum. Black solid lines and gray shadows illustrate the linear fits and their 95% confidence bands, respectively. Red dotted lines represent the theoretical mixing lines between freshwater and seawater endmembers.
4.3 Seasonal variations of non-conservative DOM in the Suixi Estuary-Zhanjiang Bay continuum
Beyond assessing DOM behavior through correlations with salinity, a multiple endmember mixing model provides insights into both conservative and nonconservative processes impacting the DOM pool in estuarine and coastal environments (Dittmar et al., 2001; Ya et al., 2015; Jiang et al., 2017). In the Suixi Estuary-Zhanjiang Bay continuum, freshwater discharge from the Suixi River and seawater intrusion from the adjacent northern SCS are the primary allochthonous DOM sources (Lao et al., 2022). Other potential sources, such as submarine groundwater discharge, atmospheric deposition, and sediment release are considered limited based on dual water isotopes (Lao et al., 2023a) and DOM-salinity relationships (Figure 5). Thus, a two-endmember mixing model was utilized in this study:
Here, fFW and fSW represent the fractions of freshwater and seawater endmembers in a given water parcel, respectively, with SalFW and SalSW denoting their salinity values. Salobs and Mobs refer to the observed salinity and DOM parameters. MFW and MSW represent the DOM values of the respective endmember (Table 2). ΔM denotes nonconservative changes in DOM, with positive ΔM values indicating additions to the DOM pool. The relative change in M due to nonconservative processes (%) was calculated as the ratio of ΔM to Mobs. To discuss the effect of nutrients on the nonconservative processes of DOM, ΔDIN and ΔSRP were also calculated using this method.
ΔDOC ranged from -30 to 685 μmol/L in summer 2021 (163 ± 168 μmol/L), -364 to 114 μmol/L in autumn 2021 (-74 ± 160 μmol/L), -116 to 56 μmol/L in winter 2021 (12 ± 17 μmol/L), -61 to 94 μmol/L in spring 2022 (22 ± 30 μmol/L), and -39 to 227 μmol/L (25 ± 65 μmol/L), respectively. Significant additions of DOC (t-test, p< 0.05) were common but varied greatly across seasons: summer 2021 (102 ± 95%), winter 2021 (7.5 ± 11.7%), spring 2022 (18.0 ± 19.0%), and summer 2022 (10.7 ± 26.9%), despite the overall conservative behaviors of DOC during these seasons (p< 0.0001, Figures 5A, C–E). Although the mean ΔDOC was not significantly different from zero in autumn 2021 (p > 0.05, Figure 6A), negative ΔDOC values (-177 ± 100 μmol/L) were found in the low-salinity waters (salinity< 25, p< 0.0001) and positive values in high-salinity waters (salinity > 25, p = 0.002), indicating remarkable DOC removal in the upper continuum (-50 ± 24%) and addition in the lower bay (25 ± 33%). Phytoplankton production was identified as the primary process responsible for the DOC addition. Positive relationships between ΔDOC and Chl-a in summer 2021 (r = 0.44, p = 0.02) and 2022 (r = 0.87, p< 0.001) support DOM accumulation during actively growing phytoplankton blooms (Søndergaard et al., 2000; Hoikkala et al., 2012; Liu et al., 2024). Maximum DOC addition occurred in area with medium-high salinity (20-30, Figures 5A, E), where nutrient inputs from urban sewage discharge (Zhang et al., 2021), extended water residence times (Cai et al., 2024), and warm temperatures favored phytoplankton blooms (maximum Chl-a > 10 μg/L in summer 2021 and 50 μg/L in summer 2022).

Figure 6. Seasonal comparison of proportions of nonconservative (A) DIN, (B) SRP, (C) DOC, (D) a350, and (E–H) four FDOM components in the Suixi Estuary-Zhanjiang Bay continuum. An asterisk (*) indicates that the mean value is significantly different from zero at the 0.05 significance level.
Higher Chl-a levels in summer 2022 coincided with greater nutrient removals but did not result in as much DOC addition as in summer 2021 (Table 1, Figure 6), suggesting regulation by other biogeochemical processes. The bio-refractory FDOMH, less directly influenced by primary production than DOC, could offer valuable insights. Substantial removals in FDOMH (-19 ± 20% for C1, -30 ± 19% for C2, -22 ± 22% for C3) were found in summer 2022 (Figures 6D–H), especially in the low salinity region (~ 13), suggesting potential flocculation of DOM in this season (McCallister et al., 2006; Asmala et al., 2014). Although the bulk DOC flocculates more slowly than humic fraction of DOM (Eckert and Sholkovitz, 1976), we speculate that this flocculation may partially offsets the enhanced DOC in summer 2022. Additionally, labile DOC from phytoplankton photosynthesis could have a priming effect on riverine terrigenous refractory DOM, facilitating its partial remobilization for microbial utilization (Bianchi, 2011; Jiao et al., 2011; Spencer and Raymond, 2024). This stimulated microbial degradation likely also contributes to the removal of FDOMH in the Suixi Estuary-Zhanjiang Bay continuum. In contrast, substantial addition of FDOMH (10 ± 25% for C1 and 11 ± 26% for C3) were observed in summer 2021, linked to the in situ production during microbial-mediated degradation of labile carbon, as evidenced by positive correlations between ΔFDOMH and ΔDIN (r: 0.53-0.63, p< 0.005).
