 Ingvill Tolas1,2*
Ingvill Tolas1,2* Zhigang Zhou3*
Zhigang Zhou3* Zhen Zhang3,4
Zhen Zhang3,4 Tsegay Teame3,5
Tsegay Teame3,5 Rolf Erik Olsen6
Rolf Erik Olsen6 Einar Ringø7
Einar Ringø7 Ivar Rønnestad1*
Ivar Rønnestad1*- 1Department of Biological Sciences, University of Bergen, Bergen, Norway
- 2Department of Biological Sciences, Norwegian University of Science and Technology (NTNU) Ålesund, Ålesund, Norway
- 3Sino-Norway Joint Lab on Fish Gut Microbiota, Institute of Feed Research, Chinese Academy of Agricultural Sciences, Beijing, China
- 4Faculty of Land and Food Systems, The University of British Columbia, Vancouver, BC, Canada
- 5Tigray Agricultural Research Institute, Mekelle, Ethiopia
- 6Norway-China Joint Lab on Fish Gastrointestinal Microbiota, Department of Biology, Norwegian University of Science and Technology, Trondheim, Norway
- 7Norwegian College of Fishery Science, Faculty of Bioscience, Fisheries and Economics, The Arctic University of Norway (UiT), Tromsø, Norway
The importance of the gastrointestinal microbiota (GM) in health and disease is widely recognized. Although less is known in fish than in mammals, advances in molecular techniques, such as 16S rRNA sequencing, have facilitated characterization of fish GM, comprising resident autochthonous and transient allochthonous bacteria. The microbial diversity and composition are strongly influenced by diet. High-protein diets, including alternative ingredients like plant and insect proteins, modify GM, impacting beneficial bacteria e.g. Cetobacterium. Lipids affect microbial metabolism and short-chain fatty acid (SCFA) production, while excessive carbohydrates can disrupt GM balance, causing enteritis. Dietary additives, including probiotics, prebiotics, and antibiotics, effectively modulate GM. Probiotics enhance immunity and growth, prebiotics support beneficial bacteria, and antibiotics, though effective against pathogens, disrupt microbial diversity and may promote antibiotic resistance. Environmental factors, such as temperature, salinity, and pollution, significantly influence GM. Elevated temperatures and salinity shifts alter microbial composition, and pollutants introduce toxins that compromise intestinal function and microbial diversity. Stress and pathogen infections further destabilize GM, often favoring pathogenic bacteria. GM communicates with the host via metabolites such as SCFAs, bile acids, and neurotransmitters, regulating appetite, energy metabolism, immunity, and neural functions. Additionally, GM influences the immune system by interacting with epithelial cells and stimulating immune responses. Despite recent advances, further research is needed to elucidate species-specific mechanisms underlying GM-host interactions, the ecological implications of GM diversity, and its applications in aquaculture to optimize fish health and performance.
1 Introduction
Given that fish-species make up nearly half of currently existing vertebrates, they have showed a great adaptability, and evolutionary traits, to different habitats including salinities, temperatures, and food preferences. This is especially true for the teleosts, the largest groups of fish existing today. It has been suggested that the adaptability in part is helped by the third gene duplication event occurring in teleosts, and the fourth in salmonids. Since each gene duplication theoretically doubles the number of paralog genes, over-represented genes can therefore be used to drive evolution through sub-functionalization and neo-functionalization (Long, 2010). Importantly, fish have shared their evolutionary journey with microorganisms living on or within them. Because these microorganisms, bacteria, yeast, fungi and virus, have evolved concomitantly with their hosts they are invariably intertwined and for many there may be a symbiotic relationship or common dependencies where fish and microbiota jointly affect each other growth and development. Understanding the role of the microbiota is therefore key to understanding the physiology of teleost fish species.
The microbiota is unique to an organism and specific to a system. Among the various populations of microorganisms in and on teleost fish, many reside within the gastrointestinal tract (GIT), making up the gut microbiota (GM). As for mammals, fish GM is believed to play important roles in regulating many physiological functions in fish including immunity, digestion, gut endocrine and neuronal signaling, drug metabolism, and more (Egerton et al., 2018). Although studies evaluating the GM of fish are few compared to those performed in humans and other mammals, the expansion of the aquaculture industry and development in microbiome research has resulted in renewed interest in the field. In fact, the discovery that the regulation, or dysregulation, of the GM may significantly affect the host welfare, growth, performance and disease resistance, has made GM investigations a hot topic of research for those interested in optimizing aquaculture procedures and promoting sustainable practices (Chen et al., 2022). In the present review, we discuss how variables such as diet, stress and environmental factors affect the GM, and how the microorganisms, and their metabolic products affect pathways involved in the communication between the GM and the host and finally the effect of GM-signaling on the host (Figure 1).
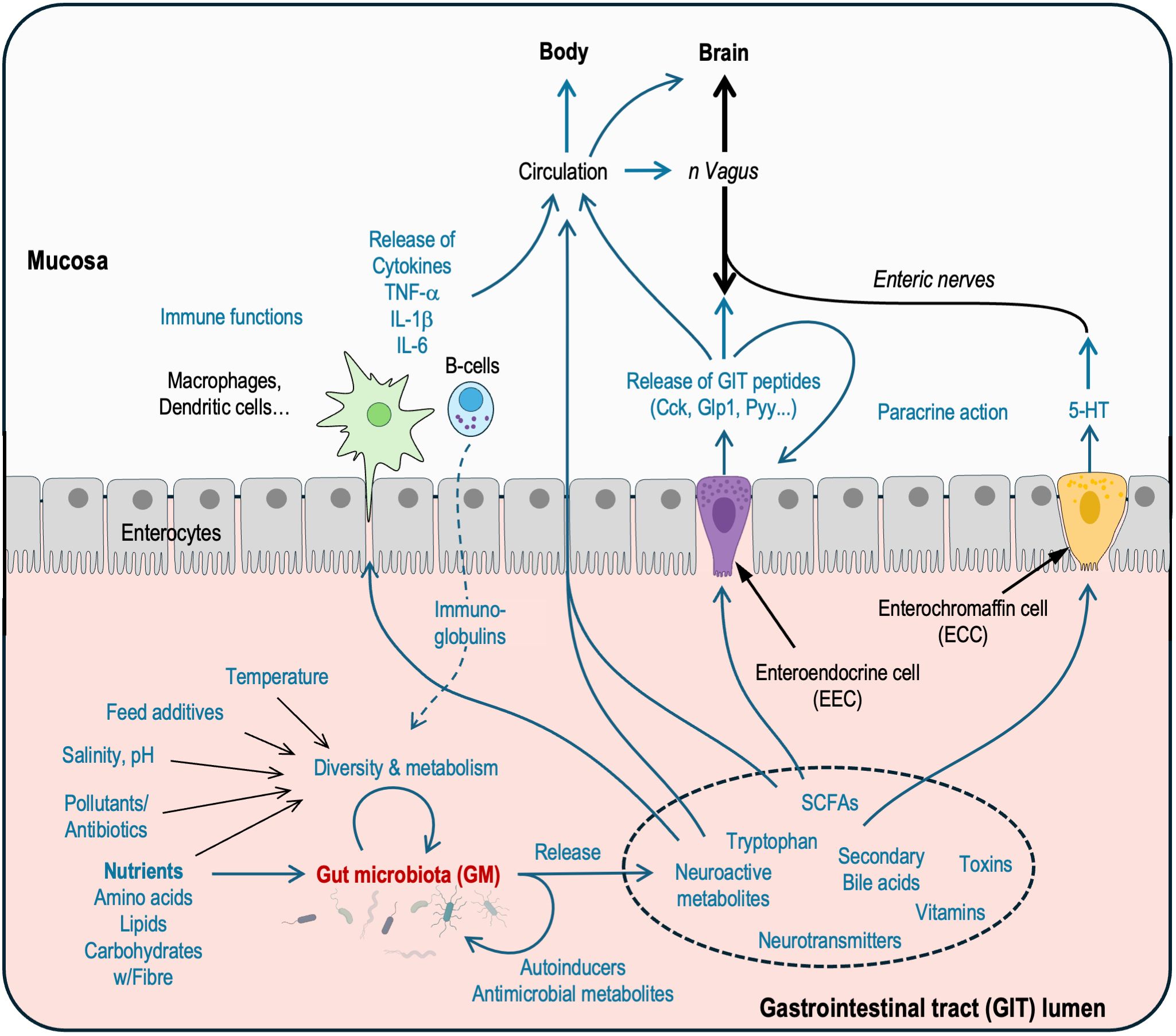
Figure 1. Schematic representation of proposed mechanisms underlying the bidirectional interactions between gut microbiota (GM) and gastrointestinal tract (GIT) tissues, other organs and the brain. GM diversity and thereby metabolism are influenced by factors such as nutrients, feed additives, temperature, salinity, and pollutants. The GM produces various metabolites, including short-chain fatty acids (SCFAs), neuroactive molecules, secondary bile acids, and vitamins, which act locally in the GIT lumen or on cells in the mucosal layers or reach other target tissues and organs via circulation and neural pathways, such as the vagus nerve. The GM released metabolites stimulate nutrient sensors on the enteroendocrine cells (EECs) modulating secretion of bioactive GIT peptides that mediate paracrine and systemic effects. Enterochromaffin cells (ECC) secrete serotonin (5-HT) which modulate activities of the enteric nervous system (ENS). The bidirectional communication via metabolites also involves ENS, immune cells such as macrophages, dendritic cells, and B-cells thereby influencing mucosal immunity and the GM. The description of mechanisms is not exhaustive, many are adapted from mammals, and remain to be fully elucidated in fish. See text for further discussions. (Based on Cryan et al, 2019; Butt and Volkoff, 2019; Blanco et al., 2021) Cck, Cholecystokinin; Glp1, Glucagon-like peptide 1; IL, Interleukin; Pyy, Peptide YY; SCFA, Short-chain fatty acid; TNF, Tumor necrosis factor.
2 Compartmentalization of the GM
Due to the development of new molecular techniques targeting the 16SrRNA, 28S rRNA genes and other genetic indicators, characterizing and analyzing of intestinal microbiota of fish and other animals has become rather straight forward. These approaches have been used to investigate the crucial role of gut microbiota to the host development and overall health (Huang et al., 2020; Johny et al., 2021). In most animals, the GM can be broadly separated into two subgroups, the autochthonous and allochthonous microbiota. The autochthonous, or resident/indigenous bacteria can often be associated to the part of mucus layer that overlies the epithelium, they can be embedded in the intestinal folds, or anchored to cells through different mechanisms, including production of adhesion molecules, formation of biofilms and/or by modulating the immune response of the intestine, which can in turn influence the anchoring of the microbes to the intestinal cells (Zhang et al., 2023a). These microorganisms will have developed mechanisms to survive the various defensive mechanisms produced by the host such as antimicrobial peptides. Like in many animal species, a small part of the autochthonous microbiota in fish is likely to become established during early life and they diversity may increase as the fish grow (Yatsunenko et al., 2012; Egerton et al., 2018). The autochthonous microbiota is known to be particularly important for the development of intestinal tissue, and intestinal immunity (de la Torre Canny et al., 2021). The other group of bacteria in the GIT are allochthonous bacteria, that are regarded as transit bacteria in the feces. This means that they are transported through the intestine and will leave when feces exit the fish. They will not colonize any habitat except under abnormal conditions. These are often quantitatively more dominant than the autochthonous bacteria. However, the autochthonous microbiota appears to be more stable than the allochthonous. For example, in gilthead seabream (Sparus aurata) changes in fishmeal (FM) was shown to have no effect on the mucosal (autochthonous) microbiota, while causing major changes in the total (allochthonous) microbiota (Fontinha et al., 2021). The population of microorganisms will also vary along the GI, affected by numerous factors such as pH, host enzymes (proteases, lipase), redox potential, mucin secretion, peristaltic activity, and nutrient availability (Hao and Lee, 2004).
3 Variables affecting the GM in teleost
Owing to the potential and demonstrated importance of the fish GM to host physiology, studies on the subject are rapidly growing. One of the most exciting topics relates to how the microbiota is established and maintained, and the mechanisms which determines its composition. There are numerous factors, both intrinsic and extrinsic, that will influence the gut microbiome of teleost fish, but the complexity of regulatory mechanisms renders it difficult to define the magnitude of influence of each separate factor. The present review aims to present an updated overview on how some selected variables may impact the GM.
3.1 Dietary effect on GM
It is well known, that the host’s diet is the key nutrient source for the GM and will therefore have a profound effect on GM density, composition and diversity (Ringø et al., 2016), and can lead to significant changes in the host’s gut function. After the introduction of next generation sequencing (NGS) our understanding of dietary effects of on GM has increased massively in all species studied such as golden pompano (Trachinotus ovatus) (Zhao et al., 2020b), Nile tilapia (Oreochromis niloticus) (Limbu et al., 2019), rainbow trout (Oncorhynchus mykiss) (Desai et al., 2012), gilthead sea bream (Sparus aurata) (Estruch et al., 2015), Arctic charr (Salvelinus alpinus) (Nyman et al., 2017), field eel (Monopterus albus) (Peng et al., 2019), yellowtail kingfish (Serio lalalandi) (Wilkes Walburn et al., 2019), olive flounder (Paralichthys olivaceus) (Niu et al., 2020). Each nutrient will have some signature effects by selecting some groups of bacteria able to utilize or utilize properties of the nutrient. However, in practical feeding, diets are composed of mixtures of many nutrient components (eg soybean meal contains soybean protein, soy lipid, many and variable antinutritional factors) so that the specific effects may be masked. In the following, we will give some typical examples of observations when trial have focused on groups of nutrients.
3.1.1 Dietary proteins
Fish require high protein diets for optimal growth and to thrive. Wild carnivorous fish get their protein largely from preying on species that are part of their natural diets, and FMs have historically been used in aquaculture feeds to mimic this. However, with limited availability of marine proteins, alternatives to FM is finding its way into fish farming. This includes proteins from plants, animal by-products and insects (Tacon and Metian, 2008; Hartviksen et al., 2014; Dani, 2018; Alfiko et al., 2022). When fed to fish, there will also be some members within a microbial population that can utilize some of the protein components better than others and have a distinct growth advantage. Since the main component of these protein sources are amino acids any variation in the content and composition will select for some microorganisms. Typical microbial use of amino acids includes synthesis of proteins needed for bacterial physiological functions, energy production through deamination, decarboxylation or desulfurization. Furthermore, the intestinal microbiota can utilize amino acids as substrates for fermentation which allows the gut microbiota to produce various essential fermentation metabolites also needed for the normal function of the host (Zhao et al., 2019). These include short-chain fatty acids (SCFAs) which will be covered in section 4.1.
The amino acid driven alteration of diversity and composition of the GM can also be modulated by the different types of metabolites produced, including nitric oxide, ammonia, polyamines, hydrogen sulfide (H2S), indoles, and phenols in both the proximal and distal intestine (Dai et al., 2011; Macfarlane and Macfarlane, 2012). For example, the use of amino acids for deamination will increase in bacterial species possessing aminotransferases like alanine- and aspartate-aminotransferases and will lead to increased accumulation of ammonia. Ammonia is toxic and can therefore limit the survivability of some microorganisms, while selecting for those microorganisms that can use ammonia as electron donors and utilize it as an energy source. Therefore, high amounts of some amino acids may promote the growth of beneficial bacteria and inhibit the growth of pathogenic bacteria by influencing the host immune system, which will be discussed in section 5.2.
Altered abundance and structure of the GM can affect apparent nutrient digestibility in fish by excreting digestive enzymes (Ray et al., 2012; Wang et al., 2015; Ringø et al., 2016; Ma et al., 2017). In rainbow trout, reductions in nutrient utilization was observed along with a reduction in the alpha diversity of GM following increasing inclusions of plant proteins (Michl et al., 2017). Plant based protein sources, especially soybean meal is commonly used as protein sources in carnivorous fish diets. They are often heavily processed to remove antinutritional factors, as in soybean concentrates. However, some antinutrients will always be present, and many yet uncharacterized are assumed to be antimicrobial. Therefore, giving diets containing plant meals to fish will often induce massive changes in the intestinal microbiome, with a selective advantage to those microorganisms that are tolerant to or even able to utilize these antinutritional factors. Following a shift in diets from animal- to plant-based diets for grass carp (Ctenopharyngodon idella), the abundance of Bacteroides, Erysipelotrichaceae and Lachnospiraceae in the GM increased while the abundance of Cetobacterium decreased (Hao et al., 2017a). Likewise, the abundance of Lactococcus lactis subsp. lactis was increased, while that of Weissella confusa was decreased in Atlantic salmon (Salmo salar) GM fed a diet supplemented with 20% soybean meal (Reveco et al., 2014). In rainbow trout, several studies have shown to a clear trend towards increased ratio of Firmicutes to Proteobacteria when FM is replaced by various plant-based protein ingredients (peas, soybean, canola) (Desai et al., 2012; Blaufuss et al., 2020; Pérez-Pascual et al., 2021). Since many common pathogens are within the Gram negative phylum Proteobacteria (like Vibrio), a reduction of members of this phylum is often regarded as advantageous for GIT health (Rimoldi et al., 2018a). The specific effects will however vary with species and detailed nature of the protein sources. In grass carp, replacing FM to plant protein increased the richness of the genus Bacteroides and families Erysipelotrichaceae and Lachnospiraceae while the abundance of the beneficial bacterium Cetobacterium of the family Fusobacteriaceae was reduced (Hao et al., 2017b).
In recent years, insect meals have become interesting alternatives to FM having good amino acid profiles and sustainable. It is also interesting to note that for many fish species, insects are part of the natural diets. In rainbow trout, replacing 50% of FM with a diet containing 15% of black soldier fly (Hermetia illucens) larvae protein increased the abundance of phylum Firmicutes, Lactobacillus and Bacillus, while reducing Proteobacteria, Aeromonas (Rimoldi et al., 2021). In Atlantic salmon replacing a commercial diet with an insect meal supplemented enriched GM with members of the Bacillaceae family (Li et al., 2022b). The study also showed that insect meal increased alpha diversity in the mucosa (autochthonous), although not in the digesta. This contrast other studies using Atlantic salmon (Li et al., 2022a) and rainbow trout reported increased alpha diversity in the digesta (Terova et al., 2019; Bruni et al., 2018). The discrepancy is probably related to external factors such as trial conditions, insect processing method, insect species or analytic approaches. When the GM response to insect feeding is compared across species (seabass, seabream and rainbow trout), responses vary significantly (Antonopoulou et al., 2023). Nevertheless, the general impression is that replacing FM with insect meal selects for a “good” GM.
There is also an increased trend to replace fishmeal with other terrestrial protein sources such as poultry by-products. The effects do vary, but in several aspects, they have elements of results from using plant proteins. For example, in Nile tilapia, animal protein increased GM Proteobacteria and reduced Bacteroidetes and Cetobacterium (Pereira et al., 2024). Replacing FM with poultry by-products in gilthead seabream (Sparus aurata) diets increased both the abundance and diversity of the intestinal microbiota of the fish (Fontinha et al., 2021), and partial replacement of FM with a combined poultry by-product and vegetable protein meal increased the ratio of Firmicutes to Proteobacteria in GM of rainbow trout (Rimoldi et al., 2018b).
3.1.2 Dietary lipids
Fish oil has always been an important oil source in fish diets due to their high content of the essential omega-3 fatty acids eicosapentaenoic acid and docosahexaenoic acid. However, with an expanding aquaculture industry, the availability is limited and the cost is high. In a previous comprehensive review, Ringø et al. (2016) discussed the dietary effect of lipid on fish gut microbiota, and today alternative lipid sources are now being included into fish diets in increasing amounts. These mainly include plant-based sources, and to a lesser extent other alternatives like algal products. Dietary lipids will surely affect the GM. The mechanisms are less well known than for proteins. However, lipids are a energy sources and building blocks for bacterial growth and metabolism. As such, different types of lipids such as saturated, unsaturated fats, and omega-3 fatty acids have varying effects on the growth and composition of gut bacteria. Moreover, lipids are digested by both host and bacterial lipases, and fatty acids can be degraded for energy release, thereby producing a long range of fatty acids (long, short, unsaturated, double bond position) which are known to be antimicrobial to many bacterial species. In this sense, the type of lipid consumed by the host constitutes a selective process for the microbes residing in its intestine (for reviews see Ringø et al., 2016; Turchini et al., 2022). Although less studies, the microbes can also have indirect effects though modulations of host the immune and other physiological effects (Zhang et al., 2012; Tanca et al., 2018).
In an interesting study, Ma et al. (2018) revealed that microencapsulated lipid increased the abundance of beneficial bacteria such as Bacillus and Paenibacillus and reduced pathogenic bacteria like Pseudoalteromonas and Roseovarius, when compared to normal lipid inclusion. Furthermore, they showed that capsules containing 60-40 fish oil (FO)-linseed oil had less effect on the GM than capsules containing 60-40 palm oil-linseed oil. In Atlantic salmon, Pseudomonas jessenii/fragi-like strains were the predominant GM species in fish fed a sunflower oil supplemented diet, while Pseudomonas submarinus/marincola-like strains and Staphylococcus spp. were dominant when fed a rapeseed oil diet (Ringø et al., 2016). This findings contrasts with the study of Atlantic salmon where Nikouli et al. (2021) reported no effect on GM composition when fish were fed fish-oil or plant oil-based diets. A general lack of effect was also noted by Huyben et al. (2020) in gilthead seabream where replacement (40%) of FO with camelina (Camelina sativa) oil had no effects on GM. However, increasing the level to 60% reduced the abundance of intestinal Lactobacillus. In juvenile golden pompano (Trachinotus ovatus) more saturated fatty acids increased GM diversity compared to fish fed more unsaturated fatty acid (Guo et al., 2021). In turbot (Scophthalmus maximus) feeding small amounts of oregano oil (1ml/kg) increased the relative abundances of some beneficial bacteria such as Akkermansia, Bifidobacterium and Faecalibacterium and reduced the abundance of Listonella and Sphingomonas compared to fish fed FOs (Guangxin et al., 2022). Feeding Tiger puffer (Takifugu rubripes) various terrestrial oils (soybean, palm, beef tallow) for 50 days impaired intestinal functions by reducing the abundance of some butyric acid-producing bacteria (Lachnospiraceae, Eubacterium, Butyricicoccus, Clostridium and Roseburia) and bacteria related to digestion and absorption (Sphingomonas, Romboutsia and Brevibacillus) (Kong et al., 2023). An interesting finding in this study was that refeeding with FO modulated and restored the GM composition.
Although most studies so far have looked at the effect of oil substitutions, there has also been some studies on the effect of single fatty acids on GM. In Arctic charr, Ringø et al. (1998) reported higher population level of intestinal lactic acid bacteria (LAB) in fish fed 7% linolenic acid (18:3 n-3) or 4% of long chain-omega-3 PUFA (LC n-3 PUFA) compared with the fish fed linoleic acid (18:2 n-6). Feeding of gilthead seabream, a combination of short and medium-chain 1-monoglycerides (SILOhealth 108Z) increased the number of beneficial gut bacteria including Lactobacillus while reducing the number of Gammaproteobacteria (phylum Proteobacteria). Atlantic salmon fed low amounts of LC-n-3PUFA had a higher abundance of Firmicutes while fish fed high levels had higher levels of Proteobacteria (Jin et al., 2019). Feeding oxidized oil to rice field eel (Monopterus albus) reduced GM alpha diversity and abundance of Clostridia, while increasing Fusobacteria (Cetobacterium), which led to gut microbiota dysbiosis (Peng et al., 2019).
From the accumulated data it appears that dietary lipids will affect GM of fish and will select for several groups of bacteria producing compounds like SCFAs that will affect gut-brain and GM signaling. However, the effects appear more variable and sometimes contradictory even within species. Some studies also vary the lipid so much that the effects observed may be more due to oxidative stress or essential fatty acid deficiency rather than a consequence of microbial metabolism.
3.1.3 Dietary carbohydrates and fiber
Carbohydrates will most certainly affect the GM of aquatic animals. This is especially true for fibers (carbohydrate chains that cannot be degraded by the host often due to β 1→4 glycosidic bonds) since many microbial species will degrade and utilize them. The bacterial utilization and selection will depend on the type of fiber. Fibers have been reported to be beneficial in both animals and fish (e.g., DeVries et al., 2001; Ringø et al., 2010; Cerezuela et al., 2013; Sonnenburg and Sonnenburg, 2014). Typical results are increasing GM diversity as in largemouth bass dietary fibers (Lin et al., 2020) and total number of bacteria as in Caspian whitefish (Rutilus frisii kutum) (Mirghaed et al., 2018). Carnivorous fish normally consume very little carbohydrates and fibers. But in farmed species, fibers have become a major feed ingredient and will affect microbiota.
Increasing the dietary content of carbohydrates will, in general, have negative consequences on the GM in fish and can cause physiological dysfunction (Boulangé et al., 2016). In largemouth bass fed 15% corn starch reduced the abundance of beneficial bacteria including Lactobacillus, while the abundance of potentially pathogenic bacteria increased such as Brevundimonas and Ralstonia (Zhou et al., 2021). Zhang et al. (2021) noted that feeding Chinese perch (Siniperca chuatsi) high levels of carbohydrates reduced gut lactate content, microbial diversity and the quantity of beneficial bacteria. In the high carbohydrate group, Mycoplasma was the predominant species while butyric acid-producing bacteria and butyric acid level was reduced. These are well recognized as triggers for enteritis. Likewise, feeding 30% of corn starch to Japanese seabass (Lateolabrax maculatus) resulted in a reduction of Firmicutes and Bacillus, while increasing the relative abundance of Proteobacteria and Bacteroidetes at the phyla level, and Photobacterium and Paraclostridium at genera level causing lipid deposition and glucose metabolism disruption (Zheng et al., 2023). But as for the other nutrients, results can vary between species and trials. In grass carp, Zhang et al. (2023c) showed that low dietary levels of carbohydrates (100g/kg of corn starch) increased the abundance of pathogenic bacteria (Ralstonia and Elizabethkingia) in the GM, while 400 g/kg increased the abundance of Flavobacterium.
3.2 Probiotics, prebiotics and antibiotics
As mentioned above, several dietary ingredients will alter the GM. In many cases, this shift may be harmful to the host, while others have beneficial effects (see Nayak, 2010; Caipang et al., 2020; Dawood et al., 2020; Hasan and Banerjee, 2020; Ringø et al., 2020, Ringø et al., 2022; Allameh et al., 2021; Vargas-Albores et al., 2021). In addition to these indirect and occasionally inadvertent effects of dietary components on the composition and abundance of the GM, intestinal microbes can also be altered via direct measures, such as through the addition of prebiotics or probiotics. These components are therefore added in a deliberate attempt to improve the health of the host. Other components such as antibiotics are used to prevent and treat bacterial diseases in fish, but will as a side effect also affect other bacteria in the GM. The known impacts of these additives are present in the following sections.
3.2.1 Probiotics
To enable beneficial microorganisms to become established in the GIT, it is becoming more common, in practical aquaculture, to add one or more “good” live bacteria directly into the diets (Druart et al., 2014; Vargas-Albores et al., 2021). This is termed a probiotic approach where these bacteria should become established in the host intestine. Here they would change the diversity, composition and function of the fish gut microbiota through several mechanisms such as competing with the harmful bacteria for attachment sites and nutrients thereby reducing the growth and colonization of pathogenic bacteria. Moreover, probiotics produce different types of antimicrobial compounds, which inhibit the growth of harmful bacteria and parasites in the intestine of the fish (Chen et al., 2024; Ferdous et al., 2024). The positive modulation of the gut microbiota in fish in turn resulted in improvement in the growth, gut health, nutrient metabolism, immune response and disease resistance of the host (Qi et al., 2024). The most commonly used probiotic species in aquaculture include Pediococcus acidilactici (Bactocell) Lactobacillus, Bacillus, Lactococcus and Saccharomyces (e.g. Nayak, 2010; Caipang et al., 2020; Ringø et al., 2020)The effects of probiotics on GM of teleosts are summarized in Table 1.
