 Lourenço Ramos-Pinto1*
Lourenço Ramos-Pinto1* Raquel Marçal2
Raquel Marçal2 André Barreto3Adriana Laranjeira3
André Barreto3Adriana Laranjeira3 Marina Machado1Sérgio Fernández-Boo1Carla Teixeira1,4,5Joana Oliveira1,6
Marina Machado1Sérgio Fernández-Boo1Carla Teixeira1,4,5Joana Oliveira1,6 Ana Couto1,6
Ana Couto1,6 Jorge Dias4Sofia Guilherme2
Jorge Dias4Sofia Guilherme2 Ana C. S. Veríssimo7
Ana C. S. Veríssimo7 Diana C. G. A. Pinto7
Diana C. G. A. Pinto7 Mário Pacheco2Rui J. M. Rocha3
Mário Pacheco2Rui J. M. Rocha3 Benjamín Costas1,5*
Benjamín Costas1,5*- 1Centro Interdisciplinar de Investigação Marinha e Ambiental (CIIMAR), Universidade do Porto, Terminal de Cruzeiros do Porto de Leixões, Avenida General Norton de Matos, Matosinhos, Portugal
- 2Centre for Environmental and Marine Studies (CESAM) and Department of Biology, University of Aveiro, Aveiro, Portugal
- 3Riasearch Lda, Cais da Ribeira de Pardelhas n°21, Murtosa, Portugal
- 4SPAROS Lda, Área Empresarial de Marim, Lote C, Olhão, Portugal
- 5Instituto de Ciências Biomédicas Abel Salazar (ICBAS-UP), Universidade do Porto, Rua de Jorge Viterbo Ferreira n° 228, Porto, Portugal
- 6Department of Biology, Faculty of Sciences, University of Porto, Rua do Campo Alegre s/n, Porto, Portugal
- 7LAQV-REQUIMTE, Department of Chemistry, University of Aveiro, Aveiro, Portugal
The whiteleg shrimp, Penaeus vannamei, is a highly valued and globally produced crustacean species. However, the rising cost of shrimp feed, exacerbated by increasing cereal prices, prompts the exploration of cost-effective and sustainable formulations. This study investigates the potential of Salicornia ramosissima biomass by-product (the non-edible part) as a substitute for wheat meal in juvenile shrimp diets, aiming to create sustainable formulations. Particularly to assess the impact of incorporating S. ramosissima into shrimp aquafeeds on various aspects of shrimp development, including growth performance, survival, immune status, and oxidative status. A commercial-like diet was formulated and served as control, whereas four other diets contained S. ramosissima stems or a combination of leaves and seeds, both at inclusion levels of 5% and 10%. Shrimps were fed the experimental diets for 31 and 55 days, followed by a bacterial bath challenge test to gauge their immune response to pathogens. At the end of the feeding period, growth performance and survival rates remained consistent across all diets. However, shrimp fed diets with S. ramosissima consumed more feed to achieve similar weights of those fed the control diet, particularly in diets containing leaves and seeds at a 10% inclusion level, likely due to lower digestibility of dry matter, lipids, and energy. While S. ramosissima biomass inclusion did not affect shrimp weight, relative growth rate, or survival, it did lead to higher feed conversion ratios and feed intake. Additionally, S. ramosissima inclusion affected shrimps’ overall body composition, particularly moisture and ash content. S. ramosissima inclusion modulated antioxidant enzyme activity in the shrimp’s hepatopancreas, indicating potential health improvements. The observed gene expression changes related to antioxidant enzymes, points to an overall down-regulation with the inclusion of S. ramosissima. Despite challenges in feeding efficiency, the inclusion of S. ramosissima, especially stems, shows promise in reducing feed costs by utilizing a food agro-industrial by-products (non edible parts). Furthermore, S. ramosissima inclusion led to subtle changes in certain plasma humoral parameters. In conclusion, this study highlights the potential of this halophyte as a functional feed ingredient capable of enhancing shrimp’s antioxidant response, aligning with global resource optimization and sustainability initiatives.
1 Introduction
The increasing challenge of soil salinization is a significant constraint on agricultural productivity globally. About 7% of the world’s land area is salt-affected to some degree, and this trend is on the rise (Wicke et al., 2011). Glycophytes, which are the primary source of food, home remedies, and drugs, are sensitive to salt and suffer yield reductions even in mildly saline conditions. Halophytes, on the other hand, have been identified as a viable alternative in recent decades. These plants are adapted to grow in saline environments, such as coastal areas and salt marshes, and can tolerate high salt concentrations of 200 mM or more in their tissues, surviving in conditions that most other plants cannot. They make up about 2% of the world’s flora (Flowers and Colmer, 2008; Flowers and Muscolo, 2015). However, it is only recently that their potential has been recognized for crop production, as they do not require as much freshwater for irrigation and can tolerate high levels of salt and other environmental stresses (Ventura and Sagi, 2013). Halophytes have a variety of applications, including food, medicine, and biofuels. Species of the genus Salicornia are particularly noteworthy due to their commercial potential, with current large-scale cultivation in Europe, North America and middle East for human consumption as a vegetable, as well as attempts to produce oilseed and animal feed using seawater irrigation (Chaturvedi et al., 2021).
The increasing interest and widespread embrace of functional foods have paved the way for a fresh market opportunity for halophytes. This stems from the revelation of various bioactive compounds found in halophytes, which not only contribute to their nutritional value but also assist in specific bodily functions. Research indicates that Salicornia spp. possess numerous nutraceutical properties attributed to their phenolic content. They have been used traditionally in the treatment of conditions such as hepatitis, constipation, nephropathy, diarrhea, and have demonstrated antihyperglycemic and antihyperlipidemic activities. Some of the phenolic compounds (quercetin and isoquercetin) in Salicornia spp. are recognized as generally safe by the United States Food and Drug Administration (USFDA) and can be administered in specific concentrations. The concentrations of phenolic compounds in Salicornia spp. can vary due to factors like biomass maturity, pre-treatment methods, and extraction techniques. In terms of antiviral properties, phenolic compounds found in Salicornia spp. have demonstrated inhibitory effects against viruses such as H1N1, HBV, HCV, and HIV-1. These effects are attributed to the compounds’ ability to interact with viral and host proteins and enzymes, preventing viral replication and infectivity (Fredsgaard et al., 2023). Salicornia species are rich in minerals (Barreira et al., 2017), flavonoids and polyphenols (Essaidi et al., 2013; Silva et al., 2021; Limongelli et al., 2022). Polyphenols have been found to be strong antioxidants that can neutralize free radicals by donating an electron or hydrogen atom (Tsao, 2010). Flavonoids possess antioxidant activity and function as a free radical scavenger and a chelator of transition metal, such as iron, a key metal in the generation of ROS through the Fenton reaction (Murota and Terao, 2003) and the quenching of singlet-oxygen atom (Mukai et al., 2005). The chemical research on various Salicornia spp., such as S. europaea, S. arthrocnemum, S. bigelovii, S. perennis, and S. disarticulata, has demonstrated their significant anti-oxidative properties. This finding supports their traditional use, as they all contain phenolic compounds (Rhee et al., 2009). Hence, functional feeds – such as diets rich in Salicornia – may play a crucial role in enhancing the overall health and performance of aquatic organisms in aquaculture, particularly in species like shrimp. By incorporating functional ingredients into their diets, it is possible to boost their immune system and improve antioxidant functions. This could help the shrimp to better combat diseases and oxidative stress, ultimately leading to improved growth, survival rates, and overall welfare. This is particularly vital in aquaculture, where maintaining the health and vitality of the stock is essential for successful and sustainable production.
Salicornia ramosissima is considered an alternative to salt and its fresh and tender green tips (leaves) are used as food ingredients for human diet, mainly in salads, canned or gourmet gastronomy (Ferreira et al., 2018). Recent research has shown that Salicornia spp. can grow in constructed wetlands and can effectively take up nutrients, such as nitrogen and phosphorus, from wastewater into its tissues (Ventura and Sagi, 2013)
Salicornia, particularly S. bigelovii and S. ramosissima, has emerged as a promising feed ingredient in aquaculture, supported by some highlighting its nutritional benefits. Ríos-Durán et al. (2013) demonstrated that S. bigelovii seed meal can be used as a source of protein in fish feed, while the health-modulating potential of Salicornia spp was demonstrated in fish (Marçal et al., 2023; Barreto et al., 2024) and farmed sheep (Taghipour et al., 2021; Nasrabadi et al., 2022), suggesting broader applicability across various species. Together, these findings emphasize the potential of Salicornia as a valuable feed source in the aquaculture industry. However, as the plant matures, it undergoes lignification and accumulates high levels of salt, rendering it less palatable and suitable for consumption. This study aims to investigate the underutilized, non-edible parts of S. ramosissima.
The whiteleg shrimp is the most farmed animal species globally in terms of the number of individuals, with an estimated 300-620 billion produced annually (Waldhorn and Autric, 2022). This industry generates over 33 billion USD per year (FAO, 2022). To sustain its rapid growth, the demand for aquafeeds, which typically comprise over 50% of production costs, has increased significantly. These feeds often rely on conventional ingredients like fish meal, soybean meal, and wheat meal, raising concerns about their sustainability (FAO, 2022).
Hence, the success of shrimp farming, and aquaculture in general, depends on the emergence of novel ingredients that can make formulated diets more economical and reduce the industry’s ecological footprint.
In summary, this study aimed to explore the viability of integrating S. ramosissima biomass into inert diets for juvenile whiteleg shrimp. Additionally, it sought to examine the potential advantages of this inclusion as a functional feed, potentially leading to positive impacts on health and welfare. The assessment encompassed parameters such as growth performance, survival rate, overall health, and oxidative status of the shrimp.
2 Materials and methods
2.1 Diets formulation
Plants were harvested in September 2019, when signs of lignification and flowering were evident, to simulate the stage of production when S. ramosissima is considered inadequate for human consumption. These were then dried in mesh mats that allowed air circulation, to prevent any degradation promoted by humidity. Plant separation occurred naturally in the drying process. Five experimental diets were evaluated in quintuplicates. A control diet (CTRL) was formulated to meet the nutritional requirements of juvenile whiteleg shrimp containing 32.5% of soybean meal, 25% of wheat meal and 15% of fish meal as main protein sources, whereas a blend of 6% of rice bran oil, 2% soybean oil and 1% fish oil was regarded as main lipid source. On the remaining treatments, four experimental variants based on the CTRL were used, differing only in the ingredient formulation by replacing wheat meal by: 1) S. ramosissima stems in 5% (SS5) and 10% (SS10) inclusion levels; and 2) S. ramosissima leaves and seeds in 5% (SL5) and 10% (SL10) inclusion levels. The term ‘stems’ in this context refers to the primary, woody structures of the plant, while leaves and seeds was considered the the softer part of the plant. These components typically have a higher lignin content, which contributes to their rigidity and indicates they are not intended for human consumption. S. ramosissima biomass was obtained in Praia da Areia Branca, Torreira, Portugal (40°46’22.1”N 8°39’29.5”W). The experimental diets composition and proximate analysis can be seen in Tables 1, 2, respectively.
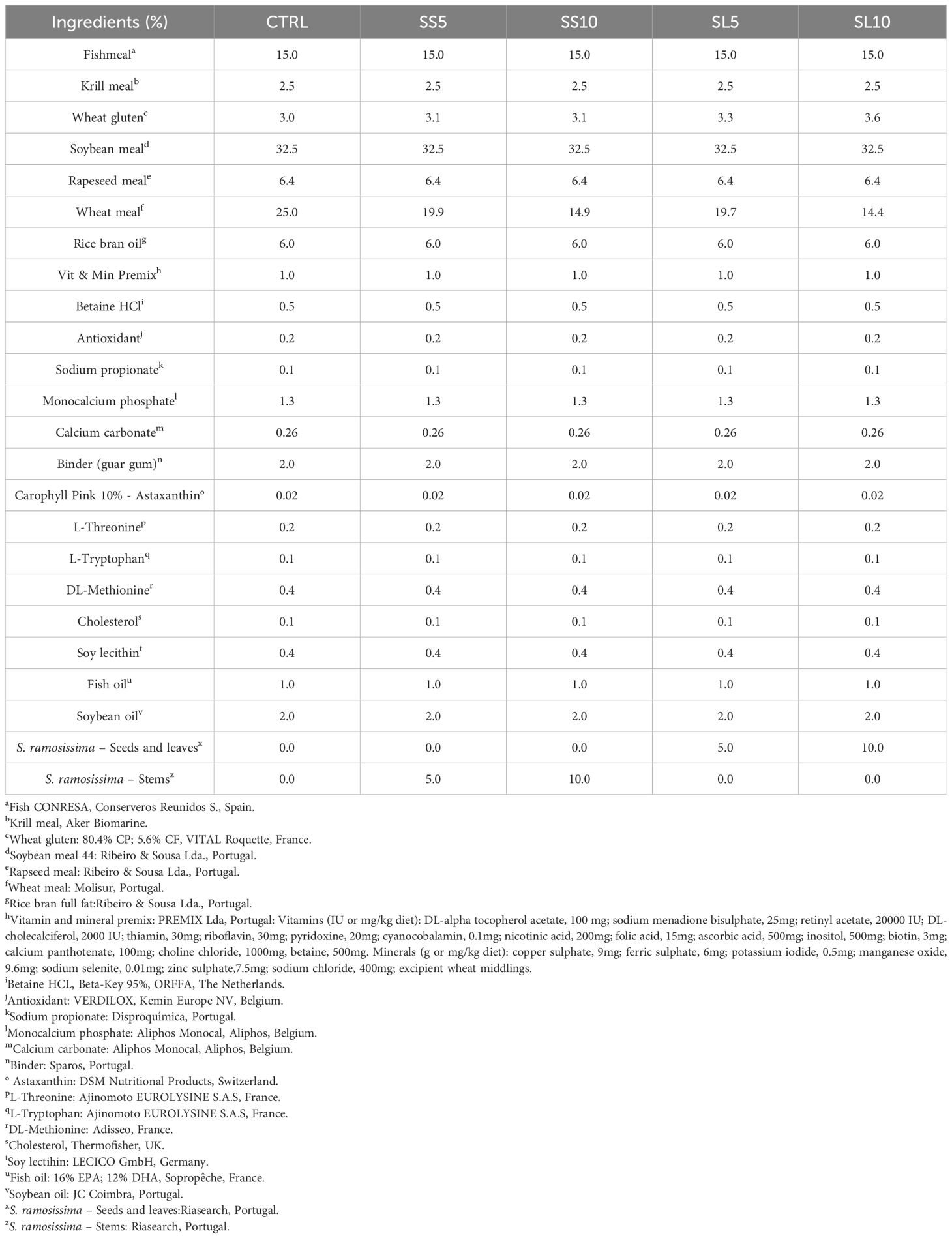
Table 1. Dietary composition of the experimental diets used to culture juvenile whiteleg shrimp (P. vannamei) for 55 days.
