 Sydney N. Zotto1*
Sydney N. Zotto1* Robin C. Dunkin2
Robin C. Dunkin2 Juliana D. Limon2Maia S. Smith2Lisabet M. Hortensius3
Juliana D. Limon2Maia S. Smith2Lisabet M. Hortensius3 Margaret E. Martinez3
Margaret E. Martinez3 Pádraig J. Duignan3
Pádraig J. Duignan3 Cara Field3
Cara Field3 Esteban Soto4Dane Whitaker4Carlos Rios3
Esteban Soto4Dane Whitaker4Carlos Rios3 Maurice Byukusenge5
Maurice Byukusenge5 Bhushan Jayarao5
Bhushan Jayarao5 Suresh V. Kuchipudi6
Suresh V. Kuchipudi6 Amy C. Kinsley1
Amy C. Kinsley1- 1Department of Veterinary Population Medicine, University of Minnesota, Saint Paul, MN, United States
- 2Long Marine Laboratory, Department of Ecology and Evolutionary Biology, University of California, Santa Cruz, Santa Cruz, CA, United States
- 3The Marine Mammal Center, Sausalito, CA, United States
- 4Department of Medicine and Epidemiology, School of Veterinary Medicine, University of California, Davis, Davis, CA, United States
- 5Animal Diagnostic Laboratory, Department of Veterinary and Biomedical Sciences, Pennsylvania State University, University Park, PA, United States
- 6Department of Infectious Diseases and Microbiology, University of Pittsburgh School of Public Health, Pittsburgh, PA, United States
Klebsiella pneumoniae is a Gram-negative, facultative anaerobic, and non-motile bacillus frequently observed in mammals. A hypermucoviscous (HMV) phenotype of K. pneumoniae is reported in stranded California sea lions (CSL) causing pleuritis and bronchopneumonia as a persistent cause of strandings. We describe an increase in cases of HMV and non-HMV K. pneumoniae in stranded CSLs along the California coast between 2017 to 2022. 2,146 live-stranded CSLs were rescued by The Marine Mammal Center (Sausalito, California). 47 of these tested positive for K. pneumoniae, with a rise in annual incidence from 0.92% (3/331) in 2017 to 8.5% (14/165) and 7.2% (23/320) in 2021 and 2022, respectively. These strandings occurred across 965 kilometers along the North-Central California coast from Bodega Bay, Sonoma, to Avila Beach, San Luis Obispo. A similar pattern was observed in beach-cast CSL carcasses by the Long Marine Laboratory (LML) Stranding Network in Santa Cruz, California who had 8 CSLs test positive for K. pneumoniae with an annual incidence rate of 2.0% (1/50) and 10.1% (7/69) in 2021 and 2022, respectively, with no cases reported prior to 2021. We applied whole-genome sequencing (WGS) to 8 isolates obtained by LML in 2022. WGS analysis revealed high nucleotide similarity between any pair of isolates. Multilocus Sequence Typing showed that all isolates belonged to Sequence type 65 and contained the K2 polysaccharide capsule and siderophore aerobactin, which are all characteristics of hypervirulent K. pneumoniae. We discuss possible causes for this trend and a need for research on K. pneumoniae in free-ranging CSLs in California.
1 Introduction
K. pneumoniae is an encapsulated, gram-negative, facultative anaerobic, and non-motile bacillus of the order Enterobacterales. K. pneumoniae is frequently observed as a ubiquitous environmental and mucosal organism, historically implicated in nosocomial infections in humans (Podschun and Ullmann, 1998; Shon et al., 2013). K. pneumoniae is recognized as a pathogen in a wide range of species with a broad spectrum of clinical manifestations such as suppurative arthritis, meningoencephalitis, peritonitis, secondary infections, respiratory and urinary tract infections, abscessation, and septicemia (Han, 1995; Podschun and Ullmann, 1998; Doud et al., 2009). K. pneumoniae is considered an emerging pathogen of concern due to its broad host, environmental, and surface range along with its high level of resistance to common disinfectants and environmental stressors (Jang et al., 2010; Clegg and Murphy, 2016; Soto et al., 2020). K. pneumoniae also possesses an intrinsic resistance to ampicillin and is able to readily develop multi-drug resistant (MDR) phenotypes contributing to its regard as a human, veterinary, and potentially zoonotic pathogen with the potential for community spread (Jang et al., 2010; Soto et al., 2012; Li et al., 2014; Zhang et al., 2019; Wang et al., 2020; Santaniello et al., 2020; Soto et al., 2020; Hu et al., 2021).
In recent years, a hypermucoviscosity phenotype (HMV) has been described and defined by its distinct capsular morphology and specific genes (rmpA, wzy, magA, siderophore genes) (Fang et al., 2004; McIver and Janda, 2008; Jang et al., 2010). First identified in humans, this phenotype is now of major concern in many marine and terrestrial mammalian species due to its invasive nature and ability to cause severe localized and systemic disease in novel sites and is readily spread in communities (McIver and Janda, 2008; Jang et al., 2010; Chang et al., 2022). HMV K. pneumoniae is now a pathogen of major concern for humans due to its ability to cause multisystemic and potentially deadly disease as well as its apparently broadening geographical range (Lederman and Crum, 2005; Shon et al., 2013; Qian et al., 2016; Russo and Marr, 2019).
Previous studies have described a variety of disease manifestations linked to HMV strains in marine mammals including California sea lions (Zalophus californianus; CSLs), endangered New Zealand sea lions (Phocarctos hookeri), Pacific harbor seals (Phoca vituline richardii), harbor porpoises (Phocoena phocoena), and threatened Southern Sea otters (Enhydra lutris nereis) (Spraker et al., 2007; Jang et al., 2010; Roe et al., 2015; Seguel et al., 2017; Whitaker et al., 2018; Chang et al., 2022). In CSLs, the HMV phenotype is most commonly associated with pleuritis and bronchopneumonia and has been considered a minor yet persistent cause of strandings in CSLs (Jang et al., 2010; Roe et al., 2015; Seguel et al., 2017). HMV K. pneumoniae has also been a leading cause of endangered New Zealand sea lion pup mortality due to septicemia and meningitis since it was first identified (Castinel et al., 2007a, b; Michael et al., 2022).