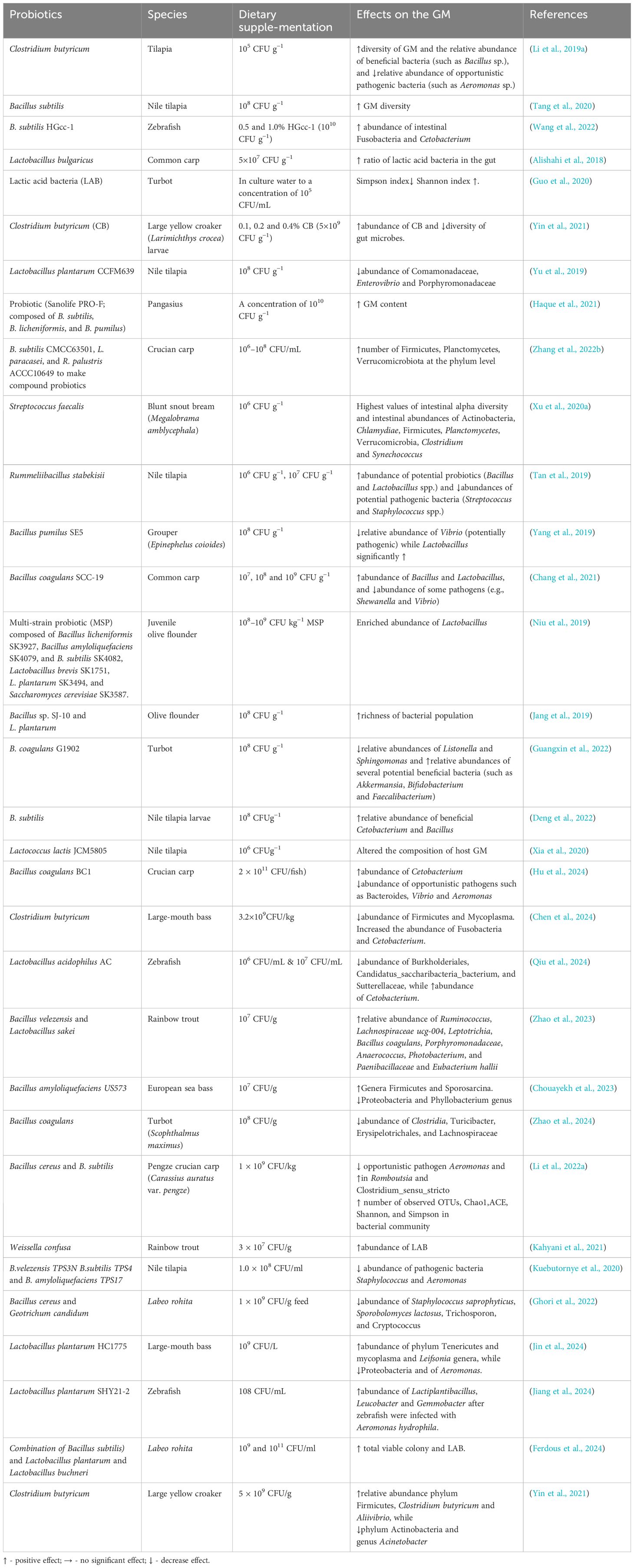
Table 1. Summary of effects of probiotics added to fish species.
3.2.2 Prebiotics
An alternative, and more practical approach than adding live bacteria, is to include the food components that the “good” probiotic bacteria would need to become established in the GI. This is the prebiotic approach. Most prebiotics are non-digestible or indigestible fibers that are utilized by beneficial bacteria (probiotics) for their growth. The presence of a normal gut microbiota enhances the efficiency of nutrient absorption and metabolism by stimulating the growth of beneficial bacteria that help break down and metabolize nutrients in the gut. Furthermore, prebiotics played a significant role in modulation of the immune response by promoting the growth of beneficial bacteria that support a balanced immune function, thereby enhancing the fish’s ability to resist pathogens and diseases. Positive modulations of gut microbiota by prebiotics also help to maintain the integrity of the gut barrier and promote the production of beneficial metabolites such as short-chain fatty acids, which have anti-inflammatory and protective effects on the gut lining and reduce gut-related disorders in the fish. Reduction of pathogen colonization of the intestine due to computing for space and nutrients with the beneficial bacteria which can promote their proliferation because of prebiotics is another scenario of the application of prebiotics in aquaculture. Many studies demonstrated that incorporating prebiotics into the diet of fish can help maintain a healthy gut microbiome and support overall fish health and performance (Merrifield et al., 2010; Ringø et al., 2010, Ringø et al., 2014; Druart et al., 2014; Davani-Davari et al., 2019; Dawood et al., 2020). Prebiotics used in aquaculture mainly include mannan-oligosaccharide (MOS), β-glucan, fructo-oligosaccharide (FOS), xylooligosaccharide (XOS), galactooligosaccharide (GOS) and inulin (Dawood and Koshio, 2016; Nawaz et al., 2018; Dawood et al., 2020; Yukgehnaish et al., 2020). Modulation of the GM following administration of prebiotics are summarized in Table 2.
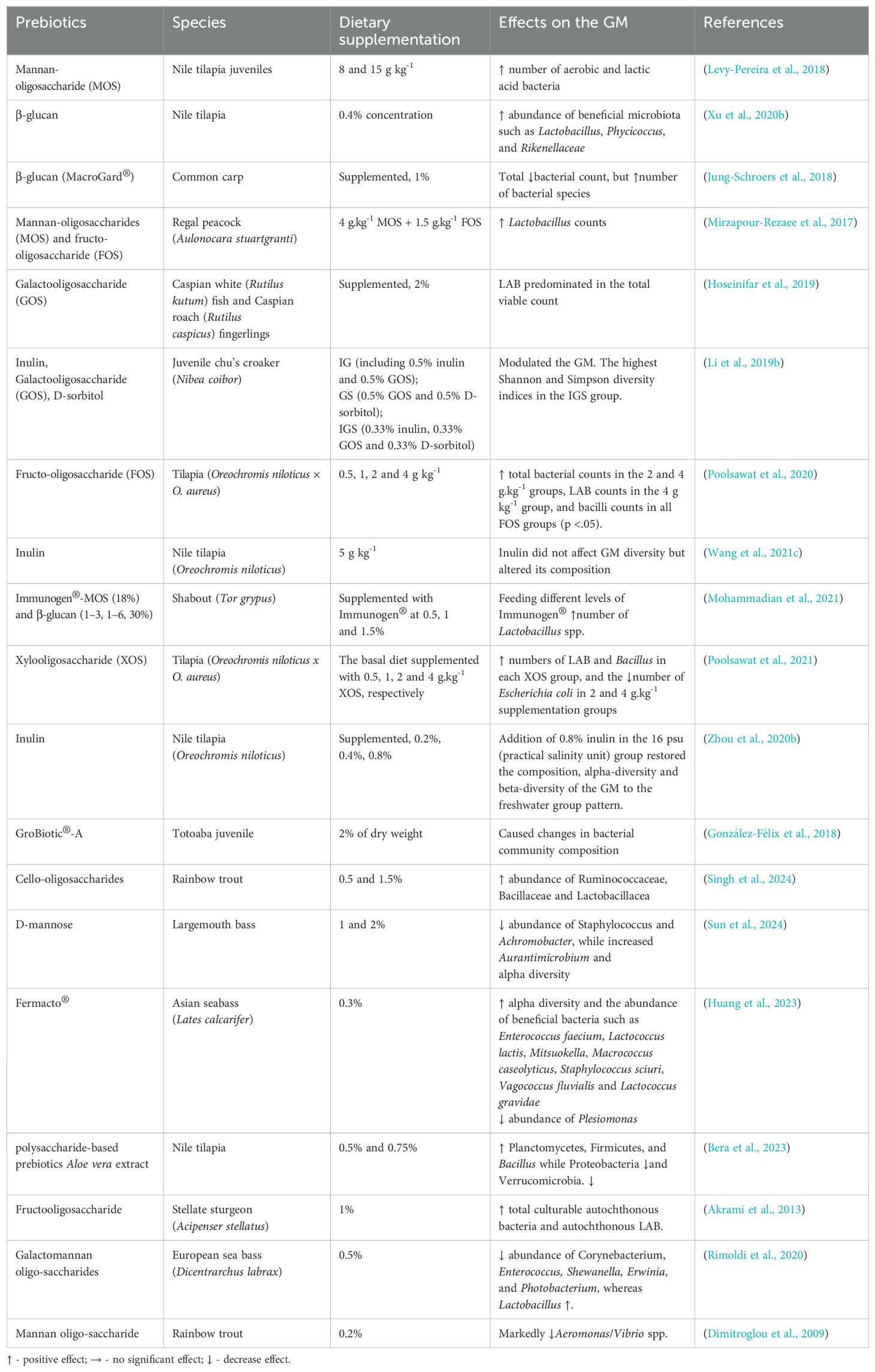
Table 2. Summary of modulatory effect of prebiotics on GM of fish species.
3.2.3 Antibiotics
With the rapid growth of aquaculture production there has been a concomitant increase in disease outbreaks, and to combat such outbreaks, antibiotics have been utilized. While the use of antibiotics in aquaculture varies substantially between different countries, antibiotics have been detected in most aquatic environments, of which human medicine, terrestrial animal farming and aquaculture are the main sources (Kim et al., 2017; Danner et al., 2019; Bojarski et al., 2020). Antibiotics function by inhibiting bacterial survival/growth through various mechanisms, such as targeting the structural integrity of the bacterial cell wall, inhibiting bacterial protein biosynthesis, inhibiting of bacterial DNA replication or inhibiting of bacterial folic acid metabolism (Kapoor et al., 2017). Importantly, these antimicrobial mechanisms target all sensitive species in the “natural” bacterial community in addition to the disease-causing agents, thus selecting for those species that are resistant to the treatment. In turn, this results in an alteration in the diversity and composition of fish GM. Indeed, the use of antibiotics cause an imbalance of fish gut microbiota and can lead to dysbiosis. Dysbiosis is defined as the loss of beneficial microbes, expansion of pathobiont microbes, or loss of the diversity of the microbes (Petersen and Round, 2014).
In line with this, Limbu et al. (2019) determined that oxytetracycline exacerbated the adverse effects of a high-fat diet on the gut of juvenile Nile tilapia, leading to microbiota dysbiosis. The effect of a combination of antibiotics containing vancomycin, enrofloxacin, florfenicol, and metronidazole at different doses on the gut microbiota of grass carp indicated that at higher doses (at 2 mg/fish/d except for vancomycin at 1 mg/fish/d) increased the abundance of Proteobacteria, while the abundance of Fusobacteria and the genus Cetobacterium was reduced (Sun et al., 2021). Furthermore, olaquindox in zebrafish, florfenicol in channel catfish and a combination of kanamycin, gentamicin, colistin, metronidazole, and vancomycin in Devils Hole pupfish (Cyprinodon diabolis) resulted in the decline of the relative abundance of Cetobacterium in the intestinal contents (He et al., 2017; Wang et al., 2019; Bhute et al., 2020). A study performed by Almeida et al. (2019) showed that treatment of zebrafish with oxytetracycline led to increased relative abundance of the Rhodobacter genus from phylum Proteobacteria. Administering oxytetracycline or amoxicillin to olive flounder reduced the diversity and quantity of GM (Kim et al., 2019). Carlson et al. (2017) showed that rifampicin reduced the diversity and composition of gut microbial communities in Western mosquitofish (Gambusia affinis). Administration of florfenicol-containing diets to catfish increased the relative abundance of Plesiomonas and Aeromonas (Wang et al., 2019). In zebrafish, olaquindox reduced the abundance of Cetobacterium and increased Enterobacter (He et al., 2017). Wang et al. (2020) established that norfloxacin caused a significant decrease in the diversity and relative abundance of Proteobacteria in the GM of juvenile large yellow croaker (Pseudosciaena crocea), while there was an increase in Tenericutes. Short-term antibiotic treatment of the hybrid grouper (Epinephelus fuscoguttatus♀ × E. lanceolatus♂) significantly reduced the number and diversity of gut microbes, the abundance of Firmicutes, and the ratio of Firmicutes/Bacteroidetes (Xu et al., 2022). Injection, oral administration and soaking of Nile tilapia with enrofloxacin for 48 h resulted in a decrease of the relative abundance Corynebacterium, Clostridium sensu stricto_3 and Bacillus and an increase in Akkermansia, Ralstonia and Romboutsia genera in the intestine of the fish (Chen et al., 2023). In addition, feeding Nile tilapia with a diet containing oxytetracycline (100 mg/kg/day) for eight days resulted in an increment in the abundance of Plesiomonas (Payne et al., 2021). Another study performed by Payne et al. (2022) demonstrated that supplementation of oxytetracycline (35 mg/kg body weight of fish/day) to the diet of rainbow trout for seven days led to a reduction of Mycoplasma and Bacillus, while increasing the abundance of Aeromonas, Deefgea and Pseudomonas in the intestine of the fish.
These and other studies clearly demonstrate that the effects of antibiotics on the gut microbiota of fish can vary depending on the type of antibiotics, dosage, duration of application, and fish species. Antibiotics can change the diversity and composition of the GM of fish, and the changes caused by different types of antibiotics may be positive or negative. In addition to their direct selection of specific microbes, antibiotics can also affect the gut microbiota indirectly by several mechanisms including, alteration of the immune system and overall health of the fish, which in turn can impact the gut microbiota diversity and structure, and this can result in alteration in metabolism of nutrients, immune functions, resistance to diseases and overall health and development of the fish. The development of antibiotic resistance microbiome in the gut of the fish will also affect the normal microbiota profile in the gut of the fish as well as the environmental microbiome structure due to the inappropriate of prolonged application of antibiotics in the fish. As in animal species, the number of antimicrobial resistance genes in pathogenic bacteria have increased, and due to this the effectiveness of antibiotics has been reduced along with increased occurrence of infectious diseases in the aquaculture environments (Santos and Ramos, 2018; Dube and Okuthe, 2024). It has been reported that mutation or horizontal transfer of resistance genes through conjugation, transformation and transduction are the main ways for the development of antibiotic resistance bacteria (Spetz, 2003; Munita and Arias, 2016). This transferring of genes and/or plasmids might contaminate the whole population in an aquaculture environment and affect the diversity and composition of gut microbiota of the fish through promoting the proliferation of potential pathogens and hindering the growth of beneficial bacteria in the gut of the fish (Ruzauskas et al., 2018; Preena et al., 2020). Therefore, it is crucial to apply antibiotics cautiously, and properly (in the time of application as well as dose) using the proper guidelines to reduce the negative effects on the gut microbiota of fish and utilization of alternative strategies will be important rather than depend entirely on antibiotics for treatment of aquatic animal diseases.
3.3 Environmental factors
While diet and dietary additives such as pro-, pre and antibiotics play major roles in the formation of the gut microbiota, the water environment provides a complex and diverse living environment for aquatic animals with many important factors affecting the GM (Spanggaard et al., 2000). Among them, temperature, salinity, inorganic salts (especially ammonia nitrogen and nitrite) and pH are important parameters. In addition, heavy metal ions, pesticides and other substances in diets or the water environment will also impact fish GM (Navarrete et al., 2008).
3.3.1 Temperature
Fish are ectotherm animals, and the diversity, structure, and abundance of GM change with water temperature. Seasonal changes are a natural part of many fishes’ life cycle and will often affect the gut microbiota. Indeed, Al-Harbi and Naim (2004) observed that the relative abundance of GM in Oreochromis niloticus × Oreochromis aureus varied significantly with seasons. Similar findings were made by Hagi et al. (2004) in common carp, silver carp, channel catfish (Ictalurus punctatus) and deepbodied crucian carp (Carassius Carassius). They noticed that the dominant LAB during summer was L. lactis while L. raffinolactis dominated during winter. In an extended experiment, Hovda et al. (2012) analyzed the GM of Atlantic salmon during a one-year cycle where water temperatures varied between 5,5 and 18,8°C. They concluded that while there were some small variances in the composition, the overall changes in the GM were not significant. In contrast to this, Neuman et al. (2016) conducted a study in Tasmanian Atlantic salmon fed two different diets at changing water temperatures. They documented that the differences in GM from the dietary groups increased when the water temperatures increased from 10.1 to 18.5°C. This showed that water temperature was the main driver of changes in the GM of these fish. These discrepancies could be owed to other environmental factors differing between the trials, such as ratio of temperature shifts, bacteria present in water etc.
Global warming and its side effects of increased frequencies of extreme weather will impact all aspects of life (Garrabou et al., 2022). This is also likely to affect numerous aspects of intestinal microbiota in affected fish species. Sánchez-Cueto et al (2023) simulated heat wave conditions and established that elevated temperatures substantially influenced the microbiota composition and decreased alpha diversity in greater amberjack (Seriola dumerili). Huyben et al (2018) showed lower bacterial diversity and abundance of LAB, especially Lactobacillus, in rainbow trout reared in 18°C rather than 11°C for 6 weeks. A decreased abundance of Lactobacillus and overall lowered bacterial diversity in rainbow trout was also demonstrated by Zhou et al. (2022), in a study which also established that the number of Mycoplasma, Firmicutes, and Tenericutes increased significantly following acute heat stress. Acute heat stress has also been shown to impact the GM of gilthead seabream by facilitating increased growth of the potential pathogenic Streptococcus in fish fed high lipid diets (Pelusio et al., 2021). Jones et al. (2018) detected significant differences in the community structure and diversity of GM of Mottled spinefoot (Siganus fuscescens) in temperate-, subtropic- and tropic-waters and Soriano et al. (2018) showed that the predominant bacterial families in yellowtail kingfish (Seriola lalandi) reared at low water temperatures (20°C) were Saprospiraceae, Comamonadaceae and Micavibrio, changing to Pseudomonadaceae, Alcaligenaceae and Microbacteriaceae at higher water temperatures (26°C). By rearing chinook salmon (Oncorhynchus tshawytscha) at 8, 12, 16 and 20°C, Steiner et al (2022) established that elevated temperature had a significant effect on the gut microbiota, with the dominant taxa gradually changing from Vibrionaceae to Fusobacteriaceae. Importantly, these changes were significantly correlated with the physiology and health of the fish, affecting parameters such as histology, fatty acids, plasma biochemistry and hematology. In chum salmon (Oncorhynchus keta) exposure to temperatures above or below the control temperature triggered significant dysbiosis in the fecal microbiota (Ghosh et al., 2022). The authors showed that Vibrio and Tenacibaculum sequence variants were highly abundant at high and low temperatures, respectively, and importantly concluded that the opportunistic growth of these pathogenic species may impede host immunity.
From the above-mentioned studies it is clearly showed that temperature is an important factor affecting the composition and diversity of the GM. This is owed to the natural ability of some bacterial species and strains to thrive at specific temperatures as well as their differing potential to adapt to shifts in surrounding temperatures. Importantly, this may in turn impact the well-being of the fish. Though temperature is a deciding factor for the microbiota, the host-specific composition and diversity of the GM means that the response to shifts in temperature is highly species dependent. For example, by exposing two invasive fish species (common carp and largemouth bass) to increased temperature, Zhang et al. (2023b) were able to determine that the GM communities in common carp changed significantly within one week, while the GM communities of the largemouth bass exhibited no significant changes. Concomitantly, the researchers identified 10 predicted bacterial functional pathways in common carp that were temperature dependent, while no functional pathways were identified in largemouth bass. In sum, they concluded that the GM of the common carp was more sensitive to temperature fluctuation and more plastic than that of largemouth bass. Notably, the plasticity of fish GM in response to temperature is not only species dependent but appears to be linked to the overall thermal tolerance of the host. In fact, Kokou et al (2018) conducted an experiment in which they performed transgenerational selective breeding cold tolerance in tropical tilapias and concluded that the microbiomes of cold-resistant fish showed higher resilience to temperature changes, indicating that the microbiome is shaped by its host’s selection.
3.3.2 Salinity
Salinity is an important environmental factor that can affect the survival and growth of fish, and also the diversity and structure of GM. Yoshimizu and Kimura (1976) discovered that the composition of salmonid GM changed in salmonids during seawater transfer. They observed that Monospora and Enterobacteriaceae were predominant in freshwater, while halophilic Vibrio was the predominant genus in seawater. More recently, Rudi et al. (2018) documented that the freshwater-to-seawater transition had a major influence on Atlantic salmon GM composition and quantity. Indeed, there was a 100-fold increase in bacterial quantity in seawater along with a relative increase of Firmicutes and a relative decrease of both Actinobacteria and Proteobacteria. Additionally, freshwater GM had a lower alpha diversity than saltwater GM. Despite the shift, the researchers also identified some unaffected gut microorganisms, namely three Clostridia and one lactobacillus-affiliated phylotype that were highly prevalent and co-occurring. This confirms suggestions that there is a stable core of microorganisms apart from the alterable population. Notably, the increased diversity observed in saltwater for Atlantic salmon is not necessarily conserved across salmonoid species. Zhao et al (2020a) conducted a trial in which chinook salmon either spent their whole lives in freshwater or were reared in freshwater facilities before being transferred to seawater. In doing so, researchers documented that species richness and diversity were significantly higher in freshwater farmed salmon than in those farmed in a marine environment. Oppositely to the anadromous fish which migrate from freshwater to saltwater for growth and development, catadromous fish migrate from saltwater to freshwater. In a recent study in Asian sea bass (Lates calcarifer), this transfer was established to affect both mucosa- and digesta-associated microbiota (Morshed et al., 2023). Plesiomonas and Cetobacterium were dominant in both the mucosa- and digesta-associated microbiota of freshwater-acclimated fish while pathogenic Vibrio, Staphylococcus, and Acinetobacter genera were dominant in the saltwater group.
It is notable that salinity appears to be among the most decisive factors for the diversity and composition of the GM: Indeed, in chinook salmon (Zhao et al., 2020a), salinity was reportedly more important than water temperature with regards to the effect on GM, and in Atlantic salmon the shift from freshwater to saltwater significantly impacted the GM while diet did not show significant associations with the microbiota (Rudi et al., 2018). Despite this documented significance, it is important to keep in mind that in both anadromous and catadromous fish the shift in habitat from fresh- to saltwater occurs in parallel with the maturation of the fish and concomitant physiological changes which allow them to acclimate to the new environmental salinity. These processes can have a separate, but simultaneous impact on the GM, which can make it very difficult to separate the changes caused directly by altered salinity. To address salinity as an isolated factor it can therefore be useful to study the impact on fish who either have not started or completed their maturation. In line with this, several recent studies have been conducted. Liu et al (2023) exposed grass carp to three different salinities (0, 2 and 6 ppt) for 8 weeks. They established that increased salinity reduced the proportion of beneficial bacteria such as Fusobacteria and Firmicutes, while the proportion of Actinobacteria, Verrucomicrobia and Acidobacteria increased. Similarly, shifts in salinity also affected the dominant intestinal microbial taxa in juvenile largemouth bass (Sun et al., 2023), and in striped catfish juveniles (Pangasianodon hypophthalmus) the Vibrio genera increased while the Akkermansia genera decreased with increasing salinity (Hieu et al., 2022). Additionally, genus Sulfurospirillium was predominantly in fish exposed to salinity treatments while alpha diversity measures generally showed a decreasing pattern as the salinities increased.
3.3.3 Pollution, inorganic salts, heavy metals and microplastics
The pollution of costal, river and lake environments are of great concern as it can have adverse effects on the health of aquatic animals. A plethora of studies have been performed describing how pollution affects both animals in general, but also the GM in particular. To study the effects of accidental oil spills, Bagi et al (2018) exposed Atlantic cod (Gadus morhua) to increasing levels of crude oil for 28 days and established that the diversity of the GM was significantly affected. Specifically, the abundance of Deferribacterales increased, while the abundance of Fusobacteriales and Alteromonadales decreased at the highest exposure level. The pollutants can be administered through several pathways. In saltwater, fish will drink seawater with any pollutants precent, and in both seawater and freshwater prey may be contaminated. The intestine is especially vulnerable as crude oil metabolites are known to be excreted into the intestinal tract via bile.
Alongside accidental oil spills, heavy metal contamination has become a severe environmental concern. Given the growing evidence that heavy metal exposure may cause various metabolic diseases partly due to heavy metal induced dysbiosis of the gut microbiota (Duan et al., 2020), the impact of various metals on the GM of fish has become a growing field of research. A 2020 study by Yan and colleagues showed that exposure to a cocktail of cadmium, lead and zinc exposure significantly influenced the diversity of the GM and intestinal function of marine medaka (Oryzias melastigma). The effects were gender dependent with the heavy metals inducing greater alterations in males. The authors interpreted this as the intestinal microbiota of adult male marine medaka being more sensitive to the heavy metals compared to the GM of the female fish, which coincides with their findings that more metabolic pathways were affected in male fish than in female. In their discussion the researchers stated that exposure to heavy metals increased abundances of Firmicutes, Proteobacteria and Bacteroidetes in females, and speculate that the relative protection seen in the female fish may be due to the reported link between the Firmicutes such as Lachnoclostridium-10, Ruminococcaceae, and Lactobacillus and SCFAs, which can enhance intestinal endocrine function, prevent infection, and protect intestinal health (Lemaire et al., 2018). These heavy metals can also influence the microbiota separately. Indeed, lead has been shown to cause gut microbiota dysbiosis and hepatic metabolic disorder in zebrafish (Xia et al., 2018a), and a 2020 study established that cadmium exposure altered the intestinal microbiota in freshwater crayfish (Procambarus clarkia) with regards to richness, diversity and composition, while also causing histological changes in the intestines (Zhang et al., 2020b). Importantly, a functional prediction analysis of intestinal microbial communities showed that Cd exposure could significantly alter the pathways related to metabolism, diseases, cellular processes, and more, thus highlighting the functional importance of such microbiome studies.
Copper is a heavy metal of particular concern since it is used as an antifouling agent in both boat paint and fish nets. Though copper is an indispensable trace element for fish, exposure to increased concentrations of can have a severe effect on the health of the fish (reviews by Liao et al., 2023a; Malhotra et al., 2020). Moreover, given its function as an antimicrobial agent and the repeated demonstration that copper exposure can cause growth impairment in several fish species (reviewed by Tavares-Dias, 2021), the effect of this heavy metal on the GM is highly relevant. Meng et al. (2018) exposed common carp to a high levels of copper (0.28 mg Cu/L), and documented that the α-and β-diversity of GM were significantly changed, and reduced the abundances of several putative SCFA-producing bacteria, including Allobaculum, Blautia, Coprococcus, Faecalibacterium, Roseburia, and Ruminococcus. The significance of SCFAs will be discussed in section 4.1, but it is notable that the authors related this to lipid metabolism and immunity. Exposing tiger puffer to 0, 50, 100 or 500 μg/L copper for three days, Liu et al (2024) noticed that increased copper concentration cause significant damage to the intestinal tissue and that the exposure also altered the structure of the intestinal microbiota. At 100 and 500 μg/L the colonization of the bacterial gut was inhibited and the intestinal barrier disrupted. This would have made the fish susceptible to infections by pathogens. The exposure also impacted the production of bacterial metabolites such as L-histidine, arachidonic acid, and L-glutamic acid, notably related to energy metabolism and immunity. The effect of copper on the GM has also been demonstrated in Nile tilapia, where exposure to subacute levels (0,5 mg/L, 1 mg/L and 2 mg/L Cu2+) for 14 days resulted in reduced bacterial diversity, a decreased abundance of Ruminococcus and Shigella and increased abundance of Edwardsiella and Aquabacterium (Zhang et al., 2022a).
In recent years the toxic properties of microplastics (MPs) have gained attention, largely due to their ubiquitous presence in marine environments. They are easily ingested by fish and have been shown to accumulate in several organs. For example, polyethylene microbeads have been shown to accumulate including eyes and kidneys of medaka (Chisada et al., 2021), while polystyrene microplastics accumulated in the gut, gills, liver and brain of Nile tilapia (Ding et al., 2018). The accumulation in the gut is of particular relevance to the present review, particularly given that ingestion of MPs has been demonstrated to cause mechanical damage to the intestine of gilthead sea beam (Varó et al., 2021), as well as impact the diversity and structure of intestinal microbiota in several species, including zebrafish (Huang et al., 2021), Nile tilapia (Zhang et al., 2022a), and marine medaka (Yan et al., 2020). Importantly, the dysbiosis of the GM caused by MPs as well as heavy metals like cadmium, copper, lead and zinc has different or exaggerated when combined. This because MPs have been proposed to function both as a toxic substance and as a transport carrier for heavy metals (Kutralam-Muniasamy et al., 2021). Indeed, Yan et al (2020), exposed marine medaka to either MPs alone (100 µg/L about 1 × 103 particles/mL 2.5 μm fluorescent polystyrene), heavy metals (Cd 10 μg/L, Pb 50 μg/L, and Zn 100 μg/L) or MPs and heavy metals combined (MPs 100 μg/L, Cd 10 μg/L, Pb 50 μg/L, and Zn 100 μg/L) for a month and concluded that, while all treatments changed the composition of the GM, the impact was highly dependent on the exposure-group as well as gender. In fact, while MPs treatment reduced the diversity and abundance of intestinal microbiota, the heavy metal and combined treatments increased them. Additionally, the combined treatment caused a higher pollution load on the gut and triggered significant changes of as Burkholderiales, Betaproteobacteria, and Corynebacteria.