In spring, negative correlations were observed between ΔDOC and Chl-a (r = -0.45, p = 0.009), and between ΔC4 and Chl-a (r = -0.49, p = 0.004), suggesting DOC and FDOMP accumulated during declining algal blooms (Lønborg et al., 2009). Previous studies demonstrated that increased primary production facilitated rapid heterotrophic microbial degradation of labile organic matter, with particulate organic matter as the major substrate (Lao et al., 2023b). During the net-anabolic phase of heterotrophic microbial communities, some traditionally defined labile DOM compounds could be released through extracellular release, protist grazing, and viral lysis, decreasing Chl-a and enhancing DOC and FDOMP levels (Kawasaki and Benner, 2006; Lønborg et al., 2009). Concurrently, the net-catabolic phase of microbial activities could release more bio-refractory compounds such as CDOM and FDOMH as the by-products (Xiao et al., 2022), contributing to their significant additions in spring 2022 (15 ± 13% for a350, 18 ± 17% for C1, 8 ± 17% for C2, and 26 ± 34% for C3, Figure 6). This is supported by significant additions of DIN (14.3 ± 16.6 μmol/L, 39 ± 24%, Figure 6A) and SRP (0.59 ± 0.44 μmol/L, 123 ± 134%, Figure 6B), coupled with linear relationships between Δ values for FDOMH and nutrient and DO levels (e.g., ΔC1 vs. ΔDIN: r = 0.44, p = 0.015; ΔC1 vs. ΔSRP: r = 0.86, p< 0.0001; ΔC1 vs. DO: r = -0.69, p< 0.0001). These findings illustrate the prevalence of microbial-mediated transformation from labile biogenic particles to refractory DOM in spring 2022, accompanied by oxygen consumption and nutrient regeneration (Xiao et al., 2022). Precisely due to considerable contributions from both primary productivity and microbial transformation, DOM in spring 2022 in the continuum experienced extensive reworking, resulting in the highest fraction of bio-labile materials, lowest humification degree and molecular weight, and greatest freshness of any season (Table 1).
Unlike the visible removal of DOC occurred in the upper continuum during autumn 2021, CDOM and FDOM remained largely unaffected (Figure 5). This pattern suggests that riverine DOM was more efficiently consumed by heterotrophic microorganism, similar to summer 2022 (Bianchi, 2011; Zhuang et al., 2021). Microbial degradation was stimulated by locally produced labile DOC under the conditions of ample nutrients and favorable temperature (Tables 1, 2). Alongside gradual depletion of degradable riverine DOM toward the sea, positive ΔDOC emerges in the lower continuum (Figure 6), most likely resulting from phytoplankton bloom (Figure 2; Carlson and Hansell, 2015). The single-peak pattern of ΔDOC-Chl-a relationship in the lower continuum during autumn 2021 suggests an uncoupling of dissolved and particulate organic materials subjected to microbial decomposition, indicating preferential utilization of locally produced POM (Lao et al., 2023b). In winter 2021, characterized by the lowest river discharge and temperatures (Table 1; Figure 2), the estuarine DOM was the least dynamic, with no significant change in CDOM and FDOM levels (p > 0.05, Figure 6). This stability suggests minimal reworking of riverine DOM, aligning with observations from other seasonal estuaries (Li et al., 2019b; Gao et al., 2020). These findings highlight the substantial seasonal and interannual variations in estuarine DOM dynamics, driven by shifts in endmembers and influenced by both abiotic and biotic processes during water masses mixing (Bianchi, 2011; Vidal et al., 2023). The results underscore the necessity for high-frequency investigations to accurately track the transport and transformation of DOM from small tropical rivers to the coastal seas (Liu et al., 2024).
4.4 Export of riverine DOM by Suixi River and global perspective
The fluxes of DOM (i.e., DOC, CDOM, and FDOM) from Suixi River to the Zhangjiang Bay were estimated as follows (Guo et al., 2014; Li et al., 2019b):
Where F represents the flux of DOM, Q denotes the Suixi River discharge, and C* represents the freshwater endmember values of DOM (Table 2). Annual riverine DOM fluxes were calculated by summing the DOM fluxes from four seasons, using the average of the two summer fluxes as one component. Specifically, the annual DOM flux of the Suixi River was calculated as follows: 3.4 × 1010 g C/yr for DOC, 3.5 × 1010 m2/yr for a350, 6.3 × 109 RU m3/yr for C1, 2.8 × 109 RU m3/yr for C2, 2.5 × 109 RU m3/yr for C3, and 1.7 × 109 RU m3/yr for C4. In summer, the Suixi River accounted for 56% of the annual water discharge, but contributed only 47-49% of the annual DOC and FDOMP fluxes, while providing 59 ± 0.2% of the annual CDOM flux and 51-56% of the annual FDOMH flux. This is attributed to stronger soil leaching and washing processes during summer, which mobilize more refractory terrigenous DOM materials into the Suixi Estuary-Zhanjiang Bay continuum, highlighting the seasonal variation in the composition and flux of riverine DOM transported by small tropical rivers.
DOC concentrations in the Suixi River are moderately high compared to other Chinese and global rivers (Figure 7A, Raymond and Spencer, 2015; Wang et al., 2022; Spencer and Raymond, 2024). These concentrations are only exceeded by the pan-Arctic Rivers, which range from 364-805 μmol/L (Stedmon et al., 2011), and rivers significantly affected by anthropogenic inputs, such as the Hai River at ~868 μmol/L (Xia and Zhang, 2011). However, the DOC levels are comparable to those found in major tropical rivers like the Amazon and Orinoco, as well as North American rivers (Spencer et al., 2013; Raymond and Spencer, 2015). In contrast, the Suixi River exhibits significantly higher concentrations than other Chinese subtropical and temperate rivers including the Yangtze River (116-174 μmol/L, Guo et al., 2014), Pearl River (82-124 μmol/L, Li et al., 2019b), Min River (88-110 μmol/L, Yang et al., 2019), Jiulong River (132-186 μmol/L, Guo et al., 2011), and mountainous Taiwan rivers (32-139 μmol/L, Yang et al., 2013). However, the CDOM levels in Suixi River (a350-based, 6.76 ± 0.83 m-1) align closely with the global average (6.18 ± 5.16 m-1, Figure 7A), resulting in the noticeable deviations from the lineal DOC-a350 relationship observed across global 52 rivers (Figure 7A).