Emerging pathogens in the marine environment are important in marine mammal ecology and health due to their significant role in mass mortality events (MME). Infectious diseases are the most common cause of MMEs in semi-aquatic species, and historically, viruses are the most common cause of MMEs in marine mammals, with environmental and anthropogenic factors playing a contributing role (Sanderson and Alexander, 2020). Outbreaks of bacterial disease including K. pneumoniae in New Zealand sea lions and leptospirosis in California sea lions have been implicated in previous MME events (Gulland et al., 1996; Roe et al., 2015; Sanderson and Alexander, 2020; Duignan, 2023).
The Marine Mammal Center (TMMC) in Sausalito, CA responds to live CSL strandings from Mendocino through San Luis Obispo counties (965 kilometers), including San Francisco Bay and Delta areas. Long Marine Lab (LML) based in Santa Cruz, CA responds to dead CSL strandings in Santa Cruz County and surrounding areas (NOAA Fisheries, 2024). In recent years reflected in this study, there has been a documented increase in K. pneumoniae-positive cultures from CSLs in rehabilitation at TMMC as well as those necropsied by LML (Figure 1). Among these positive cultures, HMV strains of K. pneumoniae have been isolated, making it important to investigate further the incidence and prevalence of these strains due to their implications as severe disease-causing organisms often linked to high mortality and morbidity outbreaks (Seguel et al., 2017; Whitaker et al., 2018).

Figure 1. (A) Monthly California Sea Lion (CSL) strandings and K. pneumoniae cases from The Marine Mammal Center (TMMC) between 2017-2022; (B) Monthly California Sea Lion strandings and K. pneumoniae cases from Long Marine Lab (LML) between 2017-2022. Cases (KP+) represent CSL strandings that tested positive for K. pneumoniae. Gray bars and n values represent CSL strandings that tested positive for K. pneumoniae. Green bars represent CSL strandings that were not tested or tested negative for K. pneumoniae.
Here we present the descriptive epidemiological features of HMV K. pneumoniae infections in stranded CSLs reported by TMMC and LML along the central and northern California coast from 2017 to 2022. The primary objectives are twofold: firstly, to describe the increase in HMV and non-HMV K. pneumoniae cases in these populations of stranded CSLs, and secondly, to conduct whole genome sequencing analysis to investigate the similarity, virulence, and resistance traits of some of the K. pneumoniae strains involved.
2 Materials and methods
The medical record databases at each respective institution were searched for confirmed cases of K. pneumoniae associated infections. A confirmed case was defined as isolation of K. pneumoniae from cultured necropsy tissues. Demographic data including sex, age, and location of stranding were extracted from the datasets for descriptive epidemiologic analysis. Sex was defined as male, female, or undetermined by the respective institutions. The age category was defined as adult, subadult, juvenile (males only), yearling, pup, or undetermined by the respective institutions. The age categories were based on straight length, dentition, and sexual maturity using the sagittal crest to identify males (Greig et al., 2005).
Aerobic culture swabs were collected from necropsy samples by both institutions with abscessation, pyothorax, pleuritis, bronchopneumonia, peritonitis, meningitis, suppurative arthritis, and/or a combination of multiple lesions (see supplement for tissue culture source breakdown). The swabs were stored on transport media/plated for up to 24 hours at 4°C then plated onto 5% sheep blood agar and MacConkey agar and incubated at 37°C and 5% CO2 with commercial bacterial identification performed on positive growths (Seguel et al., 2017).
Positive isolates from TMMC had a string test performed on the plate colonies to determine if that culture was suggestive of a hypermucoviscous phenotype (Shon et al., 2013; Seguel et al., 2017). Positive isolates from LML were submitted for whole genome sequencing by the Pennsylvania State University Genomics and Bioinformatics Service to determine their genotypes. The genomic DNA was extracted using the PureLink™ Genomic DNA Mini Kit (Thermofisher, Waltham, MA). The Illumina DNA Prep library prep kit (Illumina, San Diego, CA) was used to generate the libraries for sequencing on the Illumina MiniSeq platform. The raw reads quality assessment and quality control were performed using FastQC v. 0.11.9 (Andrews, 2019) and fastp v0.23.2 (Chen et al., 2018), respectively. SKESA v2.4.0 (Souvorov et al., 2018) was used to produce draft genome assemblies. Sequence similarity was assessed using an average nucleotide identity analysis (ANI) using FastANI (Jain et al., 2018). The Kleborate pipeline (Lam et al., 2021) was used to determine the isolates’ genotypes through the multiple sequence typing. The same pipeline was used to determine the virulence factors and antimicrobial resistance determinants.
3 Results
During the study period, 2,146 stranded CSLs were rescued by TMMC, with 47 of these culturing positive for K. pneumoniae. Of the cumulative CSL strandings reported by TMMC, 19 animals stranded more than once within the study period; however, none of the repeat strandings were positive for K. pneumoniae. Details on the location, age, and sex are reported in Figures 2, 3. None of the K. pneumoniae-positive live-stranded animals during this study period were successfully rehabilitated or released, as all of the included CSLs were either euthanized or died spontaneously. Within this timeframe, there was a steep increase in the annual incidence of K. pneumoniae cases at TMMC from 0.92% (3/331) in 2017 to 8.5% (14/165) and 7.2% (23/320) in 2021 and 2022, respectively. These strandings occurred across a broad geographic range from Bodega Bay, Sonoma, to Avila Beach, San Luis Obispo, along the central California coast (Figure 2). At the center of this range, a similar pattern was observed for the dead-stranded CSLs sampled by LML. Prior to 2021, LML had not detected any CSLs infected with K. pneumoniae, on routine cultures, but since then, eight CSLs have tested positive for K. pneumoniae with an annual incidence rate of 1.9% (1/54) and 11.3% (7/62) in 2021 and 2022, respectively (Figures 3, 4).