Another common pollutant in water ecosystems is ammonia. The major source of this pollutant is agriculture, sewage effluents and industrial waste (Randall and Tsui, 2002) along with residual feed and excretion from aquaculture. The toxic effect of ammonia on overall fish health and various organs is well documented (Wicks et al., 2002; Benli et al., 2008; Tudorache et al., 2008; McKenzie et al., 2009; Schram et al., 2010; Sinha et al., 2014), but given the increased appreciation of the GM as an important contributor to the well-being of fish, several recent studies have also included data on the impact of ammonia on the GM. A 2017 study by Qi and colleagues showed that chronic ammonia exposure (30 days) altered the composition of intestinal microbiota of crucian carp (Carassius auratus), while Yan et al (2021) established that ammonia exposure changed the abundance and evenness of intestinal microbes in hybrid grouper (♀ Epinephelus fuscoguttatus × ♂ Epinephelus lanceolatu), with increased relative abundance of Fusobacteria and a significant decrease the relative abundance of Gemmatimonadetes. This change occurred in parallel with significant up-regulation of antioxidant, inflammatory cytokines, and apoptosis genes. A chronic (28 days) ammonia exposure in yellow catfish (Pelteobagrus fulvidraco) established that the abundance of Flavobacteriaceae, Vibrio, Pseudomonas, Aeromonas and Streptococcus increased in exposed groups showing increased intestinal cell necrosis, lamina propria edema and reduced width of intestinal villi (Luo et al., 2024). In juvenile yellow catfish, Wang et al (2021a) documented that acute ammonia exposure (96h) resulted in increased ammonia accumulation in both the intestine and muscle, concomitantly with an altered amino acid composition and enzymatic activity in the muscle. Additionally, the exposure induced a reduction in diversity, richness as well as evenness and structure of the GM. At the phylum level, the abundance of Fusobacteria increased following ammonia exposure. This led the authors to speculate that Fusobacteria increased as a protection mechanism against ammonia toxicity in the intestine, although the mechanism remains unclear. Furthermore, the reduction of Firmicutes and Actinobacteria suggested to the researchers that the high concentrations of ammonia can interfere with nutrient absorption and disrupt intestinal barrier functions which could impact the host’s defense capacity against pathogen invasion. At the genus level, the population level of the beneficial bacteria Cetobacterium increased following ammonia exposure, which is similar to findings in crucian carp (Qi et al., 2017). Considering this, Wang and colleagues speculated that Cetobacterium might represent a positive factor in ammonia toxicity resistance, related to ammonia detoxification (Wang et al., 2021b). It is also notable that the two above mentioned studies in yellow catfish (Wang et al., 2021b; Luo et al., 2024) showed different effects on the GM. By comparing the two, it becomes exceedingly clear that differing concentrations, durations and fish sizes/maturities has a significant impact on how ammonia influences the GM and should be taken into account when considering studies on this topic.
Importantly, while the effects of ammonia exposure on the GM can be caused both by direct exposure, it can also stem from the acidification resulting from hydration to ammonium. One example of this comes from a study performed in the Amazon fish tambaqui (Colossoma macropomum), in which low water pH (pH 4.0) significantly reduced the abundance of gut Flavobacterium while the ratio of Firmicutes and Bacteroides were significantly increased (Sylvain et al., 2016). Notably, the authors concluded that despite these differences, the GM largely adapted to the shift in pH. Importantly, the GM was determined to be more resilient to the lowered pH than the skin microbiota, likely because the GM are naturally exposed to acidic pH from gastric sections, and the researchers speculated that the dysbiosis they observed resulted from host physiological stress rather than directly from acute acidic exposure. Notably, this may be the case for the effect seen from several of the environmental factors.
3.4 Stress
Stress is a topic separate from but related to the topic of environmental factors. Stress is a series of neuroendocrine and physical responses to a stressor that aims to optimize the animal’s ability to cope with the threat. Some argue for the necessity for the stressor to be uncontrollable and/or unpredictable (Koolhaas et al., 2011), while others have a wider perspective. The stress response is often termed a “fight-or-flight” response as one of its main goals is to provide energy to regain homeostasis or regain control. The response starts as sensing and interpretation of the stress signal by the fish’s equivalent to the limbic system such as the dorsolateral and dorsomedial pallium in the telencephalon (Wendelaar Bonga, 1997; Schreck et al., 2016; Vindas et al., 2017). This initiates a sympathetic adrenergic response both via direct innervation into tissues like the heart (often noradrenaline) and indirectly via neural activation of chromaffin cells in the head kidney to secrete adrenaline into circulation. This adrenergic-driven activation increases the capacity of energy generation and consumption and includes increased heart rate, improved gill and muscle perfusion, intestinal motility, and often reduced intestinal blood flow. To be able to maintain a high energy output, energy stores are mobilized including degradation of glycogen. Sometime after stress, typically 5-10 minutes, the hypothalamus-pituitary-interrenal axis will have activated interrenal cells to produce cortisol that will stimulate the restoration of energy supplies, and in the long term will help reduce energy costs by downregulating non-essential processes such as immune responses and growth.
Physiologically, stress will also affect the intestine through various pathways. The severity of response seems to vary with species and intestinal sections. Nutritional status when exposed to stress also seems to be important for the severity of response, often with non-fed fish mounting a more violent response than fed fish in a postprandial state (Olsen et al., 2005, 2008). A typical response to stress in mammals is the increased secretion of fluid, digestive enzymes and mucus, that combined with increased peristalsis will transport the digesta in distal direction (Gustafsson and Johansson, 2022). The barrier function also tends to be impaired often through increased paracellular permeability, and the gut microbiota will be altered (Leigh et al., 2023). This appears to be the general findings in many fish species. In salmonids, acute stress has been shown to transport intestinal mucin towards the anus (Olsen et al., 2005, Olsen et al., 2008), exhaust eicosanoid cascades (Olsen et al., 2012) and rupture of junctional complexes eventually leading to impaired barrier functions that may last for several days after exposure (Olsen et al., 2002, Olsen et al., 2005). As mentioned above, the significant species-dependent response makes it difficult to generalize responses. For example, the acute stress response in Atlantic cod (Gadus morhua) is generally less severe than in salmonids where impacts on intestinal integrity is less noticeable with lower magnitude of cortisol and glucose changes and lower increase in clinical markers for tissue damage (Olsen et al., 2008). Many of these responses to acute stress will also have a direct impact on gut microbiota (Ringø et al., 2014; Lv et al., 2021; Zhou et al., 2022). The effects are probably most important for the autochthonous microbiota which is largely embedded in the mucin layer (see section 2). Here they function in concert with antimicrobial peptides, immunoglobulins, proteins etc. to regulate microbial composition (Reverter et al., 2018). The layer will also prevent colonization on enterocyte surfaces. When mucus is detached following stress, it will mix with fecal matter and be transported out of the fish. The lack of protective mucin and antimicrobial factors will therefore lead to increased frequency of bacteria penetrating into microvillous surface of enterocytes (Olsen et al., 2002; Ringø et al., 2014) Furthermore, cellular damage and increased paracellular permeability will increase the likelihood of microbial penetration into the intestinal tissue generating inflammation and other damage. Such changes are likely to challenge the symbiotic relationship between host and microbiota. This can alter microbial fermentation patterns and the gut-brain communication axis. To what extent these changes become permanent or will return to pre-stress levels is not well documented in fish. However, there is some data suggesting that early stress in Atlantic salmon will cause lasting effects on the intestinal microbiome (Uren Webster et al., 2021).
In many cases, stress is repetitive, or the environmental changes are long lasting or permanent. This will lead to an allostatic adaptation by the fish, where setpoints are changed to minimize the load under the new conditions (Korte et al., 2007). Within limits, the fish can cope with these changes, but the new environmental factors will favor part of the microbiome causing major changes in the microbial composition. This is often seen with changes in environmental factors like temperature, salinity (Lai et al., 2020; Liu et al., 2022b). At very high load, regulatory mechanisms are exhausted resulting in allostatic overload or chronic stress. In fish, chronic stress has been reported to reduce nutrient utilization (Ringø and Olsen, 1999; Yang et al., 2022), and to induce several morphological features (varies with tissue segments) including mucosal folding, transepithelial resistance and several molecular pathways involved in tissue repair and inflammation (Ringø et al., 2014; Løvmo et al., 2020; Løvmo et al., 2022) Chronic stress also tends to alter many immune functions such as inhibiting lymphocyte activation and immunoglobulin secretion (Tort et al., 2022). All these factors will, as far as we know, affect microbiome composition and function, and in some cases open to the establishment of pathogen agents. In sturgeon hybrids (Acipenser baeri x Acipenser schrenckii), heat stress led to inhibition of digestion and increased abundance of potential pathogen bacteria (Yang et al., 2022). In addition to the physiological effects of stress on the intestine and the microbiome, cortisol will also be translocated into the intestinal lumen and come in direct contact with the microbiome. This also opens for a direct effect of cortisol on the microbiome. One example is the study of (Duran-Pinedo et al., 2018) who showed that cortisol increased the metabolic profiles of the oral microbiome, and some cultured bacteria, towards bacteria (eg genus Fusobacteria) that are involved in human diseases and the development of periodontal disease. In juvenile Atlantic salmon, Uren Webster et al (2020) suggested that the elevated fecal cortisol excretion by chronically stressed fish caused the observed reduction in prevalence of “good” LAB and increase in potential pathogens. In a previous shrimp study, Xiong et al. (2017) stated, “that stress inhibits the natural microbiota’s growth, thereby opening the door for pathogenic bacteria to multiply, thus fostering an overgrowth and disease”. In a later study, Li et al. (2018) put forward a hypothesis that reducing Pacific white shrimp (Litopenaeus vannamei) weight may be due to increased prevalence of pathogens and decreased population levels of lactobacilli, with a result; a loss of weight. Shrimp aquaculture and factors affecting the GM are reviewed in El-Saadony et al. (2022).
3.4.1 Pathogen associated stress
Bacterial disease happens when pathogen’s environment becomes suitable, which usually occurs because of an unhealthy diet or external stress. Pathogen infection is an important type of stress to aquatic organisms, and several studies, both ex vivo and in vivo, have shown that infection will affect gut morphology (e.g., Ringø et al., 2006, Ringø et al., 2007; Salinas et al., 2008) and change the GM. In a feeding experiment using Arctic charr fed soybean-, linseed- and marine oils on aerobic gut microbiota before and after challenge with Aeromonas salmonicida ssp. salmonicida, Ringø et al. (2002) revealed decrease in total viable counts of adherent bacteria in hindgut and modulation of the microbiota composition, after challenge. However, as molecular methods were not used in this study, one can question if the results present a correct picture. Aeromonas hydrophila infection has shown to gradually reduce the ratio of Firmicutes and Proteobacteria in the GM of grass carp, followed by increased proportion of Fusobacteria (Zhou et al., 2020a; Sun et al., 2022). The most common bacterial infections affecting fish include streptococcosis, vibriosis, furunculosis and mycobacteriosis (Irshath et al., 2023). In a study with European chub (Squalius cephalus) infected by the intestinal parasite Pomphorhynchus sp. revealed significantly less GM diversity than the uninfected individuals (Colin et al., 2022). In addition, the relative abundances of the main bacterial phyla, including the Firmicutes, Fusobacteria, Actinobacteria and Proteobacteria were substantial changed, confirming its pivotal role in gut microbial assemblage. Recently, Chang et al. (2023) showed the diversity and richness of the intestinal GM of common carp was significantly changed by A. hydrophila infection with an increase in the relative abundance of Vibrio, Bacteroides and Cetobacterium, in infected fish. Surprisingly, Aeromonas was not detected in neither of the three groups, resistant-, susceptible- or control fish. One interesting finding was a significant increase in the relative abundance of Lactococcus, Akkermansia and Vibrio in resistant fish vs. susceptible and control fish. Nocardiosis, an infectious disease caused by Nocardia cause high mortalities in fish culture, but the effects of Nocardia infections on fish intestinal microbiota is less investigated. In a recent study using tilapia, Medina-Felix et al. (2024) revealed that Nocardia infection caused high mortality and severe changes, loss of diversity and increase of Proteobacteria, in the stomach microbiota, but lesser effect was observed in intestine. Based on the statement by Minniti et al. (2017) that “in aquaculture, fish seem to be more susceptible to pathogens compared to wild fish”, pathogen-associated stress merits further investigations, but to avoid overlaps, readers with interest in the mechanism-driven dysbiosis by pathogenic bacteria and further information of dysbiosis by pathogens in fish are recommend having a closer look at the review paper by Medina-Felix et al. (2023).
4 Communication pathways between the microbiota and the host
Thus far, factors that impact the microbiota have been discussed. Importantly, the impact of these changes on the fish is mediated through the pathways by which the microbiota communicates with the host and vice versa (Figure 1). Indeed, the GM communicates with host cells and the host organism via a variety of metabolites with a wide range of bioactivities. Mammalian studies have shown that these metabolites include SCFAs, secondary bile acids (BAs), vitamins, gases, tryptophan and indole derivatives, and neurotransmitters such as serotonin, dopamine, and gamma-aminobutyric acid (GABA) (reviewed by Liu et al., 2022a). Studies also show that the GMs involvement in regulating metabolite production is largely conserved across vertebrates including fish (Mountfort et al., 2002; Tsuchiya et al., 2008; Borrelli et al., 2016; Xiong et al., 2022). Some metabolites, such as SCFAs, are produced by the GM directly from diets, while others, such as secondary BAs, are generated by the host and then modified by the GM. Given that signaling by these metabolites constitute the major communication pathways between the GM and the host, this section aims to provide a curated overview of the most relevant metabolites, how their production is regulated and how they interact with the host on a molecular and cellular level.
4.1 Short chained fatty acids
The ability of certain members of the GM, such as Faecalibacterium, Bifidobacteria and Lactobacilli (LeBlanc et al., 2017) to produce SCFAs is reported to be key to the interaction between the microbiota and the host, and according to Liu et al. (2022a) the SCFAs have therefore become “the cynosure of all the gut microbiota metabolites”. SCFAs are fatty acids (FFAs) with fewer than 6 carbon atoms. In both mammals and fish, acetate, propionate and butyrate have been demonstrated to be the main metabolites produced by bacterial fermentation of dietary fibers and starch in the colon (Clements et al., 1994; Zhang and Davies, 2016; Hao et al., 2017b) and the highest concentration of SCFAs in fish intestine is in the posterior region. Indeed, in herring cale (Odax cyanomelas), 85% of the total SCFAs were detected in this region (Clements et al., 1994), while a study in silver drummer (Kyphosus sydneyanus) determined that the concentrations of acetate, propionate and butyrate were 1.1, 0.0 and 0.0 mM in the stomach, 37.4, 13.9 and 2.1 mM in the proximal section and 37.55, 12.8 and 1.3 mM in the distal gut, respectively (Mountfort et al., 2002). The same study also established similar trends for butterfish (Odax pollus) and marblefish (Aplodactylus arctidens). Acetate is the most abundant SCFA in many teleosts, but the acetate: propionate: butyrate ratio is species dependent. One study demonstrated that the gut commensal microbiota in adult zebra fish are capable of synthesizing acetate, propionate and butyrate at a ratio of 90:5:5 (Cholan et al., 2020). In Atlantic salmon the ratio was 66:20:14 in fish fed a marine diet (Nimalan et al., 2022), and in grass carp it was 70:15:15 in fish fed a control FM diet, though both amount and ratio was shown to change upon alteration of feed, resulting from the altered GM that followed (Hao et al., 2017a). One study looking into the SCFA ratio in the posterior region of the intestine determined it to be 83:8:9 in herring cale and 74:17:9 in sea carp (Crinodus lophodont) (Clements et al., 1994). Notably, the same study determined the ratio in butterfish to be 64:21:14, a finding which is incongruous with the findings by Mountfort et al. (2002), who concluded the ratio to be 78:18:4 (mM levels were reportedly 20,8, 4.7 and 1.0 for acetate, propionate and butyrate, respectively) in a presumably comparable region of the intestine. This difference may be owed to variations in the diets between the experiments, or by factors such as slight variations in the gut region examined, microbiota composition, site of fermentation, host genotype, host age and various environmental factors. These variables are known to influence the amount of SCFAs and their relative proportion, and the difference in findings highlights the need to better our understanding of the mechanisms by which this is regulated.
Following their production, SCFAs can function as energy sources, regulators of gene expression and as signaling molecules recognized by specific receptors. However, the three main SCFAs differ significantly in their potential effect on host physiology. In mammas, butyrate is for example utilized as the primary energy source for colonocytes (Donohoe et al., 2011; Rivière et al., 2016), and can also orchestrate the genes encoding tight-junction proteins and regulate the redistribution of occludin to prevent abnormal intestinal permeability (Wang et al., 2012). Acetate and propionate reach the liver via the portal vein. Propionate is metabolized by hepatocytes and has been shown to increase insulin sensitivity and attenuate the lipogenic pathway, thus lowering liver triglyceride levels (Cheng and Lai, 2000; Chambers et al., 2015; Weitkunat et al., 2016). Research has also shown that propionate can be used for synthesis of glucose in the liver (den Besten et al., 2013). Acetate is reported in high concentrations in systemic circulation and functions as a versatile metabolite in a myriad of cellular pathways, including, but not limited to, mitochondrial energy metabolism, reactive oxygen species (ROS) metabolism, lipogenesis and histone-and lysine acetylation (Bose et al., 2019). The vastly different physiological functions of the SCFAs highlight the importance of studying the rate, ratio and extent of SCFA production, which results from a complex interplay between many variables, including diet, gut transit time, activity and microbiota composition (Brinkworth et al., 2009; Peng et al., 2013; Tottey et al., 2017; Bishehsari et al., 2018). Blautia, for example, is known to be an acetogen producers (Park et al., 2012), while Faecalibacterium and Roseburia are butyrate producers (Duncan et al., 2002). A full review of the functions of the gut microbiota metabolites was conducted by Liu et al. (2022a). Importantly, though much of the status quo is taken from mammalian studies, there are indications that important functions and impacts of the SCFAs are conserved in fish. A study by Zhang et al. (2020a) showed that antibacterial activity and oxygen consumption of head kidney macrophages from turbot (Scophthalmus maximus L.) increased upon incubation with sodium butyrate, sodium propionate or sodium acetate. Additionally, Liao and colleagues (Liao et al., 2023b) showed that increased acetate production by the GM in zebrafish led to enhanced immunity though upregulated expression of IL-22 and antimicrobial peptides. Nevertheless, functional characterizations of the various SCFAs in fish is a knowledge gap that should be addressed.
4.1.1 SCFA signaling
Another important aspect of SCFA signaling is the pathways through which they exert their function. In mammals, SCFAs have been demonstrated to exert their physiological functions either via the activation of G-protein coupled receptors (GPCRs) or inhibition of histone deacetylases (HDACs) (Parada Venegas et al., 2019). Although there is limited research on the SCFA signaling pathways in fish, the ubiquity of the pathways makes it likely that mechanisms and receptors are largely conserved. This section will therefore briefly summarize what is known from mammalian data before presenting the limited status quo from research in fish.
The best studied SCFA receptors in mammals are free fatty acid receptor 2 (FFAR2 – GPR43) and free fatty acid receptor 3 (FFAR3 – GPR41) (Brown et al., 2003). Studies have shown that both FFAR2 and FFAR3 are highly expressed in the intestines (Karaki et al., 2006; Dass et al., 2007; Tazoe et al., 2009), specifically in enteroendocrine cells, specialized cells of the GIT which release GI hormones that binds to local cellular and neuronal receptors and also circulate with the blood-stream producing a myriad of metabolic and immunologic effects. There are several types of enteroendocrine cells, including I cells, which secrete cholecystokinin (CCK), K cells, which secrete gastric inhibitory peptide (GIP), L cells, which secrete glucagon-like peptide 1 and 2 (GLP-1 and GLP-2) and peptide YY (PYY), and enterochromaffin cells that secrete serotonin. FFAR2 has also been located to gastric ghrelin cells (Engelstoft et al., 2013), and both FFAR2 and FFAR3 have been established to be expressed on gastric brush cells (Eberle et al., 2014). Both FFAR2 and FFAR3 signal via GPCR pathway, though while FFAR2 is reported to be capable of duel signaling via both Gαq and Gαi pathways, FFAR3 signals exclusively via Gαi/o.
In addition to FFAR2 and FFAR3, two other receptors are recognized as SCFA receptors in mammals, namely G-protein receptor 109a (GRP109a – also known as hydrocarboxylic acid receptor 2 – HCAR2) and olfactory receptor 78 (OLDR78). Neither have been extensively studied, but most is known about GRP109a. This receptor was first identified as a receptor for nicotinic acid (niacin) (Benyó et al., 2005), but has later been further characterized as a receptor for fatty acids as well. It is unique from FFAR2 and FFAR3 in its ligand selectivity, as it is activated by the longer SCFA, in such as butyrate, pen-tanoate, hexonoate and beta-hydroxybutyrate (Taggart et al., 2005; Offermanns, 2017). GRP109a is expressed in macrophages, other immune cells, adipocytes, as well as pancreatic β cells (Benyó et al., 2005; Wang et al., 2016), and activation of the receptor has been shown to have an anti-inflammatory and anti-carcinogenic effect in the intestine (Thangaraju et al., 2009; Smith et al., 2013; Singh et al., 2014).
As previously stated, much less is known in fish and functional characterizations of receptors associated with the recognition of SCFAs have not yet been widely published. In zebra fish, Cholan et al. (2020) identified a gene termed hcar1 which shared 43% identity with human hcar1, hcar2 and hcar3 as well as mouse hcar1 and hcar2 though genes directly homologous to both GPR109a (HCAR2) and OLFR78 remain unidentified in fish. Additionally, one study identified and validated ten free fatty acid receptor genes, termed gpr40L, in common carp that showed a tight relationship with mammalian SCFA receptors, both with regards to protein phylogeny and genetic synteny (Petit and Wiegertjes, 2022). Via phylogenetic analysis the authors determined that these ten genes showed a division into three subclasses names a, b and c, and they showed a closer relationship with FFAR2 than FFAR3. The authors also performed in silico investigations for genomes of 25 fish species, mostly teleosts, and largely confirmed the observations from the common carp study regarding division into subgroups and conservation of synteny and concluded that all teleosts gpr40L genes appear closest to the human GPR43 and that the teleost grp40L family originated from a single ancestral gene multiplied by tandem duplication and later divergence. The study of Petit and Wiegertjes (2022) provided indications that there may be a FFAR-based SCFA signal-relaying system in teleosts but did not provide any insight into tissue distribution of gene expression or effects of any relevant variables (such as feed intake, feed ingredients, stress, pro-, pre- or antibiotics etc.) on said expression.
Interestingly, in a study by Wentzel et al. (2020), the authors reported that macrophages from common carp can polarize into M1- and M2 phenotypes when subjected to immune stimuli (IPS and cAMP, respectively) and that the conserved functions and corresponding transcriptional profiles were comparable to that observed in mammalian macrophages. According to Petit and Wiegertjes (2022), several of the genes identified as grp40L in common carp were among the genes affected by the stimuli. This indicates that these genes play a role in immunomodulatory responses, thus strengthening the assumption that they share a functional relationship with the human FFAR2.
Aside from signaling via GPCR, SCFAs can also exert regulation by inhibiting the function of histone deacetylases. Acetylation of lysine residues within histones relaxes chromatin structure, facilitates access of transcription factors to promoter regions and increases gene transcription. Histone deacetylases (HDACs) remove acetyl groups from histones, tightening chromatin structure and preventing transcription. Several mammalian studies have shown that SCFAs can function as inhibitors of HDAC, thus promoting increased gene expression (Hinnebusch et al., 2002; Kiefer et al., 2006; Waldecker et al., 2008; Lin et al., 2015). Of the SCFAs, butyrate is the most potent inhibitor, while acetate did not inhibit HDACs. The inhibitory effects may also be tissue dependent as orally administered acetate increased acetylation and inhibit the activity and expression of HDAC2 in rodent brain (Soliman and Rosenberger, 2011). In fish, the previously addressed study by Zhang et al. (2020a) demonstrated that butyrate increased both oxygen consumption and bactericidal activity and that these effects were due to HDAC inhibition by butyrate leading to enhanced production of antibacterial effectors and increased bacterial killing of macrophages.
4.2 Bile acids
Bile salts, conjugates between bile acids (Bas) and an amino acid are the main component of bile in many fishes. In mammals, BAs are significantly affected by the gut microbiota and will therefore affect lipid absorption, as well as in intestinal and liver health. Given that several reviews have recently been written on the topics of microbial production of bile acids and mechanisms of their physiological functions (Liu et al., 2022a; Collins et al., 2023; Fogelson et al., 2023; Ridlon and Gaskins, 2024), this section will focus exclusively on relevant research that has been conducted in fish.
Interestingly, Rawls et al (2004) showed that genes involved in cholesterol metabolism and trafficking as well as in bile acid biosynthesis could be modulated by the microbiota in both mice and zebrafish. Indeed, the genome of zebrafish possesses orthologs of many mammalian genes known to be involved in bile salt homeostasis, including the FXR (named NR1H4 in fish) (Wen et al., 2021), and a 2022 study by Xiong and colleagues demonstrated that adding Citrobacter freundii GC01 isolated from grass carp intestine to the feed of mice and zebrafish resulted in altered bile acid profiles, while also affecting lipid metabolism and upregulating expression of a key gene (CYP7A1) in BA synthesis. From this the authors concluded that the impact of specific intestinal microbes on BA metabolism is conserved between higher and lower vertebrates. Notably, bile acids also impact microbiota composition in fish. For example, a trial in which a blend of bile salts was added to the diet of gilthead seabeam over the course of 90 days resulted in a modulation of the microbiota with differential effects on the anterior and posterior intestine. Specifically, the relative abundance of Desulfobacterota doubled in the anterior region, while the levels of Firmicutes increased and Proteobacteria, Actinobacteriota, and Campylobacterota were reduced in the posterior region when supplementing the diet with bile salts (Ruiz et al., 2023).
4.3 Vitamin synthesis by the microbiota
From mammalian studies it is known that the GM can synthesize certain vitamins, specifically vitamin K and B group vitamins such as biotin, cobalamin, folates, nicotinic acid, pantothenic acid, pyridoxine, riboflavin and thiamine (Hill, 1997). The vitamins synthesized by the GM are important for bacterial metabolism, but there is also evidence that vitamins will enter the organismal vitamin metabolism pathways. In an early study, Frick et al. (1967) showed that humans subjected to low vitamin K diets for 3-4 weeks did not develop vitamin deficiency, while humans treated with broad spectrum antibiotics to suppress the microbiota showed a significant decrease in plasma prothrombin levels. Later studies have used modern metagenomic sequencing to investigate pathways for vitamin synthesis by the GM. Magnúsdóttir et al. (2015) used systematic genomic assessment to investigate the presence of biosynthetic pathways for eight B vitamins, namely biotin, cobalamin, folate, nicotinic acid, pantothenate, pyridoxine, riboflavin, and thiamin, in 256 common human gut bacteria. In doing so, the authors were able to predict the proportion of each phylum to produce each vitamin. The most synthesized vitamins were riboflavin and nicotinic acid, with 166 and 162 identified potential producers, respectively. For riboflavin and biotin almost all microbes belonging to the Bacteroidota, Fusobacteriota and Proteobacteria phyla were predicted to possess the required pathways, while a much smaller portion of Firmicutes and Actinobacteria were predicted to have to potential for vitamin B biosynthesis. Upon comparing their predictions with experimental data from 16 organisms they documented that 88% of their predictions agreed with published data, providing credence to their approach. Interestingly, Magnúsdóttir and co-authors reported that partial vitamin biosynthesis from some microorganisms were completed by the biosynthetic pathways of others. This suggests that microorganisms exchange B-vitamin or B-vitamin metabolites among each other, thus enabling the survival of organisms that do not synthesize some or any of these essential cofactors. This underlines the co-evolution of the gut microbes. It also means that not all microbially produced vitamin B will be available to the host, as it is consumed by non-vitamin producing bacteria.