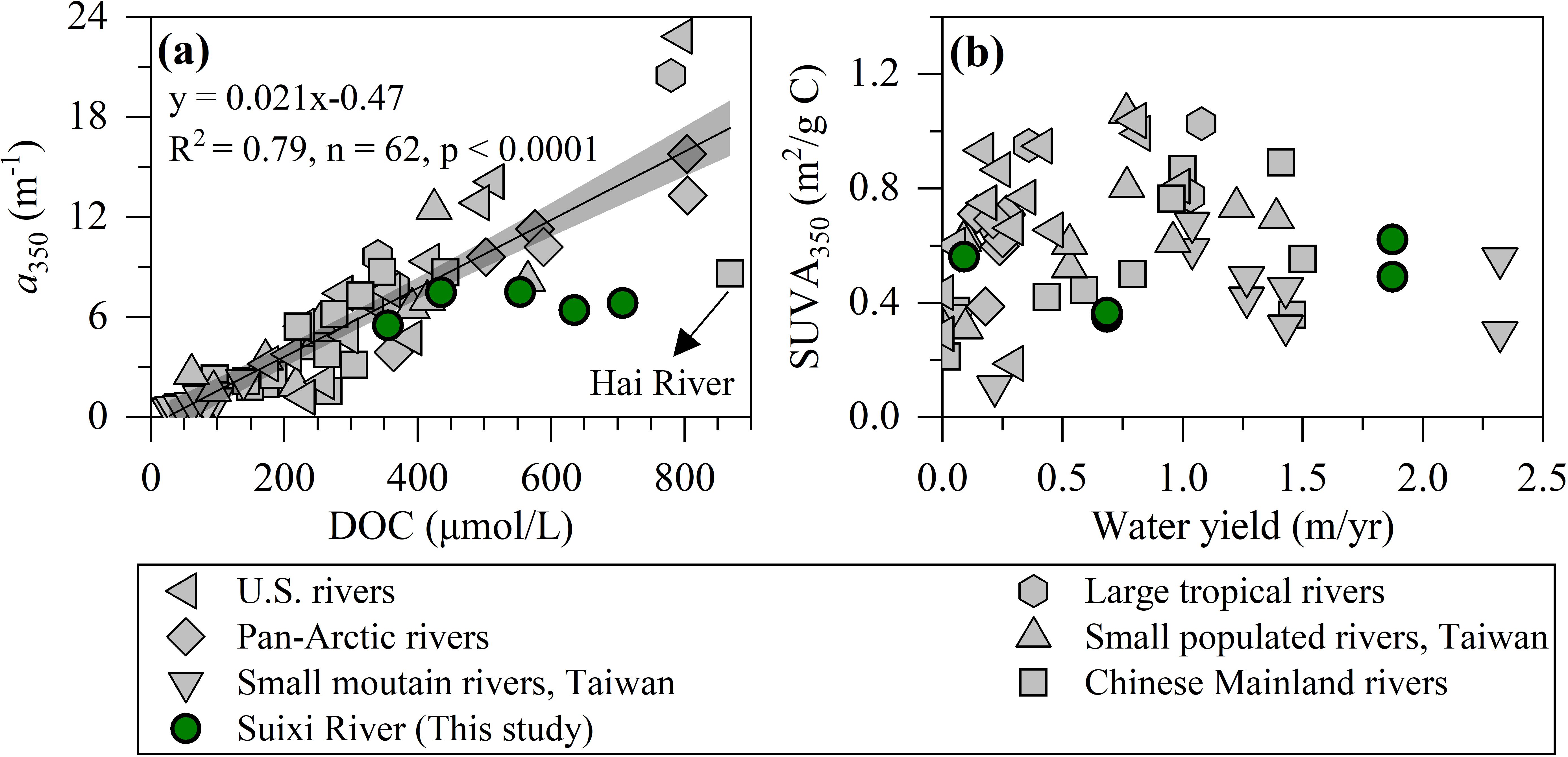
Figure 7. Relationships between (A) a350 and DOC, and between (B) SUVA350 and the ratio of water discharge to watershed area for global river endmembers. Black line and gray shadow in panel a represent the linear fit and confidence band (95%) for a total of 52 river endmembers in the globe, respectively. Data are listed in Supplementary Table S2.
In terms of DOM quality, the SUVA350 values for the Suixi River (0.48 ± 0.12 m2/g C) fall within the low to moderate range globally (0.64 ± 0.23 m2/g C, Figure 7B). These values were notably lower than those found in large tropical rivers (0.92 ± 0.13 m2/g C), North American rivers (0.71 ± 0.26 m2/g C), and pan-Arctic rivers (0.62 ± 0.12 m2/g C), yet similar to other Chinese rivers (0.47 ± 0.19 m2/g C, Supplementary Table S2). From a global perspective, peak SUVA350 values for river endmembers were observed at moderate water yields (m/yr, Figure 7B). A reasonable explanation is that larger water yield facilitates stronger interactions between river and soil, which in turn promote the release of soil organic carbon enriched in reworked CDOM. Once the soil reworked organic carbon is sufficiently mobilized, the increased water yield can enhance the contribution of labile carbon from net primary production to riverine DOM pool (Tromboni et al., 2022), as reflected by decreased SUVA350 values for river water endmembers (Figure 7B). The ongoing changes in climate (e.g., temperature and precipitation) and intensifying human activities (e.g., land utilization, dam construction) continue to affect the water yields of global tropical rivers (Wu et al., 2023; Vystavna et al., 2024), undoubtedly exert dynamic of riverine DOM composition entering the sea (Liu et al., 2022). Despite similar SUVA350 values, both the DOC yield (21.9 g C/m2/yr) and CDOM yield (a350-based, 21.9 yr-1) of the Suixi River far exceed those of other Chinese rivers (2.68 ± 4.46 g C/m2/yr and 2.78 ± 4.08 yr-1, respectively). This uncoupling is largely attributed to high watershed productivity and strong soil leaching in the Suixi River, compared to other subtropical and temperate Chinese rivers, which are affected by a low abundance of aged and leached soil organic matter, dam construction, and rapid wetlands reduction due to anthropogenic exploitation (Guo et al., 2014). This exceptionally high DOC yield from the Suixi River, more than twice the highest recorded yield from the large tropical Fly River (~8.6 g C/m2/yr, Raymond and Spencer, 2015), highlights the critical role of small, tropical rivers in estuarine and coastal carbon cycling and the need to consider these smaller systems in broader ecological and biogeochemical studies.