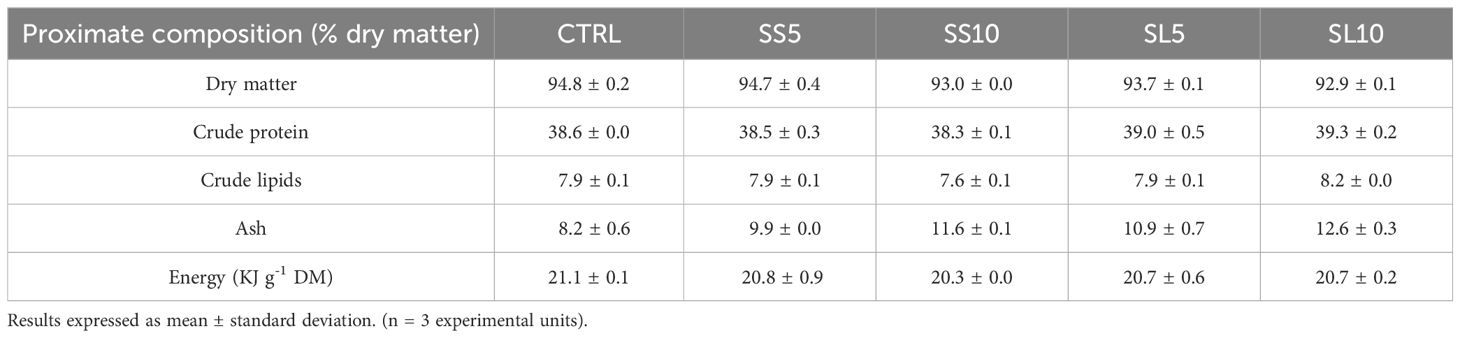
Table 2. Proximate composition of the experimental diets used to culture juvenile whiteleg shrimp (P. vannamei) for 55 days.
Diets were manufactured by extrusion at Sparos Lda (Olhão) facilities. All powder ingredients, including Salicornia, were mixed accordingly to the target formulation in a double-helix mixer (model 500 L, TGC Extrusion) and ground (below 400 µm) in a micropulveriser hammer mill (model SH1, Hosokawa-Alpine). Diets (pellet size: 1.5 and 2.0 mm) were manufactured with a twin-screw extruder (model BC45, Clextral) with a screw diameter of 55 mm. Extrusion conditions: feeder rate (80 kg/h), screw speed (255 rpm), water addition in barrel 1 (340 ml/min), temperature barrel 1 (36 °C) and temperature barrel 3 (110–112 °C). Extruded pellets were dried in a vibrating fluid bed dryer (model DR100, TGC Extrusion). After cooling, oils were added by vacuum coating (model PG-10VCLAB, Dinnissen). Coating conditions were pressure (700 mbar), spraying time under vacuum (approximately 90 s) and return to atmospheric pressure (120 s). After coating, diets were packed in sealed plastic buckets and stored at room temperature.
2.2 Shrimp rearing conditions and samplings
Whiteleg shrimp post-larvae (PL16), originated from Sea Products Development (Texas, USA), were reared for 55 days in standard conditions at Riasearch Lda. facilities (Murtosa, Portugal). After reaching approximately 6 g of body weight, shrimp were weighed individually and randomly distributed to 25 tanks with 200 L, part of a 30 m3 clear recirculating aquaculture system (RAS). Each tank was stocked with 55 individuals. Water renewal in each tank was kept at 1 renewal per hour. Water parameters were measured daily using commercial probes. During the experimental period, temperature was maintained at 28.1 ± 0.5 °C, dissolved oxygen at 5.9 ± 0.3 mg L-1, salinity at 19.4 ± 0.1 g L-1, pH at 7.6 ± 0.1, NH3 at 0.0 ± 0.0 mg L-1 and NO2 at 0.1 ± 0.0 mg L-1.
Shrimp were kept under a 12 hours light : 12 hours dark photoperiod and given 4 meals per day. Initial feed amounts (5% DW feed/g shrimp FW) were established according to Riasearch feeding tables for whiteleg shrimp juveniles. Afterwards, amounts were adjusted daily according to the feed wasted the day before, in increments or decrements of 0, 10, 20 or 30%. Feed size was 1.5 mm for the first 20 days and 2 mm for the remaining feeding period.
At the middle (Day 31) and end of the experiment (Day 55) whiteleg shrimp from each tank were weighed to determine mean body weight, specific growth rate (SGR), feed intake, feed conversion ratio (FCR) and survival. Additionally, 5 shrimp per tank were sampled for whole-body composition analysis at the end of the experiment. At the indicated times, shrimps were sorted and euthanized for tissue sampling. From three individuals per tank (15 per experimental group), fresh haemolymph was collected for haemocytes total counting and plasma for the assessment on innate immune parameters. Hepatopancreas was also collected for analysis of oxidative status and gene expression analysis of key immune related genes through RT-qPCR (Table 3A).
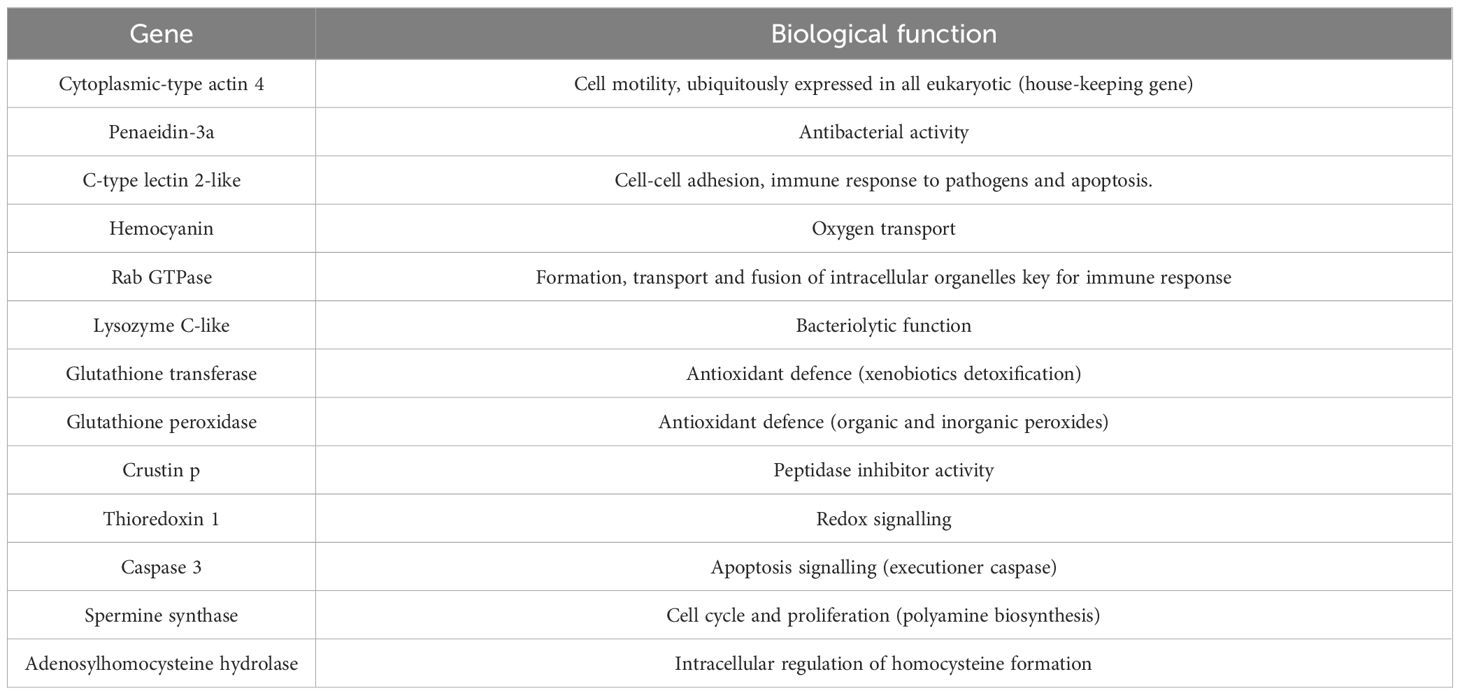
Table 3A. Immune-related genes analysed by real-time quantitative PCR and biological function.
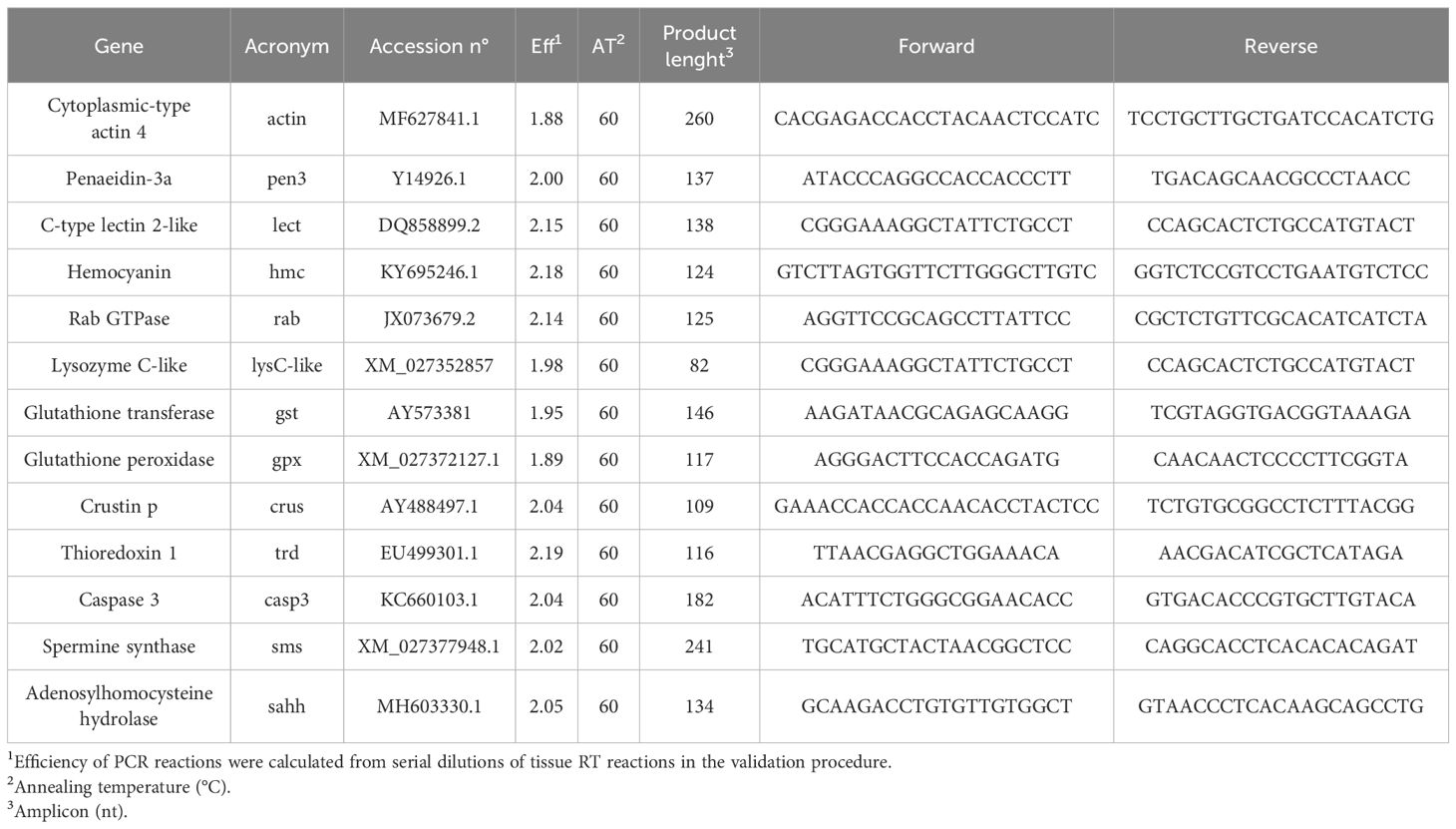
Table 3B. Forward and reverse primers for real-time quantitative PCR.
With the aim of assessing the modulatory roles of specific phytochemicals in the inflammatory processes, the inflammatory response was further studied at the end the 55 days feeding period. Twenty-five shrimps were re-allocated in a similar recirculation system where the diets were established in triplicate tanks and the individuals were submitted to a challenge bath test in 20 L tanks with Vibrio parahaemolyticus (2 x 106 CFU mL-1) for 1 h. After that, shrimp were fed following the procedures previously described for the growth trial and mortality recorded for one week. Moreover, 5 individuals from each tank (15 per group) were sampled for haemolymph and hepatopancreas at 4, 24 and 48 hours after the challenge period in order to assess cell proliferation and plasma immune mechanisms.
2.3 Bromatological analysis
Bromatological analysis of the experimental diets was performed following the Association of Official Analytical Chemists procedures (AOAC, 2000). Briefly, dry matter was determined by drying samples at 105 °C in an oven until constant weight; ash, by incineration at 450 °C for 16 h in a muffle furnace; crude protein content (N x 6.25), by the Kjeldahl method after acid digestion using a Kjeltec digestion and distillation unit; lipid content, by petroleum ether extraction (Soxtec HT System) and gross energy, by direct combustion in an adiabatic bomb calorimeter (PARR Instruments, Moline, IL, USA; PARR model 1261).
2.4 Chemical profile of shrimp carcasses
2.4.1 Pre-treatment of the samples
The samples were stored at -80 °C until starting the pre-treatment of the samples. The pre-treatment consisted of separating the muscle from the exoskeleton. A pool was made for each group. Then the muscle was cut into very small pieces and placed in an oven at 35 °C, to dry up to constant weight. Finally, the samples taken from the oven were macerated, using a mill, and stored.
2.4.2 Extracts preparation
Nearly 2.5 g of dried biomass of the sample were extracted with hexane (in a proportion of 1 g: 15 mL), at room temperature with stirring for 72 h. After the first extraction, the solvent is renewed and the extraction continues for another 72 h. The solvent from the combined hexane extractions was evaporated to dryness (Magni et al., 2023).
2.4.3 Trimethylsilyl derivatives preparation
Before analysis by GC-MS, samples were derivatized according to the known methodology and usually used in our department with slight changes (Faustino et al., 2017). To 10 mg of extract, 125 μL of pyridine, 125 μL of N, O-bis(trimethylsilyl)trifluoroacetamide (BSTFA), and 25 μL of trimethylchlorosilane (TMSCl) were added. 200 μL of 1 mg mL-1 of hexatriacontane (internal standard) and 525 μL of dichloromethane were added, making a total volume of 1mL. The mixtures were maintained at 70 °C for 30 minutes and then were transferred to vial, with needles and syringes. The samples were injected into the GC-MS apparatus. All extracts were derivatized in triplicate and injected in duplicate.