In fish, an important study by Sugita et al. (1991), investigated the vitamin B12-producing ability of the intestinal microbiota in Japanese eel, carp, goldfish, ayu, tilapia and channel catfish. In doing so they established that Bacteroides type A predominated in carp and tilapia, fish species that do not require dietary vitamin B12, as well as in goldfish and ayu. However, Bacteroides type A was not detected in the eel and channel catfish, fish which require dietary vitamin B12. The authors also reported that there is a close relationship between the amounts of vitamin B12 and viable counts of Bacteroides type A in the intestinal contents of carp, strongly indicating that it is involved in vitamin B12 production in the intestinal tract of freshwater fish. There are however no concrete studies on the relative contribution of the GM to the total fish vitamin synthesis. However, owing to established knowledge regarding the genera of bacteria responsible for producing various vitamins, studies in fish may provide some indications. For example, Cetobacterium, a known vitamin B12 producing genus has been documented to be was the most abundant genus in the intestinal microbiota of Giant Amazonian fish (Arapamia gigas) (Ramírez et al., 2018), European seabass, hybrid striped bass (Morone chrysops × Morone saxatilis), red drum (Sciaenops ocellatus), common carp, hybrid tilapia (Oreochromis niloticus x Oreochromis aureus) (Ofek et al., 2021), channel catfish, largemouth bass and bluegill (Lepomis macrochirus) (Larsen et al., 2014) and several cichlid species (Baldo et al., 2015). Cetobacterium was also reported to be a highly abundant genus in farmed adult Nile tilapia (Wu et al., 2021) and Atlantic salmon (Gajardo et al., 2017). Given that Cetobacterium isolated from the intestine of several freshwater fish has been demonstrated to be capable of producing vitamin B12 (Tsuchiya et al., 2008), the role of the GM in the vitamin homeostasis of fish species is a scientific field poised for further development. This is underscored by a recent study which established that an increase in acetate producing Cetobacterium somerae contributed to glucose homeostasis and improved carbohydrate utilization in zebrafish (Wang et al., 2021a). Though there is some data on the production of vitamins from the GM of fish, it is a research topic ripe for further development. In a commercial perspective is it of great interest to understand the role of the gut microbiota in vitamin homeostasis. For Atlantic salmon it is of particular interest because there have been significant variations in the vitamin B12 content in salmon the last 15 years (Moxness Reksten et al., 2022), which influences the commercial value of the fish, as well as growth and other physiological parameters.
4.4 Neurotransmitters
During the process of evolution, the microbes which inhabit the gut have established a symbiotic relationship with their host. The GM contribute to digests the hosts dietary components to meet its own nutritional need while simultaneously providing energy and nutrients for the host. Additionally, the GM also produces some neuroactive metabolites such as neurotransmitters or their precursors. In mammals this has been shown to affect the total concentrations of these neurotransmitters or their precursors in the brain (Gao et al., 2018), suggesting that the neurotransmitter synthesis pathway in the brain may directly or indirectly affect neuronal activity of cognitive function of the brain. The metabolites produced by the mammalian GM include the neurotransmitters glutamate, γ-aminobutyric acid (GABA), serotonin and dopamine (Strandwitz, 2018). Additionally, some bacterial metabolites can act as signaling molecules which induce the synthesis and release of neurotransmitters by enteroendocrine cells (Yano et al., 2015).
Importantly, neurotransmitters like GABA, dopamine and serotonin do not penetrate the blood-brain barrier, and as such they must be synthesized in the brain from local pools of neurotransmitter precursors. Most of these precursors are amino acids like tyrosine and tryptophan derived from diet, which are converted to functional neurotransmitters through a series of intermediate steps by various host enzymes. In this, the protein/amino acid metabolism taking place in the intestinal microbiota is a key regulator. Amino acids present in food can meet entirely different fates depending on the physiological conditions of the body and the microorganisms present in the intestines. They can be absorbed intact, or as part of di- or tripeptides (Rojas-García et al., 2016; Vacca et al., 2023), or directly incorporated into the bacterial cells providing nutrition for the intestinal flora, they can be metabolized into biogenic amines by a decarboxylation reaction, they can be metabolized through a series of reactions like fission, deamination, decarboxylation, oxidation and reduction to produce structurally related indoles and phenols (Davila et al., 2013).
5 Effects of microbiota on the host
One major focus field of research is the potential influence GM has on the host. It is often presented as established knowledge that bacteria present in the human body outnumber human cells by a ratio of 10:1 (Bäckhed et al., 2005; Gill et al., 2006; Turnbaugh et al., 2007; Hsiao et al., 2008). This remarkable ratio often serves as a compelling entry point into the field as it highlights the major role microorganisms likely play in the host physiology. While more recent studies have debunked the 10:1 ratio in favor of a much more muted ratio of 1:1 in humans (Sender et al., 2016), the explosion of research into the role of the gut microbiota has also transcended into other species. The attention paid to this “new organ” has shed some much-needed light on the important and varied roles of the intestinal bacteria. In this section we aim to highlight and discuss the knowledge of teleost GM on appetite and immune responses of the hosts.
5.1 Appetite control via the microbiota-gut-brain axis
The possible modulation of appetite and energy homeostasis by GM has been the focus of much research over the last decade (Wessels, 2022). In all vertebrates, including fish, appetite, feeding behavior and feed intake is controlled by the brain (Rønnestad et al., 2017; Soengas et al., 2018; Volkoff and Rønnestad, 2020). The key appetite controlling centers are localized in the hypothalamus where there are neural circuits that either stimulate (orexigenic) or inhibit (anorexigenic) feeding. These hypothalamic neurons release neuropeptides that acts on higher order integrating neurons that modulate appetite and the motivation to eat at any given time (Wee et al., 2019). A range of external environmental signals (e.g., presence of feed via visual and olfactory cues) as well as internal signals (e.g., stress, social interactions, circulating levels of nutrients and metabolites) will modulate this motivation (Delgado et al., 2017). The appetite controlling center(s) continuously receive peripheral inputs from the body that affects these neurons, and the GIT provides some of the key signals that results in hunger that tend to initiate a meal, or satiety that tend to terminate a meal, or satiation that tend to motivate the animal to not ingest a new meal for some time (the period between meals). The importance and range of signaling pathways affecting appetite originating in the GIT, and thus the gut-brain axis, have been the subject of studies for a long time (Wessels, 2022). However, how the GM may affect these signaling pathways has only recently been given attention. The signals originating in the GIT that reach the brain acts via the afferent n. vagus and sympathetic nerves, or via circulation after crossing the blood–brain barrier (Figure 2). In fish the main center for appetite control seems to be in the nucleus lateralis tuberis (NLT) area of the hypothalamus (Norland et al., 2023), an analogue of the arcuate nucleus in mammals. In the NLT there are neurons that express cocaine- and amphetamine-regulated transcript (cart) and proopiomelanocortin (pomc) neuropeptides. As in mammals the pomc peptide undergoes posttranslational modifications and one of its products, α-MSH, together with CART sems to inhibit appetite. Other neurons express and release neuropeptide Y (NPY) and agouti-related peptide (AgRP) that in some species seems to stimulate appetite. Together, these NLT neuronal populations comprise the key neuropeptide expression of the melanocortin system (MCS). The MCS is relatively well-conserved among vertebrates, including fish. However, several rounds of whole-genome duplication in teleosts species (Glasauer and Neuhauss, 2014) have resulted in the presence of several paralogous genes, some of which may have resulted in divergent functions. For instance, in Atlantic salmon, a salmonid-specific fourth whole-genome duplication has led to the key potential neuropeptides of the MCS repertoire being compromised of three npy, two agrp, ten cart and three pomc genes (Kalananthan et al., 2020, Kalananthan et al., 2021; Tolås et al., 2021).
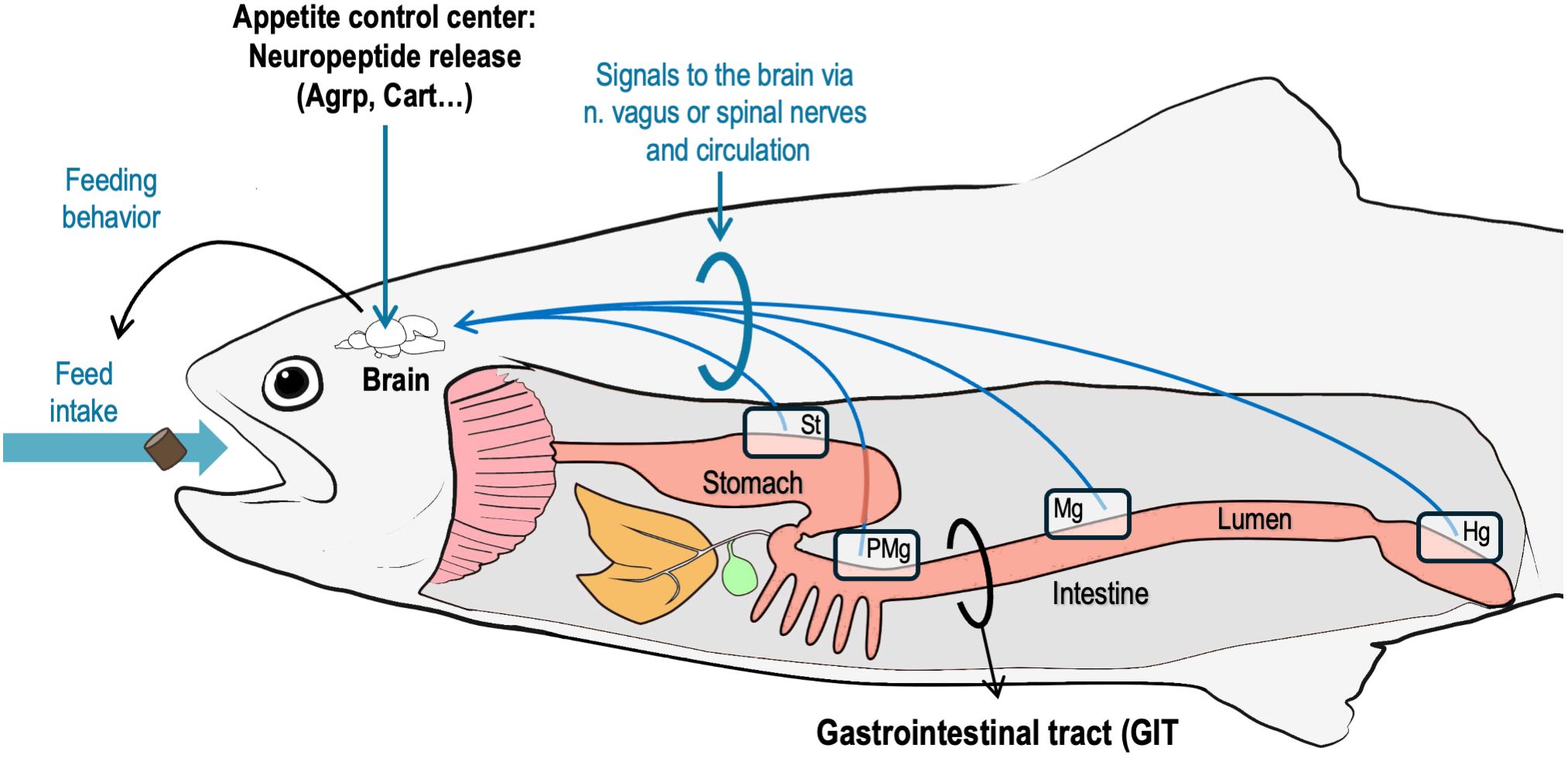
Figure 2. Possible relationship between feeding behavior and GM-derived bioactive metabolites, gut signaling pathways, and brain regulation of appetite. The GM and its metabolites are believed to influence neuropeptide secretion in the GIT (See Figure 1). The specific gut peptides released from the enteroendocrine cells varies across different GIT regions (St: stomach- ghrelin; Intestine- Cck, Pyy, Glp1). The extrinsic innervation of the gut is mainly vagal in the proximal part [St, proximal midgut (PMg)], while the midgut (Mg) and hindgut (Hg) are innervated by spinal nerves (Nilsson, 2012). The signals are transmitted to the appetite control center in the brain via the nerves or circulatory system. Glp1 may also act via modulation of insulin secretion in the endocrine pancreas. Neuropeptides (e.g., Agrp, Npy, Cart, Pomc) in the brain modulate feeding behavior and ultimately feed intake. The proposed pathways are mainly derived from mammalian studies and remain to be fully elucidated in fish. (Based on Rønnestad et al., 2017; Butt and Volkoff, 2019; Blanco et al., 2021; Pereira et al., 2024).
In mammals the MSC neurons have been shown to express receptors for several of the hormones released in the GIT and thus receive the circulating signals from the GIT. In addition, there are also important input from the GIT conveyed through the NTS in the brainstem and through the PVN area. Not all these pathways are equally well described in fish as in mammals. However, the need for a close integration of digestion and feed intake to ensure sufficient supply of nutrients and energy to an animal requires an intimate communication that includes control and feedback in the gut brain axis, suggest that these systems may be conserved in animals.
There is a broad range of afferent signals from the digestive tract that transmit the status of the filling of the digestive tract and the status of the digestive process. The signals originate in stretch and chemical receptors that sense GI-luminal content (qualitative and quantitative) and are transmitted via release of hormones from enteroendocrine cells (EEC) or through nerves, initially the enteric nervous system (ENS). There is a wide range of hormones released by EEC, and the most recently studied include ghrelin (GHRL), cholecystokinin (CCK), glucagon-like peptide-1 (GLP-1), and peptide YY (PYY) (Steinert et al., 2017). Moreover, insulin released from endocrine pancreas has a strong impact on reducing appetite after a meal. Additionally leptin that in mammals also is expressed in the GIT in addition to i.e. adipose cells may also contribute to modulate appetite (Obradovic et al., 2021) also seems to affect appetite in fish (Chisada et al., 2014; Mankiewicz et al., 2022) although the mechanisms may be different. The signaling pathways in the GIT includes the complex enteric nervous system (ENS) that consist of two neural networks located in the submucosa layer and has pacemaker cells and controls i.e., motility via release of neurotransmitters based on the amino acid tryptophan associated neurotransmitter serotonin (5-HT: Serotonin, 5-hydroxytryptamine), dopamine and GABA.
Prior to the relatively recent description of the microbiota as a potent modulator of a range of physiological and psychological factors, even controversially termed as an “extra organ” (as discussed by Riccio and Rossano, 2020), the GIT bacteria were largely ignored, even in studies of digestion. Most of the classical roles described to bacteria were related to production of some vitamins and factors affecting absorption (e.g., LeBlanc et al., 2013), effects related processing of fiber related to motility and water balance, and pathogens that affects homeostasis. Over the last two decades there has been studies that has changed in the view of the role of the luminal microbiota to also include significant impacts on host, via interactions with the GIT that may even be bidirectional. There are a range of possible mechanisms described that enables the microbiome to interact with known signaling pathways that affect the host appetite control, but also with novel pathways where specific microbial metabolites can stimulate release of appetite regulating hormones and neurotransmitters. These pathways include: 1) Direct activation of n. vagus or transmission of neurotransmitters (e.g. GABA, 5-HT) from the GM that stimulate the ENS; 2) Production or induction of metabolites that pass the intestinal barrier, enter the circulation and eventually pass the blood–brain barrier to interfere with neurological functions and 3) Microbial-associated molecular patterns (MAMPs, e.g., LPS) and metabolites produced by the microbiota that may signal to the immune system (Asano et al., 2012; Zhang and Davies, 2016; Butt and Volkoff, 2019; Wessels, 2022). If, and to what extent GM in fish may influence appetite and energy homeostasis through these and possibly also novel mechanisms still largely remains to be elucidated (Blanco et al., 2021; Volkoff, 2024; Niu et al., 2024)
Regarding 1) The short-chain fatty acids (SCFAs) are believed to be the major mediators that link the microbiome and CNS in animals. They have been shown to stimulate secretion of peptide hormones from the enteroendocrine cells such PYY and GLP-1, amino acid tryptophan with the associated neurotransmitter serotonin (5-HT: serotonin, 5-hydroxytryptamine), dopamine and GABA (Yano et al., 2015; Zhang and Davies, 2016). Regarding 2) There are numerous commensal and pathogenic bacteria that synthetize short peptide fragments that are strikingly similar to fragments of leptin, ghrelin, PYY and NPY (Fetissov et al., 2008; Alcock et al., 2014) and catecholamines (Asano et al., 2012) that might potentially affect the central regulation of appetite by triggering the respective neurons. This will reportedly affect the release of central appetite regulating neuropeptides like neuropeptide Y (NPY) and proopiomelanocortin (POMC) (Breton et al., 2016), thus making nutrient sensing of SCFAs central in the regulation of appetite and energy homeostasis (Mayer et al., 2015; Read and Holmes, 2017; Bliss and Whiteside, 2018).
The effects of gut microbiome on metabolism and appetite control in mammals continues to be discussed. Based on some initial high-profile findings in gnotobiotic mice (Bäckhed et al., 2004, Bäckhed et al., 2007) which was also reported in the popular press (e.g., Kolata, 2013) it was suggested that the GIT microbiome serve an important role in controlling feed intake, energy homeostasis and weight. Following these results there have been a huge interest to document and understand the underlying mechanisms. On whole animals, there have i.e., been several trials conducted where the GIT microbiome from lean mice have been transferred to obese mice with the aim to reduce feed intake and weight. This method is termed fecal microbiota transplant and has also been, and continues to be, tested in human intervention studies. In a critical review of trials published over the past 15 years (Dalby, 2023) failed to document consistent evidence for the role of the microbiome, as an independent factor, in regulating feed intake, adiposity and body weight in mice, rats and humans. In fact, some of the earlier studies are criticized for selective reporting of data and there is also a lack of long-term data (Dalby, 2023). Also, in the studies published over the last years most reported that they failed to show coherent differences between treatments raising questions about the previously proposed relationship between GM and obesity (Fleissner et al., 2010; Dalby et al., 2017; Yu et al., 2020; Moretti et al., 2021).
While there are several studies in mammals that reports link the GM with cognition, mental health and a range of physiological and pathological states (Tooley, 2020; Shoubridge et al., 2022), most of these studies have been done in an area where the overall work targets to understand obesity and related health issues related to metabolic syndrome and diabetes, thus focusing on identifying mechanisms that will allow a reduction of appetite and/or in combination in alterations in metabolism that reduce body fat. As such, the mechanisms described in the communication between GIT microbiota and the host involve satiety and energy expenditure inducing pathways (Fetissov, 2017). The functional significance of these appetite related signals is to induce satiety at the end of a meal, and thus terminate eating, or to prevent ingesting a new meal while there is still significant content in the GIT. As previously described of the neural and hormonals signals originating in the GIT it is only Ghrelin that has a described orexigenic role. Studies in mammals show that ghrelin also stimulates growth hormone (GH) secretion from the pituitary (hence the name, growth hormone-releasing peptide), thereby increasing adiposity and reducing energy expenditure and that that ghrelin level is linked to stomach filling and the anticipation of a meal (partly as a conditional response) and meal initiation (Howick et al., 2017). In teleost fish the role of ghrelin on appetite is still not clarified, and there are studies that indicate both orexigenic and anorexigenic effects (Jönsson, 2013; Zhong et al., 2021), whether this is due to species differences or methodological challenges remains to be determined.
The potential effect of the gut microbiome on metabolism and appetite has been examined in a number of fish species (Butt and Volkoff, 2019; Niu et al., 2024). In grass carp, many biosynthetic and metabolic pathways of carbohydrates, amino acids and lipids change as the composition of microbiota changes (Ni et al., 2014). In zebrafish, the colonization of the gut by microorganisms promotes epithelial absorption of fatty acids (Semova et al., 2012) and fish with intact microbiota have increased lipid accumulation in the intestinal epithelium, and increased expression of genes related to lipid metabolism compared to germ-free fish who lack microbiota (Sheng et al., 2018). In addition, Olive flounder fed a diet supplemented with Bacillus clausii display higher weight gain, feed efficiency and growth performance compared to fish fed control diets (Ye et al., 2011). The authors of the same publication suggest this could be attributed to increased food intake and improved nutrient digestibility. All this data suggests a strong influence of the microbiota in fish metabolism.
Still, similarly to mammals, owing to studies reporting diverging results, the influence of microbiota composition on food intake is still debated. For example, zebrafish larvae treated with Lactobacillus rhamnosus affected the GM, transcriptome and several properties of the intestinal epithelium and also led to elevated growth (Falcinelli et al., 2015). In a later study by the same group they demonstrated treatment caused reduced appetite and feed intake compared to control fish (Falcinelli et al., 2016). In this study Glp-1 (glucagon), leptin and insulin mRNA (all presumably anorexigenic) increased while the orexigenic npy, goat, cb1 decreased. A study that used the same probiotic in an encapsulated form showed significant opposite effects in juvenile Nile tilapia. In this case the fish administered the probiotic grew much better, and there was increased expression of npy, agrp and ghrelin, while there was a reduction in leptin that together supported an orexigenic drive (Gioacchini et al., 2018). A recent study demonstrates that exposure to the cyanobacterium Microcystis aeruginosa significantly alters appetite regulation in stone moroko by increasing appetite-inhibiting factors (cart, glp-1) and reducing stimulatory factors (agrp, orexin), linked to changes in lipid, amino acid, and cholesterol metabolism (Niu et al., 2024). In contrast, the macrophyte Ottelia acuminata mitigates these adverse effects, suggesting potential protective interactions via altered regulation of intestinal metabolites. Although appetite was not studied a modulation of microbiota affected anxiety in larval zebrafish larvae, and Lactobacillus plantarum (a probiotic) reduced anxiety-related behavior in conventionally-raised zebrafish larvae (Davis et al., 2016). The link between stress and appetite is well known, and reduced appetite is often the first sign of stress in fish. Taken together this shows that although there are demonstrated signaling pathways in the communication between GM and the host and that this may include appetite control, the overall effect on whole animals is not fully understood.
However, carp fed a diet supplemented with fructo-oligosaccharide (FOS) display increased levels of heterotrophic aerobic bacteria and LAB, but no changes in feeding rates compared to fish fed a control diet (Hoseinifar et al., 2014). However, it is important to keep in mind that these studies are vastly different, using different model organisms and different additives. They are therefore difficult to compare.
With regards to understanding how and to what extent the GM affects appetite via the described mechanisms above it is important to note that there are dynamic changes in the communication in the gut brain axis that correlates with the status of the digestive process and the gut transit of food and digesta. At the same time, the GM assemblies are specific per anatomical site and are highly dynamic during food digestion, indicating that digestive status and/or sampling time are factors potentially influencing the GM compositions. This was demonstrated in southern catfish (Silurus meridionalis) a carnivorous species characterized by rapid growth that typically ingest large meals and where there were large spatial and temporal variation in GM community after a meal (Zhang et al., 2017). The spatial variation of microbial diversity stomach was larger than in the gut. This suggests that the signals originating in the GM that affect appetite and energy metabolism in the host may change significantly during a meal. In conclusion, there is a need for more targeted and specialized studies investigating specific signaling pathways involved in the GM-gut-brain axis, comparable to those conducted in mammalian models
5.2 Immune system
Along with nutrient metabolism, and energy homeostasis, the GM has been generally recognized as a significant component of the intestinal mucosal immune system including regulation of innate and adaptive immunity (Figure 1). On one hand, GM can stimulate Th1 cells, regulatory T and B cells to produce Interleukin-10 (IL-10) to maintain the homeostasis of the mucosal immune system (Sun et al., 2018; Maldonado Galdeano et al., 2019; Mishima et al., 2019). GM can also regulate the host’s health by maintaining balance and inhibiting the growth of potentially pathogenic bacteria (Maldonado Galdeano et al., 2019). Furthermore, oral probiotics can induce intestinal epithelial cells to produce macrophage chemotaxis protein 1 through Toll-like receptors (TLR), and then activate the mucosal immune system (Maldonado Galdeano et al., 2019; Liu et al., 2020). Upon intestinal barrier damage, LPS, one of the most immune stimulatory components derived from the cytoderm of Gram-negative bacteria, enter the host circulation and is recognized by TLR on immune cells initiating inflammatory responses (Hug et al., 2018; Witkowski et al., 2020).
In recent years, abundant autochthonous bacteria have been isolated for immunological studies of aquatic species. Tan et al. (2019) reported that R. stabekisii used as probiotic improved innate immunity against Aeromonas hydrophila and Streptococcus iniae infections in tilapia. The mechanism appeared in part to be due to modulation f the intestinal microbiota by increasing the abundance of Bacillus sp. and Lactobacillus sp. and reducing the abundance of Staphylococcus sp. and Streptococcus sp (Abarike et al., 2018). Niu et al. (2019) isolated six probiotics (Bacillus licheniformis, Bacillus amyloliquefaciens, Bacillus subtilis, Lactobacillus brevis, L. plantarum and Saccharomyces cerevisiae) from the intestinal tract of juvenile olive founder and established that administration of multi-strain probiotics significantly improved the non-specific and specific immune responses of juvenile olive flounder by increasing the activities of myeloperoxidase (MPO), lysozyme, glutathione peroxidase (GPX) and the expression of proinflammatory genes IL-1β, IL-6 and TNF-α. Probiotic supplements (Lactobacillus rhamnosus JCM1136 and Lactococcus lactis subsp. lactis JCM5805) added together or apart not only significantly increased the length and density of intestinal microvilli, but also increased the colonization of probiotics in the intestine of Nile tilapia, which enhanced the immune response and disease resistance (Xia et al., 2018b). These observations strongly suggest that GM is a vital part of the intestinal immune system, and the structure and metabolites of GM are important in maintaining intestinal health. Indeed, the impact of the GM on host immunity is an expanding and promising research field, recently reviewed more extensively by Zhang et al. (2024) and Morshed and Lee (2023).
6 Perspectives
Though the knowledge regarding gut microbiota, metabolism and immunity has increased in fish and shrimp in recent years, several issues still merit further investigations. Indeed, there is a plethora of publications documenting the alteration in microbiota profiles upon altered conditions, including diet, dietary additives and environmental factors, but few studies explore the underlying mechanisms of how such changes impact the host. When it comes to the way in which the microbiota communicates with the host, several knowledge gaps have been described throughout the text (See also Figure 1). The take-home message is that, while knowledge from mammalian studies provide a firm starting point from which to explore these pathways, studies focused on fish species is needed to further our understanding. One example of this not brought up within the review is the topic of mechanisms or ligand-specificity for the free fatty acid receptors responsible for relaying SCFA-signaling. Crucially, there are significant species variations with regards to SCFA potency in activating FFAR2 and FFAR3 in mammals. In humans the rank order of potency for FFAR2 has been reported to be C2 = C3 > C4 > C5 = C1 whereas for FFAR3 it is C3 = C4 = C5 > C2 > C1 (Milligan et al., 2009). C2 is 20-fold more selective at human FFAR2 than at human FFAR3, while it is equipotent at both mouse orthologs. On the other hand, C3 is 12-fold more selective at mouse FFAR3, but exhibit similar potency at human orthologs (Schmidt et al., 2011). One would assume that there may be similar variation in ligand specificity between paralogs in teleosts as well as between teleost species. Experimental testing of ligand potency and specificity is therefore a highly relevant field of investigation when exploring these receptors in fish species.
In sum, the content presented in this review demonstrates that there have been significant leaps in our understanding of the GM, the factors influencing its composition and diversity, and its role in influencing host health through regulating metabolism and immunity. Still, gaps of knowledge regarding the mechanistic processes underlying its functions invites further scrutiny.