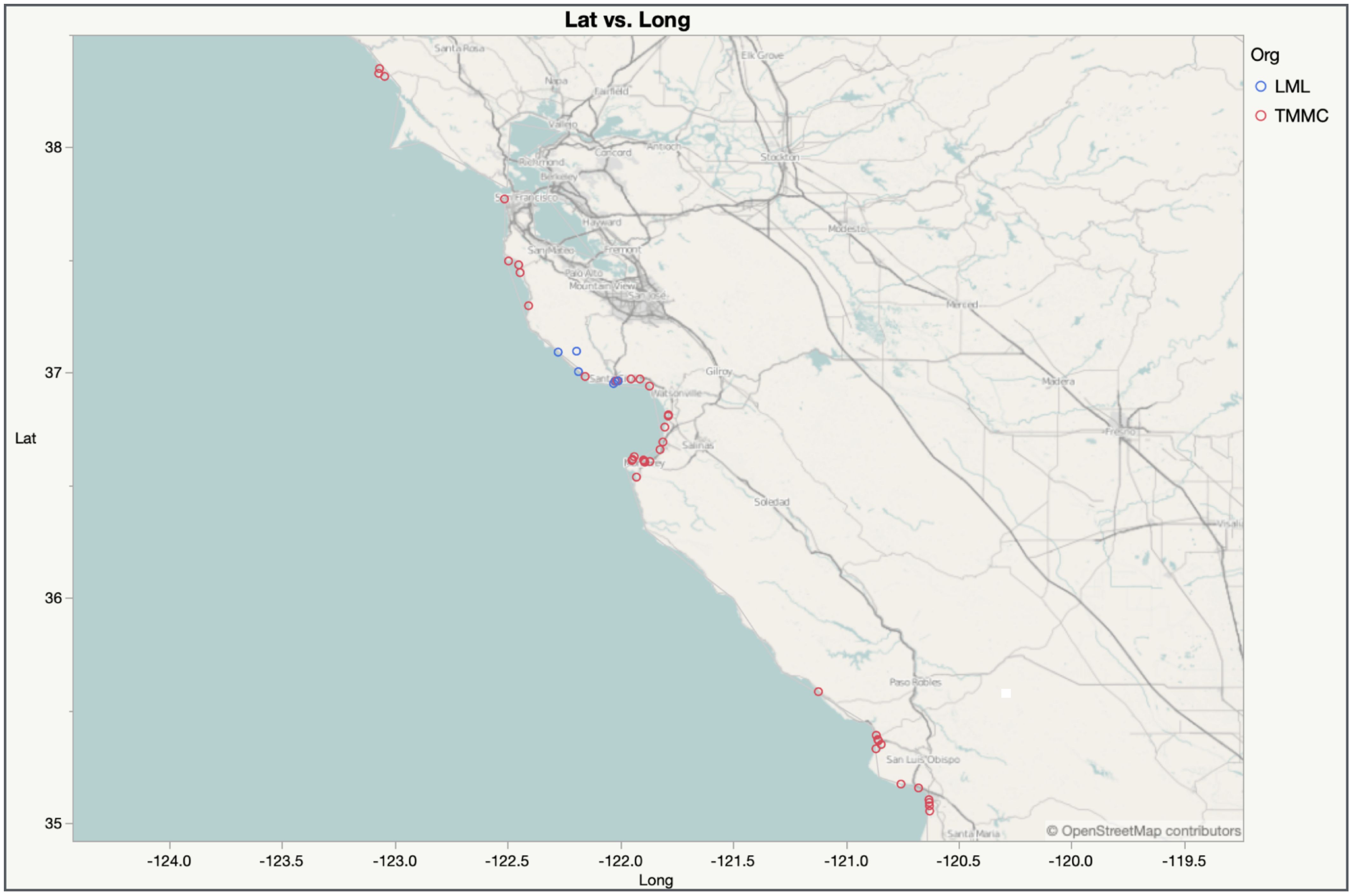
Figure 2. Map of K. pneumoniae stranding cases for The Marine Mammal Center (red dots) and Long Marine Laboratory (blue dots) between 2017 and 2022.
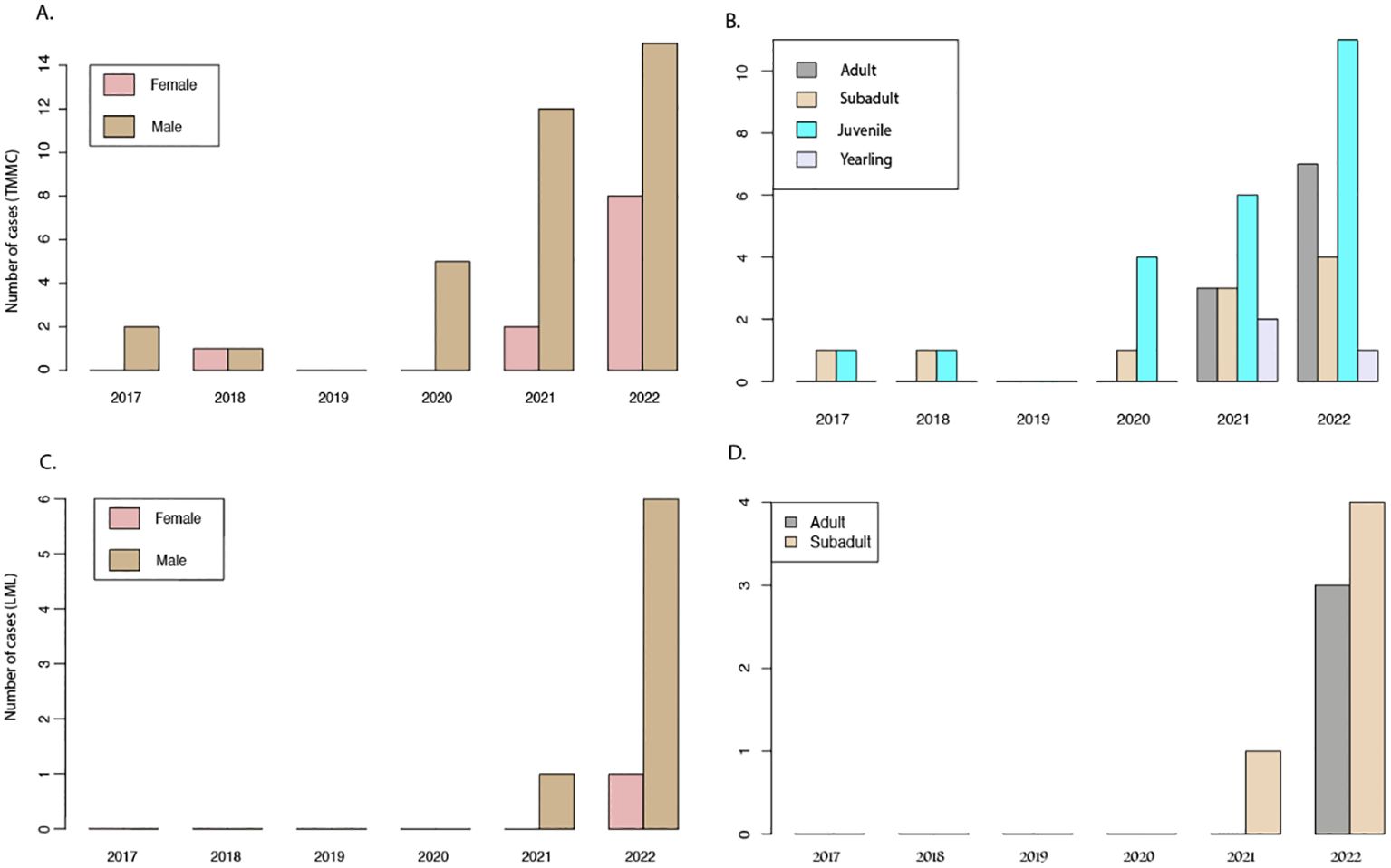
Figure 3. K. pneumoniae cases from The Marine Mammal Center (TMMC) and Long Marine Laboratory (LML) fim 2017 to 2022. (A) TMMC cases by sex; (B) TMMC cases by age; (C) LML cases by sex; (D) LML cases by age.
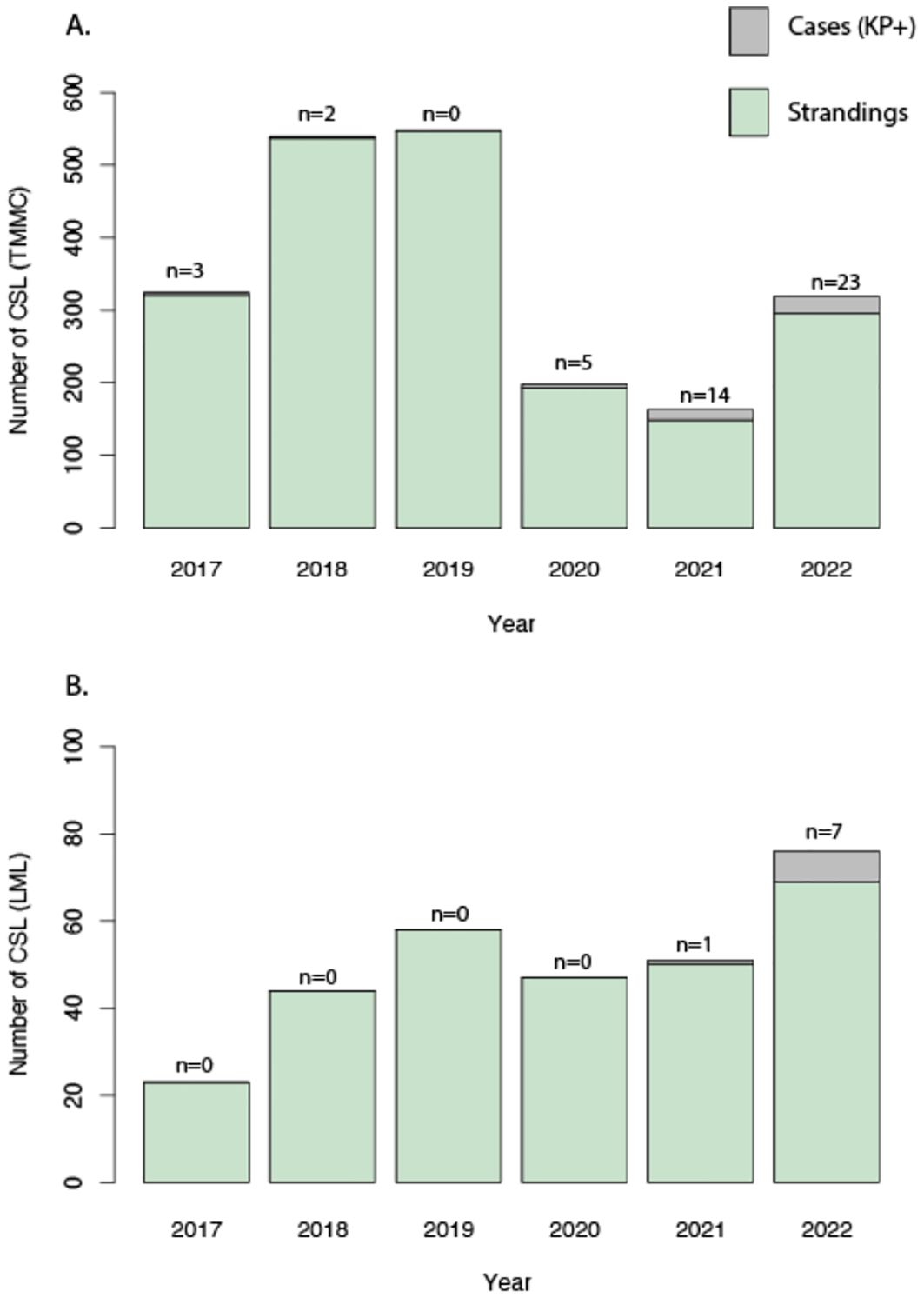
Figure 4. (A) Annual California Sea Lion (CSL) strandings and K. pneumoniae cases from The Marine Mammal Center (TMMC) between 2017 and 2022; (B) Annual California Sea Lion (CSL) strandings and K. pneumoniae cases from Long Marine Lab (LML) between 2017 and 2022. Grey bars and n values represent CSL strandings that tested positive for K. pneumoniae. Green bars represent CSL strandings that were not tested or tested negative for K. pneumoniae.