2.4.4 Gas chromatography-mass spectrometry analysis
GC–MS analysis of each silylated sample was performed using a GC–MS QP2010 Ultra Shimadzu (University of Aveiro, Portugal) equipped with a ZB-5ms J & W capillary column (30 m x 0.25 mm x 0.25 μm) as described in Ferreira et al. (2023). Samples were injected with a split ratio of 1:10 and helium as carrier gas with a flux of 1.19 mL/min. The injector temperature was at 320 °C and the transfer-line temperature was at 200 °C. The temperature of the column was maintained at 70 °C for 5 mins and then increased, first at 4 °C min-1. until 250 °C, followed by an increase of 2 °C min-1. until 300 °C, which was maintained for 5 min. The mass spectrometer was operated in the electronic impact (EI) mode with an energy of 0.1 kV, and data were collected at a rate of 1scan/s over a range of m/z 50–1000. The performed chromatography lasted 80 min and the standards were analysed separately by GC–MS under the same chromatographic conditions. The compounds were identified based on a direct comparison with the mass spectra database libraries (NIST14 Mass Spectra and WILEY Registry TM of Mass Spectra Data).
2.5 Tissue collection
Haemolymph was extracted from the ventral part of the first abdominal segment with an insulin syringe containing 200 µL of anticoagulant solution (27 mM Trisodium citrate, 385 mM sodium chloride, 115 mM glucose in mQ water, pH 7.5).
Immediately after extraction, haemolymph was diluted in anticoagulant solution 1:1 (v/v). 20 µL, 50 µL and 200 µL of haemolymph were used for oxyhemocyanin determination, total haemocyte counting and respiratory burst analysis respectively. The remaining haemolymph was then centrifuged at 2500 rpm, 10 min at 4 °C and plasma was transferred to new 1.5 mL Eppendorf and frozen immediately in liquid nitrogen.
Hepatopancreas was collected and cut in two longitudinal pieces, then one piece was placed in 1.5 mL tube with 1 mL of RNAlater (Sigma). Samples were then maintained 24 hours at 4 °C and then at -20 °C until RNA extraction.
For the oxidant status assessment, shrimp hepatopancreas was collected, immediately frozen in liquid nitrogen, and then stored in -80 °C until further analyses.
2.6 Haemocytes count
Briefly, 50 µL of 1:1 diluted haemolymph with anticoagulant solution were transferred to a new 1.5 mL tube with 50 µL of formalin according to Bautista-Covarrubias et al. (2014) to a final dilution of 1:4. Samples were counted in a Neubauer chamber under a light microscope at 400x.
2.7 Haemolymph humoral parameters
2.7.1 Oxyhemocyanin
For hemocyanin measurements, 20 µL hemolymph was immediately diluted with 180 µL distilled water. Then, 100 µL were transferred to a microplate by duplicate and and absorbance was read at 335 nm using a Synergy HT microplate reader (Biotek). Hemocyanin concentration was calculated using an extinction coefficient of 17.26 calculated on the basis of the 74000 Da functional subunit. Onyhemocyanin was measured according to the Pascual et al. (2003) method with some adaptations to microplates.
2.7.2 Respiratory burst
Intracellular production of the super oxide anion in haemocytes was quantified using the NBT (Nitroblue Tetrazolium) reduction to formazan was measured according to the Fujiki and Yano (1997) method with some adaptations. Briefly, 100 µL of hemolymph diluted in 500 µL of cacodilate buffer solution (10 mM cacodylate, 10 mM CaCl2, pH 7.0) was placed in a well of microplate (for triplicate) and centrifuged at 800 × g for 10 min. Plasma was removed and hemocytes washed with 100 μL Hank’s solution. Next, 100 μL of Vibrio anguillarum inactivated bacteria at 106 cells per mL was added and incubated for 2 h at room temperature. The bacteria solution was then removed and haemocytes washed three times with 100 μL Hank’s solution and stained for 30 min with NBT solution (0.3%) at room temperature. After fixed, haemocytes were washed three times with 100 μL methanol (70%) and dried for 5 min. Formazan was dissolved with 120 μL KOH and 140 μL DMSO, and absorbance was read at 630 nm using a Synergy HT microplate reader (Biotek).
2.8 Plasma immune parameters
Hemolymph which was not used for respiratory burst, oxyhemocyanin determination and THC was centrifuged at 2500 × rpm for 10 min at 4 °C. Plasma was placed in a new tube and hemocytes were discarded. Protein concentration in the plasma was determined using a DeNovix DS-11 spectrophotometer. Two μL of plasma were used to measure protein concentration within the instrument’s detection range. The absorbance of the plasma was measured at 280 nm, and the protein concentration was calculated using the Beer-Lambert law and the instrument’s built-in software.
Briefly, pro-phenoloxidase activity was measured as follows: 25 μL of plasma were incubated for 30 min at 25 °C with 100 μL trypsin (1 mg mL-1). Then, 100 μL L-DOPA (3 mg mL-1) were added and absorbance was measured each minute during 5 minutes at 490 nm in a Synergy HT microplate reader. Results were calculated using the Beer-Lambert law by means of the molar extinction coefficient of the L-DOPA (3700) according to Ji et al. (2009). Results are expressed as units of pro-phenoloxidase per mL of plasma.
Lysozyme activity was measured using a turbidimetric assay according to Costas et al. (2011). Lyophilized hen egg white lysozyme (Sigma-Aldrich) was serially diluted in sodium phosphate buffer (0.05 M, pH 6.2) and used to develop a standard curve. The amount of lysozyme in the sample was calculated using the formula of the standard curve.
The nitric oxide production was assayed in plasma by the Griess reaction. This reaction quantifies the nitrite content of the sample of 25 µL of plasma by adding 100 µL of 1% sulfanilamide (Sigma-Aldrich) in 2.5% of phosphoric acid to each well, followed by 100 µL of 0.1% N-naphthyl-ethylenediamine (Sigma) in 2.5% phosphoric acid. After 10 min of incubation at room temperature, the optical density was determined in a Synergy HT microplate reader (Biotek) at 540 nm. The concentration of nitrite in the sample was determined from standard curves generated using known concentrations of sodium nitrite.
Antiprotease activity was quantified using the azocasein hydrolysis assay according to Ramos-Pinto et al. (2019) with some modifications. Briefly, 20 μl of plasma were added to a microtube with 10 µL of trypsin (5 mg mL-1). After 10 minutes incubation, 90 µL of PBS and 125 µL of azocasein at 2% in a 100 mM ammonium bicarbonate buffer were added and incubate for 1 hour at 25 °C. The reaction was stopped by adding 250 µL of 10% trichloro acetic acid (TCA) and the mixture was centrifuged (10,000 × g, 5 min). Then, 100 µL of the supernatant solution were transferred to a 96-well plate in duplicate containing 100 μL well−1 of 1N NaOH. The optical density (OD) was read at 450 nm using a Synergy HT microplate reader (Biotek). Plasma was replaced by PBS as reference sample (100% of antiprotease activity), and sample with 20 µL of PBS was used as blank (0% activity). Results are expressed as the ability to suppress the trypsin activity against the reference sample.
Protease activity was quantified using the azocasein hydrolysis assay according to Ramos-Pinto et al. (2019) with some modifications. Briefly, 20 μl of plasma were incubated with 90 µL of PBS and 125 µL of azocasein at 2% in a 100 mM ammonium bicarbonate buffer for 24 hours at 24 °C in microtubes with continuous shaking, protected from direct light. The reaction was stopped by adding 250 µL of 10% TCA and the mixture was centrifuged (6.000 × g, 5 min). Then, 100 µL of the supernatant solution were transferred to a 96-well plate in duplicate containing 100 μL well−1 of 1N NaOH. The OD was read at 450 nm using a Synergy HT microplate reader (Biotek). Plasma was replaced by PBS as reference sample (100% of protease activity), and sample with 20 µL of PBS was used as blank (0% activity). Results are expressed as the ability to suppress the trypsin activity against the reference sample.
2.9 Hepatopancreas oxidant status
All measurements were carried out in a SpectraMax 190 microplate reader, at 25 °C. Shrimp hepatopancreas were homogenised using a Potter–Elvehjem homogenizer, in chilled phosphate buffer (0.1 M, pH 7.4) in a 1:11 ratio [tissue mass (mg):buffer volume (mL)]. The resulting homogenate was then divided into two aliquots, i.e. for lipid peroxidation (LPO) measurement and post-mitochondrial supernatant (PMS) preparation. The PMS was obtained by centrifugation in a refrigerated centrifuge (Eppendorf 5415R) at 13400 g for 20 min at 4 °C. Aliquots of PMS were then divided into microtubes and stored at -80 °C until further analyses. Superoxide dismutase (SOD), catalase (CAT), and glutathione peroxidase (GPx) activities as well as lipid peroxidation (LPO) levels were measured.
SOD activity was assayed in PMS with a Ransod kit (Randox Laboratories Ltd., UK). The method employs xanthine and xanthine oxidase to generate superoxide radicals, which react with 2-(4-iodo- phenyl)-3-(4-nitrophenol)-5-phenyltetrazolium chloride (INT) to form a red formazan dye determined at 505 nm. SOD activity was then measured by the degree of inhibition of this reaction, considering that one unit of SOD causes a 50% inhibition of the rate of reduction of INT, under the conditions of the assay. Results were expressed as SOD unit’s mg protein-1.
CAT activity was assayed in PMS by the method of (Clairborne, 1985), with slight modifications. Briefly, the assay mixture consisted of 0.190 mL phosphate buffer (0.05 M, pH 7.0) with hydrogen peroxide (H2O2; 0.010 M) and 0.010 mL of PMS, in a final volume of 0.2 mL. Change in absorbance was measured in appropriated UV transparent microplates (UV-Star® flat-bottom microplates, Greiner Bio-One GmbH, Germany), recorded at 240 nm and CAT activity was calculated in terms of μmol H2O2 consumed min-1 mg-1 protein using a molar extinction coefficient (ϵ) of 43.5 M-1 cm-1.
GPx activity was determined in PMS according to the method described by Mohandas et al. (1984) and modified by Athar and Iqbal (1998). The assay mixture consisted of 0.09 mL phosphate buffer (0.05 M, pH 7.0), 0.03 mL ethylenediaminetetraacetic acid (EDTA; 0.010 M), 0.03 mL sodium azide (0.010 M), 0.03 mL glutathione reductase (2.4 U mL-1), 0.03 mL reduced glutathione (GSH; 0.010 M), 0.03 mL nicotinamide adenine dinucleotide phosphate-oxidase (NADPH; 0.0015 M), 0.03 mL H2O2 (0.0025 M) and 0.03 mL of PMS in a total volume of 0.3 mL. Oxidation of NADPH to NADP+ was recorded at 340 nm in a SpectraMax 190 microplate reader and GPx activity was calculated in terms of nmol NADPH oxidized min-1 mg protein-1 using a ϵ of 6.22 × 103 M-1 cm-1.
As an estimation of LPO, TBARS quantification was carried out in the previously prepared homogenate according to the procedure of (Bird and Draper, 1984) and adapted by (Wilhelm Filho et al., 2001a, 2001b) and Torres et al. (2002). Briefly, 0.005 mL of butylatedhydroxytoluene (BHT; 4% in methanol) and 0.045 mL of phosphate buffer (0.05 M, pH 7.4) were added to 0.05 mL of homogenate and mixed well to prevent oxidation. To this aliquot, 0.250 mL of TCA (12%), 0.225 mL of Tris–HCl (0.060 M, pH 7.4 and 0.0001 M DTPA) and 0.250 mL of thiobarbituric acid (TBA; 0.73%) were added and well mixed. This mixture was heated for 1 h in a water bath set at 100 °C and then cooled to room temperature, and centrifuged at 15700 g for 5 min. The absorbance of each sample was measured at 535 nm in a SpectraMax 190 microplate reader. The rate of LPO was expressed in nmol of thiobarbituric acid reactive substances (TBARS) formed mg protein-1 using a ϵ of 1.56 × 105 M-1 cm-1.
2.10 Hepatopancreas gene expression
In a first approach, cDNA was only isolated from the hepatopancreas of shrimps sampled at 31 and 55 days fed the experimental diets. This procedure followed Machado et al. (2023) with some modifications. Briefly, hepatopancreas total RNA isolation and DNase treatment (NZY Total RNA isolation kit, MB13402, NZYTech, Portugal) and first-strand cDNA synthesis (NZY First-strand cDNA synthesis kit, MB125, NZYTech, Portugal) were performed according to manufacture guidelines. Primers design and efficiency values and quantitative PCR assays were performed. Efficiency of each primer was calculated according to a series dilution of hepatopancreas cDNA pool. Primers efficiency and DNA amplification was carried out using the CFX384 Touch Real-Time PCR Detection System with specific primers for genes that have been selected for their involvement in immune responses (Table 3A). Accession number, efficiency values, annealing temperature, product length and primers sequences are presented in Table 3B. Melting curve analysis was also performed to verify that no primer dimers were amplified. The standard cycling conditions were 95 °C initial denaturation for 10 min, followed by 40 cycles of 94 °C denaturation for 30 s, primer annealing temperature for 30 s and 72 °C extension for 30 s. All reactions were carried out as technical duplicates. The expression of the target genes was normalised using the expression of whiteleg shrimp cytoplasmic-type actin 4 (actin).
2.11 Bacterial challenge test
For the bacterial challenge, Vibrio parahaemolyticus, kindly provided by Prof. Miguel Ángel Moríñigo (University of Malaga), was used. Bacteria was routinely cultured at 25 °C in tryptic soy broth (TSB) or tryptic soy agar (TSA) supplemented with NaCl to a final concentration of 2% (w/v) and stored at -80 °C in TSB 2% NaCl with 15% (v/v) glycerol. To prepare the inoculum for the challenge bath, stock bacteria was cultured overnight at 25 °C in TSB 2% NaCl and exponentially growing bacteria were collected and adjusted to a final concentration of 2 × 108 colony forming units (cfu) mL-1, as confirmed by plating the resulting culture on TSA – 2% NaCl plates and counting of cfu. The recirculation water system was stopped, and the water volume of each tank was lowered to a final volume of 10 L. Shrimp were inoculated with the bacteria with strong aeration for 1 h. Afterwards, the rearing water was changed three times and the recirculation system was re-established.