5 Conclusion
This study reveals the complex dynamics at the tropical river-sea interface of the Suixi Estuary-Zhanjiang Bay continuum, highlighting the significant role of small tropical watersheds in global carbon cycling. Our analysis from summer 2021 to summer 2022 encompassed DOC, CDOM, and FDOM, showing pronounced seasonal variations. DOC in freshwater endmember peaked in autumn 2021, driven largely by terrestrial inputs as evidenced by strong correlations with FDOMH and enhanced by soil erosion and leaching that correlate with precipitation. Conversely, DOC in seawater endmember peaked in summer 2022, enriched in FDOMP and influenced by the West-Guangdong Coastal Current alongside in situ primary production.
Our findings indicate that while conservative mixing of freshwater and seawater predominantly influences DOM distribution within the continuum, significant non-conservative modifications occur through flocculation, phytoplankton production, and microbial transformation. These alterations vary across seasons, affecting the DOM’s qualitative and quantitative characteristics. Despite the Suixi River accounting for a substantial portion of the annual water discharge (56%), its contribution to annual DOM fluxes (47-59%) does not proportionately reflect its output, primarily due to intense seasonal soil leaching that mobilizes more refractory terrigenous DOM. This phenomenon results in remarkably high DOC and CDOM yields, substantially exceeding those recorded for other Chinese rivers and even global peak yields like the tropical Fly River.
Our analysis also revealed a distinct single-peak pattern between the SUVA350 values for river endmembers and water yield, based on a global dataset. This pattern suggests that maximum mobilization of soil-derived refractory organic carbon occurs at moderate water yields. Beyond this peak, refractory soil carbon contributions diminish while labile carbon input from net primary production increases, highlighting the complex interplay between hydrological dynamics and carbon cycling processes. The study demonstrates that the Suixi River, like other small tropical rivers, plays a crucial role in the estuarine and coastal carbon budget. Moving forward, integrating small tropical watershed data into global carbon models is essential to enhance the accuracy of carbon budget estimations.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Author contributions
PL: Formal analysis, Investigation, Visualization, Writing – original draft. KJ: Investigation, Writing – original draft. QT: Investigation, Writing – original draft. FC: Writing – review & editing. RJ: Writing – review & editing. CW: Conceptualization, Funding acquisition, Methodology, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was funded by the Guangdong Basic and Applied Basic Research Foundation (2022A1515110946), the open grant from Guangxi Key Laboratory of Beibu Gulf Marine Resources, Environment and Sustainable Development (MRESD-2022-B01), and the Tropical Ocean Environment in Western Coastal Waters Observation and Research Station of Guangdong Province (2024B1212040008).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2024.1511427/full#supplementary-material
References
Asmala E., Bowers D. G., Autio R., Kaartokallio H., Thomas D. N. (2014). Qualitative changes of riverine dissolved organic matter at low salinities due to flocculation. J. Geophysical Research-Biogeosciences 119, 1919–1933. doi: 10.1002/2014JG002722
Bauer J. E., Bianchi T. S. (2011). “Dissolved organic carbon cycling and transformation,” in Treatise on Estuarine and Coastal Science, 5 (Amsterdam, Netherlands: Academic Press, Waltham), 7–67.
Bauer J. E., Cai W. J., Raymond P. A., Bianchi T. S., Hopkinson C. S., Regnier P. A. G. (2013). The changing carbon cycle of the coastal ocean. Nature 504, 61–70. doi: 10.1038/nature12857
Bianchi T. S. (2011). The role of terrestrially derived organic carbon in the coastal ocean: A changing paradigm and the priming effect. Proc. Natl. Acad. Sci. 108, 19473–19481. doi: 10.1073/pnas.1017982108
Bittar T. B., Vieira A. A. H., Stubbins A., Mopper K. (2015). Competition between photochemical and biological degradation of dissolved organic matter from the cyanobacteria Microcystis aeruginosa. Limnology Oceanography. 60, 1172–1194. doi: 10.1002/lno.10090
Cai S., Lao Q., Cai M., Silva D. P. T. T., Lu X., Zhou X., et al. (2024). Water residence time controls seasonal nitrous oxide budget in a semi-enclosed bay: Insights from an improvement estimation method. Mar. Pollut. Bull. 206, 116701. doi: 10.1016/j.marpolbul.2024.116701
Carlson C. A., Hansell D. A (2015). “Chapter 3 - DOM sources, sinks, reactivity, and budgets,” in Biogeochemistry of marine dissolved organic matter, 2nd ed. Eds. Hansell D. A., Carlson C. A. (Academic Press, Boston), 65–126.
Carlson C. A., Liu S., Stephens B. M., Chance J. (2024). “Chapter 5 - DOM production, removal, and transformation processes in marine systems,” in Biogeochemistry of Marine Dissolved Organic Matter, 3rd ed. (Amsterdam, Netherlands: Academic Press), 137–246.