Whole Genome Sequencing analysis on all 8 LML isolates revealed a high nucleotide similarity (>99.9% average nucleotide identity) between any pair of the isolates. The Multilocus Sequence Typing (MST) showed that all eight isolates belonged to Sequence type 65 (ST65), a recently documented hypervirulent sequence of K. pneumoniae in humans (Lev et al., 2018; Zhao et al., 2021). Furthermore, the isolates contained the K2 polysaccharide capsule and siderophore aerobactin, characteristic of hypervirulent K. pneumoniae (Nassif and Sansonetti, 1986; Tan et al., 2019). Of the cases at TMMC, 70.8% (34/48) were suspected to be a hypermucoviscous form based on a positive string test on cultured colonies (Shon et al., 2013; Seguel et al., 2017).
4 Discussion
This investigation reports an increased incidence of HMV and non-HMV K. pneumoniae in stranded California sea lions in Central California, with an demonstrable increase from 2017 to 2022. Prior research on HMV K. pneumoniae in stranded CSLs has shown this pathogen to be of clinical and ecological importance, as well as extrapolatory evidence showing the potential for human health impacts and zoonoses (Jang et al., 2010; Whitaker et al., 2018; Hu et al., 2021). An increase in HMV K. pneumoniae cases is a major health concern for CSLs, other marine mammals, and public health due to concurrent research implicating this pathogen’s involvement in human disease outbreaks, as well as endangered New Zealand sea lion mortality events (Castinel et al., 2007a, b; Roe et al., 2015; Russo and Marr, 2019). Additionally, given the potential for K. pneumoniae to develop multi-drug resistance, there is a concern for community spread of antimicrobial resistance as well as the pathogen, in general (Li et al., 2013; Soto et al., 2020; Wang et al., 2020).
In this study, a higher number of K. pneumoniae cases were seen in male sea lions and more specifically, juveniles. This may be due to young, male CSLs migrating for long periods of time, long distances, and in congregations for breeding and foraging leading to increased opportunity for exposure to pathogens and high prevalence in the areas included in this study similar to trends seen in leptospirosis infections in similar populations (Gearin et al., 2017). Additionally, many of these infections could be opportunistic due to stress, co-infections, or immunosuppression in an individual, making it easier for the pathogen to cause severe illness, similar to other pathogens, climate, and environmental stressors further emphasizing the need for investigation and action into these drivers (Field et al., 2012; Banuet-Martínez et al., 2017; Peñín et al., 2018; DeRango et al., 2019). Overall, the demographics of the K. pneumoniae cases seen in this study parallel those seen in the CSL population of Northern California in general, and may represent an overall risk related to group dynamics, individual risk factors, and population bias (Gulland et al., 1996).
HMV forms of K. pneumoniae, have caused mass mortality events in endangered New Zealand sea lions (Castinel et al., 2007a, b; Roe et al., 2015) and have been tied to concerning clinical manifestations and community spread in CSLs as well (Jang et al., 2010; Whitaker et al., 2018). Longitudinal surveillance by stranding networks for emerging diseases and changes in pathogen prevalence, especially rapidly changing ones, is critical to swiftly respond to such events and assess their damage. All of the LML and 70.8% of TMMC cases were confirmed or suspected to be an HMV strain of K. pneumoniae. This suggests a more virulent phenotype of K. pneumoniae is persisting and possibly increasing in the population since being detected and characterized in a similar population in 2015-16 (Whitaker et al., 2018). This pathogen has the potential to more easily and rapidly infect hosts, is more difficult to remove from the environment and host, and, therefore, the potential to continue to spread and become problematic in sea lion populations (Whitaker et al., 2018; Soto et al., 2020).
Research to understand the ecological drivers of HMV strains, as well as how we can alter the approach to diagnosing, managing, and tracking K. pneumoniae cases, is important as a measure of preventing negative effects on the CSL population and the ecosystem in which they play a crucial role as apex predators and indicator species (Lowry et al., 1991; Laake et al., 2018; Lowry et al., 2022). The presence of both HMV and non-HMV phenotypes of K. pneumoniae in the same population poses an opportunity to further investigate the ecological dynamics and drivers of these distinct phenotypes, and potentially underlying health factors associated with infection by either. Even more importantly, understanding the broad biological and ecological behavior of K. pneumoniae and other similar emerging pathogens allows for preparation and response to potential outbreaks and spillover events. Although a limited number of isolates were evaluated, further characterization of HMV K. pneumoniae strains using WGS allowed for identification of identical virulence factors to those seen in human HMV K. pneumoniae strains, creating further opportunity for evaluation of the human, animal, and environmental players in the ecology of these pathogens (Shon et al., 2013; Seguel et al., 2017; Whitaker et al., 2018; Hu et al., 2021).
Given that K. pneumoniae is an emerging pathogen of concern in human and veterinary medicine, and its potential as a zoonotic pathogen with similar HMV strains being noted in humans as described in this study, an interdisciplinary approach is crucial to understanding possible transmission pathways and risk factors critical for effective prevention and control strategies (Jang et al., 2010; Whitaker et al., 2018; Hu et al., 2021). Future research on K. pneumoniae in CSLs should focus on unraveling the mechanisms associated with transmission, persistence, and ecology of the pathogen, host, and environment that connects them. Moreover, the presence of numerous established, emerging, and potential marine mammal zoonoses supports the role of marine mammals as sentinel organisms for monitoring human-animal spillover, in addition to the role they already serve as indicators marine ecosystem health (Bossart, 2011; Waltzek et al., 2012; Tryland, 2018).
Acknowledging the harmful impacts HMV and non-HMV K. pneumoniae phenotypes can have on humans and animals creates a unique and critical opportunity to investigate and address this human, animal, and environmental health issue sooner rather than later using an interdisciplinary, collaborative approach. Identification of this trend confirms the value of continued long-term surveillance by marine mammal stranding networks to identify and characterize major pathogens of concern and new trends in disease prevalence in their respective populations. Through interprofessional collaboration and partnerships between various stranding networks, early detection of concerning patterns and emerging pathogens is possible. Early detection may aid the prevention of spillover and disruption to the ecosystem, marine mammal populations, and human populations that are intimately connected.