2.12 Data analysis
2.12.1 Growth performance
Specific growth rate (SGR, % weight day-1) was calculated as: SGR = (eg – 1) × 100, where g = (lnWf – lnWi) × t–1. Wf and Wi correspond to the final and initial weights, respectively. Feed conversion ratio (FCR) was calculated as: FCR = (Fi/Wg), where Fi corresponds to feed intake (g) and Wg to the mean weight gain (g). Survival was expressed as percentage and calculated as: S = (Sf/Si) × 100, where Si and Sf correspond to the initial and final number of individuals in the tanks, respectively. Differences in growth performance, survival, whole-body composition, between dietary treatments were evaluated using One-way ANOVA’s, followed by Tukey HSD multiple comparison tests. Kruskal-Wallis one-way analysis of variance tests, followed by Wilcoxon pairwise comparison tests were used when data did not comply with the One-way ANOVA’s assumptions.
2.12.2 Humoral, oxidative stress and chemical profile
Differences in oxidative stress biomarkers, immune parameters and gene expression and chemical profile between dietary treatments were evaluated using One-way ANOVA’s, followed by Tukey HSD multiple comparison tests. Kruskal-Wallis one-way analysis of variance tests, followed by Wilcoxon pairwise comparison tests were used when data did not comply with the One-way ANOVA’s assumptions.
To evaluate the activation of the inflammatory mechanisms, the sampling point 55 days was used as time 0h during the time-course data analysis, as they represent unstimulated animals prior to infection. Differences between dietary treatments during the time course period were evaluated using Two-way ANOVA’s, followed by Tukey HSD multiple comparison tests.
In an attempt to discriminate and classify the dietary treatments along the experimental sampling points, a multivariate canonical discriminant analysis using XLSTAT (Addinsoft, New York, USA), was performed on the dataset to evaluate the linear combinations of the original variables that will best separate the groups (discriminant functions). Multivariate linear discriminant analyses (LDA) were performed for gene expression data to evaluate how it contributed to the dissociation of the diets in the discriminant functions generated. A MANOVA was performed to assess discriminatory significance using Wilk’s λ test, after checking data compliance to the statistics assumptions. The distance between group centroids was measured by Mahalanobis distance and its significance inferred by One-way ANOVA’s statistics. Results were expressed as means ± standard deviation (SD). In results expressed as percentage, an arcsine transformation was performed prior to any statistical test: T = ASIN (SQRT (value/100)). The significance level considered was p < 0.05 for all tests performed.
3 Results
3.1 Feeding trial
3.1.1 Growth performance
At day 31 of feeding, no significant differences between treatments were found in final body weight, SGR and survival values. However, whiteleg shrimps fed SS10 and SL10 diets had a significantly higher FCR values than those fed the CTRL diet, while shrimp fed SL10 had significantly higher FCR than those fed the remaining diets, excluding SS10. Feed intake was also higher in whiteleg shrimp fed the experimental diets rich in S. ramosissima biomasses than those fed CTRL, particularly shrimp fed diet SL10 had significantly higher feed intake values than those fed the remaining diets, excluding SS10 (Table 4).
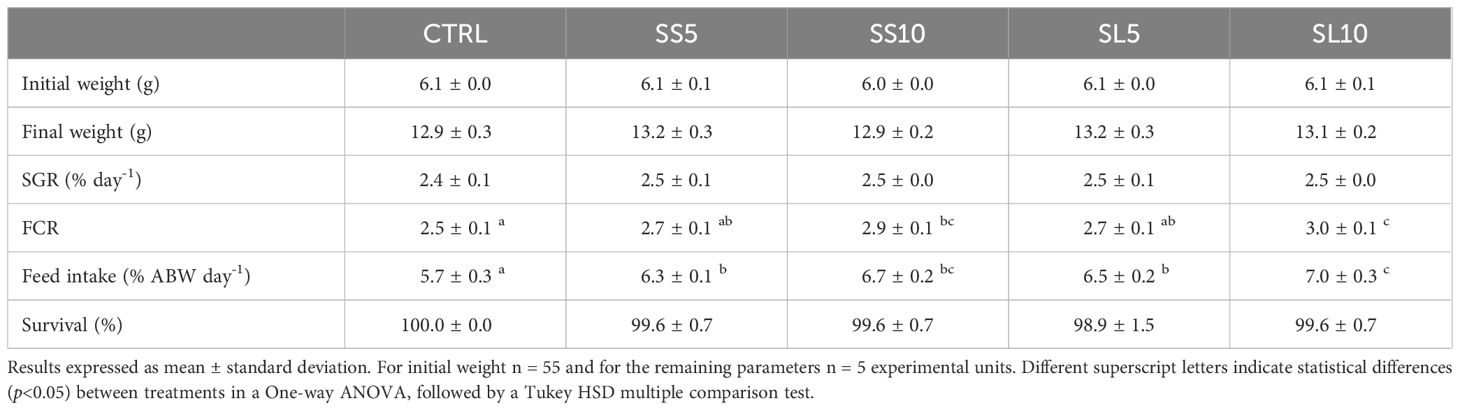
Table 4. Initial and final weight, specific growth rate (SGR), feed conversion ratio (FCR), feed intake and survival of juvenile whiteleg shrimp (P. vannamei) fed the experimental diets for 31 days.
Moreover, no significant differences between treatments were observed in final body weight, SGR and survival values after 31 days of feeding. Still, whiteleg shrimps fed the CTRL diet had significantly lower FCR and feed intake values than the experimental diets where S. ramosissima biomasses were included, while shrimp fed SL10 had significantly higher FCR and feed intake values than those fed the remaining diets (Table 5).
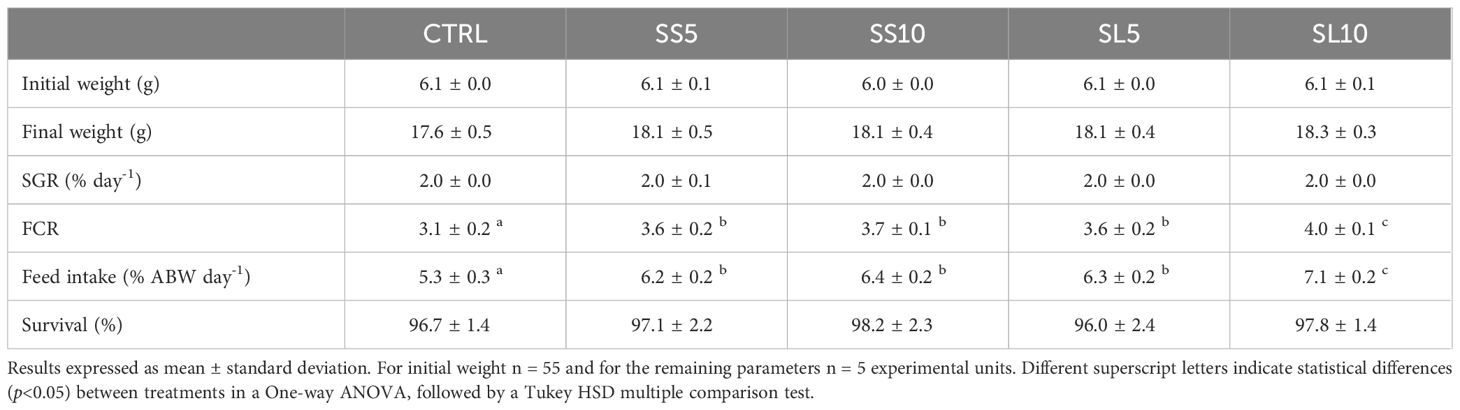
Table 5. Initial and final weight, specific growth rate (SGR), feed conversion ratio (FCR), feed intake and survival of juvenile whiteleg shrimp (P. vannamei) fed the experimental diets for 55 days.
At the end of the feeding trial (Day 55), significant differences between treatments were found in the shrimp whole-body composition. Shrimp fed the CTRL and SL5 diets had significantly higher dry matter contents than those fed SL10, while those fed SS10 had significantly higher ash contents than SL5 (Table 6).
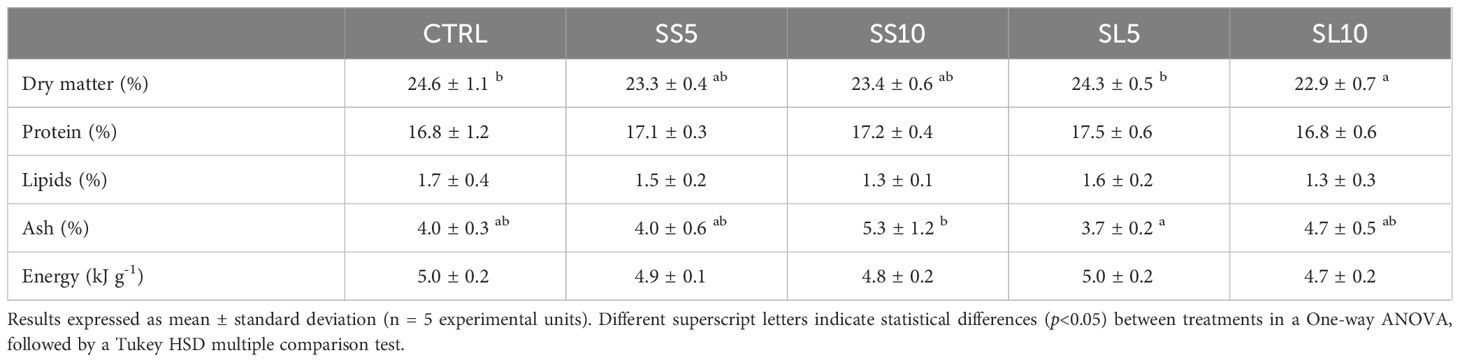
Table 6. Whole-body composition (% fresh weight) of juvenile whiteleg shrimp (P. vannamei) fed the experimental diets for 55 days.
3.1.2 Chemical profile
The phytochemical characterization of whiteleg shrimp allowed the identification of 50 and 40 compounds, after 31 days (Supplementary Table S1) and 55 days of feeding (Supplementary Table S2), respectively. The identified compounds mainly belong to 5 classes, namely alcohols, amino acids, fatty acids, organic acids and sterols. From the analyses of the chemical profile is possible to highlight that at 31 days of feeding, the total relative abundance of both alcohols and sterols is lower in the groups fed with stems and leaves and seeds than in the CTRL group. In the case of alcohols, in groups SS10, SS5 and SL5, this difference is statistically significant. For sterols, only the SL5 and SL10 groups are significantly lower than the CTRL. In organic acids and fatty acids there are significant differences between CTRL and the SL10 group: the relative abundance of organic acids is higher in the SL10 group than in the CTRL, while the relative abundance of fatty acids is lower in the SL10 than in the CTRL. Finally, the amino acids in the SS5, SL5, and SL10 groups have significantly higher relative abundances than CTRL. Regarding the chemical profile at 55 days of feeding, the total relative abundance of alcohols is significantly greater in the SS10 and SL5 groups than in the CTRL. In the case of amino acids, the SL10 group has a statistically greater abundance than in CTRL. The SL5 group, draws attention, having a total relative abundance of amino acids much lower than CTRL and even in relation to the other groups. When looking at this particular situation it appears that there is an abrupt reduction in the relative abundance of glycine in this group. Looking at the sample extraction and analysis process, and since glycine is a 1) the possibility that this compound was not correctly extracted during the hexane extraction process, thus leading to a decrease in its abundance in the extract; 2) the silylation reaction does not occur completely and does not allow the derivatization of glycine and therefore, when injected into the GC-MS, it does not volatilize. The relative abundance of fatty acids is significantly higher and lower in the SL5 and SL10 groups, respectively, in relation to the CTRL. In sterols, the total abundance is higher in the SL5 group and lower in the SL10 group, compared to CTRL. Organic acids are significantly lower in the groups fed with stems. It is also important to note that the compounds identified after 31 days are the same as those identified after 55 days. The difference in the number of compounds identified in these two situations is related to the sensitivity of the equipment used, since these compounds are present in relative abundances of less than 1. Finally, it is verified that the relative abundance of nutritionally important compounds, such as amino acids and fatty acids, are not negatively affected by the incorporation of S. ramosissima in the feed of aquaculture animals.
3.1.3 Haemocytes count
No differences in total haemocytes counts were observed during the feeding trial. In response to the bath bacterial challenge with Vibrio parahaemolyticus, shrimp fed all experimental diets decreased the number of haemocytes in circulation following exposure to the pathogen, nonetheless no differences were observed between the experimental diets (Figure 1).
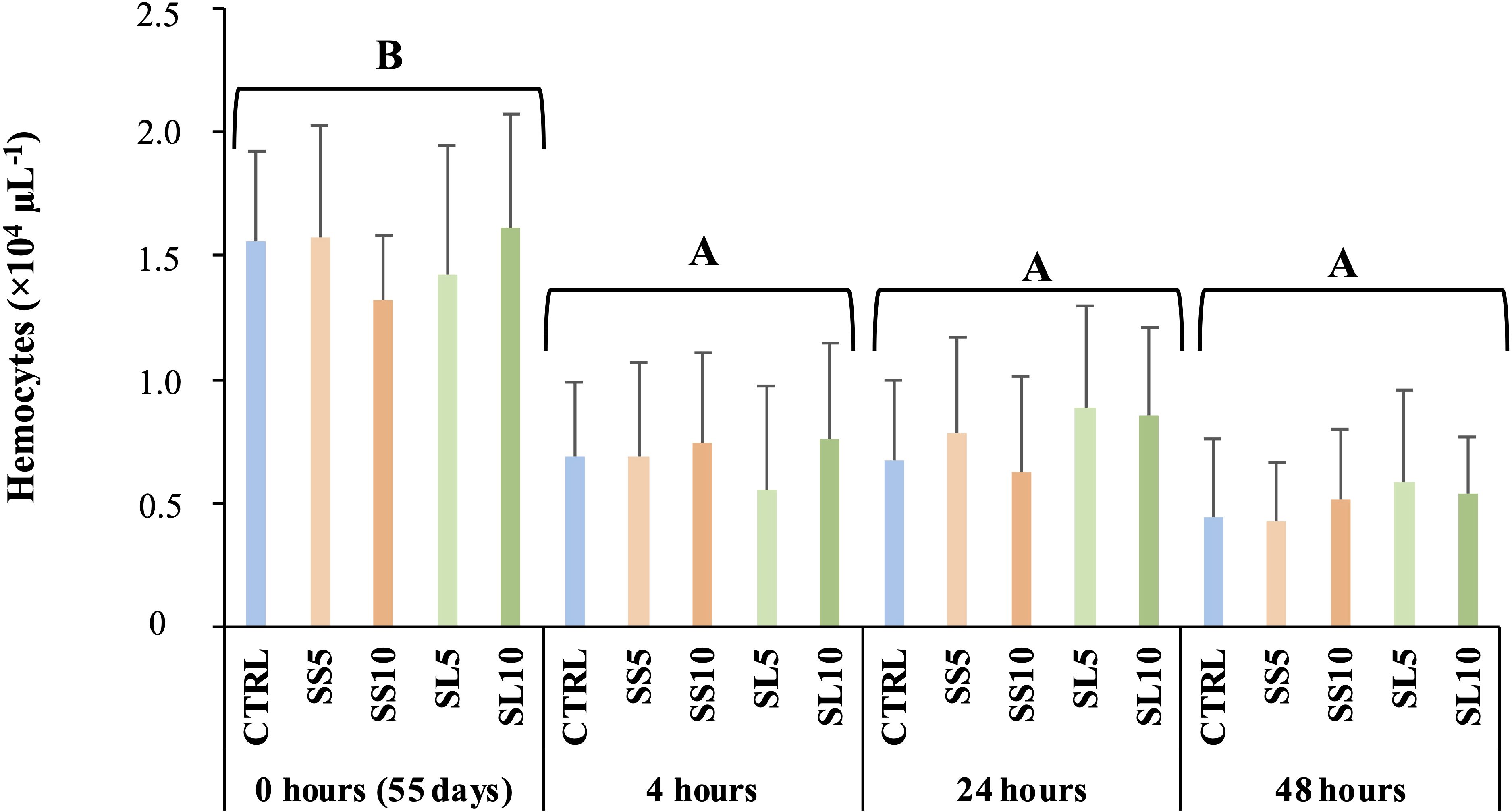
Figure 1. Haemocytes concentration during the bacterial challenge in whiteleg shrimp (P. vannamei). Experimental groups concern: control (Ctrl), fed with standard feed, and diets with S. ramosissima inclusion (SS5, SS10, SL5, and SL10). Values are means ± standard deviation (n=9). P-values from Two-way ANOVA (p ≤ 0.05). Tukey post-hoc test was used to identify differences in the experimental treatments in each sampling point. Different lowercase letters stand for significant differences between dietary treatments for the same time.