Carpenter J. H. (1965). The Chesapeake bay institute technique for the winkler dissolved oxygen method. Oceanography 10, 141–143. doi: 10.4319/lo.1965.10.1.0141
Catalá T. S., Mladenov N., Echevarría F., Reche I. (2013). Positive trends between salinity and chromophoric and fluorescent dissolved organic matter in a seasonally inverse estuary. Estuar. Coast. Shelf Sci. 133, 206–216. doi: 10.1016/j.ecss.2013.08.030
Chen R. F., Gardner G. B. (2004). High-resolution measurements of chromophoric dissolved organic matter in the Mississippi and Atchafalaya River plume regions. Mar. Chem. 89, 103–125. doi: 10.1016/j.marchem.2004.02.026
Chen F., Huang C., Lao Q., Zhang S., Chen C., Zhou X., et al. (2021). Typhoon control of precipitation dual isotopes in Southern China and its palaeoenvironmental implications. J. Geophysical Research: Atmospheres 126, e2020JD034336. doi: 10.1029/2020JD034336
Coble P. G. (2007). Marine optical biogeochemistry: The chemistry of ocean color. Chem. Rev. 107, 402–418. doi: 10.1021/cr050350+
Cohen E., Levy G. J., Borisover M. (2014). Fluorescent components of organic matter in wastewater: Efficacy and selectivity of the water treatment. Water Res. 55, 323–334. doi: 10.1016/j.watres.2014.02.040
Cory R. M., McKnight D. M. (2005). Fluorescence spectroscopy reveals ubiquitous presence of oxidized and reduced quinones in dissolved organic matter. Environ. Sci. Technol. 39, 8142–8149. doi: 10.1021/es0506962
Dai M., Su J., Zhao Y., Hofmann E. E., Cao Z., Cai W., et al. (2022). Carbon fluxes in the coastal ocean: synthesis, boundary processes, and future trends. Annu. Rev. Earth Planetary Sci. 50, 593–626. doi: 10.1146/annurev-earth-032320-090746
Dainard P. G., Guéguen C., Yamamoto-Kawai M., Williams W. J., Hutchings J. K. (2019). Interannual variability in the absorption and fluorescence characteristics of dissolved organic matter in the Canada basin polar mixed waters. J. Geophys Res: Oceans 124, 5258–5269. doi: 10.1029/2018JC014896
Ding Y., Bao X., Yao Z., Zhang C., Wan K., Bao M., et al. (2017). A modeling study of the characteristics and mechanism of the westward coastal current during summer in the northwestern South China Sea. Ocean Sci. J. 52, 11–30. doi: 10.1007/s12601-017-0011-x
Dittmar T., Lara R. J., Kattner G. (2001). River or mangrove? Tracing major organic matter sources in tropical Brazilian coastal waters. Mar. Chem. 73, 253–271. doi: 10.1016/S0304-4203(00)00110-9
Eckert J. M., Sholkovitz E. R. (1976). The flocculation of iron, aluminium and humates from river water by electrolytes. Geochim Cosmochimica Acta 40, 847–848. doi: 10.1016/0016-7037(76)90036-3
Fellman J. B., Petrone K. C., Grierson P. F. (2011). Source, biogeochemical cycling, and fluorescence characteristics of dissolved organic matter in an agro-urban estuary. Limnology Oceanography. 56, 243–256. doi: 10.4319/lo.2011.56.1.0243
Fichot C. G., Benner R. (2014). The fate of terrigenous dissolved organic carbon in a river-influenced ocean margin. Global Biogeochemical Cycles 28, 300–318. doi: 10.1002/2013GB004670
Gamrani M., Eert J., Williams W. J., Gueguen C. (2023). A river of terrestrial dissolved organic matter in the upper waters of the central Arctic Ocean. Deep-Sea Res. Part I-Oceanographic Res. Papers 196,196. doi: 10.1016/j.dsr.2023.104016
Gao L., Gao Y., Song S., Zhang F. (2020). Non-conservative behavior of dissolved organic carbon in the Changjiang (Yangtze River) Estuary and the adjacent East China Sea. Continental Shelf Res. 197, 197. doi: 10.1016/j.csr.2020.104084
Gao Z., Gueguen C. (2017). Size distribution of absorbing and fluorescing DOM in Beaufort Sea, Canada Basin. Deep-Sea Res. Part I-Oceanographic Res. Papers 121, 30–37. doi: 10.1016/j.dsr.2016.12.014
García-Martín E. E., Sanders R., Evans C. D., Kitidis V., Lapworth D. J., Rees A. P., et al. (2021). Contrasting estuarine processing of dissolved organic matter derived from natural and human-impacted landscapes. Global Biogeochem Cycles 35, 35. doi: 10.1029/2021GB007023
Guo W., Stedmon C. A., Han Y., Wu F., Yu X., Hu M. (2007). The conservative and non-conservative behavior of chromophoric dissolved organic matter in Chinese estuarine waters. Mar. Chem. 107, 357–366. doi: 10.1016/j.marchem.2007.03.006
Guo W., Yang L., Hong H., Stedmon C. A., Wang F., Xu J., et al. (2011). Assessing the dynamics of chromophoric dissolved organic matter in a subtropical estuary using parallel factor analysis. Mar. Chem. 124, 125–133. doi: 10.1016/j.marchem.2011.01.003
Guo W., Yang L., Zhai W., Chen W., Osburn C. L., Huang X., et al. (2014). Runoff-mediated seasonal oscillation in the dynamics of dissolved organic matter in different branches of a large bifurcated estuaryThe Changjiang Estuary. J. Geophysical Research-Biogeosciences 119, 776–793. doi: 10.1002/2013JG002540
Hansen A. M., Kraus T. E. C., Pellerin B. A., Fleck J. A., Downing B. D., Bergamaschi B. A. (2016). Optical properties of dissolved organic matter (DOM): Effects of biological and photolytic degradation. Limnology Oceanography 61, 1015–1032. doi: 10.1002/lno.10270