Author contributions
IT: Conceptualization, Funding acquisition, Project administration, Writing – original draft, Writing – review & editing. ZGZ: Writing – original draft, Writing – review & editing. ZZ: Writing – original draft. TT: Writing – original draft. RO: Writing – original draft, Writing – review & editing. ER: Writing – original draft, Writing – review & editing. IR: Conceptualization, Funding acquisition, Project administration, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by funding from the Research Council of Norway (325849).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abarike E. D., Cai J., Lu Y., Yu H., Chen L., Jian J., et al. (2018). Effects of a commercial probiotic BS containing Bacillus subtilis and Bacillus licheniformis on growth, immune response and disease resistance in Nile tilapia, Oreochromis niloticus. Fish Shellfish Immunol. 82, 229–238. doi: 10.1016/j.fsi.2018.08.037
Akrami R., Iri Y., Khoshbavar Rostami H., Razeghi Mansour M. (2013). Effect of dietary supplementation of fructooligosaccharide (FOS) on growth performance, survival, lactobacillus bacterial population and hemato-immunological parameters of stellate sturgeon (Acipenser stellatus) juvenile. Fish Shellfish Immunol. 35, 1235–1239. doi: 10.1016/j.fsi.2013.07.039
Alcock J., Maley C. C., Aktipis C. A. (2014). Is eating behavior manipulated by the gastrointestinal microbiota? Evolutionary pressures and potential mechanisms. Bioessays 36, 940–949. doi: 10.1002/bies.201400071
Alfiko Y., Xie D., Astuti R. T., Wong J., Wang L. (2022). Insects as a feed ingredient for fish culture: Status and trends. Aquac. Fish. 7, 166–178. doi: 10.1016/j.aaf.2021.10.004
Al-Harbi A. H., Naim M. (2004). Seasonal variation in the intestinal bacterial flora of hybrid tilapia (Oreochromis niloticus×Oreochromis aureus) cultured in earthen ponds in Saudi Arabia. Aquaculture 229, 37–44. doi: 10.1016/S0044-8486(03)00388-0
Alishahi M., Dezfuly T., Mohammadian T., Mesbah M. (2018). Effects of two probiotics, lactobacillus plantarum and lactobacillus bulgaricus on growth performance and intestinal lactic acid bacteria of cyprinus carpio. Iran. J. Vet. Med. 12, 207–218. doi: 10.22059/ijvm.2018.235444.1004816
Allameh S. K., Noaman V., boroumand jazi M., Ganjour M., Nahavandi R. (2021). Some beneficial effects of probiotics in aquaculture. J. Clin. Trials Res. 4, 215–218. Available online at: https://www.scitcentral.com/documents/86c35d5771e064c6d91650706bedcd60.pd.
Almeida A. R., Alves M., Domingues I., Henriques I. (2019). The impact of antibiotic exposure in water and zebrafish gut microbiomes: A 16S rRNA gene-based metagenomic analysis. Ecotoxicol. Environ. Saf. 186, 109771. doi: 10.1016/j.ecoenv.2019.109771
Antonopoulou E., Kolygas M., Panteli N., Gouva E., Kontogeorgiou P., Feidantsis K., et al. (2023). Breeding Substrate Containing Distillation Residues of Mediterranean Medicinal Aromatic Plants Modulates the Effects of Tenebrio molitor as Fishmeal Substitute on Blood Signal Transduction and WBC Activation of Gilthead Seabream (Sparus aurata). Anim. 13, 2537. doi: 10.3390/ani13152537
Asano Y., Hiramoto T., Nishino R., Aiba Y., Kimura T., Yoshihara K., et al. (2012). Critical role of gut microbiota in the production of biologically active, free catecholamines in the gut lumen of mice. Am. J. Physiol. Gastrointest. Liver Physiol. 303, G1288–G1295. doi: 10.1152/ajpgi.00341.2012
Bäckhed F., Ding H., Wang T., Hooper L. V., Gou Y. K., Nagy A., et al. (2004). The gut microbiota as an environmental factor that regulates fat storage. Proc. Natl. Acad. Sci. U. S. A. 101, 15718–15723. doi: 10.1073/pnas.0407076101
Bäckhed F., Ley R. E., Sonnenburg J. L., Peterson D. A., Gordon J. I. (2005). Host-bacterial mutualism in the human intestine. Science 307, 1915–1920. doi: 10.1126/science.1104816
Bäckhed F., Manchester J. K., Semenkovich C. F., Gordon J. I. (2007). Mechanisms underlying the resistance to diet-induced obesity in germ-free mice. Proc. Natl. Acad. Sci. U. S. A. 104, 979–984. doi: 10.1073/pnas.0605374104
Bagi A., Riiser E. S., Molland H. S., Star B., Haverkamp T. H. A., Sydnes M. O., et al. (2018). Gastrointestinal microbial community changes in Atlantic cod (Gadus morhua) exposed to crude oil. BMC Microbiol. 18, 25. doi: 10.1186/s12866-018-1171-2
Baldo L., Riera J. L., Tooming-Klunderud A., Albà M. M., Salzburger W. (2015). Gut microbiota dynamics during dietary shift in eastern african cichlid fishes. PloS One 10, e0127462. doi: 10.1371/journal.pone.0127462
Benli A.Ç.K., Köksal G., Özkul A. (2008). Sublethal ammonia exposure of Nile tilapia (Oreochromis niloticus L.): Effects on gill, liver and kidney histology. Chemosphere 72, 1355–1358. doi: 10.1016/j.chemosphere.2008.04.037
Benyó Z., Gille A., Kero J., Csiky M., Suchánková M. C., Nüsing R. M., et al. (2005). GPR109A (PUMA-G/HM74A) mediates nicotinic acid-induced flushing. J. Clin. Invest. 115, 3634–3640. doi: 10.1172/JCI23626
Bera A. K., Chowdhury H., Ghatak S., Malick R. C., Chakraborty N., Chakraborty H. J., et al. (2023). Microbiome analysis reveals potential for modulation of gut microbiota through polysaccharide-based prebiotic feeding in Oreochromis niloticus (Linnaeu). Front. Physiol. 14. doi: 10.3389/fphys.2023.1168284
Bhute S. S., Escobedo B., Haider M., Mekonen Y., Ferrer D., Hillyard S. D., et al. (2020). The gut microbiome and its potential role in paradoxical anaerobism in pupfishes of the Mojave Desert. Anim. Microbiome 2, 20. doi: 10.1186/s42523-020-00037-5
Bishehsari F., Engen P. A., Preite N. Z., Tuncil Y. E., Naqib A., Shaikh M., et al. (2018). Dietary fiber treatment corrects the composition of gut microbiota, promotes SCFA production, and suppresses colon carcinogenesis. Genes (Basel). 9, 102. doi: 10.3390/genes9020102
Blanco A. M., Calo J., Soengas J. L. (2021). The gut-brain axis in vertebrates: implications for food intake regulation. J. Exp. Biol. 224. doi: 10.1242/jeb.231571
Blaufuss P. C., Bledsoe J. W., Gaylord T. G., Sealey W. M., Overturf K. E. (2020). Selection on a plant-based diet reveals changes in oral tolerance, microbiota and growth in rainbow trout (Oncorhynchus mykiss) when fed a high soy diet. S. Aquaculture 525, 735287. doi: 10.1016/j.aquaculture.2020.735287
Bliss E. S., Whiteside E. (2018). The gut-brain axis, the human gut microbiota and their integration in the development of obesity. Front. Physiol. 9. doi: 10.3389/fphys.2018.00900
Bojarski B., Kot B., Witeska M. (2020). Antibacterials in aquatic environment and their toxicity to fish. Pharmaceuticals 13, 1–24. doi: 10.3390/ph13080189
Borrelli L., Aceto S., Agnisola C., De Paolo S., Dipineto L., Stilling R. M., et al. (2016). Probiotic modulation of the microbiota-gut-brain axis and behaviour in zebrafish. Sci. Rep. 6, 1–9. doi: 10.1038/srep30046
Bose S., Ramesh V., Locasale J. W. (2019). Acetate metabolism in physiology, cancer, and beyond. Trends Cell Biol. 29, 695–703. doi: 10.1016/j.tcb.2019.05.005
Boulangé C. L., Neves A. L., Chilloux J., Nicholson J. K., Dumas M.-E. (2016). Impact of the gut microbiota on inflammation, obesity, and metabolic disease. Genome Med. 8, 42. doi: 10.1186/s13073-016-0303-2
Breton J., Tennoune N., Lucas N., Francois M., Legrand R., Jacquemot J., et al. (2016). Gut commensal E. coli proteins activate host satiety pathways following nutrient-induced bacterial growth. Cell Metab. 23, 324–334. doi: 10.1016/j.cmet.2015.10.017
Brinkworth G. D., Noakes M., Clifton P. M., bird A. R. (2009). Comparative effects of very low-carbohydrate, high-fat and high-carbohydrate, low-fat weight-loss diets on bowel habit and faecal short-chain fatty acids and bacterial populations. Br. J. Nutr. 101, 1493–1502 doi: 10.1017/S0007114508094658
Brown A. J., Goldsworthy S. M., Barnes A. A., Eilert M. M., Tcheang L., Daniels D., et al. (2003). The orphan G protein-coupled receptors GPR41 and GPR43 are activated by propionate and other short chain carboxylic acids. J. Biol. Chem. 278, 11312–11319. doi: 10.1074/jbc.M211609200
Bruni L., Pastorelli R., Viti C., Gasco L., Parisi G. (2018). Characterisation of the intestinal microbial communities of rainbow trout (Oncorhynchus mykiss) fed with Hermetia illucens (black soldier fly) partially defatted larva meal as partial dietary protein source. Aquaculture 487, 56–63. doi: 10.1016/j.aquaculture.2018.01.006
Butt R. L., Volkoff H. (2019). Gut microbiota and energy homeostasis in fish. Front. Endocrinol. (Lausanne). 10. doi: 10.3389/fendo.2019.00009
Caipang C. M. A., Suharman I., Avillanosa A. L., Bargoyo V. T. (2020). Host-derived probiotics for finfish aquaculture. IOP Conf. Ser. Earth Environ. Sci. 430, 12026. doi: 10.1088/1755-1315/430/1/012026
Carlson J. M., Leonard A. B., Hyde E. R., Petrosino J. F., Primm T. P. (2017). Microbiome disruption and recovery in the fish Gambusia affinis following exposure to broad-spectrum antibiotic. Infect. Drug Resist. 10, 143–154. doi: 10.2147/IDR.S129055
Cerezuela R., Fumanal M., Tapia-Paniagua S. T., Meseguer J., Moriñigo M.Á., Esteban M.Á. (2013). Changes in intestinal morphology and microbiota caused by dietary administration of inulin and Bacillus subtilis in gilthead sea bream (Sparus aurata L.) specimens. Fish Shellfish Immunol. 34, 1063–1070. doi: 10.1016/j.fsi.2013.01.015
Chambers E. S., Viardot A., Psichas A., Morrison D. J., Murphy K. G., Zac-Varghese S. E. K., et al. (2015). Effects of targeted delivery of propionate to the human colon on appetite regulation, body weight maintenance and adiposity in overweight adults. Gut. 64, 1744–1754. doi: 10.1136/gutjnl-2014-307913
Chang X., Kang M., Shen Y., Yun L., Yang G., Zhu L., et al. (2021). Bacillus coagulans SCC-19 maintains intestinal health in cadmium-exposed common carp (Cyprinus carpio L.) by strengthening the gut barriers, relieving oxidative stress and modulating the intestinal microflora. Ecotoxicol. Environ. Saf. 228, 112977. doi: 10.1016/j.ecoenv.2021.112977
Chang S., Wang J., Dong C., Jiang Y. (2023). Intestinal microbiota signatures of common carp (Cyprinus carpio) after the infection of Aeromonas hydrophila. Aquac. Rep. 30, 101585. doi: 10.1016/j.aqrep.2023.101585
Chen X., Li P., Hou D., Li M., Chen B., Peng K., et al. (2024). Probiotic breakthrough: Clostridium butyricum improved the intestinal SCFAs content, digestive enzymes and antioxidant activities, gut morphology and microbiota composition of largemouth bass. Aquac. Rep. 36, 102090. doi: 10.1016/j.aqrep.2024.102090
Chen C.-Z., Li P., Liu L., Li Z.-H. (2022). Exploring the interactions between the gut microbiome and the shifting surrounding aquatic environment in fisheries and aquaculture: A review. Environ. Res. 214, 114202. doi: 10.1016/j.envres.2022.114202
Chen J., Li Q., Tan C., Xie L., Yang X., Zhang Q., et al. (2023). Effects of enrofloxacin’s exposure on the gut microbiota of Tilapia fish (Oreochromis niloticus). Comp. Biochem. Physiol. - Part D Genomics Proteomics 46, 101077. doi: 10.1016/j.cbd.2023.101077
Cheng H. H., Lai M. H. (2000). Fermentation of resistant rice starch produces propionate reducing serum and hepatic cholesterol in rats. J. Nutr. 130, 1991–1995. doi: 10.1093/jn/130.8.1991
Chisada S., Kurokawa T., Murashita K., Rønnestad I., Taniguchi Y., Toyoda A., et al. (2014). Leptin receptor-deficient (knockout) medaka, Oryzias latipes, show chronical up-regulated levels of orexigenic neuropeptides, elevated food intake and stage specific effects on growth and fat allocation. Gen. Comp. Endocrinol. 195, 9–20. doi: 10.1016/j.ygcen.2013.10.008
Chisada S., Yoshida M., Karita K. (2021). Polyethylene microbeads are more critically toxic to the eyes and reproduction than the kidneys or growth in medaka, Oryzias latipes. Environ. Pollut. 268, 115957. doi: 10.1016/j.envpol.2020.115957
Cholan P. M., Han A., Woodie B. R., Watchon M., Kurz A. R. M., Laird A. S., et al. (2020). Conserved anti-inflammatory effects and sensing of butyrate in zebrafish. Gut Microbes. 12, e1824563. doi: 10.1080/19490976.2020.1824563
Chouayekh H., Farhat-Khemakhem A., Karray F., Boubaker I., Mhiri N., Abdallah M., et al. (2023). Effects of dietary supplementation with bacillus amyloliquefaciens US573 on intestinal morphology and gut microbiota of european sea bass. Probiotics Antimicrob. Proteins 15, 30–43. doi: 10.1007/s12602-022-09974-w
Clements K. D., Gleeson V. P., Slaytor M. (1994). Short-chain fatty acid metabolism in temperate marine herbivorous fish. J. Comp. Physiol. B. 164, 372–377. doi: 10.1007/BF00302552
Colin Y., Molbert N., Berthe T., Agostini S., Alliot F., Decencière B., et al. (2022). Dysbiosis of fish gut microbiota is associated with helminths parasitism rather than exposure to PAHs at environmentally relevant concentrations. Sci. Rep. 12, 11084. doi: 10.1038/s41598-022-15010-2
Collins S. L., Stine J. G., Bisanz J. E., Okafor C. D., Patterson A. D. (2023). Bile acids and the gut microbiota: metabolic interactions and impacts on disease. Nat. Rev. Microbiol. 21, 236–247. doi: 10.1038/s41579-022-00805-x
Cryan J. F., O’Riordan K. J., Cowan C. S. M., Sandhu K. V., Bastiaanssen T. F. S., Codagnone B. M., et al. (2019). The microbiota-gut-brain axis. Physiol. Rev. 99, 1877–2013. doi: 10.1152/physrev.00018.2018
Dai Z.-L., Wu G., Zhu W.-Y. (2011). Amino acid metabolism in intestinal bacteria: links between gut ecology and host health. Front. Biosci. (Landmark Ed. 16, 1768–1786. doi: 10.2741/3820
Dalby M. J. (2023). Questioning the foundations of the gut microbiota and obesity. Philos. Trans. R. Soc B Biol. Sci. 378. doi: 10.1098/rstb.2022.0221
Dalby M. J., Ross A. W., Walker A. W., Morgan P. J. (2017). Dietary uncoupling of gut microbiota and energy harvesting from obesity and glucose tolerance in mice. Cell Rep. 21, 1521–1533. doi: 10.1016/j.celrep.2017.10.056
Dani D. (2018). A review on replacing fish meal in aqua feeds using plant protein sources. Int. J. Fish. Aquat. 6, 164–179. Available online at: https://www.fisheriesjournal.com/archives/2018/vol6issue2/PartC/6-1-35-823.pdf.
Danner M.-C., Robertson A., Behrends V., Reiss J. (2019). Antibiotic pollution in surface fresh waters: Occurrence and effects. Sci. Total Environ. 664, 793–804. doi: 10.1016/j.scitotenv.2019.01.406
Dass N. B., John A. K., Bassil A. K., Crumbley C. W., Shehee W. R., Maurio F. P., et al. (2007). The relationship between the effects of short-chain fatty acids on intestinal motility in vitro and GPR43 receptor activation. Neurogastroenterol. Motil. 19, 66–74. doi: 10.1111/j.1365-2982.2006.00853.x
Davani-Davari D., Negahdaripour M., Karimzadeh I., Seifan M., Mohkam M., Masoumi S. J., et al. (2019). Prebiotics: Definition, types, sources, mechanisms, and clinical applications. Foods 8, 1–27. doi: 10.3390/foods8030092
Davila A.-M., Blachier F., Gotteland M., Andriamihaja M., Benetti P.-H., Sanz Y., et al. (2013). Intestinal luminal nitrogen metabolism: role of the gut microbiota and consequences for the host. Pharmacol. Res. 68, 95–107. doi: 10.1016/j.phrs.2012.11.005
Davis D. J., Bryda E. C., Gillespie C. H., Ericsson A. C. (2016). Microbial modulation of behavior and stress responses in zebrafish larvae. Behav. Brain Res. 311, 219–227. doi: 10.1016/j.bbr.2016.05.040
Dawood M. A. O., Abo-Al-Ela H. G., Hasan M. T. (2020). Modulation of transcriptomic profile in aquatic animals: Probiotics, prebiotics and synbiotics scenarios. Fish Shellfish Immunol. 97, 268–282. doi: 10.1016/j.fsi.2019.12.054
Dawood M. A. O., Koshio S. (2016). Recent advances in the role of probiotics and prebiotics in carp aquaculture: A review. Aquaculture 454, 243–251. doi: 10.1016/j.aquaculture.2015.12.033
de la Torre Canny S. G., Mueller O., Craciunescu C. V., Blumberg B., Rawls J. F. (2021). Tributyltin exposure leads to increased adiposity and reduced abundance of leptogenic bacteria in the zebrafish intestine. bioRxiv 07, 09.451869. doi: 10.1101/2021.07.09.451869
Delgado M. J., Cerdá-Reverter J. M., Soengas J. L. (2017). Hypothalamic integration of metabolic, endocrine, and circadian signals in fish: involvement in the control of food intake. Front. Neurosci. 11. doi: 10.3389/fnins.2017.00354
den Besten G., Lange K., Havinga R., van Dijk T. H., Gerding A., van Eunen K., et al. (2013). Gut-derived short-chain fatty acids are vividly assimilated into host carbohydrates and lipids. Am. J. Physiol. - Gastrointest. Liver Physiol. 305, 900–910. doi: 10.1152/ajpgi.00265.2013
Deng Y., Verdegem M. C. J., Eding E., Kokou F. (2022). Effect of rearing systems and dietary probiotic supplementation on the growth and gut microbiota of Nile tilapia (Oreochromis niloticus) larvae. Aquaculture 546, 737297. doi: 10.1016/j.aquaculture.2021.737297
Desai A. R., Links M. G., Collins S. A., Mansfield G. S., Drew M. D., Van Kessel A. G., et al. (2012). Effects of plant-based diets on the distal gut microbiome of rainbow trout (Oncorhynchus mykiss). Aquaculture, 350–353, 134–142. doi: 10.1016/j.aquaculture.2012.04.005
DeVries J. W., Camire M. E., Cho S., Craig S., Gordon D., Jones J. M., et al. (2001). The definition of dietary fiber. Cereal Foods World. 46, 112–129. Available online at: https://ww.cerealsgrains.org/resources/definitions/Documents/DietaryFiber/DFDef.pdf.
Dimitroglou A., Merrifield D. L., Moate R., Davies S. J., Spring P., Sweetman J., et al. (2009). Dietary mannan oligosaccharide supplementation modulates intestinal microbial ecology and improves gut morphology of rainbow trout, Oncorhynchus mykiss (Walbaum). J. Anim. Sci. 87, 3226–3234. doi: 10.2527/jas.2008-1428
Ding J., Zhang S., Razanajatovo R. M., Zou H., Zhu W. (2018). Accumulation, tissue distribution, and biochemical effects of polystyrene microplastics in the freshwater fish red tilapia (Oreochromis niloticus). Environ. pollut. 238, 1–9. doi: 10.1016/j.envpol.2018.03.001
Donohoe D. R., Garge N., Zhang X., Sun W., O’Connell T. M., Bunger M. K., et al. (2011). The microbiome and butyrate regulate energy metabolism and autophagy in the mammalian colon. Cell Metab. 13, 517–526. doi: 10.1016/j.cmet.2011.02.018
Druart C., Alligier M., Salazar N., Neyrinck A. M., Delzenne N. M. (2014). Modulation of the gut microbiota by nutrients with prebiotic and probiotic properties. Adv. Nutr. 5, 624S–633S. doi: 10.3945/an.114.005835
Duan H., Yu L., Tian F., Zhai Q., Fan L., Chen W. (2020). Gut microbiota: A target for heavy metal toxicity and a probiotic protective strategy. Sci. Total Environ. 742, 140429. doi: 10.1016/j.scitotenv.2020.140429
Dube E., Okuthe G. E. (2024). Applications of antimicrobial photodynamic therapy in aquaculture: effect on fish pathogenic bacteria. Fishes 9, 99. doi: 10.3390/fishes9030099
Duncan S. H., Hold G. L., Harmsen H. J. M., Stewart C. S., Flint H. J. (2002). Growth requirements and fermentation products of Fusobacterium prausnitzii, and a proposal to reclassify it as Faecalibacterium prausnitzii gen. nov., comb. nov. Int. J. Syst. Evol. Microbiol. 52, 2141–2146. doi: 10.1099/ijs.0.02241-0
Duran-Pinedo A. E., Solbiati J., Frias-Lopez J. (2018). The effect of the stress hormone cortisol on the metatranscriptome of the oral microbiome. NPJ biofilms microbiomes 4, 25. doi: 10.1038/s41522-018-0068-z
Eberle J. A. M., Widmayer P., Breer H. (2014). Receptors for short-chain fatty acids in brush cells at the “gastric groove”. Front. Physiol. 5. doi: 10.3389/fphys.2014.00152
Egerton S., Culloty S., Whooley J., Stanton C., Ross R. (2018). The gut microbiota of marine fish. P. Front. Microbiol. 9. doi: 10.3389/fmicb.2018.00873
El-Saadony M. T., Shehata A. M., Alagawany M., Abdel-Moneim A.-M. E., Selim D. A., Abdo M., et al. (2022). A review of shrimp aquaculture and factors affecting the gut microbiome. Aquac. Int. 30, 2847–2869. doi: 10.1007/s10499-022-00936-1
Engelstoft M. S., Park W., Sakata I., Kristensen L. V., Husted A. S., Osborne-Lawrence S., et al. (2013). Seven transmembrane G protein-coupled receptor repertoire of gastric ghrelin cells. Mol. Metab. 2, 376–392. doi: 10.1016/j.molmet.2013.08.006
Estruch G., Collado M. C., Peñaranda D. S., Tomás Vidal A., Jover Cerdá M., Pérez Martínez G., et al. (2015). Impact of fishmeal replacement in diets for gilthead sea bream (Sparus aurata) on the gastrointestinal microbiota determined by pyrosequencing the 16S rRNA gene. PloS One 10, 1–22. doi: 10.1371/journal.pone.0136389
Falcinelli S., Picchietti S., Rodiles A., Cossignani L., Merrifield D. L., Taddei A. R., et al. (2015). Lactobacillus rhamnosus lowers zebrafish lipid content by changing gut microbiota and host transcription of genes involved in lipid metabolism. Sci. Rep. 5, 9336. doi: 10.1038/srep09336
Falcinelli S., Rodiles A., Unniappan S., Picchietti S., Gioacchini G., Merrifield D. L., et al. (2016). Probiotic treatment reduces appetite and glucose level in the zebrafish model. Sci. Rep. 5. doi: 10.1038/srep18061
Ferdous Z., Hossain M. K., Hadiuzzaman M., Rafiquzzaman S. M., Halim K. A., Rahman T., et al. (2024). Multi-species probiotics enhance survival, growth, intestinal microbiota and disease resistance of rohu (Labeo rohita) larvae. Water Biol. Secur. 3, 100234. doi: 10.1016/j.watbs.2023.100234
Fetissov S. O. (2017). Role of the gut microbiota in host appetite control: Bacterial growth to animal feeding behaviour. Nat. Rev. Endocrinol. 24, 348–359. doi: 10.1038/nrendo.2016.150
Fetissov S. O., Hamze Sinno M., Coëffier M., Bole-Feysot C., Ducrotté P., Hökfelt T., et al. (2008). Autoantibodies against appetite-regulating peptide hormones and neuropeptides: putative modulation by gut microflora. Nutrition 24, 348–359. doi: 10.1016/j.nut.2007.12.006
Fleissner C. K., Huebel N., Abd El-Bary M. M., Loh G., Klaus S., Blaut M. (2010). Absence of intestinal microbiota does not protect mice from diet-induced obesity. Br. J. Nutr. 104, 919–929. doi: 10.1017/S0007114510001303
Fogelson K. A., Dorrestein P. C., Zarrinpar A., Knight R. (2023). The gut microbial bile acid modulation and its relevance to digestive health and diseases. Gastroenterology 164, 1069–1085. doi: 10.1053/j.gastro.2023.02.022
Fontinha F., Magalhães R., Moutinho S., Santos R., Campos P., Serra C. R., et al. (2021). Effect of dietary poultry meal and oil on growth, digestive capacity, and gut microbiota of gilthead seabream (Sparus aurata) juveniles. Aquaculture 530, 735879. doi: 10.1016/j.aquaculture.2020.735879
Frick P. G., Riedler G., Brögli H. (1967). Dose response and minimal daily requirement for vitamin K in man. J. Appl. Physiol. 23, 387–389. doi: 10.1152/jappl.1967.23.3.387
Gajardo K., Jaramillo-Torres A., Kortner T. M., Merrifield D. L., Tinsley J., Bakke A. M., et al. (2017). Alternative protein sources in the diet modulate microbiota and functionality in the distal intestine of atlantic salmon (Salmo salar). Appl. Environ. Microbiol. 83. doi: 10.1128/AEM.02615-16
Garrabou J., Gómez-Gras D., Medrano A., Cerrano C., Ponti M., Schlegel R., et al. (2022). Marine heatwaves drive recurrent mass mortalities in the Mediterranean Sea. Glob. Change Biol. 28, 5708–5725. doi: 10.1111/gcb.16301
Gao K., Pi Y., Mu C.-L., Peng Y., Huang Z., Zhu W.-Y. (2018). Antibiotics-induced modulation of large intestinal microbiota altered aromatic amino acid profile and expression of neurotransmitters in the hypothalamus of piglets. J. Neurochem. 146, 219–234. doi: 10.1111/jnc.14333
Ghori I., Tubassam M., Ahmad T., Zuberi A., Imran M. (2022). Gut microbiome modulation mediated by probiotics: Positive impact on growth and health status of Labeo rohita. Front. Physiol. 13. doi: 10.3389/fphys.2022.949559
Ghosh S. K., Wong M. K. S., Hyodo S., Goto S., Hamasaki K. (2022). Temperature modulation alters the gut and skin microbial profiles of chum salmon (Oncorhynchus keta). Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.1027621
Gill S. R., Pop M., Deboy R. T., Eckburg P. B., Turnbaugh P. J., Samuel B. S., et al. (2006). Metagenomic analysis of the human distal gut microbiome. Science 312, 1355–1359. doi: 10.1126/science.1124234
Gioacchini G., Ciani E., Pessina A., Cecchini C., Silvi S., Rodiles A., et al. (2018). Effects of lactogen 13, a new probiotic preparation, on gut microbiota and endocrine signals controlling growth and appetite of oreochromis niloticus juveniles. Microb. Ecol. 76, 1063–1074. doi: 10.1007/s00248-018-1177-1
Glasauer S. M. K., Neuhauss S. C. F. (2014). Whole-genome duplication in teleost fishes and its evolutionary consequences. Mol. Genet. Genomics 289, 1045–1060. doi: 10.1007/s00438-014-0889-2
González-Félix M. L., Gatlin D. M., Urquidez-Bejarano P., de la Reé-Rodríguez C., Duarte-Rodríguez L., Sánchez F., et al. (2018). Effects of commercial dietary prebiotic and probiotic supplements on growth, innate immune responses, and intestinal microbiota and histology of Totoaba macdonaldi. Aquaculture 491, 239–251. doi: 10.1016/j.aquaculture.2018.03.031
Guangxin G., Li K., Zhu Q., Zhao C., Li C., He Z., et al. (2022). Improvements of immune genes and intestinal microbiota composition of turbot (Scophthalmus maximus) with dietary oregano oil and probiotics. Aquaculture 547, 737442. doi: 10.1016/j.aquaculture.2021.737442
Guo H., Chen C., Yan X., Li Y., Wen X., You C., et al. (2021). Effects of different dietary oil sources on growth performance, antioxidant capacity and lipid deposition of juvenile golden pompano Trachinotus ovatus. Aquaculture 530, 735923. doi: 10.1016/j.aquaculture.2020.735923
Guo G., Li C., Xia B., Jiang S., Zhou S., Men X., et al. (2020). The efficacy of lactic acid bacteria usage in turbot Scophthalmus maximus on intestinal microbiota and expression of the immune related genes. Fish Shellfish Immunol. 100, 90–97. doi: 10.1016/j.fsi.2020.03.003
Gustafsson J. K., Johansson M. (2022). The role of goblet cells and mucus in intestinal homeostasis. E. V Nat. Rev. Gastroenterol. Hepatol. 19, 785–803. doi: 10.1038/s41575-022-00675-x
Hagi T., Tanaka D., Iwamura Y., Hoshino T. (2004). Diversity and seasonal changes in lactic acid bacteria in the intestinal tract of cultured freshwater fish. Aquaculture 234, 335–346. doi: 10.1016/j.aquaculture.2004.01.018