3.1.4 Plasma humoral parameters
After 31 days of being fed experimental diets, shrimp that were given SS5, SL5, and SL10 exhibited lower levels of nitric oxide compared to those fed the CTRL diet. Similarly, shrimp fed SS5, SL5, and SL10 also showed lower lysozyme activity after 31 days, in contrast to those on the CTRL diet. The highest lysozyme activity was observed in shrimp that were fed a diet enriched with 10% S. ramosissima stems (refer to Figure 2; Supplementary Table S1). However, no significant differences were observed after 55 days of being fed the experimental diets (refer to Supplementary Table S1; Figure 2). The complete set of results is present in the Supplementary Table S3 of the Supplementary Materials.
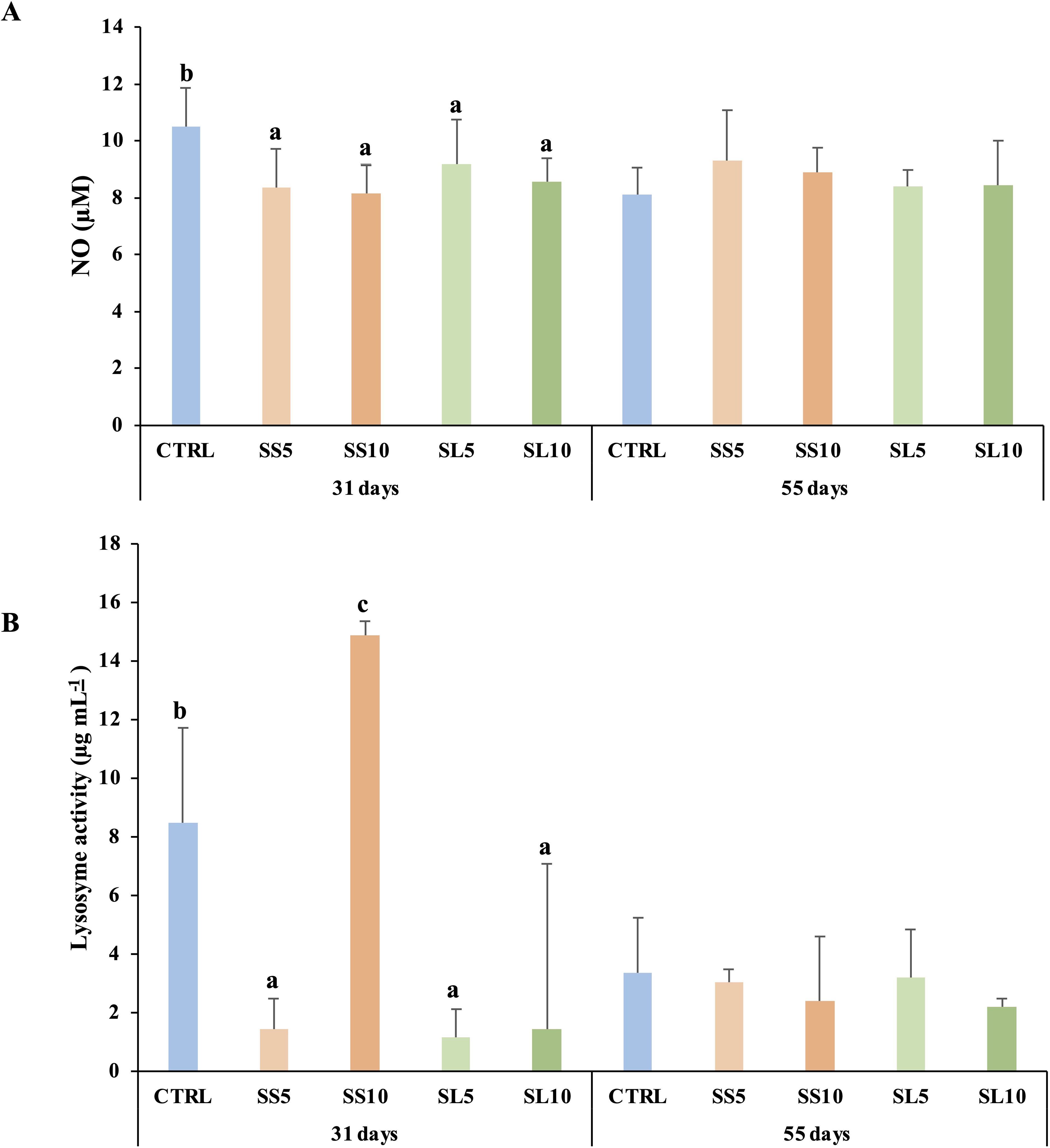
Figure 2. Plasma (A) nitric oxide (NO) and (B) lysozyme activity in whiteleg shrimp (P. vannamei) during the feeding trial. Experimental groups concern: control (Ctrl), fed with standard feed, and diets with S. ramosissima inclusion (SS5, SS10, SL5, and SL10). Values are means ± standard deviation (n=9). P-values from One-way ANOVA (p ≤ 0.05). Tukey post-hoc test was used to identify differences in the experimental treatments in each sampling point. Different lowercase letters stand for significant differences between dietary treatments for the same time.
3.1.5 Hepatopancreas oxidant status
SOD and CAT activities displayed similar response profiles after 31 days of feeding trial. In whiteleg shrimp fed with stems, SOD and CAT activities decreased significantly in comparison with the CTRL group, with the exception of SL5 (Figures 3A, B, respectively). Differently, at the end of the trial (day 55), a significant increase in SOD activity was observed in SS10 (Figure 3A), and no significant variations in CAT activity were detected, in comparison with the CTRL group (Figure 3B). Comparing the two levels leaves and seeds inclusion after 55 days, SL5 group showed lower CAT activity than SL10 (Figure 3B).

Figure 3. Mean values of hepatopancreas oxidative stress parameters in whiteleg shrimp (P. vannamei). (A) superoxide dismutase (SOD), (B) catalase (CAT), (C) lipid peroxidation (LPO). Experimental groups concern: control (Ctrl), fed with standard feed, and diets with S. ramosissima inclusion (SS5, SS10, SL5, and SL10). Bars represent standard deviations. Different letters correspond to statistically significant differences (p<0.05).
GPx activity showed no significant variations, neither when compared with the CTRL group nor among diets with S. ramosissima inclusion.
Concerning LPO levels in hepatopancreas (Figure 3C), significant alterations were restricted to the intermediate sampling (31 days), with both whiteleg shrimp groups fed with S. ramosissima stems (SS5 and SS10) displaying lower levels when compared to the CTRL group.
3.1.6 Hepatopancreas gene expression
Regarding the health and antioxidant biomarkers analysed in shrimp hepatopancreas during the feeding period, main differences between dietary treatment were observed after 31 days of feeding (Supplementary Table S4). On the one hand, it was observed a general downregulation of casp3 in shrimp fed diets rich in S. ramosissima regardless the type or inclusion level. On the other hand, it was observed a general down-regulation of gst in shrimp fed diets rich in S. ramosissima regardless the type or inclusion level after 31 days of feeding. A downregulation of gpx was observed in shrimp fed SS10. After 55 days of feeding, a consistent increase in spermine synthase (SMS) was observed in shrimp that were fed diets rich in S. ramosissima, irrespective of the type or level of inclusion (Figure 4).
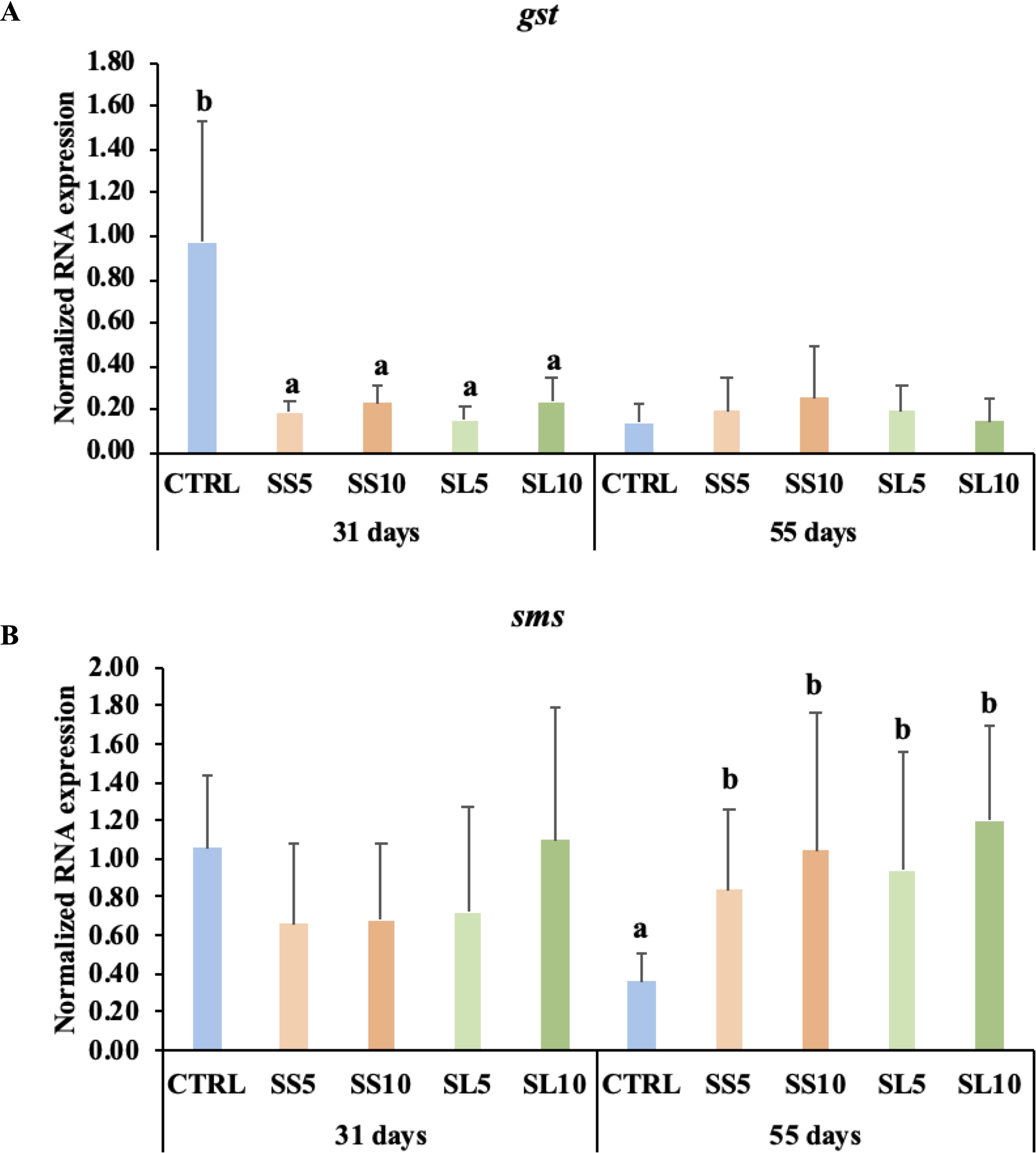
Figure 4. Relative expression of (A) gst (Glutathione transferase) and (B) sms (Spermine synthase) in the hepatopancreas gene expression of whiteleg shrimp (P. vannamei) fed dietary treatments during 55 days. Experimental groups concern: control (CTRL), fed with standard feed, and diets with S. ramosissima inclusion (SS5, SS10, SL5, and SL10). Values are presented as means ± SE (n = 9). P-values from One-way ANOVA (p ≤ 0.05). Tukey post-hoc test was used to identify differences in the experimental treatments in each sampling point. Different lowercase letters stand for significant differences between dietary treatments for the same time.
3.2 Bacterial challenge
3.2.1 Plasma humoral parameters
Whiteleg shrimp fed SS5 and SL5 diets showed a higher production of nitric oxide than those fed the SS10 diet at 24 h after infection. ROS levels were higher in shrimps fed the SL10 diet when compared to those fed the SL5 after 48 h, in general this sampling point ROS levels were higher when compared to the previous samplings. Moreover, it was also observed a higher lysozyme activity in shrimp fed SL10 than those fed the other experimental diets at 48 h post infection (Figure 5; Supplementary Table S5). The complete set of results is present in the Supplementary Table S5 of the Supplementary Materials.