Helms J. R., Stubbins A., Ritchie J. D., Minor E. C., Kieber D. J., Mopper K. (2008). Absorption spectral slopes and slope ratios as indicators of molecular weight, source, and photobleaching of chromophoric dissolved organic matter. Limnology Oceanography 53, 955–969. doi: 10.4319/lo.2008.53.3.0955
Hoikkala L., Lahtinen T., Perttila M., Lignell R. (2012). Seasonal dynamics of dissolved organic matter on a coastal salinity gradient in the northern Baltic Sea. Continental Shelf Res. 45, 1–14. doi: 10.1016/j.csr.2012.04.008
Huang X., Fu X., Zhao Z., Yin H. (2024). The telltale fluorescence fingerprints of sewer flows for interpreting the low influent concentration in wastewater treatment plant. J. Environ. Manage. 349, 349. doi: 10.1016/j.jenvman.2023.119517
Huang R., Xie L., Li M., Wang L. (2021). Observational analysis on 3D distribution and seasonal variation of thermohaline characteristics in the Zhanjiang Bay. Haiyang Xuebao 43, 46–60. doi: 10.12284/hyxb2021156
Huguet A., Vacher L., Relexans S., Saubusse S., Froidefond J. M., Parlanti E. (2009). Properties of fluorescent dissolved organic matter in the Gironde Estuary. Organic Geochemistry 40, 706–719. doi: 10.1016/j.orggeochem.2009.03.002
Jiang T., Skyllberg U., Bjorn E., Green N. W., Tang J., Wang D., et al. (2017). Characteristics of dissolved organic matter (DOM) and relationship with dissolved mercury in Xiaoqing River-Laizhou Bay estuary, Bohai Sea, China. Environ. pollut. 223, 19–30. doi: 10.1016/j.envpol.2016.12.006
Jiao N., Luo T., Chen Q., Zhao Z., Xiao X., Liu J., et al. (2024). The microbial carbon pump and climate change. Nat. Rev. Microbiol. 22, 408–419. doi: 10.1038/s41579-024-01018-0
Jiao N., Tang K., Cai H., Mao Y. (2011). Increasing the microbial carbon sink in the sea by reducing chemical fertilization on the land. Nat. Rev. Microbiol. 9, 75–75. doi: 10.1038/nrmicro2386-c2
Kawasaki N., Benner R. (2006). Bacterial release of dissolved organic matter during cell growth and decline: Molecular origin and composition. Limnology Oceanography 51, 2170–2180. doi: 10.4319/lo.2006.51.5.2170
Klinkhammer G. P., McManus J., Colbert D., Rudnicki M. D. (2000). Behavior of terrestrial dissolved organic matter at the continent-ocean boundary from high-resolution distributions. Geochimica Cosmochimica Acta 64, 2765–2774. doi: 10.1016/S0016-7037(99)00370-1
Kothawala D. N., Murphy K. R., Stedmon C. A., Weyhenmeyer G. A., Tranvik L. J. (2013). Inner filter correction of dissolved organic matter fluorescence. Limnology Oceanography: Methods 11, 616–630. doi: 10.4319/lom.2013.11.616
Lao Q., Chen F., Jin G., Lu X., Chen C., Zhou X., et al. (2023b). Characteristics and mechanisms of typhoon-induced decomposition of organic matter and its implication for climate change. J. Geophysical Research-Biogeosciences 128, 128. doi: 10.1029/2023JG007518
Lao Q., Lu X., Chen F., Chen C., Jin G., Zhu Q. (2023a). A comparative study on source of water masses and nutrient supply in Zhanjiang Bay during the normal summer, rainstorm, and typhoon periods: Insights from dual water isotopes. Sci. Total Environ. 903, 166853. doi: 10.1016/j.scitotenv.2023.166853
Lao Q., Wu J., Chen F., Zhou X., Li Z., Chen C., et al. (2022). Increasing intrusion of high salinity water alters the mariculture activities in Zhanjiang Bay during the past two decades identified by dual water isotopes. J. Environ. Manage. 320, 115815. doi: 10.1016/j.jenvman.2022.115815
Lawaetz A. J., Stedmon C. A. (2009). Fluorescence intensity calibration using the raman scatter peak of water. Appl. Spectrosc. 63, 936–940. doi: 10.1366/000370209788964548
Lehmann J., Kleber M. (2015). The contentious nature of soil organic matter. Nature 528, 60–68. doi: 10.1038/nature16069
Letscher R. T., Hansell D. A., Kadko D. (2011). Rapid removal of terrigenous dissolved organic carbon over the Eurasian shelves of the Arctic Ocean. Mar. Chem. 123, 78–87. doi: 10.1016/j.marchem.2010.10.002
Li P., Chen L., Zhang W., Huang Q. (2015). Spatiotemporal distribution, sources, and photobleaching imprint of dissolved organic matter in the Yangtze Estuary and its adjacent sea using fluorescence and parallel factor analysis. PloS One 10, e0130852. doi: 10.1371/journal.pone.0130852
Li M., Peng C., Zhou X., Yang Y., Guo Y., Shi G., et al. (2019a). Modeling global riverine DOC flux dynamics from 1951 to 2015. J. Adv. Modeling Earth Syst. 11, 514–530. doi: 10.1029/2018MS001363
Li Y., Song G., Massicotte P., Yang F., Li R., Xie H. (2019b). Distribution, seasonality, and fluxes of dissolved organic matter in the Pearl River (Zhujiang) estuary, China. Biogeosciences 16, 2751–2770. doi: 10.5194/bg-16-2751-2019
Liang H., Zhang J., Zhang J., Zhang P., Deng X., Chen J., et al. (2024). Unveiling the eutrophication crisis: 20 years of nutrient development in Zhanjiang Bay, China. Front. Mar. Sci. 11. doi: 10.3389/fmars.2024.1373716
Liu S., Maavara T., Brinkerhoff C. B., Raymond P. A. (2022). Global controls on DOC reaction versus export in watersheds: A damkohler number analysis. Global Biogeochemical Cycles 36, e2021GB007278. doi: 10.1029/2021GB007278
Liu M., Raymond P. A., Lauerwald R., Zhang Q., Trapp-Müller G., Davis K. L., et al. (2024). Global riverine land-to-ocean carbon export constrained by observations and multi-model assessment. Nat. Geosci. 17, 896–904. doi: 10.1038/s41561-024-01524-z