Data availability statement
The raw datasets presented in this article are not readily available because of permitting restrictions and sensitive case information. Requests to access the datasets should be directed to the corresponding author.
Ethics statement
Ethical approval was not required for the studies on animals in accordance with the local legislation and institutional requirements because only commercially available established cell lines were used.
Author contributions
SZ: Conceptualization, Investigation, Methodology, Project administration, Supervision, Visualization, Writing – original draft, Writing – review & editing. RD: Conceptualization, Data curation, Funding acquisition, Investigation, Project administration, Supervision, Writing – review & editing. JL: Conceptualization, Investigation, Resources, Writing – review & editing. MS: Investigation, Resources, Validation, Writing – review & editing. LH: Conceptualization, Data curation, Formal analysis, Methodology, Validation, Writing – review & editing. MM: Conceptualization, Investigation, Project administration, Resources, Writing – review & editing. PD: Investigation, Resources, Validation, Writing – review & editing. CF: Conceptualization, Investigation, Writing – review & editing. ES: Investigation, Methodology, Writing – review & editing. DW: Investigation, Methodology, Writing – review & editing. CR: Investigation, Methodology, Writing – review & editing. MB: Data curation, Formal analysis, Investigation, Methodology, Resources, Software, Writing – review & editing. BJ: Data curation, Formal analysis, Investigation, Software, Writing – review & editing. SK: Writing – review & editing, Data curation, Formal Analysis, Investigation, Software. AK: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Resources, Software, Supervision, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. The authors acknowledge funding from the John H. Prescott Grant Program (NA22NMF4390245) and the State of California for Marine Mammal Stranding Networks to the Long Marine Lab Stranding Network.
Acknowledgments
We appreciate and acknowledge the efforts and expertise of Dr. Melissa Miller at the California Department of Fish and Wildlife with assistance in setting up the whole genome sequencing and general necropsy support for the Long Marine Lab Stranding Network. The authors also thank the staff, volunteers, and donors of The Marine Mammal Center who provided live animal care and performed the diagnostic workup of these cases under NOAA permit #s 18786 and 24359.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmars.2024.1484944/full#supplementary-material
References
Andrews S. (2019). FastQC (0.11.9) [Java] (Cambridge, UK: Babraham Institute). Available at: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (Accessed November 8, 2022).
Banuet-Martínez M., Espinosa-de Aquino W., Elorriaga-Verplancken F. R., Flores-Morán A., García O. P., Camacho M., et al. (2017). Climatic anomaly affects the immune competence of California sea lions. PLoS One 12, e0179359. doi: 10.1371/journal.pone.0179359
Bossart G. D. (2011). Marine mammals as sentinel species for oceans and human health. Veterinary Pathol. 48, 676–690. doi: 10.1177/0300985810388525
Castinel A., Duignan P. J., Pomroy W. E., López-Villalobos N., Gibbs N. J., Chilvers B. L., et al. (2007a). Neonatal mortality in New Zealand sea lions (Phocarctos hookeri) at Sandy Bay, Enderby island, Auckland islands from 1998 to 2005. J. Wildlife Dis. 43, 461–474. doi: 10.7589/0090-3558-43.3.461
Castinel A., Grinberg A., Pattison R., Duignan P., Pomroy B., Rogers L., et al. (2007b). Characterization of Klebsiella pneumoniae isolates from New Zealand sea lion (Phocarctos hookeri) pups during and after the epidemics on Enderby Island, Auckland Islands. Veterinary Microbiol. 122, 178–184. doi: 10.1016/j.vetmic.2006.12.024
Chang R., Miller M., Shahin K., Batac F., Field C. L., Duignan P., et al. (2022). Genetics and pathology associated with Klebsiella pneumoniae and Klebsiella spp. Isolates from North American Pacific coastal marine mammals. Veterinary Microbiol. 265, 109307. doi: 10.1016/j.vetmic.2021.109307
Chen S., Zhou Y., Chen Y., Gu J. (2018). fastp: An ultra-fast all-in-one FASTQ preprocessor. Bioinformatics 34, i884–i890. doi: 10.1093/bioinformatics/bty560
Clegg S., Murphy C. N. (2016). Epidemiology and virulence of Klebsiella pneumoniae. Microbiol. Spectr. 4, 4.1.06. doi: 10.1128/microbiolspec.UTI-0005-2012
DeRango E. J., Prager K. C., Greig D. J., Hooper A. W., Crocker D. E. (2019). Climate variability and life history impact stress, thyroid, and immune markers in California sea lions (Zalophus californianus) during El Niño conditions. Conserv. Physiol. 7, coz010. doi: 10.1093/conphys/coz010
Doud M. S., Grimes-Zeppegno R., Molina E., Miller N., Balachandar D., Schneper L., et al. (2009). A k 2 A -positive Klebsiella pneumoniae causes liver and brain abscess in a Saint Kitt’s man. Int. J. Med. Sci. 6, 301-304. doi: 10.7150/ijms.6.301
Duignan P. (2023). “Aquatic mammals’. Chapter 6,” in Pathology and Epidemiology of Aquatic Animal Diseases for Practitioners. Eds. Urdes L., Walster C., Tepper J. (John Wiley & Sons Ltd, Singapore), 214–350.