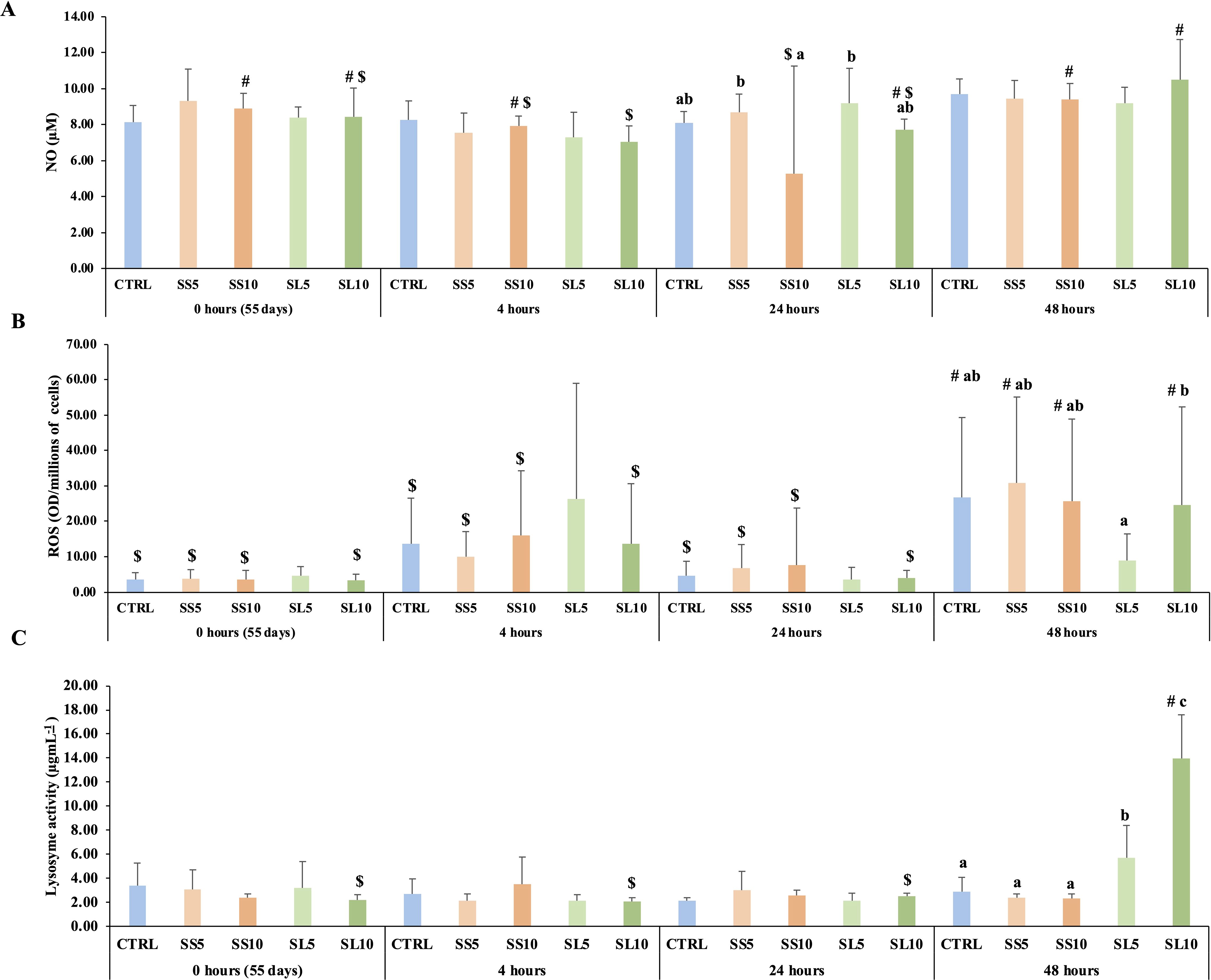
Figure 5. Plasma (A) nitric oxide (NO), (B) reactive oxygen species (ROS) levels and (C) lysozyme activity in whiteleg shrimp (P. vannamei) during the bacterial bath challenge. Experimental groups concern: control (Ctrl), fed with standard feed, and diets with S. ramosissima inclusion (SS5, SS10, SL5, and SL10). Values are means ± standard deviation (n=9). P-values from One-way ANOVA (p ≤ 0.05). Tukey post-hoc test was used to identify differences in the experimental treatments. Different lowercase letters stand for significant differences between dietary treatments for the same time. Different symbols indicate differences between times for the same dietary treatment.
3.2.2 Hepatopancreas gene expression
The same panel of biomarkers studied during the feeding trial were also analysed for whiteleg shrimp sampled during the bacterial challenge. It was observed a general up-regulation of sms in shrimp fed diets rich in S. ramosissima regardless the type or inclusion level (Supplementary Table S6). A down-regulation of lys-like was observed in shrimp fed SS10 and SL5 diets compared to those fed CTRL (Supplementary Table S6). In contrast, an up-regulation of pen3 was observed in shrimp fed SS10 regardless sampling time. Moreover, the expression of the rab was significantly up-regulated in the hepatopancreas of challenged whiteleg shrimp, firstly in those fed S. ramosissima stems (SS) at 5% and 10% after 4 h of infection and then of those fed S. ramosissima seeds and leaves (SL) at 5% and 10% inclusion levels after 48 h of bacterial challenge (Figure 6).
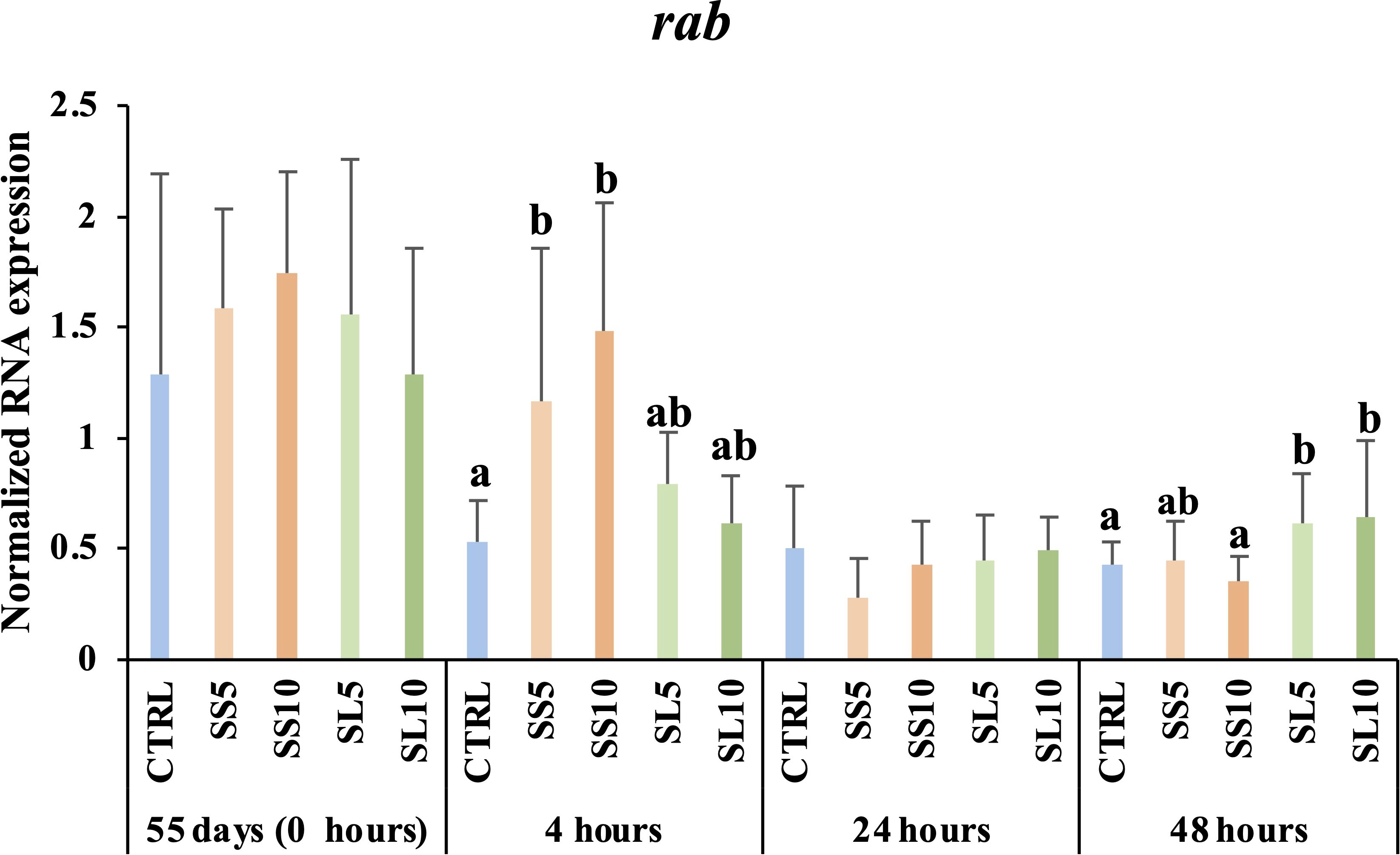
Figure 6. Relative expression of rab (Rab GTPase) in the hepatopancreas gene expression of whiteleg shrimp (P. vannamei) fed dietary treatments during the bath bacterial challenge. Experimental groups concern: control (Ctrl), fed with standard feed, and diets with S. ramosissima inclusion (SS5, SS10, SL5, and SL10). Values are presented as means ± SE (n = 9). P-values from Two-way ANOVA (p ≤ 0.05). Tukey post-hoc test was used to identify differences in the experimental treatments. Different lowercase letters stand for significant differences between dietary treatments for the same time.
3.2.3 Multivariate analysis from the hepatopancreas gene expression
An overall multivariate analysis combining raw data from hepatopancreas gene expression (using PCA‐DA) was performed to discriminate the physiological effects caused by the experimental diets both at 31 and 55 days of feeding, and following the bacterial challenge (Figure 7). The first two discriminant functions accounted for 81.79% of dataset variability for feeding trial (31 and 55 days). Group discrimination was significant (Wilk’s lambda = 0.018, p <0.0001) highlighting the differences at the first sampling point (31 days), between CTRL and the experimental groups (p <0.001). While at the final sampling point, only differences between CTRL and SL10 and SS5 diets we observed. This discrimination was loaded by the expression of hmc, casp3, gst, and gpx (Figure 7A).
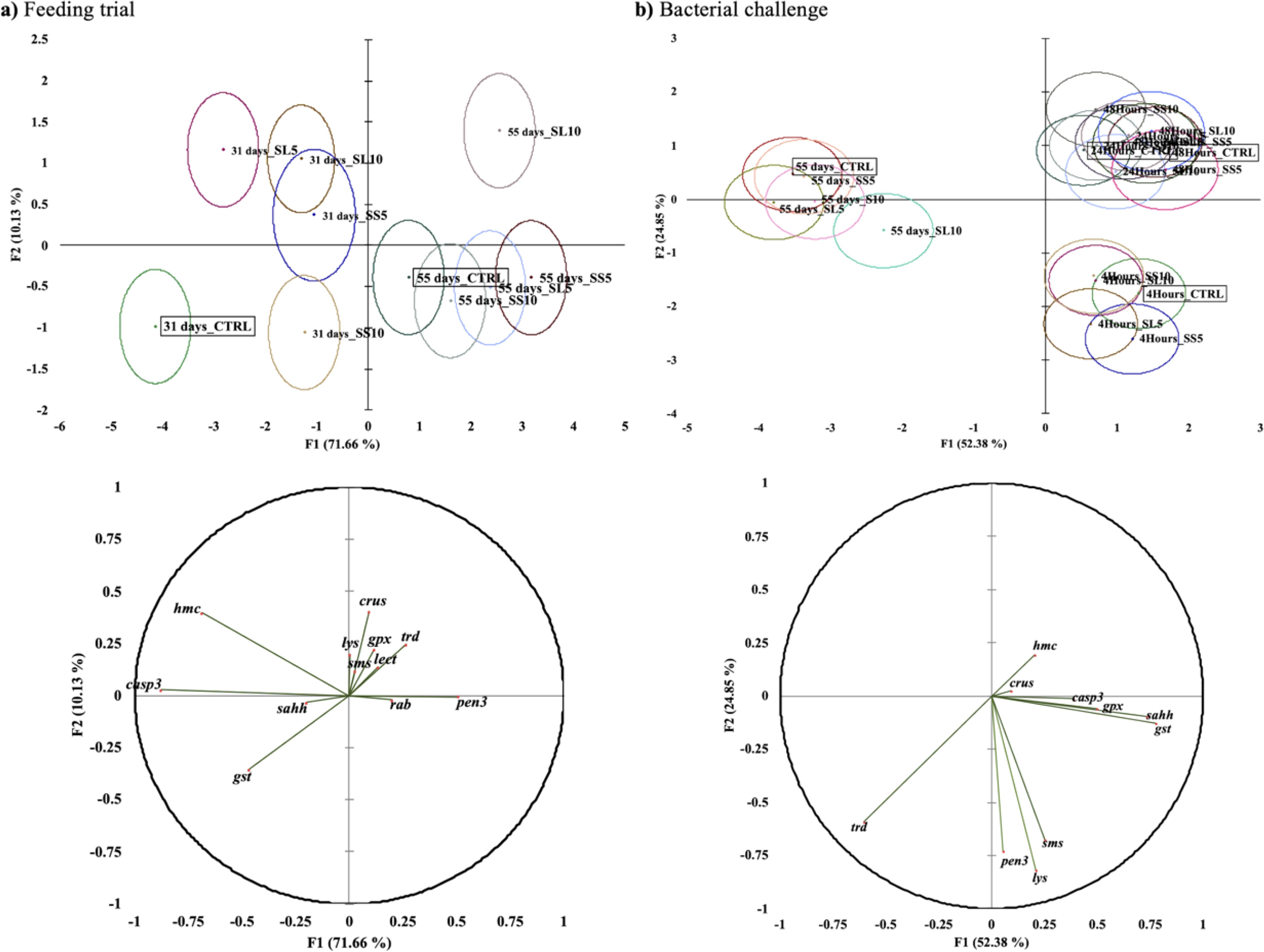
Figure 7. Canonical discriminant analysis of whiteleg shrimp (P. vannamei) immune and antioxidant responses biomarkers expression when fed with diets enriched with S. ramosissima during 31 and 55 days [(A), feeding trial]. And upon a bacterial challenge (B) (4, 24 and 48 h). Small dot marks represent group centroids, whereas ellipses indicate data distribution per group, and correlation variables/factors (factor loads) for two main discriminant functions (F1 and F2).
Following the bacterial challenge, groups were discriminated (Wilk’s lambda = 0.015; p <0.0001) and the first two discriminant factors accounted for 77.22% of dataset variability. Overall, it is possible to observe a clear significant discriminant difference between sampling points regardless of dietary treatment. Particularly between the end of the feeding trial (prior to bacterial challenge) and 4 h following bacterial bath challenge (p <0.0001), and this separation was loaded by the expression of trd, pen3, sms, and lys (Figure 7B).