Hao W.-L., Lee Y.-K. (2004). Microflora of the gastrointestinal tract: a review. Methods Mol. Biol. 268, 491–502. doi: 10.1385/1-59259-766-1:491
Hao Y. T., Wu S. G., Jakovlić I., Zou H., Li W. X., Wang T. (2017a). Impacts of diet on hindgut microbiota and short-chain fatty acids in grass carp (Ctenopharyngodon idellus). Aquac. Res., 5595–5605. doi: 10.1111/are.13381
Hao Y. T., Wu S. G., Xiong F., Tran N. T., Jakovlić I., Zou H., et al. (2017b). Succession and fermentation products of grass carp (Ctenopharyngodon idellus) hindgut microbiota in response to an extreme dietary shift. Front. Microbiol. 8. doi: 10.3389/fmicb.2017.01585
Haque M. M., Hasan N. A., Eltholth M. M., Saha P., Mely S. S., Rahman T., et al. (2021). Assessing the impacts of in-feed probiotic on the growth performance and health condition of pangasius (Pangasianodon hypophthalmus) in a farm trial. Aquac. Rep. 20, 100699. doi: 10.1016/j.aqrep.2021.100699
Hartviksen M., Vecino J. L. G., Ringø E., Bakke A. M., Wadsworth S., Krogdahl Å., et al. (2014). Alternative dietary protein sources for Atlantic salmon (Salmo salar L.) effect on intestinal microbiota, intestinal and liver histology and growth. Aquac. Nutr. 20, 381–398. doi: 10.1111/anu.12087
Hasan K. N., Banerjee G. (2020). Recent studies on probiotics as beneficial mediator in aquaculture: a review. J. Basic Appl. Zool. 81, 53. doi: 10.1186/s41936-020-00190-y
He S., Wang Q., Li S., Ran C., Guo X., Zhang Z., et al. (2017). Antibiotic growth promoter olaquindox increases pathogen susceptibility in fish by inducing gut microbiota dysbiosis. Sci. China Life Sci. 60, 1260–1270. doi: 10.1007/s11427-016-9072-6
Hieu D. Q., Hang B. T. B., Lokesh J., Garigliany M.-M., Huong D. T. T., Yen D. T., et al. (2022). Salinity significantly affects intestinal microbiota and gene expression in striped catfish juveniles. Appl. Microbiol. Biotechnol. 106, 3245–3264. doi: 10.1007/s00253-022-11895-1
Hill M. J. (1997). Intestinal flora and endogenous vitamin synthesis. Eur. J. Cancer Prev. 6, S43–S45. doi: 10.1097/00008469-199703001-00009
Hinnebusch B. F., Meng S., Wu J. T., Archer S. Y., Hodin R. A. (2002). The effects of short-chain fatty acids on human colon cancer cell phenotype are associated with histone hyperacetylation. J. Nutr. 132, 1012–1017. doi: 10.1093/jn/132.5.1012
Hoseinifar S. H., Soleimani N., Ringø E. (2014). Effects of dietary fructo-oligosaccharide supplementation on the growth performance, haemato-immunological parameters, gut microbiota and stress resistance of common carp (Cyprinus carpio) fry. Br. J. Nutr. 112, 1296–1302. doi: 10.1017/S0007114514002037
Hoseinifar S. H., Van D. H., Ashouri G. (2019). Galactooligosaccharide effects as prebiotic on intestinal microbiota of different fish species. Rudn J. Agron. Anim. Ind. 14, 266–278. doi: 10.22363/2312-797x-2019-14-3-266-278
Hovda M. B., Fontanillas R., Mcgurk C., Obach A., Rosnes J. T. (2012). Seasonal variations in the intestinal microbiota of farmed Atlantic salmon (Salmo salar L.). Aquac. Res. 43, 154–159. doi: 10.1111/j.1365-2109.2011.02805.x
Howick K., Griffin B. T., Cryan J. F., Schellekens H. (2017). From belly to brain: targeting the ghrelin receptor in appetite and food intake regulation. Int. J. Mol. Sci. 18, 273. doi: 10.3390/ijms18020273
Hsiao W. W. L., Metz C., Singh D. P., Roth J. (2008). The microbes of the intestine: an introduction to their metabolic and signaling capabilities. Endocrinol. Metab. Clin. North Am. 37, 857–871. doi: 10.1016/j.ecl.2008.08.006
Hu R. G., Yang B. T., Zheng Z. Y., Liang Z. L., Kang Y. H., Cong W. (2024). Improvement of non-specific immunity, intestinal health and microbiota of crucian carp (Carassius auratus) juvenile with dietary supplementation of Bacillus coagulans BC1. Aquaculture 580, 740327. doi: 10.1016/j.aquaculture.2023.740327
Huang K.-C., Lee J.-W., Hu Y.-F., Ballantyne R., Liu C.-H. (2023). Effects of Aspergillus-meal prebiotic diet on the growth performance, health status and gut microbiota of Asian seabass, Lates calcarifer. Fish Shellfish Immunol. 136, 108696. doi: 10.1016/j.fsi.2023.108696
Huang Q., Sham R. C., Deng Y., Mao Y., Wang C., Zhang T., et al. (2020). Diversity of gut microbiomes in marine fishes is shaped by host-related factors. Mol. Ecol. 29, 5019–5034. doi: 10.1111/mec.15699
Huang W., Song B., Liang J., Niu Q., Zeng G., Shen M., et al. (2021). Microplastics and associated contaminants in the aquatic environment: A review on their ecotoxicological effects, trophic transfer, and potential impacts to human health. J. Hazard. Mater. 405, 124187. doi: 10.1016/j.jhazmat.2020.124187
Hug H., Mohajeri M. H., La Fata G. (2018). Toll-like receptors: Regulators of the immune response in the human gut. Nutrients 10, 11–13. doi: 10.3390/nu10020203
Huyben D., Rimoldi S., Ceccotti C., Montero D., Betancor M., Iannini F., et al. (2020). Effect of dietary oil from Camelina sativa on the growth performance, fillet fatty acid profile and gut microbiome of gilthead Sea bream (Sparus aurata). PeerJ 8, e10430. doi: 10.7717/peerj.10430
Huyben D., Sun L., Moccia R., Kiessling A., Dicksved J., Lundh T. (2018). Dietary live yeast and increased water temperature influence the gut microbiota of rainbow trout. J. Appl. Microbiol. 124, 1377–1392. doi: 10.1111/jam.13738
Irshath A. A., Rajan A. P., Vimal S., Prabhakaran V.-S., Ganesan R. (2023). Bacterial pathogenesis in various fish diseases: recent advances and specific challenges in vaccine development. Vaccines 11, 470. doi: 10.3390/vaccines11020470
Jang W. J., Lee J. M., Hasan M. T., Lee B.-J., Lim S. G., Kong I.-S. (2019). Effects of probiotic supplementation of a plant-based protein diet on intestinal microbial diversity, digestive enzyme activity, intestinal structure, and immunity in olive flounder (Paralichthys olivaceus). Fish Shellfish Immunol. 92, 719–727. doi: 10.1016/j.fsi.2019.06.056
Jiang Y. H., Liang M., Yang Y. H., Xue J., Suo H. Y. (2024). Probiotic Lactobacillus plantarum SHY21-2 protected zebrafish against Aeromonas hydrophila infection by maintaining intestinal barrier integrity, inhibiting inflammatory and oxidative stress responses, and regulating intestinal microbiome. Aquaculture 582, 740506. doi: 10.1016/j.aquaculture.2023.740506
Jin Y., Angell I. L., Sandve S. R., Snipen L. G., Olsen Y., Rudi K. (2019). Atlantic salmon raised with diets low in long-chain polyunsaturated n-3 fatty acids in freshwater have a Mycoplasma-dominated gut microbiota at sea. Aquac. Environ. Interact. 11, 31–39. doi: 10.3354/aei00297
Jin W., Jiang L., Hu S., Zhu A. (2024). ). Effects of Lactobacillus plantarum and Bacillus subtilis on growth, immunity and intestinal flora of largemouth bass (Micropterus salmoides). Aquaculture 583, 740581. doi: 10.1016/j.aquaculture.2024.740581
Johny T. K., Puthusseri R. M., Bhat S. G. (2021). A primer on metagenomics and next-generation sequencing in fish gut microbiome research. Aquac. Res. 52, 4574–4600. doi: 10.1111/are.15373
Jones J., DiBattista J. D., Stat M., Bunce M., Boyce M. C., Fairclough D. V., et al. (2018). The microbiome of the gastrointestinal tract of a range-shifting marine herbivorous fish. Front. Microbiol. 9. doi: 10.3389/fmicb.2018.02000
Jönsson E. (2013). The role of ghrelin in energy balance regulation in fish. Gen. Comp. Endocrinol. 187, 79–85. doi: 10.1016/j.ygcen.2013.03.013
Jung-Schroers V., Adamek M., Harris S., Syakuri H., Jung A., Irnazarow I., et al. (2018). Response of the intestinal mucosal barrier of carp (Cyprinus carpio) to a bacterial challenge by Aeromonas hydrophila intubation after feeding with β-1,3/1,6-glucan. J. Fish Dis. 41, 1077–1092. doi: 10.1111/jfd.12799
Kahyani F., Pirali-Kheirabadi E., Shafiei S., Shenavar Masouleh A. (2021). Effect of dietary supplementation of potential probiotic Weissella confusa on innate immunity, immune-related genes expression, intestinal microbiota and growth performance of rainbow trout (Oncorhynchus mykiss). Aquac. Nutr. 27, 1411–1420. doi: 10.1111/anu.13279
Kalananthan T., Gomes A. S., Lai F., Tolås I., Jordal A.-E. O., Norland S., et al. (2021). Brain Distribution of 10 cart Transcripts and Their Response to 4 Days of Fasting in Atlantic Salmon (Salmo salar L.). Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.763766
Kalananthan T., Murashita K., Rønnestad I., Ishigaki M., Takahashi K., Silva M. S., et al. (2020). Hypothalamic agrp and pomc mRNA Responses to Gastrointestinal Fullness and Fasting in Atlantic Salmon (Salmo salar, L.). Front. Physiol. 11. doi: 10.3389/fphys.2020.00061
Kapoor G., Saigal S., Elongavan A. (2017). Action and resistance mechanisms of antibiotics: A guide for clinicians. J. Anaesthesiol. Clin. Pharmacol. 33, 300–305. doi: 10.4103/joacp.JOACP_349_15
Karaki S. I., Mitsui R., Hayashi H., Kato I., Sugiya H., Iwanaga T., et al. (2006). Short-chain fatty acid receptor, GPR43, is expressed by enteroendocrine cells and mucosal mast cells in rat intestine. Cell Tissue Res. 324, 353–360 doi: 10.1007/s00441-005-0140-x
Kiefer J., Beyer-Sehlmeyer G., Pool-Zobel B. L. (2006). Mixtures of SCFA, composed according to physiologically available concentrations in the gut lumen, modulate histone acetylation in human HT29 colon cancer cells. Br. J. Nutr. 96, 803–810. doi: 10.1017/BJN20061948
Kim S., Covington A., Pamer E. G. (2017). The intestinal microbiota: Antibiotics, colonization resistance, and enteric pathogens. Immunol. Rev. 279, 90–105. doi: 10.1111/imr.12563
Kim A., Kim N., Roh H. J., Chun W.-K., Ho D. T., Lee Y., et al. (2019). Administration of antibiotics can cause dysbiosis in fish gut. Aquaculture 512, 734330. doi: 10.1016/j.aquaculture.2019.734330
Kokou F., Sasson G., Nitzan T., Doron-Faigenboim A., Harpaz S., Cnaani A., et al. (2018). Host genetic selection for cold tolerance shapes microbiome composition and modulates its response to temperature. Elife 7. doi: 10.7554/eLife.36398
Kolata G. (2013). Gut bacteria from thin humans can slim mice down. New York Times, 3–5. Available online at: https://www.nytimes.com/2013/09/06/health/gut-bacteria-from-thin-humans-can-slim-mice-down.html#:~:text=The%20investigators%20discovered%20that%20given,fat%20mouse%20then%20loses%20weight.
Kong Y., Liao Z., Ma X., Liang M., Xu H., Mai K., et al. (2023). Response of intestinal microbiota of tiger puffer (Takifugu rubripes) to the fish oil finishing strategy. Microorganisms 11, 208. doi: 10.3390/microorganisms11010208
Koolhaas J. M., Bartolomucci A., Buwalda B., de Boer S. F., Flügge G., Korte S. M., et al. (2011). Stress revisited: a critical evaluation of the stress concept. Neurosci. Biobehav. Rev. 35, 1291–1301. doi: 10.1016/j.neubiorev.2011.02.003
Korte S. M., Olivier B., Koolhaas J. M. (2007). A new animal welfare concept based on allostasis. Physiol. Behav. 92, 422–428. doi: 10.1016/j.physbeh.2006.10.018
Kuebutornye F. K. A., Wang Z., Lu Y., Abarike E. D., Sakyi M. E., Li Y., et al. (2020). Effects of three host-associated Bacillus species on mucosal immunity and gut health of Nile tilapia, Oreochromis niloticus and its resistance against Aeromonas hydrophila infection. Fish Shellfish Immunol. 97, 83–95. doi: 10.1016/j.fsi.2019.12.046
Kutralam-Muniasamy G., Pérez-Guevara F., Martínez I. E., Shruti V. C. (2021). Overview of microplastics pollution with heavy metals: Analytical methods, occurrence, transfer risks and call for standardization. J. Hazard. Mater. 415, 125755. doi: 10.1016/j.jhazmat.2021.125755
Lai K. P., Lin X., Tam N., Ho J. C. H., Wong M. K.-S., Gu J., et al. (2020). Osmotic stress induces gut microbiota community shift in fish. Environ. Microbiol. 22, 3784–3802. doi: 10.1111/1462-2920.15150
Larsen A. M., Mohammed H. H., Arias C. R. (2014). Characterization of the gut microbiota of three commercially valuable warmwater fish species. J. Appl. Microbiol. 116, 1396–1404. doi: 10.1111/jam.12475
LeBlanc J. G., Chain F., Martín R., Bermúdez-Humarán L. G., Courau S., Langella P. (2017). Beneficial effects on host energy metabolism of short-chain fatty acids and vitamins produced by commensal and probiotic bacteria. Microb. Cell Fact. 16, 1–10. doi: 10.1186/s12934-017-0691-z
LeBlanc J. G., Milani C., de Giori G. S., Sesma F., van Sinderen D., Ventura M. (2013). Bacteria as vitamin suppliers to their host: a gut microbiota perspective. Curr. Opin. Biotechnol. 24, 160–168. doi: 10.1016/j.copbio.2012.08.005
Leigh S.-J., Uhlig F., Wilmes L., Sanchez-Diaz P., Gheorghe C. E., Goodson M. S., et al. (2023). The impact of acute and chronic stress on gastrointestinal physiology and function: a microbiota–gut–brain axis perspective. J. Physiol. 601, 4491–4538. doi: 10.1113/JP281951
Lemaire M., Dou S., Cahu A., Formal M., Le Normand L., Romé V., et al. (2018). Addition of dairy lipids and probiotic Lactobacillus fermentum in infant formula programs gut microbiota and entero-insular axis in adult minipigs. Sci. Rep. 8, 1–16. doi: 10.1038/s41598-018-29971-w
Levy-Pereira N., Yasui G. S., Cardozo M. V., Neto J. D., Farias T. H. V., Sakabe R., et al. (2018). Immunostimulation and increase of intestinal lactic acid bacteria with dietary mannan-oligosaccharide in Nile tilapia juveniles. Rev. Bras. Zootec. 47. doi: 10.1590/rbz4720170006
Li J., Fang P., Yi X., Kumar V., Peng M. (2022a). Probiotics Bacillus cereus and B. subtilis reshape the intestinal microbiota of Pengze crucian carp (Carassius auratus var. Pengze) fed with high plant protein diets. Front. Nutr. 9. doi: 10.3389/fnut.2022.1027641
Li Y., Gajardo K., Jaramillo-Torres A., Kortner T. M., Krogdahl Å. (2022b). Consistent changes in the intestinal microbiota of Atlantic salmon fed insect meal diets. Anim. Microbiome 4. doi: 10.1186/s42523-021-00159-4
Li Z., Tran N. T., Ji P., Sun Z., Wen X., Li S. (2019b). Effects of prebiotic mixtures on growth performance, intestinal microbiota and immune response in juvenile chu’s croaker. Nibea coibor. Fish Shellfish Immunol. 89, 564–573. doi: 10.1016/j.fsi.2019.04.025
Li E., Xu C., Wang X., Wang S., Zhao Q., Zhang M., et al. (2018). Gut Microbiota and its Modulation for Healthy Farming of Pacific White Shrimp Litopenaeus vannamei. Rev. Fish. Sci. Aquac. 26, 381–399. doi: 10.1080/23308249.2018.1440530
Li H., Zhou Y., Ling H., Luo L., Qi D., Feng L. (2019a). The effect of dietary supplementation with Clostridium butyricum on the growth performance, immunity, intestinal microbiota and disease resistance of tilapia (Oreochromis niloticus). PloS One 14, e0223428. doi: 10.1371/journal.pone.0223428
Liao X., Lan Y., Wang W., Zhang J., Shao R., Yin Z., et al. (2023b). Vitamin D influences gut microbiota and acetate production in zebrafish (Danio rerio) to promote intestinal immunity against invading pathogens. Gut Microbes 15. doi: 10.1080/19490976.2023.2187575
Liao W., Zhu Z., Feng C., Yan Z., Hong Y., Liu D., et al. (2023a). Toxicity mechanisms and bioavailability of copper to fish based on an adverse outcome pathway analysis. J. Environ. Sci. (China) 127, 495–507. doi: 10.1016/j.jes.2022.06.002
Limbu S. M., Ma Q., Zhang M.-L., Du Z.-Y. (2019). High fat diet worsens the adverse effects of antibiotic on intestinal health in juvenile Nile tilapia (Oreochromis niloticus). Sci. Total Environ. 680, 169–180. doi: 10.1016/j.scitotenv.2019.05.067
Lin M. Y., De Zoete M. R., Van Putten J. P. M., Strijbis K. (2015). Redirection of epithelial immune responses by short-chain fatty acids through inhibition of histone deacetylases. Front. Immunol. 6. doi: 10.3389/fimmu.2015.00554
Lin S.-M., Zhou X.-M., Zhou Y.-L., Kuang W.-M., Chen Y.-J., Luo L., et al. (2020). Intestinal morphology, immunity and microbiota response to dietary fibers in largemouth bass, Micropterus salmoide. Fish Shellfish Immunol. 103, 135–142. doi: 10.1016/j.fsi.2020.04.070
Liu Y., Cheng J., Xia Y., Li X., Liu Y., Liu P.-F. (2022b). Response mechanism of gut microbiome and metabolism of European seabass (Dicentrarchus labrax) to temperature stress. Sci. Total Environ. 813, 151786. doi: 10.1016/j.scitotenv.2021.151786
Liu P., Liu Y., Cheng J., Xia Y., Yang Y. (2024). Copper exposure causes alteration in the intestinal microbiota and metabolites in Takifugu rubripes. Ecotoxicol. Environ. Saf. 272, 116064. doi: 10.1016/j.ecoenv.2024.116064
Liu J., Tan Y., Cheng H., Zhang D., Feng W., Peng C. (2022a). Functions of gut microbiota metabolites, current status and future perspectives. Aging Dis. 13, 1106–1126. doi: 10.14336/AD.2022.0104
Liu S., Wang S., Cai Y., Li E., Ren Z., Wu Y., et al. (2020). Beneficial effects of a host gut-derived probiotic, Bacillus pumilus, on the growth, non-specific immune response and disease resistance of juvenile golden pompano, Trachinotus ovatus. Aquaculture 514, 734446. doi: 10.1016/j.aquaculture.2019.734446
Liu D., Zhang Z., Song Y., Yang J., Lu Y., Lai W., et al. (2023). Effects of salinity on growth, physiology, biochemistry and gut microbiota of juvenile grass carp (Ctenopharyngodon idella). Aquat. Toxicol. 258, 106482. doi: 10.1016/j.aquatox.2023.106482
Long J. A. (2010). The rise of fishes: 500 million years of evolution. 2nd ed (Baltimore, Maryland, USA: The Johns Hopkins University Press).
Løvmo S. D., Madaro A., Whatmore P., Bardal T., Ostensen M.-A., Sandve S. R., et al. (2020). Mid and hindgut transcriptome profiling analysis of Atlantic salmon (Salmon salar) under unpredictabl chronic stress. R. Soc Open Sci. 7, 191480. doi: 10.1098/rsos.191480
Løvmo S. D., Sundh H., Whatmore P., Nordvi M. F., Sigholt T., Madaro A., et al. (2022). Intestinal health in Atlantic salmon post-smolt (Salmo salar) when fed low- and high HUFA diets. Aquaculture 557, 738318. doi: 10.1016/j.aquaculture.2022.738318
Luo L., Liu S., Chen B., Li F., Deng Y., Huang X., et al. (2024). Chronic ammonia stress caused disorder of intestinal microbiota and damaged intestinal structure and function in yellow catfish (Pelteobagrus fulvidraco). Aquaculture 581, 740428. doi: 10.1016/j.aquaculture.2023.740428
Lv H., Liu Y., Li H., Yin X., Wang P., Qu X., et al. (2021). Modulation of antioxidant enzymes, heat shock protein, and intestinal microbiota of large yellow croaker (Larimichthys crocea) under acute cold stress. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.725899
Ma Q., Li L. Y., Le J. Y., Lu D. L., Qiao F., Zhang M. L., et al. (2018). Dietary microencapsulated oil improves immune function and intestinal health in Nile tilapia fed with high-fat diet. Aquaculture 496, 19–29. doi: 10.1016/j.aquaculture.2018.06.080
Ma N., Tian Y., Wu Y., Ma X. (2017). Contributions of the interaction between dietary protein and gut microbiota to intestinal health. Curr. Protein Pept. Sci. 18, 795–808. doi: 10.2174/1389203718666170216153505
Macfarlane G. T., Macfarlane S. (2012). Bacteria, colonic fermentation, and gastrointestinal health. J. AOAC Int. 95, 50–60. doi: 10.5740/jaoacint.sge_macfarlane
Magnúsdóttir S., Ravcheev D., De Crécy-Lagard V., Thiele I. (2015). Systematic genome assessment of B-vitamin biosynthesis suggests cooperation among gut microbes. Front. Genet. 6. doi: 10.3389/fgene.2015.00148
Maldonado Galdeano C., Cazorla S. I., Lemme Dumit J. M., Vélez E., Perdigón G. (2019). Beneficial effects of probiotic consumption on the immune system. Ann. Nutr. Metab. 74, 115–124. doi: 10.1159/000496426
Malhotra N., Ger T. R., Uapipatanakul B., Huang J. C., Chen K. H. C., Hsiao C. (2020). Review of copper and copper nanoparticle toxicity in fish. Der Nanomaterials 10, 1–28. doi: 10.3390/nano10061126
Mankiewicz J. L., Picklo M. J., Idso J., Cleveland B. M. (2022). Leptin Receptor Deficiency Results in Hyperphagia and Increased Fatty Acid Mobilization during Fasting in Rainbow Trout (Oncorhynchus mykiss). Biomolecules 12, 516. doi: 10.3390/biom12040516
Mayer E. A., Tillisch K., Gupta A. (2015). Gut/brain axis and the microbiota. J. Clin. Invest. 125, 926–938. doi: 10.1172/JCI76304
McKenzie D. J., Shingles A., Claireaux G., Domenici P. (2009). Sublethal concentrations of ammonia impair performance of the teleost fast-start escape response. Physiol. Biochem. Zool. 82, 353–362. doi: 10.1086/590218
Medina-Felix D., Garibay-Valdez E., Vargas-Albores F., Martínez-Porchas M. (2023). Fish disease and intestinal microbiota: A close and indivisible relationship. Rev. Aquac. 15, 820–839. doi: 10.1111/raq.12762
Medina-Felix D., Vargas-Albores F., Garibay-Valdez E., Martínez-Córdova L. R., Martínez-Porchas M. (2024). Gastrointestinal dysbiosis induced by Nocardia sp. infection in tilapia. Comp. Biochem. Physiol. Part D Genomics Proteomics 49, 101154. doi: 10.1016/j.cbd.2023.101154
Meng X. L., Li S., Qin C., Zhu Z. X., Hu W. P., Yang L. P., et al. (2018). Intestinal microbiota and lipid metabolism responses in the common carp (Cyprinus carpio L.) following copper exposure. Ecotoxicol. Environ. Saf. 160, 257–264. doi: 10.1016/j.ecoenv.2018.05.050
Merrifield D. L., Dimitroglou A., Foey A., Davies S. J., Baker R. T. M., Bøgwald J., et al. (2010). The current status and future focus of probiotic and prebiotic applications for salmonids. Aquaculture 302, 1–18. doi: 10.1016/j.aquaculture.2010.02.007
Michl S. C., Ratten J. M., Beyer M., Hasler M., La Roche J., Schulz C. (2017). The malleable gut microbiome of juvenile rainbow trout (Oncorhynchus mykiss): Dietdependent shifts of bacterial community structures. PloS One 12, 1–21. doi: 10.1371/journal.pone.0177735
Milligan G., Stoddart L. A., Smith N. J. (2009). Agonism and allosterism: The pharmacology of the free fatty acid receptors FFA2 and FFA3. Br. J. Pharmacol. 158, 146–153. doi: 10.1111/j.1476-5381.2009.00421.x
Minniti G., Hagen L. H., Porcellato D., Jørgensen S. M., Pope P. B., Vaaje-Kolstad G. (2017). The skin-mucus microbial community of farmed atlantic salmon (Salmo salar). Front. Microbiol. 8. doi: 10.3389/fmicb.2017.02043
Mirghaed A. T., Yarahmadi P., Hosseinifar S. H., Tahmasebi D., Gheisvandi N., Ghaedi A. (2018). The effects singular or combined administration of fermentabl fiber and probiotic on mucosal immune parameters, digestive enzyme activity, gut microbiota and growth performance of Caspian white fish (Rutilus frisii kutum) fingerlings. Fish Shellfish Immunol. 77, 194–199. doi: 10.1016/j.fsi.2018.02.007
Mirzapour-Rezaee S. S., Farhangi M., Rafiee G. (2017). Combined effects of dietary mannan- and fructo-oligosaccharide on growth indices, body composition, intestinal bacterial flora and digestive enzymes activity of regal peacock (Aulonocara stuartgranti). Aquac. Nutr. 23, 629–636. doi: 10.1111/anu.12430
Mishima Y., Oka A., Liu B., Herzog J. W., Eun C. S., Fan T. J., et al. (2019). Microbiota maintain colonic homeostasis by activating TLR2/MyD88/PI3K signaling in IL-10-producing regulatory B cells. J. Clin. Invest. 129, 3702–3716. doi: 10.1172/JCI93820
Mohammadian T., Ghanei-Motlagh R., Molayemraftar T., Mesbah M., Zarea M., Mohtashamipour H., et al. (2021). Modulation of growth performance, gut microflora, non-specific immunity and gene expression of proinflammatory cytokines in shabout (Tor grypus) upon dietary prebiotic supplementation. Fish Shellfish Immunol. 112, 38–45. doi: 10.1016/j.fsi.2021.02.012
Moretti C. H., Schiffer T. A., Li X., Weitzberg E., Carlström M., Lundberg J. O. (2021). Germ-free mice are not protected against diet-induced obesity and metabolic dysfunction. Acta Physiol. (Oxf). 231, e13581. doi: 10.1111/apha.13581
Morshed S. M., Chen Y. Y., Lin C. H., Chen Y. P., Lee T. H. (2023). Freshwater transfer affected intestinal microbiota with correlation to cytokine gene expression in Asian sea bass. Front. Microbiol. 14. doi: 10.3389/fmicb.2023.1097954
Morshed S. M., Lee T. H. (2023). The role of the microbiome on fish mucosal immunity under changing environments. Fish Shellfish Immunol. 139, 108877. doi: 10.1016/j.fsi.2023.108877
Mountfort D. O., Campbell J., Clements K. D. (2002). Hindgut fermentation in three species of marine herbivorous fish. Appl. Environ. Microbiol. 68. doi: 10.1128/AEM.68.3.1374-1380.2002
Moxness Reksten A., Ho Q. T., Nøstbakken O. J., Wik Markhus M., Kjellevold M., Bøkevoll A., et al. (2022). Temporal variations in the nutrient content of Norwegian farmed Atlantic salmon (Salmo salar), 2005–2020. Food Chem. 373, 131445. doi: 10.1016/j.foodchem.2021.131445
Munita J. M., Arias C. A. (2016). “Mechanisms of antibiotic resistance,” In: Kudva N. A. C. I. T., Plummer P. J., Zhang Q., Nicholson T. L., Bannantine J. P., Bellaire B. H., et al. Virulence Mech. Bact. Pathog. Wiley, 481–511. doi: 10.1128/9781555819286.ch17
Navarrete P., Mardones P., Opazo R., Espejo R., Romero J. (2008). Oxytetracycline treatment reduces bacterial diversity of intestinal microbiota of Atlantic salmon. J. Aquat. Anim. Health 20, 177–183. doi: 10.1577/H07-043.1
Nawaz A., Bakhsh javaid A., Irshad S., Hoseinifar S. H., Xiong H. (2018). The functionality of prebiotics as immunostimulant: Evidences from trials on terrestrial and aquatic animals. Fish Shellfish Immunol. 76, 272–278. doi: 10.1016/j.fsi.2018.03.004
Nayak S. K. (2010). Probiotics and immunity: A fish perspective. Fish Shellfish Immunol. 29, 2–14. doi: 10.1016/j.fsi.2010.02.017
Neuman C., Hatje E., Zarkasi K. Z., Smullen R., Bowman J. P., Katouli M. (2016). The effect of diet and environmental temperature on the faecal microbiota of farmed Tasmanian Atlantic Salmon (Salmo salar L.). Aquac. Res. 47, 660–672. doi: 10.1111/are.12522
Ni J., Yan Q., Yu Y., Zhang T. (2014). Factors influencing the grass carp gut microbiome and its effect on metabolism. FEMS Microbiol. Ecol. 87, 704–714. doi: 10.1111/1574-6941.12256
Nikouli E., Kormas K. A., Jin Y., Olsen Y., Bakke I. (2021). Dietary lipid effects on gut microbiota of first feeding atlantic salmon (Salmo salar L.). Front. Mar. Sci. 8, 1–10. doi: 10.3389/fmars.2021.665576
Nilsson S. (2012). Autonomic nerve function in the vertebrates (Berlin, Heidelberg, Germany: Springer).