Fang C.-T., Chuang Y.-P., Shun C.-T., Chang S.-C., Wang J.-T. (2004). A novel virulence gene in Klebsiella pneumoniae strains causing primary liver abscess and septic metastatic complications. J. Exp. Med. 199, 697–705. doi: 10.1084/jem.20030857
Field C. L., Tuttle A. D., Sidor I. F., Nyaoke A., Deering K. M., Gilbert-Marcheterre K., et al. (2012). Systemic mycosis in a California Sea Lion (Zalophus californianus) with detection of cystofilobasidiales DNA. J. Zoo Wildlife Med. 43, 144–152. doi: 10.1638/2011-0187.1
Gearin P. J., Melin S. R., DeLong R. L., Gosho M. E., Jeffries S. (2017). Migration patterns of adult male California sea lions (Zalophus californianus). U.S. Dep. Commer., NOAA Tech. Memo. NMFS-AFSC-346, 29 p. doi: 10.7289/V5/TM-AFSC-346
Greig D. J., Gulland F. M. D., Kreuder C. (2005). A decade of live California Sea Lion (Zalophus californianus) strandings along the central California coast: causes and trends 1991-2000. Aquat. Mammals 31, 11–22. doi: 10.1578/AM.31.1.2005.11
Gulland F. M. D., Koski M., Lowenstine L. J., Colagross A., Morgan L., Spraker T. (1996). Leptospirosis in California Sea Lions (Zalophus californianus) stranded along the central California coast 1981-1994. J. Wildlife Dis. 32, 572–580. doi: 10.7589/0090-3558-32.4.572
Han S. H. (1995). Review of hepatic abscess from Klebsiella pneumoniae. An association with diabetes mellitus and septic endophthalmitis. Western J. Med. 162, 220–224.
Hu Y., Anes J., Devineau S., Fanning S. (2021). Klebsiella pneumoniae: prevalence, reservoirs, antimicrobial resistance, pathogenicity, and infection: A hitherto unrecognized zoonotic bacterium. Foodborne Pathog. Dis. 18, 63–84. doi: 10.1089/fpd.2020.2847
Jain C., Rodriguez-R L. M., Phillippy A. M., Konstantinidis K. T., Aluru S. (2018). High throughput ANI analysis of 90K prokaryotic genomes reveals clear species boundaries. Nat. Commun. 9, 5114. doi: 10.1038/s41467-018-07641-9
Jang S., Wheeler L., Carey R. B., Jensen B., Crandall C. M., Schrader K. N., et al. (2010). Pleuritis and suppurative pneumonia associated with a hypermucoviscosity phenotype of Klebsiella pneumoniae in California sea lions (Zalophus californianus). Veterinary Microbiol. 141, 174–177. doi: 10.1016/j.vetmic.2009.07.032
Laake J. L., Lowry M. S., DeLong R. L., Melin S. R., Carretta J. V. (2018). Population growth and status of california sea lions. J. Wildlife Manage. 82, 583–595. doi: 10.1002/jwmg.21405
Lam M. M. C., Wick R. R., Watts S. C., Cerdeira L. T., Wyres K. L., Holt K. E. (2021). A genomic surveillance framework and genotyping tool for Klebsiella pneumoniae and its related species complex. Nat. Commun. 12, 4188. doi: 10.1038/s41467-021-24448-3
Lederman E. R., Crum N. F. (2005). Klebsiella liver abscess: A coast-to-coast phenomenon. Clin. Infect. Dis. 41, 273. doi: 10.1086/cid.2005.41.issue-2
Lev A. I., Astashkin E. I., Kislichkina A. A., Solovieva E. V., Kombarova T. I., Korobova O. V., et al. (2018). Comparative analysis of Klebsiella pneumoniae strains isolated in 2012–2016 that differ by antibiotic resistance genes and virulence genes profiles. Pathog. Global Health 112, 142–151. doi: 10.1080/20477724.2018.1460949
Li W., Sun G., Yu Y., Li N., Chen M., Jin R., et al. (2014). Increasing occurrence of antimicrobial-resistant hypervirulent (Hypermucoviscous) Klebsiella pneumoniae isolates in China. Clin. Infect. Dis. 58, 225–232. doi: 10.1093/cid/cit675
Lowry M., Nehasil S., Moore J. (2022). Spatio-temporal diet variability of the California sea lion Zalophus californianus in the southern California Current Ecosystem. Mar. Ecol. Prog. Ser. 692, 1–21. doi: 10.3354/meps14096
Lowry M. S., Stewart B. S., Heath C. B., Yochem P. K., Francis J. M. (1991). Seasonal and Annual Variability in the Diet of California Sea Lions Zalophus californianus at San Nicolas Island, California 1981-86. Fishery Bull. 89, 331–336.
McIver C. J., Janda J. M. (2008). Pathogenesis and laboratory identification of emerging hepatovirulent Klebsiella pneumoniae. Clin. Microbiol. Newslett. 30, 127–131. doi: 10.1016/j.clinmicnews.2008.08.001
Michael S. A., Hayman D. T. S., Gray R., Roe W. D. (2022). Clinical parameters of hypervirulent Klebsiella pneumoniae disease and ivermectin treatment in New Zealand sea lion (Phocarctos hookeri) pups. PLoS One 17, e0264582. doi: 10.1371/journal.pone.0264582
Nassif X., Sansonetti P. J. (1986). Correlation of the virulence of Klebsiella pneumoniae K1 and K2 with the presence of a plasmid encoding aerobactin. Infection Immun. 54, 603–608. doi: 10.1128/iai.54.3.603-608.1986
NOAA Fisheries (2024). West Coast Marine Mammal Stranding Network — California Jurisdictions. United States Department of Commerce - National Oceanographic and Atmospheric Administration. Available online at: https://www.fisheries.noaa.gov/west-coast/marine-mammal-protection/west-coast-marine-mammal-stranding-network (Accessed July 1, 2024).