4 Discussion
Salicornia ramosissima, a halophyte rich in bioactive compounds, has garnered significant attention as a potential source of natural antioxidants and other health-promoting substances. Its high content of polyphenols, including flavonoids, tannins, and phenolic acids, has been linked to various beneficial effects, such as anti-inflammatory and antioxidant properties (Najar et al., 2023). To investigate the potential of S. ramosissima as a functional feed ingredient for aquaculture, we evaluated the effects of incorporating its non-food biomass into the diet of whiteleg shrimp. This study aimed to assess the impact of S. ramosissima on growth performance, survival, immune function, and oxidative stress response. To our knowledge, this is the first study to explore the use of S. ramosissima as a growth modulator or nutraceutical ingredient in shrimp feeds.
Upon scrutinizing the chemical profile analyses, it’s noted that there were some modifications in the chemical profile on the muscle of the shrimp with the inclusion of salicornia in aquafeeds, but these results are complex and it is difficult to find a pattern that relates the inclusion of salicornia with the changes in chemical profile that we found. When we look for organic acids, succinic acid in shrimp muscle increase with salicornia inclusion. It is important to highlight this organic acid as it has been shown to have an important role in the intestinal microbiome of shrimp, modulating it and improving its digestive and immunological capabilities (Duan et al., 2018, 2020). This could indicate some health-promoting effect on shrimp. About amino acids, the groups fed with aquafeeds including salicornia, there was an increase in the total values of amino acids, as well as the essential amino acids leucine, lysine, threonine, valine and phenylalanine, which are fundamental for the normal growth and metabolism of fish (Wilson, 1986). This is beneficial for the overall health and growth of the shrimp, as amino acids are essential for protein synthesis and indicate some health-promoting effect. Intriguingly, the SL5 group exhibited a striking reduction in amino acid relative abundance, chiefly attributed to a notable decrease in glycine levels. Glycine is a key component of many biological processes. The reduction prompts questions about the overall nutritional quality of this specific diet, SL5 (Wang et al., 2013). Therefore, it is very important to look at these results very carefully.
Fatty acids manifested significant differences between the control and the SL10 group, with lower levels. However, DHA and EPA levels were maintained close to control values, and at 55 days there was an increase in DHA and EPA values in the groups fed with diets with 5% salicornia inclusion compared to the control. These two unsaturated fatty acids are the most relevant Omega-3 fatty acids, due to their recognized relevance in human nutrition (Narayan et al., 2006). The presence of essential amino acids and the omega-3 fatty acids DHA and EPA contribute to the nutritional value of shrimp as food for humans.
While S. ramosissima inclusion did not significantly affect shrimp weight, SGR, or survival after 31 and 55 days, feeding efficiency was negatively impacted. Shrimp fed S. ramosissima diets had significantly higher FCR and feed intake, particularly noticeable after 31 days in the SL10 group (leaves and seeds). This contrasts with previous work on juvenile European sea bass conducted in the framework of the same project by Barreto et al. (2024), where S. ramosissima incorporation up to 10% did not negatively impact growth or feed utilization. This species-specific difference emphasizes the importance of considering species-specific digestive capabilities and nutrient requirements when evaluating novel feed ingredients. A non-significant trend towards lower dry matter content was also observed in the SL10 group, possibly linked to increased moisture content and higher ash levels, which could have contributed to the altered feed efficiency. These findings highlight the need for further investigation into the mechanisms underlying the species-specific response to S. ramosissima and optimization of its inclusion levels in shrimp feeds to maximize benefits while mitigating any negative effects on feed efficiency.
In the present study, feeds with S. ramosissima inclusion showed the ability to modulate the shrimp hepatopancreas antioxidant enzymes, specifically SOD and CAT enzyme activities. These enzymes cooperate in transforming toxic species in harmless water products, since SOD converts superoxide radicals into hydrogen peroxide and molecular oxygen, while CAT converts hydrogen peroxide into water (Weydert and Cullen, 2010). The most pronounced effects were visible after 31 days of feeding with SS (5 and 10% inclusion levels) and with SL (10% of inclusion). Moreover, S. ramosissima stems showed the potential to diminish the oxidative damage (measured as LPO), regardless of the dose incorporated. Interestingly, a down-regulation of the glutathione S-transferase gene (gst) was observed in shrimp fed diets rich in S. ramosissima regardless the biomass source or inclusion level after 31 days of feeding. Zhou et al. (2009) in a study with whiteleg shrimp, pointed out that GSTs are important components of various detoxification, antioxidant defence and stress-tolerance pathways. Therefore, the inclusion of S. ramosissima in aquafeeds for whiteleg shrimp juveniles may be a natural way of enhancing the animal’s health status. In fact, the gene expression of antioxidant enzymes (gst and gpx) indicates an overall profile of down-regulation. There seems to be an agreement between the general antioxidant profile at 31 days of feeding, measured both in activity and gene expression, which also aligns with the lipid peroxidation data (with emphasis on the reduction in SS). Overall, the data suggest that S. ramosissima dietary inclusion acts as an enhancer of the oxidant status, particularly when stems are incorporated and after 31 days of feeding. However, this effect appears to diminish after 55 days.
These results appear to confirm the contribution of non-enzymatic antioxidants by S. ramosissima, thereby sparing endogenous resources and creating a more favourable oxidant status. While research on Salicornia spp. in aquaculture feeds is relatively scarse, with a few papers reporting that it can partially substitute fish protein in diets for O. niloticus fingerlings at levels up to 50% of the dietary protein (Ríos-Durán et al., 2013). And recent studies by Barreto et al. (2024) and Marçal et al. (2023) have revealed promising potential benefits. Barreto et al. (2024) observed significant modulatory effects on fish muscle biochemical profiles, including a decrease in lactic acid and an increase in succinic acid levels, suggesting potential health-promoting effects. Marçal et al. (2023) identified a tissue-specific antioxidant improvement and a genotoxic trigger that ultimately led to increased DNA integrity. These findings highlight the promising potential of S. ramosissima as a functional ingredient in aquaculture feeds, offering a novel, low-value raw material that can be valorized to improve fish health and sustainability. Furthermore, studies on farmed sheep have demonstrated the potential of Salicornia spp. to enhance antioxidant responses (Taghipour et al., 2021; Nasrabadi et al., 2022). Suggesting a broader potential for Salicornia as a functional feed ingredient across various aquaculture species. In this study, higher FCR values were obtained with diets where S. ramosissima biomasses were included, which would increase the final production costs. However, potential antioxidant beneficial effects were observed when using the stems, which is considered a by-product of S. ramosissima cultivation. Therefore, by using a product that would otherwise be disposed instead of wheat meal, the price of the aquafeeds could be lower.
Moreover, the inclusion of S. ramosissima in shrimp diets was able to slightly modulate some of the plasma immune parameters and gene expression analysed on the hepatopancreas. No differences were observed in cell counts during the feeding trial, and a general decrease regardless of dietary treatments was observed following the bacterial bath challenge. Regarding the immune parameters analysed in haemolymph plasma, differences between dietary treatments were only observed after 31 days during the feeding trial, suggesting that the diet effect seems to disappear with a longer feeding period regime.
Decreases in plasma nitric oxide levels and lysozyme activity were observed in shrimp fed SS5, SL5 and SL10 after 31 days of feeding. However, whiteleg shrimp fed S. ramosissima stems (SS10) showed a higher lysozyme activity at that same time, suggesting a slightly increase of the immune response boosted by stems. Lysozyme is an enzyme with bacteriolytic properties, which can be found in both prokaryotes and eukaryotes. Its main function is to break down the β-1, 4-glycosidic bonds between N-acetylmuramic acid and N-acetylglucosamine in peptidoglycan, leading to bacterial lysis (Jollès and Jollès, 1984). It has been shown that lysozyme exhibits antimicrobial activity against both Gram-negative and Gram-positive bacteria, including Vibrio species that are harmful to shrimp (Hikima et al., 2003; Supungul et al., 2010). Interestingly, it was observed that shrimp fed S. ramosissima stems at 5% and 10% inclusion levels (SS5 and SS10, respectively) had higher levels of lysozyme activity after the bacterial challenge as well as respiratory burst (RB) particularly at 10% inclusion level, and both after 48 h following bacterial bath challenge, suggesting that shrimp fed diets with S. ramosissima stems might be more prone to deal with the pathogen.
A mild modulation of gene expression during the feeding trial was observed, and particularly interesting to highlight is the up-regulation of spermine synthase after 55 days of feeding in all experimental diets with S. ramosissima regardless of the biomass type or the inclusion level. Spermine synthase is an enzyme that catalyses the conversion of spermidine to spermine, which are polyamines involved in various cellular processes such as cell growth, proliferation, and differentiation (Pegg and Michael, 2010). Interestingly, Machado et al. (2023) observed a modulation of the methionine-related pathways with the up-regulation of hepatopancreas mRNA expression of SAM-synthase (SAM-synth) and also the spermine synthase (SMS) in shrimp fed a diet rich in AQUAVI® Met-Met pointing to stimulation of polyamine biosynthesis route and key for cell proliferation.
The expression of the rab (Rab GTPase) gene responsible for the formation, transport, and fusion of intracellular organelles crucial for immune response was higher in the hepatopancreas of challenged shrimp fed S. ramosissima stems (SS) at 5% and 10% inclusion levels already at 4 h after infection. The Rab protein is an important regulatory component involved in cell biosynthesis, endocytosis, and vesicle transport (Zong et al., 2008). Recent research has also demonstrated its essential role in the development and maturation of phagocytes and innate pathways that aid in the removal of foreign pathogens (Wu and Zhang, 2007). The upregulation of Rab GTPase gene expression in response to the experimental treatment suggests increased activity of the protein. This could indicate enhanced membrane trafficking and vesicle transport, which may be important for the immune response to the challenge. For example, increased endocytosis could facilitate the uptake of pathogens or immune-stimulating molecules, while increased exocytosis could promote the secretion of immune effectors. These findings support the potential health-promoting benefits of Salicornia as mentioned earlier.
To gain a comprehensive understanding of the dietary impact on hepatopancreas gene expression, a multivariate analysis combining data from various genes was conducted. The results revealed that shrimp fed SL10 were markedly distinct from those fed the remaining experimental diets after 55 days of feeding. However, it was impossible to draw any firm conclusions from the bacterial challenge gene expression results, as no significant differences were observed among shrimp fed on different diets. Nevertheless, a clear effect of the bacterial bath challenge was evident, as demonstrated by a sharp distinction between shrimp sampled at 0 hours (55 days of feeding) and those collected after 4 hours of the bacterial bath. Furthermore, a distinct separation was noted between 4 hours and the cluster of 24- and 48-hours post-infection.
In summary, this study demonstrates that integrating S. ramosissima biomass into the diets of juvenile whiteleg shrimp does not compromise their final weight, survival, or overall health. This breakthrough offers a substantial opportunity to reinforce halophyte production and advance sustainable aquafeed practices, potentially supplanting commonplace ingredients like wheat meal, a valuable resource also utilized in human consumption. Moreover, the study’s promising outcome underscores the potential antioxidant advantages stemming from the utilization of S. ramosissima stems, conventionally viewed as cultivation by-products often relegated to waste. This innovative strategy not only taps into an overlooked resource but also presents a cost-effective alternative to traditional aquafeed components like wheat meal. Despite potential challenges related to feed efficiency, the inclusion of S. ramosissima biomass shows promise as a cost-effective and sustainable alternative to traditional feed ingredients. Although further research is needed to optimize the use of S. ramosissima biomass in shrimp feed. The cultivation of S. ramosissima is in line with the current global emphasis on resource optimization, waste reduction, and the exploration of alternative energy sources, positioning it as a viable element of a sustainable circular economy.
Data availability statement
The data that support the findings of this study are available on the following database: https://doi.org/10.6084/m9.figshare.24721608.v1.
Ethics statement
Ethical review and approval were not required for the study on animals in accordance with the local legislation and institutional requirements.
Author contributions
LR: Formal analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing. RM: Formal analysis, Investigation, Methodology, Writing – review & editing. AB: Formal analysis, Investigation, Methodology, Writing – review & editing. AL: Methodology, Writing – review & editing. MM: Investigation, Writing – review & editing. SF: Investigation, Methodology, Writing – review & editing. CT: Formal analysis, Methodology, Writing – review & editing. JO: Formal analysis, Writing – review & editing. AC: Formal analysis, Methodology, Writing – review & editing. JD: Methodology, Resources, Writing – review & editing. SG: Formal analysis, Methodology, Writing – review & editing. AV: Formal analysis, Writing – review & editing. DP: Formal analysis, Writing – review & editing. MP: Conceptualization, Methodology, Project administration, Resources, Validation, Funding acquisition, Supervision, Writing – review & editing. RR: Conceptualization, Investigation, Methodology, Resources, Validation, Funding acquisition, Supervision, Writing – review & editing. BC: Conceptualization, Funding acquisition, Investigation, Project administration, Resources, Supervision, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This work was supported by the European Union’s Horizon 2020 research and innovation programme under grant agreement No. 86283 (project AQUACOMBINE). This output reflects the views only of the authors, and the European Union cannot be held responsible for any use which may be made of the information contained therein. This research was also supported by national funds through FCT – Foundation for Science and Technology within the scope of UIDB/0443/2020 and UIDP/04423/2020. CT and BC were supported by FCT (PD/BDE/135541/2018 and 2020.00290.CEECIND, respectively). And as applicable co-financed by FEDER within the PT2020 Partnership agreement by funding the LAQV-REQUIMTE (UIDB/50006/2020+UIDP/50006/2020).
Acknowledgments
The authors would like to acknowledge technical support from Bruna Silva during shrimp rearing and sampling at Riasearch; the SPAROS feed production team for manufacturing the tested microdiets, Diogo Peixoto and Paulo Santos from CIIMAR for their help during the bacterial challenge procedures. Also, the authors acknowledge support from Renata Serradeiro in funding acquisition and in the conceptualization of this study.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2024.1491402/full#supplementary-material
References
AOAC (2000). Official Methods of Analysis of Association of Official Analytical Chemists. Arlington: Association of Official Analytical Chemists.