Nimalan N., Sørensen S. L., Fečkaninová A., Koščová J., Mudroňová D., Gancarčíková S., et al. (2022). Mucosal barrier status in Atlantic salmon fed marine or plant-based diets supplemented with probiotics. Aquaculture 547, 737516. doi: 10.1016/j.aquaculture.2021.737516
Niu M., Harshaw K., Xiang Q., Zhou Y., Xiang P., Ju Z., et al. (2024). Macrophytes mitigate Microcystis aeruginosa-induced fish appetite suppression via intestinal metabolite regulation. Ecotoxicol. Environ. Saf. 288, 117348. doi: 10.1016/j.ecoenv.2024.117348
Niu K.-M., Khosravi S., Kothari D., Lee W.-D., Lim J.-M., Lee B.-J., et al. (2019). Effects of dietary multi-strain probiotics supplementation in a low fishmeal diet on growth performance, nutrient utilization, proximate composition, immune parameters, and gut microbiota of juvenile olive flounder (Paralichthys olivaceus). Fish Shellfish Immunol. 93, 258–268. doi: 10.1016/j.fsi.2019.07.056
Niu K. M., Lee B. J., Kothari D., Lee W., Hur S. W., Lim S. G., et al. (2020). Dietary effect of low fish meal aquafeed on gut microbiota in olive flounder (Paralichthys olivaceus) at different growth stages. Microbiologyopen 9, 1–10. doi: 10.1002/mbo3.992
Norland S., Eilertsen M., Rønnestad I., Helvik J. V., Gomes A. S. (2023). Mapping key neuropeptides involved in the melanocortin system in Atlantic salmon (Salmo salar) brain. J. Comp. Neurol. 531, 89–115. doi: 10.1002/cne.25415
Nyman A., Huyben D., Lundh T., Dicksved J. (2017). Effects of microbe- and mussel-based diets on the gut microbiota in Arctic charr (Salvelinus alpinus). Aquac. Rep. 5, 34–40. doi: 10.1016/j.aqrep.2016.12.003
Obradovic M., Sudar-Milovanovic E., Soskic S., Essack M., Arya S., Stewart A. J., et al. (2021). Leptin and obesity: role and clinical implication. Front. Endocrinol. (Lausanne). 12. doi: 10.3389/fendo.2021.585887
Ofek T., Lalzar M., Laviad-Shitrit S., Izhaki I., Halpern M. (2021). Comparative study of intestinal microbiota composition of six edible fish species. Front. Microbiol. 12. doi: 10.3389/fmicb.2021.760266
Offermanns S. (2017). Hydroxy-carboxylic acid receptor actions in metabolism. Trends Endocrinol. Metab. 28, 227–236. doi: 10.1016/j.tem.2016.11.007
Olsen R. E., Sundell K., Hansen T., Hemre G.-I., Myklebust R., Mayhew T. M., et al. (2002). Acute stress alters the intestinal lining of Atlantic salmon, Salmo salar L.: An electron microscopical study. Fish Physiol. Biochem. 26, 211–221. doi: 10.1023/A:1026217719534
Olsen R. E., Sundell K., Mayhew T. M., Myklebust R., Ringø E. (2005). Acute stress alters intestinal function of rainbow trout, Oncorhynchus mykiss (Walbaum). Aquaculture 250, 480–495. doi: 10.1016/j.aquaculture.2005.03.014
Olsen R. E., Sundell K., Ringø E., Myklebust R., Hemre G.-I., Hansen T., et al. (2008). The acute stress response in fed and food deprived Atlantic cod, Gadus morhua L. Aquaculture 280, 232–241. doi: 10.1016/j.aquaculture.2008.05.006
Olsen R. E., Svardal A., Eide T., Wargelius A. (2012). Stress and expression of cyclooxygenases (cox1, cox2a, cox2b) and intestinal eicosanoids, in Atlantic salmon, Salmo salar L. Fish Physiol. Biochem. 38, 951–962. doi: 10.1007/s10695-011-9581-1
Parada Venegas D., de la Fuente M. K., Landskron G., González M. J., Quera R., Dijkstra G., et al. (2019). Short chain fatty acids (SCFAs)-mediated gut epithelial and immune regulation and its relevance for inflammatory bowel diseases. Front. Immunol. 10. doi: 10.3389/fimmu.2019.00277
Park S. K., Kim M. S., Roh S. W., Bae J. W. (2012). Blautia stercoris sp. nov., isolated from human faeces. Int. J. Syst. Evol. Microbiol. 62, 776–779. doi: 10.1099/ijs.0.031625-0
Payne C. J., Turnbull J. F., Mackenzie S., Crumlish M. (2021). Investigating the effect of an oxytetracycline treatment on the gut microbiome and antimicrobial resistance gene dynamics in Nile Tilapia (Oreochromis Niloticus). Antibiotics 10, 1213. doi: 10.3390/antibiotics10101213
Payne C. J., Turnbull J. F., MacKenzie S., Crumlish M. (2022). The effect of oxytetracycline treatment on the gut microbiome community dynamics in rainbow trout (Oncorhynchus mykiss) over time. Aquaculture 560, 738559. doi: 10.1016/j.aquaculture.2022.738559
Pelusio N. F., Scicchitano D., Parma L., Dondi F., Brini E., D’Amico F., et al. (2021). Interaction between dietary lipid level and seasonal temperature changes in gilthead sea bream sparus aurata: effects on growth, fat deposition, plasma biochemistry, digestive enzyme activity, and gut bacterial community. Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.664701
Peng X., Li S., Luo J., Wu X., Liu L. (2013). Effects of dietary fibers and their mixtures on short chain fatty acids and microbiota in mice guts. Food Funct. 4, 932–938. doi: 10.1039/c3fo60052a
Peng M., Luo H., Kumar V., Kajbaf K., Hu Y., Yang G. (2019). Dysbiosis of intestinal microbiota induced by dietary oxidized fish oil and recovery of diet-induced dysbiosis via taurine supplementation in rice field eel (Monopterus albus). Aquaculture 512, 734288. doi: 10.1016/j.aquaculture.2019.734288
Pereira G. D., Teixeira C., Couto J., Dias J., Rema P., Gonçalves A. T. (2024). Dietary protein quality affects the interplay between gut microbiota and host performance in nile tilapia. Animals 14, 1–23. doi: 10.3390/ani14050714
Pérez-Pascual D., Pérez-Cobas A. E., Rigaudeau D., Rochat T., Bernardet J. F., Skiba-Cassy S., et al. (2021). Sustainable plant-based diets promote rainbow trout gut microbiota richness and do not alter resistance to bacterial infection. Anim. Microbiome 3. doi: 10.1186/s42523-021-00107-2
Petersen C., Round J. L. (2014). Defining dysbiosis and its influence on host immunity and disease. Cell. Microbiol. 16, 1024–1033. doi: 10.1111/cmi.12308
Petit J., Wiegertjes G. F. (2022). Conservation of members of the free fatty acid receptor gene family in common carp. Dev. Comp. Immunol. 126, 104240. doi: 10.1016/j.dci.2021.104240
Poolsawat L., Li X., Xu X., Rahman M. M., Boonpeng N., Leng X. (2021). Dietary xylooligosaccharide improved growth, nutrient utilization, gut microbiota and disease resistance of tilapia (Oreochromis niloticus x O. aureus). Anim. Feed Sci. Technol. 275, 114872. doi: 10.1016/j.anifeedsci.2021.114872
Poolsawat L., Li X., Yang H., Yang P., Amirul M., Chowdhury K., et al. (2020). The potentials of fructooligosaccharide on growth, feed utilization, immune and antioxidant parameters, microbial community and disease resistance of tilapia (Oreochromis niloticus × O. aureus). 51, 4430–4442. doi: 10.1111/are.14786
Preena P. G., Swaminathan T. R., Kumar V. J. R., Singh I. S. B. (2020). Antimicrobial resistance in aquaculture: a crisis for concern. Biol. (Bratisl). 75, 1497–1517. doi: 10.2478/s11756-020-00456-4
Qi X., Xue M., Shi K., Wang G., Ling F. (2024). Evaluating Pseudomonas monteilii JK-1 as an in-feed probiotic: Enhancing growth, immune-antioxidant, disease resistance and modulating gut microflora composition in grass carp (Ctenopharyngodon idella). Aquaculture 585, 740715. doi: 10.1016/j.aquaculture.2024.740715
Qi X. Z., Xue M. Y., Yang S. B., Zha J. W., Wang G. X., Ling F. (2017). Ammonia exposure alters the expression of immune-related and antioxidant enzymes-related genes and the gut microbial community of crucian carp (Carassius auratus). Fish Shellfish Immunol. 70, 485–492. doi: 10.1016/j.fsi.2017.09.043
Qiu H., Huang L., Wang H., Tao C., Ran Z., Xu J., et al. (2024). Effects of Lactobacillus acidophilus AC on the growth, intestinal flora and metabolism of zebrafish (Danio rerio). Fish Shellfish Immunol. 149, 109570. doi: 10.1016/j.fsi.2024.109570
Ramírez C., Coronado J., Silva A., Romero J. (2018). Cetobacterium is a major component of the microbiome of giant amazonian fish (Arapaima gigas) in Ecuador. Anim. an Open Access J. MDPI 8. doi: 10.3390/ani8110189
Randall D. J., Tsui T. K. N. (2002). Ammonia toxicity in fish. Mar. pollut. Bull. 45, 17–23. doi: 10.1016/S0025-326X(02)00227-8
Rawls J. F., Samuel B. S., Gordon J. I. (2004). Gnotobiotic zebrafish reveal evolutionarily conserved responses to the gut microbiota. Proc. Natl. Acad. Sci. U. S. A. 101, 4596–4601. doi: 10.1073/pnas.0400706101
Ray A. K., Ghosh K., Ringø E. (2012). Enzyme-producing bacteria isolated from fish gut: A review. Aquac. Nutr. 18, 465–492. doi: 10.1111/j.1365-2095.2012.00943.x
Read M. N., Holmes A. J. (2017). Towards an integrative understanding of diet-host-gut microbiome interactions. Front. Immunol. 8. doi: 10.3389/fimmu.2017.00538
Reveco F. E., Øverland M., Romarheim O. H., Mydland L. T. (2014). Intestinal bacterial community structure differs between healthy and inflamed intestines in Atlantic salmon (Salmo salar L.). Aquaculture 420–421, 262–269. doi: 10.1016/j.aquaculture.2013.11.007
Reverter M., Tapissier-Bontemps N., Lecchini D., Banaigs B., Sasal P. (2018). Biological and ecological roles of external fish mucus: A review. Fishes 3, 41. doi: 10.3390/fishes3040041
Riccio P., Rossano R. (2020). The human gut microbiota is neither an organ nor a commensal. FEBS Lett. 594, 3262–3271. doi: 10.1002/1873-3468.13946
Ridlon J. M., Gaskins H. R. (2024). Another renaissance for bile acid gastrointestinal microbiology. Nat. Rev. Gastroenterol. Hepatol. 21, 348–364. doi: 10.1038/s41575-024-00896-2
Rimoldi S., Antonini M., Gasco L., Moroni F., Terova G. (2021). Intestinal microbial communities of rainbow trout (Oncorhynchus mykiss) may be improved by feeding a Hermetia illucens meal/low-fishmeal diet. Fish Physiol. Biochem. 47, 365–380. doi: 10.1007/s10695-020-00918-1
Rimoldi S., Gliozheni E., Ascione C., Gini E., Terova G. (2018a). Effect of a specific composition of short- and medium-chain fatty acid 1-Monoglycerides on growth performances and gut microbiota of gilthead sea bream (Sparus aurata). PeerJ 2018, 1–27. doi: 10.7717/peerj.5355
Rimoldi S., Terova G., Ascione C., Giannico R., Brambilla F. (2018b). Next generation sequencing for gut microbiome characterization in rainbow trout (Oncorhynchus mykiss) fed animal by-product meals as an alternative to fishmeal protein sources. PloS One 13, 1–29. doi: 10.1371/journal.pone.0193652
Rimoldi S., Torrecillas S., Montero D., Gini E., Makol A., Victoria Valdenegro V., et al. (2020). Assessment of dietary supplementation with galactomannan oligosaccharides and phytogenics on gut microbiota of European sea bass (Dicentrarchus Labrax) fed low fishmeal and fish oil based diet. PloS One 15, 1–30. doi: 10.1371/journal.pone.0231494
Ringø E., Bendiksen H. R., Gausen S. J., Sundsfjord A., Olsen R. E. (1998). The effect of dietary fatty acids on lactic acid bacteria associated with the epithelial mucosa and from faecalia of Arctic charr, Salvelinus alpinus (L.). J. Appl. Microbiol. 85, 855–864. doi: 10.1046/j.1365-2672.1998.00595.x
Ringø E., Li X., Doan H.v., Ghosh K. (2022). Interesting probiotic bacteria other than the more widely used lactic acid bacteria and bacilli in finfish. Front. Mar. Sci. 9. doi: 10.3389/fmars.2022.848037
Ringø E., Lødemel J. B., Myklebust R., Jensen L., Lund V., Mayhew T. M., et al. (2002). The effects of soybean, linseed and marine oils on aerobic gut microbiota of Arctic charr Salvelinus alpinus L. before and after challenge with Aeromonas salmonicida ssp. salmonicida. Aquac. Res. 33, 591–606. doi: 10.1046/j.1365-2109.2002.00697.x
Ringø E., Olsen R. E. (1999). The effect of diet on aerobic bacterial flora associated with intestine of Arctic charr (Salvelinus alpinus L.). J. Appl. Microbiol. 86, 22–28. doi: 10.1046/j.1365-2672.1999.00631.x
Ringø E., Olsen R. E., Gifstad T.Ø., Dalmo R. A., Amlund H., Hemre G.-I., et al. (2010). Prebiotics in aquaculture: a review. Aquac. Nutr. 16, 117–136. doi: 10.1111/j.1365-2095.2009.00731.x
Ringø E., Salinas I., Olsen R. E., Nyhaug A., Myklebust R., Mayhew T. M. (2007). Histological changes in intestine of Atlantic salmon (Salmo salar L.) following in vitro exposure to pathogenic and probiotic bacterial strains. Cell Tissue Res. 328, 109–116. doi: 10.1007/s00441-006-0323-0
Ringø E., Sperstad S., Myklebust R., Mayhew T. M., Mjelde A., Melle W., et al. (2006). The effect of dietary krill supplementation on epithelium-associated bacteria in the hindgut of Atlantic salmon (Salmo salar L.): a microbial and electron microscopical study. Aquac. Res. 37, 1644–1653. doi: 10.1111/j.1365-2109.2006.01611.x
Ringø E., Van Doan H., Lee S. H., Soltani M., Hoseinifar S. H., Harikrishnan R., et al. (2020). Probiotics, lactic acid bacteria and bacilli: interesting supplementation for aquaculture. J. Appl. Microbiol. 129, 116–136. doi: 10.1111/jam.14628
Ringø E., Zhou Z., Vecino J. L. G., Wadsworth S., Romero J., Krogdahl Å., et al. (2016). Effect of dietary components on the gut microbiota of aquatic animals. A never-ending story? Aquac. Nutr. 22, 219–282. doi: 10.1111/anu.12346
Ringø E., Zhou Z., He S., Olsen R. E. (2014). Effect of stress on intestinal microbiota of Arctic charr, Atlantic salmon, rainbow trout and Atlantic cod: A review. African Journal of Microbiology Research 8, 609–618. doi: 10.5897/AJMR2013.6395
Rivière A., Selak M., Lantin D., Leroy F., De Vuyst L. (2016). Bifidobacteria and Butyrate-Producing Colon Bacteria: Importance and Strategies for Their Stimulation in the Human Gut. Front. Microbiol. 7. doi: 10.3389/fmicb.2016.00979
Rojas-García C. R., Applebaum S. L., Morais S., Rønnestad I. (2016). Trans-intestinal absorption rates differ between free amino acids during larval development in Atlantic herring (Clupea harengus). Aquaculture 464, 222–228. doi: 10.1016/j.aquaculture.2016.06.029
Rønnestad I., Gomes A. S., Murashita K., Angotzi R., Jönsson E., Volkoff H. (2017). Appetite-controlling endocrine systems in teleosts. Front. Endocrinol. (Lausanne). 8, eCollection. doi: 10.3389/fendo.2017.00073
Rudi K., Angell I. L., Pope P. B., Vik J. O., Sandve S. R., Snipen L.-G. (2018). Stabl core gut microbiota across the freshwater-to-saltwater transition for farmed atlantic salmon. Appl. Environ. Microbiol. 84. doi: 10.1128/AEM.01974-17
Ruiz A., Andree K. B., Furones D., Holhorea P. G., Calduch-Giner J., Viñas M., et al. (2023). Modulation of gut microbiota and intestinal immune response in gilthead seabream (Sparus aurata) by dietary bile salt supplementation. Front. Microbiol. 14. doi: 10.3389/fmicb.2023.1123716
Ruzauskas M., Klimiene I., Armalyte J., Bartkiene E., Siugzdiniene R., Skerniskyte J., et al. (2018). Composition and antimicrobial resistance profile of Gram-negative microbiota prevalent in aquacultured fish. J. Food Saf. 38, 1–10. doi: 10.1111/jfs.12447
Salinas I., Myklebust R., Esteban M. A., Olsen R. E., Meseguer J., Ringø E. (2008). In vitro studies of Lactobacillus delbrueckii subsp. lactis in Atlantic salmon (Salmo salar L.) foregut: Tissue responses and evidence of protection against Aeromonas salmonicida subsp. salmonicida epithelial damage. Vet. Microbiol. 128, 167–177. doi: 10.1016/j.vetmic.2007.10.011
Sánchez-Cueto P., Stavrakidis-Zachou O., Clos-Garcia M., Bosch M., Papandroulakis N., Lladó S. (2023). Mediterranean Sea heatwaves jeopardize greater amberjack’s (Seriola dumerili) aquaculture productivity through impacts on the fish microbiota. ISME Commun. 3, 1–10. doi: 10.1038/s43705-023-00243-7
Santos L., Ramos F. (2018). Antimicrobial resistance in aquaculture: Current knowledge and alternatives to tackle the problem. Int. J. Antimicrob. Agents 52, 135–143. doi: 10.1016/j.ijantimicag.2018.03.010
Schmidt J., Smith N. J., Christiansen E., Tikhonova I. G., Grundmann M., Hudson B. D., et al. (2011). Selective orthosteric free fatty acid receptor 2 (FFA2) agonists: Identification of the structural and chemical requirements for selective activation of FFA2 versus FFA3. J. Biol. Chem. 286, 10628–10640. doi: 10.1074/jbc.M110.210872
Schram E., Roques J. A. C., Abbink W., Spanings T., de Vries P., Bierman S., et al. (2010). The impact of elevated water ammonia concentration on physiology, growth and feed intake of African catfish (Clarias gariepinus). Aquaculture 306, 108–115. doi: 10.1016/j.aquaculture.2010.06.005
Schreck C. B., Tort L., Brauner (2016). “1 - the concept of stress in fish,” in Biology of stress in fish. Eds. Schreck C. B., Tort L., Farrell A. P. (Academic Press), 1–34. doi: 10.1016/B978-0-12-802728-8.00001-1
Semova I., Carten J. D., Stombaugh J., MacKey L. C., Knight R., Farber S. A., et al. (2012). Microbiota regulate intestinal absorption and metabolism of fatty acids in the zebrafish. Cell Host Microbe. 12, 277–288. doi: 10.1016/j.chom.2012.08.003
Sender R., Fuchs S., Milo R. (2016). Are we really vastly outnumbered? Revisiting the ratio of bacterial to host cells in humans. Cell 164, 337–340. doi: 10.1016/j.cell.2016.01.013
Sheng Y., Ren H., Limbu S. M., Sun Y., Qiao F., Zhai W., et al. (2018). The presence or absence of intestinal microbiota affects lipid deposition and related genes expression in zebrafish (Danio rerio). Front. Microbiol. 9. doi: 10.3389/fmicb.2018.01124
Shoubridge A. P., Choo J. M., Martin A. M., Keating D. J., Wong M.-L., Licinio J., et al. (2022). The gut microbiome and mental health: advances in research and emerging priorities. Mol. Psychiatry 27, 1908–1919. doi: 10.1038/s41380-022-01479-w
Singh N., Gurav A., Sivaprakasam S., Brady E., Padia R., Shi H., et al. (2014). Activation of Gpr109a, receptor for niacin and the commensal metabolite butyrate, suppresses colonic inflammation and carcinogenesis. Immunity. 40, 128–139. doi: 10.1016/j.immuni.2013.12.007
Singh A., Vidakovic A., Hjertner B., Krikigianni E., Karnaouri A., Christakopoulos P., et al. (2024). Effects of dietary supplementation of lignocellulose-derived cello-oligosaccharides on growth performance, antioxidant capacity, immune response, and intestinal microbiota in rainbow trout (Oncorhynchus mykiss). Aquaculture 578, 740002. doi: 10.1016/j.aquaculture.2023.740002
Sinha A. K., AbdElgawad H., Giblen T., Zinta G., De Rop M., Asard H., et al. (2014). Anti-oxidative defences are modulated differentially in three freshwater teleosts in response to ammonia-induced oxidative stress. PloS One 9, e95319. doi: 10.1371/journal.pone.0095319
Smith P. M., Howitt M. R., Panikov N., Michaud M., Gallini C. A., Bohlooly-Y M., et al. (2013). The microbial metabolites, short-chain fatty acids, regulate colonic T reg cell homeostasis. Science. 341, 569–573. doi: 10.1126/science.1241165
Soengas J. L., Cerdá-Reverter J. M., Delgado M. J. (2018). Central regulation of food intake in fish: An evolutionary perspective. J. Mol. Endocrinol. 60, R171–R199. doi: 10.1530/JME-17-0320
Soliman M. L., Rosenberger T. A. (2011). Acetate supplementation increases brain histone acetylation and inhibits histone deacetylase activity and expression. Mol. Cell. Biochem. 352, 173–180. doi: 10.1007/s11010-011-0751-3
Sonnenburg E. D., Sonnenburg J. L. (2014). Starving our microbial self: the deleterious consequences of a diet deficient in microbiota-accessible carbohydrates. Cell Metab. 20, 779–786. doi: 10.1016/j.cmet.2014.07.003
Soriano E. L., Ramírez D. T., Araujo D. R., Gómez-Gil B., Castro L. I., Sánchez (2018). Effect of temperature and dietary lipid proportion on gut microbiota in yellowtail kingfish Seriola lalandi juveniles. Aquaculture 497, 269–277. doi: 10.1016/j.aquaculture.2018.07.065
Spanggaard B., Huber I., Nielsen J., Nielsen T., Appel K. F., Gram L. (2000). The microflora of rainbow trout intestine: a comparison of traditional and molecular identification. Aquaculture 182, 1–15. doi: 10.1016/S0044-8486(99)00250-1
Spetz A.-L. (2003). Lateral gene transfer in the genomic era: Lateral DNA Transfer: Mechanisms and Consequences. J. Cell Sci. 116, 2821. doi: 10.1242/jcs.00658
Steiner K., Laroche O., Walker S. P., Symonds J. E. (2022). Effects of water temperature on the gut microbiome and physiology of Chinook salmon (Oncorhynchus tshawytscha) reared in a freshwater recirculating system. Aquaculture 560, 738529. doi: 10.1016/j.aquaculture.2022.738529
Steinert R. E., Feinle-Bisset C., Asarian L., Horowitz M., Beglinger C., Geary N. (2017). Ghrelin, CCK, GLP-1, and PYY(3-36): secretory controls and physiological roles in eating and glycemia in health, obesity, and after RYGB. Physiol. Rev. 97, 411–463. doi: 10.1152/physrev.00031.2014
Strandwitz P. (2018). Neurotransmitter modulation by the gut microbiota. Brain Res. 1693, 128–133. doi: 10.1016/j.brainres.2018.03.015
Sugita H., Miyajima C., Deguchi Y. (1991). The vitamin B12-producing ability of the intestinal microflora of freshwater fish. Aquaculture 92, 267–276. doi: 10.1016/0044-8486(91)90028-6
Sun S., Gong C., Deng C., Yu H., Zheng D., Wang L., et al. (2023). Effects of salinity stress on the growth performance, health status, and intestinal microbiota of juvenile Micropterus salmoides. Aquaculture 576, 739888. doi: 10.1016/j.aquaculture.2023.739888
Sun B.-Y., He W., Yang H.-X., Tian D.-Y., Jian P.-Y., Wu K., et al. (2022). Increased susceptibility to Aeromonas hydrophila infection in grass carp with antibiotic-induced intestinal dysbiosis. Aquaculture 552, 737969. doi: 10.1016/j.aquaculture.2022.737969
Sun M., Wu W., Chen L., Yang W., Huang X., Ma C., et al. (2018). Microbiota-derived short-chain fatty acids promote Th1 cell IL-10 production to maintain intestinal homeostasis. Nat. Commun. 9, 3555. doi: 10.1038/s41467-018-05901-2
Sun B.-Y., Yang H.-X., He W., Tian D.-Y., Kou H.-Y., Wu K., et al. (2021). A grass carp model with an antibiotic-disrupted intestinal microbiota. Aquaculture 541, 736790. doi: 10.1016/j.aquaculture.2021.736790
Sun C., Zhang M., Feng D., Wang S., Li M. (2024). Effects of dietary D-mannose supplementation on growth performance, intestinal digestive capacity, gut microbiota, and ammonia tolerance of largemouth bass Micropterus salmoides. Aquac. Rep. 36, 102054. doi: 10.1016/j.aqrep.2024.102054
Sylvain F.É., Cheaib B., Llewellyn M., Gabriel Correia T., Barros Fagundes D., Luis Val A., et al. (2016). pH drop impacts differentially skin and gut microbiota of the Amazonian fish tambaqui (Colossoma macropomum). Sci. Rep. 6, 32032. doi: 10.1038/srep32032
Tacon A. G. J., Metian M. (2008). Global overview on the use of fish meal and fish oil in industrially compounded aquafeeds: Trends and future prospects. Aquaculture 285, 146–158. doi: 10.1016/j.aquaculture.2008.08.015