Lønborg C., Álvarez-Salgado X. A., Davidson K., Miller A. E. J. (2009). Production of bioavailable and refractory dissolved organic matter by coastal heterotrophic microbial populations. Estuar. Coast. Shelf Sci. 82, 682–688. doi: 10.1016/j.ecss.2009.02.026
Markager S., Stedmon C. A., Søndergaard M. (2011). Seasonal dynamics and conservative mixing of dissolved organic matter in the temperate eutrophic estuary Horsens Fjord. Estuar. Coast. Shelf Sci. 92, 376–388. doi: 10.1016/j.ecss.2011.01.014
Massicotte P., Asmala E., Stedmon C., Markager S. (2017). Global distribution of dissolved organic matter along the aquatic continuum: Across rivers, lakes and oceans. Sci. Total Environ. 609, 180–191. doi: 10.1016/j.scitotenv.2017.07.076
McCallister S. L., Bauer J. E., Ducklow H. W., Canuel E. A. (2006). Sources of estuarine dissolved and particulate organic matter: A multi-tracer approach. Organic Geochemistry 37, 454–468. doi: 10.1016/j.orggeochem.2005.12.005
McDonough L. K., Andersen M. S., Behnke M. I., Rutlidge H., Oudone P., Meredith K., et al. (2022). A new conceptual framework for the transformation of groundwater dissolved organic matter. Nat. Commun. 13, 2153. doi: 10.1038/s41467-022-29711-9
Medeiros P. M., Seidel M., Ward N. D., Carpenter E. J., Gomes H. R., Niggemann J., et al. (2015). Fate of the Amazon River dissolved organic matter in the tropical Atlantic Ocean. Global Biogeochemical Cycles 29, 677–690. doi: 10.1002/2015GB005115
Moran M. A., Sheldon W. M., Zepp R. G. (2000). Carbon loss and optical property changes during long-term photochemical and biological degradation of estuarine dissolved organic matter. Limnology Oceanography 45, 1254–1264. doi: 10.4319/lo.2000.45.6.1254
Murphy K. R., Stedmon C. A., Wenig P., Bro R. (2014). OpenFluor- an online spectral library of auto-fluorescence by organic compounds in the environment. Analytical Methods 6, 658–661. doi: 10.1039/C3AY41935E
Osterholz H., Kirchman D. L., Niggemann J., Dittmar T. (2016). Environmental drivers of dissolved organic matter molecular composition in the Delaware Estuary. Front. Earth Sci. 4. doi: 10.3389/feart.2016.00095
Raymond P. A., Spencer R. G. M (2015). “Chapter 11 - Riverine DOM,” in Biogeochemistry of Marine Dissolved Organic Matter, 2nd ed. Eds. Hansell D. A., Carlson C. A. (Academic Press, Boston), 509–533.
Rochelle-Newall E. J., Fisher T. R. (2002). Production of chromophoric dissolved organic matter fluorescence in marine and estuarine environments: an investigation into the role of phytoplankton. Mar. Chem. 77, 7–21. doi: 10.1016/S0304-4203(01)00072-X
Romera-Castillo C., Sarmento H., Alvarez-Salgado X. A., Gasol J. M., Marrase C. (2010). Production of chromophoric dissolved organic matter by marine phytoplankton. Limnology Oceanography 55, 446–454. doi: 10.4319/lo.2010.55.1.0446
Safieddine S. A., Heald C. L. (2017). A global assessment of dissolved organic carbon in precipitation. Geophysical Res. Lett. 44, 11,672–611,681. doi: 10.1002/2017GL075270
Søndergaard M., Borch N. H., Riemann B. (2000). Dynamics of biodegradable DOC produced by freshwater plankton communities. Aquat. Microbial Ecol. 23, 73–83. doi: 10.3354/ame023073
Spencer R. G. M., Aiken G. R., Dornblaser M. M., Butler K. D., Holmes R. M., Fiske G., et al. (2013). Chromophoric dissolved organic matter export from U.S. rivers. Geophysical Res. Lett. 40, 1575–1579. doi: 10.1002/grl.50357
Spencer R. M., Raymond P. A. (2024). “Chapter 14 - Riverine DOM,” in Biogeochemistry of Marine Dissolved Organic Matter, 3rd ed. (Amsterdam, Netherlands: Academic Press), 657–691.
Stedmon C. A., Markager S. (2005). Tracing the production and degradation of autochthonous fractions of dissolved organic matter by fluorescence analysis. Limnology Oceanography 50, 1415–1426. doi: 10.4319/lo.2005.50.5.1415
Stedmon C. A., Markager S., Bro R. (2003). Tracing dissolved organic matter in aquatic environments using a new approach to fluorescence spectroscopy. Mar. Chem. 82, 239–254. doi: 10.1016/S0304-4203(03)00072-0
Stedmon C. A., Thomas D. N., Papadimitriou S., Granskog M. A., Dieckmann G. S. (2011). Using fluorescence to characterize dissolved organic matter in Antarctic sea ice brines. J. Geophysical Research-Biogeosciences 116, G03027. doi: 10.1029/2011JG001716
Tromboni F., Hotchkiss E. R., Schechner A. E., Dodds W. K., Poulson S. R., Chandra S. (2022). High rates of daytime river metabolism are an underestimated component of carbon cycling. Commun. Earth Environ. 3, 270. doi: 10.1038/s43247-022-00607-2
Tzortziou M., Zeri C., Dimitriou E., Ding Y., Jaffe R., Anagnostou E., et al. (2015). Colored dissolved organic matter dynamics and anthropogenic influences in a major transboundary river and its coastal wetland. Limnology Oceanography 60, 1222–1240. doi: 10.1002/lno.10092
Vidal L. O., Lambert T., Cotovicz L. C. Jr., Bernardes M. C., Sobrinho R., Thompson F., et al. (2023). Seasonal and diel modulation of DOM in a mangrove-dominated estuary. Sci. Total Environ. 857, 857. doi: 10.1016/j.scitotenv.2022.159045