Athar M., Iqbal M. (1998). Ferric nitrilotriacetate promotes N-diethylnitrosamine-induced renal tumorigenesis in the rat: implications for the involvement of oxidative stress. Carcinogenesis 19, 1133–1139. doi: 10.1093/carcin/19.6.1133
Barreira L., Resek E., Rodrigues M. J., Rocha M. I., Pereira H., Bandarra N., et al. (2017). Halophytes: Gourmet food with nutritional health benefits? J. Food Composition Anal. 59, 35–42. doi: 10.1016/j.jfca.2017.02.003
Barreto A., Couto A., Jeronimo D., Laranjeira A., Silva B., Nunes C., et al. (2024). Salicornia ramosissima Biomass as a Partial Replacement of Wheat Meal in Diets for Juvenile European Seabass (Dicentrarchus labrax). Anim. (Basel) 14. doi: 10.3390/ani14040614
Bautista-Covarrubias J. C., Velarde-Montes G. J., Voltolina D., Garcia-De La Parra L. M., Soto-Jimenez M. F., Frias-Espericueta M. G. (2014). Humoral and haemocytic responses of Litopenaeus vannamei to Cd exposure. ScientificWorldJournal 2014, 903452. doi: 10.1155/2014/903452
Bird R. P., Draper H. H. (1984). Comparative studies on different methods of malonaldehyde determination. Methods Enzymology. 105: 299–305. doi: 10.1016/S0076-6879(84)05038-2
Chaturvedi T., Christiansen A. H. C., Gołębiewska I., Thomsen M. H. (2021). “Salicornia Species,” in Future of Sustainable Agriculture in Saline Environments, 461–482 Boca Raton: CRC Press.
Clairborne A. (1985). “Catalase activity,” in Handbook of Methods in Oxygen Radical Research. Ed. Greenwald R. A. (Boca Raton: CRC Press).
Costas B., Conceicao L. E. C., Aragao C., Martos J. A., Ruiz-Jarabo I., Mancera J. M., et al. (2011). Physiological responses of Senegalese sole (Solea Senegalensis Kaup 1858) after stress challenge: Effects on non-specific immune parameters, plasma free amino acids and energy metabolism. Aquaculture 316, 68–76. doi: 10.1016/j.aquaculture.2011.03.011
Duan Y., Wang Y., Ding X., Xiong D., Zhang J. (2020). Response of intestine microbiota, digestion, and immunity in Pacific white shrimp Litopenaeus vannamei to dietary succinate. Aquaculture 517. doi: 10.1016/j.aquaculture.2019.734762
Duan Y., Wang Y., Zhang J., Sun Y., Wang J. (2018). Dietary effects of succinic acid on the growth, digestive enzymes, immune response and resistance to ammonia stress of Litopenaeus vannamei. Fish Shellfish Immunol. 78, 10–17. doi: 10.1016/j.fsi.2018.04.008
Essaidi I., Brahmi Z., Snoussi A., Ben Haj Koubaier H., Casabianca H., Abe N., et al. (2013). Phytochemical investigation of Tunisian Salicornia herbacea L., antioxidant, antimicrobial and cytochrome P450 (CYPs) inhibitory activities of its methanol extract. Food Control 32, 125–133. doi: 10.1016/j.foodcont.2012.11.006
Faustino M. V., Seca A. M. L., Silveira P., Silva A. M. S., Pinto D. C. G. A. (2017). Gas chromatography–mass spectrometry profile of four Calendula L. taxa: A comparative analysis. Ind. Crops Products 104, 91–98. doi: 10.1016/j.indcrop.2017.04.029
Ferreira D., Isca V. M. S., Leal P., Seca A. M. L., Silva H., De Lourdes Pereira M., et al. (2018). Salicornia ramosissima: Secondary metabolites and protective effect against acute testicular toxicity. Arabian J. Chem. 11, 70–80. doi: 10.1016/j.arabjc.2016.04.012
Ferreira M. J., Sierra-Garcia I. N., Louvado A., Gomes N. C. M., Figueiredo S., Patinha C., et al. (2023). Domestication shapes the endophytic microbiome and metabolome of Salicornia europaea. J. Appl. Microbiol. 134. doi: 10.1093/jambio/lxad178
Flowers T. J., Colmer T. D. (2008). Salinity tolerance in halophytes. New Phytol. 179, 945–963. doi: 10.1111/j.1469-8137.2008.02531.x
Flowers T. J., Muscolo A. (2015). Introduction to the Special Issue: Halophytes in a changing world. AoB Plants 7. doi: 10.1093/aobpla/plv020
Fredsgaard M., Kaniki S. E. K., Antonopoulou I., Chaturvedi T., Thomsen M. H. (2023). Phenolic compounds in salicornia spp. and their potential therapeutic effects on H1N1, HBV, HCV, and HIV: A review. Molecules 28. doi: 10.3390/molecules28145312
Fujiki K., Yano T. (1997). Effects of sodium alginate on the non-specific defence system of the common carp (Cyprinus carpio L.). Fish Shellfish Immunol. 7, 417–427. doi: 10.1006/fsim.1997.0095
Hikima S., Hikima J., Rojtinnakorn J., Hirono I., Aoki T. (2003). Characterization and function of kuruma shrimp lysozyme possessing lytic activity against Vibrio species. Gene 316, 187–195. doi: 10.1016/S0378-1119(03)00761-3
Ji P.-F., Yao C.-L., Wang Z.-Y. (2009). Immune response and gene expression in shrimp (Litopenaeus vannamei) hemocytes and hepatopancreas against some pathogen-associated molecular patterns. Fish Shellfish Immunol. 27, 563–570. doi: 10.1016/j.fsi.2009.08.001
Jollès P., Jollès J. (1984). What’s new in lysozyme research? Always a model system, today as yesterday. Mol. Cell Biochem. 63, 165–189. doi: 10.1007/BF00285225
Limongelli F., Crupi P., Clodoveo M. L., Corbo F., Muraglia M. (2022). Overview of the polyphenols in salicornia: from recovery to health-promoting effect. Molecules 27. doi: 10.3390/molecules27227954
Machado M., Fernández-Boo S., Teixeira C., Viegas M., Serradeiro R., Dias J., et al. (2023). DL-methionyl-DL-methionine as an efficient methionine source for promoting zootechnical performance and methionine-related pathways in the whiteleg shrimp (Penaeus vannamei). Br. J. Nutr., 1–17. doi: 10.1017/S0007114522003579
Magni N. N., Verissimo A. C. S., Silva H., Pinto D. (2023). Metabolomic profile of Salicornia perennis plant’s organs under diverse in situ Stress: The Ria de Aveiro salt marshes case. Metabolites 13. doi: 10.3390/metabo13020280
Marçal R., Sousa P., Marques A., Pereira V., Guilherme S., Barreto A., et al. (2023). Exploring the antioxidant and genoprotective potential of Salicornia ramosissima incorporation in the diet of the European seabass (Dicentrarchus labrax). Anim. (Basel) 14. doi: 10.3390/ani14010093
Mohandas J., Marshall J. J., Duggin G. G., Horvath J. S., Tiller D. J. (1984). Differential distribution of glutathione and glutathione-related enzymes in rabbit kidney: Possible implications in analgesic nephropathy. Biochem. Pharmacol. 33, 1801–1807. doi: 10.1016/0006-2952(84)90353-8
Mukai K., Nagai S., Ohara K. (2005). Kinetic study of the quenching reaction of singlet oxygen by tea catechins in ethanol solution. Free Radic. Biol. Med. 39, 752–761. doi: 10.1016/j.freeradbiomed.2005.04.027
Murota K., Terao J. (2003). Antioxidative flavonoid quercetin: implication of its intestinal absorption and metabolism. Arch. Biochem. Biophys. 417, 12–17. doi: 10.1016/S0003-9861(03)00284-4
Najar A. M., Romero-Bernal M., Del Rio C., Montaner J. (2023). A review on polyphenols in salicornia ramosissima with special emphasis on their beneficial effects on brain ischemia. Nutrients 15. doi: 10.3390/nu15030793
Narayan B., Miyashita K., Hosakawa M. (2006). Physiological effects of eicosapentaenoic acid (EPA) and docosahexaenoic acid (DHA)—A review. Food Rev. Int. 22, 291–307. doi: 10.1080/87559120600694622
Nasrabadi M., Rouzbehan Y., Fazaeli H., Rezaei J. (2022). Influence of partial replacement of alfalfa with two Salicornia forages on digestion, rumen variables, blood biochemistry metabolites and antioxidant capacity in sheep. Small Ruminant Res. 214. doi: 10.1016/j.smallrumres.2022.106744
Pascual C., Gaxiola G., Rosas C. (2003). Blood metabolites and hemocyanin of the white shrimp, Litopenaeus vannamei: the effect of culture conditions and a comparison with other crustacean species. Mar. Biol. 142, 735–745. doi: 10.1007/s00227-002-0995-2
Pegg A. E., Michael A. J. (2010). Spermine synthase. Cell Mol. Life Sci. 67, 113–121. doi: 10.1007/s00018-009-0165-5
Ramos-Pinto L., Martos-Sitcha J. A., Reis B., Azeredo R., Fernandez-Boo S., Pérez-Sánchez J., et al. (2019). Dietary tryptophan supplementation induces a transient immune enhancement of gilthead seabream (Sparus aurata) juveniles fed fishmeal-free diets. Fish Shellfish Immunol. 93, 240–250. doi: 10.1016/j.fsi.2019.07.033
Rhee M., Park H.-J., Cho J. (2009). Salicornia herbacea: Botanical, chemical and pharmacological review of halophyte marsh plant. J. Medicinal Plants Res. 3, 548–555.
Ríos-Durán M. G., Valencia I. R., Ross L. G., Martínez-Palacios C. A. (2013). Nutritional evaluation of autoclaved Salicornia bigelovii Torr. seed meal supplemented with varying levels of cholesterol on growth, nutrient utilization and survival of the Nile tilapia (Oreochromis niloticus). Aquaculture Int. 21, 1355–1371. doi: 10.1007/s10499-013-9638-5
Silva A. M., Lago J. P., Pinto D., Moreira M. M., Grosso C., Cruz Fernandes V., et al. (2021). Salicornia ramosissima Bioactive Composition and Safety: Eco-Friendly Extractions Approach (Microwave-Assisted Extraction vs. Conventional Maceration). Appl. Sci. 11. doi: 10.3390/app11114744
Supungul P., Rimphanitchayakit V., Aoki T., Hirono I., Tassanakajon A. (2010). Molecular characterization and expression analysis of a c-type and two novel muramidase-deficient i-type lysozymes from Penaeus monodon. Fish Shellfish Immunol. 28, 490–498. doi: 10.1016/j.fsi.2010.01.003
Taghipour M., Rouzbehan Y., Rezaei J. (2021). Influence of diets containing different levels of Salicornia bigelovii forage on digestibility, ruminal and blood variables and antioxidant capacity of Shall male sheep. Anim. Feed Sci. Technol. 281. doi: 10.1016/j.anifeedsci.2021.115085
Torres M. A., Pires Testa C., Gáspari C., Beatriz Masutti M., Maria Neves Panitz C., Curi-Pedrosa R., et al. (2002). Oxidative stress in the mussel Mytella guyanensis from polluted mangroves on Santa Catarina Island, Brazil. Mar. pollut. Bull. 44, 923–932. doi: 10.1016/S0025-326X(02)00142-X
Tsao R. (2010). Chemistry and biochemistry of dietary polyphenols. Nutrients 2, 1231–1246. doi: 10.3390/nu2121231
Ventura Y., Sagi M. (2013). Halophyte crop cultivation: The case for Salicornia and Sarcocornia. Environ. Exp. Bot. 92, 144–153. doi: 10.1016/j.envexpbot.2012.07.010
Waldhorn D., Autric E. (2022). Shrimp production: Understanding the scope of the problem. Center for Open Science.
Wang W., Wu Z., Dai Z., Yang Y., Wang J., Wu G. (2013). Glycine metabolism in animals and humans: implications for nutrition and health. Amino Acids 45, 463–477. doi: 10.1007/s00726-013-1493-1
Weydert C. J., Cullen J. J. (2010). Measurement of superoxide dismutase, catalase and glutathione peroxidase in cultured cells and tissue. Nat. Protoc. 5, 51–66. doi: 10.1038/nprot.2009.197
Wicke B., Smeets E., Dornburg V., Vashev B., Gaiser T., Turkenburg W., et al. (2011). The global technical and economic potential of bioenergy from salt-affected soils. Energy Environ. Sci. 4, 2669–2681. doi: 10.1039/C1EE01029H
Wilhelm Filho D., Torres M., Tribess T., Pedrosa R., Soares C. H. L. (2001a). Influence of season and pollution on the antioxidant defenses of the cichlid fish acará (Geophagus brasiliensis). Braz. J. Med. Biol. Res. 34, 719–726. doi: 10.1590/S0100-879X2001000600004
Wilhelm Filho D., Tribess T., Gáspari C., Claudio F. D., Torres M. A., Magalhães A. R. M. (2001b). Seasonal changes in antioxidant defenses of the digestive gland of the brown mussel (Perna perna). Aquaculture 203, 149–158. doi: 10.1016/S0044-8486(01)00599-3
Wilson R. P. (1986). Protein and amino acid requirements of fishes. Annu. Rev. Nutr. 6, 225–244. doi: 10.1146/annurev.nu.06.070186.001301
Wu W., Zhang X. (2007). Characterization of a Rab GTPase up-regulated in the shrimp Peneaus japonicus by virus infection. Fish Shellfish Immunol. 23, 438–445. doi: 10.1016/j.fsi.2007.01.001
Zhou J., Wang W. N., Wang A. L., He W. Y., Zhou Q. T., Liu Y., et al. (2009). Glutathione S-transferase in the white shrimp Litopenaeus vannamei: Characterization and regulation under pH stress. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 150, 224–230. doi: 10.1016/j.cbpc.2009.04.012
Keywords: aquaculture, halophytes, immune status, oxidative stress, circular economy
Citation: Ramos-Pinto L, Marçal R, Barreto A, Laranjeira A, Machado M, Fernández-Boo S, Teixeira C, Oliveira J, Couto A, Dias J, Guilherme S, Veríssimo ACS, Pinto DCGA, Pacheco M, Rocha RJM and Costas B (2024) Inclusion of Salicornia ramosissima biomass in diets for juvenile whiteleg shrimp (Penaeus vannamei) induces favourable but transient effects in the immune and oxidative status. Front. Mar. Sci. 11:1491402. doi: 10.3389/fmars.2024.1491402
Received: 04 September 2024; Accepted: 12 November 2024;
Published: 12 December 2024.
Edited by:
Gladstone Sagada, Victory Farms, KenyaReviewed by:
Yuwen Dong, University of Pennsylvania, United StatesAntonio Luca Langellotti, University of Naples Federico II, Italy
Copyright © 2024 Ramos-Pinto, Marçal, Barreto, Laranjeira, Machado, Fernández-Boo, Teixeira, Oliveira, Couto, Dias, Guilherme, Veríssimo, Pinto, Pacheco, Rocha and Costas. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Lourenço Ramos-Pinto, bG91cmVuY28ucGludG9AY2lpbWFyLnVwLnB0; Benjamín Costas, YmNvc3Rhc0BjaWltYXIudXAucHQ=