Taggart A. K. P., Kero J., Gan X., Cai T.-Q., Cheng K., Ippolito M., et al. (2005). (D)-beta-Hydroxybutyrate inhibits adipocyte lipolysis via the nicotinic acid receptor PUMA-G. J. Biol. Chem. 280, 26649–26652. doi: 10.1074/jbc.C500213200
Tan H. Y., Chen S.-W., Hu S.-Y. (2019). Improvements in the growth performance, immunity, disease resistance, and gut microbiota by the probiotic Rummeliibacillus stabekisii in Nile tilapia (Oreochromis niloticus). Fish Shellfish Immunol. 92, 265–275. doi: 10.1016/j.fsi.2019.06.027
Tanca A., Abbondio M., Palomba A., Fraumene C., Marongiu F., Serra M., et al. (2018). Caloric restriction promotes functional changes involving short-chain fatty acid biosynthesis in the rat gut microbiota. Sci. Rep. 8, 1–13. doi: 10.1038/s41598-018-33100-y
Tang S., Liu S., Zhang J., Zhou L., Wang X., Zhao Q., et al. (2020). Relief of hypersaline stress in Nile tilapia Oreochromis niloticus by dietary supplementation of a host-derived Bacillus subtilis strain. Aquaculture 528, 735542. doi: 10.1016/j.aquaculture.2020.735542
Tavares-Dias M. (2021). Toxic, physiological, histomorphological, growth performance and antiparasitic effects of copper sulphate in fish aquaculture. Aquaculture 535, 736350. doi: 10.1016/j.aquaculture.2021.736350
Tazoe H., Otomo Y., Karaki S. I., Kato I., Fukami Y., Terasaki M., et al. (2009). Expression of short-chain fatty acid receptor GPR41 in the human colon. Biomed. Res. 30, 149–156. doi: 10.2220/biomedres.30.149
Terova G., Rimoldi S., Ascione C., Gini E., Ceccotti C., Gasco L. (2019). Rainbow trout (Oncorhynchus mykiss) gut microbiota is modulated by insect meal from Hermetia illucens prepupae in the diet. Rev. Fish Biol. Fish. 29, 465–486. doi: 10.1007/s11160-019-09558-y
Thangaraju M., Cresci G. A., Liu K., Ananth S., Gnanaprakasam J. P., Browning D. D., et al. (2009). GPFM 09A is a G-protein-coupled receptor for the bacterial fermentation product butyrate and functions as a tumor suppressor in colon. Cancer Res. 69, 2826–2832. doi: 10.1158/0008-5472.CAN-08-4466
Tolås I., Kalananthan T., Gomes A. S., Lai F., Norland S., Murashita K., et al. (2021). Regional Expression of npy mRNA Paralogs in the Brain of Atlantic Salmon (Salmo salar, L.) and Response to Fasting. Front. Physiol. 12. doi: 10.3389/fphys.2021.720639
Tooley K. L. (2020). Effects of the human gut microbiota on cognitive performance, brain structure and function: A narrative review. Nutrients 12, 3009. doi: 10.3390/nu12103009
Tort L., Balasch J. C., Buchmann K. (2022). “Stress and immunity in fish BT - principles of fish immunology : from cells and molecules to host protection,” Ed. Secombes C. J. Principles of Fish Immunology : From Cells and Molecules to Host Protection. (Springer International Publishing, Cham). doi: 10.1007/978-3-030-85420-1_20
Tottey W., Feria-Gervasio D., Gaci N., Laillet B., Pujos E., Martin J. F., et al. (2017). Colonic transit time is a driven force of the gut microbiota composition and metabolism: In vitro evidence. J. Neurogastroenterol. Motil. 23, 124–134. doi: 10.5056/jnm16042
Tsuchiya C., Sakata T., Sugita H. (2008). Novel ecological niche of Cetobacterium somerae, an anaerobic bacterium in the intestinal tracts of freshwater fish. Lett. Appl. Microbiol. 46, 43–48. doi: 10.1111/j.1472-765X.2007.02258.x
Tudorache C., Blust R., De Boeck G. (2008). Social interactions, predation behaviour and fast start performance are affected by ammonia exposure in brown trout (Salmo trutta L.). Aquat. Toxicol. 90, 145–153. doi: 10.1016/j.aquatox.2008.08.009
Turchini G. M., Francis D. S., Du Z.-Y., Olsen R. E., Ringø E., Tocher D. R. (2022). Chapter 5 - the lipids. Eds. Hardy R. W., Kaushik F. E. (Academic Press), 303–467. doi: 10.1016/B978-0-12-819587-1.00003-3
Turnbaugh P. J., Ley R. E., Hamady M., Fraser-Liggett C. M., Knight R., Gordon J. (2007). The human microbiome project. I.Nature 449, 804–810. doi: 10.1038/nature06244
Uren Webster T. M., Consuegra S., Garcia de Leaniz C. (2021). Early life stress causes persistent impacts on the microbiome of Atlantic salmon. Comp. Biochem. Physiol. Part D. Genomics Proteomics 40, 100888. doi: 10.1016/j.cbd.2021.100888
Uren Webster T. M., Rodriguez-Barreto D., Castaldo G., Gough P., Consuegra S., Garcia de Leaniz C. (2020). Environmental plasticity and colonisation history in the Atlantic salmon microbiome: A translocation experiment. Mol. Ecol. 29, 886–898. doi: 10.1111/mec.15369
Vacca F., Gomes A. S., De Gennaro M., Rønnestad I., Bossi E., Verri T. (2023). The teleost fish PepT1-type peptide transporters and their relationships with neutral and charged substrates. Front. Physiol. 14. doi: 10.3389/fphys.2023.1186475
Vargas-Albores F., Martínez-Córdova L. R., Hernández-Mendoza A., Cicala F., Lago-Lestón A., Martínez-Porchas M. (2021). Therapeutic modulation of fish gut microbiota, a feasible strategy for aquaculture? Aquaculture 544, 737050. doi: 10.1016/j.aquaculture.2021.737050
Varó I., Osorio K., Estensoro I., Naya-Català F., Sitjà-Bobadilla A., Navarro J. C., et al. (2021). Effect of virgin low density polyethylene microplastic ingestion on intestinal histopathology and microbiota of gilthead sea bream. Aquaculture 545, 737245. doi: 10.1016/j.aquaculture.2021.737245
Vindas M. A., Gorissen M., Höglund E., Flik G., Tronci V., Damsgård B., et al. (2017). How do individuals cope with stress? Behavioural, physiological and neuronal differences between proactive and reactive coping styles in fish. J. Exp. Biol. 220, 1524–1532. doi: 10.1242/jeb.153213
Volkoff H. (2024). The effects of environmental changes on the endocrine regulation of feeding in fishes. Philos. Trans. R. Soc. B: Biol. Sci. 379, 20220503. doi: 10.1098/rstb.2022.0503
Volkoff H., Rønnestad I. (2020). Effects of temperature on feeding and digestive processes in fish. Temperature 7, 307–320. doi: 10.1080/23328940.2020.1765950
Waldecker M., Kautenburger T., Daumann H., Busch C., Schrenk D. (2008). Inhibition of histone-deacetylase activity by short-chain fatty acids and some polyphenol metabolites formed in the colon. J. Nutr. Biochem. 19, 587–593. doi: 10.1016/j.jnutbio.2007.08.002
Wang N., Guo D., Tian X., Lin H., Li Y., Chen S., et al. (2016). Niacin receptor GPR109A inhibits insulin secretion and is down-regulated in type 2 diabetic islet beta-cells. Gen. Comp. Endocrinol. 227, 112932. doi: 10.1016/j.ecoenv.2021.112932
Wang X., Hu M., Gu H., Zhang L., Shang Y., Wang T., et al. (2020). Short-term exposure to norfloxacin induces oxidative stress, neurotoxicity and microbiota alteration in juvenile large yellow croaker Pseudosciaena crocea. Environ. pollut. 267, 115397. doi: 10.1016/j.envpol.2020.115397
Wang S., Li X., Zhang M., Jiang H., Wang R., Qian Y., et al. (2021b). Ammonia stress disrupts intestinal microbial community and amino acid metabolism of juvenile yellow catfish (Pelteobagrus fulvidraco). Ecotoxicol. Environ. Saf. 227, 112932. doi: 10.1016/j.ecoenv.2021.112932
Wang A., Meng D., Hao Q., Xia R., Zhang Q., Ran C., et al. (2022). Effect of supplementation of solid-state fermentation product of Bacillus subtilis HGcc-1 to high-fat diet on growth, hepatic lipid metabolism, epidermal mucus, gut and liver health and gut microbiota of zebrafish. Aquaculture 560, 738542. doi: 10.1016/j.aquaculture.2022.738542
Wang H. B., Wang P. Y., Wang X., Wan Y. L., Liu Y. C. (2012). Butyrate enhances intestinal epithelial barrier function via up-regulation of tight junction protein claudin-1 transcription. Dig. Dis. Sci. 57, 3126–3135. doi: 10.1007/s10620-012-2259-4
Wang W., Yang Q., Sun Z., Chen X., Yang C., Ma X. (2015). Editorial: advance of interactions between exogenous natural bioactive peptides and intestinal barrier and immune responses. Curr. Protein Pept. Sci. 16, 574–575. doi: 10.2174/138920371607150810124927
Wang E., Yuan Z., Wang K., Gao D., Liu Z., Liles M. R. (2019). Consumption of florfenicol-medicated feed alters the composition of the channel catfish intestinal microbiota including enriching the relative abundance of opportunistic pathogens. Aquaculture 501, 111–118. doi: 10.1016/j.aquaculture.2018.11.019
Wang A., Zhang Z., Ding Q., Yang Y., Bindelle J., Ran C., et al. (2021a). Intestinal Cetobacterium and acetate modify glucose homeostasis via parasympathetic activation in zebrafish. Gut Microbes 13, 1–15. doi: 10.1080/19490976.2021.1900996
Wang T., Zhang N., Yu X.-B., Qiao F., Chen L.-Q., Du Z.-Y., et al. (2021c). Inulin alleviates adverse metabolic syndrome and regulates intestinal microbiota composition in Nile tilapia (Oreochromis niloticus) fed with high-carbohydrate diet. Br. J. Nutr. 126, 161–171. doi: 10.1017/S000711452000402X
Wee C. L., Song E. Y., Johnson R. E., Ailani D., Randlett O., Kim J. Y., et al. (2019). A bidirectional network for appetite control in larval zebrafish. Elife 8, 1–37. doi: 10.7554/eLife.43775
Weitkunat K., Schumann S., Nickel D., Kappo K. A., Petzke K. J., Kipp A. P., et al. (2016). Importance of propionate for the repression of hepatic lipogenesis and improvement of insulin sensitivity in high-fat diet-induced obesity. Mol. Nutr. Food Res. 60, 2611–2621. doi: 10.1002/mnfr.201600305
Wen J., Mercado G. P., Volland A., Doden H. L., Lickwar C. R., Crooks T., et al. (2021). Fxr signaling and microbial metabolism of bile salts in the zebrafish intestine. Sci. Adv. 7, 1–19. doi: 10.1126/sciadv.abg1371
Wendelaar Bonga S. E. (1997). The stress response in fish. Physiol. Rev. 77, 591–625. doi: 10.1152/physrev.1997.77.3.591
Wentzel A. S., Petit J., van Veen W. G., Fink I. R., Scheer M. H., Piazzon M. C., et al. (2020). Transcriptome sequencing supports a conservation of macrophage polarization in fish. Sci. Rep. 10, 13470. doi: 10.1038/s41598-020-70248-y
Wessels A. G. (2022). Influence of the gut microbiome on feed intake of farm animals. Microorganisms 10. doi: 10.3390/microorganisms10071305
Wicks B. J., Joensen R., Tang Q., Randall D. J. (2002). Swimming and ammonia toxicity in salmonids: The effect of sub lethal ammonia exposure on the swimming performance of coho salmon and the acute toxicity of ammonia in swimming and resting rainbow trout. Aquat. Toxicol. 59, 55–69. doi: 10.1016/S0166-445X(01)00236-3
Wilkes Walburn J., Wemheuer B., Thomas T., Copeland E., O’Connor W., Booth M., et al. (2019). Diet and diet-associated bacteria shape early microbiome development in Yellowtail Kingfish (Seriola lalandi). Microb. Biotechnol. 12, 275–288. doi: 10.1111/1751-7915.13323
Witkowski M., Weeks T. L., Hazen S. L. (2020). Gut microbiota and cardiovascular disease. Circ. Res. 127, 553–570. doi: 10.1161/CIRCRESAHA.120.316242
Wu Z., Zhang Q., Lin Y., Hao J., Wang S., Zhang J., et al. (2021). Taxonomic and functional characteristics of the gill and gastrointestinal microbiota and its correlation with intestinal metabolites in NEW GIFT strain of farmed adult nile tilapia (Oreochromis niloticus). Microorganisms 9, 617. doi: 10.3390/microorganisms9030617
Xia Y., Lu M., Chen G., Cao J., Gao F., Wang M., et al. (2018b). Effects of dietary Lactobacillus rhamnosus JCM1136 and Lactococcus lactis subsp. lactis JCM5805 on the growth, intestinal microbiota, morphology, immune response and disease resistance of juvenile Nile tilapia, Oreochromis niloticus. Fish Shellfish Immunol. 76, 368–379. doi: 10.1016/j.fsi.2018.03.020
Xia J., Lu L., Jin C., Wang S., Zhou J., Ni Y., et al. (2018a). Effects of short term lead exposure on gut microbiota and hepatic metabolism in adult zebrafish. Comp. Biochem. Physiol. Part - C Toxicol. Pharmacol. 209, 209. doi: 10.1016/j.cbpc.2018.03.007
Xia Y., Yu E., Lu M., Xie J. (2020). Effects of probiotic supplementation on gut microbiota as well as metabolite profiles within Nile tilapia, Oreochromis niloticus. Aquaculture 527, 735428. doi: 10.1016/j.aquaculture.2020.735428
Xiong F., Chen S., Jakovlić I., Li W., Li M., Zou H., et al. (2022). The role of intestinal microbiota in regulating the metabolism of bile acids is conserved across vertebrates. Front. Microbiol. 13. doi: 10.3389/fmicb.2022.824611
Xiong J., Zhu J., Dai W., Dong C., Qiu Q., Li C. (2017). Integrating gut microbiota immaturity and disease-discriminatory taxa to diagnose the initiation and severity of shrimp disease. Environ. Microbiol. 19, 1490–1501. doi: 10.1111/1462-2920.13701
Xu C., Liu J., Gao J., Wu X., Cui C., Wei H., et al. (2020a). Combined soluble fiber-mediated intestinal microbiota improve insulin sensitivity of obese mice. Nutrients. 12, 351. doi: 10.3390/nu12020351
Xu C., Suo Y., Wang X., Qin J. G., Chen L., Li E. (2020b). Recovery from Hypersaline-Stress-Induced Immunity Damage and Intestinal-Microbiota Changes through Dietary β-glucan Supplementation in Nile tilapia (Oreochromis niloticus). Animals. 10, 2243. doi: 10.3390/ani10122243
Xu J., Xie S., Chi S., Zhang S., Cao J., Tan B. (2022). Short-term dietary antibiotics altered the intestinal microbiota and improved the lipid metabolism in hybrid grouper fed medium and high-lipid diets. Aquaculture 547, 737453. doi: 10.1016/j.aquaculture.2021.737453
Yan X., Chen Y., Dong X., Tan B., Liu H., Zhang S., et al. (2021). Ammonia Toxicity Induces Oxidative Stress, Inflammatory Response and Apoptosis in Hybrid Grouper (♀ Epinephelus fuscoguttatus × ♂ E. lanceolatu). Front. Mar. Sci. 8. doi: 10.3389/fmars.2021.667432
Yan W., Hamid N., Deng S., Jia P. P., Pei D. S. (2020). Individual and combined toxicogenetic effects of microplastics and heavy metals (Cd, Pb, and Zn) perturb gut microbiota homeostasis and gonadal development in marine medaka (Oryzias melastigma). J. Hazard. Mater. 397, 122795. doi: 10.1016/j.jhazmat.2020.122795
Yang H.-L., Sun Y.-Z., Hu X., Ye J.-D., Lu K.-L., Hu L.-H., et al. (2019). Bacillus pumilus SE5 originated PG and LTA tuned the intestinal TLRs/MyD88 signaling and microbiota in grouper (Epinephelus coioides). Fish Shellfish Immunol. 88, 266–271. doi: 10.1016/j.fsi.2019.03.005
Yang S., Zhang C., Xu W., Li D., Feng Y., Wu J., et al. (2022). Heat stress decreases intestinal physiological function and facilitates the proliferation of harmful intestinal microbiota in sturgeons. Front. Microbiol. 13. doi: 10.3389/fmicb.2022.755369
Yano J. M., Yu K., Donaldson G. P., Shastri G. G., Ann P., Ma L., et al. (2015). Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell 161, 264–276. doi: 10.1016/j.cell.2015.02.047
Yatsunenko T., Rey F. E., Manary M. J., Trehan I., Dominguez-Bello M. G., Contreras M., et al. (2012). Human gut microbiome viewed across age and geography. Nature 486, 222–227. doi: 10.1038/nature11053
Ye J. D., Wang K., Li F. D., Sun Y. Z. (2011). Single or combined effects of fructo- and mannan oligosaccharide supplements and Bacillus clausii on the growth, feed utilization, body composition, digestive enzyme activity, innate immune response and lipid metabolism of the Japanese flounder Paralichthys olivaceus. Aquac. Nutr. 17, e902–e911. doi: 10.1111/j.1365-2095.2011.00863.x
Yin Z., Liu Q., Liu Y., Gao S., He Y., Yao C., et al. (2021). Early Life Intervention Using Probiotic Clostridium butyricum Improves Intestinal Development, Immune Response, and Gut Microbiota in Large Yellow Croaker (Larimichthys crocea) Larvae. Front. Immunol. 12. doi: 10.3389/fimmu.2021.640767
Yoshimizu M., Kimura T. (1976). Study on the intestinal microflora of salmonids. Fish Pathol. 10, 243–259. doi: 10.3147/jsfp.10.243
Yu E. W., Gao L., Stastka P., Cheney M. C., Mahabamunuge J., Torres Soto M., et al. (2020). Fecal microbiota transplantation for the improvement of metabolism in obesity: The FMT-TRIM double-blind placebo-controlled pilot trial. PloS Med. 17, e1003051. doi: 10.1371/journal.pmed.1003051
Yu L., Qiao N., Li T., Yu R., Zhai Q., Tian F., et al. (2019). Dietary supplementation with probiotics regulates gut microbiota structure and function in Nile tilapia exposed to aluminum. PeerJ 7, e6963. doi: 10.7717/peerj.6963
Yukgehnaish K., Kumar P., Sivachandran P., Marimuthu K., Arshad A., Paray B. A., et al. (2020). Gut microbiota metagenomics in aquaculture: factors influencing gut microbiome and its physiological role in fish. Rev. Aquac. 12, 1903–1927. doi: 10.1111/raq.12416
Zhang L. S., Davies S. S. (2016). Microbial metabolism of dietary components to bioactive metabolites: Opportunities for new therapeutic interventions. Genome Med. 8, 46. doi: 10.1186/s13073-016-0296-x
Zhang Y., Ji T., Jiang Y., Zheng C., Yang H., Liu Q. (2022b). Long-term effects of three compound probiotics on water quality, growth performances, microbiota distributions and resistance to Aeromonas veronii in crucian carp Carassius auratus gibelio. Fish Shellfish Immunol. 120, 233–241. doi: 10.1016/j.fsi.2021.11.036
Zhang Y., Li Z., Kholodkevich S., Sharov A., Chen C., Feng Y., et al. (2020b). Effects of cadmium on intestinal histology and microbiota in freshwater crayfish (Procambarus clarkii). Chemosphere 242, 125105. doi: 10.1016/j.chemosphere.2019.125105
Zhang Z., Li D., Refaey M. M., Xu W. (2017). High Spatial and Temporal Variations of Microbial Community along the Southern Catfish Gastrointestinal Tract: Insights into Dynamic Food Digestion. Front. Microbiol. 8, 152099. doi: 10.3389/fmicb.2017.01531
Zhang F., Li D., Yang Y., Zhang H., Zhu J., Liu J., et al. (2022a). Combined effects of polystyrene microplastics and copper on antioxidant capacity, immune response and intestinal microbiota of Nile tilapia (Oreochromis niloticus). Sci. Total Environ. 808, 152099. doi: 10.1016/j.scitotenv.2021.152099
Zhang Y., Liang X., He S., Wang J., Li L., Zhang Z., et al. (2021). Metabolic responses of Chinese perch (Siniperca chuatsi) to different levels of dietary carbohydrate. Fish Physiol. Biochem. 47, 1449–1465. doi: 10.1007/s10695-021-00965-2
Zhang Y., Liu Y., Ma H., Sun M., Wang X., Jin S., et al. (2023c). Insufficient or excessive dietary carbohydrates affect gut health through change in gut microbiota and regulation of gene expression of gut epithelial cells in grass carp (Ctenopharyngodon idella). Fish Shellfish Immunol. 132, 108442. doi: 10.1016/j.fsi.2022.11.030
Zhang C., Liu H., Sun L., Wang Y., Chen X., Du J., et al. (2023a). An overview of host-derived molecules that interact with gut microbiota. iMeta 2, 1–21. doi: 10.1002/imt2.88
Zhang B., Yang H., Cai G., Nie Q., Sun Y. (2024). The interactions between the host immunity and intestinal microorganisms in fish. Appl. Microbiol. Biotechnol. 108, 1–14. doi: 10.1007/s00253-023-12934-1
Zhang L., Yang Z., Yang F., Wang G., Zeng M., Zhang Z., et al. (2023b). Gut microbiota of two invasive fishes respond differently to temperature. Front. Microbiol. 14. doi: 10.3389/fmicb.2023.1087777
Zhang J., Zhang H., Liu M., Lan Y., Sun H., Mai K., et al. (2020a). Short-Chain Fatty Acids Promote Intracellular Bactericidal Activity in Head Kidney Macrophages From Turbot (Scophthalmus maximus L.) via Hypoxia Inducible Factor-1α. Front. Immunol. 11. doi: 10.3389/fimmu.2020.615536
Zhang C., Zhang M., Pang X., Zhao Y., Wang L., Zhao L. (2012). Structural resilience of the gut microbiota in adult mice under high-fat dietary perturbations. ISME J. 6, 1848–1857. doi: 10.1038/ismej.2012.27
Zhao C., Guo G., Li Z., Chen J., Ren Y. (2024). Effects of probiotics (Bacillus coagulans) supplementation after antibiotic administration on growth, immunity, and intestinal microflora in turbot Scophthalmus maximus. Aquac. Int. 32, 1473–1491. doi: 10.1007/s10499-023-01225-1
Zhao C., Men X., Dang Y., Zhou Y., Ren Y. (2023). Probiotics mediate intestinal microbiome and microbiota-derived metabolites regulating the growth and immunity of rainbow trout (Oncorhynchus mykiss). Microbiol. Spectr. 11, e0398022. doi: 10.1128/spectrum.03980-22
Zhao R., Symonds J. E., Walker S. P., Steiner K., Carter C. G., Bowman J. P., et al. (2020a). Salinity and fish age affect the gut microbiota of farmed Chinook salmon (Oncorhynchus tshawytscha). Aquaculture 528, 735539. doi: 10.1016/j.aquaculture.2020.735539
Zhao W., Xie J. J., Fang H. H., Liu Y. J., Tian L. X., Niu J. (2020b). Effects of corn starch level on growth performance, antioxidant capacity, gut morphology and intestinal microflora of juvenile golden pompano, Trachinotus ovatus. Aquaculture 524, 735197. doi: 10.1016/j.aquaculture.2020.735197
Zhao J., Zhang X., Liu H., Brown M. A., Qiao S. (2019). Dietary protein and gut microbiota composition and function. Curr. Protein Pept. Sci. 20, 145–154. doi: 10.2174/1389203719666180514145437
Zheng L., Wang Z., Zhang B., Yan L., Wang P., Zhao C., et al. (2023). Effects of high dietary carbohydrate levels on growth performance, enzyme activities, expression of genes related to liver glucose metabolism, and the intestinal microbiota of lateolabrax maculatus juveniles. Fishes 8, 431. doi: 10.3390/fishes8090431
Zhong H., Hu Y., Yu F. (2021). A review on ghrelin and fish reproduction. Reprod. Breed. 1, 128–135. doi: 10.1016/j.repbre.2021.07.004
Zhou Y. L., He G. L., Jin T., Chen Y. J., Dai F. Y., Luo L., et al. (2021). High dietary starch impairs intestinal health and microbiota of largemouth bass, Micropterus salmoides. Aquaculture 534, 736261. doi: 10.1016/j.aquaculture.2020.736261
Zhou L., Wei J., Lin K., Gan L., Wang J., Sun J., et al. (2020a). Intestinal microbial profiling of grass carp (Ctenopharyngodon idellus) challenged with Aeromonas hydrophila. Aquaculture 524, 735292. doi: 10.1016/j.aquaculture.2020.735292
Zhou C., Yang S., Ka W., Gao P., Li Y., Long R., et al. (2022). Association of gut microbiota with metabolism in rainbow trout under acute heat stress. Front. Microbiol. 13. doi: 10.3389/fmicb.2022.846336
Keywords: gut microbiota, metabolism, fish diet, aquaculture, gut-brain axis, teleosts
Citation: Tolas I, Zhou Z, Zhang Z, Teame T, Olsen RE, Ringø E and Rønnestad I (2025) A fishy gut feeling – current knowledge on gut microbiota in teleosts. Front. Mar. Sci. 11:1495373. doi: 10.3389/fmars.2024.1495373
Received: 12 September 2024; Accepted: 05 December 2024;
Published: 10 January 2025.
Edited by:
Neil Ross McEwan, Scotland’s Rural College, United KingdomReviewed by:
Xiaodan Wang, East China Normal University, ChinaEric Amenyogbe, University of Environment and Sustainable Development, Ghana
Copyright © 2025 Tolas, Zhou, Zhang, Teame, Olsen, Ringø and Rønnestad. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ingvill Tolas, aW5ndmlsbC50b2xhc0BudG51Lm5v; Zhigang Zhou, emhvdXpoaWdhbmcwM0BjYWFzLmNu; Ivar Rønnestad, aXZhci5yb25uZXN0YWRAdWliLm5v