Vystavna Y., Chavanne L., Harjung A., Soto D. X., Watson A., Miller J., et al. (2024). Predicting river flow dynamics using stable isotopes for better adaptation to climate and land-use changes. Nat. Water 2, 741–748. doi: 10.1038/s44221-024-00280-z
Wang C., Hu X., Liu Y., Jin G. (2022). Sources, spectral characteristics, and fluxes of dissolved organic matter in coastal groundwater and river water in western Guangdong, China. Front. Environ. Sci. 10. doi: 10.3389/fenvs.2022.995190
Weishaar J. L., Aiken G. R., Bergamaschi B. A., Fram M. S., Fujii R., Mopper K. (2003). Evaluation of specific ultraviolet absorbance as an indicator of the chemical composition and reactivity of dissolved organic carbon. Environ. Sci. Technol. 37, 4702–4708. doi: 10.1021/es030360x
Welschmeyer N. A. (1994). Fluorometric analysis of chlorophyll a in the presence of chlorophyll b and pheopigments. Limnology Oceanography 39, 1985–1992. doi: 10.4319/lo.1994.39.8.1985
Wen Z., Shang Y., Lyu L., Liu G., Hou J., He C., et al. (2021). Sources and composition of riverine dissolved organic matter to marginal seas from mainland China. J. Hydrology 603, 127152. doi: 10.1016/j.jhydrol.2021.127152
Wu Q., Ke L., Wang J., Pavelsky T. M., Allen G. H., Sheng Y., et al. (2023). Satellites reveal hotspots of global river extent change. Nat. Commun. 14, 1587. doi: 10.1038/s41467-023-37061-3
Xia B., Zhang L. (2011). Carbon distribution and fluxes of 16 rivers discharging into the Bohai Sea in summer. Acta Oceanologica Sin. 30, 43–54. doi: 10.1007/s13131-011-0118-3
Xiao X., Powers L. C., Liu J., Gonsior M., Zhang R., Zhang L., et al. (2022). Biodegradation of Terrigenous Organic Matter in a Stratified Large-Volume Water Column: Implications of the removal of terrigenous organic matter in the coastal ocean. Environ. Sci. Technol. 56, 5234–5246. doi: 10.1021/acs.est.1c08317
Ya C., Anderson W., Jaffe R. (2015). Assessing dissolved organic matter dynamics and source strengths in a subtropical estuary: Application of stable carbon isotopes and optical properties. Continental Shelf Res. 92, 98–107. doi: 10.1016/j.csr.2014.10.005
Yamashita Y., Fichot C. G., Shen Y., Jaffe R., Benner R. (2015). Linkages among fluorescent dissolved organic matter, dissolved amino acids and lignin-derived phenols in a river-influenced ocean margin. Front. Mar. Sci. 2, G00F10. doi: 10.3389/fmars.2015.00092
Yamashita Y., Jaffe R., Maie N., Tanoue E. (2008). Assessing the dynamics of dissolved organic matter (DOM) in coastal environments by excitation emission matrix fluorescence and parallel factor analysis (EEM-PARAFAC). Limnology Oceanography 53, 1900–1908. doi: 10.4319/lo.2008.53.5.1900
Yamashita Y., Maie N., Briceno H., Jaffe R. (2010). Optical characterization of dissolved organic matter in tropical rivers of the Guayana Shield, Venezuela. J. Geophysical Research-Biogeosciences 115. doi: 10.1029/2009JG000987
Yamashita Y., Tanoue E. (2003). Chemical characterization of protein-like fluorophores in DOM in relation to aromatic amino acids. Mar. Chem. 82, 255–271. doi: 10.1016/S0304-4203(03)00073-2
Yang L., Cheng Q., Zhuang W.-E., Wang H., Chen W. (2019). Seasonal changes in the chemical composition and reactivity of dissolved organic matter at the land-ocean interface of a subtropical river. Environ. Sci. pollut. Res. 26, 24595–24608. doi: 10.1007/s11356-019-05700-2
Yang L., Hong H., Chen C.-T. A., Guo W., Huang T.-H. (2013). Chromophoric dissolved organic matter in the estuaries of populated and mountainous Taiwan. Mar. Chem. 157, 12–23. doi: 10.1016/j.marchem.2013.07.002
Zhang J., Zhang Y., Zhang P., Li Y., Li J., Luo X., et al. (2021). Seasonal phosphorus variation in coastal water affected by the land-based sources input in the eutrophic Zhanjiang Bay, China. Estuarine Coast. Shelf Sci. 252, 107277. doi: 10.1016/j.ecss.2021.107277
Zhao C., Zhou Y., Pang Y., Zhang Y., Huang W., Wang Y., et al. (2021). The optical and molecular signatures of DOM under the eutrophication status in a shallow, semi-enclosed coastal bay in southeast China. Sci. China-Earth Sci. 64, 1090–1104. doi: 10.1007/s11430-020-9728-4
Zhong J., Pan P., Xiao S., Ouyang X. (2022). Influence of eucalyptus plantation on soil organic carbon and its fractions in severely degraded soil in Leizhou Peninsula, China. Forests 13, 1606. doi: 10.3390/f13101606
Zhuang W.-E., Chen W., Cheng Q., Yang L. (2021). Assessing the priming effect of dissolved organic matter from typical sources using fluorescence EEMs-PARAFAC. Chemosphere 264, 128600. doi: 10.1016/j.chemosphere.2020.128600
Keywords: Suixi River, Zhanjiang Bay, dissolved organic carbon, chromophoric dissolved organic matter, fluorescent dissolved organic matter, mixing behavior
Citation: Liu P, Jiang K, Tan Q, Chen F, Jia R and Wang C (2025) Seasonal dynamics of dissolved organic matter in a small tropical estuary-coastal bay continuum: distribution, transformation, flux, and global perspectives. Front. Mar. Sci. 11:1511427. doi: 10.3389/fmars.2024.1511427
Received: 15 October 2024; Accepted: 23 December 2024;
Published: 21 January 2025.
Edited by:
Meilin Wu, Chinese Academy of Sciences (CAS), ChinaCopyright © 2025 Liu, Jiang, Tan, Chen, Jia and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Chao Wang, Y2hhb3dhbmdAZ2RvdS5lZHUuY24=