Peñín I., Levin M., Acevedo-Whitehouse K., Jasperse L., Gebhard E., Gulland F. M. D., et al. (2018). Effects of polychlorinated biphenyls (PCB) on California sea lion (Zalophus californianus) lymphocyte functions upon in vitro exposure. Environ. Res. 167, 708–717. doi: 10.1016/j.envres.2018.08.028
Podschun R., Ullmann U. (1998). Klebsiella spp. as nosocomial pathogens: epidemiology, taxonomy, typing methods, and pathogenicity factors. Clin. Microbiol. Rev. 11, 589–603. doi: 10.1128/CMR.11.4.589
Qian Y., Wong C. C., Lai S., Chen H., He X., Sun L., et al. (2016). A retrospective study of pyogenic liver abscess focusing on Klebsiella pneumoniae as a primary pathogen in China from 1994 to 2015. Sci. Rep. 6, 38587. doi: 10.1038/srep38587
Roe W. D., Rogers L., Pinpimai K., Dittmer K., Marshall J., Chilvers B. L. (2015). Septicaemia and meningitis caused by infection of New Zealand sea lion pups with a hypermucoviscous strain of Klebsiella pneumoniae. Veterinary Microbiol. 176, 301–308. doi: 10.1016/j.vetmic.2015.01.019
Russo T. A., Marr C. M. (2019). Hypervirulent Klebsiella pneumoniae. Clin. Microbiol. Rev. 32. doi: 10.1128/CMR.00001-19
Sanderson C. E., Alexander K. A. (2020). Unchartered waters: Climate change likely to intensify infectious disease outbreaks causing mass mortality events in marine mammals. Global Change Biol. 26, 4284–4301. doi: 10.1111/gcb.15163
Santaniello A., Sansone M., Fioretti A., Menna L. F. (2020). Systematic review and meta-analysis of the occurrence of ESKAPE bacteria group in dogs, and the related zoonotic risk in animal-assisted therapy, and in animal-assisted activity in the health context. Int. J. Environ. Res. Public Health 17, 3278. doi: 10.3390/ijerph17093278
Seguel M., Gottdenker N. L., Colegrove K., Johnson S., Struve C., Howerth E. W. (2017). Hypervirulent Klebsiella pneumoniae in California Sea Lions (Zalophus californianus): Pathologic Findings in Natural Infections. Veterinary Pathol. 54, 846–850. doi: 10.1177/0300985817705172
Shon A. S., Bajwa R. P. S., Russo T. A. (2013). Hypervirulent (hypermucoviscous) Klebsiella pneumoniae: A new and dangerous breed. Virulence 4, 107–118. doi: 10.4161/viru.22718
Soto E., Abdelrazek S. M. R., Basbas C., Duignan P. J., Rios C., Byrne B. A. (2020). Environmental persistence and disinfectant susceptibility of Klebsiella pneumoniae recovered from pinnipeds stranded on the California Coast. Veterinary Microbiol. 241, 108554. doi: 10.1016/j.vetmic.2019.108554
Soto E., LaMon V., Griffin M., Keirstead N., Beierschmitt A., Palmour R. (2012). Phenotypic and genotypic characterization of Klebsiella pneumoniae isolates recovered from nonhuman primates. J. Wildlife Dis. 48, 603–611. doi: 10.7589/0090-3558-48.3.603
Souvorov A., Agarwala R., Lipman D. J. (2018). SKESA: Strategic k-mer extension for scrupulous assemblies. Genome Biol. 19, 153. doi: 10.1186/s13059-018-1540-z
Spraker T. R., DeLong R. L., Lyons E. T., Melin S. R. (2007). Hookworm enteritis with bacteremia in California Sea Lion pups on San Miguel island. J. Wildlife Dis. 43, 179–188. doi: 10.7589/0090-3558-43.2.179
Tan D., Zhang Y., Cheng M., Le S., Gu J., Bao J., et al. (2019). Characterization of Klebsiella pneumoniae ST11 isolates and their interactions with lytic phages. Viruses 11, 1080. doi: 10.3390/v11111080
Tryland M. (2018). “Zoonoses and public health,” in CRC Handbook of Marine Mammal Medicine, 3rd ed. Eds. Gulland F. M. D., Dierauf L. A., Whitman K. L. (Boca Raton, FL, USA: CRC Press: Taylor & Francis Group), 47–62.
Waltzek T. B., Cortés-Hinojosa G., Wellehan J. J.F.X., Gray G. C. (2012). Marine mammal zoonoses: A review of disease manifestations. Zoonoses Public Health 59, 521–535. doi: 10.1111/j.1863-2378.2012.01492.x
Wang G., Zhao G., Chao X., Xie L., Wang H. (2020). The characteristic of virulence, biofilm and antibiotic resistance of Klebsiella pneumoniae. Int. J. Environ. Res. Public Health 17, 6278. doi: 10.3390/ijerph17176278
Whitaker D. M., Reichley S. R., Griffin M. J., Prager K., Richey C. A., Kenelty K. V., et al. (2018). Hypermucoviscous Klebsiella pneumoniae isolates from stranded and wild-caught marine mammals of the US pacific coast: prevalence, phenotype, and genotype. J. Wildlife Dis. 54, 659–670. doi: 10.7589/2017-07-178
Zhang W., Zhu Y., Wang C., Liu W., Li R., Chen F., et al. (2019). Characterization of a Multidrug-Resistant Porcine Klebsiella pneumoniae Sequence Type 11 Strain Coharboring blaKPC-2 and fosA3 on Two Novel Hybrid Plasmids. mSphere 4, e00590–e00519. doi: 10.1128/mSphere.00590-19
Keywords: California sea lion (Zalophus californianus), Klebsiella pneumoniae, hypermucoviscosity, whole-genome sequencing (WGS), Central California Coast
Citation: Zotto SN, Dunkin RC, Limon JD, Smith MS, Hortensius LM, Martinez ME, Duignan PJ, Field C, Soto E, Whitaker D, Rios C, Byukusenge M, Jayarao B, Kuchipudi SV and Kinsley AC (2025) Klebsiella pneumoniae in stranded California sea lions (Zalophus californianus) of Central California between 2017-2022. Front. Mar. Sci. 11:1484944. doi: 10.3389/fmars.2024.1484944
Received: 22 August 2024; Accepted: 20 December 2024;
Published: 16 January 2025.
Edited by:
Lyne Morissette, M – Expertise Marine, CanadaReviewed by:
Wanchun Guan, Wenzhou Medical University, ChinaAdrianne Akmajian, Makah Tribal Council, United States
Copyright © 2025 Zotto, Dunkin, Limon, Smith, Hortensius, Martinez, Duignan, Field, Soto, Whitaker, Rios, Byukusenge, Jayarao, Kuchipudi and Kinsley. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sydney N. Zotto, am9oMTc3NThAdW1uLmVkdQ==